
Submitted:
28 November 2023
Posted:
28 November 2023
You are already at the latest version
Abstract
Histone deacetylase 6 (HDAC6) regulates many physiological processes that are involved in cancer development and progression such as cell proliferation, apoptosis, motility, epithelial to mesenchymal transition, by deacetylation of multiple substrates and association with interacting proteins. Due to its the ability to remove misfolded proteins, induce autophagy, and regulate unfolded protein response, HDAC6 plays a protective role in responses to stress and enables tumor cells survival. HDAC6 is often overexpressed in tumors, including colorectal cancer (CRC) and is correlated with poor disease prognosis. However, it has been shown in animal models that HDAC6 is not essential for normal mammalian development. Therefore, it represents a good target for the development of novel cancer therapeutics. Selective inhibition of HDAC6 impairs growth and progression of many tumors without inducing major adverse events in experimental animals. In CRC, HDAC6 inhibitors have shown potential to reduce tumor progression and to enhance the therapeutic effect of other drugs. As HDAC6 is involved in regulation of immune responses, HDAC6 inhibitors have shown potential to improve antitumor immunity by increasing immunogenicity of tumor cells, augmenting immune cell activity and alleviating immunosuppression in tumor microenvironment. Therefore, HDAC6 inhibitors may represent promising candidates to improve the effect and to overcome resistance to immunotherapy.
Keywords:
HDAC6
; histone deacetylases inhibitors
; colorectal cancer
1. Introduction
Histone deacetylases (HDAC)s are originally characterized as nuclear enzymes that catalyze the removal of acetyl groups from the ϵ-amino groups of lysine (Lys) residues in histone proteins and thereby regulate the higher order chromatin structure and repress gene expression. Together with their functional counterparts, the histone acetyl transferases (HAT)s, HDACs play a key role in epigenetic regulation of gene expression. HDACs also deacetylate a variety of non-histone nuclear and cytoplasmic proteins and regulate their activity, stability, and localization. Based on their structure and enzymatic activity HDACs are divided into four major classes. The class II HDACs are known to shuttle between the nucleus and the cytoplasm. HDAC6 is a class II subclass b HDAC, mainly localized in the cytoplasm of differentiated cells [1,2].
HDAC6 has raised a lot of attention since its discovery in 1999 [3]. It consists of 1215 amino acids and possesses 5 functional domains (Figure 1). Starting from its N- to the C- terminus, HDAC6 comprises the following regions: nuclear localization sequence (NLS) which is rich in arginine (Arg) and Lys, nuclear export sequence (NES) that is rich in leucine (Leu), the two catalytic deacetylase domains (CD1 and CD2), the cytoplasmic anchoring serin (Ser) glutamin (Glu)-containing tetrapeptide (SE14) and a ubiquitin binding zinc finger motif domain (ZnF-UBP). NLS and NES together control the trafficking of HDAC6 between the nucleus and the cytoplasm, while SE14 is responsible for intracellular retention of HDAC6. HDAC6 also contains a dynein motor binding sequence (DMBS) between CD1 and CD2 catalytic domains [2-5].
With the two catalytic domains, HDAC6 is a unique class IIb HDAC, responsible for deacetylation of a number of non-histone substrates involved in regulation of crucial physiological processes, including cell proliferation, survival, apoptosis, autophagy, motility, intracellular transport and stress responses. HDAC6 regulates deacetylation of multiple cytoplasmic substrates and affects their activity, cellular location, and protein-protein interactions. The deacetylase activity of both catalytic domains is Zn2+ dependent. CD1 catalytic domain mostly deacetylates C-terminal acetyl-Lys residues and cannot independently exert catalytic activity but needs CD2 assistance. Aside from its deacetylase activity, HDAC6 through the catalytic CD1 domain exhibits E3 ubiquitin ligase activity [4]. Proteins in the cytoplasm that are identified as substrates for deacetylation by HDAC6 include α-tubulin, cortactin, heath shock protein (Hsp)90, heat shock transcription facto-1 (HSF-1), Ku70, p53, peroxiredoxins, signal transducer and activator of transcription (STAT)3, fork- head box protein O1 (FOXO1), and β-Catenin [2,6-9] (Table 1).
HDAC6 is partially localized in the nucleus and interacts with histone H4 [7] and non-histone nuclear proteins, including HDAC11 [10,11], the transcriptional corepressor LCoR [11], and transcription factors such as nuclear factor-kappa B (NF-κB) and Runx [2,12]. Aside from its deacetylase activity, HDAC6 exhibits its biological function by nonenzymatic interactions with other proteins (Table 2).
In this sense, via its ZnF-UBP domain, HDAC6 binds to ubiquitin as well as to sequence-specific proteins via its C-terminal glycine (Gly)-Gly motif that regulates cellular functions such as autophagic degradation, formation, and assembly of inflammasomes [13].
Catalytic activity of HDAC6 is affected by its subcellular localization and regulated by posttranslational mechanisms such as phosphorylation and acetylation that increase its activity. Phosphorylation of threonine (Thr), Ser or Tyr residues of HDAC6 by various kinases (including extracellular signal-regulated kinase (ERK), glycogen synthase kinase (GSK)3β, G protein-coupled receptor kinase (GRK)2, protein kinase C (PKC)) and growth factor receptors such as epidermal growth factor receptor (EGFR), augment its enzymatic activity [2, 14]. Furthermore, HDAC6 activity can be regulated by modifications of Lys residues, including sumoylation, ubiquitination, and acetylation. Although the biological consequence of sumoylation is not known, recent evidence shows that the ubiquitination may alter the activity of the target protein without inducing its degradation [15], while Lys acetylation attenuates enzymatic activity of HDAC6 [16,17]. Predominantly cytoplasmic localization of HDAC6 is influenced by NES and this process is dependent on the chromosome region maintenance protein 1 (CRM1), which is the main protein receptor that facilitates export of molecules from the nucleus to the cytoplasm [18,19]. Furthermore, the acetylation of NLS sequence of HDAC6 by histone acetyltransferase p300 contributes to retention of HDAC6 in cytoplasm as it blocks the interaction with the nuclear import protein importin-α [20].
HDAC6 is involved in a plethora of physiological processes that are often dysregulated in cancer.
2. Roles of HDAC6 in physiological processes
2.1. Cytoskeleton organization
HDAC6 affects cytoskeletal structure and dynamics by modifying microtubules and actin organization and thereby is involved in the maintenance of cellular shape, cell division, cell migration, intracellular transport, and angiogenesis. Cytoskeletal component, α-tubulin, is acetylated at Lys40 by α-tubulin acetyltransferase (αTAT) that enables tubulin polymerization and formation of microtubules, while its deacetylation by HDAC6 promotes microtubule disassembly [21,22]. The overexpression of HDAC6 is associated with tubulin deacetylation, proclivity to chemotactic movement and increased cell motility [23]. Furthermore, HDAC6 deacetylates another cytoskeletal protein cortactin that is present at areas of dynamic actin assembly such as at the leading edge of migrating cells. Deacetylated cortactin subsequently binds to F-actin through the small GTPase Rac1 and the actin nucleating complex Arp2/3, that enhances actin polymerization and branching leading to increased cell motility. Cortactin is often overexpressed in tumors [24], including colorectal cancer (CRC), where it promotes CRC cell proliferation by activating the EGFR-MAPK pathway. Its expression was reported to correlate with the metastatic potential of a tumor [25,26]. The acetylation of cortactin prevents its translocation to the cell periphery, blocks its association with F-actin and impairs cell motility, while its deacetylation by HDAC6 increases tumor cell motility and hence the invasiveness [20,25].
2.2. Degradation of damaged proteins
HDAC6 mediates the clearance of misfolded and damaged proteins that are often accumulated in malignant cells due to intense protein synthesis, gene mutations, incomplete translation in ribosomes or changes in cellular environment. Although the majority of defective proteins are labeled with ubiquitin chains and degraded by 26S proteasomes, if the proteasome system is overwhelmed or if protein aggregates are unsuitable for proteasomal degradation, they are degraded in autophagosomes. While the ubiquitin–proteasome (UP) pathway is mostly responsible for the degradation of short-lived proteins and soluble misfolded proteins, autophagy eliminates long-lived proteins, insoluble protein aggregates and organelles [27]. Cancer cells are characterized by extensive autophagic activity that modulates recycling of cellular proteins, regulates cellular homeostasis and energy metabolism, enables uncontrollable proliferation, and promotes survival of malignant cells.
Misfolded or aggregated proteins that cannot be degraded in proteasomes are transported along microtubules towards the microtubule-organizing center (MTOC) through motor protein dynein [28]. During this process, HDAC6 by its UBP domain, simultaneously interacts with polyubiquitinated proteins [29] while by its DMBS interacts with dynein motor complex, namely with its p150glued component [21] (Table 2). This protein complex is transported via the microtubule network to MOTC where HDAC6 mediates formation of aggresomes around misfolded protein aggregates by deacetylating cortactin. Subsequently, the deacetylated cortactin interacts with F-actin, triggering actin polymerization that is associated with fusion of aggresomes with autophagosomes and lysosomes to lyse proteins [30]. The ability of HDAC6 to mediate formation and removal of aggresomes is regulated by casein kinase (CK)2, which directly phosphorylates HDAC6 and increases its deacetylase activity [31].
HDAC6 coordinates UP system and autophagy maintaining them in complementary relationship that is essential to protect cells from stress imposed by accumulated damaged proteins. HDAC6 and the scaffolding protein sequestosome 1(SQSTM1)/P62, play an important role in coordinating the two pathways. P62 is a ubiquitin-recognizing receptor that binds ubiquitinated protein aggregates and encapsulates them into aggresomes where they fuse with lysosome and become degraded in a process designated as aggrephagy [32]. Furthermore, E3-ubiquitin ligase TRIM50 promotes the recruitment and aggregation of polyubiquitinated (poly-U) proteins into aggresome by facilitating the interactions of HDAC6 and p62 [33]. In this way HDAC6 improves the efficiency and selectivity of autophagic degradation [34,35]. While TRIM50 promotes the clearance of ubiquitinated proteins in aggresomes, an HDAC6-interacting chaperone, valosin-containing protein p97/(VCP) ATPase induces dissociation of HDAC6 and poly-U proteins and protein delivery to proteasomes. In this sense, the excess of p97/VCP favors protein degradation in proteasomes [36]. Recent studies have revealed the colocalization of HDAC6 and p62 with ubiquitinated mitochondria, as well as with the ubiquitinated protein aggregates associated with the E3 ubiquitin ligase TRIM50 [33]. Hence the balance of HDAC6 and p97/VCP levels determines whether the misfolded proteins and damaged mitochondria are going to be degraded by autophagy [36].
2.3. Autophagy
Aside from the formation of aggresomes, transportation and degradation of autophagosomes, HDAC6 regulates autophagy by deacetylating autophagy-related transcription factors and proteins [37]. In this sense, HDAC6 deacetylates transcription factor EB (TFEB) [38] and forkhead box O1 (FOXO1) [39] to decrease their activity and to inhibit autophagy. TFEB and FOXO transcription factors are inactivated by phosphorylation. In this sense, mTORC1 (mechanistic target of rapamycin complex 1) and ERK2 phosphorylate TFEB, while AKT (protein kinase B) phosphorylates FOXO1. Conversely, when unphosphorylated, TFEB and FOXO1 are translocated to the nucleus to initiate the transcription of target genes [40]. HDAC6 inhibition promotes the acetylation of TFEB, which then enhances the expression of autophagy-related protein Beclin-1 [41]. However, there are opposing data on the effect of HDAC6 on Beclin-1 expression obtained on liver cancer cells that showed that HDAC6 overexpression activated c-Jun NH2-terminal kinase (JNK) and increased the phosphorylation of c-Jun, which induced Beclin 1 dependent autophagy [43,44]. Acetylated FOXO1 transcription factor is also required for the T cell differentiation into regulatory T (Treg) cells expressing Foxp3 transcription factors [44]. It was reported that pharmacological inhibition of HDAC6 enhanced the transcriptional activity of acetylated FOXO1 and facilitated the autophagy process [39,43].
Furthermore, HDAC6 regulates the activity of an autophagy-related protein, the microtubule-associated protein 1 light chain 3 (LC3). LC3-I forms conjugate with phosphatidylethanolamine (PE) and becomes LC3-II which is then transported by HDAC6 to the MTOC to promote autophagosome formation [43,45]. It was reported that the deacetylation of LC3-II by HDAC6 promotes its translocation to cytoplasm and autophagy thereby inducing the survival of nutrient-deprived tumor cells [43,46]. This suggests that HDAC6 functions as a scaffold protein and deacetylase in regulation of LC3 that promotes autophagy [43]. Cytoskeletal modifications by HDAC6 are relevant for the progression of autophagy. While the actin-remodeling induced by cortactin deacetylation by HDAC6 is essential for the fusion of autophagosomes and lysosomes [30] the deacetylation of α-tubulin and microtubule disassembly inhibits their fusion with lysosome [47].
2.4. Regulation of molecular chaperons and other stress-related proteins
HDAC6 affects protein degradation in proteasomes by deacetylation of Hsp90 molecular chaperone whose primary physiological role is to protect proteins from degradation. Hsp90 that has been deacetylated by HDAC6 stabilizes protein tertiary structure, regulates transportation, and prevents protein degradation in proteasomes, thus allowing biological functions of its client protein. Multiple client proteins are regulated by Hsp90 such as steroid hormone receptors, growth factor receptors (EGFR, Vascular endothelial growth factor (VEGFR)), molecules engaged in the regulation of cell cycle and apoptotic pathway, transcription factors such as hypoxia-inducible factor 1-α (HIF-1α), signaling molecule c-Raf Ser/Thr-protein kinase, etc. Acetylation of Hsp90 impairs its chaperone activity and therefore induces degradation of the client protein. Simultaneously, HDAC6 is also a client protein of Hsp90 and the stability of HDAC6 is modulated by Hsp90 [48,49]. Moreover, degradation of HDAC6 is regulated by Cullin 3SPOP ubiquitin E3 ligase that has been shown to promote HDAC6 polyubiquitination and degradation in proteasomes in multiple cancer cell lines, including CRC [50].
HDAC6 regulates the function of Hsp90 also through the ZnF-UBP domain. HDAC6 senses ubiquitinated protein aggregates but also indirectly induces the expression of molecular chaperons, as it can activate p97/VCP that by its enzymatic activity induces the release of the HSF-1 from the Hsp90/HSF1 complex [29,36]. The released HSF1 induces the expression of genes for molecular chaperones Hsp70 and Hsp90 [51] which is followed by the release of HDAC6 from the complex and its binding to ubiquitinated proteins [27,48].
In CRC, the expression of Hsp90 in tumor tissue was inversely associated with survival outcomes and could represent a potential unfavorable prognostic factor for CRC patients [52]. As HDAC6 protects tumor cells from cytotoxic effects caused by defective and misfolded proteins, the inhibition of HDAC6 and Hsp90 may have therapeutic potential in cancer. In this sense, inhibition of Hsp90 has shown antitumor effect in animal models of CRC and in CRC cell lines as it caused the depletion of B-Raf and K-Ras oncoproteins that are major oncogenic drivers in CRC associated with poor disease prognosis [53].
HDAC6 via its deacetylase and ubiquitin binding activity is also involved in the regulation of mitochondrial metabolic activity. In this sense, the downregulation of HDAC6 causes impairment in the activity of mitochondrial enzymes, including respiratory complex II and citrate synthase. Hsp90 may be involved in this process as it is located in the inner space of mitochondrial membrane when deacetylated by HDAC6 [54].
HDAC6 plays an active role in response to environmental stress imposed by newly synthetized secretory proteins in the endoplasmic reticulum (ER) in order to eliminate misfolded proteins before protein aggregation becomes lethal for the cell [55]. In this sense, HDAC6 targets 78 kDa glucose-regulated protein (GRP78) and Hsp70, the molecular chaperones that are involved in unfolded protein response (UPR) that implies regulation of proper folding, conformational maturation, assembly of proteins in ER, and the control of the overall quality of proteins. Aside from its role in UPR, GRP78 displays antiapoptotic properties, promotes tumor proliferation, survival, and metastasis, and confers resistance to chemotherapy. The level of GRP78 was inversely associated with sensitivity of CRC cells to alkylating agents, including cisplatin and 5-flourouracil [56]. In solid tumors, hypoxic conditions, acidosis, and glucose deficiency induce GRP78 expression [57]. It has been shown that colon cancer cells secrete GRP78 via exosomes and that this process is dependent on the activity of HDAC6[58] which is often overexpressed in CRC [59]. HDAC6 inhibition increases GRP78 acetylation. Subsequently, the acetylated GRP78 dissociates from HDAC6 and then binds to VPS34, a class III phosphoinositol-3 kinase, thus preventing the sorting of GRP78 into multivesicular bodies and GRP78 release that induces its aggregation in the ER that further inhibits tumor growth [58]. Moreover, by inhibiting the release of exosomes containing GRP78 from cancer cells, HDAC6 inhibition also inhibits angiogenesis as GRP78 is involved in blood vessel formation in growing tumors through activation of HIF-1α and VEGF/VEGFR, as well as PI3K/AKT, ERK, FAK signaling pathways [60].
HDAC6 has an important role in redox regulation in response to cellular stress. HDAC6 deacetylates redox regulatory antioxidant enzymes peroxiredoxin (Prx) I and Prx II [61]. Prx are often present at high levels in cancer and neurodegenerative disorders and play a protective role against the oxidative damage. Acetylation of Prx increases its reductase activity, thus, HDAC6 and Prx may be considered as therapeutic targets for modulating intracellular redox status in cancer [5].
2.5. Apoptosis
HDAC6 affects cell cycle progression and apoptosis by modulating activity of multiple proteins, including p53. Upon DNA damage p53 is activated by kinases ATM, ATR, Chk1, and Chk2. This leads to the disruption of the interaction between p53 and Mouse double minute 2 homolog (MDM2) which is an E3 ubiquitin-protein ligase [62]. P53 can be acetylated by acetyltransferases CBP and p300 which prevent its ubiquitination, enhance its stability and transcriptional activity towards the expression of proapoptotic proteins Bax and Puma [65,66]. Moreover, acetylated p53 releases apoptotic molecule Bax from the nuclear p53/Bax complex which is then translocated to mitochondria to induce cytochrome C release and apoptosis. [63,64]. HDAC6 has been found to deacetylate p53 and repress its function as a tumor suppressor [65-67]. It was reported that HDAC6 inhibition increases acetylation of p53 in tumor cells, which leads to upregulated expression of genes related to cell cycle control and apoptosis, including p21 cyclin-dependent kinase (CDK) inhibitor, which can be induced with HDAC inhibition in p53-dependent and independent ways [66,68]. Moreover, acetylated cytoplasmic p53 inhibits autophagy by inducing Beclin-1 degradation via the ubiquitin-specific peptidase USP10 and by inhibiting AMPK mTOR pathway [67]. HDAC6 can also interact with p53 and attenuate its transcriptional activity through promotion of its degradation [66].
p53 is mutated in 43% of CRC [69] indicating that the induction of degradation of mutant p53 may represent a potential therapeutic approach. In this sense, several Hsp90 and HDAC inhibitors have been shown to destabilize p53 mutant proteins. Hsp90 inactivates the p53 E3 ubiquitin ligases, MDM2 and CHIP, thereby increasing mutant p53 levels, while Hsp90 chaperone activity is enhanced by HDAC6-mediated deacetylation [70,71]. Thus, targeting of Hsp90 or HDAC6 induces degradation of mutant p53. Whereas several Hsp90 inhibitors have so far been only investigated in clinical trials, HDAC inhibitors have already been approved by the FDA for the use in cancer therapy [66,69,72].
Due to increased demand for degradation in cancer, proteasomes participate in the maintenance of homeostasis in cancer cells. It has been reported that during the progression of CRC the level of ubiquitin-conjugating enzymes (E2) that are involved in various tumor-promoting processes and specifically the newly identified UBE2Q1, is elevated. UBE2Q1 suppresses the transcriptional activities of p53 by inducing its ubiquitination and degradation, and thereby may contribute to survival of tumor cells [73].
Due to frequent p53 mutations in CRC, the apoptotic effect of HDAC inhibition is not always p-53 dependent [74]. Recent studies indicate that the deacetylation of the DNA repair protein Ku70 by HDAC6 induces its binding to proapoptotic proteins Bax and Mcl-1 in cytoplasm to increase their stability and protect cells from apoptosis [75]. However, in vitro treatments of CRC cell lines with nonselective and selective HDAC6 inhibitors resulted in increased acetylation of Ku70 and induction of apoptosis by releasing Bax which was subsequently translocated to mitochondria and induced cytochrome release [9,76]. Interestingly, the dissociation of the complexes formed between Ku70 and antiapoptotic FLIP protein following the acetylation of Ku70 was found to trigger FLIP polyubiquitination and degradation in proteasomes in CRC cells [77].
Moreover, HDAC6 regulates apoptosis by deacetylation of antiapoptotic protein survivin. When acetylated by CBP, survivin is located in the nucleus and its function is inhibited. It was reported in breast cancer [78] and colon cancer cells [79] that deacetylation of survivin by HDAC6 promotes its exit from the nucleus and inhibits apoptosis.
Runt-related transcription factor-2 (Runx2), although initially defined as a transcription factor responsible for osteogenic differentiation in mammals, is closely related to proliferation, invasion, and bone metastasis of multiple cancer types. Interaction between Runx2 and HDAC6 leads to recruitment of HDAC6 from the cytoplasm to chromatin and repression of p21 gene promoter that induces proliferation [80]. In CRC cells with high Wnt signaling activity Runx2 was designated as a critical transcription factor to trigger the expression of genes that regulate epithelial to mesenchymal transition in vitro through orchestration of chromatin organization [81]. Moreover, clinical data showed that Runx2 was more abundant in CRC patients with metastasis and associated with shorter survival [82].
2.6. Regulation of signal transduction molecules
HDAC6 is also involved in the regulation of Ras-Raf-MEK-ERK and PI3K/AKT/mTOR signaling pathways. It was first discovered that ERK1 phosphorylates and activates HDAC6 [14]. Further investigations revealed that HDAC6 deacetylates ERK1/2 and augments its activity while acetylation by CREB-binding protein and p300 decreases its activity towards the transcription factor ELK1. Accordingly, HDAC6 inhibition has been shown to suppress tumor proliferation and induce apoptosis via deactivation of AKT and ERK signaling [83]. Moreover, HDAC6 affects AKT signaling indirectly, by deacetylating Hsp90 which subsequently binds to AKT and protects it from phosphatases and preserves its activity. By stimulating AKT signaling pathway HDAC6 contributes to cancer cell migration, and angiogenesis [84]. HDAC6 inhibition has been shown to decrease AKT binding to PIP3 and activity [85]. This is relevant for CRC as this tumor may show increased activation of this signaling pathway as well as increased expression of HDAC6 [86].
HDAC6 regulates EGFR endocytosis and degradation by controlling the acetylation status of α-tubulin and subsequently receptor trafficking along microtubules. A negative feedback loop consisting of EGFR-mediated phosphorylation of HDAC6 on Tyr570 reduces the deacetylase activity and increases acetylation of α-tubulin [87].
2.7. Regulation of NLRP3 inflammasome
HDAC6 is involved in the assembly, priming and activation of inflammasomes, the cytoplasmic protein complexes that are a crucial part of the innate immune system. The NLRP3 inflammasome contains NLRP3 pattern recognition receptor (PPR), apoptosis-associated spike-like protein containing caspase recruitment domain for caspase-1 (ASC), and caspase-1 itself. In response to activation of PPR by products of damaged or dying cancer cells and activation of NF-κB transcription factor, inflammasomes induce expression of NLRP3, pro-IL-1β, and pro-IL-18 [88]. This is followed by caspase-1 induced maturation of proactive inflammatory cytokines to IL-1β and IL-18 and subsequent cleaving of gasdermin D to induce pyroptosis [43]. HDAC6 facilitates the priming of NLRP3 inflammasome most prominently by deacetylating and activating p65 subunit of NF-κB that subsequently induces transcription of genes for NLRP3, pro-IL-1β and pro-IL-18. HDAC6 also promotes the activation of NF-κB via interaction with myeloid differentiation primary response protein 88 (Myd88) [43,89]. Moreover, HDAC6 induces the activation of inflammasomes by activating PrxII that increases the level of reactive oxygen species which are important activators of inflammasome [90]. It has been shown that HDAC6 inhibition upregulates p65 expression in the cytoplasm and reduces p65 expression in the nuclei of macrophages to attenuate the transcription of NLRP3 and reduce pyroptosis [91]. However, HDAC6 negatively regulates inflammasome activation through its interaction with ubiquitinated NLRP3 [43].
NLRP3 can impact CRC development due to its broad activity in shaping immune responses, apoptosis, and the gut microbiome. In this sense, role of inflammasomes in colitis and colitis-associated CRC have been shown in animal models [92,93]. Moreover, the activation of NLRP3 inflammasome in macrophages has been shown to promote the invasion of CRC cells by regulating epithelial-mesenchymal transition via secretion of IL-1β from activated macrophages [93,94]. This finding, together with a clinical finding that suggested the positive correlation of NLRP3 expression with advanced disease and poor prognosis in patients with CRC, indicates potential relevance of NLRP3 inflammasome as therapeutic target [95].
2.8. Role of HDAC6 in tumor invasiveness
HDAC6 is involved in multiple phases of epithelial-mesenchymal transition (EMT) by which tumor cells lose epithelial characteristics, cell to cell junctions, reorganize their cytoskeleton, increase cell motility, and acquire properties that are typical of mesenchymal cells. HDAC6 plays an important role in EMT by deacetylating α-tubulin and augmenting cell motility. Moreover, high level of acetylated α-tubulin was correlated with epithelial morphology while the deacetylated form corresponded to EMT transition [96].
The most potent inducer of EMT, transforming growth factor (TGF)-β, has the ability to activate HDAC6. TGF-β and HDAC6 pathways intercept and induce phosphorylation and activation of Smad3 molecule that inhibits the transcription of E-cadherin leading to mesenchymal-like phenotype of malignant cell [97]. It has been suggested that HDAC6 may induce activity of signal transducer Smad3 molecule directly by deacetylation [98], or indirectly by deacetylating α-tubulin and promoting Smad3 release [99,100]. Moreover, the selective HDAC6 inhibition was reported to downregulate the expression of TGF-βRⅠ, phosphorylation of Smad3, and EMT- inducing transcription factor Snail that led to preserved expression of E-cadherin in cultured cancer cells [96,101].
IL-6, has been reported to induce the expression of HDAC6, concomitantly with increased proliferation, migration, and EMT of tumor cells. Moreover, the IL-6-induced HDAC6 not only upregulated IL-6 downstream JAK2/STAT3 but also co-activated the TGF-β/Smad3 signaling [99]. The increased level of circulating IL-6 has been related to metastatic disease and poor prognostic outcome in cancer patients with diverse histological tumor types as it was reported in numerous studies [102-104].
Through deacetylation of cortactin and tubulin HDAC6 is involved in formation of invadopodia, the actin-rich proteolytic structures specialized in degradation of extracellular matrix that mediate invasion of malignant cells to distant tissues and organs [105]. Invadopodia have been observed in CRC [106]. Hypoxic conditions that are prevailing in solid tumors, enhance HDAC6 deacetylase activity by EGFR, resulting in enhanced Smad phosphorylation and nuclear accumulation that influences invadopodia formation [107,108]. Therefore, considering the role of HDAC6 in hypoxia-induced metastatic invasion to regional lymphatics, the therapeutic targeting of HDAC6 may have important therapeutic implications for the treatment of metastasis in cancer patients [108].
Furthermore, HDAC6 as an enzyme that regulates cytoskeletal dynamics, contributes to angiogenesis by regulating the polarization and migration of vascular endothelial cells in a microtubule end-binding protein (EBP)1-dependent manner and generation of capillary-like structures [109]. In support of this, the upregulation of HDAC6 mRNA level and protein level has been shown in endothelial cells under hypoxic conditions [110]. Furthermore, HDAC6 was found to associate with HIF-1α [111] and with its transcriptional target the VEGF receptor, and thereby increase their stability and activity in cancer cells [112]. Moreover, it was reported that HDAC6 affects Hsp90-mediated regulation of VEGFR in tumor cells [113].
HDAC6 modifies β-catenin that plays an essential role in cell-to-cell adherens junctions as it links E-cadherin to actin filaments. β-catenin is a key player in Wnt cascade signaling pathway that induces EMT, cancer cell motility, and cancer stem cell maintenance [114]. The activation of HDAC6 upon simulation of EGFR leads to deacetylation of β-catenin as well as the breakup of cell-to-cell junctions which increases the level of nuclear β-catenin, either by direct release of the junctional β-catenin from the cell membrane or by activating E-cadherin endocytosis. Nuclear localization of β-catenin increases proliferative potential of tumor cell by activation of target genes for c-myc and cyclin D1 [115]. It was reported that HDAC6 negatively regulates EGFR endocytosis and degradation in lysosomes by controlling the acetylation status of α-tubulin and hence the receptor trafficking along microtubules. However, the phosphorylation of HDAC6 by activated EGFR was found to reduce deacetylase activity and create a negative feedback loop, leading to increased degradation of activated EGFRs [87]. The Wnt/β-catenin signaling is involved in tumorogenesis of CRC. The presence of mutation in APC (adenomatous polyposis coli), induces β-catenin stabilization and nuclear localization and expression of Wnt target genes that promote tumor progression in CRC [115,116].
2.9. Involvement of HDAC6 in immune responses
HDAC6 has been shown to intervene in many aspects of the innate and adaptive immune responses. It affects antigen (Ag) uptake and presentation by antigen presenting cells (APC), dendritic cells (DC) or macrophages and cytotoxic function of natural killer (NK) cells, thus influencing the innate immune cells. Furthermore, HDAC6 partially affects T-cell activation and antitumor cytotoxicity in adaptive immune responses [117].
HDAC6 displays many of its immune-related effects by affecting STAT3 signaling pathway which is involved in the development of malignancies and in the induction and maintenance of immune tolerance and inhibition of immune responses [117,118]. STAT3 signaling can be activated with cytokines IL-6, IL-10, IL-21, TNF and growth factor GM-CSF that in DCs downregulate the expression of MHC class II and costimulatory molecules, thereby inducing the tolerogenic immune response [119]. Furthermore, STAT3 plays a key role in suppressing signal transduction mediated by Toll-like receptors (TLR)s in mature phagocytic cells. Accordingly, it has been shown that Stat3-deficient macrophages and DCs produce increased levels of proinflammatory cytokines (TNF, IL-1β, IL-6, IL-12) which activate immune response upon TLR4 activation, while reducing the amount of anti-inflammatory IL-10 and losing responsiveness to this cytokine which inhibits TLR4-dependent pro-inflammatory cytokine production [120].
HDAC6 is also involved in the regulation of macrophages [121]. In this sense, after stimulation of macrophages with lipopolysaccharide (LPS), HDAC6 was shown to translocate to the cell periphery where it induces cortactin deacetylation and the formation of inavadopodia protrusions that increase cell mobility and enable macrophage infiltration into tissue [122]. Under physiological conditions, macrophages activated by LPS, IFN-γ or TNF polarize into M1-type that secrete pro-inflammatory cytokines (TNF, IL-1β, IL-6, and IL-12), and activate the antitumor immune response. Conversely, in tumors, macrophages after exposure to immunosuppressive cytokines such as IL-10, and TGF-β in TME, often differentiate into M2-type that are known to produce high concentrations of the immunosuppressive cytokine IL-10. However, HDAC6 inhibition in macrophages and DCs results in diminished production of IL-10 and induction of inflammatory APCs that are able to effectively activate Ag-specific naive T cells. HDAC6 forms a molecular complex with STAT3 that was detected in cytoplasmic and nuclear compartments of the APCs [118].
It has been reported that colon cancer specimens with high HDAC6 expression show increased infiltration of immunosuppressive M2 macrophages that can be attributed to the HDAC6 activity [123]. In this setting, HDAC6 deacetylates TGF-β-activated kinase 1 (TAK1), (also known as MAPK kinase 7 (MAP3K7)), which subsequently activates p38 MAPK leading to phosphorylation and activation of A disintegrin and metalloproteinase-17 (ADAM17). ADAM17 by its proteolytic activity is responsible for shedding of IL-6 receptor [123] from the cell membrane, resulting in the release of soluble IL-6 receptor (sIL-6R). Aside from the classical IL-6 signaling that involves IL-6 ligation to membrane-bound IL-6R and gp130 transmembrane receptor dimerization, “IL-6 trans-signaling” is mediated by sIL-6R, which forms a complex with IL-6 and directly engages gp130 [99]. Since ADAM17 is more abundant in CRC cells compared to normal tissue, it contributes to increased levels of soluble IL-6R that promotes M2 macrophage polarization [123,124].
As HDAC6 is involved in intracellular trafficking of granules, the HDAC6-deficient CD8+ cytotoxic T lymphocytes (CTLs) were reported to display defective in vitro cytolytic activity due to altered dynamics, inhibited transport of lytic granules to the immune synapse, and deficient exocytosis, while target cell recognition, T cell receptor (TCR) activation or IFN-γ production were not inhibited [125].
HDAC6 affects the development and activity of regulatory T cells (Treg) that have a physiological function of suppressing excessive immune responses to maintain immune homeostasis. In tumor immunity, Tregs are involved in tumor development and progression as they impair T cell function through secretion of immune suppressive cytokines (IL-10, TGF-β, IL-35), consumption of IL-2 that leads to its depletion in TME, and expression of inhibitory checkpoint receptor CTLA-4. HDAC6 inhibits Treg differentiation as it deacetylates Foxp3 transcription factor and inhibits the transcription of Foxp3-induced genes. It has been reported in several murine models that treatment with HDAC6-specific inhibitors, increased activity, and induced differentiation of suppressive Tregs [126,127]. Inhibition of HDAC6 by tubastatin A increased the acetylation of Hsp90 in Tregs inducing the release of HSF-1 and upregulation of Treg- related genes [127]. Given that acetylation reduces the proteosomal degradation of FoxP3, HDAC6 inhibition would be expected to increase FoxP3 expression and increase the Treg number or function [128,129]. Regardless of these findings there are conflicting reports on the role of HDAC6 in Treg cell. In this sense, one study reported that pharmacological inhibition of HDAC6 inhibited Treg cell differentiation and suppressive function in TGF-β-induced murine Treg cell differentiation by inhibiting their proliferation [130]. It has not yet been elucidated why different studies reported different effects of HDAC6 inhibition on Tregs. Increased accumulation of Treg cells is generally associated with CRC progression and metastasis, immunotherapy failure, and a poorer prognosis, although this correlation is not conclusive [131].
HDAC6 activity in tumor cells and immune cells in tumor microenvironment (TME) has been shown to regulate the expression of tumor associated antigens, MHC class I molecules, costimulatory molecules, and cytokines [132].
3. HDAC6 inhibition in colorectal cancer
HDAC6 regulates signaling pathways which are involved in tumor cell growth, survival, and invasiveness and are often overexpressed in the majority of malignancies, including CRC. The increased expression of HDAC6 was reported in colon cancer tissue compared to the adjacent noncancerous tissue and is often associated with unfavorable disease prognosis [7,59]. The oncogenic potential of HDAC6 has been well established as its inactivation by genetic manipulations reduced oncogenic transformation and tumor growth in vitro as well as in in vivo models. More importantly, data obtained on HDAC6 null mice show that HDAC6 is not an essential gene for the development of an adult organism and that physiological functions of normal cells are not affected with deletion of HDAC6 gene [22,133]. Furthermore, it has been shown that unlike the other HDACs, selective inhibition of HDAC6 impaired tumor growth and progression without inducing major adverse events in experimental animals [22,61,110,134]. Altogether, these characteristics make HDAC6 a highly desirable target for cancer treatment. [135,136]. In experimental models of CRC, the HDAC6 gene knockdown and pharmacological HDAC6 inhibition reduced cell viability and migration of tumor cells by inhibiting the MAPK/ERK pathway [59,137]. It has been reported that patients with CRC show significantly lower expression of SET7 Lys methyltransferase in cancer tissue than in adjacent tissue. SET7 catalyzes the methylation of histone H3K4 that affects chromatin remodeling and regulates genes that are involved in cell cycle regulation, differentiation, DNA damage response and thereby plays a significant role in tumorigenesis. Moreover, SET7 functions as a tumor suppressor by inhibiting deacetylating activity of HDAC6, partially through ERK signaling pathway in colon cancer cells [137]. Downregulation of SET7 expression was closely correlated with poor prognosis in CRC [138] which is relevant for the oncogenic potential of HDAC6 in CRC. Furthermore, high expression of HDAC6 in CRC tissue was reported to be associated with reduced levels of acetylation at the 12th Lys residue of the histone H4 protein (H4K12ac). This histone residue was also reported to be highly sensitive to HDAC6 inhibition in several cancer types that subsequently induced chromatin relaxation [7].
HDAC inhibitors (HDACi)s can be classified according to their chemical structure and based on their ability to inhibit the specific HDAC isoform or distinct HDAC classes. In this sense, HDAC6 activity can be inhibited by unselective or pan-HDACis that inhibit the majority of HDAC classes, and selective inhibitors that target specifically HDAC6.
Pan-HDACis have been extensively investigated in preclinical and clinical studies and have shown therapeutic benefit in hematological malignancies (multiple myeloma (MM), cutaneous T-cell lymphoma, peripheral T-cell lymphoma) that led to their approval for therapeutic applications. In this sense, suberoylanilide hydroxamic acid (SAHA, vorinostat) was the first approved pan-HDACi by Food and Drug Administration (FDA) initially for the treatment of relapsed and refractory cutaneous T cell lymphoma [139-41]. Following the successful clinical results with vorinostat, pan-HDACis romidepsin and belinostat have been approved for the treatment of T cell lymphoma and adult leukemia [142,143]. Anther pan-HDACi panobinostat (LBH589, Farydak), was approved and used in the combination with proteasome inhibitor bortezomib and corticosteroid dexamethasone for the therapy of patients of with recurrent MM and showed some therapeutic benefit [144].
Regarding the antitumor effect of pan-HDACis in CRC, in vitro studies have shown that treatment with vorinostat induced cell death of CRC cell lines (Table 3) regardless of the p53 mutational status and led to downregulation of mutated p53 and upregulation of the wild type (wt) p53. Moreover, synergistic antiproliferative effect was shown when vorinostat was used in combination with the thymidylate synthase inhibitors 5-fluorouracil (5-FU) or raltitrexed, both commonly used in the treatment of CRC [145]. One study reported that vorinostat showed preferential cytotoxicity in cancer cells with mutated p53 by destabilizing mutant p53 through the inhibition of HDAC6-Hsp90 chaperone axis [71]. Also, vorinostat showed apoptotic and antiproliferative effect on CRC cell lines in vitro in combination with another pan-HDAC inhibitor trichostatin A. This treatment induced attenuation of Wnt signaling due to proteasome-dependent degradation of Wnt transcription factor TCF7L that was HDAC6- dependent [146].
Unlike clinical studies in hematological malignancies, in solid tumor the use of a single pan-HDACi has led to limited therapeutic success due to considerable adverse effects. This was most likely the consequence of pharmacological targeting of more than one of the eighteen HDAC isoforms that affects multiple survival-related cellular pathways [147]. Vorinostat was the first drug from the group of pan-HDACis to be used in clinical trial in combination with conventional chemotherapy, in combination with 5-FU in patients with metastatic CRC who previously failed to respond to 5-FU-based chemotherapy [148]. Results of a small phase I clinical study on the combination therapy with vorinostat and DNA methyl transferase inhibitor decitabine in patients with advanced CRC showed antitumor activity with prolonged disease stabilization [149].
Considering that HDAC6 is involved in multiple cancer-related pathways and that there was no severe toxicity with HDAC6 inhibition in animal models [134], some therapeutic advantages have been postulated for selective therapeutic targeting of HDAC6. In recent years, intensive investigations were conducted with the aim to develop small molecular modulators that selectively target HDAC6 and are effective in solid tumors as well as in hematological malignancies. In this sense, several drug candidates have been evaluated in the clinic. HDAC6is tubacin and tubastatin A were the first HDAC6is used in experimental settings [135]. Unfortunately, as they possess nondrug-like properties such as high lipophilicity, unfavorable pharmacokinetics, and rapid metabolic inactivation, they are only used in research as tools to validate HDAC6 as a target, but not in the clinical settings [135,136,150].
Ricolinostat (ACY-1215) is the first effective selective HDAC6i that became orally available, and it was shown to be ten times more selective against HDAC6 than other HDACs [151,152]. Since its first application in MM [151], ACY-1215 has shown some efficacy in various tumors [153]. By increasing the level of acetylated tubulin, cortactin, and Hsp90, ACY-1215 inhibits cell cycle progression, motility and invasiveness, processing of misfolded proteins, ubiquitin proteasome pathway, and autophagy. Furthermore, it renders GRP78 in acetylated form thus preventing it from transporting misfolded proteins from ER lumen, hence increasing ER stress. This effect, together with increased accumulation of protein aggregates, polyubiquitinated and misfolded proteins, induced tumor cell death by apoptosis [154]. Furthermore, ACY-1215 exerts its antitumor effect by inhibition of proliferation and induction of apoptosis by inhibiting MAPK/ERK, PI3K/AKT cell signaling pathways in CRC [59,155].
Combinations of selective HDAC6is with different alkylating agents have shown synergistic antitumor effects in various tumors including CRC. ACY-1215 showed antiproliferative effect on CRC cell lines in combination with 5-FU [156]. Also, in combination with oxaliplatin, ACY-1215 promotes cell apoptosis via activation of caspase-3, elevation of the Bak to Bcl-xL ratio, and downregulation of p-ERK and p-AKT in CRC cell lines [155]. Collectively, these findings suggest that targeting HDAC6 activity using ACY-1215 may present a promising therapeutic opportunity.
Ryu investigated the anticancer mechanisms of a novel potent and selective HDAC6i, A452, and compared its effect on CRC cell lines with the clinically tested HDAC6i ACY-1215. In the aforementioned study, A452 showed antiproliferative effect irrespective of p53 status and induced apoptosis by activating caspase-3 and enzyme involved in DNA repair poly ADP-ribose polymerase (PARP), increasing Bak and Bax while decreasing Bcl-xL levels. It also inhibited AKT and ERK pathways and triggered DNA damage by increasing the activation of the checkpoint kinase Chk2 that induced cell cycle arrest and apoptosis [157]. Further in vitro studies indicated that A452 combined with vorinostat was more effective in induction of apoptosis than either drug alone [158]. Furthermore, some natural products such as Aceroside VIII, which is a diarylheptanoid isolated from plant Betula platyphylla, were reported to enhance the anticancer activity of HDAC inhibitor A452 in CRC cell lines [159].
Another selective HDAC6i, citarinostat (ACY-241), is under clinical development for the treatment of relapsed and refractory MM [160] and more recently solid tumors. ACY-241 showed improved antitumor activity in combination with paclitaxel in mouse xenograft model of ovarian and pancreatic carcinoma [161] that was later confirmed in a clinical trial conducted in patients with previously treated advanced solid tumors [162].
Nexturastat A (Nex A) is a selective inhibitor of HDAC6, that was first investigated in melanoma and showed antitumor activity in vitro and in vivo experimental models [132]. This next-generation HDAC6i is unique compared to other selective HDAC6is as it mainly affects immune-related traits and functions in tumors and immune cells with minimal cytotoxic effects [121]. Furthermore, Nex A exerted its antitumor effect mostly by inducing changes in TME, including decreased infiltration of immunosuppressive cells and upregulation of adhesion molecules such as E-cadherin, thus inhibiting EMT process as shown in a breast cancer model [163].
MPT0G612 is a novel HDAC6i that has been investigated in experimental in vitro settings and exhibited promising antitumor activity against several solid tumors, including CRC. Antiproliferative and proapoptotic effects of this compound were accompanied with activation of autophagy mediated with LC3B-II formation and p62 degradation [164].
3.1. HDAC6 inhibition in combination with other therapeutic modalities
In order to explore the options for the most optimal antitumor activity while minimizing the side effects of the maximally tolerated doses, the selective HDAC6is have been further evaluated in combination with different anticancer agents such as proteasome inhibitors, tyrosine kinase inhibitors, radiotherapy, and immunotherapy.
The afore mentioned ACY-1215 has been reported to act more effectively in combination with the protease inhibitor bortezomid which is currently used in the treatment of MM and Hodgkin and non-Hodgkin lymphoma [151]. In CRC, ACY1215 showed improved antitumor effect in combination with carfilzomib (proteasome inhibitor) by inducing accumulation of protein aggregates, ER stress, and subsequently apoptosis of treated CRC cell lines harboring BRAFV600E mutation in in vitro and in xenograft murine models [165]. Another selective HDAC6 inhibitor C1A also showed potential antitumor activity against KRAS-mutated CRC in murine xenograft models [150] in combination with bortezomib that was characterized with inhibited degradation of misfolded proteins in proteasomes and decreased autophagy [166].
BRAFV600E mutations that lead to constitutive activation of BRAF kinase and increased RAS/RAF/MEK/ERK signaling have been reported in 10% of patients with CRC [167]. However, in CRC harboring BRAFV600E mutations, response rates to BRAF inhibitors vemurafenib or dabrafenib and MEK1/2 inhibition with trametinib were low due to adaptive feedback reactivation of upstream RTKs and RAS. In these tumors, BRAF inhibition enhances RAF dimerization and thus results in adaptive feedback re-activation of MAPK signaling, often mediated by EGFR activation [168]. HDAC inhibitors have been tested as potential agents to reduce the resistance to BRAF/MEK inhibition in CRC. Accordingly, one study in mouse xenograft CRC model and cell lines showed the ability of vorinostat to overcome resistance to treatment with MEK1/2 inhibitor trametinib. The study also identified a novel resistance mechanism that is mediated via STAT3 and the anti-apoptotic protein c-FLIPL [169]. Increasing experimental evidence shows enhanced antitumor effects of simultaneous application of HDACis and RAF/MEK/ERK targeting therapeutic agents, in other tumors with BRAF mutation. Synergistic antitumor effect of vorinostat in combination with PLX4720 BRAF inhibitor [170] and the ability of vorinostat to eliminate BRAF inhibitor resistant and senescencent cells was reported in BRAF mutated melanoma cells [171]. Similarly, panobinostat in combination with MAPK and BRAF inhibitor dabrafenib has shown synergistic antitumor effect in BRAF mutated thyroid carcinoma cells [172].
As HDAC inhibition often induces cell cycle arrest and inhibits DNA repair, the ability of HDAC inhibitors to improve the therapeutic outcome of radiotherapy and to sensitize the tumor cells to ionizing radiation has been investigated in experimental settings [173]. In this sense pan-HDAC inhibitors panobinostat and vorinostat have shown enhanced antitumor effect when used in combination with radiation therapy in clinical trials in prostate and gastrointestinal tract carcinoma [174]. Furthermore, one study reported that treatment with the selective HDAC6i SP-2-225 resulted in decreased tumor growth and increased infiltration of M1 macrophages within tumors. These finding support further investigation of the use of selective HDAC6is to improve antitumor immune responses and prevent post-radiation therapy tumor relapse [175].
3.2. Modulation of antitumor immunity with HDAC6 inhibition
HDAC inhibitors have shown considerable immunomodulatory effects by influencing many aspects of the immune response in tumors. However, some differences were reported depending on weather pan-HDAC or selective HDAC6 inhibitors were used and on the experimental model used in a study (Table 4) [117].
Pan-HDAC inhibitors have been reported to decrease the expression of costimulatory molecules (CD40, CD80, CD83 and CD86) on DCs and secretion of Th1 polarizing cytokines (IL-6, IL-12, TNF) after stimulation of TLR on DCs that led to inhibition of T cell activity. Panobinostat reduced the expression of costimulatory molecules on DCs and impaired IFN-γ production by T cells [176]. Similarly, pan-HDACi vorinostat was reported to inhibit T cell functions by inducing the transcription of enzyme indoleamine 2,3-dioxygenase (IDO) in DCs which regulates the catabolism of tryptophan which is essential for T cell activation [177]. HDAC inhibition by vorinostat was shown to promote the transcription of IDO through acetylation and activation of STAT3 and hence to inhibit T cell functions [178]. Contrary to pan-HDACis, selective HDAC6 inhibitors have shown immunostimulatory effects on DCs. It has been reported that ACY241 increased the expression of costimulatory and MHC molecules on DCs [179]. Furthermore, it was shown that tubastatin A impairs the production of immunosuppressive cytokine IL-10 by DCs and macrophages by disrupting the complex between HDAC6 and STAT3 and by impairing STAT3 signaling that in turn increased the production of IFN-γ by CD4 T cells [118,163]. All these factors indicate that HDACis by affecting APCs may regulate innate and adaptive immune response and inflammation in TME. Moreover, tubastatin A showed antiinflammatory effect by inhibiting IL-6 synthesis, nitric oxide (NO) secretion, cell viability and motility in human macrophages stimulated with LPS [180]. This effect is also relevant for antitumor effect of tubastatin A, as IL-6 represents a potent EMT triggering factor in the TME involved in tumor progression, metastatic invasion, and chemoresistance [99].
The effects of pan-HDACis on T cell activation differ from the effects of specific HDAC6is. Pan-HDACis trichostatin A and rodempsin inhibit activation-induced proliferation of naïve T cells and IL-2 production [181], unfavorably affect metabolic reprograming of recently activated T cells, impair T cell receptor signaling, and induce T cell apoptosis [117,182,183]. However, reports on the impact of pan-HDACis on previously activated effector T cells show that when applied after initial activation of CD4 T cells, pan-HDACi trichostatin A prevents FasL-driven activation-induced cell death, increases infiltration of CD4 T cells into tumor and reduces tumor growth in lymphoma and melanoma murine models [117,184]. Regarding selective HDAC6is, Laino et al. found that peripheral blood T cells of melanoma patients in vitro treated with selective HDAC6is ACY-1215 and ACY-241 showed decreased production of immunosuppressive Th2 cytokines (IL-4, IL-5, IL-6, IL-10 and IL-13) with concomitant downregulation of the Th2 transcription factor GATA3, upregulation of the Th1 transcription factor T-BET, and favorized accumulation of central memory phenotype T cells. This report indicated the immunostimulating potential of selective HDAC6is on T cell cells [185].
CD8 T cells directly kill tumor cells by secretion of perforin and granzymes and facilitating antitumor immune responses by production of IFN-γ and TNF that activate local APCs and increase immunogenicity of tumor cells by inducing MHC expression, and activation of immunoproteasomes. It was reported that treatment with pan-HDACi vorinostat increased the proliferation and function of CD8 T cells, particularly the frequency of IFN-γ or perforin-producing T cells in mammary tumor bearing mice [186]. Contrary to pan-HDAC inhibitor vorinostat, selective inhibition of HDAC6 may impair the cytotoxic capacity of CD8 T cells, as tumor-specific CD8 T cells from mice treated with HDAC6-specific inhibitor tubastatin A and the HDAC6 deficient mice showed reduced lytic capacity of CD8 T cells, probably due to interrupted intracellular trafficking and exocytosis of perforin [125]. However, one study reported that another HDAC6 selective inhibitor, ACY241 increased perforin and IFN-γ production in CD8 T cells [179]. Evidently, due to the conflicting evidence on the effect of different selective HDAC6is on T cell activity, further investigations are needed [187].
Treatment with selective HDAC6is has been shown to affect the susceptibility of tumor cells to T and NK cell mediated killing, as they affect the expression of MHC class I molecules on tumor cells. In this sense, HADC6 inhibitor tubastatin A has been reported to induce expression of MHC class I molecules on melanoma cells and increase their susceptibility to CD8 T cell mediated-lysis [132]. Moreover, the treatment of colon cancer cells with pan-HDACi vorinostat increased the expression of death receptor Fas that led to enhanced Fas-dependent cytotoxicity activity of T cells [149]. Furthermore, it has been reported that treatment with pan-HDACi panobinostat increased the expression of genes involved in cell adhesion and junction and formation of conjugates between NK and tumor cells, and modulated the expression of NK cell-activating receptors and ligands on tumor cells, thus contributing to the increased cytolysis of tumor [188]. Regarding NK cell antitumor activity, it was reported that pan-HDACis vorinostat and trichostatin A induced the expression of MHC class I-related chain A (MICA) and B ligands for activating NK cell receptor NKG2D in hepatocellular carcinoma and Ewing sarcoma and thereby increased the susceptibility of treated tumor cells to NK cell-mediated lysis [189,190]. Furthermore, it was reported that HDAC6i Nex A increased tumor infiltration with NK cells, an effect that has been associated with improved prognosis and survival in tumors [121,187].
Moreover, some HDACis enhance T cell migration to tumor site. Pan-HDACi rodempsin was reported to increase the expression of chemokines CCL5, CXCL9, 10 by tumor and stromal cells, increase tumor infiltration with T cells and thereby improve antitumor immune response [191].
Selective HDAC6 inhibition contributes to antitumor immunity by inhibiting differentiation and influx of suppressive immune cells into TME. Ricolinostat was reported to inhibit the activity of myeloid-derived suppressor cells (MDSC) and inhibit tumor growth [192] while Nex A treatment has been shown to induce differentiation of immunostimulating M1 macrophages [121] similarly to the pan-HDACi trichostatin A [193].
In summary, the HDAC6is have shown potential inherent to immunomodulatory agents as they have the ability to improve antitumor immunity by stimulating the immunogenicity of tumors and activity of immune cells.
The expression of immune checkpoint (IC) molecules is often upregulated in immune cells in TME as a consequence of antitumor immune responses and due to elevated levels of immunosuppressive mediators produced by tumor cells, suppressive immune cells, and stromal cells [194]. In this sense in TME, T and NK cells may express programed cell death receptor (PD)-1, cytotoxic T lymphocyte antigen (CTL)-4, T cell immunoglobulin and mucin domain-containing protein 3 (TIM3), lymphocyte activation gene-3 (LAG-3), and TIGIT, that inhibit antitumor immune responses [104,195]. On the other hand, tumor cells often express ligands for ICs that can be induced with oncogenic pathways and extrinsic factors in TME such as cytokines IFN-γ, IL-6, TNF-α [196,197] that contribute to upregulation of PD-1 ligands (L)1 (B7H1), and L2 (PD-L2, B7DC) on tumor cells. In CRC, MAPK, PI3K, JAK/STAT3, and phospholipase Cγ signaling pathways have been related to the upregulation of PD-L1 expression [198].
It has been well established that metastases of CRC show increased expression of PD-L1 compared to primary tumors [199] that is associated with unfavorable disease prognosis [200]. Moreover, the expression of PD-L2 in tumor cells, which is more inherent to immune cells, was recently associated with poor patient survival in CRC [200,201]. Therapeutic blockade of PD-1 and its ligand with anti-PD-1 and anti-PD-L1 antibodies has shown a considerable clinical benefit in some metastatic CRC [199,202]. However, blockade of PD-1 enhances T cell function and the subsequent production of inflammatory cytokines, most notably IFN-γ that enhances PD-L1 and PD-L2 expression on tumor cells thereby inducing negative feedback as well as other immunosuppressive pathways [194,196,203].
As tumors during their evolution develop mechanisms allowing them to evade immune responses, it is of importance to identify treatments that can increase immunogenicity, minimize immune-related adverse events, and maximize therapeutic benefits of IC inhibition [104]. Among the emerging new therapeutic targets HDACs have raised great interest, especially HDAC6 since it is involved in the control of immunomodulatory pathways and expression of IC molecules. It has been shown recently that HDAC6 may induce the expression of PD-L1 in cancer cells via activation and recruitment of STAT3 transcription factor, as it was shown in the experimental models of pharmacological impairment of HDAC6 or by its genetic abrogation in melanoma [204] and breast cancer [163,205]. However, there are differences in existing published data between the effects of some selective pharmacological HDAC6 inhibitors on PD-L1 expression. It was reported that in vitro treatment with Nex A decreased the expression of PD-L1 in melanoma [121] and breast [163] animal tumor models. Similar effect was reported for the in vitro treatment with the novel selective HDAC6i MPTOG612 in CRC cells [164]. However, some publications report the opposing effect for certain selective HDAC6is. One study reported that the treatment with novel small molecule HDAC6i A452 and ACY-1215 increased PD-L1 expression in CRC tumor cells [157]. Another study reported that ACY-1215 alone and in combination with alkylating agents upregulated PD-L1 in CRC cells in vitro and that was achieved irrespective of p-STAT3 status [155]. However, ACY-1215 and A452 induced PD-L1 expression may increase susceptibility of tumor cells to PD-1/PD-L1 axis IC blockade therapy.
The use of nonspecific HDACi, such as panobinostat [206] as well as low doses of trichostatin A [193] was reported to increase the expression of PD-L1 and PD-L2 on cell surface of tumor cells in murine models of melanoma and breast cancer. Moreover, panobinostat was shown to synergize with PD-L1 blockade by different mechanisms such as promotion of NK cell-target cell conjugation formation by increasing the expression of cell adhesion and tight junction-related genes and by increasing the expression of CD80, CD86 (ligands for CD28), and CD112 (PVRL2/nectin-2 ligand for activating DNAM-1 NK cell receptor) on tumor cells [188]. These results also indicate the antitumor potential of HDAC inhibition in the context of NK cell-based immunotherapy. It was also reported that panobinostat augmented the expression of MHC I and costimulatory molecules (CD40, CD80) on melanoma cells in vitro that led to increased activation of antigen-specific T cells [207].
Aside from the immune cells, PD-1 is also expressed on malignant cells including colon cancer [208]. Moreover, tumor cells expressing PD-1 exhibit higher ability of proliferation and tumorigenicity [209]. Recently it has been reported that the transcription of PD-1 gene in cancer cells is regulated via acetylation of p53 tumor suppressor by HATs p300, CBP and Tip60 in a manner that acetylated p53 recruits the acetyltransferase cofactors to interact with PD-1 promotor and induce the expression of PD-1 [210]. Although the validation on larger sample size is needed, the expression of PD-1 on tumor cells versus its expression on immune cells may have some relevance for IC blockade selection in patients with CRC [208]. However, there are scarce studies on the effect of HDACis on PD-1 compared to PD-L1 in tumor cells [208].
Selective HDAC6 inhibition also affects IC expression on immune cells. In this sense, one study reported that ACY-1215 and ACY-241 decreased the expression of LAG-3, TIM-3, and PD-1 on peripheral blood T cells of melanoma patients. These findings indicate that inhibition of HDAC6 activity may be effective in alleviating T cell suppression and enhancement of the cytotoxic function of T cells and may provide theoretical basis for further evaluation of potential clinical efficacy of joint HDAC6 and IC inhibition [185]. These effects of HDAC6 inhibition may be relevant for CRC cells as TIM-3 [211], along with PDL-1/2 [186,200], has been identified on tumor cells and has been designated as a negative prognostic biomarker in colon cancer.
3.3. Combination therapies with HDAC6 and IC inhibitors
Although the IC blockade therapies targeting PD-1 and CTLA-4 have shown considerable therapeutic benefit, they have proved ineffective for some patients presumably due to the development of resistance to therapy, immunosuppressive nature of TME, and lack of antitumor T cell response prior to therapy. As selective HDAC6 inhibition has shown in preclinical settings the ability to alter the expression of PD-L1 and PD-L2 on tumor cells, increase immunogenicity of tumor cells, induce more effective antigen presentation, relieve T cell suppression by downregulating the expression of ICs (PD-1, TIM-3, LAG-3) on immune cells [121,184], and reverse TME by increasing infiltration of immunostimulating and inhibiting differentiation of suppressive immune cells, the combined application of HDAC6i and immunotherapy has recently emerged as significant approach in the field of cancer treatment [212]. Therefore, the use of HDAC inhibitors to augment antitumor T cell responses following the application of IC inhibitors may increase the number of patients that respond to IC blockade therapy.
Experimental data obtained on HDAC6 inhibition in combination with immunotherapeutic agents in the settings of tumor cell lines and animal models have shown improved antitumor effect of this drug combination compared to each agent alone. The examples that support this are investigations of ACY1215 in combination with anti-PD-L1 therapy in ovarian carcinoma [213], ACY-241 in combination with anti-PD-L1 antibody in MM [214], Nex A in combination with anti-PD-1 antibodies in melanoma [121], etc. Based on the similar and growing experimental data a considerable number of clinical studies were conducted in advanced solid tumors. The synergistic effect of pan-HDACi vorinostat with anti-PD-1 agent pembrolizumab was shown in metastatic non-small cell lung cancer (NSCLC) patients with good tolerance of both agents in investigated patients [215]. Another phase 2 clinical trial showed that combined application of vorinostat with pembrolizumab and ER antagonist tamoxifen in ER-positive breast cancer patients resulted in prolonged progression free survival of treated patients [216]. Phase 2 trial involving combined application of pembrolizumab and vorinostat in recurrent metastatic head and neck showed significant efficacy although with a relatively higher grade of toxicity compared with anti-PD-1 monotherapy [217]. Regarding the studies on the selective HDAC6i in solid tumors, some therapeutic benefit was shown in patients with previously treated advanced NSCLC treated with ACY-241 in combination with PD-1 blocking drug nivolumab [218].
Regarding CRC, in patients with microsatellite stable (MSS) CRC that do not respond to IC inhibitors, anti-tumor activity of HDACi CXD101 and nivolumab (anti-PD-1) was assessed in phase II clinical and showed good tolerance and efficacy in the treatment of advanced or metastatic microsatellite-stable CRC [219]. Furthermore, there is an ongoing phase I clinical study on joint application of pan-HDACi romidepsin in combination with anti-PD-1 agent pembrolizumab in mismatch repair proficient CRC [220].
Therefore, there is a need for broadening of the research on the role of HDACi and IC inhibitors in antitumor treatment to better evaluate their therapeutic potential and broaden synergistic application in a wider range of tumors.
3.4. Novel HDAC6 based therapeutical approaches
The use of drug combinations can provide efficacy by targeting different signaling pathways and may reverse drug resistance. Although preclinical studies showed that HDACis in combination with other anticancer agents have a better antitumor effect, results obtained in clinical trials have not been in accordance with this. Simultaneous disruption of different signaling pathways and processes aims to reduce tumor cell proliferation and induce tumor cell death but unfortunately some interactions between simultaneously applied drugs cannot be predicted. Desirable additive or synergistic effects of drug combinations may thus impose a risk of inducing adverse effects due to unwanted drug to drug interactions [221,222].
Given the advantages and disadvantages of combinational therapies, a novel approach in pharmacology is the design of a drug that can interact with two different targets that are common in a certain disease. Therefore, new treatments have been developed by conjugating two distinct therapeutic compounds in a single molecule for dual- targeting strategies. This may provide increased efficacy of the drug by targeting additional disease-related pathways. In this sense, dual HDAC and kinase inhibitors have been tested in preclinical and clinical settings [222,223]. At preclinical level the research on HDACis has evolved and compounds that HDACs interact with receptor tyrosine kinases including PI3K, Src, CDKs, JAKs as well as HDAC have been developed [221,223] Class II HDAC6 selective inhibitors that simultaneously target JAK via conjugation of JAK2 selective inhibitor pacritinib with SAHA, has shown improved selectivity for JAK2 [224].
As in CRC harboring BRAF-V600E mutations, the response rate to BRAF inhibitors is low due to the emergence of resistance. With the aim to effectively target BRAF mutated CRC, novel series of hydroxamate acid and 2-aminopyridinyl-containing compounds such as BRAF and HDAC dual-targeted inhibitors were synthetized. The compounds exerted enzymatic inhibitory activities against BRAFV600E, HDAC1/6 and suppressed the proliferation of CRC cells harboring both wild-type BRAF and mutated BRAFV600E [225]. Therefore, in theory, the optimal pathway blockade can be achieved by simultaneously targeting multiple steps of the pathway.
Other combinations in development include dual inhibitors that contain DNA targeting agents such as DNA-alkylating agents temozolomide [221]. Furthermore, dual HDAC6 inhibitor that contained Hsp90 inhibitor, the compound identified as compound 12 (dual HDAC6 Hsp90) displayed inhibitory effects towards the HDAC6 isoform and 246-fold higher selectivity for HDAC6 over HDAC 1, 3 and 8 isoforms and was endowed with significant cytotoxic effects against CRC cell lines [159].
As was stated before, multi-target drug design approach is aiming to enhance drug activity, selectivity, and overcome drug resistance. Recently, the proteolysis targeting chimera (PROTAC) has become a revolutionary technology in modern drug discovery. PROTACs are bifunctional small molecules consisting of an E3 ubiquitin ligase recognition motif and a ligand for the target protein of interest connected by a suitable linker. PROTACS regulate the expression of the target protein of interest at the posttranslational level by inducing its degradation in proteasomes. The advantages of this catalytic mode of action are enhanced selectivity and improved potency. Moreover, this mode of action results in acute post-translational depletion of pathological protein (target) and eliminates the risk of therapeutic resistance due to physiological feedback mechanisms that upregulate the target protein. In the field of HDAC targeting PROTACs, HDAC6 was designated as the promising target. Several examples in the literature have demonstrated that HDAC6 could be selectively degraded by converting either HDAC6-specific or even pan-HDAC inhibitors into PROTACs. The first HDAC6 degrader was generated based on nonselective HDAC inhibitor and pomalidomide as a ligand for the E3 ubiquitin ligase cereblon (CRBN). CRBN ligands rely on the structure of the anticancer drug thalidomide and its derivatives and have proved to be efficient and selective degraders of HDAC6 [226]. This may be due to cellular localization of HDAC6 or to formation of more efficient ternary complex [221,226,227]. Until recently, most research on HDAC PROTACs was focused on hematological malignancies that are more sensitive to HDAC6 degraders regarding degradation of HDAC6 [227] although some encouraging results were observed in tumor cell lines derived from solid tumors [228]. However, since this research area has very recently emerged it is too early to draw any conclusions.
4. Concluding remarks
HDAC6 has an essential role in many cellular signaling pathways that enable cancer cells to survive and maintain their malignant phenotype. Overexpression of HDAC6 in multiple tumors identifies HDAC6 inhibition as a promising strategy for cancer therapy. Considering the role of HDAC6 in metastatic invasion to regional lymphatics and distant organs, HDAC6 inhibition has important therapeutic implications for the treatment of metastasis in cancer patients. However, despite the promising results shown in experimental settings, high selectivity of HDAC6 inhibition and low toxicity, clinical results were not always promising. The effect of selective HDAC6 inhibition has, in combination with other antitumor treatments, yielded better treatment outcome in some tumors. Furthermore, as selective HDAC6is increase immunogenicity of tumor cells, effective antigen presentation, immune cell functions, and reduce immunosuppression in TME, HDAC6is could potentially improve antitumor immune response in the context of other therapeutic modalities as well as reverse resistance to targeted therapy and IC inhibitors. Further studies should be focused on the improvement of HDAC6 inhibitors to increase their selectivity and accumulation in solid tumors so that adverse effects exerted by high doses on healthy cells would be minimized. Novel approaches in drug design that are still under development such as bifunctional inhibitors and PROTACs represent a growing field for future investigations of HDAC6-based antitumor therapy.
Funding
This study was funded by the Horizon Europe STEPUPIORS Project (HORIZON-WIDERA-2021-ACCESS-03, European Commission, Agreement No. 101079217) and the Ministry of Education and Science of the Republic of Serbia (Agreement No. 451-03-47/2023-01/200043).
References
- Seto, E.; Yoshida, M. Erasers of histone acetylation: the histone deacetylase enzymes. Cold Spring Harb Perspect Biol. 2014, 6:a018713. [CrossRef]
- Seidel, C,; Schnekenburger, M.; Dicato M, Diederich M. Histone deacetylase 6 in health and disease. Epigenomics. 2015, 7:103-18. [CrossRef]
- Grozinger, CM.; Hassig, CA.; Schreiber, SL. Three proteins define a class of human histone deacetylases related to yeast Hda1p. Proc Natl Acad Sci U S A. 1999, 96:4868-73. [CrossRef]
- Hai, Y.; Christianson, D.W. Histone deacetylase 6 structure and molecular basis of catalysis and inhibition. Nat Chem Biol. 2016, 12:741-7. [CrossRef]
- Li, Y.; Shin, D.; Kwon, SH. Histone deacetylase 6 plays a role as a distinct regulator of diverse cellular processes. FEBS J. 2013 280(3):775-93. [CrossRef]
- Zhang, Y.; Li, N.; Caron, C.; Matthias, G.; Hess, D.; Khochbin, S.; Matthias, P. HDAC-6 interacts with and deacetylates tubulin and microtubules in vivo. EMBO J. 2003, 22:1168-79. [CrossRef]
- García-Domínguez, D. J.; Hontecillas-Prieto, L.; Kaliszczak, M.; He, M.; Burguillos, M. A.; Bekay, R.; Abdul-Salam, V.B.; Khozoie, C.; Shah, K.; O’Neill, K.; de Álava, E.; Silver, A.; Syed, N.; Aboagye, E. O.; Hajji, N. Novel nuclear role of HDAC6 in prognosis and therapeutic target for colorectal cancer. bioRxiv 2020.11.02.356121. [CrossRef]
- Pulya, S.; Amin, S.A.; Adhikari, N.; Biswas, S.; Jha, T.; Ghosh, B. HDAC6 as privileged target in drug discovery: A perspective. Pharmacol Res. 2021, 163:105274. [CrossRef]
- Subramanian, C.; Jarzembowski, J.A.; Opipari, A.W. Jr.; Castle, V.P.; Kwok, R.P. HDAC6 deacetylates Ku70 and regulates Ku70-Bax binding in neuroblastoma. Neoplasia. 2011, 13:726-34. [CrossRef]
- Gao, L.; Cueto, M.A.; Asselbergs, F.; Atadja, P. Cloning and functional characterization of HDAC11, a novel member of the human histone deacetylase family. J Biol Chem. 2002, 277:25748-55. [CrossRef]
- Fernandes, I.; Bastien, Y.; Wai, T.; Nygard, K.; Lin, R., Cormier; Lee, H.S., Eng; Bertos, N.R.; Pelletier, N.; Mader, S.; Han, V.K.; Yang, X.J.; White, J.H. Ligand-dependent nuclear receptor corepressor LCoR functions by histone deacetylase-dependent and -independent mechanisms. Mol Cell. 2003, 11:139-50. [CrossRef]
- Yang, C.J.; Liu, Y.P.; Dai, H.Y.; Shiue, Y.L.; Tsai, C.J.; Huang, M.S.; Yeh, Y.T. Nuclear HDAC6 inhibits invasion by suppressing NF-κB/MMP2 and is inversely correlated with metastasis of non-small cell lung cancer. Oncotarget. 2015, 6:30263-76. [CrossRef]
- Zhu, Y.; Feng, M.; Wang, B.; Zheng, Y.; Jiang, D.; Zhao, L.; Mamun, M.A.A.; Kang, H.; Nie, H., Zhang; Guo, N.; Qin, S.; Wang, N.; Liu, H.; Gao, Y. New insights into the non-enzymatic function of HDAC6. Biomed Pharmacother. 2023, 161:114438. [CrossRef]
- Williams, K.A.; Zhang, M.; Xiang, S.; Hu, C.; Wu, J.Y.; Zhang, S.; Ryan, M.; Cox, A.D.; Der, C.J.; Fang, B.; Koomen, J.; Haura, E.; Bepler, G.; Nicosia, S.V.; Matthias, P.; Wang, C.; Bai, W.; Zhang, X. Extracellular signal-regulated kinase (ERK) phosphorylates histone deacetylase 6 (HDAC6) at serine 1035 to stimulate cell migration. J Biol Chem. 2013, 288:33156-70. [CrossRef]
- Hook, S.S.; Orian, A.; Cowley, S.M.; Eisenman, R.N. Histone deacetylase 6 binds polyubiquitin through its zinc finger (PAZ domain) and copurifies with deubiquitinating enzymes. Proc Natl Acad Sci U S A. 2002, 99:13425-30. [CrossRef]
- Han, Y.; Jeong, H.M.; Jin, Y.H.; Kim, Y.J.; Jeong, H.G.; Yeo, C.Y.; Lee, K.Y. Acetylation of histone deacetylase 6 by p300 attenuates its deacetylase activity. Biochem Biophys Res Commun. 2009, 383:88-92. [CrossRef]
- Li, G.; Tian, Y.; Zhu, W.G. The Roles of Histone Deacetylases and Their Inhibitors in Cancer Therapy. Front Cell Dev Biol. 2020, 8:576946. [CrossRef]
- McKinsey, T.A.; Zhang, C.L.; Olson, E.N. Identification of a signal-responsive nuclear export sequence in class II histone deacetylases. Mol Cell Biol. 2001, 21:6312-21. [CrossRef]
- McKinsey, T.A.; Kuwahara, K.; Bezprozvannaya, S.; Olson, E.N. Class II histone deacetylases confer signal responsiveness to the ankyrin-repeat proteins ANKRA2 and RFXANK. Mol Biol Cell. 2006, 17:438-47. [CrossRef]
- Liu, Y.; Peng, L.; Seto, E.; Huang, S.; Qiu, Y. Modulation of histone deacetylase 6 (HDAC6) nuclear import and tubulin deacetylase activity through acetylation. J Biol Chem. 2012, 287:29168-74. [CrossRef]
- Hubbert, C.; Guardiola, A.; Shao, R.; Kawaguchi, Y.; Ito, A.; Nixon, A.; Yoshida, M.; Wang, X.F.; Yao, T.P. HDAC6 is a microtubule-associated deacetylase. Nature. 2002, 417:455-8. [CrossRef]
- Zhang, Y.; Kwon, S.; Yamaguchi, T.; Cubizolles, F.; Rousseaux, S.; Kneissel, M.; Cao, C.; Li, N.; Cheng, H.L.; Chua, K.; Lombard, D.; Mizeracki, A.; Matthias, G.; Alt, F.W.; Khochbin, S.; Matthias, P. Mice lacking histone deacetylase 6 have hyperacetylated tubulin but are viable and develop normally. Mol Cell Biol. 2008, 28:1688–701. [CrossRef]
- Valenzuela-Fernández, A.; Cabrero, J.R.; Serrador, J.M.; Sánchez-Madrid, F. HDAC6: a key regulator of cytoskeleton, cell migration and cell-cell interactions. Trends Cell Biol. 2008, 18:291-7. [CrossRef]
- Chuma, M.; Sakamoto, M.; Yasuda, J.; Fujii, G.; Nakanishi, K.; Tsuchiya, A.; Ohta, T.; Asaka, M.; Hirohashi, S. Overexpression of cortactin is involved in motility and metastasis of hepatocellular carcinoma. J Hepatol. 2004, 41:629-36. [CrossRef]
- Zhang, X.; Yuan, Z.; Zhang, Y.; Yong, S.; Salas-Burgos, A.; Koomen, J.; Olashaw, N.; Parsons, J.T.; Yang, X.J.; Dent, S.R.; Yao, T.P.; Lane, W.S.; Seto, E. HDAC6 modulates cell motility by altering the acetylation level of cortactin. Mol Cell. 2007, 27:197-213. [CrossRef]
- Zhang, X.; Liu, K.; Zhang, T.; Wang, Z.; Qin, X.; Jing, X.; Wu, H.; Ji, X.; He, Y.; Zhao, R. Cortactin promotes colorectal cancer cell proliferation by activating the EGFR-MAPK pathway. Oncotarget. 2017, 8:1541-1554. [CrossRef]
- Boyault, C.; Zhang, Y.; Fritah, S.; Caron, C.; Gilquin, B.; Kwon, S.H.; Garrido, C.; Yao, T.P.; Vourc'h, C.; Matthias, P.; Khochbin, S. HDAC6 controls major cell response pathways to cytotoxic accumulation of protein aggregates. Genes Dev. 2007, 21:2172-81. [CrossRef]
- Johnston, H.E.; Samant, R.S. Alternative systems for misfolded protein clearance: life beyond the proteasome. FEBS J. 2021, 288:4464-4487. [CrossRef]
- Seigneurin-Berny, D.; Verdel, A.; Curtet, S.; Lemercier, C.; Garin, J.; Rousseaux, S.; Khochbin, S. Identification of components of the murine histone deacetylase 6 complex: link between acetylation and ubiquitination signaling pathways. Mol Cell Biol. 2001, 21:8035-44. [CrossRef]
- Lee, J.Y.; Koga, H.; Kawaguchi, Y.; Tang, W.; Wong, E.; Gao, Y.S.; Pandey, U.B.; Kaushik, S.; Tresse, E.; Lu, J.; Taylor, J.P.; Cuervo, A.M.; Yao, T.P. HDAC6 controls autophagosome maturation essential for ubiquitin-selective quality-control autophagy. EMBO J. 2010, 29:969-80. [CrossRef]
- Watabe, M.; Nakaki, T. Protein kinase CK2 regulates the formation and clearance of aggresomes in response to stress. J Cell Sci. 2011, 124:1519-32. [CrossRef]
- Watanabe, Y.; Tanaka, M. p62/SQSTM1 in autophagic clearance of a non-ubiquitylated substrate. J Cell Sci. 2011, 124:2692-701. [CrossRef]
- Fusco, C.; Micale, L.; Egorov, M.; Monti, M.; D'Addetta, E.V.; Augello, B.; Cozzolino, F.; Calcagnì, A.; Fontana, A.; Polishchuk, R.S.; Didelot, G.; Reymond, A.; Pucci, P.; Merla, G. The E3-ubiquitin ligase TRIM50 interacts with HDAC6 and p62, and promotes the sequestration and clearance of ubiquitinated proteins into the aggresome. PLoS One. 2012, 7:e40440. [CrossRef]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell Mol Biol Lett. 2016, 21:29. [CrossRef]
- Yan, J.; Seibenhener, M.L.; Calderilla-Barbosa, L.; Diaz-Meco, M.T.; Moscat, J.; Jiang, J.; Wooten, M.W.; Wooten, M.C. SQSTM1/p62 interacts with HDAC6 and regulates deacetylase activity. PLoS One. 2013, 8:e76016. [CrossRef]
- Boyault, C.; Gilquin, B.; Zhang, Y.; Rybin, V.; Garman, E.; Meyer-Klaucke, W.; Matthias, P.; Müller, C.W.; Khochbin, S. HDAC6-p97/VCP controlled polyubiquitin chain turnover. EMBO J. 2006, 25:3357-66. [CrossRef]
- Passaro, E.; Papulino, C.; Chianese, U.; Toraldo, A.; Congi, R.; Del Gaudio, N.; Nicoletti, M.M.; Benedetti, R.; Altucci, L. HDAC6 Inhibition Extinguishes Autophagy in Cancer: Recent Insights. Cancers (Basel). 2021, 13:6280. [CrossRef]
- Zhang, J.; Wang, J.; Zhou, Z.; Park, J.E.; Wang, L.; Wu, S.; Sun, X.; Lu, L.; Wang, T.; Lin, Q.; Sze, S.K.; Huang, D.; Shen, H.M. Importance of TFEB acetylation in control of its transcriptional activity and lysosomal function in response to histone deacetylase inhibitors. Autophagy. 2018, 14:1043-1059. [CrossRef]
- Zhang, J.; Ng, S.; Wang, J.; Zhou, J.; Tan, S.H.; Yang, N.; Lin, Q.; Xia, D.; Shen, H.M. Histone Deacetylase Inhibitors Induce Autophagy Through FOXO1-Dependent Pathways. Autophagy 2015, 11:629–42. [CrossRef]
- Li, C.; Wang, X.; Li, X.; Qiu, K.; Jiao, F.; Liu, Y.; Kong, Q.; Liu, Y.; Wu, Y. Proteasome Inhibition Activates Autophagy-Lysosome Pathway Associated With TFEB Dephosphorylation and Nuclear Translocation. Front Cell Dev Biol. 2019, 22;7:170. [CrossRef]
- Brijmohan, A.S.; Batchu, S.N.; Majumder, S.; Alghamdi, T.A.; Thieme, K.; McGaugh, S.; Liu, Y.; Advani, S.L.; Bowskill, B.B.; Kabir, M.G.; Geldenhuys, L.; Siddiqi, F.S.; Advani, A. HDAC6 Inhibition Promotes Transcription Factor EB Activation and Is Protective in Experimental Kidney Disease. Front Pharmacol. 2018, 9:34. [CrossRef]
- Jung, K.H.; Noh, J.H.; Kim, J.K.; Eun, J.W.; Bae, H.J.; Chang, Y.G.; Kim, M.G.; Park, W.S.; Lee, J.Y.; Lee, S.Y.; Chu, I.S.; Nam, S.W. Histone deacetylase 6 functions as a tumor suppressor by activating c-Jun NH2-terminal kinase-mediated beclin 1-dependent autophagic cell death in liver cancer. Hepatology. 2012, 56(2):644-57. [CrossRef]
- Chang, P.; Li, H.; Hu, H.; Li, Y.; Wang, T. The Role of HDAC6 in Autophagy and NLRP3 Inflammasome. Front Immunol. 2021, 12:763831. [CrossRef]
- Kerdiles, Y.M.; Stone, E.L.; Beisner, D.R.; McGargill, M.A.; Ch'en, I.L.; Stockmann, C.; Katayama, C.D.; Hedrick, S.M. Foxo transcription factors control regulatory T cell development and function. Immunity. 2010, 33(6):890-904. [CrossRef]
- Huang, R.; Liu, W. Identifying an Essential Role of Nuclear LC3 for Autophagy. Autophagy 2015, 11:852–3. [CrossRef]
- Liu, K.P.; Zhou, D.; Ouyang, D.Y.; Xu, L.H.; Wang, Y.; Wang, L.X.; Pan, H.; He, X.H. LC3B-II deacetylation by histone deacetylase 6 is involved in serum-starvation-induced autophagic degradation. Biochem Biophys Res Commun. 2013, 441:970-5. [CrossRef]
- Xie, R.; Nguyen, S.; McKeehan, W.L.; Liu, L. Acetylated microtubules are required for fusion of autophagosomes with lysosomes. BMC Cell Biol. 2010, 11:89. [CrossRef]
- Liu, P.; Xiao, J.; Wang, Y.; Song, X.; Huang, L.; Ren, Z.; Kitazato, K.; Wang, Y. Posttranslational modification and beyond: interplay between histone deacetylase 6 and heat-shock protein 90. Mol Med. 2021, 27(1):110. [CrossRef]
- Kovacs, J.J.; Murphy, P.J.; Gaillard, S.; Zhao, X.; Wu, J.T.; Nicchitta, C.V.; Yoshida, M.; Toft, D.O.; Pratt, W.B.; Yao, T.P. HDAC6 regulates Hsp90 acetylation and chaperone-dependent activation of glucocorticoid receptor. Mol Cell. 2005, 18:601-7. [CrossRef]
- Tan, Y.; Ci, Y.; Dai, X.; Wu, F.; Guo, J.; Liu, D.; North, B.J.; Huo, J.; Zhang, J. Cullin 3SPOP ubiquitin E3 ligase promotes the poly-ubiquitination and degradation of HDAC6. Oncotarget. 2017, 8:47890-47901. [CrossRef]
- Li, Z.Y.; Zhang, C.; Zhang, Y.; Chen, L.; Chen, B.D.; Li, Q.Z.; Zhang, X.J.; Li, W.P. A novel HDAC6 inhibitor Tubastatin A: controls HDAC6-p97/VCP-mediated ubiquitination-autophagy turnover and reverses Temozolomide-induced ER stress-tolerance in GBM cells. Cancer Lett. 2017, 39:89-99. [CrossRef]
- Zhang, S.; Guo, S.; Li, Z.; Li, D.; Zhan, Q. High expression of HSP90 is associated with poor prognosis in patients with colorectal cancer. PeerJ. 2019, 31;7:e7946. [CrossRef]
- Moser, C.; Lang, S.A.; Stoeltzing, O. Heat-shock protein 90 (Hsp90) as a molecular target for therapy of gastrointestinal cancer. Anticancer Res. 2009, 29:2031-42.
- Kamemura, K.; Ogawa, M.; Ohkubo, S.; Ohtsuka, Y.; Shitara, Y.; Komiya, T.; Maeda, S.; Ito, A.; Yoshida, M. Depression of mitochondrial metabolism by downregulation of cytoplasmic deacetylase, HDAC6. FEBS Lett. 2012, 586(9):1379-83. [CrossRef]
- Sergi, C.M. Targeting the 'garbage-bin' to fight cancer: HDAC6 inhibitor WT161 has an anti-tumor effect on osteosarcoma and synergistically interacts with 5-FU. Biosci Rep. 2021,41:BSR20210952. [CrossRef]
- Mhaidat, N.M.; Alzoubi, K.H.; Khabour, O.F.; Banihani, M.N.; Al-Balas, Q.A.; Swaidan, S. GRP78 regulates sensitivity of human colorectal cancer cells to DNA targeting agents. Cytotechnology. 2016 May;68(3):459-67. [CrossRef]
- Li, Z.; Li, Z. Glucose regulated protein 78: a critical link between tumor microenvironment and cancer hallmarks. Biochim Biophys Acta. 2012, 1826:13-22. [CrossRef]
- Li, Z.; Zhuang, M.; Zhang, L.; Zheng, X.; Yang, P.; Li, Z. Acetylation modification regulates GRP78 secretion in colon cancer cells. Sci Rep. 2016, 7;6:30406. [CrossRef]
- Zhang, S.L.; Zhu, H.Y.; Zhou, B.Y.; Chu, Y.; Huo, J.R.; Tan, Y.Y.; Liu, D.L. Histone deacetylase 6 is overexpressed and promotes tumor growth of colon cancer through regulation of the MAPK/ERK signal pathway. Onco Targets Ther. 2019,12:2409-2419. [CrossRef]
- Cai, H.; Gong, L.; Liu, J.; Zhou, Q.; Zheng, Z. Diosgenin inhibits tumor angiogenesis through regulating GRP78-mediated HIF-1α and VEGF/VEGFR signaling pathways. Pharmazie. 2019, 74:680-684. [CrossRef]
- Parmigiani, R.B.; Xu, W.S.; Venta-Perez, G.; Erdjument-Bromage, H.; Yaneva, M.; Tempst, P.; Marks, P.A. HDAC6 is a specific deacetylase of peroxiredoxins and is involved in redox regulation. Proc Natl Acad Sci U S A. 2008, 105:9633-8. [CrossRef]
- Harris, S.; Levine, A. The p53 pathway: positive and negative feedback loops. Oncogene 2005, 24:2899–2908. [CrossRef]
- Ito, A.; Lai, C.H.; Zhao, X.; Saito, S.; Hamilton, M.H.; Appella, E.; Yao, T.P. p300/CBP-mediated p53 acetylation is commonly induced by p53-activating agents and inhibited by MDM2. EMBO J. 2001, 20:1331-40. [CrossRef]
- Brooks, C.L.; Gu, W. The impact of acetylation and deacetylation on the p53 pathway. Protein Cell. 2011, 2:456-62. [CrossRef]
- Ryu, H.W.; Shin, D.H.; Lee, D.H.; Choi, J.; Han, G; Lee, K.Y., Kwon. HDAC6 deacetylates p53 at lysines 381/382 and differentially coordinates p53-induced apoptosis. Cancer Lett. 2017, 391:162-171. [CrossRef]
- Ding, G.; Liu, H.D.; Huang, Q.; Liang, H.X.; Ding, Z.H.; Liao, Z.J.; Huang., G. HDAC6 promotes hepatocellular carcinoma progression by inhibiting P53 transcriptional activity. FEBS Lett. 2013, 587:880-6. [CrossRef]
- Mrakovcic, M.; Bohner, L.; Hanisch, M.; Fröhlich, L.F. Epigenetic Targeting of Autophagy via HDAC Inhibition in Tumor Cells: Role of p53. Int J Mol Sci. 2018, 19:3952. [CrossRef]
- Zopf, S.; Neureiter, D.; Bouralexis, S.; Abt, T.; Glaser, K.B.; Okamoto, K.; Ganslmayer, M.; Hahn, E.G.; Herold, C.; Ocker, M. Differential response of p53 and p21 on HDAC inhibitor-mediated apoptosis in HCT116 colon cancer cells in vitro and in vivo. Int J Oncol. 2007,31:1391-402. [CrossRef]
- Liebl, M.C.; Hofmann, T.G. The Role of p53 Signaling in Colorectal Cancer. Cancers (Basel). 2021, 13:2125. [CrossRef]
- Li, D.; Marchenko, N.D.; Schulz, R.; Fischer, V.; Velasco-Hernandez, T.; Talos, F.; Moll, U.M. Functional inactivation of endogenous MDM2 and CHIP by HSP90 causes aberrant stabilization of mutant p53 in human cancer cells. Mol Cancer Res. 2011, 9:577-88. [CrossRef]
- Li, D.; Marchenko, N.D.; Moll, U.M. SAHA shows preferential cytotoxicity in mutant p53 cancer cells by destabilizing mutant p53 through inhibition of the HDAC6-Hsp90 chaperone axis. Cell Death Differ. 2011, 18:1904-13. [CrossRef]
- Parrales, A.; Iwakuma, T. Targeting oncogenic mutant p53 for cancer therapy. Front. Oncol. 2015, 5, 288. [CrossRef] [PubMed]
- Rasouli, M.; Khakshournia, S.; Vakili, O.; Dastghaib, S.; Seghatoleslam, A.; Shafiee, S.M. The crosstalk between ubiquitin-conjugating enzyme E2Q1 and p53 in colorectal cancer: An in vitro analysis. Med Oncol. 2023, 40:199. [CrossRef]
- Hirose, T.; Sowa, Y.; Takahashi, S.; Saito, S.; Yasuda, C.; Shindo, N.; Furuichi, K.; Sakai, T. p53-independent induction of Gadd45 by histone deacetylase inhibitor: coordinate regulation by transcription factors Oct-1 and NF-Y. Oncogene. 2003, 22:7762-73. [CrossRef]
- Gong, P.; Wang, Y.; Jing, Y. Apoptosis Induction by Histone Deacetylase Inhibitors in Cancer Cells: Role of Ku70. Int J Mol Sci. 2019, 20:1601. [CrossRef]
- Meng, J.; Zhang, F.; Zhang, X.T.; Zhang, T.; Li, Y.H.; Fan, L.; Sun, Y.; Zhang, H.L.; Mei, Q.B. Ku70 is essential for histone deacetylase inhibitor trichostatin A-induced apoptosis. Mol Med Rep. 2015, 2:581-6. [CrossRef]
- Kerr, E.; Holohan, C.; McLaughlin, K.M.; Majkut, J.; Dolan, S.; Redmond, K.; Riley, J.; McLaughlin, K.; Stasik, I.; Crudden, M.; Van Schaeybroeck, S.; Fenning, C.; O'Connor, R.; Kiely, P.; Sgobba, M.; Haigh, D.; Johnston, P.G.; Longley, D.B. Identification of an acetylation-dependant Ku70/FLIP complex that regulates FLIP expression and HDAC inhibitor-induced apoptosis. Cell Death Differ. 2012, 19:1317-27. [CrossRef]
- Riolo, M.T.; Cooper, Z.A.; Holloway, M.P.; Cheng, Y.; Bianchi, C.; Yakirevich, E.; Ma, L.; Chin, Y.E.; Altura, R.A. Histone deacetylase 6 (HDAC6) deacetylates survivin for its nuclear export in breast cancer. J Biol Chem. 2012, 287:10885-93. [CrossRef]
- Jin, J.S.; Tsao, T.Y.; Sun, P.C.; Yu, C.P.; Tzao, C. SAHA inhibits the growth of colon tumors by decreasing histone deacetylase and the expression of cyclin D1 and survivin. Pathol Oncol Res. 2012, 18:713-20. [CrossRef]
- Westendorf, J.J.; Zaidi, S.K.; Cascino, J.E.; Kahler, R.; van Wijnen, A.J.; Lian, J.B.; Yoshida, M.; Stein, G.S.; Li, X. Runx2 (Cbfa1, AML-3) interacts with histone deacetylase 6 and represses the p21(CIP1/WAF1) promoter. Mol Cell Biol. 2002, 22:7982-92. [CrossRef]
- Yi, H.; Li, G.; Long, Y.; Liang, W.; Cui, H.; Zhang, B.; Tan, Y.; Li, Y.; Shen, L.; Deng, D.; Tang, Y.; Mao, C.; Tian, S.; Cai, Y.; Zhu, Q.; Hu, Y.; Chen, W.; Fang, L. Integrative multi-omics analysis of a colon cancer cell line with heterogeneous Wnt activity revealed RUNX2 as an epigenetic regulator of EMT. Oncogene. 2020 Jul;39(28):5152-5164. [CrossRef]
- Lin, Tsung-Chieh. 2023. "RUNX2 and Cancer" International Journal of Molecular Sciences 24, no. 8: 7001. [CrossRef]
- Wu, J.Y.; Xiang, S.; Zhang, M.; Fang, B.; Huang, H.; Kwon, O.K.; Zhao, Y.; Yang, Z.; Bai, W.; Bepler, G.; Zhang, X.M. Histone deacetylase 6 (HDAC6) deacetylates extracellular signal-regulated kinase 1 (ERK1) and thereby stimulates ERK1 activity. J Biol Chem. 2018, 293:1976-1993. [CrossRef]
- Tsutsumi, S.; Beebe, K.; Neckers, L. Impact of heat-shock protein 90 on cancer metastasis. Future Oncol. 2009, 5:679-88. [CrossRef]
- Iaconelli, J.; Lalonde, J.; Watmuff, B.; Liu, B.; Mazitschek, R.; Haggarty, S.J.; Karmacharya, R. Lysine Deacetylation by HDAC6 Regulates the Kinase Activity of AKT in Human Neural Progenitor Cells. ACS Chem Biol. 2017, 12:2139-2148. [CrossRef]
- Johnson, S.M.; Gulhati, P.; Rampy, B.A.; Han, Y.; Rychahou, P.G.; Doan, H.Q.; Weiss, H.L.; Evers, B.M. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J Am Coll Surg. 2010, 210:767-76, 776-8. [CrossRef]
- Deribe, Y.L.; Wild, P.; Chandrashaker, A.; Curak, J.; Schmidt, M.H.H.; Kalaidzidis, Y.; Milutinovic, N.; Kratchmarova, I.; Buerkle, L.; Fetchko, M.J.; Schmidt, P.; Kittanakom, S.; Brown, K.R.; Jurisica, I.; Blagoev, B.; Zerial, M.; Stagljar, I.; Dikic, I. Regulation of epidermal growth factor receptor trafficking by lysine deacetylase HDAC6. Sci Signal. 2009, 2:ra84. [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci. 2019, 20:3328. [CrossRef]
- Moreno-Gonzalo, O.; Ramírez-Huesca, M.; Blas-Rus, N.; Cibrián, D.; Saiz, M.L.; Jorge, I.; Camafeita, E.; Vázquez, J.; Sánchez-Madrid, F. HDAC6 controls innate immune and autophagy responses to TLR-mediated signalling by the intracellular bacteria Listeria monocytogenes. PLoS Pathog. 2017, 13:e1006799. [CrossRef]
- Youn, G.S.; Lee, K.W.; Choi, S.Y.; Park, J. Overexpression of HDAC6 Induces Pro-Inflammatory Responses by Regulating ROS-MAPK-NF-Kappab/AP-1 Signaling Pathways in Macrophages. Free Radic Biol Med. 2016, 97:14–23. [CrossRef]
- Xu, S.; Chen, H.; Ni, H.; Dai, Q. Targeting HDAC6 attenuates nicotine-induced macrophage pyroptosis via NF-κB/NLRP3 pathway. Atherosclerosis. 2021, 317:1-9. [CrossRef]
- Sharma, B.R.; Kanneganti, T.D. Inflammasome signaling in colorectal cancer. Transl Res. 2023, 252:45-52. [CrossRef]
- Zhang, L.; Wang, Y.; Liu, X.; Zhang, Y. NLRP3 Inflammasome Activation in MΦs-CRC Crosstalk Promotes Colorectal Cancer Metastasis. Ann Clin Lab Sci. 2022, 52:571-579.
- Vafaei, S.; Taheri, H.; Hajimomeni, Y.; Fakhre Yaseri, A.; Abolhasani Zadeh, F. The role of NLRP3 inflammasome in colorectal cancer: potential therapeutic target. Clin Transl Oncol. 2022, 24:1881-1889. [CrossRef]
- Shi, F.; Wei, B.; Lan, T.; Xiao, Y.; Quan, X.; Chen, J.; Zhao, C.; Gao, J. Low NLRP3 expression predicts a better prognosis of colorectal cancer. Biosci Rep. 2021, 41:BSR20210280. [CrossRef]
- Gu, S.; Liu, Y.; Zhu, B.; Ding, K.; Yao, T.P.; Chen, F.; Zhan, L.; Xu, P.; Ehrlich, M.; Liang, T.; Lin, X.; Feng, X.H. Loss of α-Tubulin Acetylation Is Associated with TGF-β-induced Epithelial-Mesenchymal Transition. J Biol Chem. 2016, 291:5396-405. [CrossRef]
- Shan, B.; Yao, T.P.; Nguyen, H.T.; Zhuo, Y.; Levy, D.R.; Klingsberg, R.C.; Tao, H.; Palmer, M.L.; Holder, K.N.; Lasky, J.A. Requirement of HDAC6 for transforming growth factor-beta1-induced epithelial-mesenchymal transition. J Biol Chem. 2008, 283:21065-73. [CrossRef]
- Osseni, A.; Ravel-Chapuis, A., Belotti; Scionti, I.; Gangloff, Y.G.; Moncollin, V.; Mazelin, L.; Mounier, R.; Leblanc, P.; Jasmin, B.J.; Schaeffer, L. Pharmacological inhibition of HDAC6 improves muscle phenotypes in dystrophin-deficient mice by downregulating TGF-β via Smad3 acetylation. Nat Commun. 2022, 13:7108. [CrossRef]
- Shi, Y.; Tao, M.; Ni, J.; Tang, L.; Liu, F.; Chen, H.; Ma, X.; Hu, Y.; Zhou, X.; Qiu, A.; Zhuang, S; Liu, N. Requirement of Histone Deacetylase 6 for Interleukin-6 Induced Epithelial-Mesenchymal Transition, Proliferation, and Migration of Peritoneal Mesothelial Cells. Front Pharmacol. 2021, 12:722638. [CrossRef]
- Dong, C.; Li, Z.; Alvarez, R. Jr.; Feng, X.H. Goldschmidt-Clermont PJ. Microtubule binding to Smads may regulate TGF beta activity. Mol Cell. 2000, 5:27-34. [CrossRef]
- Xu, L.; Liu, N.; Gu, H.; Wang, H.; Shi, Y.; Ma, X.; Ma, S.; Ni, J.; Tao, M.; Qiu, A.; Zhuang, S. Histone deacetylase 6 inhibition counteracts the epithelial–mesenchymal transition of peritoneal mesothelial cells and prevents peritoneal fibrosis. Oncotarget. 2017, 8: 88730-88750. [CrossRef]
- Mirjačić Martinović, K.; Vuletić, A.; Tišma Miletić, N.; Matković, S; Gavrilović, D.; Ninković, A.; Jurišić, V.; Babović, N. Circulating IL-6 is associated with disease progression in BRAFwt metastatic melanoma patients receiving anti-PD-1 therapy. J Clin Pathol. 2023,jcp-2022-208615. [CrossRef]
- Waldner, M.J.; Foersch, S.; Neurath, M.F. Interleukin-6--a key regulator of colorectal cancer development. Int J Biol Sci. 2012, 8:1248-53. [CrossRef]
- Vuletić, A.; Mirjačić Martinović, K.; Tišma Miletić, N.; Zoidakis, J.; Castellvi-Bel, S.; Čavić, M. Cross-Talk Between Tumor Cells Undergoing Epithelial to Mesenchymal Transition and Natural Killer Cells in Tumor Microenvironment in Colorectal Cancer. Front Cell Dev Biol. 2021, 9:750022. [CrossRef]
- Gould, C.M.; Courtneidge, S.A. Regulation of invadopodia by the tumor microenvironment. Cell Adh Migr. 2014, 8:226-35. [CrossRef]
- Schoumacher, M.; Goldman, R.D.; Louvard, D.; Vignjevic, D.M. Actin, microtubules, and vimentin intermediate filaments cooperate for elongation of invadopodia. J Cell Biol. 2010, 189:541-56. [CrossRef]
- Wang, J.; Lin, A.; Lu, L. Effect of EGF-induced HDAC6 activation on corneal epithelial wound healing. Invest Ophthalmol Vis Sci. 2010, 51:2943-8. [CrossRef]
- Arsenault, D.; Brochu-Gaudreau, K.; Charbonneau, M.; Dubois, C.M. HDAC6 deacetylase activity is required for hypoxia-induced invadopodia formation and cell invasion. PLoS One. 2013, 8:e55529. [CrossRef]
- Li, D.; Xie, S.; Ren, Y.; Huo, L.; Gao, J.; Cui, D.; Liu, M.; Zhou, J. Microtubule-associated deacetylase HDAC6 promotes angiogenesis by regulating cell migration in an EB1-dependent manner. Protein Cell. 2011, 2:150-60. [CrossRef]
- Kaluza, D.; Kroll, J.; Gesierich, S.; Yao, T.P.; Boon, R.A.; Hergenreider, E.; Tjwa, M.; Rössig, L.; Seto, E.; Augustin, H.G.; Zeiher, A.M.; Dimmeler, S.; Urbich, C. Class IIb HDAC6 regulates endothelial cell migration and angiogenesis by deacetylation of cortactin. EMBO J. 2011, 30:4142-56. [CrossRef]
- Qian, D.Z.; Kachhap, S.K.; Collis, S.J.; Verheul, H.M.; Carducci, M.A.; Atadja, P.; Pili, R. Class II histone deacetylases are associated with VHL-independent regulation of hypoxia-inducible factor 1 alpha. Cancer Res. 2006, 66:8814-21. [CrossRef]
- Ellis, L.; Hammers, H.; Pili, R. Targeting tumor angiogenesis with histone deacetylase inhibitors. Cancer Lett. 2009, 280:145-53. [CrossRef]
- Park, J.H.; Kim, S.H.; Choi, M.C.; Lee, J.; Oh, D.Y.; Im, S.A.; Bang, Y.J.; Kim, T.Y. Class II histone deacetylases play pivotal roles in heat shock protein 90-mediated proteasomal degradation of vascular endothelial growth factor receptors. Biochem Biophys Res Commun. 2008, 368:318-22. [CrossRef]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of β-catenin. EMBO J. 2012, 31:2714-36. [CrossRef]
- Li, Y.; Zhang, X.; Polakiewicz, R.D.; Yao, T.P.; Comb, M.J. HDAC6 is required for epidermal growth factor-induced beta-catenin nuclear localization. J Biol Chem. 2008, 283:12686-90. [CrossRef]
- Noe, O.; Filipiak, L.; Royfman, R.; Campbell, A.; Lin, L.; Hamouda, D.; Stanbery, L.; Nemunaitis, J. Adenomatous polyposis coli in cancer and therapeutic implications. Oncol Rev. 2021, 15:534. [CrossRef]
- McCaw, T.R.; Randall, T.D.; Forero, A.; Buchsbaum, D.J. Modulation of antitumor immunity with histone deacetylase inhibitors. Immunotherapy. 2017, 9:1359-1372. [CrossRef]
- Cheng, F.; Lienlaf, M.; Wang, H.W.; Perez-Villarroel, P.; Lee, C.; Woan, K.; Rock-Klotz, J.; Sahakian, E.; Woods, D.; Pinilla-Ibarz, J.; Kalin, J.; Tao, J.; Hancock, W.; Kozikowski, A.; Seto, E.; Villagra, A.; Sotomayor, E.M. A novel role for histone deacetylase 6 in the regulation of the tolerogenic STAT3/IL-10 pathway in APCs. J Immunol. 2014, 193:2850-2862. [CrossRef]
- Sulczewski, F.B.; Martino, L.A.; Salles, D.; Yamamoto, M.M.; Rosa, D.S.; Boscardin, S.B. STAT3 signaling modulates the immune response induced after antigen targeting to conventional type 1 dendritic cells through the DEC205 receptor. Front Immunol. 2022, 13:1006996. [CrossRef]
- Hillmer, E.J.; Zhang, H.; Li, H.S.; Watowich, S.S. STAT3 signaling in immunity. Cytokine Growth Factor Rev. 2016, 31:1-15. [CrossRef]
- Knox, T.; Sahakian, E.; Banik, D.; Hadley, M.; Palmer, E.; Noonepalle, S.; Kim, J.; Powers, J.; Gracia-Hernandez, M.; Oliveira, V.; Cheng, F.; Chen, J.; Barinka, C.; Pinilla-Ibarz, J.; Lee, N.H.; Kozikowski, A.; Villagra, A. Selective HDAC6 inhibitors improve anti-PD-1 immune checkpoint blockade therapy by decreasing the anti-inflammatory phenotype of macrophages and down-regulation of immunosuppressive proteins in tumor cells. Sci Rep. 2019, 9:6136. Erratum in: Sci Rep. 2019 Oct 10;9(1):14824. [CrossRef]
- Zhang, Q.Q.; Zhang, W.J.; Chang, S. HDAC6 inhibition: a significant potential regulator and therapeutic option to translate into clinical practice in renal transplantation. Front Immunol. 2023, 14:1168848. [CrossRef]
- Xu, G.; Niu, L.; Wang, Y.; Yang, G.; Zhu, X.; Yao, Y.; Zhao, G.; Wang, S.; Li, H. HDAC6-dependent deacetylation of TAK1 enhances sIL-6R release to promote macrophage M2 polarization in colon cancer. Cell Death Dis. 2022, 13:888. [CrossRef]
- Chen, L.; Wang, S.; Wang, Y.; Zhang, W.; Ma, K.; Hu, C.; Zhu, H.; Liang, S.; Liu, M.; Xu, N. IL-6 influences the polarization of macrophages and the formation and growth of colorectal tumor. Oncotarget. 2018, 9:17443-17454. [CrossRef]
- Nunez-Andrade, N.; Iborra, S.; Trullo, A.; Moreno-Gonzalo, O.; Calvo, E.; Catalán, E.; Menasche, G.; Sancho, D.; Vázquez, J.; Yao, T.P.; Martín-Cófreces, N.B.; Sánchez-Madrid, F. HDAC6 regulates the dynamics of lytic granules in cytotoxic T lymphocytes. J. Cell Sci. 2016, 129: 1305–1311. [CrossRef]
- Beier, U.H.; Wang, L.; Han, R.; Akimova, T.; Liu, Y.; Hancock, W.W. Histone deacetylases 6 and 9 and sirtuin-1 control Foxp3+ regulatory T cell function through shared and isoform-specific mechanisms. Sci Signal. 2012, 5:ra45. [CrossRef]
- de Zoeten, E.F.; Wang, L.; Butler, K.; Beier, U.H.; Akimova, T.; Sai, H.; Bradner, J.E.; Mazitschek, R.; Kozikowski, A.P.; Matthias, P.; Hancock, W.W. Histone deacetylase 6 and heat shock protein 90 control the functions of Foxp3(+) T-regulatory cells. Mol Cell Biol. 2011, 31:2066-78. [CrossRef]
- Xiao, Y.; Li, B.; Zhou, Z.; Hancock, W.W.; Zhang, H.; Greene, M.I. Histone acetyltransferase mediated regulation of FOXP3 acetylation and Treg function. Curr. Opin. Immunol. 2010, 22:583–591. [CrossRef]
- Josefowicz, S.Z.; Lu, L.F.; Rudensky, A.Y. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol. 2012, 30:531-64. [CrossRef]
- Lee, J.H.; Kim, H.S.; Jang, S.W.; Lee, G.R. Histone deacetylase 6 plays an important role in TGF-β-induced murine Treg cell differentiation by regulating cell proliferation. Sci Rep. 2022, 12:22550. [CrossRef]
- Aristin Revilla, S.; Kranenburg, O.; Coffer, P. J. Colorectal Cancer-Infiltrating Regulatory T Cells: Functional Heterogeneity, Metabolic Adaptation, and Therapeutic Targeting. Front Immunol. 2022, 13: 903564. [CrossRef]
- Woan, K.V.; Lienlaf, M.; Perez-Villaroel, P.; Lee, C.; Cheng, F.; Knox, T.; Woods, D.M.; Barrios, K.; Powers, J.; Sahakian, E.; Wang, H.W.; Canales, J.; Marante, D.; Smalley, K.S.M.; Bergman, J.; Seto, E.; Kozikowski, A.; Pinilla-Ibarz, J.; Sarnaik, A.; Celis, E.; Weber, J.; Sotomayor, E.M.; Villagra, A. Targeting histone deacetylase 6 mediates a dual anti-melanoma effect: Enhanced antitumor immunity and impaired cell proliferation. Mol Oncol. 2015, 9:1447-1457. [CrossRef]
- Lee, Y.S.; Lim, K.H.; Guo, X.; Kawaguchi, Y.; Gao, Y.; Barrientos, T.; Ordentlich, P.; Wang, X.F.; Counter, C.M.; Yao, T.P. The cytoplasmic deacetylase HDAC6 is required for efficient oncogenic tumorigenesis. Cancer Res. 2008, 68:7561-9. [CrossRef]
- Aldana-Masangkay, G.I.; Sakamoto, K.M. The role of HDAC6 in cancer. J Biomed Biotechnol. 2011, 2011:875824. [CrossRef]
- Haggarty, S.J.; Koeller, K.M.; Wong, J.C.; Grozinger, C.M; Schreiber, S.L. Domain-selective small-molecule inhibitor of histone deacetylase 6 (HDAC6)-mediated tubulin deacetylation. Proc Natl Acad Sci U S A. 2003, 100:4389-94. [CrossRef]
- Butler, K.V.; Kalin, J.; Brochier, C.; Vistoli, G.; Langley, B.; Kozikowski, A.P. Rational design and simple chemistry yield a superior, neuroprotective HDAC6 inhibitor, tubastatin A. J Am Chem Soc. 2010, 132:10842-6. [CrossRef]
- Zhou, B.; Liu, D.; Tan, Y. Role of HDAC6 and Its Selective Inhibitors in Gastrointestinal Cancer. Front Cell Dev Biol. 2021, 9:719390. [CrossRef]
- Zhang, S.L.; Du, X.; Tan, L.N.; Deng, F.H.; Zhou, B.Y.; Zhou, H.J.; Zhu, H.Y.; Chu, Y.; Liu, D.L.; Tan, Y.Y. SET7 interacts with HDAC6 and suppresses the development of colon cancer through inactivation of HDAC6. Am J Transl Res. 2020, 12:602-61.
- Duvic, M.; Vu, J. Vorinostat: a New Oral Histone Deacetylase Inhibitor Approved for Cutaneous T-Cell Lymphoma. Expert Opin. Investig. Drugs. 2007, 16: 1111–1120. [CrossRef]
- Gryder, B.E.; Sodji, Q.H.; Oyelere, A.K. Targeted cancer therapy: giving histone deacetylase inhibitors all they need to succeed. Future Med Chem. 2012, 4:505-24. Erratum in: Future Med Chem. 2012 Jun;4(10):1369-70. [CrossRef]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int J Mol Sci. 2017, 18:1414. [CrossRef]
- Whittaker, S.J.; Demierre, M.F.; Kim, E.J.; Rook, A.H.; Lerner, A.; Duvic, M.; Scarisbrick, J.; Reddy, S.; Robak, T.; Becker, J.C.; Samtsov, A.; McCulloch, W.; Kim, Y.H. Final results from a multicenter, international, pivotal study of romidepsin in refractory cutaneous T-cell lymphoma. J Clin Oncol. 2010, 28:4485-91. [CrossRef]
- Wang, P.; Wang, Z.; Liu, J. Role of HDACs in normal and malignant hematopoiesis. Mol Cancer. 2020, 19, 5. [CrossRef] [PubMed]
- Garnock-Jones, K.P. Panobinostat: First Global Approval. Drugs. 2015, 75:695–704. [CrossRef]
- Di Gennaro, E.; Bruzzese, F.; Pepe, S.; Leone, A.; Delrio, P.; Subbarayan, P.R.; Avallone, A.; Budillon, A. Modulation of thymidilate synthase and p53 expression by HDAC inhibitor vorinostat resulted in synergistic antitumor effect in combination with 5FU or raltitrexed. Cancer Biol Ther. 2009, 8:782-91. [CrossRef]
- Götze, S.; Coersmeyer, M.; Müller, O.; Sievers, S. Histone deacetylase inhibitors induce attenuation of Wnt signaling and TCF7L2 depletion in colorectal carcinoma cells. Int J Oncol. 2014, 45:1715-23. [CrossRef]
- Munster, P.N.; Marchion, D.; Thomas, S.; Egorin, M.; Minton, S.; Springett, G.; Lee, J.H.; Simon, G.; Chiappori, A.; Sullivan, D.; Daud, A. Phase I trial of vorinostat and doxorubicin in solid tumours: histone deacetylase 2 expression as a predictive marker. Br J Cancer. 2009, 101:1044-50. [CrossRef]
- Wilson, P.M.; El-Khoueiry, A.; Iqbal, S.; Fazzone, W.; LaBonte, M.J.; Groshen, S.; Yang, D.; Danenberg, K.D.; Cole, S.; Kornacki, M.; Ladner, R.D.; Lenz, H.J. A phase I/II trial of vorinostat in combination with 5-fluorouracil in patients with metastatic colorectal cancer who previously failed 5-FU-based chemotherapy. Cancer Chemother Pharmacol. 2010, 65:979-88. [CrossRef]
- Yang, D.; Torres, C.M.; Bardhan, K.; Zimmerman, M.; Mcgaha, T.L.; Liu, K. Decitabine and vorinostat cooperate to sensitize colon carcinoma cells to Fas ligand-induced apoptosis in vitro and tumor suppression in vivo. J. Immunol. 2012, 188:4441–4449. [CrossRef]
- Kaliszczak, M.; Trousil, S.; Åberg, O.; Perumal, M.; Nguyen, Q.D.; Aboagye, E.O. A novel small molecule hydroxamate preferentially inhibits HDAC6 activity and tumour growth. Br J Cancer. 2013,108:342-50. [CrossRef]
- Santo, L.; Hideshima, T.; Kung, A.L.; Tseng, J.C.; Tamang, D.; Yang, M.; Jarpe, M.; van Duzer, J.H.; Mazitschek, R.; Ogier, W.C.; Cirstea, D.; Rodig, S.; Eda, H.; Scullen, T.; Canavese, M.; Bradner, J.; Anderson, K.C.; Jones, S.S.; Raje, N. Preclinical activity, pharmacodynamic, and pharmacokinetic properties of a selective HDAC6 inhibitor, ACY-1215, in combination with bortezomib in multiple myeloma. Blood. 2012, 119:2579-89. [CrossRef]
- Amengual, J.E.; Lue, J.K.; Ma, H.; Lichtenstein, R.; Shah, B.; Cremers, S.; Jones, S.; Sawas, A. First-in-Class Selective HDAC6 Inhibitor (ACY-1215) Has a Highly Favorable Safety Profile in Patients with Relapsed and Refractory Lymphoma. Oncologist. 2021, 26:184-e366. [CrossRef]
- Li, J.; Yu, M.; Fu, S.; Liu, D.; Tan, Y. Role of Selective Histone Deacetylase 6 Inhibitor ACY-1215 in Cancer and Other Human Diseases. Front Pharmacol. 2022, 16;13:907981. doi: 10.3389/fphar.2022.907981. Erratum in: Front Pharmacol. 2022 13:1117936. Erratum in: Front Pharmacol. 2022 13:1117936. [CrossRef]
- Kim, C.; Lee, S.; Kim, D.; Lee, D.S.; Lee, E.; Yoo, C.; Kim, K.P. Blockade of GRP78 Translocation to the Cell Surface by HDAC6 Inhibition Suppresses Proliferation of Cholangiocarcinoma Cells. Anticancer Res. 2022, 42:471-482. [CrossRef]
- Lee, D.H.; Won, H.R.; Ryu, H.W.; Han, J.M.; Kwon, S.H. The HDAC6 inhibitor ACY-1215 enhances the anticancer activity of oxaliplatin in colorectal cancer cells. Int J Oncol. 2018, 53:844-854. [CrossRef]
- Tan, Y.; Zhang, S.; Zhu, H.; Chu, Y.; Zhou, H.; Liu, D.; Huo, J. Histone deacetylase 6 selective inhibitor ACY1215 inhibits cell proliferation and enhances the chemotherapeutic effect of 5-fluorouracil in HCT116 cells. Ann Transl Med. 2019, 7:2. [CrossRef]
- Ryu, H.W.; Shin, D.H.; Lee, D.H.; Won, H.R.; Kwon, S.H. A potent hydroxamic acid-based, small-molecule inhibitor A452 preferentially inhibits HDAC6 activity and induces cytotoxicity toward cancer cells irrespective of p53 status. Carcinogenesis. 2018, 39:72-83. [CrossRef]
- Won, H.R.; Ryu, H.W.; Shin, D.H.; Yeon, S.K.; Lee, D.H.; Kwon, S.H. A452, an HDAC6-selective inhibitor, synergistically enhances the anticancer activity of chemotherapeutic agents in colorectal cancer cells. Mol Carcinog. 2018, 57:1383-1395. [CrossRef]
- Ojha, R.; Huang, H.L.; HuangFu, W.C.; Wu, Y.W.; Nepali, K.; Lai, M.J.; Su, C.J.; Sung, T.Y.; Chen, Y.L.; Pan, S.L.; Liou, J.P. 1-Aroylindoline-hydroxamic acids as anticancer agents, inhibitors of HSP90 and HDAC. Eur J Med Chem. 2018, 150:667-677. [CrossRef]
- North, B.J.; Almeciga-Pinto, I.; Tamang, D.; Yang, M.; Jones, S.S.; Quayle, S.N. Enhancement of pomalidomide anti-tumor response with ACY-241, a selective HDAC6 inhibitor. PLoS One. 2017,12:e0173507. [CrossRef]
- Huang, P.; Almeciga-Pinto, I.; Jarpe, M.; van Duzer, J.H.; Mazitschek, R.; Yang, M.; Jones, S.S.; Quayle, S.N. Selective HDAC inhibition by ACY-241 enhances the activity of paclitaxel in solid tumor models. Oncotarget. 2017, 8:2694-2707. [CrossRef]
- Gordon, M.S.; Shapiro, G.I.; Sarantopoulos, J.; Juric, D.; Lu, B.; Zarotiadou, A.; Connarn, J.N.; Le Bruchec, Y.; Dumitru, C.D.; Harvey, R.D. Phase Ib Study of the Histone Deacetylase 6 Inhibitor Citarinostat in Combination With Paclitaxel in Patients With Advanced Solid Tumors. Front Oncol. 2022, 11:786120. [CrossRef]
- Banik, D.; Noonepalle, S.; Hadley, M.; Palmer, E.; Gracia-Hernandez, M.; Zevallos-Delgado, C.; Manhas, N.; Simonyan, H.; Young, C.N.; Popratiloff, A.; Chiappinelli, K.B.; Fernandes, R.; Sotomayor, E.M.; Villagra, A. HDAC6 Plays a Noncanonical Role in the Regulation of Antitumor Immune Responses, Dissemination, and Invasiveness of Breast Cancer. Cancer Res. 2020,80:3649-3662. [CrossRef]
- Chen, M.C.; Lin, Y.C.; Liao, Y.H.; Liou, J.P.; Chen, C.H. MPT0G612, a Novel HDAC6 Inhibitor, Induces Apoptosis and Suppresses IFN-γ-Induced Programmed Death-Ligand 1 in Human Colorectal Carcinoma Cells. Cancers (Basel). 2019,11:1617. [CrossRef]
- Forsythe, N.; Refaat, A.; Javadi, A.; Khawaja, H.; Weir, J.-A.; Emam, H.; Allen, W.L.; Burkamp, F.; Popovici, V.; Jithesh, P.V.; Isella, C.; Labonte, M.J.; Mills, I.G.; Johnston, P.G.; Van Schaeybroeck, S. The Unfolded Protein Response: A Novel Therapeutic Target for Poor Prognostic BRAF Mutant Colorectal Cancer. Mol. Cancer Ther. 2018, 17:1280–1290. [CrossRef]
- Kaliszczak, M.; van Hechanova, E.; Li, Y.; Alsadah, H.; Parzych, K.; Auner, H.W.; Aboagye, E.O. The HDAC6 inhibitor C1A modulates autophagy substrates in diverse cancer cells and induces cell death. Br J Cancer. 2018, 119:1278-1287. [CrossRef]
- Fransén, K.; Klintenäs, M.; Osterström, A.; Dimberg, J.; Monstein, H.J.; Söderkvist, P. Mutation analysis of the BRAF, ARAF and RAF-1 genes in human colorectal adenocarcinomas. Carcinogenesis. 2004, 25:527-33. [CrossRef]
- Ros, J.; Baraibar, I.; Sardo, E.; Mulet, N.; Salvà, F.; Argilés, G.; Martini, G.; Ciardiello, D.; Cuadra, J.L.; Tabernero, J.; Élez, E. BRAF, MEK and EGFR inhibition as treatment strategies in BRAF V600E metastatic colorectal cancer. Ther Adv Med Oncol. 2021, 13:1758835921992974. [CrossRef]
- Carson, R.; Celtikci, B.; Fenning, C.; Javadi, A.; Crawford, N.; Carbonell, L.P.; Lawler, M.; Longley, D.B.; Johnston, P.G.; Van Schaeybroeck, S. HDAC Inhibition Overcomes Acute Resistance to MEK Inhibition in BRAF-Mutant Colorectal Cancer by Downregulation of c-FLIPL. Clin Cancer Res. 2015, 21:3230-3240. [CrossRef]
- Lai, F.; Guo, S.T.; Jin, L.; Jiang, C.C.; Wang, C.Y.; Croft, A.; Chi, M.N.; Tseng, H.Y.; Farrelly, M.; Atmadibrata, B.; Norman, J.; Liu, T.; Hersey, P.; Zhang, X.D. Cotargeting histone deacetylases and oncogenic BRAF synergistically kills human melanoma cells by necrosis independently of RIPK1 and RIPK3. Cell Death Dis. 2013, 4:e655. [CrossRef]
- Madorsky Rowdo, F.P.; Barón, A.; Gallagher, S.J.; Hersey, P.; Emran, A.A.; Von Euw, E.M.; Barrio, M.M.; Mordoh, J. Epigenetic inhibitors eliminate senescent melanoma BRAFV600E cells that survive long-term BRAF inhibition. Int J Oncol. 2020, 56:1429-1441. [CrossRef]
- Fu, H.; Cheng, L.; Jin, Y.; Cheng, L.; Liu, M.; Chen, L. MAPK Inhibitors Enhance HDAC Inhibitor-Induced Redifferentiation in Papillary Thyroid Cancer Cells Harboring BRAFV600E: An In Vitro Study. Mol Ther Oncolytics. 2019, 12:235-245. [CrossRef]
- Groselj, B.; Sharma, N.L.; Hamdy, F.C.; Kerr, M.; Kiltie, A.E. Histone deacetylase inhibitors as radiosensitisers: effects on DNA damage signalling and repair. Br J Cancer. 2013,108:748-754. [CrossRef]
- Suraweera, A.; O'Byrne, K.J.; Richard, D.J. Combination Therapy With Histone Deacetylase Inhibitors (HDACi) for the Treatment of Cancer: Achieving the Full Therapeutic Potential of HDACi. Front Oncol. 2018, 8:92. [CrossRef]
- Noonepalle, S.K.R.; Grindrod, S.; Aghdam, N.; Li, X.; Gracia-Hernandez, M.; Zevallos-Delgado, C.; Jung, M.; Villagra, A.; Dritschilo, A. Radiation therapy-induced immune response enhanced by selective HDAC6 Inhibition. Mol Cancer Ther. 2023, MCT-23-0215. [CrossRef]
- Song, W.; Tai, Y.T.; Tian, Z.; Hideshima, T.; Chauhan, D.; Nanjappa, P.; Exley, M.A.; Anderson, K.C.; Munshi, N.C. HDAC inhibition by LBH589 affects the phenotype and function of human myeloid dendritic cells. Leukemia. 2011, 25:161-8. [CrossRef]
- Reddy, P.; Sun, Y.; Toubai, T.; Duran-Struuck, R.; Clouthier, S.G.; Weisiger, E.; Maeda, Y.; Tawara, I.; Krijanovski, O.; Gatza, E.; Liu, C.; Malter, C.; Mascagni, P.; Dinarello, C.A.; Ferrara, J.L. Histone deacetylase inhibition modulates indoleamine 2,3-dioxygenase-dependent DC functions and regulates experimental graft-versus-host disease in mice. J. Clin. Invest. 2008, 118:2562–2573. [CrossRef]
- Sun, Y.; Chin, Y.E.; Weisiger, E.; Malter, C.; Tawara, I.; Toubai, T.; Gatza, E.; Mascagni, P.; Dinarello, C.A.; Reddy, P. Cutting edge: Negative regulation of dendritic cells through acetylation of the nonhistone protein stat-3. J. Immunol. 2009, 182:5899–5903. [CrossRef]
- Bae, J.; Hideshima, T.; Tai, Y.T; Song, Y.; Richardson, P.; Raje, N.; Munshi, N.C.; Anderson, K.C. Histone deacetylase (HDAC) inhibitor ACY241 enhances anti-tumor activities of antigen-specific central memory cytotoxic T lymphocytes against multiple myeloma and solid tumors. Leukemia. 2018, 32:1932-1947. [CrossRef]
- Vishwakarma, S.; Iyer, L.R.; Muley, M.; Singh, P.K.; Shastry, A.; Saxena, A.; Kulathingal, J.; Vijaykanth, G.; Raghul, J.; Rajesh, N.; Rathinasamy, S.; Kachhadia, V.; Kilambi, N.; Rajgopal, S.; Balasubramanian, G.; Narayanan, S. Tubastatin, a selective histone deacetylase 6 inhibitor shows anti-inflammatory and anti-rheumatic effects. Int Immunopharmacol. 2013, 16:72-8. [CrossRef]
- Skov, S.; Rieneck, K.; Bovin, L.F.; Skak, K.; Tomra, S.; Michelsen, B.K.; Ødum, N. Histone deacetylase inhibitors: a new class of immunosuppressors targeting a novel signal pathway essential for CD154 expression. Blood. 2003, 101:1430-8. [CrossRef]
- Peart, M.J.; Smyth, G.K.; van Laar, R.K.; Bowtell, D.D.; Richon, V.M.; Marks, P.A.; Holloway, A.J.; Johnstone, R.W. Identification and functional significance of genes regulated by structurally different histone deacetylase inhibitors. Proc Natl Acad Sci U S A. 2005, 102:3697-702. [CrossRef]
- Kelly-Sell, M.J.; Kim, Y.H.; Straus, S.; Benoit, B.; Harrison, C.; Sutherland, K.; Armstrong, R.; Weng, W.K.; Showe, L.C.; Wysocka, M.; Rook, A.H. The histone deacetylase inhibitor, romidepsin, suppresses cellular immune functions of cutaneous T-cell lymphoma patients. Am J Hematol. 2012, 87:354-60. [CrossRef]
- Cao, K.; Wang, G.; Li, W.; Zhang, L.; Wang, R.; Huang, Y.; Du, L.; Jiang, J.; Wu, C.; He, X.; Roberts, A.I.; Li, F.; Rabson, A.B.; Wang, Y.; Shi, Y. Histone deacetylase inhibitors prevent activation-induced cell death and promote antitumor immunity. Oncogene, 2015, 34:5960–5970. [CrossRef]
- Laino, A.S.; Betts, B.C.; Veerapathran, A.; Dolgalev, I.; Sarnaik, A.; Quayle, S.N.; Jones, S.S.; Weber, J.S.; Woods, D.M. HDAC6 selective inhibition of melanoma patient T-cells augments anti-tumor characteristics. J Immunother Cancer. 2019, 7:33. [CrossRef]
- Wang, H.F.; Ning, F.; Liu, Z.C.; Wu, L.; Li, Z.Q.; Qi, Y.F.; Zhang, G.; Wang, H.S.; Cai, S.H.; Du, J. Histone deacetylase inhibitors deplete myeloid-derived suppressor cells induced by 4T1 mammary tumors in vivo and in vitro. Cancer Immunol Immunother. 2017, 66:355-366. [CrossRef]
- Moran, B.; Davern, M.; Reynolds, J.V.; Donlon, N.E.; Lysaght, J. The impact of histone deacetylase inhibitors on immune cells and implications for cancer therapy. Cancer Lett. 2023, 559:216121. [CrossRef]
- Afolabi, L.O.; Bi, J.; Li, X.; Adeshakin, A.O.; Adeshakin, F.O.; Wu, H.; Yan, D.; Chen, L.; Wan, X. Synergistic Tumor Cytolysis by NK Cells in Combination With a Pan-HDAC Inhibitor, Panobinostat. Front Immunol. 2021, 12:701671. [CrossRef]
- Berghuis, D.; Schilham, M.W.; Vos, H.I.; Santos, S.J.; Kloess, S.; Buddingh', E.P.; Egeler, R.M.; Hogendoorn, P.C.; Lankester, A.C. Histone deacetylase inhibitors enhance expression of NKG2D ligands in Ewing sarcoma osteosarcoma and sensitize for natural killer cell-mediated cytolysis. Clin Sarcoma Res. 2012,2:8. [CrossRef]
- López-Soto, A.; Folgueras, A.R.; Seto, E.; Gonzalez, S. HDAC3 represses the expression of NKG2D ligands ULBPs in epithelial tumour cells: potential implications for the immunosurveillance of cancer. Oncogene. 2009, 28:2370-82. [CrossRef]
- Zheng, H.; Zhao, W.; Yan, C.; Watson, C.C.; Massengill, M.; Xie, M.; Massengill, C.; Noyes, D.R.; Martinez, G.V.; Afzal, R.; Chen, Z.; Ren, X.; Antonia, S.J.; Haura, E.B.; Ruffell, B.; Beg, A.A. HDAC Inhibitors Enhance T-Cell Chemokine Expression and Augment Response to PD-1 Immunotherapy in Lung Adenocarcinoma. Clin Cancer Res. 2016, 22:4119-32. [CrossRef]
- Hashimoto, A.; Fukumoto, T.; Zhang, R.; Gabrilovich, D. Selective targeting of different populations of myeloid-derived suppressor cells by histone deacetylase inhibitors. Cancer Immunol Immunother. 2020, 69:1929-1936. [CrossRef]
- Li, X.; Su, X.; Liu, R.; Pan, Y.; Fang, J.; Cao, L.; Feng, C.; Shang, Q.; Chen, Y.; Shao, C.; Shi, Y. HDAC inhibition potentiates anti-tumor activity of macrophages and enhances anti-PD-L1-mediated tumor suppression. Oncogene. 2021, 40:1836-1850. [CrossRef]
- Romeo, E.; Caserta, C. A.; Rumio, C.; Marcucci, F. The vicious crosstalk between tumor cells with an EMT phenotype and cells of the immune system. Cells. 2019, 8:460. [CrossRef]
- Beldi-Ferchiou, A.; Caillat-Zucman, S. Control of NK Cell Activation by Immune Checkpoint Molecules. Int. J. Mol. Sci. 2017, 18:2129. [CrossRef]
- Gao, Y.; Yang, J.; Cai, Y.; Fu, S.; Zhang, N.; Fu, X.; Li, L. IFN-gammamediated inhibition of lung cancer correlates with PD-L1 expression and is regulated by PI3K-AKT signaling. Int. J. Cancer. 2018, 143:931–943. [CrossRef]
- Ju, X.; Zhang, H.; Zhou, Z.; Chen, M.; Wang, Q. Tumor-associated macrophages induce PD-L1 expression in gastric cancer cells through IL-6 and TNF-a signaling. Exp. Cell Res. 2020, 396:112315. [CrossRef]
- Li, P.; Huang, T.; Zou, Q.; Liu, D.; Wang, Y.; Tan, X.; Wei, Y.; Qiu, H. FGFR2 promotes expression of PD-L1 in colorectal cancer via the JAK/STAT3 signaling pathway. J. Immunol. 2019, 202:3065–3075. [CrossRef]
- Wang, H.B.; Yao, H.; Li, C. S.; Liang, L. X.; Zhang, Y.; Chen, Y. X.; Fang, J.Y.; Xu, J. Rise of PD-L1 expression during metastasis of colorectal cancer: implications for immunotherapy. J. Dig. Dis. 2017, 18:574–581. [CrossRef]
- Chen, Q. Y.; Chen, Y. X.; Han, Q. Y.; Zhang, J. G.; Zhou, W. J.; Zhang, X.; Ye, Y.H.; Yan, W.H.; Lin, A. Prognostic significance of immune checkpoints HLA-G/ILT-2/4 and PD-L1 in colorectal cancer. Front. Immunol. 2021, 12:679090. [CrossRef]
- Wang, H.; Yao, H.; Li, C.; Liang, L.; Zhang, Y.; Shi, H.; Zhou, C.; Chen, Y.; Fang, J.Y.; Xu, J. PD-L2 expression in colorectal cancer: independent prognostic effect and targetability by deglycosylation. Oncoimmunology, 2017, 6:e1327494. [CrossRef]
- Zou, W.; Wolchok, J. D.; Chen, L. PD-L1 (B7-H1) and PD- 1 pathway blockade for cancer therapy: mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 2016, 8:328rv4. [CrossRef]
- Konjević, G.M.; Vuletić, A.M.; Mirjačić Martinović, K.M.; Larsen, A.K.; Jurišić, V.B. The role of cytokines in the regulation of NK cells in the tumor environment. Cytokine. 2019,117:30-40. [CrossRef]
- Lienlaf, M.; Perez-Villarroel, P.; Knox, T.; Pabon, M.; Sahakian, E.; Powers, J.; Woan, K. V.; Lee, C.; Cheng, F.; Deng, S.; Smalley, K. S. M.; Montecino, M.; Kozikowski, A.; Pinilla-Ibarz, J.; Sarnaik, A.; Seto, E.; Weber, J.; Sotomayor, E. M.; Villagra, A. Essential role of HDAC6 in the regulation of PD-L1 in melanoma. Mol Oncol. 2016, 10:735-750. [CrossRef]
- Keremu, A.; Aimaiti, A.; Liang, Z.; Zou, X. Role of the HDAC6/STAT3 pathway in regulating PD-L1 expression in osteosarcoma cell lines. Cancer Chemother Pharmacol. 2019, 83:255-264. [CrossRef]
- Woods, D.M.; Sodré, A.L.; Villagra, A.; Sarnaik, A.; Sotomayor, E.M.; Weber, J. HDAC Inhibition Upregulates PD-1 Ligands in Melanoma and Augments Immunotherapy with PD-1 Blockade. Cancer Immunol Res. 2015, 3:1375-85. [CrossRef]
- Woods, D.M.; Woan, K.; Cheng, F.; Wang, H.; Perez-Villarroel, P.; Lee, C.; Lienlaf, M.; Atadja, P.; Seto, E.; Weber, J.; Sotomayor, E.M.; Villagra, A. The antimelanoma activity of the histone deacetylase inhibitor panobinostat (LBH589) is mediated by direct tumor cytotoxicity and increased tumor immunogenicity. Melanoma Res. 2013,23:341-8. [CrossRef]
- Ieranò, C.; Righelli, D.; D'Alterio, C.; Napolitano, M.; Portella, L.; Rea, G.; Auletta, F.; Santagata, S.; Trotta, A.M.; Guardascione, G.; Liotti, F.; Prevete, N.; Maiolino, P.; Luciano, A.; Barbieri, A.; Di Mauro, A.; Roma, C.; Esposito Abate, R.; Tatangelo, F.; Pacelli, R.; Normanno, N.; Melillo, R.M.; Scala, S. In PD-1+ human colon cancer cells NIVOLUMAB promotes survival and could protect tumor cells from conventional therapies. J Immunother Cancer. 2022, 10:e004032. [CrossRef]
- Kleffel, S.; Posch, C.; Barthel, S.R.; Mueller, H.; Schlapbach, C.; Guenova, E.; Elco, C.P.; Lee, N.; Juneja, V.R.; Zhan, Q.; Lian, C.G.; Thomi, R; Hoetzenecker, W.; Cozzio, A.; Dummer, R.; Mihm, M.C., Jr.; Flaherty., K.T.; Frank, M.H.; Murphy, G.F.; Sharpe, A.H.; Kupper, T.S.; Schatton, T. Melanoma Cell-Intrinsic PD-1 Receptor Functions Promote Tumor Growth. Cell. 2015, 162:1242-56. [CrossRef]
- Cao, Z.; Kon, N.; Liu, Y.; Xu, W.; Wen, J.; Yao, H.; Zhang, M.; Wu, Z.; Yan, X.; Zhu, W.G.; Gu, W.; Wang, D. An unexpected role for p53 in regulating cancer cell-intrinsic PD-1 by acetylation. Sci Adv. 2021,7:eabf4148. [CrossRef]
- Zhou, E.; Huang, Q.; Wang, J.; Fang, C.; Yang, L.; Zhu, M.; Chen, J.; Chen, L.; Dong, M. Up-regulation of Tim-3 is associated with poor prognosis of patients with colon cancer. Int J Clin Exp Pathol. 2015, 8:8018-27.
- Hontecillas-Prieto, L.; Flores-Campos, R.; Silver, A.; de Álava, E.; Hajji, N.; García-Domínguez, D.J. Synergistic Enhancement of Cancer Therapy Using HDAC Inhibitors: Opportunity for Clinical Trials. Front Genet. 2020, 11:578011. [CrossRef]
- Fukumoto, T.; Fatkhutdinov, N.; Zundell, J.A.; Tcyganov, E.N.; Nacarelli, T.; Karakashev, S.; Wu, S.; Liu, Q.; Gabrilovich, D.I.; Zhang, R. HDAC6 Inhibition Synergizes with Anti-PD-L1 Therapy in ARID1A-Inactivated Ovarian Cancer. Cancer Res. 2019,79:5482-5489. [CrossRef]
- Ray, A.; Das, D.S.; Song, Y.; Hideshima, T.; Tai, Y.T.; Chauhan, D.; Anderson, K.C. Combination of a novel HDAC6 inhibitor ACY-241 and anti-PD-L1 antibody enhances anti-tumor immunity and cytotoxicity in multiple myeloma. Leukemia. 2018, 32:843-846. [CrossRef]
- Gray, J.E.; Saltos, A.; Tanvetyanon, T.; Haura, E.B.; Creelan, B.; Antonia, S.J.; Shafique, M.; Zheng, H.; Dai, W.; Saller, J.J.; Chen, Z.; Tchekmedyian, N.; Goas, K.; Thapa, R.; Boyle, T.A.; Chen, D.T.; Beg, A.A. Phase I/Ib Study of Pembrolizumab Plus Vorinostat in Advanced/Metastatic Non-Small Cell Lung Cancer. Clin Cancer Res. 2019, 25:6623-6632. [CrossRef]
- Terranova-Barberio, M.; Pawlowska, N.; Dhawan, M.; Moasser, M.; Chien, A.J.; Melisko, M.E.; Rugo, H.; Rahimi, R.; Deal, T.; Daud, A.; Rosenblum, M.D.; Thomas, S.; Munster, P.N. Exhausted T cell signature predicts immunotherapy response in ER-positive breast cancer. Nat Commun. 2020,11:3584. [CrossRef]
- Rodriguez, C.P.; Wu, Q.V.; Voutsinas, J.; Fromm, J.R.; Jiang, X.; Pillarisetty, V.G.; Lee, S.M.; Santana-Davila, R.; Goulart, B.; Baik, C.S.; Chow, L.Q.M.; Eaton, K.; Martins, R. A Phase II Trial of Pembrolizumab and Vorinostat in Recurrent Metastatic Head and Neck Squamous Cell Carcinomas and Salivary Gland Cancer. Clin Cancer Res. 2020, 26:837-845. [CrossRef]
- Awad, M.M.; Le Bruchec, Y.; Lu, B.; Ye, J.; Miller, J.; Lizotte, P.H., Cavanaugh; Rode, A.J.; Dumitru, C.D.; Spira, A. Selective Histone Deacetylase Inhibitor ACY-241 (Citarinostat) Plus Nivolumab in Advanced Non-Small Cell Lung Cancer: Results From a Phase Ib Study. Front Oncol. 2021, 11:696512. [CrossRef]
- Saunders, M.P.; Graham, J.; Cunningham, D.; Plummer, R.; Church, D.; Kerr, R.; Cook, S.; Zheng, S.; La Thangue, N.; Kerr, D. CXD101 and nivolumab in patients with metastatic microsatellite-stable colorectal cancer (CAROSELL): a multicentre, open-label, single-arm, phase II trial. ESMO Open. 2022, 7:100594. [CrossRef]
- Borcoman, E.; Kamal, M.; Marret, G.; Dupain, C.; Castel-Ajgal, Z.; Le Tourneau, C. HDAC Inhibition to Prime Immune Checkpoint Inhibitors. Cancers (Basel). 2021, 14:66. [CrossRef]
- Jenke, R.; Reßing, N.; Hansen, F.K.; Aigner, A.; Büch, T. Anticancer Therapy with HDAC Inhibitors: Mechanism-Based Combination Strategies and Future Perspectives. Cancers (Basel). 2021, 13:634. [CrossRef]
- Ruzic, D.; Djoković, N.; Srdić-Rajić, T.; Echeverria, C.; Nikolic, K.; Santibanez, J.F. Targeting Histone Deacetylases: Opportunities for Cancer Treatment and Chemoprevention. Pharmaceutics. 2022, 14:209. [CrossRef]
- Liu, T.; Wan, Y.; Xiao, Y.; Xia, C.; Duan, G. Dual-Target Inhibitors Based on HDACs: Novel Antitumor Agents for Cancer Therapy. J. Med. Chem. 2020. [CrossRef]
- Yang, E.G.; Mustafa, N.; Tan, E.C.; Poulsen, A.; Ramanujulu, P.M.; Chng, W.J.; Yen, J.J.; Dymock, B.W. Design and Synthesis of Janus Kinase 2 (JAK2) and Histone Deacetlyase (HDAC) Bispecific Inhibitors Based on Pacritinib and Evidence of Dual Pathway Inhibition in Hematological Cell Lines. J Med Chem. 2016, 59:8233-62. [CrossRef]
- Li, Y.; Huang, Y.; Cheng, H.; Xu, F.; Qi, R.; Dai, B.; Yang, Y.; Tu, Z.; Peng, L.; Zhang, Z. Discovery of BRAF/HDAC Dual Inhibitors Suppressing Proliferation of Human Colorectal Cancer Cells. Front Chem. 2022, 10:910353. [CrossRef]
- Yang, K.; Song, Y.; Xie, H.; Wu, H.; Wu, Y.T.; Leisten, E.D.; Tang, W. Development of the first small molecule histone deacetylase 6 (HDAC6) degraders. Bioorg Med Chem Lett. 2018, 28:2493-2497. [CrossRef]
- Wu, H.; Yang, K.; Zhang, Z.; Leisten, E.; LI, Z.; Xie, H.; Tang, W. Development of Multi-Functional Histone Deacetylase 6 Degraders with Potent Anti-Myeloma Activity. Journal of Medicinal Chemistry. 2019. [CrossRef] [PubMed]
- Yang, H.; Lv, W.; He, M.; Deng, H.; Li, H.; Wu, W.; Rao, Y. Plasticity in Designing PROTACs for Selective and Potent Degradation of HDAC6. Chem. Commun. 2019, 55, 14848–1485. [CrossRef] [PubMed]
Figure 1.
Functional domains of HDAC6: NLS- nuclear localization sequence, NES- nuclear export sequence; two catalytic domains (CD1 and CD2), DMBS- dynein motor binding sequence, ZnF-UBP- zinc finger ubiquitin binding domain, (U- ubiquitin). Acetylation of NLS by p300 inhibits HDAC6 interaction with importin-α. Ras/Raf/MEK/ERK, EFFR, GRK2, CK2, GSK3β signaling phosphorylate and activate HDAC6.
Figure 1.
Functional domains of HDAC6: NLS- nuclear localization sequence, NES- nuclear export sequence; two catalytic domains (CD1 and CD2), DMBS- dynein motor binding sequence, ZnF-UBP- zinc finger ubiquitin binding domain, (U- ubiquitin). Acetylation of NLS by p300 inhibits HDAC6 interaction with importin-α. Ras/Raf/MEK/ERK, EFFR, GRK2, CK2, GSK3β signaling phosphorylate and activate HDAC6.
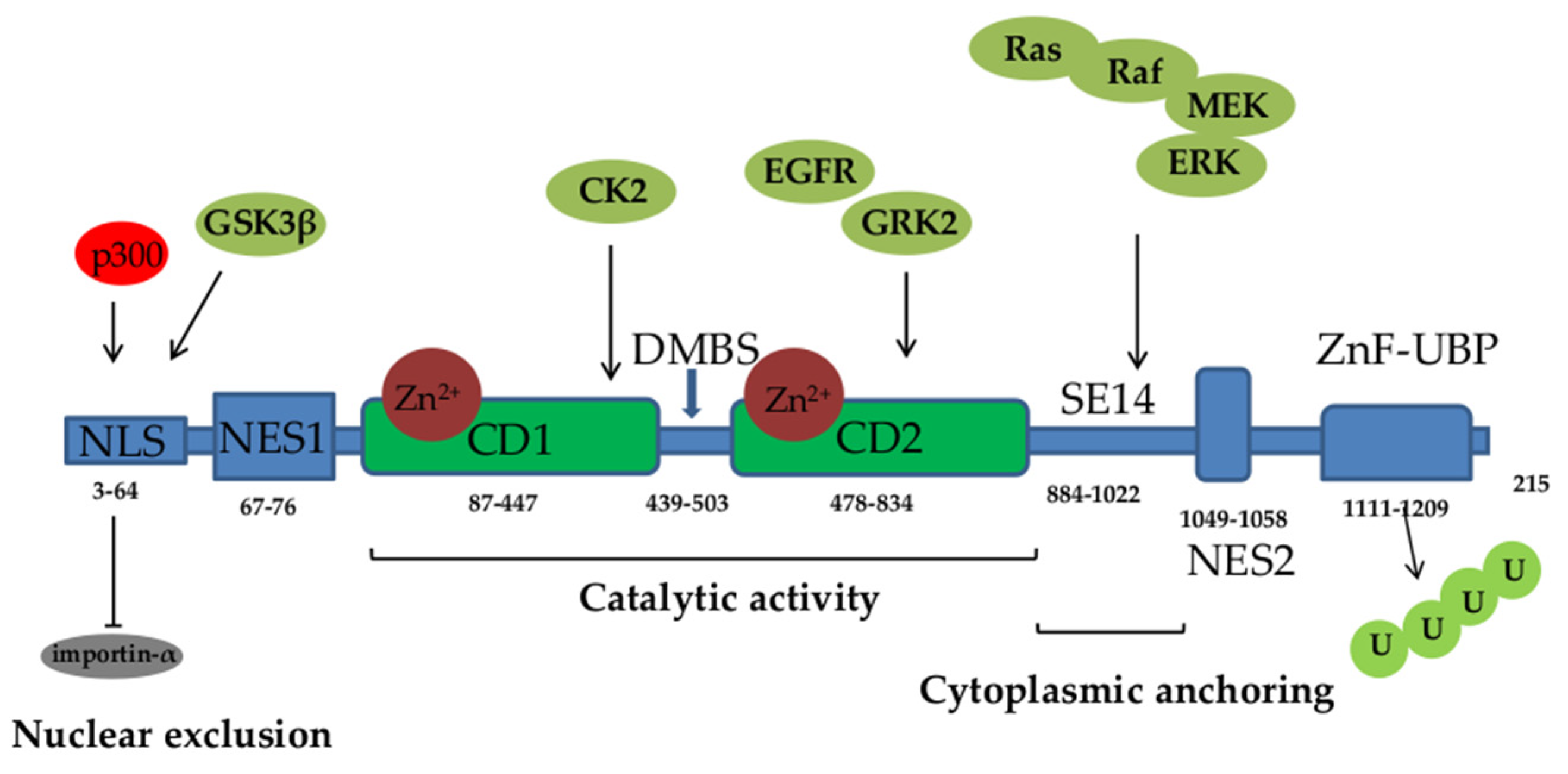

Table 1.
HDAC6 substrates and physiological functions of HDAC6-mediated deacetylation.
| Protein | Localization | Function | Reference |
|---|---|---|---|
| α-tubulin | Cytoplasm | Microtubule disassembly, increases cell motility | [21], [22], [23] |
| Cortactin | Cytoplasm | Actin polymerization and branching, increases cell motility | [25] |
| TFEB | Cytoplasm | Autophagy | [38] |
| FOXO1 | Cytoplasm | Autophagy | [39] |
| Hsp90 | Cytoplasm | Degradation of misfolded proteins | [49] |
| GRP78 | Cytoplasm and nucleus | ER stress regulation, tumor progression via secretion of exosomes | [58] |
| NF-κB | Nucleus | Transcription of genes for NLRP3, pro-IL-1β, pro-IL-18, inflammasome activity | [12] |
| P53 | Cytoplasm | Cell cycle progression, inhibition of apoptosis, induced autophagy via upregulation of Beclin-1 | [65], [66], [67] |
| Ku70 | Cytoplasm | Suppression of apoptosis | [9], [77] |
| Survivin | Nucleus | Suppression of apoptosis | [78] |
| Peroxidins | Cytoplasm and nucleus | Antioxidant activity | [61] |
| Smad3 | Cytoplasm | Downregulation of E-cadherin expression, EMT | [97], [98] |
| β-catenin | Cytoplasm | Translocation into nucleus and tumor cell invasion | [115] |
| STAT3 | Cytoplasm | Activation of JAK/STAT3 signaling and inflammatory responses | [118] |
| TAK1 | Cytoplasm | Activation of ADAM17 MMP enhances sIL-6R release and M2 macrophage differentiation | [123] |
| ERK1 | Cytoplasm | Activation of ERK1, proliferation, survival, and increased cell motility | [83] |
| AKT | Cytoplasm | Activation of AKT pathway, cell migration | [85] |
*EMT- epithelial-mesenchymal transition; sIL-6R soluble IL-6 receptor; MMP- matrix metalloproteinase.
Table 2.
HDAC6 substrates and physiological functions of HDAC6-mediated deacetylation.
| HDAC6 domain | Protein | Function | References |
|---|---|---|---|
| DMBS | Dynein/ p150glued | Aggresome formation and autophagy | [21] |
| ZnF-UBP | Ubiquitin chain | Protein degradation | [33] |
| Not defined | TRIM50 | E3-Ubiquitin ligase activity promotes recruitment of polyU proteins to aggresome and degradation | [33] |
| Not defined | P97/VCP | Dissociation of HDAC6 and polyU protein and protein delivery to proteasomes | [29], [36] |
| Not defined | HSF1 | Release of HSF1 and transcription of genes for Hsp90 and Hsp70 molecular chaperons | [48], [51] |
| Not defined | Runx2 | Proliferation, inhibition of apoptosis | [80] |
| Not defined | STAT3 | Activation of JAK/STAT3 signaling and inflammatory responses | [118], [205] |
Table 3.
HDAC6 inhibiting agents and their effects in CRC.
| HDAC6 inhibitor | Effect | Reference |
|---|---|---|
| Vorinostat (SAHA) | Inhibition of proliferation, downregulation of mutated p53, upregulation of wtp53, inhibition of HDAC6-Hsp90 axis, apoptosis | [71], [77], [145] |
| Vorinostat + 5-fluorouracil | Inhibition of proliferation, downregulation of mutated p53, | [145], [148] |
| Vorinostat + decitabine | Inhibition of proliferation and migration, apoptosis, decreased pMEK and pERK | [149]. |
| Vorinostat + trametinib | Inhibition of proliferation, apoptosis | [169] |
| Vorinostat + trichostatin A | Attenuation of Wnt signaling, apoptosis | [146] |
| Trichostatin A | Increased acetylation of Ku70 and apoptosis by releasing Bax | [76] |
| ACY-1215 | Inhibition of MAPK/ERK and PI3K/AKT signaling; acetylated tubulin, cortactin, Hsp90, and GRP78 |
[155] |
| ACY-1215 + oxaliplatin | Apoptosis, downregulation of p-ERK and p-AKT | [155] |
| ACY-1215 + carfilzomib | Accumulation of protein aggregates, ER stress, apoptosis | [165] |
| ACY-1215 + 5- fluorouracil | Inhibition of proliferation | [156] |
| ACY-241 + paclitaxel | Growth arrest | [162] |
| A452 | Activation of caspase-3 and PARP; increased Bak and Bax, decreased Bcl-xL level, increased PD-L1 expression | [157] |
| A452 + Vorinostat | Inhibition of proliferation, apoptosis | [158] |
| A452 + Aceroside VIII | Inhibition of proliferation, apoptosis | [159] |
| C1A | Inhibition of proliferation, apoptosis acetylation of α-tubulin and HSP90 | [150] |
| C1A + bortezomib | Accumulation of misfolded proteins and decreased autophagy | [166] |
| Tubacin | Ku70 acetylation and suppression of FLIP, apoptosis | [77] |
| MPT0G612 | Inhibition of proliferation, apoptosis, decreased PD-L1 expression | [164] |
| Romidepsin + pembrolizumab | Synergistic antitumor effect | [220] |
| CXD101 + nivolumab | Synergistic antitumor effect | [219] |
| BRAF/HDAC dual inhibitor compound 14b |
Inhibition of proliferation | [234] |
| compound 12 | Inhibition of HDAC6 and Hsp90 | [159] |
Table 4.
Immune-related effects of HDAC6 inhibition.
| Inhibitor | Cell | Effect | References |
|---|---|---|---|
| Vorinostat (SAHA) | DCs T cells Tumor cells |
↓CD40, CD80, CD83, ↓ TNF, IL-6, IL-12 ↑IDO ↑T cell proliferation ↑Cytotoxicity, IFNγ ↑Fas mediated cytotoxicity ↑PD-L1 ↑MICA/MICB (NK cell ligands) |
[177] [177] [177], [178] [186], [149] [186], [149] [214] [190] |
| Panobinostat | DCs | ↓CD40, CD83, ↓MHC I | [176] |
| (LBH589) | ↓ TNF, IL-6, IL-10, IL-12, IL-23 | [176] | |
| CD4 T cells | ↓IFN-γ | [176] | |
| Tumor cell | ↑ CD80, CD86, CD112(↑ NK cell synapsis) | [188] | |
| ↑PD-L1, | [188], [206] | ||
| ↑MHC I, CD40, CD80 | [207] | ||
| Rodempsin | T cells Tumor cells |
↓ proliferation, activation ↑apoptosis ↑CCL5, CCXL9,10 |
[181] [183] [191] |
| Trichostatin A | Naïve T cells | ↓T cell proliferation, activation | [181], [184] |
| ↑T cell infiltration, ↑ apoptosis | [184] | ||
| Macrophages | ↑M1 differentiation | [193] | |
| Tumor cells | ↑MICA/MICB | [189] | |
| ↑PD-L1 | [193] | ||
| Ricolinostat (ACY-1215) | T cells | activation (CD38) ↑perforin, IFN-γ/IL-2 ↓PD-1, TIM3, LAG-1 |
[185] |
| [185] | |||
| [185] | |||
| ↓IL-4, IL-5, IL-6, IL-10, IL-13 | [185] | ||
| MDSC | ↓MDSC | [192] | |
| Tumor cells | ↑CD80, CD86, MHC I, MHC II ↑PD-L1 |
[155] [155], [157] |
|
| Citarinostat (ACY241) | Tumor cells and DCs T cells |
↑CD80, CD86, MHC I, MHC II ↑co-stimulatory (CD28, 41BB, CD40L, OX40) ↓IL-4, IL-5, IL-6, IL-10, IL-13 |
[179] [179] [185] |
| ↑perforin, IFN-γ/IL-2 | [179], [185] | ||
| ↓PD-1, TIM3, LAG-1 | [185] | ||
| Tubastatin A | DCs T cells Macrophages Tumor cells Treg |
↓IL-10 ↓perforin secretion ↓TNF, IL-6, NO ↑MHC I ↑FoxP3, CTL-4, IL-10, PD-1 |
[118] [125] [180] [132] [127] |
| Tubacin | Treg | ↑FoxP3, CTL-4, IL-10, PD-1 | [127] |
| Nexturastat A | NK cells | ↑NK cell infiltration | [121], |
| macrophages | ↑M1 differentiation | [121] | |
| Tumor cells | ↓PD-L1 | [121], [163], [204], | |
| MPTOG612 | Tumor cells | ↓PD-L1 | [164] |
| A452 | Tumor cells | ↑PD-L1 | [157] |
| SP-2-225 | Macrophages | ↑M1 differentiation | [175] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



