
Submitted:
29 November 2023
Posted:
30 November 2023
You are already at the latest version
Abstract
Hepatitis C virus (HCV) is diffused worldwide, and it is responsible for potentially severe chronic liver disease and primary liver cancer. Chronic infection remains for life if not spontaneously eliminated and viral persistence profoundly impairs the efficiency of host’s immunity. Attempts have been made to develop an effective vaccine, but efficacy trials have met with failure. The availability of highly efficacious direct acting antivirals (DAA) has shed hopes for progressive elimination of chronic HCV infection; however, this approach requires a global monumental effort. Moreover, DAA treatment does not completely restore the normal immunologic homeostasis. Here we discuss the main immunological features of immune responses to HCV and the epigenetic scars that chronic viral persistence leaves behind.
Keywords:
HCV
; Innate immunity
; Adaptive immunity
1. Introduction
Hepatitis C virus (HCV) is responsible for persistent infection in most exposed individuals. After several years of a largely asymptomatic course, chronic hepatitis may develop into cirrhosis and hepatocellular carcinoma (HCC) in a proportion of patients [1]. Symptomatic acute infection is rarely observed and recovers spontaneously in about a third of cases. The mechanisms responsible for chronically evolving infection are still poorly understood, despite host’s immune responses to HCV having been extensively explored (reviewed in [2]). One of the reasons that may explain the aura of mystery surrounding HCV is the lack of a reliable animal model of disease that accurately reproduces the natural history of infection in humans. Indeed, this rather peculiar Flaviviridae family member has evolved with the human host like enemies living under one roof, ensuring pathogen persistence but, at the same time, refraining from being aggressive to secure long-term survival of the human species. In line with this view, HCV usually causes chronic low-level inflammation in the host while, at the same time, outpaces the host’s immune surveillance [3] by establishing a swarm of diverse viral variants (quasispecies) and via suppression or alterations of innate and adaptive immunity. Interestingly, HCV does not confer immunity to reinfection, and it is common thought that this is mainly due to virus variability, resulting in impaired recognition of variants which patients have never been exposed to. However, virus-induced long-term impairment of immune responses results in T-cell exhaustion which often persists even after cure [4,5]. Moreover, reduced production of interferon (IFN)-γ has been shown to contribute to increased susceptibility to HCV reinfection [6].
The complex strategies employed by HCV to evade the immune system's detection can lead to alterations in how immune cells function and interact, leaving behind residual effects, akin to scars, within the immune system's memory. The advent of Direct-Acting Antivirals (DAAs) marks a pivotal turning point in the management of HCV infection, particularly following the introduction of second-generation DAAs that target different viral proteins. This treatment regimen is well-tolerated, of short duration (8-12 weeks), and results in eradication rates nearing 100%. However, there is still a residual risk of developing HCC [7,8]. Moreover, previous studies performed after IFN-based therapies, showed a complete or partial persistence of T cell dysfunction after SVR [9,10], raising the question of whether DAA therapies can restore viral-induced immune dysfunction after years of chronic infection. Theoretically, diverse mechanisms may be triggered to reverse immune dysfunctions, including DAA-induced diminution of viral antigens and viral load [11]. However, a body of evidence indicates that not all consequences of chronic hepatitis C are completely reversible following sustained virologic response.
The purpose of this review to examine immunological aspects of HCV-host relationship and residual impairment of selected cells involved in antiviral immunity after cure with DAAs.
2. HCV-Host relationship: a mutual endurance
2.1. Innate immunity: friend or foe?
Viruses, particularly those responsible for persistent infection, have a remarkable ability to become adapted to different environments. In the case of RNA viruses, such as HCV, this is in part mediated by their high mutational rates, allowing for the rapid selection of variants that overcome hostile environments. These are initially represented by early innate defense mechanisms which are triggered immediately after infection and have the function to limit the extent of microbial spread. Recognition of pathogens occurs through a series of receptors that sense regular patterns of molecular structure shared by many micro-organisms but are not present on the host’s own cells. These patterns and the receptors involved in their recognition are called pathogen-associated molecular patterns [PAMP] and pathogen recognition receptors [PRR], respectively. The innate signaling receptors consist of a tetrad of PRR relevant to viruses: i) Toll-like receptors [TLRs] which sense all microbes; ii) retinoic acid-inducible gene I [RIG-I]-like RNA helicases [RLH] and iii) melanoma differentiation-associated 5 [MDA-5] both of which predominantly sense viruses; iv) nucleotide-binding oligomerization domain [NOD]-like receptors [NLRs] which sense bacteria and viruses [12]. When entering the cells, viruses usually elicit a robust innate immune response unleashing the intracellular IFN cascade leading to type I IFN production [13]. HCV is no exception since cytosolic HCV RNA induces a brisk interferon-stimulated gene response in the liver.
Upon acute HCV infection of hepatocytes, early HCV-RNA recognition by PRR induces interferon (IFN) type I (IFN-α/β) and type III (IFN-λ) gene transcription through IFN regulatory factor (IRF) 3 phosphorylation, dimerization, and nuclear translocation. Following recognition of secreted IFNs by cognate receptors, JAK/STAT-mediated pathways activate the expression of IFN-stimulated genes (ISGs), to induce an antiviral state. Interestingly, HCV seems to ignore early innate defense mechanisms, as it replicates almost immediately after penetration into target cells, suggesting that innate immunity is unable to control virus infection. Indeed, despite HCV flooding the hepatocytes with type I IFN, some HCV proteins interfere with IFN regulatory proteins resulting in impaired signal transmission. Indeed, NS3/4A cleaves mitochondrial antiviral signaling (MAVS) [14] and TIR-domain containing adapter-inducing IFN-β (TRIF) adapter protein [15] resulting in failure to activate IRF3 with consequent impaired activation of downstream target genes, including IFN-β. Moreover NS3/4A protein may reduce E3 ubiquitin ligase Riplet-induced RIG-I activation [16,17] and NS3 is able to block interaction between TBK1 and IRF3 inhibiting IRF3 activation [18]. Another mechanism exploited by the virus to inactivate IFN-β production is mediated by the NS4B protein, which blocks RIG-I-induced activation of IFN-β production through binding to stimulator of interferon genes (STING) and blocking STING–MAVS interaction [19,20]. NS4B may also inhibit TLR3-mediated interferon signaling pathway inducing the degradation of TRIF [21]. Moreover, NS5A plays a role in the reduction of RIG I and TLR3 mediated responses [22,23]. It has been recently shown that NS5A is able to interfere with MAVS activity through binding to mitochondrial-associated protein leucine-rich pentatricopeptide repeat-containing (LRPPRC) by exploiting the ability of LRPPRC to inhibit MAVS-regulated antiviral signaling [24]. Other mechanisms of immune evasion are mediated by the HCV core protein, which can directly bind to STAT1, blocking STAT1/STAT2 heterodimerization and thus inhibiting IFN signal transduction [25-27]. A role in the inhibition of antiviral functions has been also reported for E2 [28] and p7 [29] proteins.
However, it is important to emphasize that interference of HCV proteins with type I IFN signaling described above has been observed upon expression of single HCV proteins in cell lines and not in virus-infected cells, thus limiting the biological significance of such experiments.
2.2. T-cell exhaustion disrupts adaptive immunity and sets the stage for HCV persistence
As in other virus infections, a rapid and efficient activation of the different components of the innate immune system is crucial not only for the initial containment of virus replication and spread, but also for a timely and efficient promotion of downstream adaptive responses, which require more time for their induction but are essential for eradication or at least long-term control of infection [30]. The evidence that HCV can affect innate immune function [31], as outlined above, may have important implications with respect to priming and maturation of HCV-specific adaptive responses.
The importance of HCV-specific T cells in facilitating viral clearance is supported by the observations that self-limiting HCV infection is associated with vigorous and multi-specific T helper and cytotoxic T cell responses [32,33] and that depletion of CD8 T cells in HCV infected chimpanzees prevents HCV eradication in this animal model [34]. This evidence is further supported by work that identified transcriptional differences between HCV-specific CD8 T cells from patients with chronically evolving acute HCV infection and those with self-limiting infection [35]. CD4+ T cells were found to play a key role in infection control, being required for CD8 T cell-mediated HCV clearance. However, in patients unable to clear HCV infection HCV-specific CD4 T cells rapidly developed proliferative defects followed by their deletion [36] in patients with persisting viremia.
However, signs of progressive CD8+ T-cell exhaustion start to appear after persisting viremia is established, with viral escape mutations in CD8 epitopes regions leading to diminished recognition of the increasingly complex circulating quasispecies [37]. T-cell exhaustion is typically induced by prolonged exposure to antigens during chronic infection. Mechanisms that may reverse T cell exhaustion include the reduction of inhibitory molecules expression, such as PD1, CTLA-4, TIGIT, and TIM-3 which are known to inhibit immune responses via binding to their ligands expressed on infected cells [38]. Indeed, successful immune therapies based on the use of antibodies blocking these molecules have demonstrated the potential for the reversal of T cell exhaustion, also in the context of HCV infection [39,40].
2.3. B-cell responses are not protective but play a role in lymphoproliferative disorders
The role of antibodies in controlling HCV is far from being clarified. Antibody responses to structural and non-structural viral proteins appear 1-2 months after acute infection and their presence appears to correlate with ongoing infection rather than protection [41]. Neutralizing antibodies are certainly generated in the HCV infected host; however, they do not seem to represent the main driving force of HCV control. This poses a serious challenge to the development of an effective traditional prophylactic vaccine and cast doubts as to the role of humoral immunity in recovery. For these reasons, the ideal objective of attaining sterilizing protective immunity remains elusive. Indeed, HCV has developed many strategies to evade humoral control [42] and although there are reasons to believe in the development of a HCV vaccine instead of pursuing difficult to realize elimination programs with DAAs, one attempt at assessing the protective efficacy of a vaccine has met with failure [43].
Beside a possible role of B cells in generating HCV neutralizing antibodies, there is evidence that B cells are chronically activated in persistent HCV infection and that this phenomenon may be responsible for the extrahepatic manifestations, particularly leukocytoclastic vasculitis and lymphoproliferative disorders, arising from chronic antigenic stimulation. Current evidence points to the possible role of HCV E2 envelope protein-CD81 interaction which may reduce the activation threshold of B cells, potentially leading to polyclonal and, eventually, oligo and monoclonal expansion of B lymphocytes. Polyclonal B cell activation is a typical feature of chronic HCV infection and is associated with upregulation of B-cell activation molecules [44,45]. In depth analysis of B-cell subsets in patients with chronic HCV infection reveals increments in the frequencies of atypical memory B cells resulting in a skewed B-cell profile characterized by exhaustion markers, in keeping with continuous antigenic stimulation [46]. Surprisingly, however, bulk memory B cells maintain the ability to be activated in vitro upon exposure to HCV and efficiently respond to innate and adaptive immune stimuli [45,47], suggesting that chronic HCV infection does not pervasively affect the B-cell function.
3. Can direct-acting antivirals (DAAs) reverse HCV-altered immune responses?
3.1. Effect of DAA treatment on liver inflammation and fibrosis
Upon development of chronic infection, the effort to contrast viral spread yields an exaggerated and persistent activation of antiviral mechanisms which set the stage for the development of an inflammatory environment within the liver, ultimately contributing to the onset of fibrosis. Indeed, HCV-induced liver injury determines activation of hepatic stellate cells, leading to deposition of extracellular matrix components, which are responsible of liver fibrosis [48]. Many patients who achieve HCV eradication following DAA treatment have endured several years of chronic infection and may have developed advanced stages of liver fibrosis. Of note, DAA therapy can lead to a reduction in ISG expression [49,50], indicating a potential restoration of the HCV-associated inflammatory environment to a more typical state. However, some studies have shown that soluble inflammatory mediators remain elevated even after successful completion of DAA treatment [51], including cases in whom treatment was initiated during the early phase of infection [52]. In addition, certain molecules, such as IL17, which were shown to be reduced during HCV infection, did not increase after DAA treatment [51]. Similar findings have been reported in cases of viral clearance following liver transplantation [53]. Notably, an inflammatory milieu was associated with HCC development in cirrhotic HCV patients after DAA treatment [54,55].
Different studies documented significant fibrosis regression following DAA therapy [56-59]. Interestingly, HCV eradication by DAAs was shown to reduce systemic oxidative stress in patients with advanced liver fibrosis. [60], while others identified serum angiopoietin-2 [Ang-2] levels as a predictor of regression of liver fibrosis after successful HCV eradication by DAAs [61]. Moreover, serum CXCL10 levels predicted fibrosis regression after DAA treatment in patients with baseline levels of Mac-2 binding protein glycosylation isomer (M2BPGi) higher than the cut off index 2 [62]. Recently, Ferreira et al evaluated liver fibrosis before and after DAA-induced HCV clearance and found that the reduction of liver fibrosis after DAA treatment was associated with a specific TNFα and IL10 genotype. Specifically, the frequency of patients in whom an improved fibrosis stage was documented was higher in those with a TNFα 308 G/A genotype or TT for IL10 1082 T/C and GG genotype for 592 G/T [63].
3.2. Controversial recovery of NK cell function after HCV cure
Beside type I IFN-mediated immunity, innate immune responses include several cellular lineages. Natural killer (NK) cells play an important role in the control of viral infections and have been shown to undergo significant phenotypic and functional changes at all stages of HCV infection, contributing to liver cell damage. In the acute phase of HCV infection, NK cells display an activated phenotype with enhanced cytotoxicity and IFN-γ production [64]. In chronic infection, prolonged exposure to large amounts of endogenous IFN-α produced by infected hepatocytes results in polarization toward cytotoxicity with deficient IFN-γ secretion [65]. This is caused by type I IFN-induced phosphorylation of signal transducer and activation of transcription [STAT] 1, which displaces STAT4 at the IFN-α/β receptor resulting in decreased pSTAT4-dependent IFN-γ production and increased pSTAT1-dependent cytotoxicity [66]. Increased expression of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is also upregulated in this context. The consequence of this virus-induced altered signaling is a NK cell “functional dichotomy” characterized by enhanced NK cytolytic activity and a failure to produce adequate amounts of IFN-γ and tumor necrosis factor [TNF]-α, contributing to the inability to eradicate HCV [65,67]. This profound alteration of NK cell homeostasis has been shown to be reversible upon viral eradication by treatment with DAAs [68-73], although NK cell functional recovery has not been confirmed in cirrhotic patients [74]. On the other hand, phenotypic and functional recovery of a particular NK cell subset named adaptive/memory was observed in patients with advanced fibrosis treated with DAAs [75]. Upregulation of PD-1 and reduced antibody-dependent cell-mediated cytotoxicity (ADCC) was completely restored after viral cure [76] Although most studies suggest NK cell functional recover after cure, as expected from NK cells that react promptly to virus induced activation, there is still uncertainty as to possible residual functional defects after HCV eradication. For instance, a stochastic neighbor embedding analysis showed that HCV infection leaves an imprint on NK cells that is not reversed by DAA [77] and down-regulation of FcγRIII, typically observed in chronic HCV infection was not completely restored several months after DAA cure [78].
3.3. T-cell exhaustion is only partially reversed after HCV cure and leaves indelible epigenetic scars
Initial reports documented a reversal of T cell exhaustion post-DAA treatment, characterized by enhanced proliferation of HCV-specific CD8+ T cells [79], diminished PD-1 expression [80-82], and a transition toward a TCF-1+CD127+ memory-like T cell phenotype [83]. However, one study found that only partial restoration of immune responses could be obtained in HCV patients treated with DAA, with increased proliferation of HCV-specific CD4 and CD8 T-cells, accompanied by an impaired ability to secrete IFNγ and IL2 [84]. Also, a recent study highlighted the importance of early DAA treatment commencement, during the acute phase of HCV infection, which led to a reduction of immune exhaustion and to stronger HCV-specific T-cell responses after treatment, thus reducing the risk of possible reinfection [85].
A previous study also revealed that recovery from exhaustion is related to the duration of T cell stimulation, with prolonged antigenic stimulation being associated with an irreversible state of exhaustion [86]. The authors showed that antigen removal by successful DAA treatment induced phenotypic changes in exhausted T cells, which seem differentiated toward a memory like profile. However, these changes did not translate into a functional recovery, since the level of transcriptional regulators typical of exhausted cells, such as TOX, a key driver of T cell exhaustion, did not normalize after cure [86]. Failure of normalization of transcriptional regulators of T cell exhaustion represents an immunological scar which, intriguingly, is not detectable in T cells exposed to antigen for a limited time [86]. In line with this, another study used single-cell transcriptomics to demonstrate that, despite terminally exhausted HCV specific CD8 T cells decreased after DAA, a signature of exhaustion persisted after cure [87]. Epigenetic studies analyzing the chromatin accessibility explained the reasons behind this apparently irreversible state of exhaustion. For instance, Yates et al discovered a similar epigenetic profile in response to chronic stimulation in antigen-specific exhausted CD8 T cells in multiple human viral infections, including HCV, and showed that this epigenetic state of exhaustion signature persists for over a year after DAA-induced virus eradication [88]. Epigenetic scars included super-enhancer elements near exhaustion-associated key transcription factors TOX and HIF1A [88]. In agreement with the above, other studies showed that checkpoint-blockade immunotherapy did not fundamentally reverse exhaustion-associated epigenetic changes, supporting the epigenetic irreversibility of exhausted T cells [89,90]. Therefore, HCV infection leaves an epigenetic signature on the host chromatin that is not fully reversed following DAA-induced virus eradication, resulting in a state of exhaustion in HCV-specific CD8 T cells that does not offer protection from subsequent viral reinfection, as demonstrated in a non-human model [91]. Interestingly, among the epigenetic changes, DNA methylation status has been associated with aging. Specifically, it has been shown that the gain or loss of CpG methylation over time is a valid method to estimate biological age [92]. Analysis of the DNA-methylation status in PBMCs to calculate the epigenetic age acceleration [EAA] in chronic HCV infection showed that HCV infection induces an acceleration of epigenetic age which was only partially reverted by DAA treatment during a long-term follow-up. Moreover, individuals who developed HCC post-eradication showed the most pronounced epigenetic aging acceleration without signs of reversal [93].
Interestingly, the severity of T cell exhaustion was shown to be associated with the metabolism of HCV specific T cells, identifying enolase as a metabolic regulator of severely exhausted T cells. Thus, in chronic HCV infection, severely exhausted CD8 T cells exhibited mitochondrial impairment linked to elevated liver inflammation, and reduced enolase activity [94]. Notably, DAA therapy partially improved mitochondrial polarization in HCV-specific CD8+ T cells [94]. These findings are akin to those of others who also observed improved mitochondrial polarization in HCV-specific CD8+ T cells following viral eradication [5]. In contrast, others did not observe a mitochondrial functional recovery in HCV-specific CD8+ T cell metabolism [95].
Few data are available on resolution of CD4 T cell perturbations after HCV cure. In one study, DAA treatment did not lead to any increment in the proliferative ability and cytokine production of HCV specific CD4 T cells [33] while others showed a significantly increased frequency of HCV specific CD4 T cells with reduced PD1 expression [96,97].
3.4. B cell activation may be reversed by DAAs even though cryoglobulins may persist for years after recovery
DAA treatment of cryoglobulinemia arising from chronic B-cell activation leads to remission in approximately half of the patients, as shown in a large Italian study, which may be associated with a significant relapse rate [98]. Indeed, complete remission is reached in only about 40% of patients. Of note, the presence of pretreatment peripheral neuropathy, weakness, sicca syndrome and renal involvement have been found to be reliable predictors of poor clinical response in patients with HCV-related cryoglobulinemic vasculitis [99]. After HCV eradication, circulating B-cell clones can persist in the absence of clinical or biochemical relapse, suggesting that those clones could have switched to a quiescent state [100]. Infections, cancer or other endogenous or exogenous factors could trigger the proliferations of those B-cells [101], in which co-stimulation of BCR by surrogate immune complexes and of TLR9 by CpG DNA sequences has led to clonal expansion of pathogenic quiescent B cells from HCV-cured patients with cryoglobulinemia.
The effect of DAA treatment on B cells in patients with chronic HCV infection has been explored in a limited number of studies. T-bet+ B cells are usually expanded during chronic HCV infection and coincide with a tissue-like memory, or atypical memory, phenotype [46] and return to physiologic levels after HCV cure after DAA therapy [102]. Similarly, HCV-specific memory B cells persisted, displayed a restored resting phenotype, normalized chemokine receptor expression and preserved the ability to differentiate into antibody-secreting cells after cure [103].
4. Conclusions
In conclusion, despite DAA treatment ability to easily eradicate HCV, numerous unresolved questions persist. Figure 1 summarizes phenotypic and functional features of host’s immunity in chronic HCV infection before and after cure. First, eradicating HCV does not confer protective immunity against subsequent infections or eliminate the risk of developing hepatocellular carcinoma (HCC). Impaired immune responses developed over years of chronic infection do not fully recover after DAA treatment. Indeed, despite the partial reversal of T-cell exhaustion markers after HCV cure, there is evidence suggesting an incomplete reversal of epigenetic changes associated with T-cell exhaustion. These changes persist in HCV-specific CD8 T cells after successful DAA-induced virus eradication, leaving individuals potentially vulnerable to reinfection. Furthermore, DAA treatment appears to improve Natural Killer (NK) cell functions, particularly in non-cirrhotic patients. However, complete functional recovery of NK cells in cirrhotic patients remains uncertain, leaving certain subsets unable to fully restore their functionality post-DAA treatment. Additionally, while DAA treatment holds promise in reducing liver inflammation by potentially reversing the HCV-associated inflammatory environment, some studies indicate persistently elevated levels of soluble inflammatory mediators even after successful DAA treatment. The connection between the inflammatory milieu and the development of HCC post-DAA treatment in cirrhotic patients leave significant concerns over long-term prognosis in this setting.
In summary, while the beneficial effects of DAA treatment of HCV infection are undisputable, concerns persist regarding complete recovery of innate and adaptive immunity, especially in cirrhotic patients, with possible implications on susceptibility to hepatic and extra-hepatic cancer.
Author Contributions
SV and MUM provided substantial contribution to the conception and design of the review article and wrote the manuscript. BO, SM, SO, AC, DM revised the manuscript critically and provided important intellectual input. All authors critically read, edited, and approved the final version of the manuscript.
Conflicts of Interest
None of the authors declared a conflict of interest.
References
- Martinello, M.; Naggie, S.; Rockstroh, J.; Matthews, G. V. Direct-Acting Antiviral Therapy for Treatment of Acute and Recent Hepatitis C Virus Infection: A Narrative Review. Clinical Infectious Diseases 2023, 77 (Supplement_3), S238–S244. [CrossRef]
- Ferrari, C.; Barili, V.; Varchetta, S.; Mondelli, M.U. Immune mechanisms of viral clearance and disease pathogenesis during viral hepatitis. In Liver Biology and Pathobiology, 6th ed.; Arias IM, Alter HJ, Boyer JL, Cohen DE, Shafritz DA, Thorgeirsson SS, Wolkoff AW, Eds.; Wiley Blackwell, Oxford UK, 2020; pp. 821-850.
- Thimme, R.; Oldach, D.; Chang, K.M.; Steiger, C.; Ray, S.C.; Chisari, F.V. Determinants of viral clearance and persistence during acute hepatitis C virus infection. J Exp Med. 2001, 19;194(10):1395-406. [CrossRef]
- Penna, A.; Pilli, M.; Zerbini, A.; Orlandini, A.; Mezzadri, S.; Sacchelli, L.; Missale, G.; Ferrari, C. Dysfunction and functional restoration of HCV-specific CD8 responses in chronic hepatitis C virus infection. Hepatology. 2007, 45(3):588-601. [CrossRef]
- Barili, V.; Fisicaro, P.; Montanini, B.; Acerbi, G.; Filippi, A.; Forleo, G.; Romualdi, C.; Ferracin, M.; Guerrieri, F.; Pedrazzi, G.; Boni, C.; Rossi, M.; Vecchi, A.; Penna, A.; Zecca, A.; Mori, C.; Orlandini, A.; Negri, E.; Pesci, M.; Massari, M.; Missale, G.; Levrero, M.; Ottonello, S.; Ferrari, C. Targeting P53 and Histone Methyltransferases Restores Exhausted CD8+ T Cells in HCV Infection. Nature Communications 2020, 11 (1). [CrossRef]
- Osburn, W.O.; Fisher, B.E.; Dowd, K.A.; Urban, G.; Liu, L.; Ray, S.C.; Thomas, D.L.; Cox, A.L. Spontaneous control of primary hepatitis C virus infection and immunity against persistent reinfection. Gastroenterology. 2010, 138(1):315-24. [CrossRef]
- Ono, A.; Goossens, N.; Finn, R.S.; Schmidt, W.N.; Thung, S.N.; Im, G.; Hoshida, Y. Precision Liver Cancer Prevention Consortium. Persisting risk of hepatocellular carcinoma after hepatitis C virus cure monitored by a liver transcriptome signature. Hepatology. 2017 66(4):1344-1346. [CrossRef]
- Romano, A.; Angeli, P.; Piovesan, S.; Noventa, F.; Anastassopoulos, G.; Chemello, L.; Cavalletto, L.; Gambato, M.; Russo, F. P.; Burra, P.; Vincenzi, V.; Scotton, P. G.; Panese, S.; Tempesta, D.; Bertin, T.; Carrara, M.; Carlotto, A.; Capra, F.; Carolo, G.; Scroccaro, G.; Alberti, A. Newly Diagnosed Hepatocellular Carcinoma in Patients with Advanced Hepatitis C Treated with DAAs: A Prospective Population Study. Journal of Hepatology 2018, 69 (2), 345–352. [CrossRef]
- Missale, G.; Pilli, M.; Zerbini, A.; Penna, A.; Ravanetti, L.; Barili, V.; Orlandini, A.; Molinari, A.; Fasano, M.; Santantonio, T.; Ferrari, C. Lack of Full CD8 Functional Restoration after Antiviral Treatment for Acute and Chronic Hepatitis C Virus Infection. Gut 2012, 61 (7), 1076–1084. [CrossRef]
- Abdel-Hakeem, M. S.; Bédard, N.; Badr, G.; Ostrowski, M.; Sékaly, R. P.; Bruneau, J.; Willems, B.; Heathcote, E. J.; Shoukry, N. H. Comparison of Immune Restoration in Early versus Late Alpha Interferon Therapy against Hepatitis C Virus. Journal of Virology 2010, 84 (19), 10429–10435. [CrossRef]
- Heim, M. H.; Thimme, R. Innate and Adaptive Immune Responses in HCV Infections. Journal of Hepatology 2014, 61 (1), S14–S25. [CrossRef]
- Kawai, T.; Akira, S. The Roles of TLRs, RLRs and NLRs in Pathogen Recognition. International Immunology 2009, 21 (4), 317–337. [CrossRef]
- Su, A. I.; Pezacki, J. P.; Wodicka, L.; Brideau, A. D.; Supeková, L.; Thimme, R.; Wieland, S.; Bukh, J.; Purcell, R. H.; Schultz, P. G.; Chisari, F. V. Genomic Analysis of the Host Response to Hepatitis C Virus Infection. Proceedings of the National Academy of Sciences of the United States of America 2002, 99 (24), 15669–15674. [CrossRef]
- Loo, Y.; Owen, D. M.; Li, K.; Erickson, A. K.; Johnson, C.; Fish, P. M.; Carney, D. S.; Wang, T.; Ishida, H.; Yoneyama, M.; Fujita, T.; Saito, T.; Lee, W. M.; Hagedorn, C. H.; Lau, D. T. Y.; Weinman, S. A.; Lemon, S. M.; Gale, M. Viral and Therapeutic Control of IFN-β Promoter Stimulator 1 during Hepatitis C Virus Infection. Proceedings of the National Academy of Sciences of the United States of America 2006, 103 (15), 6001-6006. [CrossRef]
- Horner, S. M.; Liu, H. M.; Park, H. S.; Briley, J.; Gale, M. Mitochondrial-Associated Endoplasmic Reticulum Membranes (MAM) Form Innate Immune Synapses and Are Targeted by Hepatitis C Virus. Proceedings of the National Academy of Sciences of the United States of America 2011, 108 (35), 14590- 14595. [CrossRef]
- Oshiumi, H.; Miyashita, M.; Matsumoto, M.; Seya, T. A Distinct Role of Riplet-Mediated K63-Linked Polyubiquitination of the RIG-I Repressor Domain in Human Antiviral Innate Immune Responses. PLOS Pathogens 2013, 9 (8), e1003533. [CrossRef]
- Vazquez, C.; Tan, C. Y.; Horner, S. M. Hepatitis C Virus Infection Is Inhibited by a Noncanonical Antiviral Signaling Pathway Targeted by NS3-NS4A. Journal of Virology 2019, 93 (23). [CrossRef]
- Otsuka, M.; Kato, N.; Moriyama, M.; Taniguchi, H.; Wang, Y.; Dharel, N.; Kawabe, T.; Omata, M. Interaction between the HCV NS3 Protein and the Host TBK1 Protein Leads to Inhibition of Cellular Antiviral Responses. Hepatology 2005, 41 (5), 1004–1012. [CrossRef]
- Nitta, S.; Sakamoto, N.; Nakagawa, M.; Kakinuma, S.; Mishima, K.; Kusano-Kitazume, A.; Kiyohashi, K.; Murakawa, M.; Nishimura-Sakurai, Y.; Azuma, S.; Tasaka-Fujita, M.; Asahina, Y.; Yoneyama, M.; Fujita, T.; Watanabe, M. Hepatitis C Virus NS4B Protein Targets STING and Abrogates RIG-I-Mediated Type I Interferon-Dependent Innate Immunity. Hepatology 2013, 57 (1), 46–58. [CrossRef]
- Ding, Q.; Cao, X.; Lu, J.; Huang, B.; Liu, Y. J.; Kato, N.; Shu, H.; Zhong, J. Hepatitis C Virus NS4B Blocks the Interaction of STING and TBK1 to Evade Host Innate Immunity. Journal of Hepatology 2013, 59 (1), 52–58. [CrossRef]
- Liang, Y.; Cao, X.; Ding, Q.; Zhao, Y.; He, Z.; Zhong, J. Hepatitis C Virus NS4B Induces the Degradation of TRIF to Inhibit TLR3-Mediated Interferon Signaling Pathway. PLOS Pathogens 2018, 14 (5), e1007075. [CrossRef]
- Hiet, M.; Bauhofer, O.; Zayas, M.; Roth, H.; Tanaka, Y.; Schirmacher, P.; Willemsen, J.; Grünvogel, O.; Bender, S.; Binder, M.; Lohmann, V.; Lotteau, V.; Ruggieri, A.; Bartenschlager, R. Control of Temporal Activation of Hepatitis C Virus-Induced Interferon Response by Domain 2 of Nonstructural Protein 5A. Journal of Hepatology 2015, 63 (4), 829–837. [CrossRef]
- Çevik, R. E.; Cesarec, M.; Da Silva Filipe, A.; Licastro, D.; McLauchlan, J.; Marcello, A. Hepatitis C Virus NS5A Targets Nucleosome Assembly Protein NAP1L1 To Control the Innate Cellular Response. Journal of Viro ogy 2017, 91 (18). [CrossRef]
- Refolo, G.; Ciccosanti, F.; Di Rienzo, M.; Perdomo, A. B.; Romani, M.; Alonzi, T.; Tripodi, M.; Ippolito, G.; Piacentini, M.; Fimia, G. M. Negative Regulation of Mitochondrial Antiviral Signaling Protein–Mediated Antiviral Signaling by the Mitochondrial Protein LRPPRC during Hepatitis C Virus Infection. Hepatology 2018, 69 (1), 34–50. [CrossRef]
- Heim, M. H.; Moradpour, D.; Blum, H. E. Expression of Hepatitis C Virus Proteins Inhibits Signal Transduction through the Jak-STAT Pathway. Journal of Virology 1999, 73 (10), 8469–8475. [CrossRef]
- Lin, W.; Kim, S. S.; Yeung, E.; Kamegaya, Y.; Blackard, J. T.; Kim, K.-A.; Holtzman, M. J.; Chung, R. T. Hepatitis C Virus Core Protein Blocks Interferon Signaling by Interaction with the STAT1 SH2 Domain. Journal of Virology 2006, 80 (18), 9226–9235. [CrossRef]
- Lin, W.; Choe, W. H.; Hiasa, Y.; Kamegaya, Y.; Blackard, J. T.; Schmidt, E. V.; Chung, R. T. Hepatitis C Virus Expression Suppresses Interferon Signaling by Degrading STAT1. Gastroenterology 2005, 128 (4), 1034–1041. [CrossRef]
- Taylor, D. J.; Shi, S. T.; Romano, P. R.; Barber, G. N.; Lai, M. M. C. Inhibition of the Interferon- Inducible Protein Kinase PKR by HCV E2 Protein. Science 1999, 285 (5424), 107–110. [CrossRef]
- Qi, H.; Chu, V.; Wu, N. C.; Chen, Z.; Truong, S.; Brar, G.; Su, S.; Du, Y.; Arumugaswami, V.; Olson, C. A.; Chen, S. H.; Lin, C. Y.; Wu, T.; Sun, R. Systematic Identification of Anti-Interferon Function on Hepatitis C Virus Genome Reveals P7 as an Immune Evasion Protein. Proceedings of the National Academy of Sciences of the United States of America 2017, 114 (8), 2018–2023. [CrossRef]
- Rehermann, B.; Nascimbeni, M. Immunology of Hepatitis B Virus and Hepatitis C Virus Infection. Nature Reviews Immunology 2005, 5 (3), 215–229. [CrossRef]
- Gale, M.; Foy, E. Evasion of Intracellular Host Defence by Hepatitis C Virus. Nature 2005, 436 (7053), 939–945. [CrossRef]
- Neumann-Haefelin, C.; Thimme, R. Success and Failure of Virus-Specific T Cell Responses in Hepatitis C Virus Infection. Digestive Diseases 2011, 29 (4), 416–422. [CrossRef]
- Hartnell, F.; Esposito, I.; Swadling, L.; Brown, A.; Phetsouphanh, C.; De Lara, C.; Gentile, C.; Turner, B. L.; Dorrell, L.; Capone, S.; Folgori, A.; Barnes, E.; Klenerman, P. Characterizing Hepatitis C Virus–Specific CD4+ T Cells Following Viral-Vectored Vaccination, Directly Acting Antivirals, and Spontaneous Viral Cure. Hepatology 2020, 72 (5), 1541–1555. [CrossRef]
- Ramalingam, R.; Meyer-Olson, D.; Shoukry, N. H.; Bowen, D. G.; Walker, C. M.; Kalams, S. A. Kinetic Analysis by Real-Time PCR of Hepatitis C Virus (HCV)-Specific T Cells in Peripheral Blood and Liver after Challenge with HCV. Journal of Virology 2008, 82 (21), 10487–10492. [CrossRef]
- Wolski, D.; Foote, P. K.; Chen, D. Y.; Lewis-Ximenez, L. L.; Fauvelle, C.; Aneja, J.; Walker, A.; Tonnerre, P.; Torres-Cornejo, A.; Kvistad, D.; Imam, S.; Waring, M. T.; Tully, D. C.; Allen, T. M.; Chung, R. T.; Timm, J.; Haining, W. N.; Kim, A. Y.; Baumert, T. F.; Lauer, G. M. Early Transcriptional Divergence Marks Virus-Specific Primary Human CD8+ T Cells in Chronic versus Acute Infection. Immunity 2017, 47 (4), 648-663.e8. [CrossRef]
- Wiesch, J. S. Z.; Ciuffreda, D.; Lewis-Ximenez, L. L.; Kasprowicz, V.; Nolan, B. E.; Streeck, H.; Aneja, J.; Reyor, L. L.; Allen, T. M.; Lohse, A. W.; McGovern, B.; Chung, R. T.; Kwok, W. W.; Kim, A. Y.-S.; Lauer, G. M. Broadly Directed Virus-Specific CD4+ T Cell Responses Are Primed during Acute Hepatitis C Infection, but Rapidly Disappear from Human Blood with Viral Persistence. Journal of Experimental Medicine 2012, 209 (1), 61–75. [CrossRef]
- Wildner, N. H.; Walker, A.; Brauneck, F.; Ditt, V.; Peine, S.; Huber, S.; Haag, F.; Beisel, C.; Timm, J.; Wiesch, J. S. Z. Transcriptional Pattern Analysis of Virus-Specific CD8+ T Cells in Hepatitis C Infection: Increased Expression of TOX and Eomesodermin during and after Persistent Antigen Recognition. Frontiers in Immunology 2022, 13. [CrossRef]
- Jenkins, E.; Whitehead, T.; Fellermeyer, M.; Davis, S. J.; Sharma, S. The Current State and Future of T-Cell Exhaustion Research. Oxford Open Immunology 2023, 4 (1). [CrossRef]
- Gardiner, D. F.; Lalezari, J.; Lawitz, E.; DiMicco, M. A.; Ghalib, R.; Reddy, K. R.; Chang, K.; Sulkowski, M. S.; Marro, S. O.; Anderson, J.; He, B.; Kansra, V.; McPhee, F.; Wind-Rotolo, M.; Grasela, D. M.; Selby, M.; Korman, A. J.; Lowy, I. A Randomized, Double-Blind, Placebo-Controlled Assessment of BMS-936558, a Fully Human Monoclonal Antibody to Programmed Death-1 (PD-1), in Patients with Chronic Hepatitis C Virus Infection. PLOS ONE 2013, 8 (5), e63818. [CrossRef]
- Sangro, B.; Gómez-Martín, C.; De La Mata, M.; Iñarrairaegui, M.; Garralda, E.; Barrera, P.; Riezu-Boj, J. I.; Larrea, E.; Alfaro, C.; Sarobe, P.; Lasarte, J. J.; Pérez-Gracia, J. L.; Melero, I.; Prieto, J. A Clinical Trial of CTLA-4 Blockade with Tremelimumab in Patients with Hepatocellular Carcinoma and Chronic Hepatitis C. Journal of Hepatology 2013, 59 (1), 81–88. [CrossRef]
- Takaki, A.; Wiese, M.; Maertens, G.; Depla, E.; Seifert, U.; Liebetrau, A.; Miller, J. L.; Manns, M. P.; Rehermann, B. Cellular Immune Responses Persist and Humoral Responses Decrease Two Decades after Recovery from a Single-Source Outbreak of Hepatitis C. Nature Medicine 2000, 6 (5), 578–582. [CrossRef]
- Cashman, S. B.; Marsden, B. D.; Dustin, L. B. The Humoral Immune Response to HCV: Understanding Is Key to Vaccine Development. Frontiers in Immunology 2014, 5. [CrossRef]
- Page, K.; Melia, M. T.; Veenhuis, R. T.; Winter, M. E.; Rousseau, K. E.; Massaccesi, G.; Osburn, W. O.; Forman, M.; Thomas, E. D.; Thornton, K.; Wagner, K.; Vassilev, V.; Lin, L.; Lum, P. J.; Giudice, L. C.; Stein, E.; Asher, A.; Chang, S.; Gorman, R. L.; Ghany, M. G.; Liang, T. J.; Wierzbicki, M. R.; Scarselli, E.; Nicosia, A.; Folgori, A.; Capone, S.; Cox, A. L. Randomized Trial of a Vaccine Regimen to Prevent Chronic HCV Infection. The New England Journal of Medicine 2021, 384 (6), 541–549. [CrossRef]
- Rosa, D.; Saletti, G.; De Gregorio, E.; Zorat, F.; Comar, C.; D’Oro, U.; Nuti, S.; Houghton, M.; Barnaba, V.; Pozzato, G.; Abrignani, S. Activation of Naïve B Lymphocytes via CD81, a Pathogenetic Mechanism for Hepatitis C Virus-Associated B Lymphocyte Disorders. Proceedings of the National Academy of Sciences of the United States of America 2005, 102 (51), 18544–18549. [CrossRef]
- Oliviero, B.; Cerino, A.; Varchetta, S.; Paudice, E.; Pai, S. S.; Ludovisi, S.; Zaramella, M.; Michelone, G.; Pugnale, P.; Negro, F.; Barnaba, V.; Mondelli, M. U. Enhanced B-Cell Differentiation and Reduced Proliferative Capacity in Chronic Hepatitis C and Chronic Hepatitis B Virus Infections. Journal of Hepatology 2011, 55 (1), 53–60. [CrossRef]
- Oliviero, B.; Mantovani, S.; Ludovisi, S.; Varchetta, S.; Mele, D.; Paolucci, S.; Baldanti, F.; Mondelli, M. U. Skewed B Cells in Chronic Hepatitis C Virus Infection Maintain Their Ability to Respond to Virus-induced Activation. Journal of Viral Hepatitis 2014, 22 (4), 391–398. [CrossRef]
- Sugalski, J. M.; Rodrı́Guez, B.; Moir, S.; Anthony, D. D. Peripheral Blood B Cell Subset Skewing Is Associated with Altered Cell Cycling and Intrinsic Resistance to Apoptosis and Reflects a State of Immune Activation in Chronic Hepatitis C Virus Infection. Journal of Immunology 2010, 185 (5), 3019–3027. [CrossRef]
- Friedman, S. L. Mechanisms of Hepatic Fibrogenesis. Gastroenterology 2008, 134 (6), 1655 1669. [CrossRef]
- Holmes, J. A.; Carlton-Smith, C.; Kim, A. Y.; Dumas, E. O.; Brown, J.; Gustafson, J.; Lauer, G. M.; Silva, S. T.; Robidoux, M.; Kvistad, D.; Alatrakchi, N.; Tonnerre, P.; Cohen, D. E.; Zhang, H.; Shulman, N. S.; Chung, R. T. Dynamic Changes in Innate Immune Responses during Direct-Acting Antiviral Therapy for HCV Infection. Journal of Viral Hepatitis 2019, 26 (3), 362–372. [CrossRef]
- Owusu Sekyere S, Suneetha PV, Hardtke S, Falk CS, Hengst J, Manns MP, Cornberg M, Wedemeyer H, Schlaphoff V. Type I Interferon Elevates Co-Regulatory Receptor Expression on CMV- and EBV-Specific CD8 T Cells in Chronic Hepatitis C. Front Immunol. 2015 Jun 10;6:270. [CrossRef]
- Hengst, J.; Falk, C. S.; Schlaphoff, V.; Deterding, K.; Manns, M. P.; Cornberg, M.; Wedemeyer, H. Direct-Acting Antiviral–Induced Hepatitis C Virus Clearance Does Not Completely Restore the Altered Cytokine and Chemokine Milieu in Patients with Chronic Hepatitis C. The Journal of Infectious Diseases 2016, 214 (12), 1965–1974. [CrossRef]
- Khera, T.; Du, Y.; Todt, D.; Deterding, K.; Strunz, B.; Hardtke, S.; Aregay, A.; Port, K.; Hardtke-Wolenski, M.; Steinmann, E.; Björkström, N. K.; Manns, M. P.; Hengst, J.; Cornberg, M.; Wedemeyer, H. Long-Lasting Imprint in the Soluble Inflammatory Milieu despite Early Treatment of Acute Symptomatic Hepatitis C. The Journal of Infectious Diseases 2021, 226 (3), 441–452. [CrossRef]
- Aregay, A.; Engel, B.; Port, K.; Vondran, F. W. R.; Bremer, B.; Niehaus, C.; Khera, T.; Richter, N.; Jaeckel, E.; Cornberg, M.; Taubert, R.; Wedemeyer, H. Distinct Immune Imprints of Post–Liver Transplantation Hepatitis C Persist despite Viral Clearance. Liver Transplantation 2021, 27 (6), 887–899. [CrossRef]
- Sekyere, S. O.; Port, K.; Deterding, K.; Cornberg, M.; Wedemeyer, H. Inflammatory Patterns in Plasma Associate with Hepatocellular Carcinoma Development in Cured Hepatitis C Cirrhotic Patients. United European Gastroenterology Journal 2021, 9 (4), 486–496. [CrossRef]
- Debes, J. D.; Van Tilborg, M.; Groothuismink, Z. M. A.; Hansen, B. E.; Wiesch, J. S. Z.; Von Felden, J.; De Knegt, R. J.; Boonstra, A. Levels of Cytokines in Serum Associate with Development of Hepatocellular Carcinoma in Patients with HCV Infection Treated with Direct-Acting Antivirals. Gastroenterology 2018, 154 (3), 515-517.e3. [CrossRef]
- Knop, V.; Hoppe, D.; Welzel, T. M.; Vermehren, J.; Herrmann, E.; Vermehren, A.; Friedrich-Rust, M.; Sarrazin, C.; Zeuzem, S.; Welker, M.-W. Regression of Fibrosis and Portal Hypertension in HCV-Associated Cirrhosis and Sustained Virologic Response after Interferon-Free Antiviral Therapy. Journal of Viral Hepatitis 2016, 23 (12), 994–1002. [CrossRef]
- Bachofner, J. A.; Valli, P. V.; Kröger, A.; Bergamin, I.; Künzler, P.; Baserga, A.; Braun, D. L.; Seifert, B.; Moncsek, A.; Fehr, J.; Semela, D.; Magenta, L.; Müllhaupt, B.; Beretta-Piccoli, B. T.; Mertens, J. C. Direct Antiviral Agent Treatment of Chronic Hepatitis C Results in Rapid Regression of Transient Elastography and Fibrosis Markers Fibrosis-4 Score and Aspartate Aminotransferase-platelet Ratio Index. Liver International 2016, 37 (3), 369–376. [CrossRef]
- Rosato, V.; Ascione, A.; Nevola, R.; Fracanzani, A. L.; Piai, G.; Messina, V.; Claar, E.; Coppola, C.; Fontanella, L.; Lombardi, R.; Staiano, L.; Valente, G.; Fascione, M. C.; Giorgione, C.; Mazzocca, A.; Galiero, R.; Perillo, P.; Marrone, A.; Sasso, F. C.; Adinolfi, L. E.; Rinaldi, L. Factors Affecting Long-term Changes of Liver Stiffness in Direct-acting Anti-hepatitis C Virus Therapy: A Multicentre Prospective Study. Journal of Viral Hepatitis 2021, 29 (1), 26–34. [CrossRef]
- Yoo, H. W.; Park, J. Y.; Kim, S. G.; Jung, Y. K.; Lee, S. H.; Kim, M. Y.; Jun, D. W.; Jang, J. Y.; Lee, J. W.; Kwon, O. S. Regression of Liver Fibrosis and Hepatocellular Carcinoma Development after HCV Eradication with Oral Antiviral Agents. Scientific Reports 2022, 12 (1). [CrossRef]
- Salomone, F.; Petta, S.; Micek, A.; Pipitone, R. M.; Distefano, A.; Castracani, C. C.; Rini, F.; Di Rosa, M.; Gardi, C.; Calvaruso, V.; Di Marco, V.; Volti, G. L.; Grimaudo, S. Hepatitis C Virus Eradication by Direct Antiviral Agents Abates Oxidative Stress in Patients with Advanced Liver Fibrosis. Liver International 2020, 40 (11), 2820–2827. [CrossRef]
- Suzuki, T.; Matsuura, K.; Nagura, Y.; Ogawa, S.; Fujiwara, K.; Nojiri, S.; Watanabe, T.; Kataoka, H.; Tanaka, Y. Serum Angiopoietin-2 Levels Predict Regression of Mac-2 Binding Protein Glycosylation Isomer-based Liver Fibrosis after Hepatitis C Virus Eradication by Direct-acting Antiviral Agents. Hepatology Research 2022, 52 (11), 919–927. [CrossRef]
- Nagura, Y.; Suzuki, T.; Matsuura, K.; Ogawa, S.; Kawamura, H.; Kuno, K.; Fujiwara, K.; Nojiri, S.; Nonomura, K.; Iio, E.; Watanabe, T.; Kataoka, H.; Tanaka, Y. Serum Inducible Protein 10 kDa/C-X-C Motif Chemokine 10 Levels Predict Regression of M2BPGi-based Liver Fibrosis after Hepatitis C Virus Eradication by Direct-acting Antiviral Agents. Hepatology Research 2023. [CrossRef]
- Ferreira, J.; Oliveira, M. J. S.; Bicho, M.; Serejo, F. Role of Inflammatory/Immune Response and Cytokine Polymorphisms in the Severity of Chronic Hepatitis C (CHC) before and after Direct Acting Antiviral (DAAs) Treatment. International Journal of Molecular Sciences 2023, 24 (2), 1380. [CrossRef]
- Amadei, B.; Urbani, S.; Cazaly, A.; Fisicaro, P.; Zerbini, A.; Ahmed, P. S.; Missale, G.; Ferrari, C.; Khakoo, S. I. Activation of Natural Killer Cells during Acute Infection with Hepatitis C Virus. Gastroenterology 2010, 138 (4), 1536–1545. [CrossRef]
- Oliviero, B.; Varchetta, S.; Paudice, E.; Michelone, G.; Zaramella, M.; Mavilio, D.; Filippi, F.; Bruno, S.; Mondelli, M. U. Natural Killer Cell Functional Dichotomy in Chronic Hepatitis B and Chronic Hepatitis C Virus Infections. Gastroenterology 2009, 137 (3), 1151-1160.e7. [CrossRef]
- Miyagi, T.; Takehara, T.; Nishio, K.; Shimizu, S.; Kohga, K.; Li, W.; Tatsumi, T.; Hiramatsu, N.; Kanto, T.; Hayashi, N. Altered Interferon-α-Signaling in Natural Killer Cells from Patients with Chronic Hepatitis C Virus Infection. Journal of Hepatology 2010, 53 (3), 424–430. [CrossRef]
- Ahlenstiel, G.; Titerence, R.; Koh, C.; Edlich, B.; Feld, J. J.; Rotman, Y.; Ghany, M. G.; Hoofnagle, J. H.; Liang, T. J.; Heller, T.; Rehermann, B. Natural Killer Cells Are Polarized toward Cytotoxicity in Chronic Hepatitis C in an Interferon-Alfa–Dependent Manner. Gastroenterology 2010, 138 (1), 325-335.e2. [CrossRef]
- Spaan, M.; Van Oord, G.; Kreefft, K.; Hou, J.; Hansen, B. E.; Janssen, H. L. A.; De Knegt, R. J.; Boonstra, A. Immunological Analysis during Interferon-Free Therapy for Chronic Hepatitis C Virus Infection Reveals Modulation of the Natural Killer Cell Compartment. The Journal of Infectious Diseases 2015, 213 (2), 216–223. [CrossRef]
- Wang, X.; Luo, B.; Jiang, H.-J.; Xu, C.; Qian, J.; Ma, D.-L.; Wei, L.; Feng, B. Recovery of Natural Killer Cells Is Mainly in Post-Treatment Period in Chronic Hepatitis C Patients Treated with Sofosbuvir plus Ledipasvir. World Journal of Gastroenterology 2018, 24 (40), 4554–4564. [CrossRef]
- Serti, E.; Chepa-Lotrea, X.; Kim, Y. J.; Keane, M.; Fryzek, N.; Liang, T. J.; Ghany, M. G.; Rehermann, B. Successful Interferon-Free Therapy of Chronic Hepatitis C Virus Infection Normalizes Natural Killer Cell Function. Gastroenterology 2015, 149 (1), 190-200.e2. [CrossRef]
- Bozzano, F.; Picciotto, A.; Costa, P.; Marras, F.; Fazio, V.; Hirsch, I.; Olive, D.; Moretta, A.; De Maria, A. Activating NK Cell Receptor Expression/Function (NKp30, NKp46, DNAM-1) during Chronic Viraemic HCV Infection Is Associated with the Outcome of Combined Treatment. European Journal of Immunology 2011, 41 (10), 2905–2914. [CrossRef]
- Jiang, H.-J.; Wang, X.; Luo, B.; Xu, C.; Qian, J.; Qin, H.; Zhang, H.; Kong, X.; Wei, L.; Feng, B. Direct Antiviral Agents Upregulate Natural Killer Cell Potential Activity in Chronic Hepatitis C Patients. Clinical and Experimental Medicine 2019, 19 (3), 299–308. [CrossRef]
- Golden–Mason, L.; McMahan, R. H.; Kriss, M.; Kilgore, A.; Cheng, L.; Dran, R. J.; Wieland, A.; Rosen, H. R. Early and Late Changes in Natural Killer Cells in Response to Ledipasvir/Sofosbuvir Treatment. Hepatology Communications 2018, 2 (4), 364–375. [CrossRef]
- Perpiñán, E.; Pérez-del-Pulgar, S.; Londoño, M.-C.; Mariñó, Z.; Bartrés, C.; González, P.; García-López, M.; Pose, E.; Lens, S.; Maini, M. K.; Forns, X.; Koutsoudakis, G. Cirrhosis Hampers Early and Rapid Normalization of Natural Killer Cell Phenotype and Function in Hepatitis C Patients Undergoing Interferon-Free Therapy. Frontiers in Immunology 2020, 11. [CrossRef]
- Wijaya, R. S.; Read, S. A.; Selvamani, S. P.; Schibeci, S. D.; Azardaryany, M. K.; Ong, A.; Van Der Poorten, D.; Lin, R.; Douglas, M. W.; George, J.; Ahlenstiel, G. Hepatitis C Virus (HCV) Eradication with Interferon-Free Direct-Acting Antiviral-Based Therapy Results in KLRG1+ HCV-Specific Memory Natural Killer Cells. The Journal of Infectious Diseases 2020, 223 (7), 1183–1195. [CrossRef]
- Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Lombardi, A.; Genco, F.; Gulminetti, R.; Novati, S.; Mondelli, M. U.; Varchetta, S. Adaptive Natural Killer Cell Functional Recovery in Hepatitis C Virus Cured Patients. Hepatology 2020, 73 (1), 79–90. [CrossRef]
- Strunz, B.; Hengst, J.; Deterding, K.; Manns, M. P.; Cornberg, M.; Ljunggren, H.; Wedemeyer, H.; Björkström, N. K. Chronic Hepatitis C Virus Infection Irreversibly Impacts Human Natural Killer Cell Repertoire Diversity. Nature Communications 2018, 9 (1). [CrossRef]
- Oliviero, B.; Mantovani, S.; Varchetta, S.; Mele, D.; Grossi, G.; Ludovisi, S.; Nuti, E.; Rossello, A.; Mondelli, M. U. Hepatitis C Virus-Induced NK Cell Activation Causes Metzincin-Mediated CD16 Cleavage and Impaired Antibody-Dependent Cytotoxicity. Journal of Hepatology 2017, 66 (6), 1130–1137. [CrossRef]
- Martin, B.; Hennecke, N.; Lohmann, V.; Kayser, A.; Neumann-Haefelin, C.; Kukolj, G.; Böcher, W. O.; Thimme, R. Restoration of HCV-Specific CD8+ T Cell Function by Interferon-Free Therapy. Journal of Hepatology 2014, 61 (3), 538–543. [CrossRef]
- Burchill, M. A.; Golden-Mason, L.; Wind-Rotolo, M.; Rosen, H. R. Memory Re-Differentiation and Reduced Lymphocyte Activation in Chronic HCV-Infected Patients Receiving Direct-Acting Antivirals. Journal of Viral Hepatitis 2015, 22 (12), 983–991. [CrossRef]
- Romani, S.; Stafford, K. A.; Nelson, A.; Bagchi, S.; Kottilil, S.; Poonia, B. Peripheral PD-1+ T Cells Co-Expressing Inhibitory Receptors Predict SVR with Ultra Short Duration DAA Therapy in HCV Infection. Frontiers in Immunoogy 2019, 10. [CrossRef]
- Han, J. W.; Sung, P. S.; Kim, K. H.; Hong, S.-H.; Shin, E.; Song, M. J.; Park, S. Dynamic Changes in Ex Vivo T-Cell Function after Viral Clearance in Chronic HCV Infection. The Journal of Infectious Diseases 2019, 220 (8), 1290–1301. [CrossRef]
- Wieland, D.; Kemming, J.; Schuch, A.; Emmerich, F.; Knolle, P. A.; Neumann-Haefelin, C.; Held, W.; Zehn, D.; Hofmann, M.; Thimme, R. TCF1+ Hepatitis C Virus-Specific CD8+ T Cells Are Maintained after Cessation of Chronic Antigen Stimulation. Nature Communications 2017, 8 (1). [CrossRef]
- Llorens-Revull, M.; Costafreda, M. I.; Rico, A.; Guerrero-Murillo, M.; Soria, M. E.; Píriz-Ruzo, S.; Vargas-Accarino, E.; Gabriel-Medina, P.; Rodríguez-Frías, F.; Riveiro-Barciela, M.; Perales, C.; Quer, J.; Sauleda, S.; Esteban, J. I.; Bes, M. Partial Restoration of Immune Response in Hepatitis C Patients after Viral Clearance by Direct-Acting Antiviral Therapy. PLOS ONE 2021, 16 (7), e0254243. [CrossRef]
- Casey, J. R.; Dore, G. J.; Grebely, J.; Matthews, G.; Cherepanov, V.; Martinello, M.; Marks, P.; Janssen, H. L. A.; Hansen, B. E.; Kaul, R.; MacParland, S. A.; Gehring, A. J.; Feld, J. J. Hepatitis C Virus-specific Immune Responses Following Direct-acting Antivirals Administered during Recent Hepatitis C Virus Infection. Journal of Viral Hepatitis 2023, 30 (1), 64–72. [CrossRef]
- Tonnerre, P.; Wolski, D.; Subudhi, S.; Aljabban, J.; Hoogeveen, R. C.; Damasio, M.; Drescher, H. K.; Bartsch, L. M.; Tully, D. C.; Sen, D. R.; Bean, D. J.; Brown, J.; Torres-Cornejo, A.; Robidoux, M.; Kvistad, D.; Alatrakchi, N.; Cui, A.; Lieb, D.; Cheney, J. A.; Gustafson, J.; Lewis-Ximenez, L. L.; Massenet-Regad, L.; Eisenhaure, T.; Aneja, J.; Haining, W. N.; Chung, R. T.; Hacohen, N.; Allen, T. M.; Kim, A. Y.; Lauer, G. M. Differentiation of Exhausted CD8+ T Cells after Termination of Chronic Antigen Stimulation Stops Short of Achieving Functional T Cell Memory. Nature Immunology 2021, 22 (8), 1030–1041. [CrossRef]
- Hensel, N.; Gu, Z.; Sagar; Wieland, D.; Jechow, K.; Kemming, J.; Llewellyn-Lacey, S.; Gostick, E.; Sogukpinar, O.; Emmerich, F.; Price, D. A.; Bengsch, B.; Böettler, T.; Neumann-Haefelin, C.; Eils, R.; Conrad, C.; Bartenschlager, R.; Grün, D.; Ishaque, N.; Thimme, R.; Hofmann, M. Memory-like HCV-Specific CD8+ T Cells Retain a Molecular Scar after Cure of Chronic HCV Infection. Nature Immunology 2021, 22 (2), 229–239. [CrossRef]
- Yates, K. B.; Tonnerre, P.; Martin, G.; Gerdemann, U.; Abosy, R. A.; Comstock, D. E.; Weiss, S. A.; Wolski, D.; Tully, D. C.; Chung, R. T.; Allen, T. M.; Kim, A. Y.; Fidler, S.; Fox, J.; Frater, J.; Lauer, G. M.; Haining, W. N.; Sen, D. R. Epigenetic Scars of CD8+ T Cell Exhaustion Persist after Cure of Chronic Infection in Humans. Nature Immunology 2021, 22 (8), 1020–1029. [CrossRef]
- Pauken, K. E.; Sammons, M. A.; Odorizzi, P. M.; Manne, S.; Godec, J.; Khan, O.; Drake, A.; Chen, Z.; Sen, D. R.; Kurachi, M.; Barnitz, R. A.; Bartman, C.; Bengsch, B.; Huang, A. C.; Schenkel, J. M.; Vahedi, G.; Haining, W. N.; Berger, S. L.; Wherry, E. J. Epigenetic Stability of Exhausted T Cells Limits Durability of Reinvigoration by PD-1 Blockade. Science 2016, 354 (6316), 1160–1165. [CrossRef]
- Miller, B. C.; Sen, D. R.; Abosy, R. A.; Bi, K.; Virkud, Y.; LaFleur, M. W.; Yates, K. B.; Lako, A.; Felt, K. D.; Naik, G. S.; Manos, M. P.; Gjini, E.; Kuchroo, J. R.; Ishizuka, J. J.; Collier, J. L.; Griffin, G. K.; Maleri, S.; Comstock, D. E.; Weiss, S. A.; Brown, F. D.; Panda, A.; Zimmer, M. D.; Manguso, R. T.; Hodi, F. S.; Rodig, S. J.; Sharpe, A. H.; Haining, W. N. Subsets of Exhausted CD8+ T Cells Differentially Mediate Tumor Control and Respond to Checkpoint Blockade. Nature Imunology 2019, 20 (3),326336. [CrossRef]
- Callendret, B.; Eccleston, H. B.; Hall, S.; Satterfield, W. C.; Capone, S.; Folgori, A.; Cortese, R.; Nicosia, A.; Walker, C. M. T-Cell Immunity and Hepatitis C Virus Reinfection after Cure of Chronic Hepatitis C with an Interferon-Free Antiviral Regimen in a Chimpanzee. Hepatology 2014, 60 (5), 1531–1540. [CrossRef]
- Horvath, S. DNA Methylation Age of Human Tissues and Cell Types. GenomeBiology.com (London. Print) 2013, 14 (10), R115. [CrossRef]
- Oltmanns, C.; Liu, Z.; Mischke, J.; Tauwaldt, J.; Mekonnen, Y. A.; Urbanek-Quaing, M.; Debarry, J.; Maasoumy, B.; Wedemeyer, H.; Kraft, A.; Xu, C.; Cornberg, M. Reverse Inflammaging: Long-Term Effects of HCV Cure on Biological Age. Journal of Hepatology 2023, 78 (1), 90–98. [CrossRef]
- Winkler, F.; Hipp, A. M.; Ramı́Rez, C.; Martin, B.; Villa, M.; Neuwirt, E.; Gorka, O.; Aerssens, J.; Johansson, S. E.; Rana, N.; Llewellyn-Lacey, S.; Price, D. A.; Panning, M.; Groß, O.; Pearce, E. L.; Hermann, C.; Schumann, K.; Hannibal, L.; Neumann-Haefelin, C.; Böettler, T.; Knolle, P.; Hofmann, M.; Wohlleber, D.; Thimme, R.; Bengsch, B. Enolase Represents a Metabolic Checkpoint Controlling the Differential Exhaustion Programmes of Hepatitis Virus-Specific CD8+T Cells. Gut 2023, 72 (10), 1971–1984. [CrossRef]
- Aregay, A.; Sekyere, S. O.; Deterding, K.; Port, K.; Dietz, J.; Berkowski, C.; Sarrazin, C.; Manns, M. P.; Cornberg, M.; Wedemeyer, H. Elimination of Hepatitis C Virus Has Limited Impact on the Functional and Mitochondrial Impairment of HCV-Specific CD8+ T Cell Responses. Journal of Hepatology 2019, 71 (5), 889–899. [CrossRef]
- Smits, M. J.; Zoldan, K.; Ishaque, N.; Gu, Z.; Jechow, K.; Wieland, D.; Conrad, C.; Eils, R.; Fauvelle, C.; Baumert, T. F.; Emmerich, F.; Bengsch, B.; Neumann-Haefelin, C.; Hofmann, M.; Thimme, R.; Böettler, T. Follicular T Helper Cells Shape the HCV-Specific CD4+ T Cell Repertoire after Virus Elimination. Journal of Clinical Investigation 2020, 130 (2), 998–1009. [CrossRef]
- Osuch, S.; Laskus, T.; Berak, H.; Perlejewski, K.; Metzner, K. J.; Paciorek, M.; Radkowski, M.; Cortés, K. C. Decrease of T-Cells Exhaustion Markers Programmed Cell Death-1 and T-Cell Immunoglobulin and Mucin Domain-Containing Protein 3 and Plasma IL-10 Levels after Successful Treatment of Chronic Hepatitis C. Scientific Reports 2020, 10 (1). [CrossRef]
- Kondili, L. A.; Monti, M.; Quaranta, M. G.; Gragnani, L.; Panetta, V.; Brancaccio, G.; Mazzaro, C.; Persico, M.; Masarone, M.; Gentile, I.; Andreone, P.; Madonia, S.; Biliotti, E.; Filomia, R.; Puoti, M.; Fracanzani, A. L.; Laccabue, D.; Ieluzzi, D.; Coppola, C.; Rumi, M. G.; Benedetti, A.; Verucchi, G.; Coco, B.; Chemello, L.; Iannone, A.; Ciancio, A.; Russo, F. P.; Barbaro, F.; Morisco, F.; Chessa, L.; Massari, M.; Blanc, P.; Zignego, A. L. A Prospective Study of Direct-acting Antiviral Effectiveness and Relapse Risk in HCV Cryoglobulinemic Vasculitis by the Italian PITER Cohort. Hepatology 2022, 76 (1), 220–232. [CrossRef]
- Gragnani, L.; Lorini, S.; Marri, S.; Vacchi, C.; Madia, F.; Monti, M.; Ferri, C.; Zignego, A. L. Predictors of Long-Term Cryoglobulinemic Vasculitis Outcomes after HCV Eradication with Direct-Acting Antivirals in the Real-Life. Autoimmunity Reviews 2022, 21 (1), 102923. [CrossRef]
- Visentini, M.; Del Padre, M.; Colantuono, S.; Yang, B.; Minafò, Y. A.; Antonini, S.; Carnovale, M.; De Santis, A.; Pulsoni, A.; De Sanctis, G. M.; Gragnani, L.; Zignego, A. L.; Fiorilli, M.; Casato, M. Long-lasting Persistence of Large B-cell Clones in Hepatitis C Virus-cured Patients with Complete Response of Mixed Cryoglobulinaemia Vasculitis. Liver International 2019, 39 (4), 628–632. [CrossRef]
- Del Padre, M.; Marrapodi, R.; Minafò, Y. A.; Mortari, E. P.; Radicchio, G.; Bocci, C.; Gragnani, L.; Camponeschi, A.; Colantuono, S.; Stefanini, L.; Basili, S.; Carsetti, R.; Fiorilli, M.; Casato, M.; Visentini, M. Dual Stimulation by Autoantigen and CpG Fosters the Proliferation of Exhausted Rheumatoid Factor-Specific CD21low B Cells in Hepatitis C Virus-Cured Mixed Cryoglobulinemia. Frontiers in Immunology 2023, 14. [CrossRef]
- Chang, L.-Y.; Li, Y.; Kaplan, D. E. Hepatitis C Viraemia Reversibly Maintains Subset of Antigen-Specific T-Bet+ Tissue-like Memory B Cells. Journal of Viral Hepatitis 2016, 24 (5), 389–396. [CrossRef]
- Nishio, A.; Hasan, S.; Park, H.; Park, N.; Salas, J.; Salinas, E.; Kardava, L.; Juneau, P.; Frumento, N.; Massaccesi, G.; Moir, S.; Bailey, J. R.; Grakoui, A.; Ghany, M. G.; Rehermann, B. Serum Neutralization Activity Declines but Memory B Cells Persist after Cure of Chronic Hepatitis C. Nature Communications 2022, 13 (1). [CrossRef]
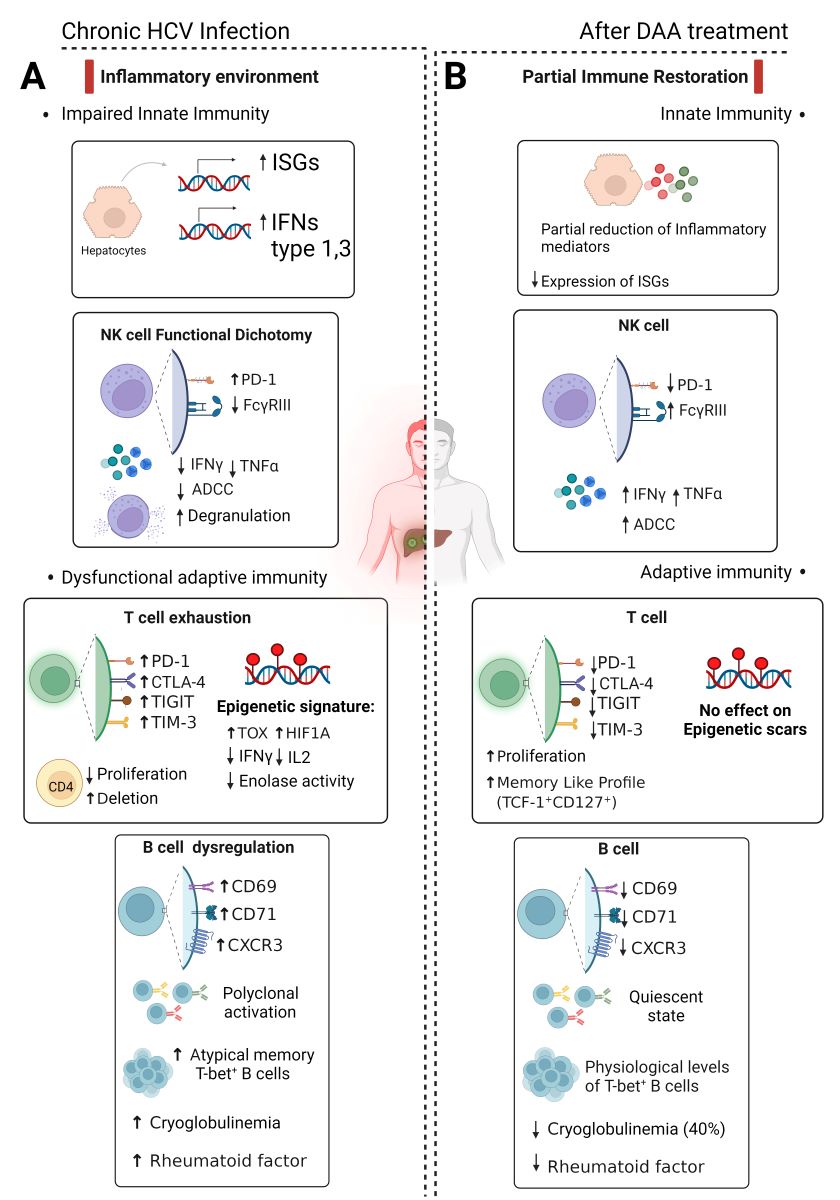
Figure 1.
Schematic overview depicting the impact of chronic HCV infection and Direct-Acting Antivirals (DAA) on the immune response. A. During chronic infection, there is heightened production of type I and III interferon and IFN-stimulated genes (ISGs). NK cells show a functional impairment marked by increased inhibitory molecule PD-1, reduced FcγRIII, decreased ADCC activity, IFN-γ, and TNF-α secretion, despite increased degranulation. T cell exhaustion occurs during chronic infection with increased expression of inhibitory molecules (PD-1, CTLA-4, TIGIT, TIM-3) and epigenetic imprints. B cells exhibit chronic activation, polyclonal expansion, and elevated markers of atypical memory cells. Cryoglobulinemia and Rheumatoid factors may be present. B. DAA treatment reduces ISG expression, while maintaining a notable presence of soluble inflammatory mediators. NK cell status reverts to physiological homeostasis after viral eradication. However, complete functional restoration remains uncertain, leaving residual defects such as suboptimal antibody-dependent cell-mediated cytotoxicity (ADCC). T-cell recovery post-DAA cure is also only partial, leaving long-lasting epigenetic scars. B cell phenotype and physiological levels of T-bet+ B cells are restored, yet oligoclonality of B-cells may persist even after HCV eradication, potentially in a quiescent state. DAA treatment for cryoglobulinemia results remission in only about half of the patients.
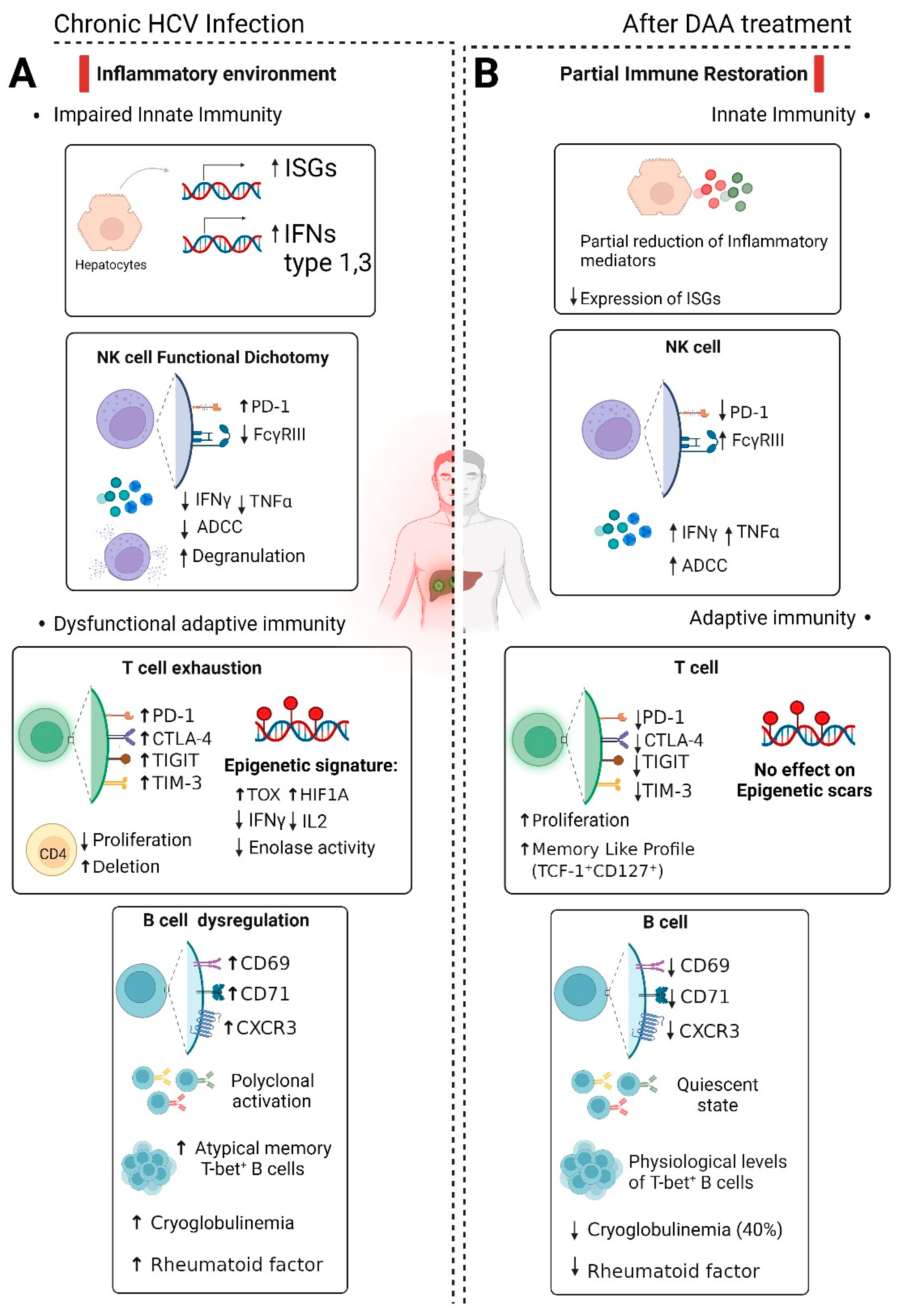
Figure 1.
Schematic overview depicting the impact of chronic HCV infection and Direct-Acting Antivirals (DAA) on the immune response. A. During chronic infection, there is heightened production of type I and III interferon and IFN-stimulated genes (ISGs). NK cells show a functional impairment marked by increased inhibitory molecule PD-1, reduced FcγRIII, decreased ADCC activity, IFN-γ, and TNF-α secretion, despite increased degranulation. T cell exhaustion occurs during chronic infection with increased expression of inhibitory molecules (PD-1, CTLA-4, TIGIT, TIM-3) and epigenetic imprints. B cells exhibit chronic activation, polyclonal expansion, and elevated markers of atypical memory cells. Cryoglobulinemia and Rheumatoid factors may be present. B. DAA treatment reduces ISG expression, while maintaining a notable presence of soluble inflammatory mediators. NK cell status reverts to physiological homeostasis after viral eradication. However, complete functional restoration remains uncertain, leaving residual defects such as suboptimal antibody-dependent cell-mediated cytotoxicity (ADCC). T-cell recovery post-DAA cure is also only partial, leaving long-lasting epigenetic scars. B cell phenotype and physiological levels of T-bet+ B cells are restored, yet oligoclonality of B-cells may persist even after HCV eradication, potentially in a quiescent state. DAA treatment for cryoglobulinemia results remission in only about half of the patients.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



