
Submitted:
28 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
The gut-liver axis plays a pivotal role in the development of liver diseases, necessitating exploration into substances capable of modulating the microbiota to prevent dysbiosis. Over the past decade, diverse therapeutic approaches have emerged to target pathogenic factors involved in the hepatic gut microbiota axis. This study investigated the impact of a supplement with hepatoprotective activity, containing extracts of Silybum marianum, prebiotics, probiotics, n-3 polyunsaturated fatty acids, minerals, and vitamins on hematological markers of liver functions and on the intestinal microbiota of 10 adult dogs with metabolic liver disease over a 35-day time span. Animals underwent clinical and laboratory evaluations every 7 days, before administration of the supplement (T0) and after 7, 14, 21, 28 and 35 days (T1, T2, T3, T4 and T5). In comparison to T0, a significant (P<0.05) decrease of AST activity was measured at T2, T3, T4 and T5. The activity of ALP, glucose and CRP significantly decreased (P<0.05) at T3, T4, and T5. The alpha diversity of the fecal microbiota significantly decreased (P<0.05) only at T1, a high variability was observed between dogs. The total short-chain fatty acid and lactic acid were also lower at T1 (P<0.05) in comparison to the other times of sampling. The beta diversity of fecal microbiota failed to find a clear pattern in relation to the sampling times. These results underscore the usefulness of the supplement on liver function and highlight high individual variability in its interaction with the fecal microbiome.
Keywords:
Gut-Liver Axis
; Supplement
; Dogs
1. Introduction
The liver plays a vital role in regulating the body’s internal antioxidant levels, primarily through the synthesis of glutathione, a significant intracellular antioxidant. An essential factor in the development of hepatitis, both acute and chronic, is the disrupted oxidative balance in the liver.
Oxidative stress arises when the production of reactive oxygen species (ROS) exceeds the body’s capacity to counteract them with antioxidant defense mechanisms. To manage liver diseases effectively, antioxidant supplementation with vitamins E and C is often recommended. These vitamins aid in building a robust antioxidant network, replenishing glutathione levels, and resorting the activity of glutathione peroxidase in the liver [1,2,3].
In the past decade, the hepatoprotective activity has been observed also for several nutraceuticals, including Silybum marianum [4], also in canine medicine [5,6,7]. These findings underline the importance of a well-rounded diet in achieving a favorable oxidative balance in dogs. Furthermore, Di Cerbo et al. [8] and Sechi et al. [9] findings strongly indicate that dietary intervention reduced d-ROMs and regulated neuroendocrine parameters, along with a general improvement in behavioral disruptions in dogs.
Nutritionists have become increasingly aware of the importance of prevention in maintaining the health of companion animals. This heightened awareness has led to a surge in the popularity of dietary supplements for pets [10], typically consisting of one or more ingredients intended to complement their diet. The supplements serve the purpose of preventing nutritional deficiencies and often contain essential components like vitamins, minerals, amino acids, and essential fatty acids. They can also provide other functional compounds such as herbs, nutraceutical plants, enzymes, and metabolites. In addition to their preventive role in disease development, nutritional and functional supplements also fall under the realm of complementary or alternative medicine (CAM). These supplements provide an alternative or adjunct approach to drug therapies, offering a means to minimize side effects [11].
In recent years, there has been a significant focus on understanding the role of gut microbiota in animal health [12,13], particularly the intricate relationship between the intestine and liver. This connection, known as the intestine-liver axis [14], is maintained through a complex interplay of metabolic, immune, neuroendocrine, and microbial interactions, ensuring the regulation and stability of the gut microbiota-liver relationship via the portal circulation [15]. The tight junctions of the intestinal epithelium act as a natural barrier against bacteria and their metabolic by-products. When antigens, whether from pathogenic microorganisms or food, cross these junctions, they trigger an immune response that alters the behavior of T lymphocytes (T-regulatory cells and T-helper 17 cells), resulting in the release of inflammatory cytokines and chemokines that ultimately enter the portal circulation [16].
Given the influence of the gut-liver axis on the onset and development of metabolic liver diseases, it becomes crucial to explore substances that can positively modulate the microbiota, preventing dysbiosis and inflammatory processes at the intestinal mucosa level.
Probiotics, prebiotics, and synbiotics have gained significant recognition as potential treatment options for liver disease [17]. The concept of “synbiotics” refers to the combination of prebiotics and probiotics, acting synergistically by providing the former with substrates for the latter to better survive within the gut, thus promoting the yield of short chain fatty acids (SCFAs). These SCFAs can serve as energy substances for intestinal cells, mimicking the effects of probiotics. In this way, prebiotics contribute to promoting a healthy intestinal environment and fostering a symbiotic relationship between the host and the beneficial microorganisms [18,19]. In particular, prebiotics are substances that stimulate the growth or activity of microorganisms, functioning as undigested foods that aid in intestinal peristalsis and selectively promote the growth of beneficial intestinal bacteria. Certain prebiotics, such as fructo-oligosaccharides (FOS), have demonstrated potential in preventing liver damage in human by resorting levels of beneficial bacteria, making them a promising therapeutic option [20,21]. Considering the positive activity of several natural compounds on liver function and gut health, in the present study, we wanted to investigate the safety and efficacy of a supplement made with a mix of nutraceuticals in promoting liver function in dogs affected by chronic metabolic hepatitis. We evaluated changes in nutritional and clinical parameters and assessed the impact on the composition of the gut microbiome.
2. Materials and Methods
2.1. Animals and Study Design
A total of 10 adult dogs were carefully selected from a veterinary clinic in Italy (Table 1). All dogs were diagnosed with chronic metabolic hepatitis following clinical and laboratory examinations conducted by the same veterinarian. The study was approved by the Ethical Committee of the University of Turin (Protocol N° 1234 of 14/4/2022). At the time of enrolment, all the animals presented pathological conditions referring to chronic metabolic hepatitis, ensuring consistency and comparability among the subjects participating in the study. After obtaining informed consent from the pet owners, the dogs remained domiciled at their usual site for the entire duration of the study, ensuring minimal disruption to their daily routines.
To establish the daily portion of the maintenance diet, the estimated requirements were computed following NRC guidelines [22] using the value of 110 kcal of metabolizable energy per kg metabolic live weight. At the commencement of the trial (T0) and subsequently every week of treatment, comprehensive nutritional assessments and physical examinations were conducted by the clinicians on the subjects to evaluate their overall health and monitor any changes. Body weight (kg) and body condition score (BCS) were recorded at the beginning of the study. BCS assessments was conducted by a trained veterinarian who assigned values ranging from 1 to 9 [23], with a BCS score of 4 or 5 considered the ideal value according to FEDIAF guidelines [24].
Throughout a period of 42 days, dogs were closely monitored and observed, with regular check-ups conducted at the veterinary facility. After an initial 7 days adaptation period, the subjects were introduced to a combination of their regular maintenance diet and a supplementary feed, consisting of 1 tablet (2gr) per 15 kg of body weight given every 24 hours, which continued to be provided throughout the study duration.
The supplement contained a combination of natural ingredients as reported in Table 2. The formulation of supplements was based on the published data by Marchegiani et al., [25].
At weekly intervals, the attending veterinarian conducted thorough examinations of the subjects to monitor any potential changes in the animal health. Faecal and blood samples were collected at the beginning of the study (T0) before the administration of the supplement, and then after 7, 14, 21, 28, 35 days (T1, T2, T3, T4 and T5, respectively).
2.2. Laboratory Analysis
2.2.1. Blood Analysis
Blood tests on serum samples were conducted to assess specific parameters linked to the liver functions and metabolism, which included aspartate transaminase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), gamma glutamyl transaminase (GGT), glucose, triglycerides, total bilirubin and C-reactive protein (CRP). For the biochemical parameters we used the Catalyst I (Ideex) and for the CRP parameter we utilized the rapid test TestICT (diagnosticavet).
2.2.2. Microbiome analysis
Faecal samples were collected by the owner on the day of the enrolment (T0) and before each of the other scheduled weekly appointments. The owners were informed on how to collect stools and how to store before the appointment (-20°C). At the end of the 35 days of the trial, all faecal samples were delivered to the veterinary clinic and immediately sent to the laboratory for analysis.
To quantify the content of SCFAs, namely acetate, propionate, butyrate, isobutyrate, and isovalerate, and lactic acid in faecal samples, a High-Performance Liquid Chromatography (HPLC) equipment was employed (Shimadzu Corporation, Kyoto, Japan). Faecal samples (1 g) were diluted 1:20 with H2SO4 0.1 N (Carlo Erba, Milan, Italy) and briefly vortexed, then centrifuged at 4000 g for 15 min on a Centrifuge (5804R, Eppendorf AG, Germany). Supernatants were filtered through syringe filters (RC 0.45 μm, 25 mm, DTO Servizi Srl, Venice, Italy) and the filtrate was transferred into an autosampler vial. Twenty microliters (20 μL) were injected into HPLC Shimadzu LC-20AT, equipped with a Prominence SPD-M20A photodiode-array detector. For separation, an Aminex HPX-87H column (300 mm x 7.8 mm) and a pre-column (Bio-Rad, Hercules, CA, USA) were used. Peaks of analytes were identified by comparing retention times of standard mixtures to those of the samples, and quantification was based on peak area measurements by external standard method. Standards of acetic, propionic, butyric, isobutyrric, isovaleric, valeric and lactic acid were obtained from Merck (Darmstadt, Germany).
Total DNA was extracted from 150 mg of faeces using a faecal DNA miniprep kit (Zymo Research; Irvine, CA, US), following the manufacter’s instructions. After the extraction, quantification and quality assessment were performed using a QubitTM 3 Fluorometer (Thermo Scientific; Waltham, MA, USA). Libraries were prepared by amplifying the V3 and V4 regions of the 16S rRNA gene, incorporating Indexes for sequencing. This was accomplished using a Nextera DNA Library Prep kit (Illumina; San Diego, CA, USA), following the manufacturer’s instructions and utilizing specific primers [26]. The resulting amplicons were sequenced on a MiSeq platform (Illumina; San Diego, CA, USA) in 2×250 paired-end mode, adhering to standard procedures and generating a sequencing depth of 50,000 reads.
The effectiveness of the entire pipeline, from DNA extraction to taxonomic annotation, was assessed using a ZymoBIOMICSTM Microbial Community Standard (Zymo Research, Irvine, CA, USA). According to the manufacturer’s specifications, the mock community included eight bacterial species with their corresponding percentages: Pseudomonas aeruginosa (4.2%), Escherichia coli (10.1%), Salmonella enterica (10.4%), Lactobacillus fermentum (18.4%), Enterococcus faecalis (9.9%), Staphylococcus aureus (15.5%), Listeria monocytogenes (14.1%), and Bacillus subtilis (17.4%).
2.3. Bioinformatic and Statistical Analysis
The initial raw sequences (FASTQ) from the 10 DNA isolation samples were processed using the bioinformatic tool Quantitative Insights Into Microbial Ecology 2 (QIIME 2) [27]. After demultiplexing, the sequenced reads meeting the quality threshold (Phred score ≥30) were identified, and chimeras were filtered out. The remaining high-quality sequences were then clustered into amplicon sequence variants (ASV) against the Greengenes database [28] for 16S rRNA. Alpha diversity between dogs and times of treatment was reported as Shannon rarefaction curve, and comparison of evenness was also computed. To evaluate beta diversity, the phylogeny was constructed based on the weighted UniFrac distance metric [29], and the results were visualized using Principal Coordinate Analysis (PCoA) plots. The differences in community composition were assessed by performing a permutational multivariate analysis of variance (PERMANOVA) using weighted UniFrac distances.
The normalized relative abundance profiles (RAs) of taxa were obtained for annotated sequences for each sample. Data were uploaded into the MicrobiomeAnalyst suite [30] for further statistical evaluation, including LefSe, single analysis, and multifactorial analysis, with times of sampling as the main factors. In the multifactorial analysis, the dog was considered as covariate to determine its impact on community composition. The raw sequence data were subsequently uploaded to the NCBI Sequence Read Archive.
3. Results
The mean values of enzymatic activity (AST, ALT, ALP, GGT) and concentrations of glucose, triglycerides, total bilirubin, and CRP in the serum sampled from the dogs before and after the administration of a supplement over a time span of 35 days are reported in Table 3.
The activity of AST at the start of the study, before supplement administration (T0), was 49.5 UI/l and decreased to 31.0 UI/l after 14 days (T2; P<0.05). The ALT activity was abnormal at T0 (107.2) and decreased from T0 to T5, with the lowest values at T1, but differences were not significant. The mean ALP decreased from the initial values (T0) at T3, T4 and T5. The mean GGT activity did not vary between sampling times, although a slight decrease was observed. At T0 and T1, the mean glucose concentrations were significantly (P<0.05) higher than at T3, T4 and T5, but all the values were within the reference range. Triglycerides showed a similar trend, with constant decrease from T0 to T5, but due to wide individual variability, the changes in concentration were not significantly different. At time point T0, the total bilirubin concentration was 0.6 mg/100ml, which was above the normal reference range. At T1 and T2, the total bilirubin concentration was reduced to 0.4 mg/100ml and to 0.3 mg/100ml, respectively, falling within the normal reference range for T3, T4 and T5. The initial CRP concentration (T0) was 1.2 mg/l, exhibiting a significant decrease (P<0.05) until the sampling time T3 (0.6 mg/l), and it maintained stability throughout the remaining duration of the study.
The evaluation of the effect of supplementation on the gut microbiome was considered in terms of the composition of the bacterial population and the main products of fermentation, namely SCFAs and lactic acid.
Figure 1A contains data representing the values of Shannon index of alpha diversity in the intestinal microbiota of the 10 adult dogs over the course of a complementary diet intervention at different time points (T0, T1, T2, T3, T4, and T5). There was not a large variation in the index for each time of sampling, and the p-value of the ANOVA test indicates that there were no statistically significant differences in these values, suggesting limited dynamic changes in the taxa abundances during the dietary intervention. The data provided in Figure 1B represents the alpha diversity values for each dog during the study and highlight the wide variations between the subjects (p < 0.05). Some dogs exhibit alpha diversity values higher than 3 (D8, D1, D10), while others have values around 2 (D6, D2). Conversely, the rarefaction curves clearly indicated that the sequence depth was appropriate to describe the diversity of the microbiome in the faecal samples.
The evenness, calculated starting from the alpha biodiversity, was significantly lower (P<0.05) at T1 compared to T0. For the samples collected after T1 (Figure 2A), the evenness was not significantly different from T0. However, the variability between dogs was also significant (Figure 2A), with D6 having the lowest and D8 the highest values. Of note, the within dog variability of evenness was very reduced for 4 dogs (D5, D3, D8, D10) in comparison to the other subjects, confirming the large individual variability.
The beta diversity was assessed using weighted UniFrac distances (Figure 3A) for adult dogs over 35 days (T0 to T5). The graphical appraisal showed that the samples from T0 to T5 did not cluster and were almost scattered, underlining limited variations during the time course of the study. The results of beta diversity distances for different dogs showed two distinct clusters of samples representing dogs D7 and D4, while the samples from other dogs were more scattered and tended to overlap (Figure 3B).
As a final evaluation of the variation of the faecal microbiome in relation to the supplement administration, the comparison of RAs between times of sampling was computed and features significantly different are reported in Figure 4.
Ruminococcus gnavus showed a large fluctuation over the specified time points. Notably, the highest normalized count of Ruminococcus gnavus was at T3 and T5 (58.4 and 58.6, respectively) and these values were higher than those measured at T0 and T1 (25.6 and 23.7, respectively; P<0.05). Apart for the decrease from T3 to T4, we can consider that this specie increased after the administration of the supplement. The variations in normalized counts of Blautia in adult dogs with liver diseases after the administration of the supplement dramatically decreased (P<0.05) from T0 to the other time points of sampling. At (T0) the highest normalized counts of Blautia accounted for 45.5, indicating a high presence of this microbial species at the beginning of the study, and the lowest was 11.8 at T3. The RAs of Streptococcus were very low from T0 to T3 and sharply increased at T4 and T5, reaching values of 9.0 and 5.7 (P<0.05).
In relation to the initial RA at T0, Turicibacter showed a fluctuation, decreasing until T3 (P<0.05), followed by an increase to basal value at T5 and the mean values ranged from 7.1 to 25.1. The RA of the family of Peptostreptococcaceae remained relatively stable at T0 and T1, notably increased at T2 (33.5), and had the lowest values at T3 (15.5) and T4 (17.1). These differences were statistically significant (P<0.05).
Table 4 reports the concentrations of total SCFA and lactic acid in mmol/l at the different time points (T0 to T5) in 10 adult dogs. The concentrations of SCFA and lactic acid varied significantly across the observed time points. T0 (0-day) indicated a total concentration of 213.9 mmol/l, and at T1(7-day), the total concentration was the lowest and decreased to 173.5 mmol/l, significantly different (P<0.05) from T3 (249.7 mmol/l).
Lactic acid concentrations decreased at T1 (7 days) in comparison to T0 (before supplementation) and then increased significantly (P<0.05) at T3 (21 days). There were not significant differences between T0, T2, T4, and T5. During supplement administration, lactic acid levels displayed dynamic fluctuations, initially declining and subsequently increasing, hinting at possible shifts in metabolic processes. Meanwhile, the levels of acetic acid, propionic acid, isobutyric acid, butyric acid, and isovaleric acid remained relatively constant throughout the study, showing no significant changes.
4. Discussion
The study involved 10 adult dogs subjected to a complementary diet intervention over the course of 35 days, with a focus on assessing the diet’s impact on liver function, overall well-being, and various nutritional and clinical parameters. Additionally, the study aimed to evaluate changes in the composition of the intestinal microbiota.
According to the veterinary diagnosis and blood laboratory data (Table 2), dog suffered from hepatocellular damage, as indicated by the enzyme activities of ALT. The dogs were also affected by metabolic chronic liver disease, as supported by the high blood concentrations of triglycerides and glycemia (Table 3) associated to overweight (Table 1). Even though a biopsy was not collected, it is likely that a concomitant hepatic steatosis was also present, excluding an acute form of liver disease related to infectious and toxicity. In summary the initial values of ALT, ALP, GGT and bilirubin are indicative of liver pathology.
As reported by Gommeren et al. [31] and Torrente et al. [32], a high concentration of CRP is associated to a systemic inflammatory condition and can be used to monitor the progress of recovery during treatments. From a metabolic point of view, the linear decrease in glycemia and triglycerides suggests a recovery from a condition likely associated to fatty liver, even though the latter was not significant from T0 to T5.
After the supplementation with the mixture of nutraceuticals, vitamin, minerals, and n-3 PUFA (EPA and DHA) a significant improvement for AST, ALP and CRP was observed (Table 3). The results suggest a positive effect of the supplement on the liver conditions observed in the 10 dogs.
Silybum marianum is a biennial plant belonging to the Asteraceae family. The predominant active constituent of the extract is silymarin, a phytocomplex composed of various flavonoids, with silybin, isosilybin, silychristin, and silydianin being the most significant ones, primarily concentrated in its fruits and seeds [33]. Silymarin possesses multiple beneficial effects beneficial for hepatobiliary disease treatment, encompassing antioxidant, anti-inflammatory, and antifibrotic properties [34,35]. The positive effect of the supplementation with Silybum marianum for the support of the treatment of liver diseases was also reported in dogs [36,37,38].
Other compounds with anti-inflammatory activity are the EPA and DHA, that are involved in the biosynthesis of resolvins, a family of molecules that mediated the release of proinflammatory cytokines [39]. More specifically, EPA stimulates the biosynthesis of resolvins, and DHA the biosynthesis of resolvins, maresins and protectins. In humans, dietary administration of n-3 PUFA is beneficial for the treatment of non-alcoholic fatty liver disease [40,41]. In dogs, there is not proven evidence for the efficacy of EPA and DHA on liver disease, but the anti-inflammatory activity is reported for other conditions, such as atopic dermatitis [42] and atrial fibrillation [43].
If the blood parameters indicted a recovery of the dogs from the liver inflammatory and metabolic chronic disease, the faecal microbiome showed only a transient decrease in alpha diversity after 7 days (T0) of supplement administration (Figure 2, P<0.05).
The administration of Bacillus subtilis to weaned puppies, together with other ingredients, caused only minor modification of RA of faecal microbiota [44]. The specie is considered positive for gut health of dogs and affecting the production of SCFAs [45,46] but in the present study the changes of microbiome was not in line with the previous studies. The supplement fed to the dogs in the present study also contained FOS and beta glucans from barley, which are prebiotics known to modify faecal microbiota and end products of fermentation in dogs [47,48,49]. In another study, FOS were administered either independently or in conjunction with mannan-oligosaccharide to dogs whose diet primarily consisted of meat, exhibiting elevated ileal immunoglobulin A concentrations but demonstrating reduced concentrations of faecal total indole and phenol [50]. Flickinger et al. [51] reported that in adult male beagles, an oligofructose-enriched diet decreased faecal ammonia and Clostridium perfringens concentrations, while total aerobes increased, thus ameliorating the overall dog health.
Nutraceuticals from plant extracts can also modify gut microbiota, and polyphenols have gained the attention of researchers in dogs [52,53,54]. However, to the best of our knowledge, no results are available in the literature concerning the effect of Silybum marianum on faecal microbiota of dogs, although silybin have been reported to positively affect the microbial population of the gut. Interestingly, studies conducted by Wang et al. [55] indicated a beneficial effect of silybin on the gut microbiota of mice.
The slight variation observed for the faecal microbiome of dogs was probably due to the different effects of each compound contained in the supplement on microbial populations or to confounding factors such as breed and sex. This can be deducted from the large individual variability of faecal microbiome at the beginning of the study and later after feeding the supplement (Figure 2 and Figure 3).
In our study, the fluctuation of the relative abundance of Turicibacter, the linear decrease of Blautia with the time of sampling, and the higher relative abundance of Streptococcus at T4 and T5 (Figure 4) would suggest that the supplement decreased the health condition of the gut. In the dysbiosis index proposed by AlShawaqfeh et al. [56], Blautia and Turicibacter contribute positively to the gut health, while Streptococcus has a negative impact. Conversely, the relative abundance of Ruminococcus gnavus, was higher at the end of the study (T5) in comparison to T0 and T1. This species, or at least some strains, is thought to have beneficial roles in the gut of humans [57] and possibly in dogs [2].
Blautia, Turicibacter and Ruminococcus gnavus are considered butyrate producing bacteria [13,58,59,60] and their variation would impact on SCFA faecal content. However, the results reported in Table 4 did not show any variation in individual and total SCFA, apart from T1. It is likely that in the complex microbiota ecosystem of the gut, the ecological niche of these bacteria was occupied by other taxa.
5. Conclusions
Our study and results highlight the individualized nature of the diet-microbiota relationship in adult dogs. The dietary supplement had positive effects on blood parameters, ameliorating the liver conditions. The dogs displayed considerable inter-individual variation in microbiota composition, and overall, the supplement’s impact on the gut microbiota was complex and not uniform. These findings underscore the importance of considering individual variability when assessing the effects of dietary interventions on canine health and emphasize the need for further research to understand the mechanisms behind these observed changes.
Acknowledgments
We would like to thank the Clinica Veterinaria Mattalia (Margarita, CN, Italy) for the support during the trial.
References
- Margaritelis, N.V.; Veskoukis, A.S.; Paschalis, V.; Vrabas, I.S.; Dipla, K.; Zafeiridis, A.; Kyparos, A.; Nikolaidis, M.G. Blood reflects tissue oxidative stress: a systematic review. Biomarkers. 2015, 20, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lauber, C.L.; Czarnecki-Maulden, G.; Pan, Y.; Hannah, S.S. Effects of the Dietary Protein and Carbohydrate Ratio on Gut Microbiomes in Dogs of Different Body Conditions. mBio 2017, 24, 8–e01703. [Google Scholar] [CrossRef] [PubMed]
- Barry-Heffernan, C.; Ekena, J.; Dowling, S.; Pinkerton, M.E.; Viviano, K. Biomarkers of oxidative stress as an assessment of the redox status of the liver in dogs. J Vet Intern Med. 2019, 33, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Shirzad, H.; Rafieian, S.; Rafieian-Kopaei, M. Silybum marianum: Beyond Hepatoprotection. J Evid Based Complementary Altern Med. 2015, 20, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Martello, E.; Perondi, F.; Bisanzio, D.; Lippi, I.; Meineri, G.; Gabriele, V. Antioxidant Effect of a Dietary Supplement Containing Fermentative S-Acetyl-Glutathione and Silybin in Dogs with Liver Disease. Vet Sci. 2023, 8, 10–131. [Google Scholar] [CrossRef] [PubMed]
- Perondi, F.; Bisanzio, D.; Adami, R.; Lippi, I.; Meineri, G.; Cutrignelli, M.I.; Massa, S.; Martello, E. The effect of a diet supplement containing S-acetyl-glutathione (SAG) and other antioxidant natural ingredients on glutathione peroxidase in healthy dogs: a pilot study. Italian Journal of Animal Science 2023, 22, 589–593. [Google Scholar] [CrossRef]
- Marchegiani, A.; Fruganti, A.; Spaterna, A.; Dalle Vedove, E.; Bachetti, B.; Massimini, M.; Di Pierro, F.; Gavazza, A.; Cerquetella, M. Impact of Nutritional Supplementation on Canine Dermatological Disorders. Vet Sci. 2020, 7, 38. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Sechi, S.; Canello, S.; Guidetti, G.; Fiore, F.; Cocco, R. Behavioral disturbances: an innovative approach to monitor the modulatory effects of a nutraceutical diet. J. Vis. Exp. 2017, 119. [Google Scholar]
- Sechi, S.; Di Cerbo, A.; Canello, S.; Guidetti, G.; Chiavolelli, F.; Fiore, F.; Cocco, R. Effects in dogs with behavioural disorders of a commercial nutraceutical diet on stress and neuroendocrine parameters. Vet. Rec. 2017, 180, 18. [Google Scholar] [CrossRef]
- Krawczyk, M.; Grąt, M.; Barski, K.; Ligocka, J.; Antczak, A.; Kornasiewicz, O.; Skalski, M.; Patkowski, W.; Nyckowski, P. , et al. 1000 liver transplantations at the Department of General, Transplant and Liver Surgery, Medical University of Warsaw--analysis of indications and results. Pol Przegl Chir. 2012, 84, 304–12. [Google Scholar] [CrossRef]
- De Almeida Andrade, F.; Schlechta Portella, C.F. Research methods in complementary and alternative medicine: an integrative review. J Integr Med. 2018, 16, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front Vet Sci. 2020, 14, 6–498. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria, and the gut-heart axis in atherosclerosis. Clinica chimica acta; international journal of clinical chemistry. 2020, 507, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Shalaby, N.; Samocha-Bonet, D.; Kaakoush, N. O.; Danta, M. The Role of the Gastrointestinal Microbiome in Liver Disease. Pathogens (Basel, Switzerland) 2023, 12, 1087. [Google Scholar] [CrossRef]
- Chassaing, B.; Etienne-Mesmin, L.; Gewirtz, A.T. Microbiota-liver axis in hepatic disease. Hepatology. 2014, 59, 328–339. [Google Scholar] [CrossRef]
- Konturek, P.C.; Harsch, I.A.; Konturek, K.; Schink, M.; Konturek, T.; Neurath, M.F.; Zopf, Y. Gut–Liver Axis: How Do Gut Bacteria Influence the Liver? Med. Sci. 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Minemura, M.; Shimizu, Y. Gut microbiota and liver diseases. World J Gastroenterol. 2015, 14, 1691–702. [Google Scholar] [CrossRef] [PubMed]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef]
- Desdouets, C. Oxidative stress promotes pathologic polyploidization in non-alcoholic fatty liver disease. J. Clin. Investig. 2015, 125, 981–992. [Google Scholar]
- Wan, M.L.Y.; El-Nezami, H. Targeting gut microbiota in hepatocellular carcinoma: Probiotics as a novel therapy. Hepatobiliary Surg. Nutr. 2018, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- National Research Council; Division on Earth and Life Studies; Board on Agriculture and Natural Resources; Subcommittee on Dog and Cat Nutrition; Committee on Animal Nutrition. https://nap.nationalacademies.org/catalog/10668.
- Laflamme, D.P. Development and validation of a body condition score system for dogs. Canine Pract. 1997, 22, 10–15. [Google Scholar]
- FEDIAF Nutritional Guidelines for Complete and Complementary Pet Food, (europeanpetfood.org).
- Marchegiani, A.; Fruganti, A.; Gavazza, A.; Mangiaterra, S.; Candellone, A.; Fusi, E.; Rossi, G.; Cerquetella, M. Evidences on Molecules Most Frequently Included in Canine and Feline Complementary Feed to Support Liver Function. Vet Med Int. 2020, 9, 9185759. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic. Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable, and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Help with greengenes gg_13_8 reference otus - Other Bioinformatics Tools - QIIME 2 Forum.
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knigh, R. Quantitative and qualitative diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- (https://www.microbiomeanalyst.ca/).
- Gommeren, K.; Desmas, I.; Garcia, A.; Bauer, N.; Moritz, A.; Roth, J.; Peeters, D. Inflammatory cytokine and C-reactive protein concentrations in dogs with systemic inflammatory response syndrome. Journal of veterinary emergency and critical care (San Antonio, Tex.: 2001). 2018, 28, 9–19. [Google Scholar] [CrossRef]
- Torrente, C.; Manzanilla, E. G.; Bosch, L.; Fresno, L.; Rivera Del Alamo, M.; Andaluz, A.; Saco, Y.; Ruiz de Gopegui, R. Plasma iron, C-reactive protein, albumin, and plasma fibrinogen concentrations in dogs with systemic inflammatory response syndrome. Journal of veterinary emergency and critical care (San Antonio, Tex.: 2001). 2015, 25, 611–619. [Google Scholar] [CrossRef]
- Pradhan, S.C.; Girish, C. Hepatoprotective herbal drug, silymarin from experimental pharmacology to clinical medicine. Indian J Med Res. 2006, 124, 491–504. [Google Scholar]
- Trappoliere, M.; Caligiuri, A.; Schmid, M.; Bertolani, C.; Failli, P.; Vizzutti, F.; Novo, E.; di Manzano, C.; Marra, F.; Loguercio, C.; Pinzani, M. Silybin, a component of sylimarin, exerts anti-inflammatory and anti-fibrogenic effects on human hepatic stellate cells. J Hepatol. 2009, 50, 1102–11. [Google Scholar] [CrossRef]
- Verma, S.; Thuluvath, P.J. Complementary and alternative medicine in hepatology: review of the evidence of efficacy. Clin Gastroenterol Hepatol. 2007, 5, 408–16. [Google Scholar] [CrossRef] [PubMed]
- Magdalan, J.; Ostrowska, A.; Piotrowska, A.; Izykowska, I.; Nowak, M.; Szelag, A.; Dziegiel, P. Failure of benzylpenicillin, N-acetylcysteine and silibinin to reduce alpha-amanitin hepatotoxicity. In Vivo 2009, 23, 393–9. [Google Scholar] [PubMed]
- Sgorlon, S.; Stefanon, B.; Sandri, M.; Colitti, M. Nutrigenomic activity of plant derived compounds in health and disease: Results of a dietary intervention study in dog. Res. Vet. Sci. 2016, 109, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Vandeweerd, J.M.; Cambier, C.; Gustin, P. Nutraceuticals for canine liver disease: assessing the evidence. Vet Clin North Am Small Anim Pract. 2013, 43, 1171–9. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C. N.; Dalli, J.; Colas, R. A.; Winkler, J. W.; Chiang, N. Protectins and maresins: New pro-resolving families of mediators in acute inflammation and resolution bioactive metabolome. Biochimica et biophysica acta. 2015, 1851, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids and non-alcoholic fatty liver disease: Evidence of efficacy and mechanism of action. Mol Aspects Med. 2018, 64, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B.; Lytle, K.A.; Depner, C.M.; Tripathy, S. Omega-3 polyunsaturated fatty acids as a treatment strategy for non-alcoholic fatty liver disease. Pharmacol Ther. 2018, 181, 108–25. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Kusakabe, M.; Imanishi, I.; Hisano, T.; Fukamachi, T.; Taguchi, N.; Iyori, K.; Hsiao, Y. H. A randomised, double-blinded, controlled trial to determine the efficacy of combined therapy of oclacitinib and marine oil extract PCSO-524 in dogs with atopic dermatitis. Veterinary dermatology. 2023, 34, 523–531. [Google Scholar] [CrossRef]
- Tu, T.; Li, B.; Li, X.; Zhang, B.; Xiao, Y.; Li, J.; Qin, F.; Liu, N.; Sun, C.; Liu, Q.; Zhou, S. Dietary ω-3 fatty acids reduced atrial fibrillation vulnerability via attenuating myocardial endoplasmic reticulum stress and inflammation in a canine model of atrial fibrillation. Journal of cardiology. 2022, 79, 194–201. [Google Scholar] [CrossRef]
- Meineri, G.; Cocolin, L.; Morelli, G.; Schievano, C.; Atuahene, D.; Ferrocino, I. Effect of an Enteroprotective Complementary Feed on Faecal Markers of Inflammation and Intestinal Microbiota Composition in Weaning Puppies. Veterinary sciences. 2023, 10, 434. [Google Scholar] [CrossRef]
- Schauf, S.; Nakamura, N.; Castrillo, C. Effect of Calsporin (Bacillus subtilis C-3102) addition to the diet on faecal quality and nutrient digestibility in healthy adult dogs. J. Appl. Anim. Nutr. 2019, 7. [Google Scholar]
- de Lima, D.C.; Souza, C.M.M.; Nakamura, N.; Mesa, D.; de Oliveira, S.G.; Félix, A.P. Dietary supplementation with Bacillus subtilis C-3102 improves gut health indicators and fecal microbiota of dogs. Anim. Feed. Sci. Technol. 2020, 270, 114672. [Google Scholar] [CrossRef]
- Panasevich, M.R.; Daristotle, L.; Quesnell, R.; Reinhart, G.A.; Frantz, N.Z. Altered fecal microbiota, IgA, and fermentative end-products in adult dogs fed prebiotics and a nonviable Lactobacillus acidophilus. J Anim Sci. 2021, 99, skab347. [Google Scholar] [CrossRef] [PubMed]
- Pinna, C. , Vecchiato, C.G.; Bolduan, C.; Grandi, M.; Stefanelli, C.; Windisch, W.; Zaghini, G.; Biagi, G. Influence of dietary protein and fructooligosaccharides on fecal fermentative end-products, fecal bacterial populations, and apparent total tract digestibility in dogs. BMC veterinary research. 2018, 14, 106. [Google Scholar]
- Van den Abbeele, P.; Duysburgh, C.; Rakebrandt, M.; Marzorati, M. Dried yeast cell walls high in beta-glucan and mannan-oligosaccharides positively affect microbial composition and activity in the canine gastrointestinal tract in vitro. Journal of animal science. 2020, 98, skaa173. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; Grieshop, C.M.; Flickinger, E.A.; Bauer, L.L.; Healy, H.P.; Dawson, K.A.; Merchen, N.R.; Fahey, G.C.j.r. Supplemental fructooligosaccharides and mannanoligosaccharides influence immune function, ileal and total tract nutrient digestibilities, microbial populations and concentrations of protein catabolites in the large bowel of dogs. J. Nutr. 2002, 132, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Flickinger, E.A.; Schreijen, E.M.; Patil, A.R.; Hussein, H.S.; Grieshop, C.M.; Merchen, N.R.; Fahey, G.C. J.r. . Nutrient digestibilities, microbial populations, and protein catabolites as affected by fructan supplementation of dog diets. J. Anim. Sci. 2003, 81, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Ephraim, E.; Brockman, J. A.; Jewell, D. E. A Diet Supplemented with Polyphenols, Prebiotics and Omega-3 Fatty Acids Modulates the Intestinal Microbiota and Improves the Profile of Metabolites Linked with Anxiety in Dogs. Biology, 2022, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Rahman, S. U.; Huang, Y.; Zhang, Y.; Ming, P.; Zhu, L.; Chu, X.; Li, J.; Feng, S.; Wang, X.; Wu, J. Green tea polyphenols decrease weight gain, ameliorate alteration of gut microbiota, and mitigate intestinal inflammation in canines with high-fat-diet-induced obesity. The Journal of nutritional biochemistry. 2020, 78, 108324. [Google Scholar] [CrossRef]
- Scarsella, E.; Cintio, M.; Iacumin, L.; Ginaldi, F.; Stefanon, B. Interplay between Neuroendocrine Biomarkers and Gut Microbiota in Dogs Supplemented with Grape Proanthocyanidins: Results of Dietary Intervention Study. Animals: an open access journal from MDPI. 2020, 10, 531. [Google Scholar] [CrossRef]
- Wang, W.; Zhai, T.; Luo, P.; Miao, X.; Wang, J.; Chen, Y. Beneficial effects of silibinin on serum lipids, bile acids, and gut microbiota in methionine-choline-deficient diet-induced mice. Frontiers in nutrition. 2023, 10, 1257158. [Google Scholar] [CrossRef] [PubMed]
- AlShawaqfeh, M. K.; Wajid, B.; Minamoto, Y.; Markel, M.; Lidbury, J. A.; Steiner, J. M.; Serpedin, E.; Suchodolski, J. S. A dysbiosis index to assess microbial changes in fecal samples of dogs with chronic inflammatory enteropathy. FEMS microbiology ecology. 2017, 93, 10. [Google Scholar] [CrossRef] [PubMed]
- Crost, E.H.; Tailford, L.E.; Le Gall, G.; Fons, M.; Henrissat, B.; Juge, N. Utilisation of mucin glycans by the human gut symbiont Ruminococcus gnavus is strain dependent. PLoS One. 2013, 8, e76341. [Google Scholar] [CrossRef] [PubMed]
- Caspani, G.; Kennedy, S.; Foster, J.A.; Swann, J. Gut microbial metabolites in depression: Understanding the biochemical mechanisms. Microb. Cell. 2019, 6, 454–481. [Google Scholar] [CrossRef]
- Sandri, M.; Dal Monego, S.; Conte, G.; Sgorlon, S.; Stefanon, B. Raw meat-based diet influences faecal microbiome and end products of fermentation in healthy dogs. BMC Vet. Res. 2017, 13, 65. [Google Scholar] [CrossRef]
- Gilley, S.P.; Ruebel, M.L.; Sims, C.; Zhong, Y.; Turner, D.; Lan, R.S.; Pack, L.M.; Piccolo, B.D.; Chintapalli, S.V.; Abraham, A.; Bode, L.; Andres, A.; Shankar, K. Associations between maternal obesity and offspring gut microbiome in the first year of life. Pediatr Obes. 2022, 17, e12921. [Google Scholar] [CrossRef]
Figure 1.
Rarefaction curves of the alpha diversity Shannon index between days of sampling (A), between dogs participating to the study (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: Each rarefaction curve corresponds to the 6 samples for each of the dogs reported in the legend.
Figure 1.
Rarefaction curves of the alpha diversity Shannon index between days of sampling (A), between dogs participating to the study (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: Each rarefaction curve corresponds to the 6 samples for each of the dogs reported in the legend.
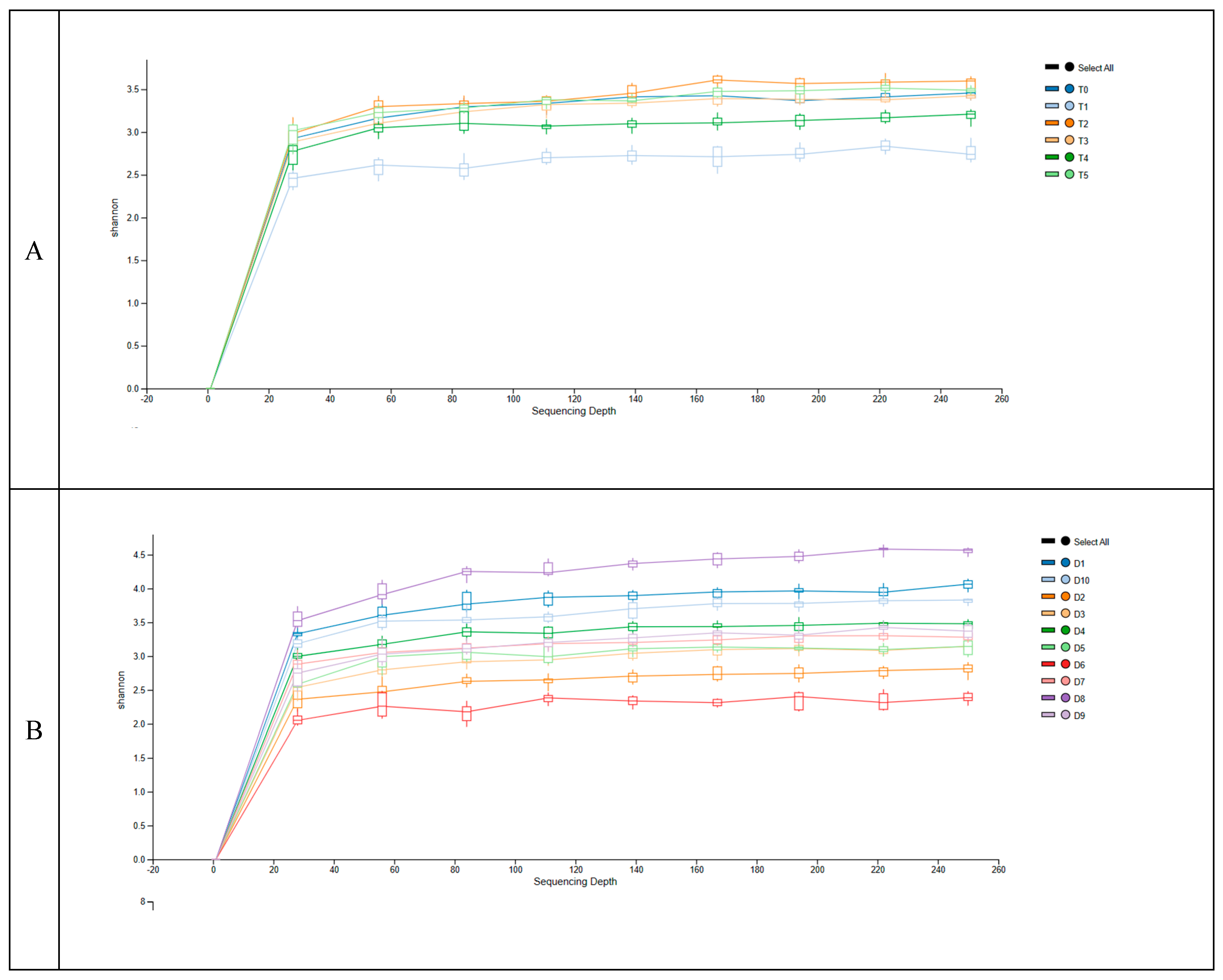
Figure 2.
Evenness of the samples between days of sampling (A); between dogs participating to the study (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: The box-plots correspond to the 6 samples for each of the dogs.
Figure 2.
Evenness of the samples between days of sampling (A); between dogs participating to the study (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: The box-plots correspond to the 6 samples for each of the dogs.
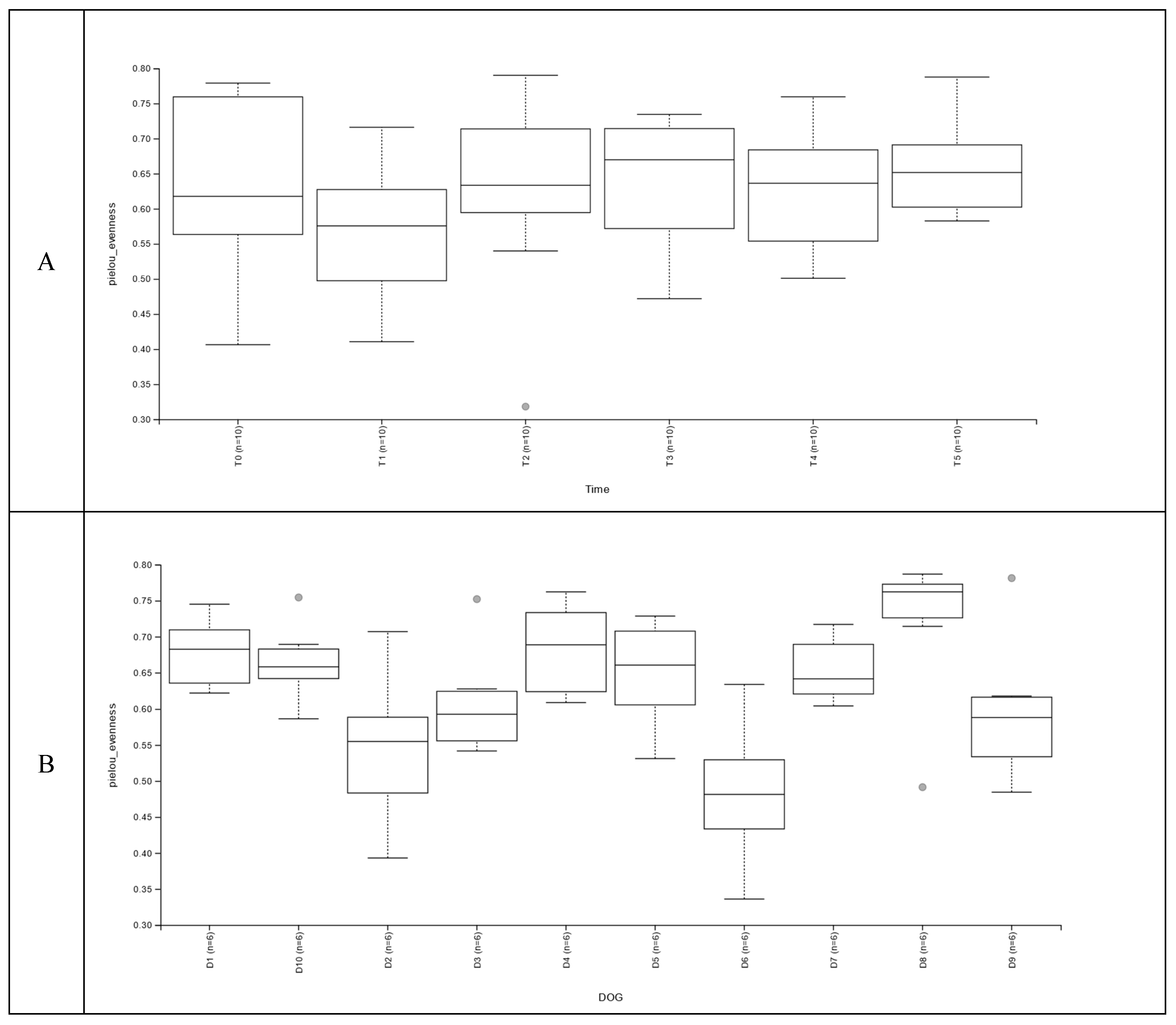
Figure 3.
Principal coordinates analysis of weighted unifrac distance metric for beta diversity between days of sampling (A); between dogs (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: The box-plots correspond to the 6 samples for each of the dogs.
Figure 3.
Principal coordinates analysis of weighted unifrac distance metric for beta diversity between days of sampling (A); between dogs (B). Panel A: T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; Panel B: The box-plots correspond to the 6 samples for each of the dogs.
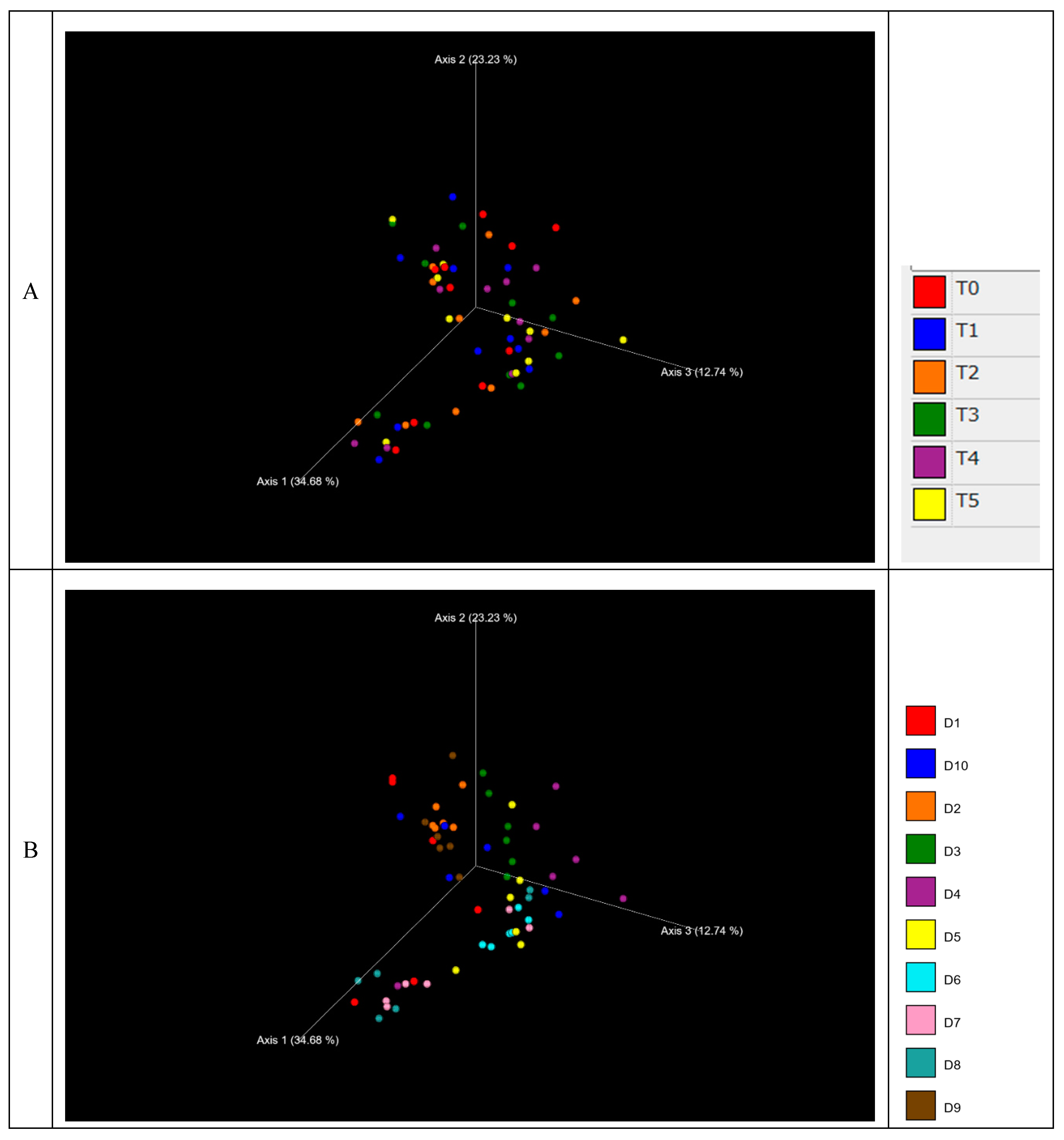
Figure 4.
Normalized counts (y axis) of Ruminococcus gnavus, Blautia, Streptococcus, Turicibacter and Peptostreptococcaceae in 10 adult dogs with liver diseases at various time points (x axis).. T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; a, b, denotes mean that significantly difer for p < 0.05.
Figure 4.
Normalized counts (y axis) of Ruminococcus gnavus, Blautia, Streptococcus, Turicibacter and Peptostreptococcaceae in 10 adult dogs with liver diseases at various time points (x axis).. T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; a, b, denotes mean that significantly difer for p < 0.05.
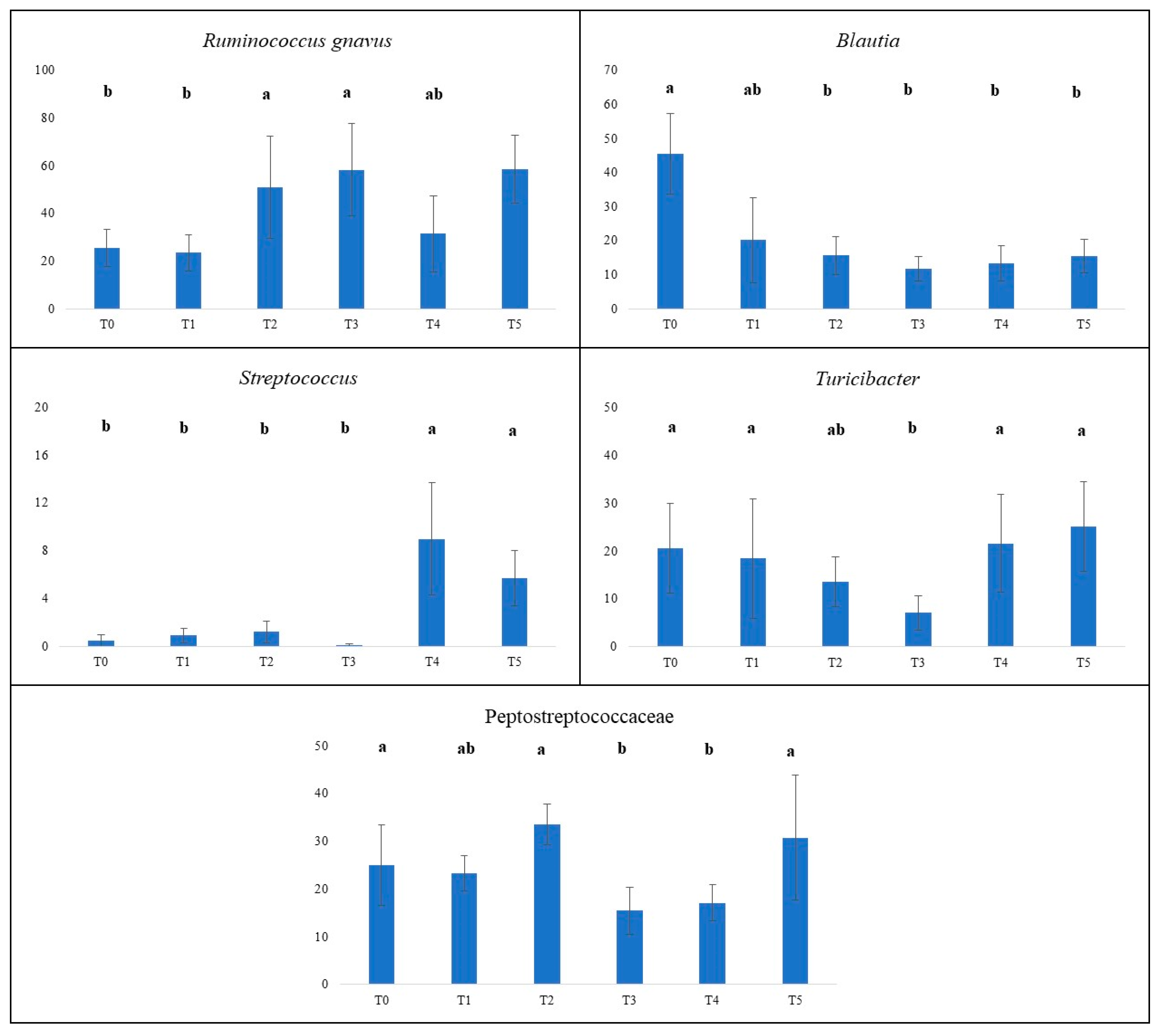

Table 1.
Data on Included Dogs. Age (years), Sex, Breed, Weight, Body Condition Score.
| Dog number | Age | Sex | Breed | Weight (kg) | BCS |
|---|---|---|---|---|---|
| D1 | 12 | F | American staffordshire terrier | 30 | 7 |
| D2 | 10 | F | Dachshund | 9 | 8 |
| D3 | 8 | M | English Bulldog | 30 | 8 |
| D4 | 8 | M | Shar Pei | 35,5 | 7 |
| D5 | 12 | M | Staffordshire bull terrier | 22,1 | 7 |
| D6 | 11 | F | Beagle | 17,5 | 8 |
| D7 | 13 | M | Bullterrier | 38,1 | 7 |
| D8 | 10 | F | Dalmatian | 28,8 | 7 |
| D9 | 9 | F | German shepherd | 34,8 | 6 |
| D10 | 8 | M | Bassethound | 30,5 | 8 |
Data were Recorded at the Beginning of the Trial (T0); BCS: scores 0-9; Sex: M=male; F=female.
Table 2.
Formulation of the Supplement Administered to the Dogs for the Study.
| Diet | Amount |
|---|---|
| Sylibum marianum | 2.5% |
| Choline chloride | 0.8% |
| Bacillus subtilis | 6% |
| Fructoligosaccharides | 0.5% |
| Beta glucans from barley | 0.5% |
| Betaine | 0.9% |
| n-3 polyunsaturated fatty acids (n-3 PUFA) in fish oil | 6% |
| Vitamin B12 | 0.00002% |
| Vitamin C | 0.15% |
| Alpha tocopherol | 3% |
| Zinc | 0.9% |
| Magnesium | 1.6% |
| Arginine | 0.4% |
| Taurine | 2% |
| Hydrolysed proteins | 24% |
| Maltodextrine | 48.74998% |
Table 3.
Plasma enzymatic activity, metabolites concentration and Reactive C protein concentration in the plasma of dogs with liver disease before and after the administration of supplement.
Table 3.
Plasma enzymatic activity, metabolites concentration and Reactive C protein concentration in the plasma of dogs with liver disease before and after the administration of supplement.
| Time | AST UI/l |
ALT UI/l |
ALP UI/l |
GGT UI/l |
Glucose mg/100ml |
Trygl mg/100ml |
Total bilirubin mg/100ml |
CRP mg/l |
|---|---|---|---|---|---|---|---|---|
| 23-66 | 21-102 | 20-156 | 1.2-6.0 | 65-110 | 20-112 | 0.1-0.5 | ||
| T0 | 49.5 a | 107.2 ns | 302.1 a | 6.3 ns | 106.4 a | 162.7 ns | 0.6 a | 1.2 a |
| T1 | 32.8 ab | 82.4 ns | 246.0 ab | 6.2 ns | 108.6 a | 119.1 ns | 0.4 ab | 0.9 a |
| T2 | 31.0 b | 91.2 ns | 242.9 ab | 6.3 ns | 97.4 ab | 150.6 ns | 0.4 ab | 0.8 ab |
| T3 | 34.3 ab | 91.6 ns | 208.6 b | 6.2 ns | 92.9 b | 110.5 ns | 0.3 b | 0.6 b |
| T4 | 36.0 ab | 96.0 ns | 213.4 b | 5.4 ns | 93.9 b | 114.6 ns | 0.3 b | 0.6 b |
| T5 | 40.5 ab | 91.1 ns | 204.6 b | 5.2 ns | 91.2 b | 103.1 ns | 0.3 b | 0.6 b |
| RMSE | 2.1 | 7.4 | 27.5 | 0.4 | 2.8 | 9.9 | 0.0 | 4.5 |
Abbreviations: ALT, alanine aminotransferase; ALP, alkaline phosphatase; BUN, blood urea nitrogen; GGT, gamma-glutamyl transferase; T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; a, b on the same column denotes statistical differences for P<0.05.
Table 4.
Sum of short chain fatty acid, and lactic acid (Total) and molar partitioning (%) in the faeces of dog with liver diseas before and after the administration of supplement.
Table 4.
Sum of short chain fatty acid, and lactic acid (Total) and molar partitioning (%) in the faeces of dog with liver diseas before and after the administration of supplement.
| Time | Total mmol/l |
Lactic % |
Acetic % |
Propionic % |
Isobutyric % |
Butyric % |
Isovaleric % |
|---|---|---|---|---|---|---|---|
| 23-66 | 21-102 | 20-156 | 1.2-6.0 | 65-110 | 20-112 | 0.1-0.5 | |
| T0 | 213.9 ab | 2.1 ab | 66.7 ns | 15.9 ns | 3.3 ns | 3.5 ns | 8.5 ns |
| T1 | 173.5 b | 0.4 b | 66.9 ns | 20.9 ns | 3.9 ns | 2.5 ns | 5.4 ns |
| T2 | 227.1 ab | 1.4 ab | 69.6 ns | 16.4 ns | 4.0 ns | 4.5 ns | 4.1 ns |
| T3 | 249.7 a | 2.6 a | 72.1 ns | 14.0 ns | 3.8 ns | 4.2 ns | 3.4 ns |
| T4 | 225.1 ab | 1.9 ab | 66.6 ns | 17.4 ns | 4.6 ns | 3.2 ns | 6.3 ns |
| T5 | 210.0 ab | 2.5 a | 67.4 ns | 16.7 ns | 4.3 ns | 4.1 ns | 5.0 ns |
| RMSE | 8.9 | 0.3 | 1.8 | 1.3 | 0.4 | 0.3 | 0.9 |
T0, before administration of supplement; T1, T2, T3, T4 and T5 correspond to 7, 14, 21, 28 and 35 days from the administration of supplement; a, b on the same column denotes statistical differences for P<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



