
Submitted:
28 November 2023
Posted:
29 November 2023
You are already at the latest version
Abstract
Grapevine trunk diseases (GTDs) attack the vine's wood, devastating vineyards worldwide. Chile is the world's fourth largest wine exporter, and Cabernet Sauvignon leads the red wine varieties. Botryosphaeria dieback is an important GTDs and Diplodia seriata is one of the main pathogenic species. Biocontrol studies of these pathogens are commonly carried out at different incubation times but at a single temperature. This study aimed to evaluate the biocontrol effect of native Chilean bacteria against D. seriata isolates at different temperatures. We analyzed the biocontrol effects of Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b and Rhodococcus sp. PU4, with three D. seriata isolates (PUCV 2120, PUCV 2142 and PUCV 2183) at 8, 22 and 35°C. Agar plug diffusion and double plate methods were used to evaluate the effect of diffusible and volatile organic compounds (DOC and VOC), and an in vivo test was performed with Cabernet Sauvignon cuttings. Pseudomonas strains showed the most significant inhibition in the DOC assay, and a demelanizing activity was observed. Rhodococcus sp. PU4 showed VOC biocontrol potential at high temperatures (35°C). In vivo, a biocontrol effect was observed against two D. seriata isolates, while the PUCV 2142 isolate proved to be more resistant to being controlled. The biocontrol ability at low temperatures is promising for effective control in the field, where infections occur primarily in winter.
Keywords:
Biocontrol
; Diplodia seriata
; Botryosphaeria dieback
; Grapevine trunk diseases
; GTDs
; Botryosphaeriaceae
; Pseudomonas
; Rhodococcus
; Demelanizing activity
1. Introduction
The cultivation of grapevines (Vitis vinifera) is an important and widespread agricultural activity worldwide that dates back more than 7000 years. Grapevines are highly appreciated for their fresh fruits and wines [1]. However, one of the most significant challenges in growing grapevines is their susceptibility to various pathogens, including viruses, bacteria, fungi, and nematodes [2]. Grapevine trunk diseases (GTDs) attack grapevine wood and devastate vineyards worldwide. Although GTDs have been known for more than a century, their impact has increased significantly in recent decades [3,4,5,6], causing significant production losses [4,7]. The three main GTDs are Botryosphaeria dieback, Eutypa dieback, and Esca disease, which generally attack perennial organs at all stages of vine growth [2,3,8]. Botryosphaeria dieback is one of the most important GTDs worldwide and has been associated with 26 botryosphaeriaceous taxa [9]; the most frequently reported species are Diplodia seriata, Diplodia mutila, Lasiodiplodia theobromae, and Neofusicoccum parvum [2,4,9,10].
Chile is currently the top wine exporter in America and the fourth wine exporter worldwide, surpassed only by France, Spain, and Italy [11]. The total area of Chilean vineyards currently occupies more than 141 thousand hectares, which has a wine production potential of close to 1200 million liters [11]. According to the National Viticultural Registry of Chile 2020, Cabernet Sauvignon leads the red wine varieties with more than 40,000 hectares [12]. A study of Botryosphaeria dieback disease in Cabernet Sauvignon vineyards of central Chile showed that the overall disease incidences were 87% in 2010 and 84% in 2018, the severities of the damage were 49% in 2010 and 47% in 2018, and yield losses were 39% in 2010 and 46% in 2018, wherein D. seriata was the most prevalent fungus isolated from symptomatic plants [4]. Other studies demonstrated that D. seriata (anamorph of Botryosphaeria obtusa) is one of the most frequently isolated species from diseased grapevines in Chile [4,13,14,15] and other regions, such as California [16], Mexico [17], Spain [18,19], Portugal [20], France [21], Iran [22], Lebanon [23], Algeria [23], Tunisia [25], and Australia [26].
Compared to other species of Botryosphaeriaceae (Botryosphaeria dothidea, Diplodia corticola, D. mutila, Dothiorella iberica, L. theobromae, N. parvum, and Spencermartinsia viticola), conidia of D. seriata showed the highest germination under a wide range of temperatures (10 to 40°C) [27]. More isolates of D. seriata than N. parvum were isolated from colder places in Iran [22]. Diplodia seriata strains were isolated at temperatures between 5 and 40°C. These studies suggest that D. seriata is probably the most cosmopolitan botryosphaeriaceous fungus infecting grapevines [27]. Climate change, including extreme heat or cold, will continue to increase stress on plant communities, and some species of Botryosphaeriaceae, such as D. seriata, will probably cause large-scale damage [28] due to their wide growth temperature range. Considering this, it is essential to know, on the one hand, how the Chilean strains of D. seriata behave at different temperatures and their possible biocontrol agents. During the pruning season, there is dispersion of spores, which coincides with rainfall [15], and the average temperatures oscillate between 8 and 22°C in the Maule region of Central Chile. During the summer, the maximum average temperatures on some days exceed 35°C (https://agrometeorología.cl).
Several species of Botryosphaeriaceae are endophytes or have an endophytic stage, including D. seriata [28]. Currently, there are no efficient tools to eradicate infections caused by these pathogens other than surgical removal of the infected organs, so they are managed mainly by practices that aim to prevent infections [9,29]. In recent years, synthetic products for controlling Botryosphaeria dieback in Vitis vinifera, such as benzimidazoles (benomyl and carbendazim), have been limited and banned in several countries [29]. Biocontrol of GTDs using microorganisms is a promising alternative [30,31] that appears to be a response to the increase in wood diseases, product restrictions and low efficacy of some chemicals [29]. Bacteria from the phylum Actinomycetota [32,33] and the genus Pseudomonas [30] have shown high antifungal activity against fungi associated with GTDs.
Biocontrol in vitro and in vivo studies of grapevine trunk pathogens are commonly performed at different incubation times but only at a single temperature, usually 25°C [30,31,32,33]. In vitro and in vivo studies have shown that the fungus Diplodia seriata is able to grow at a wide range of temperatures [22,27]. Therefore, its biocontrol at different temperatures should be studied, considering those that occur in winter and summer during vineyard cultivation. The objective of this study was to evaluate the biocontrol effects of native bacteria from Chile against different isolates of Diplodia seriata at low (8°C), medium (22°C), and high temperatures (35°C). The potential diffusible organic compounds (DOCs) and volatile organic compounds (VOCs) were evaluated in vitro using agar plug diffusion and double plate methods, and the in vivo assay with Cabernet Sauvignon cuttings. We analyzed the biocontrol effects of three native bacteria, Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b, and Rhodococcus sp. PU4, on three D. seriata isolates at 8, 22, and 35°C.
2. Materials and Methods
2.1. Chemicals, reagents, and culture media
D-Glucose was obtained from Merck (Darmstadt, Germany). Yeast extract and Bacto Proteose Peptone No. 3 were purchased from Difco Laboratories (Franklin Lakes, NJ, USA). Malt extract and potato dextrose agar (PDA) were obtained from HiMedia Laboratories (West Chester, PA, USA).
2.2. Microorganisms
The psychrotolerant plant growth-promoting bacteria (PGPB) for biocontrol studies were Pseudomonas sp. GcR15a [34], Pseudomonas sp. AMCR2b (protected use under patent request), and the endophyte Rhodococcus sp. PU4, which was obtained from a healthy and nongrafted Cabernet Sauvignon plant. Three phytopathogenic D. seriata isolates were used in this study: PUCV 2120, PUCV 2142, and PUCV 2183, obtained from diseased grapevine plants [4]. These microorganisms were obtained from the culture collections of the Escuela de Agronomía (Pontificia Universidad Católica de Valparaíso, Quillota, Chile) and the Molecular Microbiology and Environmental Biotechnology Laboratory (Universidad Técnica Federico Santa María, Valparaíso, Chile).
2.3. In vitro biocontrol assay of diffusible organic compounds
The effects of DOCs on D. seriata isolates were evaluated using the agar plug diffusion method as described by Balouiri et al. [34], with modifications. Yeast malt (YM) agar culture medium (10 g L−1 glucose, 3 g L−1 malt extract, 5 g L−1 peptone, 3 g L−1 yeast extract, and 15 g L−1 agar) was used for biocontrol bacteria (strains GcR15a, AMCR2b, and PU4), and PDA (HiMedia) was used for D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183). Bacteria were grown in YM medium for ~12 h and then adjusted to a turbidity at 600 nm of 1. Three drops of 100 µL of each isolate were placed on a plate, allowed to dry, and incubated for 72 h at room temperature (22 ± 3°C). D. seriata isolates were grown in PDA medium for 4–5 days at 26°C. An 8 mm mycelial disc of colony under active growth of each D. seriata isolate was obtained with a sterilized metal punch and deposited in a PDA plate at 4 cm of an 8 mm piece of YM agar with bacterial growth also obtained from a colony in active growth. A negative control (YM medium without bacteria) and a positive inhibition control (tebuconazole, 0.3 mg L−1) [36] were incorporated. This assay was performed twice in triplicate. Inhibitions were observed at 7 and 14 days at 8, 22, and 35°C. The inner radius of fungal growth was recorded.
2.4. In vitro biocontrol assay of volatile organic compounds
The effects of VOCs on D. seriata isolates were analyzed by the double plate method as reported by Delgado et al. [37], with modifications according to Gao et al. [38]. YM culture medium was used for biocontrol bacteria (strains GcR15a, AMCR2b, and PU4), and PDA was used for D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183). The biocontrol bacteria were streaked onto YM plates, 8 mm pieces of D. seriata isolates were placed in the center of PDA plates, and the two plates were sealed with parafilm. This assay was performed twice in triplicate. Fungal growth inhibition was observed after 7 and 14 days at 8, 22, and 35°C. The fungal growth diameter was measured.
2.5. In vivo biocontrol assay
The in vivo biocontrol capacity of the grapevine endophytic bacterium Rhodococcus sp PU4 and the PGPBs Pseudomonas sp. GcR15a and Pseudomonas sp. AMCR2b was determined with three D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183) employing cuttings following the methodology of Haidar et al. [39] and Kotze et al. [40]. From a vineyard cv. Cabernet Sauvignon, 18 cm long one-year-old semi-lignified cuttings were taken, disinfected with 1% sodium hypochlorite for 5 min and 95% ethanol for 30 s, and washed three times with sterile distilled water (SDW). Finally, it was dry at room temperature inside a laminar flow chamber. The cuttings were kept at 5°C for two weeks before use. A fresh wound was made and was immediately inoculated with 40 µL of each bacterial suspension (1*108 CFU mL–1) in YM medium, YM medium (negative control, C-), and Tebuconazole (0.5% p v–1, positive control, C+). Bacterial suspensions were prepared as described previously in Vega-Celedón et al. [34]. After inoculation with the treatments, the cuttings were left at room temperature until the total absorption of the inoculum. Then, each cutting was placed in a humid chamber with a disinfected plastic grid including a sterile absorbent moistened paper (5 mL of SDW) and placed at 8, 22°C and 35°C. One day later, the cuttings were inoculated with a 50 µL suspension of D. seriata at a concentration of 1*105 conidia µL-1 at the same end where the previous inoculation was performed. Fungal suspensions were prepared as described previously in Larach et al. [6,41]. Cuttings inoculated with each D. seriata isolate and their treatments (GcR15a, AMCR2b, PU4, C- and C+) were placed in humid chambers at 8, 22, and 35°C. Each treatment consisted of 6 replicates, and the assay was evaluated two months after inoculation with D. seriata. The length of the vascular lesions was measured in every detached cutting.
2.6. Statistical analysis
One-way ANOVA was used to analyze the effects of the biocontrol strains. After performing one-way ANOVA, Tukey's test was used to detect significant differences (p < 0.05) for the in vitro biocontrol assay, and LSD was used to evaluate the in vivo biocontrol assay using grapevine cuttings.
3. Results
3.1. In vitro biocontrol by diffusible organic compounds
The biocontrol effect by native biocontrol bacteria (Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b, and Rhodococcus sp. PU4) against D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183) was evaluated through DOC activity. The activity was evaluated by the agar plug diffusion method at different temperatures (Figure 1, Figure 2, Figure 3 and Figure 4), where the fungal mycelial growth inhibition was determined by measuring the inner radius of the fungal growth area.
For the biocontrol by DOCs of D. seriata PUCV 2120 (Figure 1), strain GcR15a exhibited a significant decrease in growth after 7 and 14 days at 8 and 22°C (Figure 4a,d), while at 35°C, no biocontrol effect was observed. Strain GcR15a reached 75% and 56% inhibition after 14 days at 8 and 22°C, respectively. Strain AMCR2b presented a significant inhibitory effect on growth after 7 and 14 days at 8 and 22°C (Figure 4a, d). At 35°C, inhibition was observed only after 14 days (Figure 1g). Strain AMCR2b reached 20%, 65%, and 43% inhibition after 14 days at 8, 22, and 35°C, respectively. With strain AMCR2b, changes in the color of the fungal growth were also observed after 7 and 14 days at 22°C (Figure 1b). For strain PU4, a significant decrease in D. seriata growth was observed after 7 and 14 days only at 8°C (Figure 4a), reaching 74% inhibition after 14 days. Strain AMCR2b presented the highest decrease in the inner radius of the isolate PUCV 2120 (Figure 1). The positive control (tebuconazole) presented complete inhibition after 7 and 14 days at 8°C (Figure 4a), while at 7 days at 22°C, there was a significant effect; however, the effect was lost after 14 days (Figure 4d). Significant biocontrol effects were observed at high temperatures only after 14 days at 35°C (Figure 4g).
For the biocontrol by DOCs of D. seriata PUCV 2142 (Figure 2), strain GcR15a exhibited a significant decrease in growth after 7 and 14 days at 8 and 22°C (Figure 4a,d), while at 35°C, a biocontrol effect was not observed. Strain GcR15a reached 55% and 46% inhibition after 14 days at 8 and 22°C, respectively. Strain AMCR2b showed a significant inhibitory effect on growth after 14 days at 8°C (Figure 4b), while at 22 and 35°C, a decrease was observed after 7 and 14 days (Figure 4e,h). Strain AMCR2b reached 20%, 65%, and 43% inhibition after 14 days at 8, 22, and 35°C, respectively. For strain PU4, a significant decrease at 8°C was observed after 7 and 14 days (Figure 4b), while at 35°C, a significant decrease was observed only after 14 days (Figure 1h). In contrast, no effect was observed at 22°C. Strain PU4 reached 65% and 27% inhibition after 14 days at 8 and 35°C, respectively. Strain AMCR2b presented the highest decrease in the inner radius of strain PUCV 2142 (Figure 2). The positive control presented a significant inhibitory effect at 8 and 35°C after 7 and 14 days (Figure 4b, h), while at 22°C, its effect was significant only after 7 days (Figure 4d).
The biocontrol by DOCs of D. seriata PUCV 2183 (Figure 3) showed the same effects of strains GcR15a and AMCR2b, significantly decreasing growth after 14 days at 8°C (Figure 4c) and after 7 and 14 days at 22 and 35°C (Figure 4f,i). Strain GcR15a reached 40%, 59%, and 20% inhibition after 14 days at 8, 22, and 35°C, respectively. Strain AMCR2b reached 27%, 61%, and 31% inhibition after 14 days at 8, 22, and 35°C, respectively. For strain PU4, a significant decrease at 8°C and 35°C was observed after 14 days (Figure 4c, 4i), while no inhibition was observed at 22°C. Strain PU4 reached 55% and 35% inhibition after 14 days at 8 and 35°C, respectively. Strains GcR15a and AMCR2b presented the highest decrease in the inner radius of strain PUCV2183 (Figure 3). The positive control presented complete inhibition after 7 and 14 days at 8°C (Figure 4c), while at 22°C, its effect was significant after 7 days (Figure 4f) and at 35°C after 7 and 14 days (Figure 4i).
From the negative controls, we observed that all D. seriata isolates showed higher growth at 22°C, followed by 35 and 8°C (Figure 1, Figure 2 and Figure 3), highlighting that the isolate PUCV 2120 presented warped growth at 35°C with all treatments (Figure 1c). On the other hand, at 22°C, a noticeable change in the color of the growth (demelanizing effect) of all D. seriata strains (PUCV 2120, PUCV 2142, and PUCV 2183) was also observed with the biocontrol strains GcR15a and AMCR2b (Figure 1b, Figure 2b, Figure 3b).
3.2. In vitro biocontrol by volatile organic compounds
The biocontrol effect by native biocontrol bacteria (Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b, and Rhodococcus sp. PU4) against D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183) through VOCs was evaluated by the double plate method at different temperatures (Figure 5, Figure 6, Figure 7 and Figure 8). The fungal mycelial growth inhibition was determined by measuring the diameter of the fungal growth. With respect to the biocontrol of all the D. seriata isolates at 22°C, there was no significant biocontrol effect observed for any strain.
For the biocontrol effect by VOCs of D. seriata PUCV 2120 (Figure 5), strains GcR15a and AMCR2b showed the same effect, significantly decreasing the growth after 7 and 14 days at 8°C (Figure 8a), while no effect was observed at 35°C (Figure 8d). For strain PU4, a significant decrease at 8°C was observed after 14 days (Figure 8a). At 35°C, a significant decrease was determined after 7 and 14 days (Figure 8d). Strain PU4 presented the highest decrease in the growth diameter of the isolate PUCV 2120 (Figure 5).
Biocontrol assays of D. seriata PUCV 2142 VOCs (Figure 6) showed no significant decrease in the diameter of the fungal growth with any of the treatments (Figure 8b,e).
For biocontrol by D. seriata PUCV 2183 VOCs (Figure 7), the strains GcR15a and AMCR2b did not present significant decreases in the growth of the fungus at 8 and 35°C (Figure 8c,f). For strain PU4, only a significant decrease at 35°C was observed at 14 days of evaluation (Figure 8f), while at 8°C, no effect was observed (Figure 8d).
In this assay, the bacterial strains studied did not have a significant effect through VOCs, contrasting with the results observed with DOCs. Similar to the DOC assay at 22°C, a notable color change in the growth, demelanizing effect, of the isolates PUCV 2142 and PUCV 2183 was observed with strains GcR15a and AMCR2b (Figure 6 and Figure 7). In addition, irregular growth of the isolate PUCV 2120 at 35°C was observed (Figure 7c).
3.3. Biocontrol on grapevine cuttings
The effects of D. seriata isolates on grapevine cuttings preinoculated with native biocontrol bacteria (Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b, and Rhodococcus sp. PU4) at 8 and 22°C were evaluated. The damage to the grapevine cuttings was observed by measuring the vascular lesion length after two months.
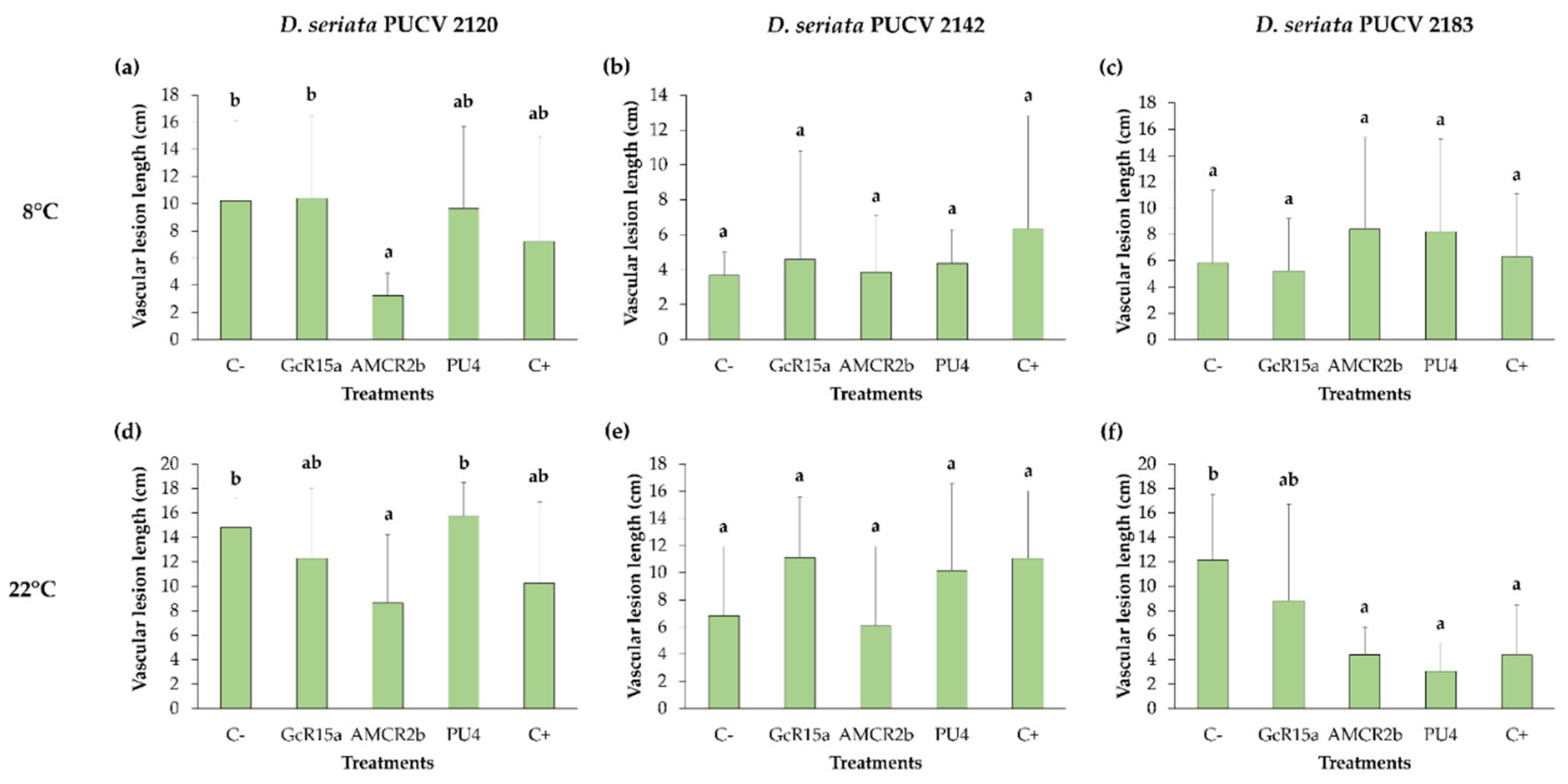
For the biocontrol effect of D. seriata PUCV 2120 (Figure 9), only the strain AMCR2b showed a significant decrease in the vascular lesion length at 8 and 22°C with respect to the negative control (Figure 9a,d).
For the biocontrol effect of D. seriata PUCV 2142 (Figure 9), none of the treatments exhibited significant differences with respect to the negative control at 8 and 22°C (Figure 9b, e).
For the biocontrol effect of D. seriata PUCV 2183 (Figure 9), at 22°C, the strains AMCR2b and PU4 showed a significant decrease in the vascular lesion length with respect to the negative control (Figure 9e), while none of the treatments presented significant differences with respect to the negative control at 8°C (Figure 9c).
4. Discussion
D. seriata is one of the most frequently isolated fungal species from diseased grapevines in different countries and states, such as Chile, California, Mexico, Spain, Portugal, France, Iran, Lebanon, Algeria, Tunisia, and Australia [4,13,14,15,16,17,18,19,20,21,22,23,24,25,26]. The biocontrol of D. seriata by microorganisms is a promising alternative to the use of chemical fungicides [29,30,31]. Due to the wide temperature growth range of this pathogenic fungus [22,27], its biocontrol should be evaluated at different temperatures. In this study, we evaluated the capability of native bacteria from Chile to inhibit the growth of different D. seriata isolates from Mediterranean zones of Chile at 8, 22, and 35°C. This is the first report of in vitro and in vivo biocontrol against D. seriata at different temperatures. In previous work, we determined that D. seriata is the most abundant fungus in V. vinifera cv. Cabernet Sauvignon wood samples with canker lesions [4]. Three D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183) and the endophyte Rhodococcus sp. PU4 were selected for biocontrol assays. In addition, Pseudomonas sp. GcR15a and Pseudomonas sp. AMRC2b (protected use under patent request), whose biocontrol of phytopathogenic bacteria has been reported previously [34], were selected for biocontrol assays. In the assays of this work, we observed a different inner radius or diameter of growth at different temperatures and evaluation times of the three D. seriata isolates. Previous reports support the importance of evaluating the effect on different D. seriata isolates that presented different radius lengths (mycelial growth) at 22°C, where differences in necrosis length in detached canes and potted vines can be detected [42].
Antagonistic effects against phytopathogenic fungi at different temperatures have rarely been reported [43,44,45,46]. In the present work, the biocontrol effect on the growth of three D. seriata isolates (PUCV 2120, PUCV 2142, and PUCV 2183) at 8, 22, and 35°C was analyzed. The best results were obtained by testing in vitro DOCs, where for the three isolates of D. seriata, Pseudomonas sp. GcR15a presented linear growth inhibition at 14 days between 40-75% at 8°C, 46-59% at 22°C, and 10-20% at 35°C, while Pseudomonas sp. AMCR2b presented values from 20-54% at 8°C, 56-65% at 22°C, and 31-43% at 35°C, and Rhodococcus sp. PU4 presented values of 55-74% at 8°C, 0% at 22°C, and 27-35% at 35°C. Álvarez-Pérez et al. [32] observed inhibition by Streptomyces and Saccharopopolyspora strains against D. seriata CBS 112555, with inhibition indexes between 29% and 61% after 12 days of incubation at 25°C. Niem et al. [30] observed mycelial growth inhibition of Pseudomonas strains against D. seriata A142a (12-47%) after 7 days of incubation at 25°C. Silva-Valderrama et al. [31] observed growth inhibition between 15-100% after 21 days of incubation at 25°C with fungal isolates of Chaetomium sp., Cladosporium sp., Clonostachys rosea, Epicoccum nigrum, Purpureocillium lilacinum, and Trichoderma sp. against D. seriata strain 117 Molina. Regarding reports at different temperatures, in a temperature range of 25 to 35°C, Pseudomonas fluorescens RG-26 inhibits the growth (≥50%) of Fusarium oxysporum f. sp. ciceris race 5, whereas Trichoderma sp. isolate Td-1 inhibited (36-56%) the growth of Sclerotium rolfsii [45]. Guetsky et al. [44] and Manaa & Kim [46] highlighted the biocontrol effect at temperatures ≤10°C, observing a significant decrease in the fungus Aspergillus flavus KCCM 60330 growth with the bacterium Pseudomonas protegens AS15 at 10, 20, 30, and 40°C [46], and a similar effect was reported against B. cinerea with a bacterial consortium composed of Pichia guilermondii and Bacillus mycoides at 4, 10, 15, 20, 25, 30, and 36°C, with spore germination inhibition between 20 and 80% [44]. Knowing biocontrol at low temperatures is important for the protection of exposed wounds of vine plants generated by pruning during winter [47], which is consistent with most dispersion times of spores of causal fungi of dieback-type trunk diseases in Mediterranean climate zones such as Chile [15] and California [16,48]. In addition, the importance of demonstrating the biocontrol effect at different temperatures is in accordance with the increasing evidence that suggests that extreme temperatures (cold or heat) will increase in the coming years [28]. Therefore, microbial products that alleviate the stress caused by climate change in crops are required for agriculture [49,50].
In this study, after seven days, tebuconazole inhibited mycelial growth of isolates of D. seriata in PDA agar plates 100% at 8°C, 42-48% at 22°C, and 35-50% at 35°C, maintaining an inhibition at 14 days of 88-100% to 8°C, 0-12% at 22°C, and 15-48% at 35°C. Therefore, tebuconazole presented high efficacy at 8°C. A tebuconazole-based treatment was able to inhibit the conidial and mycelial germination of three D. seriata isolates (Vid 1472, Vid 1468, and Vid 1270) at temperatures of 22 and 25°C [36]. Treatments with Pseudomonas strains GcR15a and AMCR2b at 22°C maintained biocontrol for 14 days for the three D. seriata isolates. This study highlighted the importance of carrying out tests on plants naturally infected or inoculated with D. seriata conidia to determine how infection occurs in nature [51,52].
In the present work, we observed that the greatest biocontrol effect was exhibited by Pseudomonas strains, which were isolated from cold environments [34]. The biocontrol potential of Pseudomonas strains against phytopathogens has been previously reported [30,34,46,53,54,55,56,57]. The assay of VOCs suggested that the biocontrol effect exhibited by the bacteria is probably produced by DOCs. In accordance, DOCs were more effective than VOCs in in vitro assays using Pseudomonas strains against B. cinerea [53]. Niem et al. [30] showed that Pseudomonas is a healthy vineyard's predominant endophytic bacterial genus. In contrast, this genus decreased significantly in diseased plants with GTDs, suggesting a possible effect of Pseudomonas strains against D. seriata and other fungi associated with GTDs. On the other hand, the biocontrol potential of Rhodococcus spp. against phytopathogens has rarely been reported [58,59]. In our work, Rhodococcus sp. PU4 presented a significant inhibitory effect by DOCs in some cases. Still, its inhibitory effect on the growth of D. seriata strains PUCV 2120 and PUCV 2183 by VOCs at high temperatures (35°C) was higher than the effect of Pseudomonas strains. A potential bacterial consortium composed of these three bacteria seems attractive for future studies, presenting various biocontrol mechanisms against D. seriata. The strategy of consortia may increase the efficacy and improve the biocontrol effects due to synergistic mechanisms [56].
In some cases, in in vitro assays, no growth inhibition, just a change of color, was observed for D. seriata growth by the biocontrol bacteria. A darker phenotype was observed in the control at longer incubation times at the optimal temperature (22°C) and 35°C, possibly due to melanin production. Genes involved in melanin syntheses, such as DOPA-melanin (production of aerial mycelium and protection against enzymatic lysis and oxidative stress), DHN-melanin (ramification of mycelium when exposed to nutrient deficiency), and pyomelanin (hyphae development), were conserved among Botryosphaeriaceae, highlighting the importance of melanin in pathogenesis [60]. Melanization is not an essential factor for fungal growth but contributes to the survival of cells under environmental stress conditions and may confer virulence in pathogens [60,61]. Demelanizing activity was observed with Ganoderma lucidum extracts against Aspergillus niger, indicating that the decrease in pigmentation could reduce this virulence factor [63].
When considering the results obtained in vine cuttings, there were differences in biocontrol at 8 and 22°C depending on the type of D. seriata isolate used. Pseudomonas sp. AMCR2b presented a biocontrol effect against the isolate PUCV 2120 at both temperatures and against the isolate PUCV 2183 only at 22°C, and Rhodococcus sp. PU4 showed a biocontrol effect only against the isolate PUCV 2183 at 22°C, while none of the bacterial strains presented activity against the isolate PUCV 2142, demonstrating to be a D. seriata isolate more resistant to be controlled, even by the fungicide tebuconazole. Previous studies performed by Larach et al. [6] showed that the isolate PUCV 2142 presented a greater lesion diameter on grape berries than the isolates PUCV 2120 and PUCV 2183. This aspect reinforces the need to evaluate more than one isolate per pathogen, an aspect not previously considered by some authors [30,31,32]. Therefore, the biocontrol effects of the bacteria studied in this work could prevent the fungus from establishing itself in grapevine plants through direct inhibition or demelanization. In future trials, it will be interesting to test the biocontrol effects of these bacteria in plants with a mixture of D. seriata isolates at different temperatures.
5. Conclusions
In this study, the growth inhibition of three D. seriata isolates by beneficial bacteria was observed. The highest inhibition was observed with both Pseudomonas strains using the DOC assay at different temperatures. In assays without fungal inhibition, antifungal demelanizing activity was observed. Rhodococcus sp. PU4 showed biocontrol potential by producing VOCs capable of inhibiting D. seriata growth at high temperatures (35°C). The in vivo biocontrol assay with cuttings corroborated the biocontrol effect of Pseudomonas sp. AMRC2b in two isolates from D. seriata, while Rhodococcus sp. PU4 presented biocontrol effect in only one isolate. None of the bacterial strains evaluated in the in vivo biocontrol assay showed an effect against the isolated PUCV 2142, demonstrating to be a highly resistant isolate to be controlled. The study of different pathogen isolates should be mostly evaluated to better understand their different behaviors and to improve their control and biocontrol strategies. The results obtained at low temperatures of in vitro and in vivo assays is promising for effective control in the field, where infections occur mainly in winter.
Author Contributions
A.L., P.V.-C, L.T., D.C.-N, and X.B. conceived and designed the experiments; A.L., P.V.-C, L.T., and D.C.-N performed the experiments; A.L., P.V.-C., L.T, D.C.-N., M.S., and X.B. analyzed the data; M.S. and X.B. contributed reagents, materials, and analysis tools; P.V.-C., A.L., D.C.-N., M.S., and X.B. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fondecyt 1211094 (A.L., P.V.-C., L.T., D.C.-N., M.S., X.B.), Fondecyt 1200756 (P.V.-C., M.S.), Millennium Nucleus BioGEM NCN2023_054, PhD USM fellowship (D.C.-N.), and PhD ANID fellowship (A.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The partial 16S rRNA gene sequences of Pseudomonas sp. GcR15a, Pseudomonas sp. AMCR2b, and Rhodococcus sp. PU4 are deposited in GenBank under the accession numbers MW548343, OQ244037, and OQ244039, respectively.
Acknowledgments
The authors thank Dr. Beatriz Cámara, UTFSM researcher, for the suggestions on in vitro assays, Dr. Eduardo Salgado for the statistical support, and the Phytopathology team for their collaboration in preparing the laboratory material.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sargolzaei, M.; Rustioni, L.; Cola, G.; Ricciardi, V.; Bianco, P. A.; Maghradze, D.; Failla, O.; Quaglino, F.; Toffolatti, S. L.; de Lorenzis, G. Georgian grapevine cultivars: Ancient biodiversity for future viticulture. Front. Plant Sci. 2021, 12, 630122. [CrossRef]
- Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.; El Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlali, R.; Barka, E. A. A panoramic view on Grapevine Trunk Diseases threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease. J. Fungi 2022, 8(6), 595. [CrossRef]
- Kovács, C.; Sándor, E. The increasing importance of grapevine trunk diseases. Int. J. Hortic. Sci. 2016, 22(3-4), 21–30. [CrossRef]
- Larach, A.; Torres, C.; Riquelme, N.; Valenzuela, M.; Salgado, E.; Seeger, M.; Besoain, X. Yield loss estimation and pathogen identification from Botryosphaeria dieback in vineyards of Central Chile over two growing seasons. Phytopathol. Mediterr. 2020, 59(3), 537–548.
- Lade, S. B.; Štraus, D.; Oliva, J. Variation in fungal community in grapevine (Vitis vinifera) nursery stock depends on nursery, variety and rootstock. J. Fungi 2022, 8(1), 47.
- Larach, A.; Vega-Celedón, P.; Salgado, E.; Salinas, A.; Riquelme, N.; Castillo-Novales, D.; Sanhueza, P.; Seeger, M.; Besoain, X. Higher virulence of Diplodia seriata isolates on vines of cv. Cabernet Sauvignon associated with 10-Year-old wood compared to young tissue. Plants 2023, 12, 2984. [CrossRef]
- Kaplan J.; Travadon R.; Cooper M.; Hillis V.; Lubell M.; Baumgartner K. Identifying economic hurdles to early adoption of preventative practices: The case of trunk diseases in California winegrape vineyards. Wine Econ. Policy 2016, 5, 127–141. [CrossRef]
- Besoain, X. Grapevine Trunk Diseases (GTDs): Impact in table grapes and wine vineyards in Chile. In: Grapes and Wines- Advances in production, processing, analysis and valorization (A.M. Jordao, F. Cosme, ed.). Intechopen, Londres, 2018, 43–58.
- Gramaje, D.; Urbez-Torres, J. R.; Sosnowski, M. R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102(1), 12–39. [CrossRef]
- Úrbez-Torres, J. R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50(4), 5–45.
- ODEPA. Oficina de Estudios y Políticas Agrarias. Rubros. Vinos. 2022. Available on: https://www.odepa.gob.cl/rubros/vinos-y-alcoholes (accessed on 06 September 2023).
- SAG. Catastro Vitícola Nacional. 2020. Available on: https://www.sag.gob.cl/noticias/sag-presenta-catastro-viticola-nacional-2020 (accessed on 06 September 2023).
- Morales, A.; Latorre, B. A.; Piontelli, E.; Besoain, X. Botryosphaeriaceae species affecting table grape vineyards in Chile and cultivar susceptibility. Cienc. Inv. Agr. 2012, 39(3), 445–458. [CrossRef]
- Díaz, G. A.; Auger, J.; Besoain, X.; Bordeu, E.; Latorre, B. A. Prevalence and pathogenicity of fungi associated with grapevine trunk diseases in Chilean vineyards. Cien. Inv. Agr. 2013, 40(2), 327–339. [CrossRef]
- Valencia, D.; Torres, C.; Camps, R.; López, E.; Celis-Diez, J. L.; Besoain, X. Dissemination of Botryosphaeriaceae conidia in vineyards in the semiarid Mediterranean climate of the Valparaíso Region of Chile. Phytopathol. Mediterr. 2015, 54(2), 394–402.
- Úrbez-Torres, J. R.; Battany, M.; Bettiga, L. J.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R. J.; Verdegaal, P.; Gubler, W. D. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant Dis. 2010a, 94(6), 717–724. [CrossRef]
- Úrbez-Torres, J. R.; Leavitt, G. M.; Guerrero, J. C.; Guevara, J.; Gubler, W. D. Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of grapevines in Mexico. Plant Dis. 2008, 92(4), 519–529.
- Martín, M. T.; Cobos, R. Identification of fungi associated with grapevine decline in Castilla y León (Spain). Phytopathol. Mediterr. 2007, 46, 18–25.
- Luque, J.; Elena, G.; Garcia-Figueres, F.; Reyes, J.; Barrios, G.; Legorburu, F. J. Natural infections of pruning wounds by fungal trunk pathogens in mature grapevines in Catalonia (Northeast Spain). Aust. J. Grape Wine Res. 2014, 20(1), 134–143. [CrossRef]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Lourenço, I.; Saramago, J.; Mota, M.; Rego, C. Early season symptoms on stem, inflorescences and flowers of grapevine associated with Botryosphaeriaceae species. Plants 2020, 9(11), 1427.
- Kuntzmann, P.; Villaume, S.; Larignon, P.; Bertsch, C. Esca, BDA and Eutypiosis: foliar symptoms, trunk lesions and fungi observed in diseased vinestocks in two vineyards in Alsace. Vitis 2010, 49, 71–76.
- Mohammadi, H.; Gramaje, D.; Banihashemi, Z.; Armengol, J. Characterization of Diplodia seriata and Neofusicoccum parvum associated with grapevine decline in Iran. J. Agr. Sci. Tech. 2013, 15, 603–616.
- Choueiri, E.; Jreijiri, F.; Chlela, P.; Louvet, G.; Lecomte, P. Occurrence of grapevine declines and first report of black dead arm associated with Botryosphaeria obtusa in Lebanon. Plant Dis. 2006, 90(1):115. [CrossRef]
- Ammad, F.; Benchabane, M.; Toumi, M.; Belkacem, N.; Guesmi, A.; Ameur, C.; Lecomte, P.; Merah, O. (2014). Occurrence of Botryosphaeriaceae species associated with grapevine dieback in Algeria. Turk. J. Agric. For. 2014, 38(6), 865–876. [CrossRef]
- Chebil, S.; Fersi, R.; Bouzid, M.; Quaglino, F.; Chenenaoui, S.; Melki, I.; Durante, G.; Zacchi, E.; Bahri, B. A.; Bianco, P. A.; Rhouma, A. Fungi from the Diaporthaceae and Botryosphaeriaceae families associated with grapevine decline in Tunisia. Cienc. Investig. Agrar. 2017, 44(2), 127–138.
- Savocchia, S.; Steel, C. C.; Stodart, B. J. Pathogenicity of Botryosphaeria species isolated from declining grapevines in sub tropical regions of eastern Australia Grapevine Trunk Disease. Vitis 2007, 46(1), 27–32.
- Úrbez-Torres, J. R.; Bruez, E.; Hurtado, J.; Gubler, W. D. Effect of temperature on conidial germination of Botryosphaeriaceae species infecting grapevines. Plant Dis. 2010b, 94(12), 1476–1484. [CrossRef]
- Slippers, B.; Wingfield, M. J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21(2–3), 90–106. [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102(7), 1189–1217. [CrossRef]
- Niem, J. M.; Billones-Baaijens, R.; Stodart, B.; Savocchia, S. Diversity profiling of grapevine microbial endosphere and antagonistic potential of endophytic Pseudomonas against Grapevine Trunk Diseases. Front. Microbiol. 2020, 11, 477. [CrossRef]
- Silva-Valderrama, I.; Toapanta, D.; Miccono, M. de los A.; Lolas, M.; Díaz, G. A.; Cantu, D.; Castro, A. Biocontrol potential of grapevine endophytic and rhizospheric fungi against trunk pathogens. Front. Microbiol. 2021, 11, 614620. [CrossRef]
- Álvarez-Pérez, J. M.; González-García, S.; Cobos, R.; Olego, M. Á.; Ibañez, A.; Díez-Galán, A.; Garzón-Jimeno, E.; Coque, J. J. R. Use of endophytic and rhizosphere actinobacteria from grapevine plants to reduce nursery fungal graft infections that lead to young grapevine decline. Appl. Environ. Microbiol. 2017, 83(24). [CrossRef]
- Laassami, A.; Yekkour, A.; Meklat, A.; Djemouai, N.; Zitouni, A.; Mokrane, S.; Lecomte, P.; Rey, P.; Berraf-Tebbal, A. Actinobacteria associated with vineyard soils of Algeria: Classification, antifungal potential against grapevine trunk pathogens and plant growth-promoting features. Curr. Microbiol. 2020, 77(10), 2831–2840. [CrossRef]
- Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F. P.; Valenzuela, M.; Ramírez, I.; Vasconez, I. N.; Álvarez, I.; Jorquera, M. A.; Seeger, M. Microbial diversity of psychrotolerant bacteria isolated from wild flora of Andes Mountains and Patagonia of Chile towards the selection of plant growth-promoting bacterial consortia to alleviate cold stress in plants. Microorganisms 2021, 9(3), 1–28. [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S. K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6(2), 71–79. [CrossRef]
- Torres, C.; Latorre, B. A.; Undurraga, P.; Besoain, X. Evaluation of DMI fungicides against species of Diplodia and Neofusicoccum associated with Botryosphaeria canker of grapevine. Cienc. Inv. Agr. 2013, 40(1), 131–138. [CrossRef]
- Delgado, N.; Olivera, M.; Cádiz, F.; Bravo, G.; Montenegro, I.; Madrid, A.; Fuentealba, C.; Pedreschi, R.; Salgado, E.; Besoain, X. Volatile Organic Compounds (VOCs) produced by Gluconobacter cerinus and Hanseniaspora osmophila displaying control effect against table grape-rot pathogens. Antibiotics 2021, 10, 663. [CrossRef]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [CrossRef]
- Haidar, R.; Deschamps, A.; Roudet, J.; Calvo-Garrido, C.; Bruez, E.; Rey, P.; Fermaud, M. Multi-organ screening of efficient bacterial control agents against two major pathogens of grapevine. Biol. Control 2016, 92, 55–65. [CrossRef]
- Kotze, C.; van Niekerk, J.; Halleen, F.; Mostert, L.; Pourie, P. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, 247–263.
- Larach, A.; Riquelme, N.; Salinas, A.; Rolshausen, P. E.; Seeger, M.; Besoian, X. First report of Diaporthe ambigua associated with dead arm disease on grapevine in Chile. Plant Dis. 2021, 106(7). [CrossRef]
- Elena, G.; Garcia-Figueres, F.; Reigada, S.; Luque, J. Intraspecific variation in Diplodia seriata isolates occurring on grapevines in Spain. Plant Pathol. 2015, 64(3), 680–689. [CrossRef]
- Mukherjee, P. K.; Raghu, K. Effect of temperature on antagonistic and biocontrol potential of Trichoderma sp. on Sclerotium rolfsii. Mycopathologia 1997, 139(3), 151–155.
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Dinoor, A. Combining biocontrol agents to reduce the variability of biological control. Phytopathology 2001, 91(7), 621–627. [CrossRef]
- Landa, B. B.; Navas-Cortés, J. A.; Jiménez-Díaz, R. M. Influence of temperature on plant-rhizobacteria interactions related to biocontrol potential for suppression of fusarium wilt of chickpea. Plant Pathol. 2004, 53, 341–352. [CrossRef]
- Mannaa, M.; Kim, K. D. Effect of temperature and relative humidity on growth of Aspergillus and Penicillium spp. and biocontrol activity of Pseudomonas protegens AS15 against aflatoxigenic Aspergillus flavus in stored rice grains. Mycobiology 2018, 46(3), 287–295.
- Rolshausen, P. E., Úrbez-Torres, J. R.; Rooney-Latham, S.; Eskalen, A.; Smith, R. J. Evaluation of pruning wound susceptibility and protection against fungi associated whith grapevine trunk diseases. Am. J. Enol. Vitic. 2009, 60, 113–119.
- Fujiyoshi, P. T.; Lawrence, D. P.; Travadon, R.; Cooper, M.; Verdegaal, P.; Schwebs, S.; Baumgartner, K. Detection of spores of causal fungi of dieback-type trunk diseases in young, asymptomatic vineyards and mature, symptomatic vineyards. Crop Prot. 2021, 150:105798. [CrossRef]
- Naamala, J.; Smith, D. L. Relevance of plant growth promoting microorganisms and their derived compounds, in the face of climate change. Agronomy 2020, 10, 1179. [CrossRef]
- Fadiji, A. E.; Babalola, O. O.; Santoyo, G.; Perazzolli, M. The potential role of microbial biostimulants in the amelioration of climate change-associated abiotic stresses on crops. Front. Microbiol. 2022, 12:829099. [CrossRef]
- van Niekerk J. M.; Fourie; P. H.; Halleen, F.; Crous, P. Botryosphaeria spp. as grapevine trunk disease pathogens. Phytopathol. Mediterr. 2006, 45, 43–54.
- Úrbez-Torres, J. R.; Gubler, W. B. Susceptibility of grapevine pruning wounds to infection by Lasiodiplodia theobromae and Neofusicoccum parvum. Plant Pathol. 2011, 60, 261–270. [CrossRef]
- Hernández-León, R.; Rojas-Solís, D.; Contreras-Pérez, M.; Orozco-Mosqueda, M. del C.; Macías-Rodríguez, L. I.; Reyes-de la Cruz, H.; Valencia-Cantero, E.; Santoyo, G. Characterization of the antifungal and plant growth-promoting effects of diffusible and volatile organic compounds produced by Pseudomonas fluorescens strains. Biol. Control 2015, 81, 83–92. [CrossRef]
- Simionato, A. S.; Navarro, M. O. P.; de Jesus, M. L. A.; Barazetti, A. R.; da Silva, C. S.; Simões, G. C.; Balbi-Peña, M. I.; de Mello, J. C. P.; Panagio, L. A.; de Almeida, R. S. C.; Andrade, G.; de Oliveira, A. G. The effect of phenazine-1-carboxylic acid on mycelial growth of Botrytis cinerea produced by Pseudomonas aeruginosa LV strain. Front. Microbiol. 2017, 8, 1102.
- Kong, W. L.; Li, P. S.; Wu, X. Q.; Wu, T. Y.; Sun, X. R. Forest tree associated bacterial diffusible and volatile organic compounds against various phytopathogenic fungi. Microorganisms 2020, 8(4), 590. [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as biological control agents of plant diseases. Microorganisms 2022, 10, 1759. [CrossRef]
- Win, K. T.; Kobayashi, M.; Tanaka, F.; Takeuchi, K.; Oo, A. Z.; Jiang, C. J. Identification of Pseudomonas strains for the biological control of soybean red crown root rot. Sci. Rep. 2022, 12, 14510. [CrossRef]
- Chane, A.; Barbey, C.; Robert, M.; Merieau, A.; Konto-Ghiorghi, Y.; Beury-Cirou, A.; Feuilloley, M.; Pátek, M.; Gobert, V.; Latour, X. Biocontrol of soft rot: Confocal microscopy highlights virulent pectobacterial communication and its jamming by rhodococcal quorum-quenching. Mol. Plant Microbe Interact. 2019, 32(7), 802–812. [CrossRef]
- Zhou, Z.; Wu, X.; Li, J.; Zhang, Y.; Huang, Y.; Zhang, W.; Shi, Y.; Wang, J.; Chen, S. A novel quorum quencher, Rhodococcus pyridinivorans XN-36, is a powerful agent for the biocontrol of soft rot disease in various host plants. Biol. Control 2022, 169, 104889. [CrossRef]
- Rangel-Montoya, E. A.; Paolinelli, M.; Rolshausen, P.; Hernandez-Martinez, R. The role of melanin in the grapevine trunk disease pathogen Lasiodiplodia gilanensis. Phytopathol. Mediterr. 2020, 59(3), 549–563.
- Eisenman, H. C.; Greer, E. M.; McGrail, C. W. The role of melanins in melanotic fungi for pathogenesis and environmental survival. Appl. Microbiol. Biotechnol. 2020, 104(19), 4247–4257. [CrossRef]
- Heleno, S. A.; Ferreira, I. C. F. R.; Esteves, A. P.; Ćirić, A.; Glamočlija, J.; Martins, A.; Soković, M.; Queiroz, M. J. R. P. Antimicrobial and demelanizing activity of Ganoderma lucidum extract, p-hydroxybenzoic and cinnamic acids and their synthetic acetylated glucuronide methyl esters. Food Chem. Toxicol. 2013, 58, 95–100. [CrossRef]
Figure 1.
Biocontrol by native bacteria against Diplodia seriata PUCV 2120 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2120 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2120 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2120 at 35°C after 7 and 14 days. Abbreviations: 2120, D. seriata PUCV 2120; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Figure 1.
Biocontrol by native bacteria against Diplodia seriata PUCV 2120 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2120 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2120 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2120 at 35°C after 7 and 14 days. Abbreviations: 2120, D. seriata PUCV 2120; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).

Figure 2.
Biocontrol by native bacteria against Diplodia seriata PUCV 2142 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2142 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2142 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2142 at 35°C after 7 and 14 days. Abbreviations: 2142, D. seriata PUCV 2142; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Figure 2.
Biocontrol by native bacteria against Diplodia seriata PUCV 2142 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2142 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2142 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2142 at 35°C after 7 and 14 days. Abbreviations: 2142, D. seriata PUCV 2142; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).

Figure 3.
Biocontrol by native bacteria against Diplodia seriata PUCV 2183 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2183 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2183 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2183 at 35°C after 7 and 14 days. Abbreviations: 2183, D. seriata PUCV 2183; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Figure 3.
Biocontrol by native bacteria against Diplodia seriata PUCV 2183 by the agar plug diffusion method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2183 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2183 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2183 at 35°C after 7 and 14 days. Abbreviations: 2183, D. seriata PUCV 2183; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
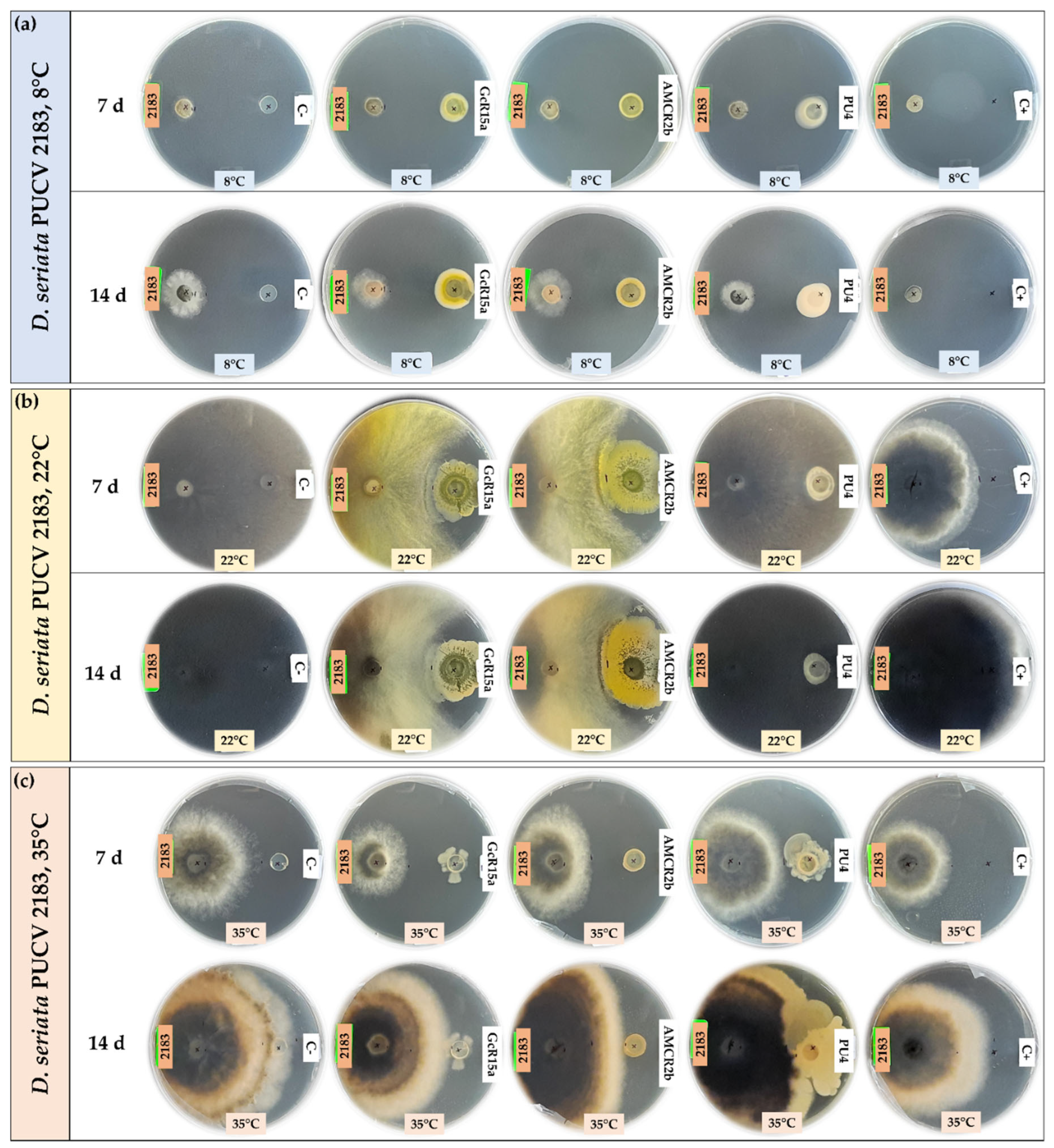
Figure 4.
Effects of native bacteria on the inner radius of Diplodia seriata isolates by the agar plug diffusion method at different temperatures. (a-c) Effects of native bacteria on the inner radius of the isolates of D. seriata at 8°C after 7 and 14 days. (d-f) Effects of native bacteria on the inner radius of D. seriata isolates at 22°C after 7 and 14 days. (g-i) Effects of native bacteria on the inner radius of D. seriata isolates at 35°C after 7 and 14 days. Means with different letters indicate significant differences (p < 0.05), and uppercase and lowercase letters correspond to 7 and 14 days, respectively. Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Figure 4.
Effects of native bacteria on the inner radius of Diplodia seriata isolates by the agar plug diffusion method at different temperatures. (a-c) Effects of native bacteria on the inner radius of the isolates of D. seriata at 8°C after 7 and 14 days. (d-f) Effects of native bacteria on the inner radius of D. seriata isolates at 22°C after 7 and 14 days. (g-i) Effects of native bacteria on the inner radius of D. seriata isolates at 35°C after 7 and 14 days. Means with different letters indicate significant differences (p < 0.05), and uppercase and lowercase letters correspond to 7 and 14 days, respectively. Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
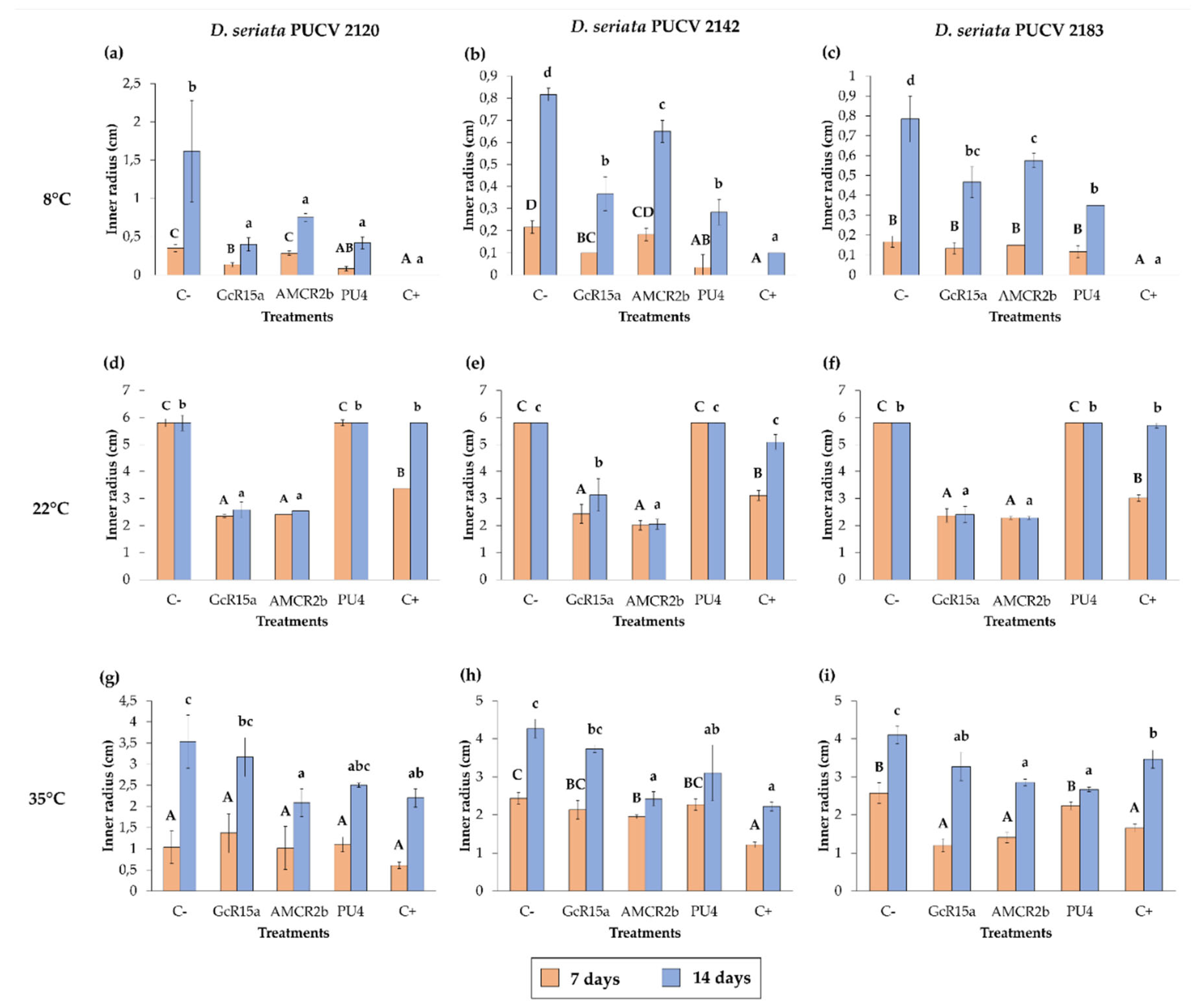
Figure 5.
Biocontrol by native bacteria against Diplodia seriata PUCV 2120 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2120 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2120 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2120 at 35°C after 7 and 14 days. Abbreviations: 2120, D. seriata PUCV 2120; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
Figure 5.
Biocontrol by native bacteria against Diplodia seriata PUCV 2120 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2120 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2120 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2120 at 35°C after 7 and 14 days. Abbreviations: 2120, D. seriata PUCV 2120; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
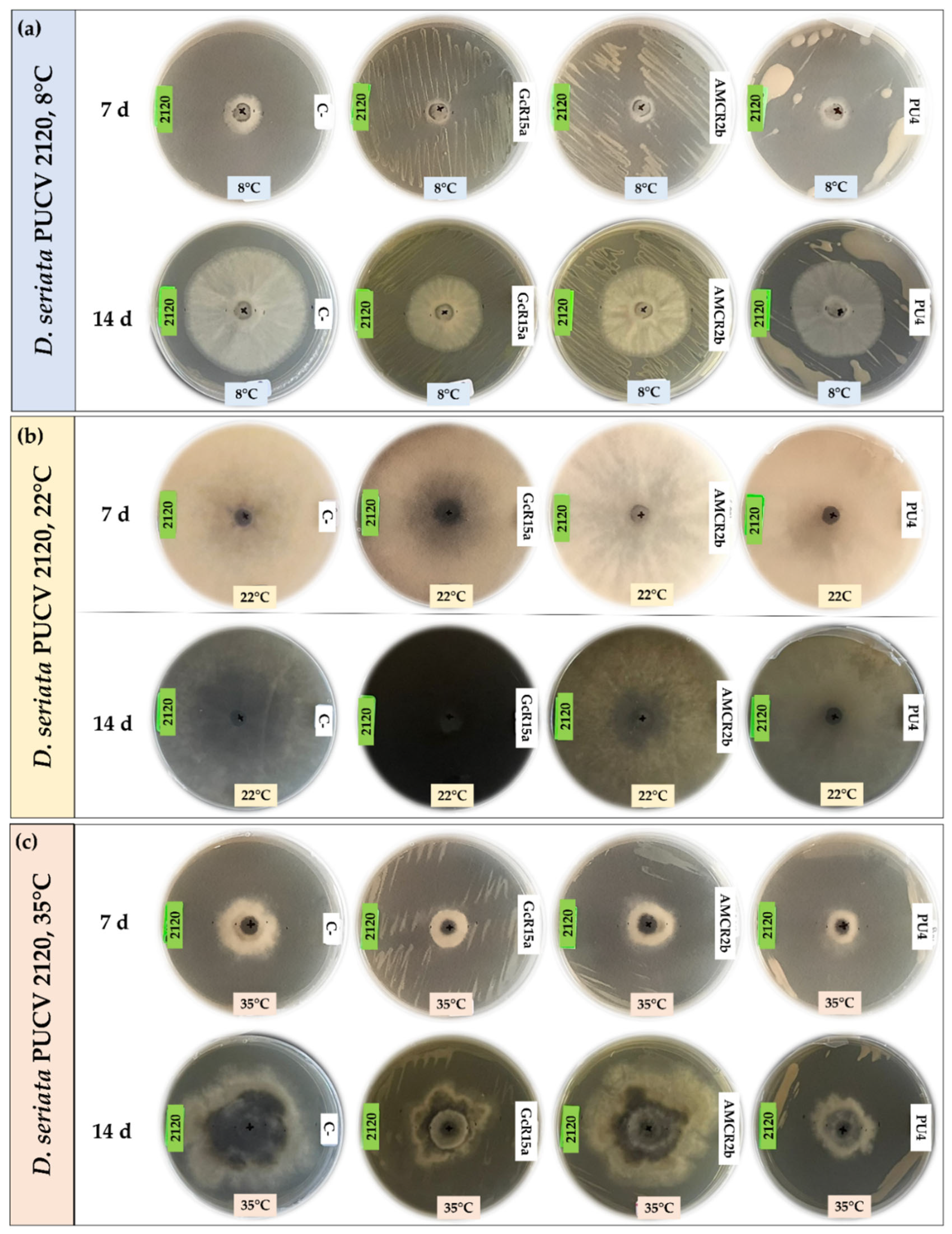
Figure 6.
Biocontrol by native bacteria against Diplodia seriata PUCV 2142 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2142 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2142 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2142 at 35°C after 7 and 14 days. Abbreviations: 2142, D. seriata PUCV 2142; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
Figure 6.
Biocontrol by native bacteria against Diplodia seriata PUCV 2142 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2142 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2142 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2142 at 35°C after 7 and 14 days. Abbreviations: 2142, D. seriata PUCV 2142; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
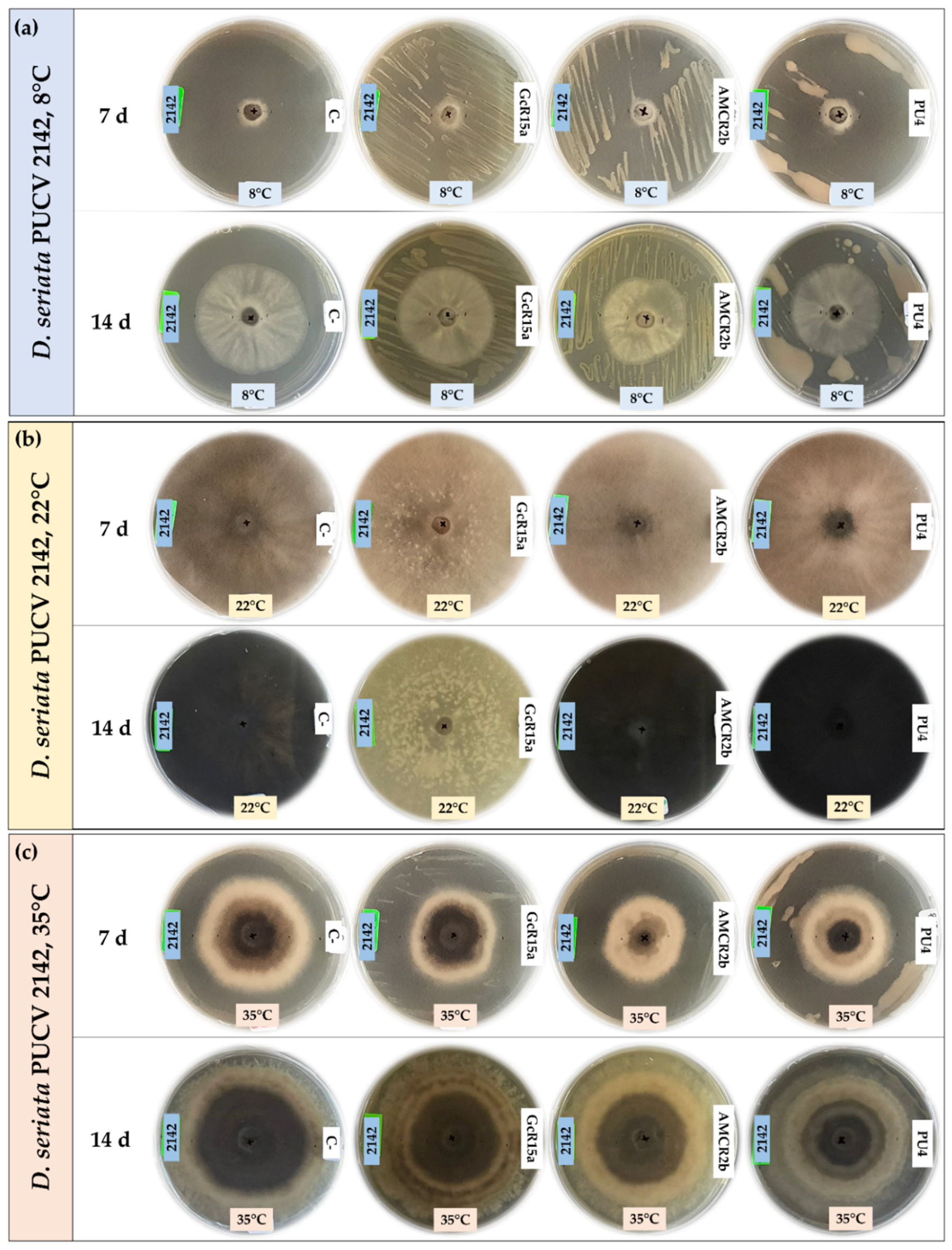
Figure 7.
Biocontrol by native bacteria against Diplodia seriata PUCV 2183 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2183 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2183 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2183 at 35°C after 7 and 14 days. Abbreviations: 2183, D. seriata PUCV 2183; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
Figure 7.
Biocontrol by native bacteria against Diplodia seriata PUCV 2183 by the double plate method at different temperatures. (a) Biocontrol by native bacteria against D. seriata PUCV 2183 at 8°C after 7 and 14 days. (b) Biocontrol by native bacteria against D. seriata PUCV 2183 at 22°C after 7 and 14 days. (c) Biocontrol by native bacteria against D. seriata PUCV 2183 at 35°C after 7 and 14 days. Abbreviations: 2183, D. seriata PUCV 2183; C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
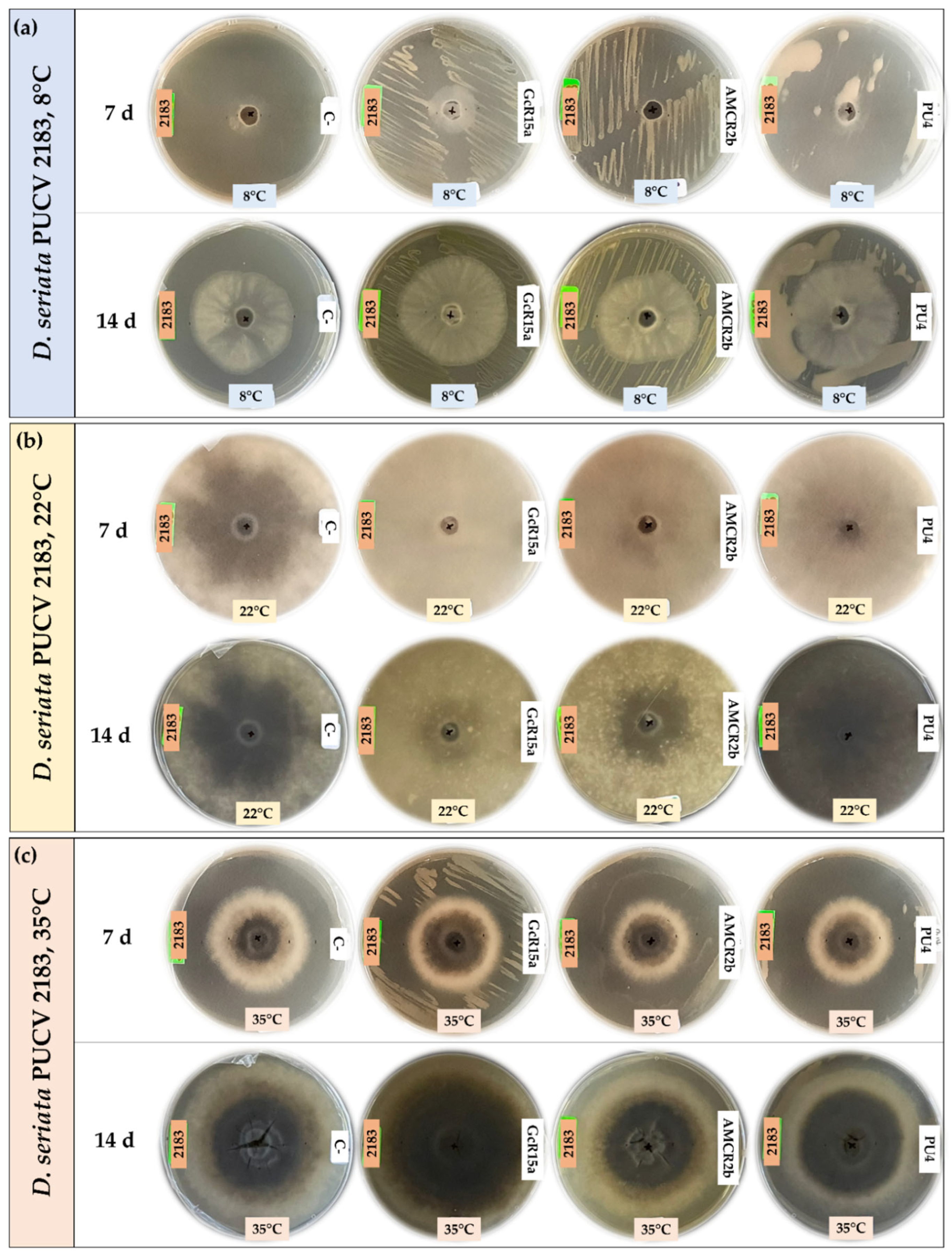
Figure 8.
Effects of native bacteria on the diameter of Diplodia seriata isolates by the double plate method at different temperatures. (a-c) Effects of native bacteria on the diameter of the isolates of D. seriata at 8°C after 7 and 14 days. (d-f) Effects of native bacteria on the inner radius of the isolates of D. seriata at 35°C after 7 and 14 days. There was no decrease in the diameter of the isolates of D. seriata with any treatments at 22°C, and the fungi grew entirely from 7 days (8.5 cm). Means with different letters indicate significant differences (p < 0.05), and uppercase and lowercase letters correspond to 7 and 14 days, respectively. Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
Figure 8.
Effects of native bacteria on the diameter of Diplodia seriata isolates by the double plate method at different temperatures. (a-c) Effects of native bacteria on the diameter of the isolates of D. seriata at 8°C after 7 and 14 days. (d-f) Effects of native bacteria on the inner radius of the isolates of D. seriata at 35°C after 7 and 14 days. There was no decrease in the diameter of the isolates of D. seriata with any treatments at 22°C, and the fungi grew entirely from 7 days (8.5 cm). Means with different letters indicate significant differences (p < 0.05), and uppercase and lowercase letters correspond to 7 and 14 days, respectively. Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4.
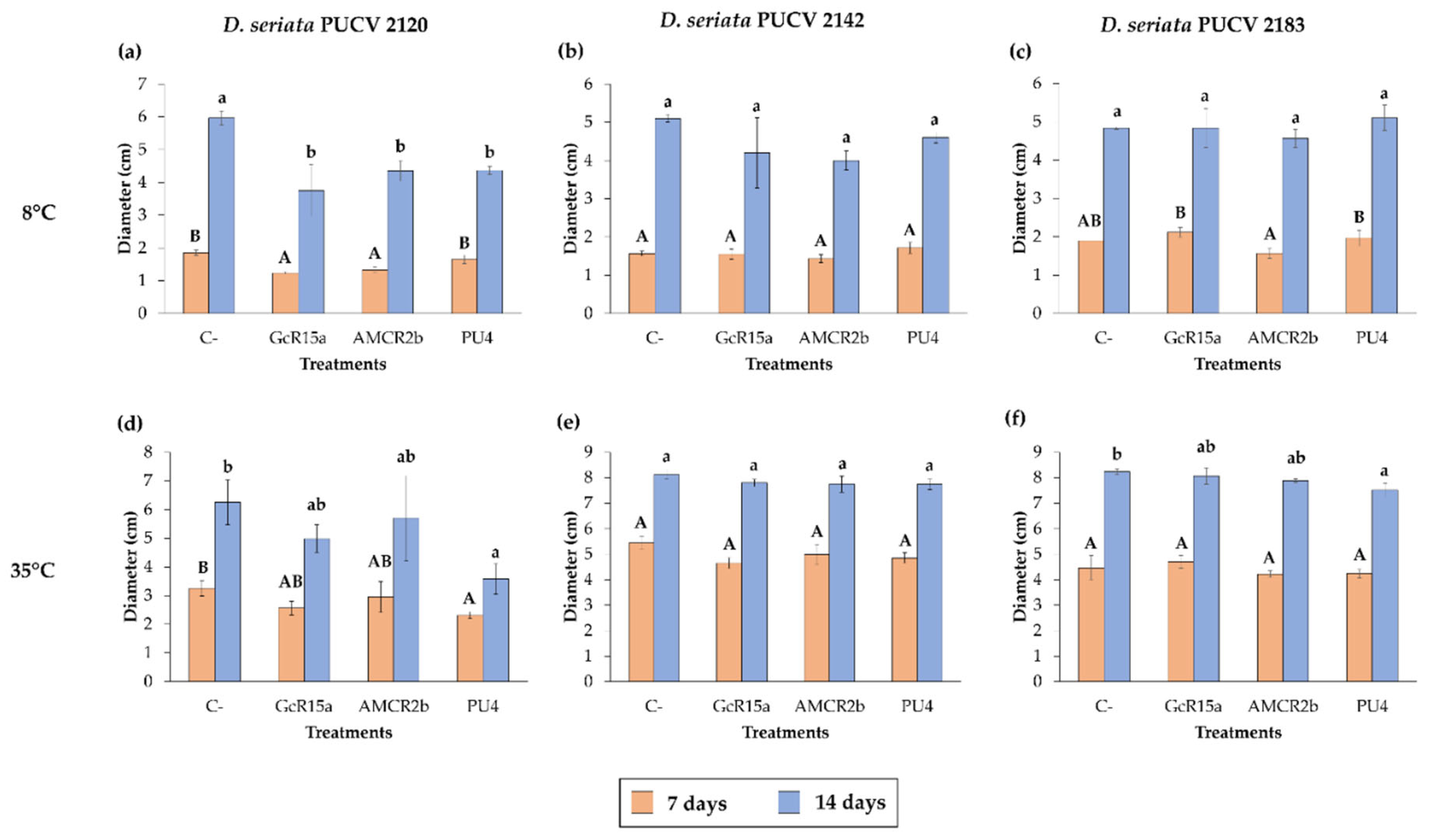
Figure 9.
Effects of Diplodia seriata isolates on grapevine pruning material preinoculated with native bacteria at different temperatures. (a-c) Effects of preinoculation with native bacteria on the vascular lesion length of grapevine pruning material inoculated with D. seriata isolates at 8°C. (d-f) Effects of preinoculation with native bacteria on the vascular lesion length of grapevine pruning material inoculated with D. seriata isolates at 22°C. Means with different letters indicate significant differences (p < 0.05). Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Figure 9.
Effects of Diplodia seriata isolates on grapevine pruning material preinoculated with native bacteria at different temperatures. (a-c) Effects of preinoculation with native bacteria on the vascular lesion length of grapevine pruning material inoculated with D. seriata isolates at 8°C. (d-f) Effects of preinoculation with native bacteria on the vascular lesion length of grapevine pruning material inoculated with D. seriata isolates at 22°C. Means with different letters indicate significant differences (p < 0.05). Abbreviations: C-, negative control; GcR15a, Pseudomonas sp. GcR15a; AMCR2b, Pseudomonas sp. AMCR2b; PU4, Rhodococcus sp. PU4; C+, positive control (tebuconazole).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



