
Submitted:
28 November 2023
Posted:
28 November 2023
You are already at the latest version
Abstract
Background: Inflammasomes recognize cytosolic danger signals, including pathogens, and subsequently induce the secretion of pro-inflammatory cytokines, such as interleukin (IL)-1β. Studying inflammasomes in the red fox (Vulpes vulpes) is crucial from a wildlife veterinary and public health perspective, as it can be used to help control and prevent the transmission of zoonotic pathogens. The understanding of how diversity in inflammasomes across species affects pathogen carriage in the red fox is still limited. Methods: We investigated the activation and intracellular mechanisms of three inflammasomes (NLRP3, AIM2, and NLRC4) in fox peripheral blood mononuclear cells (PBMCs), using established triggers and inhibitors derived from humans and mice. Results: Fox PBMCs exhibited normal activation and induction of IL-1β secretion in response to representative inflammasome triggers (ATP and nigericin for NLRP3, dsDNA for AIM2, flagellin for NLRC4). However, in inhibitor studies, MCC950, a selective NLRP3 inhibitor, suppressed IL-1β secretion mediated by the fox AIM2 and NLRC4 inflammasomes. Conclusions: These findings suggest that NLRP3 may have a common role in dsDNA- and flagellin-mediated inflammasome activation in the red fox. In conclusion, this fox inflammasome biology can be applied to the control of zoonotic infections and the treatment of inflammasome-mediated diseases in the red fox.
Keywords:
Red foxes
; Vulpes vulpes
; inflammasome
; cytokine
; interleukin-1beta
1. Introduction
Inflammasomes are multiprotein complexes that are assembled by cytoplasmic pattern recognition receptors (PRRs) in innate immune cells, such as monocytes and macrophages, responsible for innate immunity [1,2]. They recognize pathogens and other danger signals and trigger inflammatory responses through the activation of inflammatory caspases [3,4]. Inflammasomes are composed of sensor proteins, adaptor proteins (apoptosis-associated speck-like protein containing a caspase recruitment domain [ASC]), and effector proteins (e.g., inflammatory caspases). Inflammasomes are named based on the sensor protein they utilize [1,2]. The nucleotide-binding oligomerization domain, leucine rich repeat, and pyrin domain containing 3 (NLRP3) sensor detects various endogenous and exogenous danger signals, the absent in melanoma 2 (AIM2) and AIM2-like receptors recognize cytoplasmic double-stranded DNA (dsDNA), and the nod-like receptor (NLR) family caspase recruitment domains (CARD) domain-containing protein 4 (NLRC4) specifically detects bacterial proteins (e.g., flagellin), all of which trigger the assembly of their respective inflammasomes: NLRP3, AIM2, and NLRC4 [1,3]. The activation of inflammasomes plays a crucial role in driving the maturation and secretion of pro-inflammatory cytokines, such as interleukin (IL)-1β and IL-18. Additionally, inflammasomes can induce inflammatory cell death known as pyroptosis by forming pores in the cell membrane, leading to the release of inflammatory mediators [1].
The activation of inflammasomes requires two steps: priming and activation [5]. During the priming step, the expression of genes encoding inflammasome components is upregulated, preparing the cell to form inflammasome complexes [3]. In the activation step, each component binds together to assemble the inflammasome, leading to the activation of caspase-1 and subsequent release of cytokines, as well as the induction of pyroptosis [3]. Lipopolysaccharide (LPS), which is known to induce the priming step, specifically binds to toll-like receptor (TLR) 4, leading to the activation of nuclear factor kappa B (NF-κB) signaling and subsequent expression of target genes [5]. Triggers that induce the activation step of the NLRP3 inflammasome include adenosine triphosphate (ATP), nigericin, and calcium chloride [6]. Additionally, the NLRC4 and AIM2 inflammasomes are activated by the cytosolic delivery of flagellin and dsDNA [6,7]. Furthermore, inflammasomes selectively operate in response to various bacterial pathogens, such as Staphylococcus aureus, Salmonella Typhimurium, Listeria monocytogenes, and Escherichia coli [4]. The mechanism of action of inflammasomes has been predominantly elucidated in the NLRP3 inflammasome [4]. It is known that intracellular K+ efflux and the generation of reactive oxygen species (ROS) serve as cellular signaling mechanisms that trigger the assembly of the inflammasome complex [1]. Inflammasomes also play a crucial role in immune responses that drive age-related and metabolic disorders, such as Alzheimer’s disease and diabetes [3]. Additionally, inflammasomes have an impact on the prevention and management of infectious diseases [8].
The red fox, Vulpes vulpes (Carnivora, Canidae) is a small carnivorous mammal that is widely distributed worldwide. Red foxes were known to inhabit Korea in the past, but became extinct due to indiscriminate hunting and illegal killing [9]. The red fox has been one of the animal species included in the endangered species restoration project in the Republic of Korea since 2011 [9]. Due to this project, red foxes could inhabit human residential areas, making them potential hosts for transmitting various diseases, including rabies, to human populations, similar to raccoon dogs [10]. The study of the inflammasome biology of wildlife including the red fox plays a crucial role in understanding and addressing transmission of zoonotic infectious diseases [11]. For instance, bats exhibit a dampened inflammatory response to viral infections due to alterations in their inflammasomes [12,13]. Moreover, ducks possess an active retinoic acid-inducible gene (RIG)-I inflammasome which grants them a heightened resistance against influenza viruses [14,15]. Conversely, chickens lack the RIG-I, rendering them highly susceptible to influenza viruses [14]. Therefore, the current study sought to characterize fox inflammasomes, to obtain essential insights for the development of treatments for inflammasome-related diseases and the control of zoonotic infectious diseases spread through red foxes.
2. Materials and Methods
2.1. Cell preparation
Peripheral blood monocytes (PBMCs) were isolated from freshly drawn blood from Korean red foxes (Wildlife Medical Center, Korea National Park Research Institute, Republic of Korea) and beagle dogs (Veterinary Medical Teaching Hospital, Kangwon National University, Republic of Korea) as per the procedure followed in a previous study [7]. Briefly, the blood was drained into heparin tubes and moved to the laboratory. The blood was mixed with an equal volume of phosphate-buffered saline (PBS) in centrifuge tubes (SPL Life Science Co., Gyeonggi-do, Republic of Korea), and then lymphocyte separation medium (LSM; Thermo Fisher Scientific, Waltham, MA, USA) was added into the tube (blood: PBS: LSM = 1: 1: 1). After centrifuging (500 × g, room temperature [RT], 30 min), the interphase of the mixture containing monocytes were harvested, and placed in a new tube. After centrifuging (500 × g, RT, 10 min), the remaining cell pellets were added to Red Blood Cell Lysis Buffer (iNtRON Biotech., Sengnam-si, Republic of Korea) and subjected to centrifuging (500 × g, RT, 10 min). PBMCs were used for experiments immediately on the day they were prepared. Mouse bone marrow-derived macrophages (BMDMs) were differentiated from marrow cells (C57BL/6, Narabio Co., Seoul, Republic of Korea) in media containing 30% L929 cell-conditioned media, 10% fetal bovine serum (FBS), and antibiotics for 7 days as per the procedure followed in the previous study [16]. Animal experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals, National Research Council Committee for the Update of the Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care, and Use Committee of Kangwon National University (IACUC; approval KW-210317-2 for mice, KW-210331-2 for foxes, and KW-210504-1 for dogs). All cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) or Roswell Park Memorial Institute (RPMI) medium (Welgene Inc., Gyeongsangbuk-do, Republic of Korea) containing 10% FBS (Welgene Inc.) and antibiotics (penicillin and streptomycin; Welgene Inc.) at 37°C in a 5% CO2 atmosphere.
2.2. Inflammasome activation and inhibition
Cells (2 × 106 cells per well for PBMCs, or 1 × 106 cells per well for BMDMs) were plated in 12-well plates (SPL Life Science), and then treated with lipopolysaccharide (LPS; #L4130, Sigma-Aldrich, St. Louis, MI, USA) in RPMI media containing 10% FBS and antibiotics to induce the priming step. The LPS-primed cells were further treated with the following triggers to activate inflammasome; ATP (InvivoGen, San Diego, CA, USA) for 1 h, nigericin (NG; Tocris Bioscience, Bristol, UK) for 1 h, calcium chloride (CaCl2; iNtRON Biotech.) for 3 h, dsDNA with jetPRIME™ (Polyplus-transfection Inc., Illkirch, France) for 1 h, flagellin (InvivoGen) with Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) for 3h, and bacterial triggers (Staphylococcus aureus for 6 h, Salmonella Typhimurium for 1 h, Listeria monocytogenes for 3 h, and Escherichia coli for 6 h) [1]. The bacteria were cultured at 37°C on Luria-Bertani media (Condalab, Torrejón de Ardoz, Madrid, Spain) for S. aureus, S. Typhimurium, E. coli, or Brain Heart Infusion media (Condalab) for L. monocytogenes. For inflammasome inhibition assessment, the LPS-primed cells were treated with diphenyleneiodonium (DPI, 100 μM, Tocris Bioscience). high potassium solution (KCl, 50 mM; iNtRON Biotech.), glibenclamide (150 μM; Santa Cruz Biotech., Santa Cruz, CA, USA), or MCC950 (200 nM; InvivoGen) in the presence of NG (40 μM) for 1 h, dsDNA (4 μg/mL) with jetPRIME (5 μL/mL) for 1 h, or flagellin (500 ng/mL) with Lipofectamin 2000 (10 μL/mL) for 3 h [17,18,19,20]. After the activation step, the cellular supernatant and lysate were prepared for use in further analyses.
2.3. Immunoblotting
The supernatant and lysate samples were separated with electrophoresis using 10% or 16% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels (Mini-PROTEAN®® TETRA Handcast system; BIO-RAD, Hercules, CA, USA). Subsequently, the gels were transferred onto a polyvinylidene difluoride membrane (PVDF, Thermo Fisher Scientific). The membranes were blocked with 3% skim milk and probed with anti-dog IL-1β sera (AF3747, R&D Systems, Minneapolis, MN, USA) overnight at 4°C. In addition, the membrane was probed with horseradish peroxidase (HRP) secondary antibody (Thermo Fisher Scientific) for 1 h and then visualized by applying a chemiluminescence solution (Abfrontier, Seoul, Republic of Korea) and a chemiluminescence imaging system (EZ-Capture II, ATTO Technology, Tokyo, Japan). The membranes were stripped and re-probed with anti-actin sera (#sc-1615, Santa Cruz Biotechnology).
2.4. Assay for IL-1β secretion
The IL-1β levels in the supernatant were measured by using an enzyme-linked immunosorbent assay (ELISA) kit (DY3747 for foxes and DY401 for mice, R&D Systems). The plates were analyzed using a multi-microplate spectrophotometer (Synergy™ H1 Hybrid Multi-Mode Reader, BioTek, Winooski, VT, USA).
2.5. Statistical analysis
Statistical software (GraphPad Prism 6, San Diego, CA) was used to perform analyses using the Mann-Whitney or Kruskal-Wallis tests. The p-value is shown in the figures.
3. Results
3.1. Optimization of inflammasome priming and activation in fox PBMCs
The gene expression of inflammasome components and inflammatory cytokines in red foxes was analyzed using specific primers designed for dogs due to limited information on the fox genes (Supplemental Figure S1 and Table S1). Although the gene sequences of red foxes and dogs are not identical (Supplemental Data S1), it was confirmed that the dog primers used are functional in red foxes as well. Upon treatment with LPS, both dog and fox PBMCs exhibited increased expression of inflammasome component genes (pro-IL-1β and NLRP3) and inflammatory cytokine genes (IL-1α, tumor necrosis factor [TNF]α, IL-6, and IL-10) except for ASC. It indicates that LPS induces gene expression through the TLR4/NF-κB signaling pathway in fox PBMCs, suggesting that LPS can serve as an inducer for the priming step in fox PBMCs, similar to other animals. Moreover, these findings imply that antibodies designed for dogs can be used to assess inflammasome activation in red foxes.
Both dog and fox PBMCs were primed with LPS and subsequently their activation step was induced by inflammasome triggers, such as NG and ATP (Figure 1A). As shown in Figure 1B, LPS-primed cells of dogs and red foxes exhibited the expression of pro-IL-1β protein in the cellular lysate. The pro-IL-1β (around 31 kDa) was cleaved into IL-1β (p17) upon the NG- and ATP-mediated inflammasome activation, and subsequently released from the cells into the cellular supernatant. This indicates that dog IL-1β antibodies can detect the secreted fox IL-1β (Supplemental Data S2), a readout of inflammasome activation. Furthermore, it was revealed that the known triggers of NLRP3 inflammasome, namely NG and ATP, can induce activation of inflammasomes in red foxes.
3.2. NLRP3 inflammasome triggers in fox PBMCs
To optimize the priming step, which is crucial for NLRP3 inflammasome activation [5], fox PBMCs were treated with various concentrations of LPS in the presence or absence of NLRP3 triggers (ATP and NG). Treatment with LPS in a concentration-dependent manner led to a small but detectable secretion of IL-1β through LPS priming alone (Figure 2A). However, no significant increase in IL-1β secretion was observed. Furthermore, there were no significant differences in IL-1β secretion when ATP and NG were administered across different LPS concentrations. ELISA-based studies consistently demonstrated the selective detection of cleaved fox IL-1β (p17) in the cellular supernatant using the dog IL-1β ELISA kit (Supplemental Figure S2A). Therefore, priming with 1ng/mL LPS ensures robust NLRP3 inflammasome activation in fox PBMCs. Subsequently, the concentration-dependent effects of NLRP3 triggers, including ATP, NG, and CaCl2, were evaluated for NLRP3 inflammasome activation in fox PBMCs (Figure 2B). The results revealed that all three NLRP3 triggers significantly induced the secretion of IL-1β (p17) in a concentration-dependent manner. In conclusion, the NLRP3 inflammasome is effectively activated by the NLRP3 triggers in red foxes.
3.3. AIM2 and NLRC4 inflammasome triggers in fox PBMCs
The NLRP3 inflammasome can recognize both danger- and pathogen-associated molecular patterns (DAMPs and PAMPs), while the AIM2 and NLRC4 inflammasomes specifically specialize in detecting PAMPs [1]. To activate the fox AIM2 inflammasome, cytoplasmic dsDNA was introduced into the cells. Similarly, the fox NLRC4 inflammasome was activated by delivering flagellin into the cytoplasm. The optimal concentration of LPS for priming the AIM2 inflammasome in red foxes was determined by evaluating various concentrations of LPS, and it was found that 1 ng/mL of LPS priming resulted in the optimal secretion of IL-1β (Figure 3A and Supplemental Figure S2B). In LPS-primed fox PBMCs, the introduction of dsDNA into the cytoplasm led to a concentration-dependent secretion of IL-1β (Figure 3B). Similarly, the appropriate LPS priming concentration for fox NLRC4 inflammasome activation was determined to be 1 ng/mL, which induced optimal secretion of IL-1β (Figure 3C). As shown in Figure 3D, LPS-primed fox PBMCs treated with various concentrations of flagellin introduced into the cytoplasm exhibited a concentration-dependent secretion of IL-1β. In conclusion, dsDNA and flagellin, the well-known triggers of AIM2 and NLRC4, were effective in activating the AIM2 and NLRC4 inflammasomes in red foxes.
3.4. Bacterial inflammasome triggers in fox PBMCs
The host detects pathogen invasion through cytosolic PRRs such as NLRP3, AIM2, and NLRC4, and induces immune responses to resistant infection through the inflammasome assembly [4]. Various types of inflammasomes have been identified that are activated depending on the type of pathogen [1,3]. In this study, the LPS-primed fox PBMCs and mouse BMDMs were inoculated with the well-characterized bacteria (i.e., Staphylococcus aureus, Salmonella Typhimurium, Listeria monocytogenes, and Escherichia coli) to trigger the inflammasome activation. These bacteria can simultaneously stimulate more than one type of inflammasome [4]. The secretion of IL-1β was measured as an indicator of inflammasome activation (Figure 4). All the pathogens induced IL-1β secretion from fox PBMCs in a dose-dependent manner, comparable to that from mouse BMDMs (Supplemental Figure S3). Taken together, it was found that the bacterial triggers, known for inflammasome activation in the humans and mice, were capable of activating inflammasomes in fox cells.
3.5. Mechanistic studies of fox inflammasomes
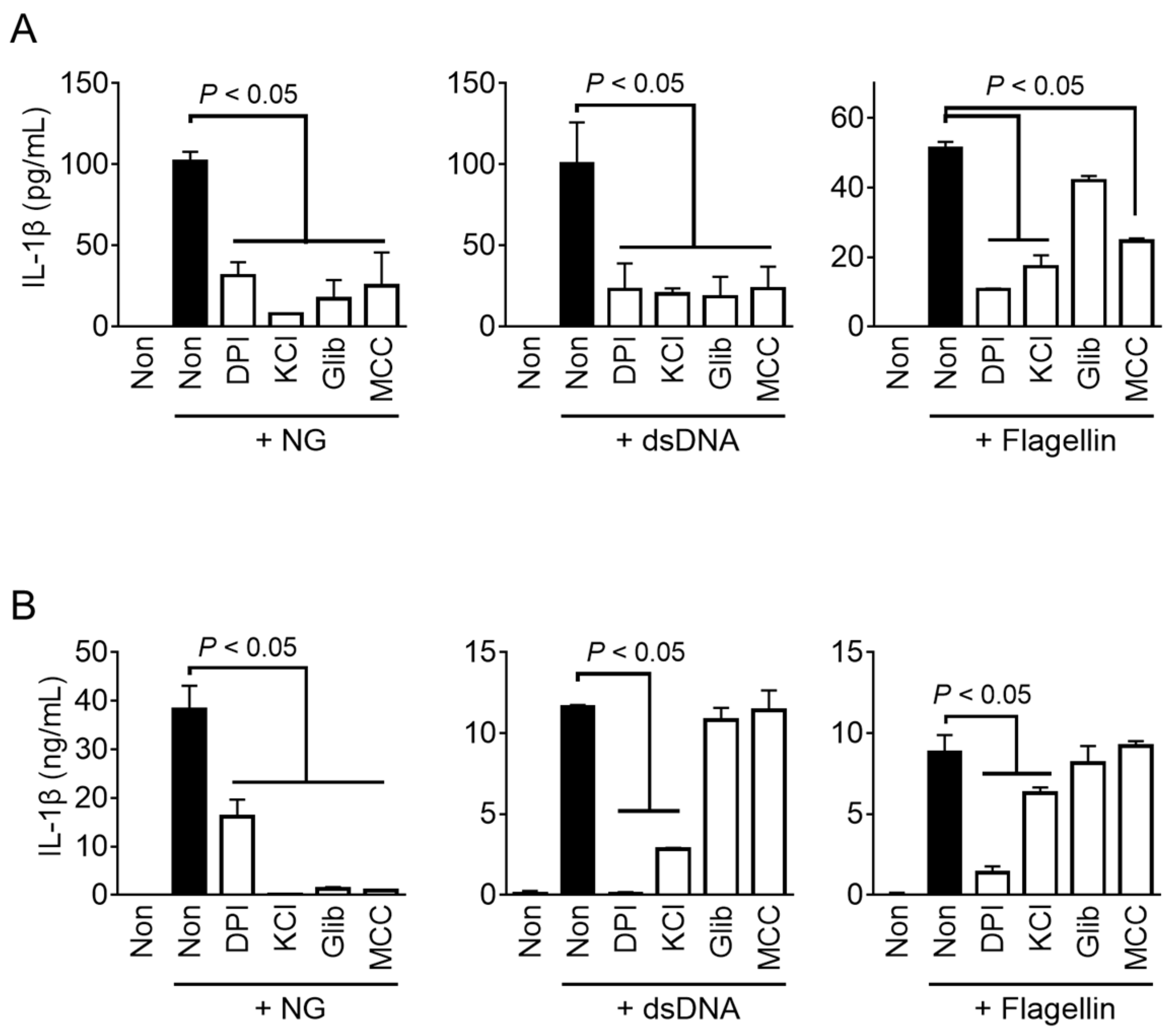
Inflammasome sensors directly or indirectly recognize danger signals (e.g., ROS or K+ efflux) or danger from specific factors (e.g., dsDNA or flagellin) in the cytoplasm, and initiate the assembly of the inflammasomes [1,2]. To investigate the cytosolic signaling involved in fox inflammasome activation, representative inhibitors of inflammasome activation were utilized to treat fox PBMCs during the activation step of NLRP3, AIM2, and NLRC4 inflammasomes (Figure 5A). The results showed that the activation of NLRP3 and AIM2 inflammasomes in fox PBMCs was inhibited when treated with a scavenger of ROS (DPI), inhibitors of potassium efflux (KCl and glibenclamide), and a selective inhibitor of NLRP3 inflammasome assembly (MCC950). Conversely, the activation of the fox NLRC4 inflammasome mediated by flagellin was suppressed by DPI, KCl, and MCC950, while no significant change was observed with glibenclamide. The results (Figure 5B) showed that all inhibitors suppressed activation of NLRP3 inflammasome mediated by NG in both foxes and mice. However, glibenclamide and MCC950 were unable to inhibit AIM2 and NLRC4 inflammasome activation in mice. Interestingly, the NLRC4 and AIM2 inflammasomes in red foxes were found to be suppressed by MCC950, which is known as a selective inhibitor of NLRP3 inflammasome in humans and mice [21]. Overall, the NLRP3 inflammasome shares the same intracellular signaling pathway in both mice and red foxes, but the pathway of AIM2 and NLRC4 inflammasomes in red foxes may operate differently from those in mice.
4. Discussion
In this study, the response of fox PBMCs to LPS priming and inflammasome triggers for the priming and activation steps was similar to that of other species [5,7,22,23]. However, differences were observed in inhibitor studies (Figure 5). Both fox and mouse NLRP3 showed similar responses to inhibitors. Specifically, the ROS scavenger (DPI), K+ efflux inhibitors (KCl and glibenclamide), and NLRP3 selective inhibitor (MCC950) suppressed IL-1β secretion induced by NG-mediated NLRP3 inflammasome activation in both foxes and mice. These intracellular molecular pathways have been validated as NLRP3 inflammasome activation pathways in humans and mice [2]. However, for the AIM2 inflammasome triggered by dsDNA transfection, the results observed for red foxes and mice were different. In red foxes, all the inhibitors (DPI, KCl, glibenclamide, and MCC950) suppressed dsDNA-induced IL-1β secretion, whereas, in mice, suppression was only observed with DPI and KCl. Both KCl and glibenclamide inhibit K+ efflux, the upstream pathway of NLRP3 inflammasome activation [2], but a high concentration of KCl inhibits it via high hydrostatic pressure [17], while glibenclamide inhibits it via ATP-sensitive potassium channels [18]. Therefore, glibenclamide is selective for NLRP3, while KCl inhibits the AIM2 inflammasome [17,18]. MCC950, known as a selective NLRP3 inhibitor [19,21], directly inhibits NLRP3 but does not inhibit IL-1β secretion induced by the AIM2 inflammasome in mice. However, MCC950 inhibited IL-1β secretion induced by dsDNA transfection in red foxes in the current study. Regarding NLRC4 inflammasome activation triggered by flagellin, DPI, KCl, and MCC950 inhibited IL-1β secretion in red foxes. In mice, DPI inhibited IL-1β secretion mediated by flagellin, and partial inhibition was observed with high-concentration KCl as well. Glibenclamide had no impact on IL-1β secretion induced by NLRC4 inflammasome in both mice and red foxes. Interestingly, MCC950 did not affect mouse NLRC4 inflammasome activation, but it inhibited IL-1β secretion induced by flagellin in foxes. Thus, MCC950 inhibited NLRP3, AIM2, and NLRC4 inflammasomes in red foxes. Based on these results, we hypothesized that fox inflammasome activation shares a common NLRP3 inflammasome pathway with mice and humans, while having different intracellular signaling pathways for AIM2 and NLRC4 inflammasomes compared to the known pathways in humans and mice. MCC950 inhibits the NLRP3 inflammasome by directly targeting the NLRP3 NACHT domain and interfering with the Walker B motif function, preventing conformational changes and oligomerization of NLRP3 [19]. Previous studies have reported that MCC950 does not affect AIM2 and NLRC4 inflammasome activation [21]. Gene deficiencies in AIM2 and NLRC4 may be seen in various animals other than humans and mice [11,24]. Therefore, it appears that alternative proteins in red foxes compensate for the loss of AIM2 and NLRC4, the cytosolic sensors of dsDNA and flagellin, and collectively activate the NLRP3 inflammasome through a unique mechanism.
Genomic studies have been conducted to investigate the expression and function of the inflammasome components in various animal species, including red foxes [11,25]. It was found that the AIM2 gene is not present in red foxes [11,24]. AIM2 is a sensor protein that recognizes intracellular dsDNA, and AIM2-like receptors (i.e., myeloid cell nuclear differentiation antigen [MNDA], pyrin and HIN domain family member 1 [PYHIN1], and interferon-gamma-inducible protein 16 [IFI16]) with similar functions exist [25]. Therefore, the absence of AIM2 in red foxes does not imply the lack of functioning AIM2 inflammasomes. This study revealed that red foxes possess an AIM2-like receptor that can sense the presence of cytosolic dsDNA and activate the inflammasome, leading to the secretion of mature IL-1β. Similar results were obtained in a previous study on porcine AIM2 inflammasome [22]. Like red foxes, pigs also lack the AIM2 gene, but the AIM2 inflammasome can be activated through dsDNA transfection [22]. In porcine PBMCs, treatment with KCl and glibenclamide did not inhibit AIM2 inflammasome activation, but DPI did [22]. These results indicate that the regulation of the AIM2 inflammasome may vary among different animal species. In addition to AIM2, the NLRC4 and NLR family of apoptosis inhibitory proteins (NAIP) genes are also not expressed in red foxes [11]. The NLRC4 inflammasome is assembled upon the detection of intracellular flagellin by NAIP (e.g., Naip 5 and 6) [1]. Therefore, NLRC4 and NAIP are essential components for NLRC4 inflammasome activation. However, the current study successfully induced IL-1β secretion through the cellular uptake of flagellin in red foxes (Figure 3). Salmonella Typhimurium, a representative trigger for NLRC4 inflammasome [26], also successfully induced IL-1β secretion in fox PBMCs. Thus, there may be alternative sensors in the red fox cells that can substitute for NLRC4/NAIP. In addition, inhibitor studies showed that KCl and MCC950 inhibited fox NLRC4 inflammasomes (Figure 5), suggesting the involvement of a different form of inflammasome activation, rather than the canonical NLRC4 inflammasome. In particular, based on this study, it is anticipated that the NLRP3 inflammasome may play a common role in the activation of both AIM2 and NLRC4 inflammasomes in foxes, as evidenced by the inhibitory effects of the NLRP3 selective inhibitor, MCC950, on dsDNA- and flagellin-mediated IL-1β secretion in red foxes.
5. Conclusions
In this study, we investigated the activation of three representative inflammasomes (i.e., NLRP3, AIM2, and NLRC4) and their molecular regulatory mechanisms in red foxes. Red foxes, are members of the Canidae family and share a similar nucleotide sequence and amino acid sequence of pro-IL-1β with dogs. Through this study, we have revealed that the use of antibodies and primers derived from dogs can be applied to fox inflammasome research. Additionally, it has been observed that fox PBMCs secrete mature IL-1β (p17) in response to ATP, NG, and CaCl2, which are typical triggers of NLRP3 inflammasome activation [6]. Additionally, we found that fox PBMCs increased secretion of IL-1β (p17) upon the cytosolic introduction of dsDNA and flagellin, which are triggers for AIM2 and NLRC4 inflammasomes, respectively [17,26]. To induce the activation of inflammasomes, we used LPS, a well-known inducer of inflammasome priming [5]. Fox PBMCs were capable of completing the inflammasome priming process through the LPS/TLR4 signaling pathway, similar to other species [5,7,23]. Moreover, fox PBMCs increased IL-1β secretion in response to inflammasome-inducing bacteria [1]. Although the degree of IL-1β secretion by inflammasome activation differed between fox PBMCs and mouse BMDMs, the pattern of IL-1β secretion induced by known inflammasome triggers was similar to mice. This indicates that the three inflammasomes in red foxes may be presumed to function normally. To investigate the intracellular signaling pathways of fox inflammasomes, we used known inflammasome inhibitors [17,18,19,20]. The results revealed that the fox NLRP3 inflammasome was inhibited by the inhibitors similar to humans and mice, but there were differences observed for AIM2 and NLRC4 inflammasomes. Therefore, red foxes appear to have distinct intracellular signaling pathways for AIM2 and NLRC4 inflammasomes compared to the pathways known in humans and mice. These findings provide valuable information on the inflammasome biology in red foxes. Since red foxes share human habitats and serve as potential reservoirs for pathogens causing zoonotic infections, this information can be utilized for the treatment of inflammasome-mediated diseases and the control of zoonotic infections spread through red foxes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Comparison of the gene expression of inflammasome components and inflammatory cytokines between dog and fox PBMCs; Figure S2: The IL-1β secretion of fox inflammasome activation; Figure S3: Effect of bacteria on inflammasome activation in mouse BMDMs; Figure S4: Effect of Inhibitors on the fox inflammasome activation; Table S1: Primers; Data S1: The sequence comparison of pro-IL-1β mRNA between dogs (GenBank ID, NM_001037971) and red foxes (XM_025994662); Data S2: The sequence comparison of pro-IL-1β amino acids between dogs (GenBank ID, NM_001037971) and red foxes (XM_025994662).
Author Contributions
H.A., D.-H.J. and G.-S.L. Conceptualization; H.A., D.-H.J., S.-J.L., J.-J.Y., Y.-H.K., T.-W.H. and S.C. Methodology; H.A., S.-J.L, J.-J.Y. and G.L. Validation, Investigation; H.A. and G.-S.L. Funding acquisition; G.-S.L. Supervision, Writing—Original Draft; H.A., J.K., D.-H.J. and G.-S.L. Writing—Review & Editing.
Funding
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korean government (No. RS-2023-00208354, and RS-2023-00244078), and by a Korea Basic Science Institute (National Research Facilities and Equipment Center) grant funded by the Ministry of Education (No. 2022R1A6C101A739).
Institutional Review Board Statement
The animal study was conducted according to the National Institutes of Health Guide for the Care and Using of Laboratory Animals and approved by the Institutional Animal Care and Use Committee of Kangwon National University (approval no. KW-210331-2 for foxes, and KW-210504-1 for dogs).
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Man, S.M.; Kanneganti, T.D. Regulation of inflammasome activation. Immunological reviews 2015, 265, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.V.; Deng, M.; Ting, J.P. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nature reviews. Immunology 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; Lee, G.S. Korean Red Ginseng, a regulator of NLRP3 inflammasome, in the COVID-19 pandemic. Journal of ginseng research 2022, 46, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Karki, R.; Kanneganti, T.D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunological reviews 2017, 277, 61–75. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.M.; Coll, R.C. NLRP3 inflammasome priming: A riddle wrapped in a mystery inside an enigma. Journal of leukocyte biology 2020, 108, 937–952. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The calcium-sensing receptor regulates the NLRP3 inflammasome through Ca2+ and cAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Kim, J.; Lee, H.; Lee, E.; Lee, G.S. Characterization of equine inflammasomes and their regulation. Veterinary research communications 2020, 44, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Deets, K.A.; Vance, R.E. Inflammasomes and adaptive immune responses. Nature immunology 2021, 22, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.N.; Han, S.H.; Kim, B.H.; Kryukov, A.P.; Kim, S.; Lee, B.Y.; Kwak, M. Insights into Korean red fox (Vulpes vulpes) based on mitochondrial cytochrome b sequence variation in East Asia. Zoological science 2012, 29, 753–760. [Google Scholar] [CrossRef]
- Cho, H.K.; Shin, Y.J.; Shin, N.S.; Chae, J.S. Efficient distribution of oral vaccines examined by infrared triggered camera for advancing the control of raccoon dog rabies in South Korea. The Journal of veterinary medical science 2020, 82, 1685–1692. [Google Scholar] [CrossRef]
- Digby, Z.; Tourlomousis, P.; Rooney, J.; Boyle, J.P.; Bibo-Verdugo, B.; Pickering, R.J.; Webster, S.J.; Monie, T.P.; Hopkins, L.J.; Kayagaki, N.; et al. Evolutionary loss of inflammasomes in the Carnivora and implications for the carriage of zoonotic infections. Cell reports 2021, 36, 109614. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Chen, V.C.; Rozario, P.; Ng, W.L.; Kong, P.S.; Sia, W.R.; Kang, A.E.Z.; Su, Q.; Nguyen, L.H.; Zhu, F.; et al. Bat ASC2 suppresses inflammasomes and ameliorates inflammatory diseases. Cell 2023, 186, 2144–2159.e2122. [Google Scholar] [CrossRef]
- Goh, G.; Ahn, M.; Zhu, F.; Lee, L.B.; Luo, D.; Irving, A.T.; Wang, L.F. Complementary regulation of caspase-1 and IL-1beta reveals additional mechanisms of dampened inflammation in bats. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 28939–28949. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.; Aldridge, J.R., Jr.; Webster, R.G.; Magor, K.E. Association of RIG-I with innate immunity of ducks to influenza. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 5913–5918. [Google Scholar] [CrossRef] [PubMed]
- Poeck, H.; Bscheider, M.; Gross, O.; Finger, K.; Roth, S.; Rebsamen, M.; Hannesschlager, N.; Schlee, M.; Rothenfusser, S.; Barchet, W.; et al. Recognition of RNA virus by RIG-I results in activation of CARD9 and inflammasome signaling for interleukin 1 beta production. Nature immunology 2010, 11, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Lee, G.; Han, B.C.; Lee, S.H.; Lee, G.S. Maltol, a Natural Flavor Enhancer, Inhibits NLRP3 and Non-Canonical Inflammasome Activation. Antioxidants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Alnemri, T.; Yu, J.W.; Juliana, C.; Solorzano, L.; Kang, S.; Wu, J.; Datta, P.; McCormick, M.; Huang, L.; McDermott, E.; et al. The AIM2 inflammasome is critical for innate immunity to Francisella tularensis. Nature immunology 2010, 11, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Mueller, J.L.; Vitari, A.C.; Misaghi, S.; Fedorova, A.; Deshayes, K.; Lee, W.P.; Hoffman, H.M.; Dixit, V.M. Glyburide inhibits the Cryopyrin/Nalp3 inflammasome. The Journal of cell biology 2009, 187, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Abellan, A.; Angosto-Bazarra, D.; Martinez-Banaclocha, H.; de Torre-Minguela, C.; Ceron-Carrasco, J.P.; Perez-Sanchez, H.; Arostegui, J.I.; Pelegrin, P. MCC950 closes the active conformation of NLRP3 to an inactive state. Nature chemical biology 2019, 15, 560–564. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nature immunology 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Munoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nature medicine 2015, 21, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Kim, J.; Kwon, S.; Kim, P.H.; Kwon, H.M.; Lee, E.; Lee, G.S. Triggers of NLRC4 and AIM2 inflammasomes induce porcine IL-1beta secretion. Veterinary research communications 2018, 42, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ahn, H.; Woo, H.M.; Lee, E.; Lee, G.S. Characterization of porcine NLRP3 inflammasome activation and its upstream mechanism. Veterinary research communications 2014, 38, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Brunette, R.L.; Young, J.M.; Whitley, D.G.; Brodsky, I.E.; Malik, H.S.; Stetson, D.B. Extensive evolutionary and functional diversity among mammalian AIM2-like receptors. The Journal of experimental medicine 2012, 209, 1969–1983. [Google Scholar] [CrossRef] [PubMed]
- Dawson, H.D.; Smith, A.D.; Chen, C.; Urban, J.F., Jr. An in-depth comparison of the porcine, murine and human inflammasomes; lessons from the porcine genome and transcriptome. Veterinary microbiology 2017, 202, 2–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature 2011, 477, 596–600. [Google Scholar] [CrossRef]
Figure 1.
Comparison of inflammasome activation between dog and fox PBMCs. (A) Schematic diagram representing the priming and activation steps of inflammasomes. (B), LPS-primed PBMCs were treated with NG (40 μM) and ATP (5 mM) for 1 h, and the expression of pro-IL-1β in the cellular lysate (Lys) and the secretion of IL-1β (p17) in the cellular supernatant (Sup) were detected by immunoblotting. Lys: Cellular lysate and Sup: Cellular supernatant.
Figure 1.
Comparison of inflammasome activation between dog and fox PBMCs. (A) Schematic diagram representing the priming and activation steps of inflammasomes. (B), LPS-primed PBMCs were treated with NG (40 μM) and ATP (5 mM) for 1 h, and the expression of pro-IL-1β in the cellular lysate (Lys) and the secretion of IL-1β (p17) in the cellular supernatant (Sup) were detected by immunoblotting. Lys: Cellular lysate and Sup: Cellular supernatant.
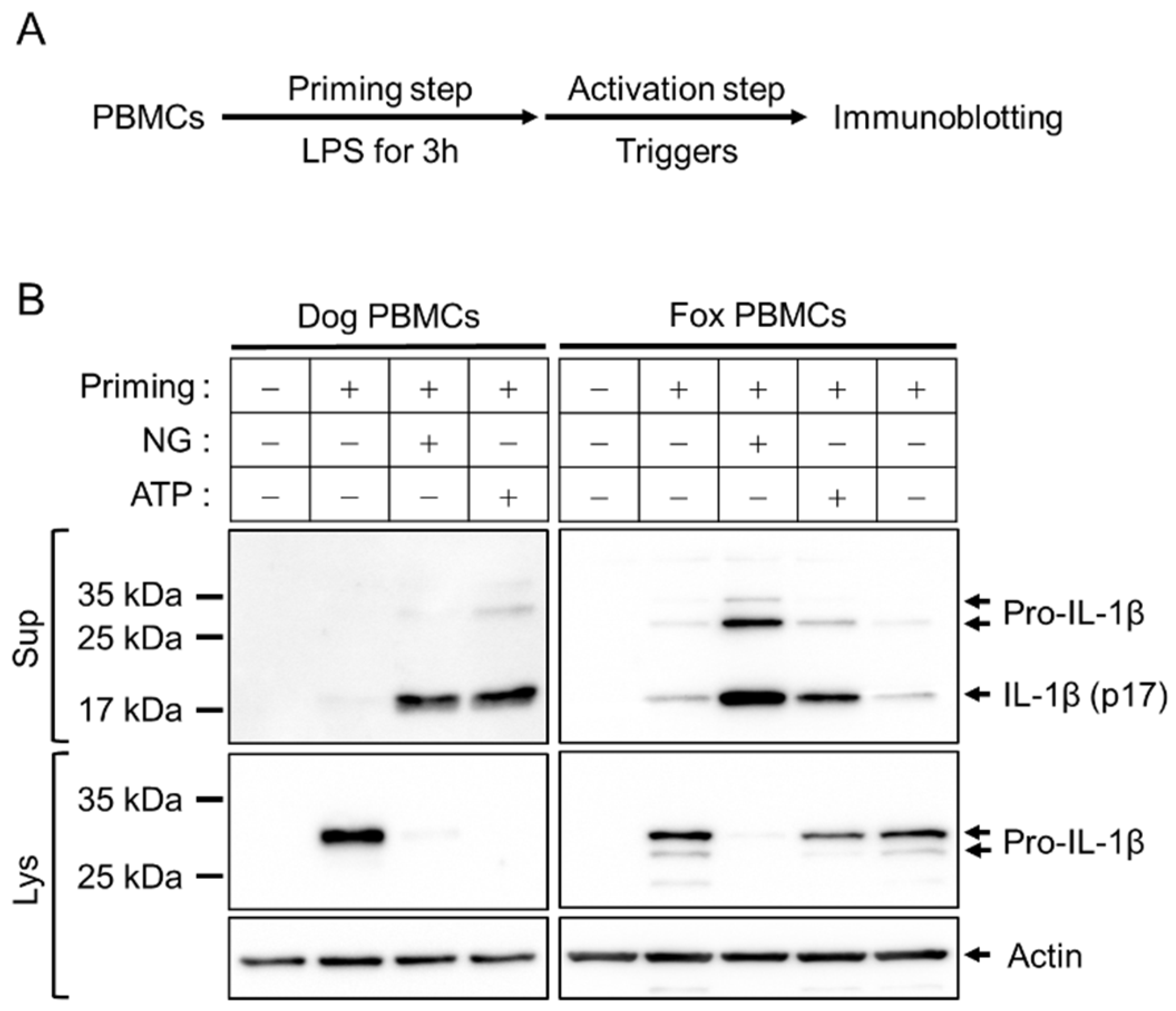
Figure 2.
Optimization of the NLRP3 inflammasome in red foxes. (A) Fox PBMCs were primed with LPS (1 to 1000 ng/mL), and further treated with ATP (5mM) or NG (40 μM). (B) Fox PBMCs primed with LPS (1 ng/mL) were treated with ATP, NG, and CaCl2 at indicated concentrations. The secreted IL-1β was analyzed by immunoblotting. Images are representative of two independent experiments. Lys: Cellular lysate and Sup: Cellular supernatant.
Figure 2.
Optimization of the NLRP3 inflammasome in red foxes. (A) Fox PBMCs were primed with LPS (1 to 1000 ng/mL), and further treated with ATP (5mM) or NG (40 μM). (B) Fox PBMCs primed with LPS (1 ng/mL) were treated with ATP, NG, and CaCl2 at indicated concentrations. The secreted IL-1β was analyzed by immunoblotting. Images are representative of two independent experiments. Lys: Cellular lysate and Sup: Cellular supernatant.
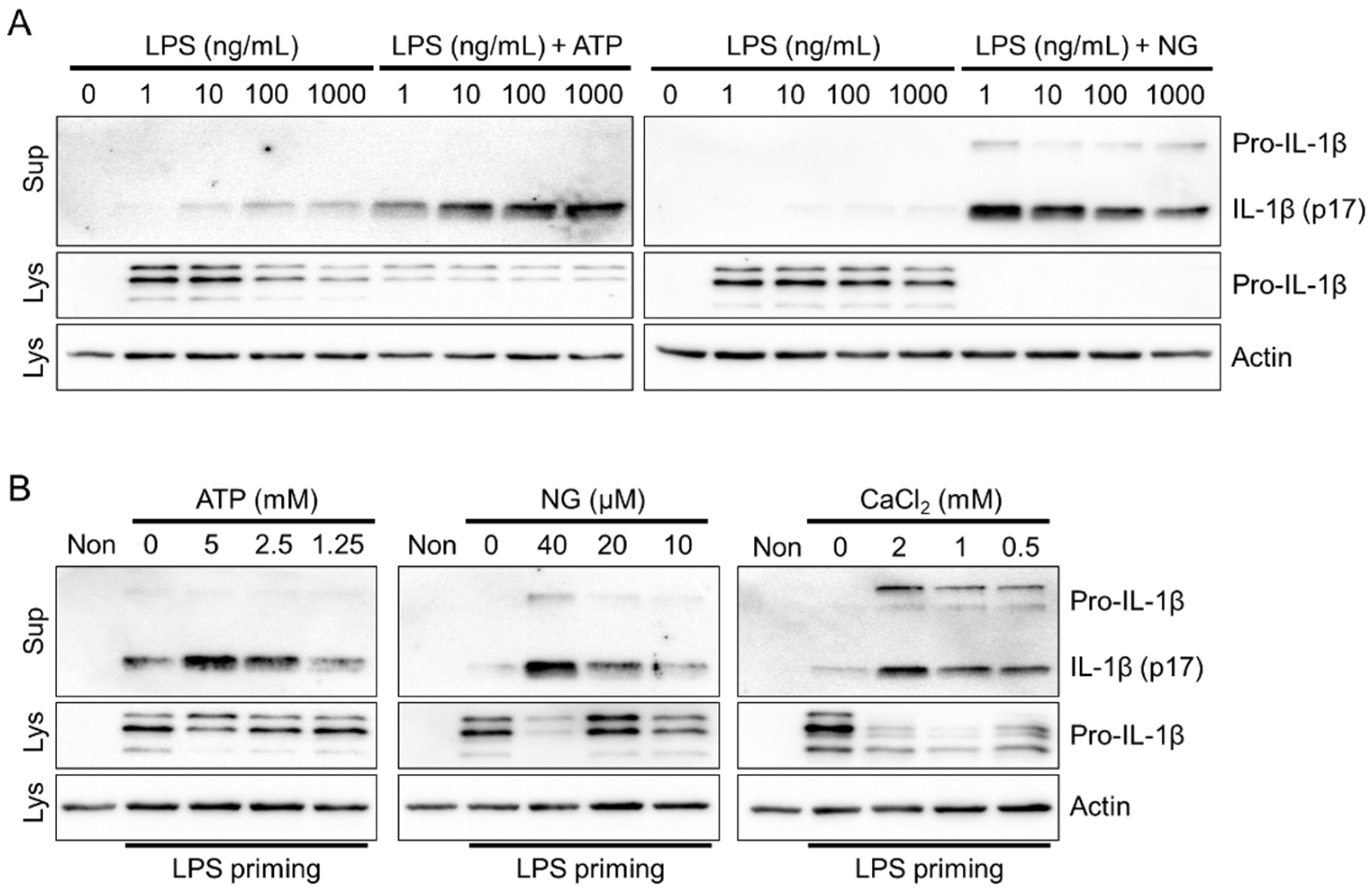
Figure 3.
Optimization of AIM2 and NLRC4 inflammasomes in red foxes. (A) Fox PBMCs were primed with LPS (1 to 1000 ng/mL), and further transfected with dsDNA (4 μg/mL). (B) LPS (1 ng/mL)-primed fox PBMCs were transfected with dsDNA at indicated concentrations. (C) Fox PBMCs were treated with LPS (1 to 1000 ng/mL), followed by the introduction of flagellin (500 ng/mL). (D) LPS-primed PBMCs of red foxes were treated with flagellin as presented. IL-1β secretion was measured using immunoblotting or ELISA. Images are representative of two independent experiments, and bar graphs present the mean ± standard deviation (SD) of at least three independent experiments. Lys: Cellular lysate and Sup: Cellular supernatant.
Figure 3.
Optimization of AIM2 and NLRC4 inflammasomes in red foxes. (A) Fox PBMCs were primed with LPS (1 to 1000 ng/mL), and further transfected with dsDNA (4 μg/mL). (B) LPS (1 ng/mL)-primed fox PBMCs were transfected with dsDNA at indicated concentrations. (C) Fox PBMCs were treated with LPS (1 to 1000 ng/mL), followed by the introduction of flagellin (500 ng/mL). (D) LPS-primed PBMCs of red foxes were treated with flagellin as presented. IL-1β secretion was measured using immunoblotting or ELISA. Images are representative of two independent experiments, and bar graphs present the mean ± standard deviation (SD) of at least three independent experiments. Lys: Cellular lysate and Sup: Cellular supernatant.
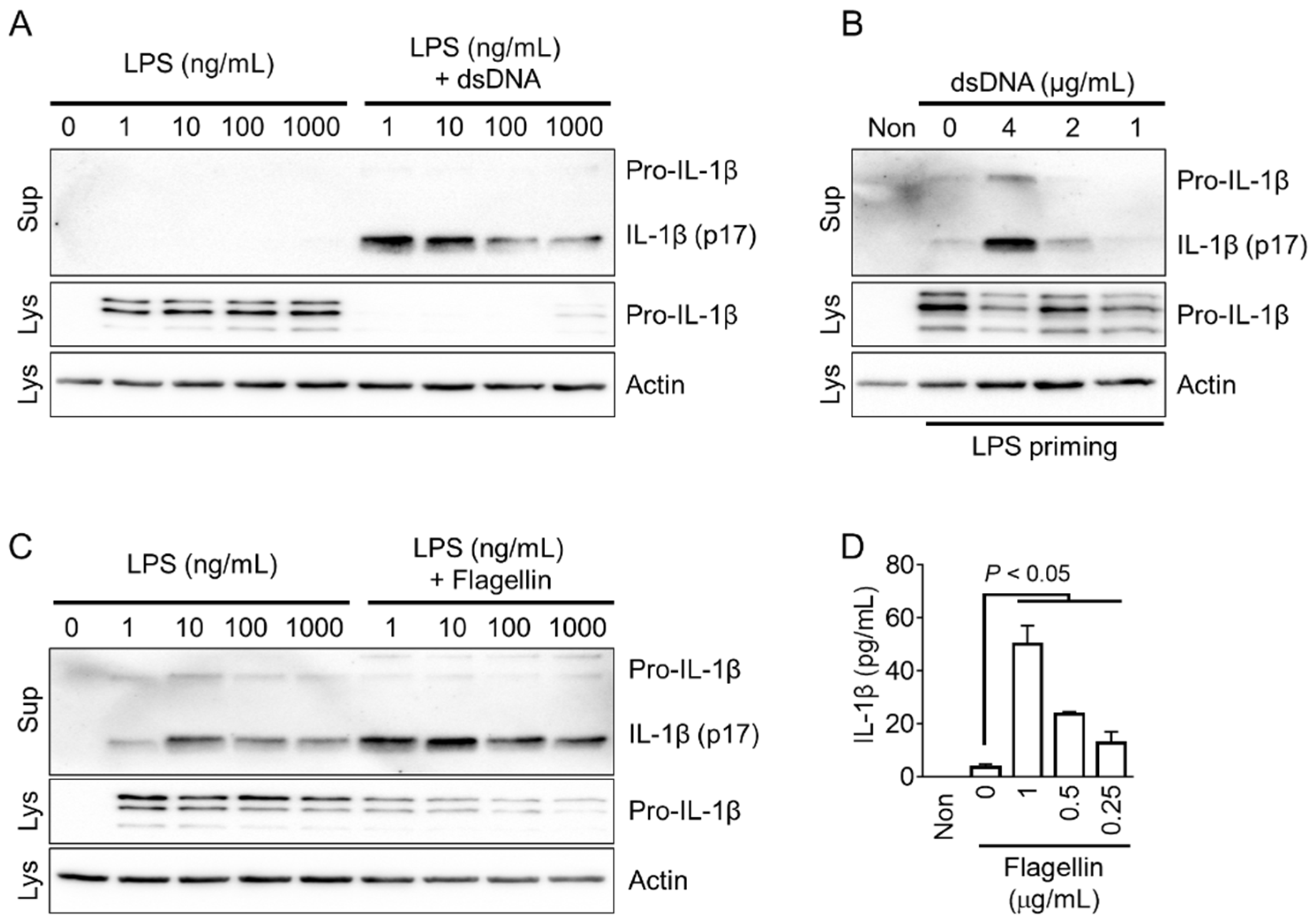
Figure 4.
Effect of bacteria on inflammasome activation in fox PBMCs. Fox PBMCs were primed with LPS (1 ng/mL) and then inoculated with bacteria at the indicated multiplicity of infection (MOI) for the presented times. The release of IL-1β was measured using ELISA. The bar graphs present the mean ± SD of at least three independent experiments.
Figure 4.
Effect of bacteria on inflammasome activation in fox PBMCs. Fox PBMCs were primed with LPS (1 ng/mL) and then inoculated with bacteria at the indicated multiplicity of infection (MOI) for the presented times. The release of IL-1β was measured using ELISA. The bar graphs present the mean ± SD of at least three independent experiments.
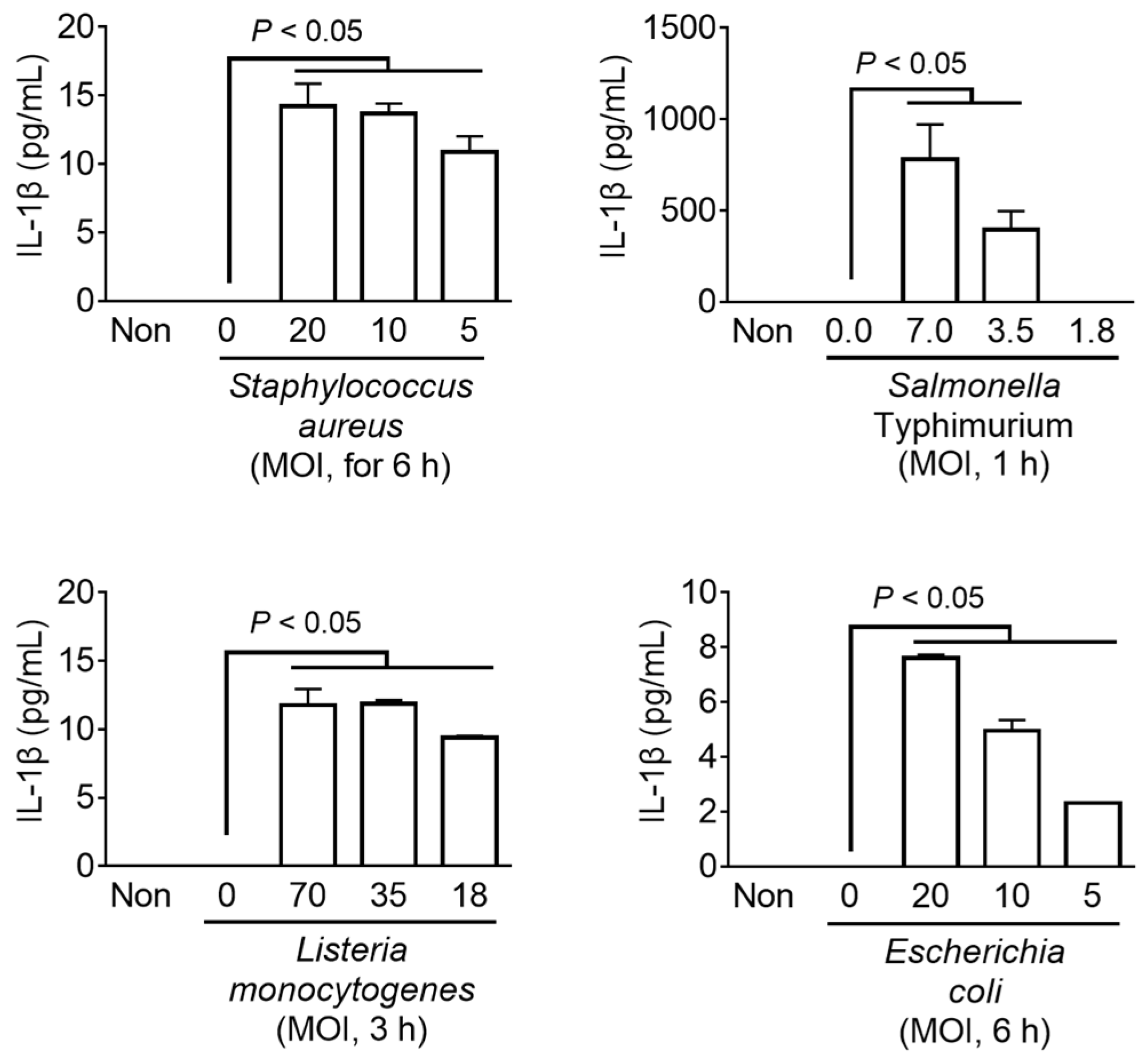
Figure 5.
Effect of inhibitors on the activation of fox and mouse inflammasomes. LPS-primed fox PBMCs (A) and mouse BMDMs (B) were treated with a ROS scavenger (diphenyleneiodonium [DPI]), a potassium efflux inhibitor (KCl and glibenclamide [Glib]), and an NLRP3 selective inhibitor (MCC950 [MCC]) in the presence of inflammasome triggers such as NG for NLRP3, dsDNA for AIM2, and flagellin for NLRC4. The secretion of IL-1β was analyzed by ELISA. Bar graphs present the mean ± SD of at least three independent experiments.
Figure 5.
Effect of inhibitors on the activation of fox and mouse inflammasomes. LPS-primed fox PBMCs (A) and mouse BMDMs (B) were treated with a ROS scavenger (diphenyleneiodonium [DPI]), a potassium efflux inhibitor (KCl and glibenclamide [Glib]), and an NLRP3 selective inhibitor (MCC950 [MCC]) in the presence of inflammasome triggers such as NG for NLRP3, dsDNA for AIM2, and flagellin for NLRC4. The secretion of IL-1β was analyzed by ELISA. Bar graphs present the mean ± SD of at least three independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



