
Submitted:
29 November 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
Sustainability enhancement is one of the main challenges of aquaculture. Since feeds represent one of the major costs from an environmental point of view, it is priority to find sustainable alterna-tive ingredients for aquaculture diet. Insect meals have some advantages as ingredients for aqua-culture, like its sustainability and nutritional value. However, the biggest drawback of full-fat in-sect meal is its fatty acid profile. The objective of this work was to assess the effects of partial plant or marine-derived ingredients replacements with full-fat Tenebrio molitor meal (TM) at two differ-ent levels on growth performance and fatty acids profiles of Solea senegalensis. For this purpose, a control diet and four experimental diets were tested, two of them contained 5 and 10% w/w TM that replaced mostly fish meal. Two other experimental diets included 10 and 15% w/w TM that replaced mostly plant meals. The inclusion of insect meal resulted in an improvement in growth rate and feed efficiency in both cases. Moreover, dietary inclusion of insect meal increased muscle total protein and decreased total lipid, without changes in phospholipids, and maintaining the relative level of n-3 PUFA. In conclusion, our study demonstrated that full-fat TM inclusion up to 15% in S. senegalensis diets had no negative effects or even positive effects on fish survival, growth performance, nutrient utilization and flesh quality.
Keywords:
Tenebrio molitor
; insect meal
; sustainable protein sources
; Solea senegalensis
; fatty acids profile
1. Introduction
The growth of global demand for fish together with limitations of wild fish capture led to a rapid expansion of aquaculture and a subsequent exponential increase in prices for raw materials used in aquafeeds, primarily fish meal (FM) and fish oil (FO) [1]. Furthermore, aquaculture competes with other animal production systems for feed ingredients, which increases the demand for raw materials. To satisfy all these requirements in animal feed production, extensive research has been conducted to identify new ingredients. Moreover, since feeds represent one of the major costs from an environmental point of view [2], there is an urgent need to find sustainable alternative ingredients for animal diets. The most common alternatives to FM are plant meals, mainly soy and corn meals. These alternatives began to be competitive since 2006 when the price of FM notably increased [3]. The main problems associated with these meals are their unbalanced essential amino acid (EAA) profile, in particular the deficiency in methionine and lysine; and the high content of anti-nutritional factors (ANFs), causing pro-inflammatory effects in fish intestine [4,5]. However, in recent years the intense competition for most currently used plant protein resources (for both human consumption and terrestrial animal feeds) has resulted in a significant price increase, limiting their use as an aquaculture alternative [1]. In this context, it is crucial to develop alternative, non-traditional protein and oil sources, such as seaweed, algae and microalgae, single-cell proteins, microbial biomass and insects, and to recycle food waste, in order to meet the demand for aquaculture feed in the future and to promote a sustainable growth of aquaculture [6].
Insect meals (IM) are gaining popularity as a primary ingredient in aquafeeds. Several insect species can be raised efficiently in organic streams, while exhibiting relatively low carbon footprint and land usage [7,8]. Moreover, most edible insect species appear to be good sources of not only valuable nutrients but also compounds that modulate animal microbiota and improve animal health [9]. In general, IM are good sources of protein (ranging from 45% to 70%), lipids (8% to 35%), some essential amino acids and, most minerals and B-complex vitamins [10]. Among the different candidate species to produce IM for aquaculture, Tenebrio molitor (TM) (commonly known as yellow mealworm) has been considered as one of the most promising protein sources for replacing FM in aquafeeds [11]. TM are rich in crude protein (53.2%) and fat (34.5%) [10,12], have an adequate amino acid profile, although with a limited amount of total sulfur amino acids methionine and cystine [10,13]. Regarding micronutrients, TM is an excellent source of zinc, selenium, riboflavin, biotin, pantothenic acid and folic acid [10]. However, deficiencies in calcium, vitamin D3, vitamin A, vitamin B12, thiamine, vitamin E, iodine, manganese and sodium are also possible [10]. However, it is interesting to note that many of them are influenced by the insect diet, or can be modified with UVB exposure (e.g. vitamin B12) [14]. While others, such as vitamin A content, cannot generally be significantly modified in insects [14]. Regarding the fatty acid (FA) profile, TM meals have a high content in oleic acid (OA, 18:1n-9), palmitic acid (PA, 16:0) and linoleic acid (LA, 18:2n-6) [15]. However, the high lipid content and the lack of n-3 long-chain polyunsaturated FA (LC-PUFA), such as eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3), are two major disadvantages [16]. Use of full-fat TM can worsen the lipid health indices (n-3/n-6 ratio, atherogenicity, and thrombogenicity indices) of fillets [17,18,19] Although it is possible to defat TM meals to mitigate potential adverse effects of insect lipids, the processes for fat extraction and protein purification should be carefully considered since they reduce environmental sustainability and profitability of IM and may diminish its nutritive and functional value [20]. Therefore, it is essential to understand the effects of insect meal inclusion on growth performance, survival and FA profiles of edible fish parts.
Mediterranean aquaculture is highly demanding of sustainable, cost-effective feeds to support a competitive growth. Previous studies in European sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) have demonstrated the feasibility of using full-fat TM meals up to 25% of inclusion in diets with a low content of terrestrial plant (<25%), without affecting growth performance [15,21,22]. In diets with a medium plant component content (40%) for the European sea bass, the inclusion rates can be increased in defatted TM meals up to 80% without detrimental effects on nutrient digestibility and growth performance [23]. Regarding the effects of full-fat TM meal on fish FA profile, it has been reported an increase in OA and LA, as well as, a decrease of n-3 LC-PUFA [17,19,24]. Unlike D. labrax and S. aurata that are strict carnivorous fish with limited de novo FA biosynthesis capacity, the flatfish Solea senegalensis is an interesting targeted model in Europe for new feedstuffs. This species is highly efficient in biosynthesising LC-PUFA such as DHA, showing optimal survival and growth rates when fed with diets with low fish-oil inclusion and reduced levels of DHA and EPA [25,26]. To our knowledge, there are no studies that have dealt with the use of TM meals as feed ingredients in Senegalese sole (Solea senegalensis) yet. The aim of this study was to assess the effects of partial plant or marine-derived ingredients replacements with full-fat TM meal at two different levels on growth performance and FA profiles of S. senegalensis.
2. Materials and Methods
2.1. Source and composition of TM meal
Insect meal from T. molitor was provided by Beetle Genius S.L. (Brussels, Belgium). Proximate and lipid compositions are depicted in Table 1 and Table 2. Total lipid content was 35.1% with high levels of triacylglycerides (TAG) and free FA (FFA) (Table 2). The most abundant FA were OA (51.7% Total FA), LA (18.0% Total FA) and PA (18.0% Total FA) (Table 2).
2.2. Experimental diets
Five iso-nitrogenous and isolipidic experimental diets were formulated (Table 1). Two of them contained 5 and 10% w/w TM meal that replaced mostly fish meal. They were named as FM5 and FM10, respectively. Two other experimental diets included 10 and 15% w/w TM meal that replaced mostly plant meals (designed as PP10 and PP15, respectively). The fifth diet was TM meal free and used as the control (CTRL). All diets including TM meal were adjusted to achieve iso-lipidic diets by modulating oil content (mainly soybean oil). The experimental diets were formulated and manufactured by the Service of Experimental Diets from CEIMAR-University of Almería (Almeria, Spain) using standard aquafeed processing procedures. Briefly, all ingredients were mixed in a 120 L mixer, ground with a hammer mill (UPZ 100, Hosokawa-Alpine, Augsburg, Germany) to 0.5 mm. The diets were extruded in a twin-screw extruder (Evolum 25, Clextral, Firminy, France), fitted with adequate die plates for manufacturing 2 and 3 mm sinking pellets. The extruder barrel consisted of four sections and the temperature profile in each section (from inlet to outlet) was 100°C, 95°C, 95°C and 90°C, respectively. The pellets were dried after extrusion at 30°C using a 12m3-drying chamber with forced-air circulation (Airfrio, Almería), and cooled at ambient temperature. Vacuum oil coating was done on the following day in a Pegasus PG-10VC LAB vacuum coater (Dinnissen, The Netherlands). Then, feeds were kept in sealed plastic bags at −20°C until use.
The formulation and chemical composition of the experimental diets are shown in Table 1 and Table 2. Crude protein and total lipids of all experimental diets were approximately 53% and 16% on a dry matter basis, respectively (Table 1). All diets had a similar content of total saturated FA (Table 2), with slight differences in PA that were proportional to the TM meal inclusion. The greatest differences between diets were associated with total monounsaturated FA (MUFA), and n-6 and n-3 PUFA. The control diet (CTRL) had the lowest content of MUFA and the highest levels of n-3 LC-PUFA (mostly DHA and EPA) and n-6 PUFA (mostly LA). The inclusion of TM meal increased OA and FFA and decreased LA content.
2.3. Fish and experimental design
Senegalese sole specimens were provided by the Aquaculture company Cupimar (San Fernando, Cádiz, Spain). Fish were transported to IFAPA El Toruño (El Puerto de Santa Maria, Cadiz, Spain) and acclimated in 5,000 L tanks for six months in a flow-through circuit. Before the trial, all animals were sampled and intraperitoneally injected with an electronic Passive Integrated Transponder (PIT)-tag transponder (Trovan, Fish-Tags©, Melton, UK) as previously described [27].
To evaluate the effect of experimental diets on growth, a total of 1,050 fish were randomly distributed into fifteen tanks (total volume 360 L; three replicates per treatment) at an initial density of 70 fish tank−1. All the tanks were connected to a recirculation system (RAS) equipped with a cooling, mechanical filter, skimmer, ultraviolet lights and biofilter. During the trial, temperature, salinity and oxygen were daily monitored. Values oscillated in the ranges 19-24ºC, 38-41 ppt and 3-7 ppm. Mean weight at the beginning of the experiment was 215 ± 5g without statistically significant differences across tanks (Table 3).
Diets were supplied with automatic feeders (Mirafeed©; Innovaqua, Lebrija, Spain) between 01:00 and 19:00h in 72 doses. The amount of feed supplied was weekly adjusted to fit the expected total biomass (based on the lasted sampling, 0.5-1% of total biomass). Moreover, the daily feed left over in tanks were considered to adjust tank feed supplied.
All procedures were previously authorized by the Bioethics and Animal Welfare Committee of IFAPA and given the registration number 22/11/2021/182 by the National authorities for regulation of animal care and experimentation.
2.4. Fish sampling
Individual weight was monthly recorded from the onset of the trial up to three months. Four samples were carried out: starting point (t0), day 43 days (t1), day 64 (t2) and day 98 (t3). Before each sampling, specimens were fasted for one day and anesthetized before handling (2- MS-222, 200 ppm). Weight and PIT-Tags were automatically registered using a FISH Reader Weight (Zeuss, Trovan, Spain).
In the last sampling, thirty fish (six fish by diet) were sacrificed with an anesthesia overdose (MS-222, >500 mg/L). Liver and muscle samples were frozen in liquid nitrogen and kept at −80 ºC until analysis.
2.5. Growth performance and nutrient utilization
Growth performance was assessed by different parameters according to the following formulae: daily gain (DG, g day-1) = (Wf − Wi) / ∆days; specific growth rate (SGR, % d-1) = (Ln (Wf) − Ln (Wi) / ∆days) × 100, where Wf and Wi were the final and initial fish weight. Nutrient utilization indices were estimated as follows: feed conversion ratio (FCR) = total feed intake on dry basis (g) weight gain (g)-1 and protein efficiency ratio (PER) =WG total protein ingested (g)-1, where WG was the weight gain (g).
2.6. Biochemical Analysis
2.6.1. Proximate composition of diets and tissues
Gross proximate compositions of feeds (protein, lipid, ash and moisture) and fish tissues (protein and lipid) were determined according to standard procedures [28]. Briefly, moisture contents were obtained after drying the sample in an oven at 110 °C for 24 h and ash contents were determined after incineration at 600 °C for 16 h. Crude protein was measured by determining nitrogen content (N × 6.25) using automated Kjeldahl analysis (Tecator Kjeltec Auto 1030 analyser, Foss, Warrington, UK) and total lipids were extracted from feeds and tissues of the experimental fish and quantified according to the method of Folch, et al. [29].
2.6.2. Total lipids, lipid classes and FA analyses
For lipids extraction approximately 200 mg of ground feed, or fish tissues were placed in ice-cold chloroform/methanol (2:1, by vol) and homogenised with an Ultra-Turrax tissue disrupter (Fisher Scientific, Loughborough, UK). Next, the non-lipid and lipid layers were separated by addition of 0.88% (w/v) KCl. The upper aqueous layer was then aspirated and discarded, whereas the lower organic layer was dried under oxygen-free nitrogen. The lipid content was determined gravimetrically after drying overnight in a vacuum desiccator.
Separation of main lipid classes was realized in 20 × 10 cm plates by double development high-performance thin-layer chromatography (HPTLC) using the technics described by Olsen and Henderson [30]. Firstly, plates were pre-run in diethyl ether and then activated at 120 °C for 1 h. The lipid classes were visualized after spraying with 3% (w/v) copper acetate, containing 8% (v/v) phosphoric acid by charring at 160 °C for 20 min. Quantification was made by densitometry using a CAMAG-3 TLC scanner (Version Firmware 1.14.16; CAMAG, Muttenz, Switzerland) with winCATS Planar Chromatography Manager. Samples and authentic standards run alongside, in the same conditions, on high-performance thin-layer chromatography (HPTLC) plates, as the way to determine the identities of individual lipid classes by contrasting Rf values.
Fatty acid methyl esters (FAME) of total lipids were prepared by acid-catalysed transesterification at 50 °C for 16 h according to Christie [31]. Firstly, the FAME were separated and quantified by gas-liquid chromatography (Agilent Technologies 7890B GC System) using a 30 m × 0.32 mm i.d. fused silica capillary column (SUPELCOWAXTM-10, Supelco Inc., Bellefonte, PA, USA) and on-column injection at 50 °C. Hydrogen was used as carrier gas and temperature programming was from 50 °C to 150 °C at 40 °C per min and then to 230 °C at 2.0 °C per min. Then, individual methyl esters were identified by comparison with known standards and by reference to published data [32,33]. Agilent Technologies Openlab CDS Chemstation for Windows (version A.02.05.21, Santa Clara, CA, USA) was used to collect and process data.
2.7. Statistical analysis
Mean and standard error of the mean (SEM) were calculated using SPSS statistics v22 software (IBM, Armonk, USA). A two-way repeated measures ANOVA was performed to test the effect of the five diets and the four sampling points on growth using the fish weight and SGR. Tank was added to the model as a random factor. A one-way ANOVA was performed to evaluate the effect of diets on initial and final fish weight and on the nutritional utilization parameters (FCR and PER). Two-way ANOVAs were run to test the effect of diets and gender on muscle fillets and liver protein and lipid composition at the end of trial. Where ANOVA indicated a significant difference (p<0.05) for a given factor, the source of the difference was identified using a Tukey test. Moreover, principal component analysis (PCA) was conducted to FA matrixes for the ordination of samples, using PRIMER6 packages.
BeforE running all parametrict tests, the normality was confirmed with Kolmogorov–Smirnov test (p>0.05) and the homogeneity of variances with Levene test (p>0.05). The proportions were transformed by arcsine transformation before analysis [34]. If variances remained heterogeneous even after data transformation, untransformed data were still analysed, as ANOVA is a robust statistical test and is relatively unaffected by the heterogeneity of variances [35]. In such cases, the level of significance was reduced to <0.01 to avoid type I error.
3. Results
3.1. Growth performance and proximate composition
No mortality was detected through the whole trial for any of the diets. Fish growth was monitored by weight and SGR are depicted in Figure 1. Using a longitudinal approach, we identified a statistically significant interaction diet × time (within-subjects) for weight and SGR, with diet PP15 showing a higher weight gains and different growth rates between periods than CTRL. As average, we found a significantly higher mean SGR (between-effects) in the period for PP15 compared to CTRL. Moreover, dietary inclusion of TM meal slightly improved nutrient utilization as determined by FCR and PER, although these differences were not statistically significant (p > 0.05; Table 3).
Total lipid and protein content of fish fillet muscle are shown in Figure 2. In general, dietary inclusion of TM meal reduced lipid and increased protein contents in sole fillets (p < 0.05). The PP15 diet showed the lowest levels of total lipids in muscle (8.6%). Regarding liver, the average total lipid content was 59% (in dw), without differences between experimental groups (p > 0.05) (data not shown). No differences in lipid or protein associated with gender were detected.
3.2. Lipid classes and fatty acid profile
Since we found differences in muscle total lipids between dietary treatments, we analysed the lipid classes composition in this tissue (Figure 3). Interestingly, no significant differences were detected for polar lipids among dietary treatments (p < 0.05). However, we observed a lower content of neutral lipids such as sterols, triacylglycerols and others neutral lipids (mainly free FA and sterol esters) in sole fed TM-based diets. This depletion was more important at the highest level of insect inclusion, regardless of the kind of dietary ingredients replaced (fish or plant meals).
Changes in hepatic and muscle FA profile were explored by a Principal Component Analysis (PCA) multivariate analysis. The first two components of PCA explained 86% of total variance in liver (Figure 4A). PC1 (70.4% total variance) correlated positively with OA (r = 0.800; p < 0.0005) and negatively with LA (r = -0.503; p < 0.005). PC2 (16.0% total variance) correlated negatively with DHA (r = −0.801, p < 0.0005) and positively with LA (r = 0.517, p < 0.0025). The two-way ANOVA analyses confirmed the significant differences for FA that correlated with PCA axis, except for DHA (Figure 4B). In this regard it should be noted that dietary inclusion of insect meal increased OA and decreased LA content in liver.
Regarding the PCA analysis for muscle, the first two PCA components explained 88% of total variance (Figure 5A). PC1 (49.4% total variance) correlated positively with DHA (r = 0.648, p < 0.0005) and OA (r = 0.373, p < 0.025), and negatively with LA (r = −0.575, p < 0.0005); clustering the samples in two main groups according to dietary inclusion of TM meal: i) fish fed diets CTRL and FM5 (higher content of LA); and ii) fish fed FM10, PP10 and PP15 (higher content of OA). PC2 (40.3% total variance) correlated negatively with OA (r = −0.797, p < 0.0005) and positively with DHA (r = 0.648, P < 0.0005). The two-way ANOVA confirmed differences for linoleic and oleic acids (Figure 5B). TM-fed fishes showed lower levels of LA and higher levels of OA in muscle. For DHA, differences related with dietary treatments were not clearly associated to dietary inclusion of insect meal, and no differences were found between fish fed CTRL and diets PP10, PP15, FM5 and FM10.
Hepatic and fillet muscle FA profiles of fish fed dietary treatments are shown in detail in Table 4 and Table 5. The most abundant FA found in Senegalese sole liver and muscle were PA, OA, LA and DHA, reflecting those of experimental diets. In liver, OA and hexadecenoic fatty acid (16:1n-9) were higher in fish fed TM-based diets. In contrast, LA and eicosadienoic acid (20:2n-6), as well as linolenic acid (LNA, 18:3n-3) and eicosatrienoic acid (20:3n-3) were higher in liver of fish fed CTRL diet. However, no differences were found between fish fed CTRL and TM-based diets for eicosapentanoic acid (EPA, 20:5n-3), docosapentanoic acid (DPA, 22:5n-3), DHA, n-3 PUFA and ratio n-3 PUFA/n-6 PUFA. In muscle, the effect of dietary inclusion of insects in the FA profile was similar to that observed in liver, detecting higher level of hexadecenoic acid (16:1n-9) and OA, and lower of vaccenic acid (18:1n- 7), LA, LNA, eicosatrienoic acid, eicosatetraenoic acid (20:4n-3), and EPA in fish fed TM-based diets. While no significant differences were found between fish fed CTRL and TM-based diets for DPA, DHA and n-3 PUFA. Interestingly, the higher ratio n-3 PUFA/n-6 PUFA was detected in fish fed diets FM10, PP10 y PP15.
4. Discussion
Recently the use of insect meal as a dietary ingredient in aquaculture has received an increasing interest. As ingredients for aquaculture diets, insects have some advantages like its sustainability and nutritional values [36]. However, one of the biggest drawbacks of full-fat insect meal is its fatty acid profile. In this study we investigated the effects on growth performance and lipid profile of partial replacement of plant or marine-derived ingredients with full-fat TM meal in the flatfish S. senegalensis. Our results showed that partially replacement of fish meal up to 10% replacement had no negative effect on growth (as determined by fish weight and SGR), nutrient utilization (FCR and PER) and fish survival. More interesting, substitution of plant meals by TM meal up to 15% significantly improved growth rates. In other studies in marine species, authors did not find adverse effects on growth parameters of full-fat TM meal up to total ingredient replacement values of 25% [15,22,37]. In general, it is not easy to compare the present results with those previous reports due to the species-specific characteristics and differences between tested diets. In order to put our results in context, it is important to point out that our control group (CTRL) had a low content in total marine ingredients (46%), in order to reflect the current levels used in commercial diets. A previous study with rainbow trout, tench and sea bream used experimental diets similar to those of the present study, in terms of levels of marine ingredient inclusion and fish meal replacement, and they did not find either statistic differences for weight gain among control and fish fed diets that included TM meal in percentages of up to 10% [37]. Instead, others works with European sea bass [15] and sea bream [22] formulated reference diets with a high content in marine ingredients (over 64%), and both concluded that the optimal full-fat TM meal dietary inclusion level was 25%.
Some authors have explained dietary inclusion of insects modulates growth performance by modifying fish gut microbiota through the prebiotic activity of chitin [38,39,40]. The effect of chitin on protein digestibility is well established in literature due to the high protein binding capacity of this polysaccharide [22]. Nevertheless, the effect of chitin on fish nutrition is dose-dependent and, moreover, chitin might be considered as an antinutritive compound at high doses which may decrease feed palatability, nutrient digestibility and fish growth [41]. In this study, chitin content of TM-based diets is lower than 1% dry weight (data not shown), below the minimum level of dietary chitin that may cause a reduction in fish growth [15,22]. In view of these results, it would be interesting to test higher dietary inclusion of TM meal, since we observed in both cases (marine and plant ingredients replacement) a trend to improve growth performance when we increased the TM inclusion. However, the biggest limitation to rise the dietary inclusion of full-fat TM meal is its high lipid content, and the drastic decrease of dietary n-3 PUFA.
Regarding the effect of full-fat TM inclusion on muscle and liver of Senegalese sole, dietary inclusion of full-fat insect meal decreased total lipid and increased muscle total protein content without changes in liver total lipids content. It is interesting to highlight that reduction in muscle total lipids was due to a decrease in the content of the principal neutral lipids: triacylglycerols and cholesterol; without changes in phospholipids. Similarly, Jeong, et al. [19] and Belforti, et al. [42] detected a decrease in muscle total lipids in olive flounder and rainbow trout, respectively, while this effect was not fully consistent and other authors failed to find changes in the fillet composition of blackspot sea bream [18], mandarin fish [43], and rainbow trout [44]. We hypothetize that differences in muscle lipid content in sole might be related to the effect of chitin reducing fat digestibility and lipid absorption [41,45]. The use of chitinous polymers as a feed additive to animal diets have shown lower cholesterol and triacylglycerol values in rabbits and rats without any adverse effect on normal growth pattern [46,47]. The chemical properties of chitin can reduce activity of digestive enzymes as demonstrated for porcine pancreatic lipase [48]. In this sense, Hansen, et al. [49] found a decrease in bile acid concentration in the pyloric intestine in fish fed a diet with a high chitin content. These authors conclude that bile acids probably reduced lipid digestibility because the pyloric region is the main area for lipid digestion and bile acids are essential for lipase activation as well as for efficient fatty acid absorption. A reduction in lipid absorption and fat digestibility could also explain the enhancement of growth observed in sole in this experiment. In that sense, Borges, et al. [26] demonstrated that low dietary lipid levels improved nutrient retention and growth in S. senegalensis, without major effects on whole-body composition, and Campos, et al. [50] suggest that dietary lipid level has a great impact on the expression of genes related with growth in S. senegalensis.
Fish that were fed diets containing TM progressively enriched in TM-associated FA, such as oleic acid, at the expense of linoleic acid, the major component of soybean oil (adjusted in experimental diets to achieve iso-lipidic diets). However, we found no significant differences in n-3 PUFA and DHA content between fish fed TM-based diets and CTRL group. The same pattern was observed in both tissues analysed: liver and muscle. Some authors have pointed that dietary inclusion of TM meal increased the content of oleic and linoleic acid and decreased n-3 PUFA in fish tissues [15,17,19,42,43,51]. Intriguingly, we observed a different pattern for LA content in sole tissues. This discrepancy can be explained by: i) the relatively low content of LA in TM meal used in this work (18% TFA) compared to others studies (35% TFA) [15,24,42]; and ii) the fact that in the present work we lower dietary soy oil to compensate the high lipid content of TM meal. Moreover, it is interesting that n-3 PUFA and DHA content in sole muscle were maintained, despite the reduction in dietary n-3 PUFA (max reduction: 26%) and specifically in DHA (max reduction: 23%). This result could be explained through the DHA biosynthesis and/or selective deposition of the dietary DHA, since this FA is usually accumulated in tissues at higher levels than those present in diets [52]. Regarding the PUFA biosynthesis capacity, Senegalese sole possesses a ∆4 Fads2 that enables this species to biosynthesise DHA from docosapentaenoic acid (22:5n-3) via the so-called “∆4 pathway” [53]. So, dietary replacement of marine ingredients with terrestrial sources has shown good growth performance in Senegalese sole without affecting flesh content in n-3 PUFA [25,54].
5. Conclusions
In conclusion, our study demonstrated that full-fat TM meal inclusion up to 15% in Senegalese sole diets had no negative or even positive effects on fish survival, growth performance, nutrient utilization and flesh quality. Fish fed TM-based diets reduced its content in total and neutral lipids and increased the total protein in fillets, maintaining the relative level of n-3 PUFA and DHA and improving the lipid health indices n-3:n-6.
Author Contributions
Conceptualization, I.H.-C and F.J.A.; methodology, I.H.-C. and M.M.; validation, I.H.-C.; formal analysis, I.H.-C., M.M. and M.B.B.; investigation, I.H.-C. and M.B.B.; data curation, I.H.-C. and M.B.B.; writing—original draft preparation, I.H.-C.; writing—review and editing, I.H.-C., F.J.A., M.B.B. and M.M.; supervision, I.H.-C.; project administration, I.H.-C. and A.C.; funding acquisition, I.H.-C. and A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-funded 75% by European Maritime and Fisheries Fund (EMFF) project FEM.PPA201900.001 “Ecointensificación de la acuicultura mediante tecnologías innovador” and contract 101/2021. Authors acknowledge the support of the University of Almeria (Experimental feeds Service, grant EQC2019-006380-P) on aquafeed elaboration and project AquaTech4Feed project (grant # PCI2020-112204 by AEI within the ERA-NET BioBlue COFUND).
Institutional Review Board Statement
All procedures were authorized by the Bioethics and Ani- mal Welfare Committee of IFAPA and given the registration number 22-11-2021-182 by the National authorities for regulation of animal care and experimentation.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon any reasonable request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation. 2022. [CrossRef]
- Nijdam, D.; Rood, T.; Westhoek, H. The price of protein: Review of land use and carbon footprints from life cycle assessments of animal food products and their substitutes. Food Policy 2012, 37, 760-770. [CrossRef]
- Hardy, R.W. Utilization of plant proteins in fish diets: effects of global demand and supplies of fishmeal. Aquaculture Research 2010, 41, 770-776. [CrossRef]
- Montero, D.; Benitez-Dorta, V.; Caballero, M.J.; Ponce, M.; Torrecillas, S.; Izquierdo, M.; Zamorano, M.J.; Manchado, M. Dietary vegetable oils: Effects on the expression of immune-related genes in Senegalese sole (Solea senegalensis) intestine. Fish & Shellfish Immunology 2015, 44, 100-108. [CrossRef]
- Bakke-McKellep, A.M.; Penn, M.H.; Salas, P.M.; Refstie, S.; Sperstad, S.; Landsverk, T.; Ringø, E.; Krogdahl, Å. Effects of dietary soyabean meal, inulin and oxytetracycline on intestinal microbiota and epithelial cell stress, apoptosis and proliferation in the teleost Atlantic salmon (Salmo salar L.). British Journal of Nutrition 2007, 97, 699-713. [CrossRef]
- Cottrell, R.S.; Blanchard, J.L.; Halpern, B.S.; Metian, M.; Froehlich, H.E. Global adoption of novel aquaculture feeds could substantially reduce forage fish demand by 2030. Nature Food 2020, 1, 301-308. [CrossRef]
- Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. EDIBLE INSECTS future prospects fo food and feed security; 2013; Volume 171.
- Quang Tran, H.; Van Doan, H.; Stejskal, V. Environmental consequences of using insect meal as an ingredient in aquafeeds: A systematic view. Reviews in Aquaculture 2022, 14, 237-251. [CrossRef]
- Gasco, L.; Finke, M.; Huis, A. Can diets containing insects promote animal health? Journal of Insects as Food and Feed 2018, 4, 1-4. [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biology 2002, 21, 269-285. [CrossRef]
- Hua, K. A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 2021, 530, 735732. [CrossRef]
- Marco, M.; Martinez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S.; et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Animal Feed Science and Technology 2015, 209. [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innovative Food Science & Emerging Technologies 2013, 17, 1-11. [CrossRef]
- Koutsos, L.; McComb, A.; Finke, M. Insect Composition and Uses in Animal Feeding Applications: A Brief Review. Annals of the Entomological Society of America 2019, 112, 544-551. [CrossRef]
- Gasco, L.; Henry, M.; Piccolo, G.; Marono, S.; Gai, F.; Renna, M.; Lussiana, C.; Antonopoulou, E.; Mola, P.; Chatzifotis, S. Tenebrio molitor meal in diets for European sea bass (Dicentrarchus labrax L.) juveniles: Growth performance, whole body composition and in vivo apparent digestibility. Animal Feed Science and Technology 2016, 220, 34-45. [CrossRef]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a feed ingredient for fish culture: Status and trends. Aquaculture and Fisheries 2022, 7, 166-178. [CrossRef]
- Fabrikov, D.; Barroso, F.G.; Sánchez-Muros, M.J.; Hidalgo, M.C.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F.; Guil-Guerrero, J.L. Effect of feeding with insect meal diet on the fatty acid compositions of sea bream (Sparus aurata), tench (Tinca tinca) and rainbow trout (Oncorhynchus mykiss) fillets. Aquaculture 2021, 545, 737170. [CrossRef]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49-58. [CrossRef]
- Jeong, S.-M.; Khosravi, S.; Yoon, K.-Y.; Kim, K.-W.; Lee, B.-J.; Hur, S.-W.; Lee, S.-M. Mealworm, Tenebrio molitor, as a feed ingredient for juvenile olive flounder, Paralichthys olivaceus. Aquaculture Reports 2021, 20, 100747. [CrossRef]
- Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization. Animals 2020, 10. [CrossRef]
- Mastoraki, M.; Mollá Ferrándiz, P.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A comparative study on the effect of fish meal substitution with three different insect meals on growth, body composition and metabolism of European sea bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [CrossRef]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Bovera, F.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Animal Feed Science and Technology 2017, 226, 12-20. [CrossRef]
- Basto, A.; Valente, L.M.P.; Conde-Sieira, M.; Soengas, J.L. Central regulation of food intake is not affected by inclusion of defatted Tenebrio molitor larvae meal in diets for European sea bass (Dicentrarchus labrax). Aquaculture 2021, 544, 737088. [CrossRef]
- Iaconisi, V.; Bonelli, A.; Pupino, R.; Gai, F.; Parisi, G. Mealworm as dietary protein source for rainbow trout: Body and fillet quality traits. Aquaculture 2018, 484, 197-204. [CrossRef]
- Villalta, M.; Estévez, A.; Bransden, M.P.; Bell, J.G. The effect of graded concentrations of dietary DHA on growth, survival and tissue fatty acid profile of Senegal sole (Solea senegalensis) larvae during the Artemia feeding period. Aquaculture 2005, 249, 353-365. [CrossRef]
- Borges, P.; Oliveira, B.; Casal, S.; Dias, J.; Conceição, L.; Valente, L.M.P. Dietary lipid level affects growth performance and nutrient utilisation of Senegalese sole (Solea senegalensis) juveniles. British Journal of Nutrition 2009, 102, 1007-1014. [CrossRef]
- Carballo, C.; Berbel, C.; Guerrero-Cózar, I.; Jiménez-Fernández, E.; Cousin, X.; Bégout, M.L.; Manchado, M. Evaluation of different tags on survival, growth and stress response in the flatfish Senegalese sole. Aquaculture 2018, 494, 10-18. [CrossRef]
- AOAC, C. Official methods of analysis of the Association of Analytical Chemists International. Official Methods: Gaithersburg, MD, USA 2005.
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. Journal of Biological Chemistry 1957, 226, 497-509. [CrossRef]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double-development HPTLC and scanning densitometry. Journal of Experimental Marine Biology and Ecology 1989, 129, 189-197. [CrossRef]
- Christie, W.W. Lipid Analysis: Isolation, Separation, Identification, and Structural Analysis of Lipids; Amer Oil Chemists Society: 2003.
- Hachero-Cruzado, I.; Rodriguez-Rua, A.; Roman-Padilla, J.; Ponce, M.; Fernandez-Diaz, C.; Manchado, M. Characterization of the genomic responses in early Senegalese sole larvae fed diets with different dietary triacylglycerol and total lipids levels. Comp. Biochem. Physiol. Part D Genomics Proteomics 2014, 12, 61-73. [CrossRef]
- Rodiles, A.; Herrera, M.; Hachero-Cruzado, I.; Ruiz-Jarabo, I.; Mancera, J.M.; Cordero, M.L.; Lall, S.P.; Alarcón, F. Tissue composition, blood biochemistry and histology of digestive organs in Senegalese sole (Solea senegalensis) juveniles fed diets containing different plant protein ingredients. Aquaculture Nutrition 2014, 21. [CrossRef]
- Sokal, R.; Rohlf, F. Biometry : the principles and practice of statistics in biological research / Robert R. Sokal and F. James Rohlf; 2013.
- Underwood, A.J. Experiments in ecology: their logical design and interpretation using analysis of variance; Cambridge university press: 1997.
- Gasco, L.; Biasato, I.; Enes, P.; Gai, F. Potential and challenges for the use of insects as feed for aquaculture. 2023; pp. 465-492.
- Fabrikov, D.; Sánchez-Muros, M.J.; Barroso, F.G.; Tomás-Almenar, C.; Melenchón, F.; Hidalgo, M.C.; Morales, A.E.; Rodriguez-Rodriguez, M.; Montes-Lopez, J. Comparative study of growth performance and amino acid catabolism in Oncorhynchus mykiss, Tinca tinca and Sparus aurata and the catabolic changes in response to insect meal inclusion in the diet. Aquaculture 2020, 529, 735731. [CrossRef]
- Bruni, L.; Pastorelli, R.; Viti, C.; Gasco, L.; Parisi, G. Characterisation of the intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens (black soldier fly) partially defatted larva meal as partial dietary protein source. Aquaculture 2018, 487, 56-63. [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Reviews in Fish Biology and Fisheries 2019, 29, 465-486. [CrossRef]
- Henry, M.A.; Gai, F.; Enes, P.; Peréz-Jiménez, A.; Gasco, L. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish & Shellfish Immunology 2018, 83, 308-313. [CrossRef]
- Shiau, S.-Y.; Yu, Y.-P. Dietary supplementation of chitin and chitosan depresses growth in tilapia, Oreochromis niloticus×O. aureus. Aquaculture 1999, 179, 439-446. [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I.; et al. Tenebrio Molitor Meal in Rainbow Trout (Oncorhynchus Mykiss) Diets: Effects on Animal Performance, Nutrient Digestibility and Chemical Composition of Fillets. Italian Journal of Animal Science 2015, 14, 4170. [CrossRef]
- Sankian, Z.; Khosravi, S.; Kim, Y.-O.; Lee, S.-M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juveniles. Aquaculture 2018, 496, 79-87. [CrossRef]
- Jeong, S.-M.; Khosravi, S.; Mauliasari, I.R.; Lee, S.-M. Dietary inclusion of mealworm (Tenebrio molitor) meal as an alternative protein source in practical diets for rainbow trout (Oncorhynchus mykiss) fry. Fisheries and Aquatic Sciences 2020, 23, 12. [CrossRef]
- Zacour, A.C.; Silva Me Fau - Cecon, P.R.; Cecon Pr Fau - Bambirra, E.A.; Bambirra Ea Fau - Vieira, E.C.; Vieira, E.C. Effect of dietary chitin on cholesterol absorption and metabolism in rats. 1992.
- Hirano, S. Chitin Biotechnology Applications. In Biotechnology Annual Review, El-Gewely, M.R., Ed.; Elsevier: 1996; Volume 2, pp. 237-258.
- Zacour, A.C.; Silva, M.E.; Cecon, P.R.; Bambirra, E.A.; Vieira, E.C. Effect of Dietary Chitin on Cholesterol Absorption and Metabolism in Rats. Journal of Nutritional Science and Vitaminology 1992, 38, 609-613. [CrossRef]
- Kılınç, A.; Teke, M.; Önal, S.; Telefoncu, A. Immobilization of Pancreatic Lipase on Chitin and Chitosan. Preparative Biochemistry & Biotechnology 2006, 36, 153-163. [CrossRef]
- Hansen, J.Ø.; Penn, M.; Øverland, M.; Shearer, K.D.; Krogdahl, Å.; Mydland, L.T.; Storebakken, T. High inclusion of partially deshelled and whole krill meals in diets for Atlantic salmon (Salmo salar). Aquaculture 2010, 310, 164-172. [CrossRef]
- Campos, C.; Valente, L.M.P.; Borges, P.; Bizuayehu, T.; Fernandes, J.M.O. Dietary lipid levels have a remarkable impact on the expression of growth-related genes in Senegalese sole (Solea senegalensis Kaup). Journal of Experimental Biology 2010, 213, 200-209. [CrossRef]
- Basto, A.; Calduch-Giner, J.; Oliveira, B.; Petit, L.; Sá, T.; Maia, M.R.G.; Fonseca, S.C.; Matos, E.; Pérez-Sánchez, J.; Valente, L.M.P. The Use of Defatted Tenebrio molitor Larvae Meal as a Main Protein Source Is Supported in European Sea Bass (Dicentrarchus labrax) by Data on Growth Performance, Lipid Metabolism, and Flesh Quality. Frontiers in Physiology 2021, 12.
- Brodtkorb, T.; Rosenlund, G.; Lie, Ø. Effects of dietary levels of 20:5n-3 and 22:6n-3 on tissue lipid composition in juvenile Atlantic salmon, Salmo salar, with emphasis on brain and eye. Aquaculture Nutrition 1997, 3, 175-187. [CrossRef]
- Morais, S.; Castanheira, F.; Martinez-Rubio, L.; Conceição, L.E.C.; Tocher, D.R. Long chain polyunsaturated fatty acid synthesis in a marine vertebrate: Ontogenetic and nutritional regulation of a fatty acyl desaturase with Δ4 activity. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2012, 1821, 660-671. [CrossRef]
- Borges, P.; Reis, B.; Fernandes, T.J.R.; Palmas, Â.; Castro-Cunha, M.; Médale, F.; Oliveira, M.B.P.P.; Valente, L.M.P. Senegalese sole juveniles can cope with diets devoid of supplemental fish oil while preserving flesh nutritional value. Aquaculture 2014, 418-419, 116-125. [CrossRef]
Figure 1.
Weight (A) and specific growth rates (SGR) (B) of Senegalese sole fed with the experimental diets. Lines represent the estimated marginal means (EMM) for weight or the SGR in the evaluation periods as calculated by repeated measures ANOVA. Asterisks on the horizontal denote significant differences for interaction diet × time (within-subjects). Bars on the right represent the EMM for weight and SGR in the whole period. Different letter denotes statistically significant differences according to the between-subjects. FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
Figure 1.
Weight (A) and specific growth rates (SGR) (B) of Senegalese sole fed with the experimental diets. Lines represent the estimated marginal means (EMM) for weight or the SGR in the evaluation periods as calculated by repeated measures ANOVA. Asterisks on the horizontal denote significant differences for interaction diet × time (within-subjects). Bars on the right represent the EMM for weight and SGR in the whole period. Different letter denotes statistically significant differences according to the between-subjects. FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
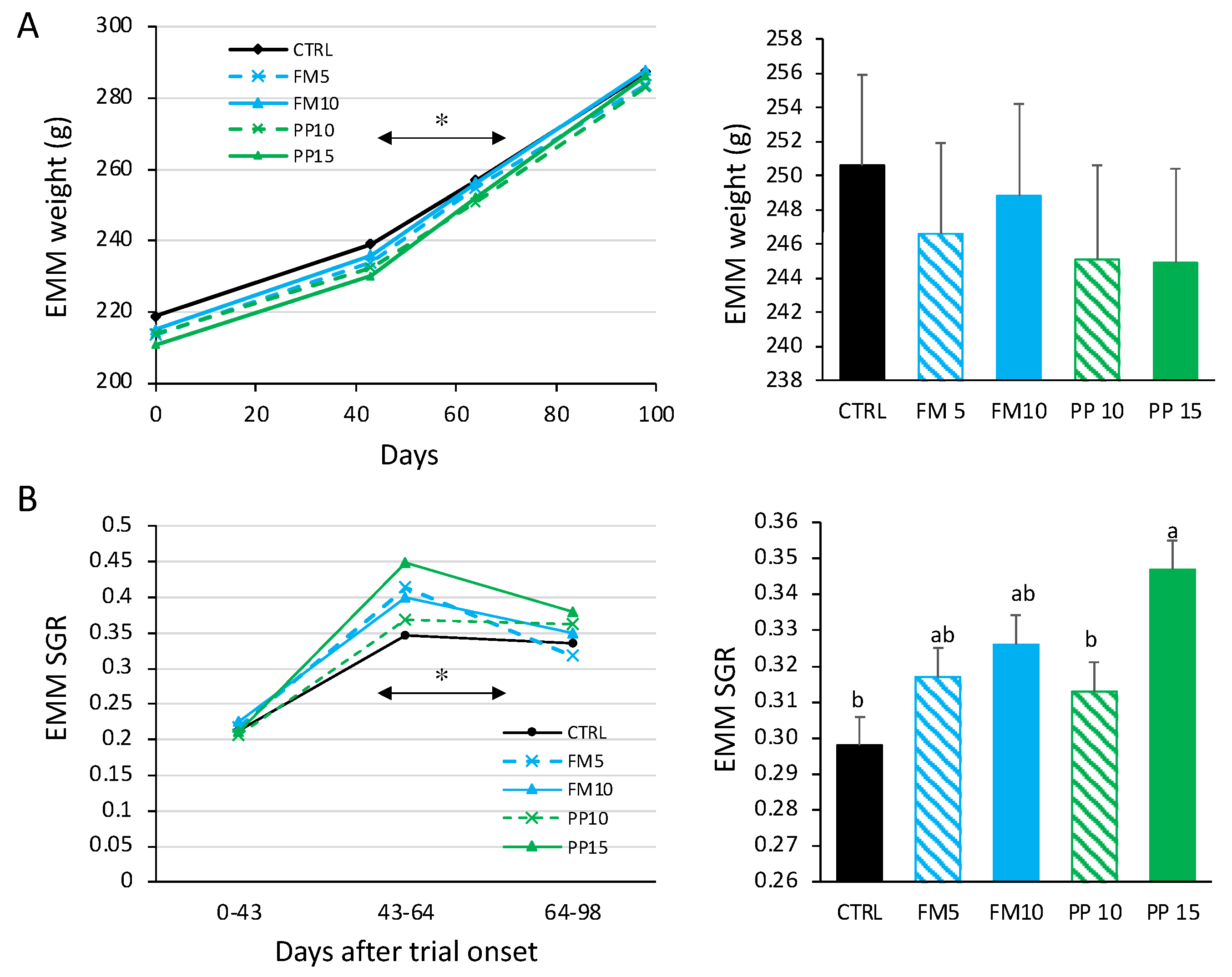
Figure 2.
Total lipids (in % dw) (A) and total protein (in % dw) (B) in fillets of Senegalese sole fed with the experimental diets after 98 days. Data are presented as mean ± SEM (n = 6). Different letters above bars indicate significant differences between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
Figure 2.
Total lipids (in % dw) (A) and total protein (in % dw) (B) in fillets of Senegalese sole fed with the experimental diets after 98 days. Data are presented as mean ± SEM (n = 6). Different letters above bars indicate significant differences between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
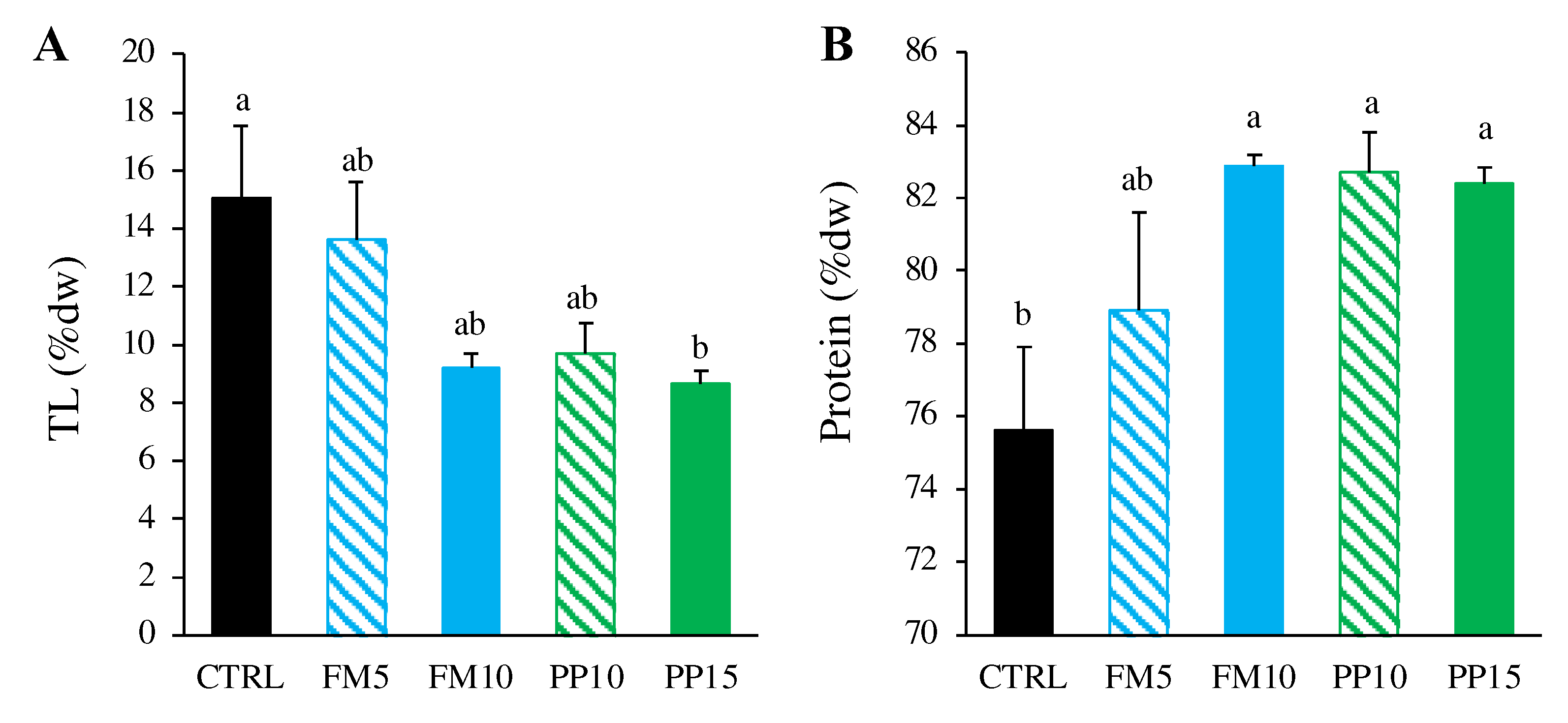
Figure 3.
Lipid classes composition (in % dw) in muscle of sole fed with the experimental diets. Data are presented as mean ± SEM (n = 6). Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). TPL, total polar lipids; ST, sterols; TAG, triacylglycerols; Others, others neutral lipids. TPL include lyso-phosphatidylcholine, sphingomyeline, phosphatidylcholine, phosphatidylserine, phosphatidylinositol, phosphatidylglycerol, phosphatidylethanolamine; others include diacylglycerols, free fatty acids and sterol esters.
Figure 3.
Lipid classes composition (in % dw) in muscle of sole fed with the experimental diets. Data are presented as mean ± SEM (n = 6). Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). TPL, total polar lipids; ST, sterols; TAG, triacylglycerols; Others, others neutral lipids. TPL include lyso-phosphatidylcholine, sphingomyeline, phosphatidylcholine, phosphatidylserine, phosphatidylinositol, phosphatidylglycerol, phosphatidylethanolamine; others include diacylglycerols, free fatty acids and sterol esters.
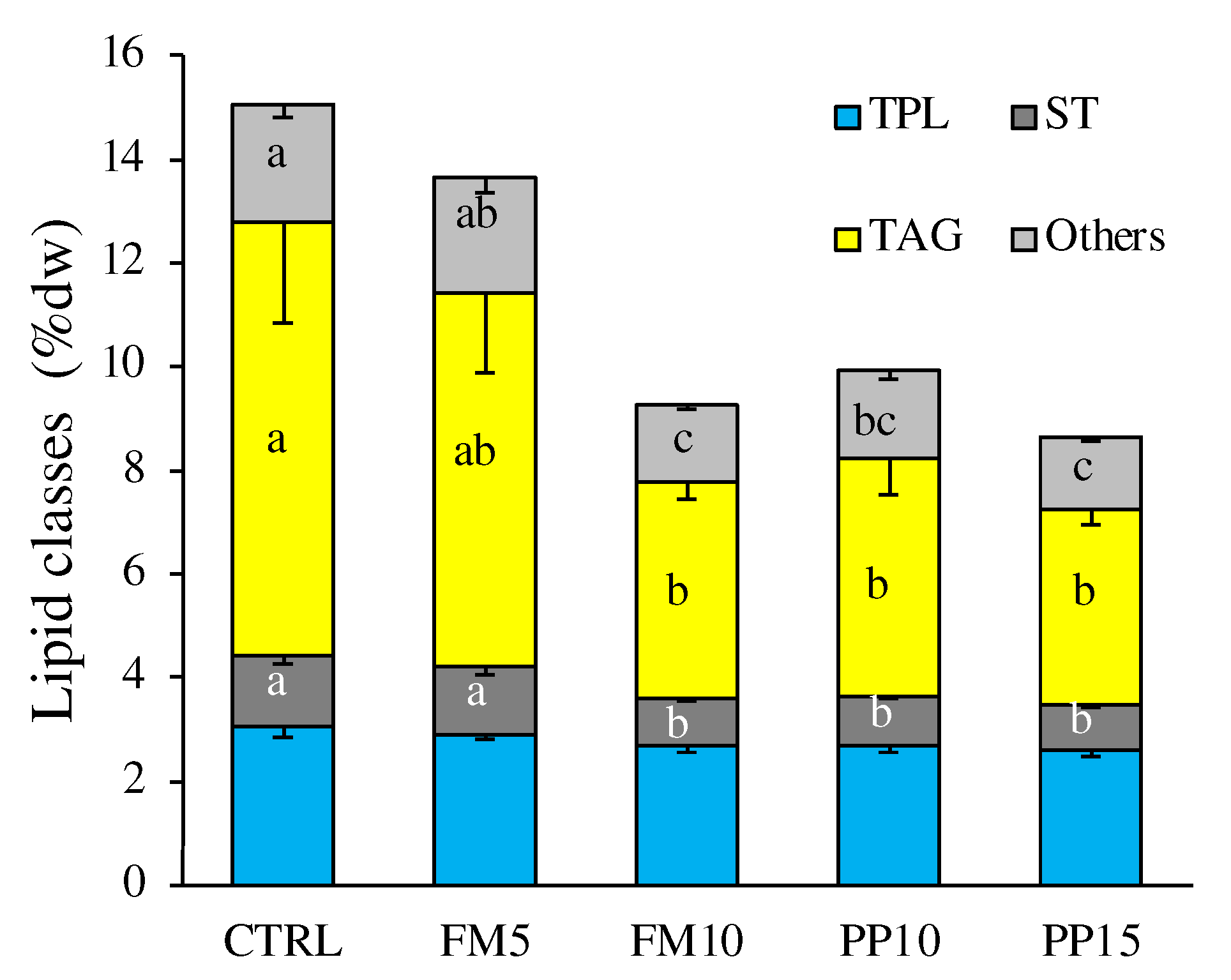
Figure 4.
Hepatic FA content in fish fed with the control (CTRL, black) or TM-based diets replacing fish meal (blue) or replacing plant meal (green). (a) Principal component análisis (PCA) plot based on FA composition (% TFA); (b) hepatic content of FA significantly correlated with PC1: 18:1n-9 and 18:2n-6; and PC2: 22:6n-3. Data are expressed as mean ± SEM (n=6). Results of two-way ANOVA are presented in the square. Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
Figure 4.
Hepatic FA content in fish fed with the control (CTRL, black) or TM-based diets replacing fish meal (blue) or replacing plant meal (green). (a) Principal component análisis (PCA) plot based on FA composition (% TFA); (b) hepatic content of FA significantly correlated with PC1: 18:1n-9 and 18:2n-6; and PC2: 22:6n-3. Data are expressed as mean ± SEM (n=6). Results of two-way ANOVA are presented in the square. Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
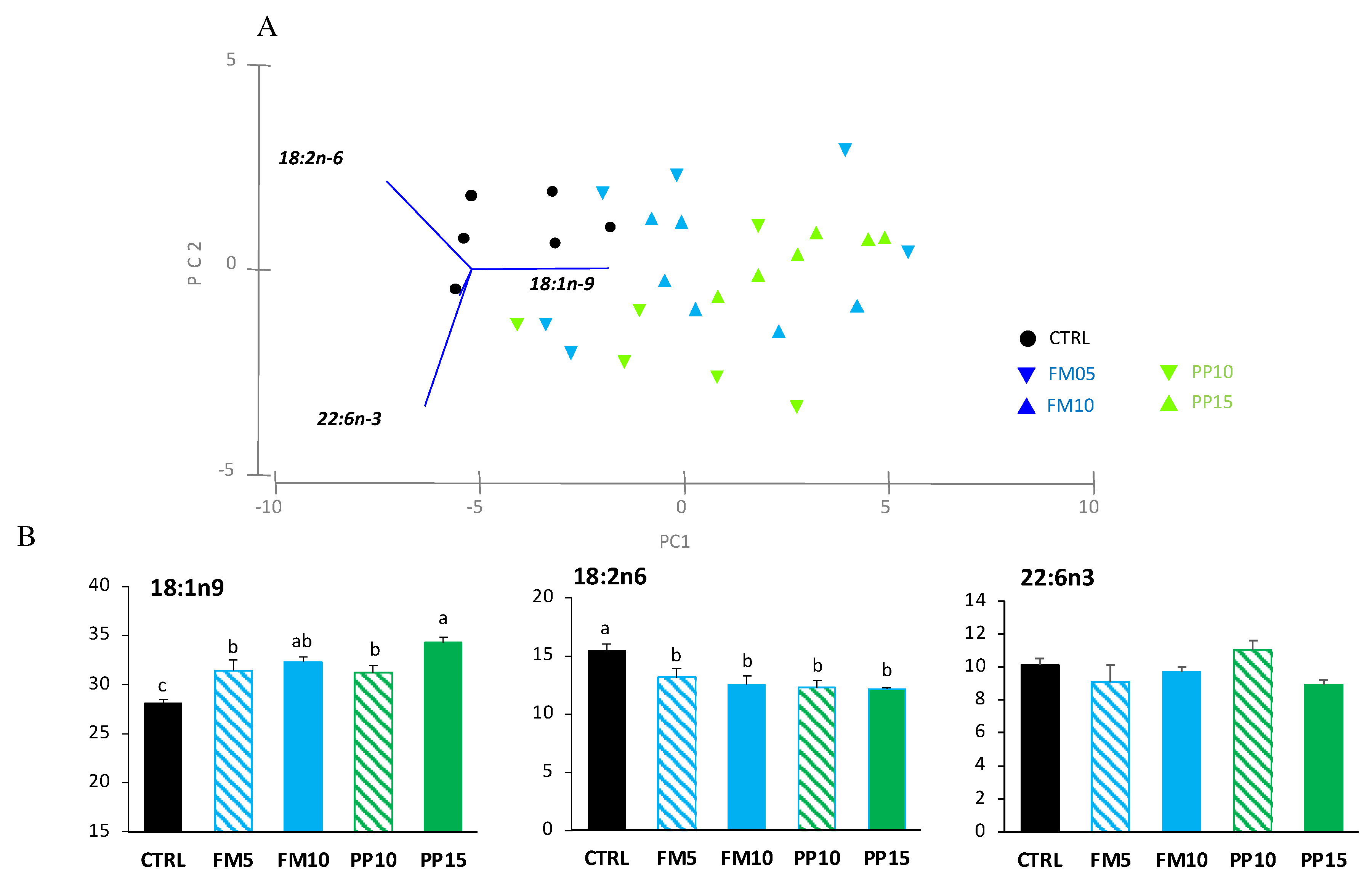
Figure 5.
Muscle FA content in fish fed with the control (CTRL, black) or TM-based diets replacing fish meal (blue) or replacing plant meal (green). (a) Principal component analysis (PCA) plot based on FA composition (% TFA); (b) muscle content of FA significantly correlated with PC1: 22:6n-3, 18:1n-9 and 18:2n-6; and PC2: 18:1n-9 and 22:6n-3. Data are expressed as mean ± SEM (n=6). Results of two-way ANOVA are presented in the square. Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
Figure 5.
Muscle FA content in fish fed with the control (CTRL, black) or TM-based diets replacing fish meal (blue) or replacing plant meal (green). (a) Principal component analysis (PCA) plot based on FA composition (% TFA); (b) muscle content of FA significantly correlated with PC1: 22:6n-3, 18:1n-9 and 18:2n-6; and PC2: 18:1n-9 and 22:6n-3. Data are expressed as mean ± SEM (n=6). Results of two-way ANOVA are presented in the square. Different letters above bars indicate significant different between dietary treatments (Tukey test; p < 0.05). FM diets are in blue and PP diets in green. The lowest TM substitution within each dietary group is indicated with dashed lines.
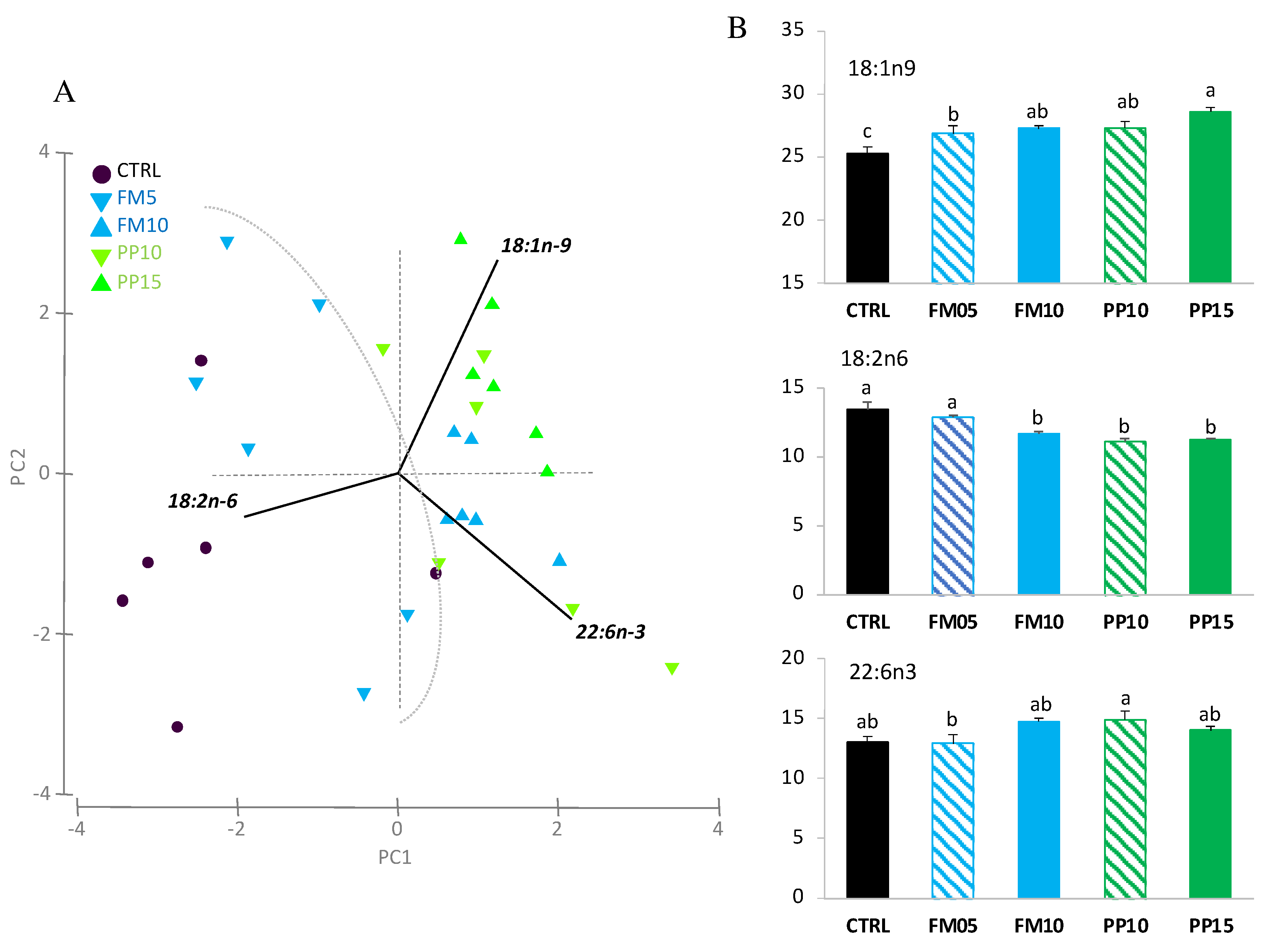

Table 1.
Formulation and chemical composition (% dw) of Senegalese sole experimental diets (CTRL, FM5, FM10, PP10, PP15) and T. molitor (TM) meal.
Table 1.
Formulation and chemical composition (% dw) of Senegalese sole experimental diets (CTRL, FM5, FM10, PP10, PP15) and T. molitor (TM) meal.
| TM | Diets | |||||||
| CTRL | FM5 | FM10 | PP10 | PP15 | ||||
| Ingredients (% dw) | ||||||||
| Fish meal LT94 1 | - | 30.00 | 26.50 | 23.20 | 30.00 | 30.00 | ||
| Squid meal 2 | - | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | ||
| CPSP90 3 | - | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | ||
| Krill meal 4 | - | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | ||
| Wheat gluten 5 | - | 8.70 | 8.70 | 8.70 | 6.80 | 5.90 | ||
| Soybean protein concentrate 6 | - | 12.00 | 12.00 | 12.00 | 9.90 | 8.90 | ||
| Pea protein concentrate 7 | - | 9.00 | 9.00 | 9.00 | 7.10 | 6.20 | ||
| Wheat flour 8 | - | 12.10 | 12.00 | 11.60 | 11.40 | 10.20 | ||
| Tenebrio molitor meal 9 | - | 0.00 | 5.00 | 10.00 | 10.00 | 15.00 | ||
| Fish oil 10 | - | 5.40 | 5.40 | 5.40 | 5.00 | 4.00 | ||
| Soybean oil 11 | - | 3.00 | 1.60 | 0.30 | 0.00 | 0.00 | ||
| Soy lecithin 12 | - | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | ||
| Methionine 13 | - | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | ||
| Lysine 14 | - | 1.20 | 1.20 | 1.20 | 1.20 | 1.20 | ||
| Betaine 15 | - | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | ||
| Choline chloride 16 | - | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | ||
| Digestive system improver 17 | - | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | ||
| Vitamins and minerals premix 18 | - | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | ||
| Vitamin C 19 | - | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | ||
| Guar gum 20 | - | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | ||
| Proximate composition (% dw) | ||||||||
| Moisture | 7.78 | 6.49 | 6.16 | 6.18 | 7.29 | 6.50 | ||
| Ash | 2.92 | 8.07 | 7.68 | 7.37 | 8.03 | 8.17 | ||
| Protein | 43.77 | 54.03 | 53.81 | 53.68 | 52.95 | 52.73 | ||
| Lipid | 35.05 | 15.59 | 15.46 | 16.51 | 16.16 | 17.11 | ||
TM: Tenebrio molitor; CTRL: control; FM5 and FM10: 5% and 10% of marine animal ingredients replaced by TM meal; PP10 and PP15: 10 and 15% of plant ingredients replaced by TM meal. 1 69.4% crude protein, 12.3% crude lipid (Norsildemel, Bergen, Norway); 2, 3, 4 purchased from Bacarel (UK). CPSP90 is enzymatically pre-digested fish meal; 5 78% crude protein (Lorca Nutrición Animal SA, Murcia, Spain; 6 50% crude protein, 8% crude lipid (LorcaNutrition, Spain); 7 85% crude protein, 1.5% crude lipid (Emilio Peña SA, Spain); 8 Local provider (Almería, Spain); 9 Beetle Genius S.L. (Brussels, Belgium); 10 AF117DHA (Afamsa, Spain); 11 Soybean oil (Aceites el Niño, Spain); 12 P700IP (Lecico, DE); 13, 14, 15, 16 Lorca Nutrición Animal SA (Murcia, Spain); 17 LBGutHealth, LifeBioencapsulation S.L. (Almería, Spain); 18 Lifebioencapsulation SL (Almería, Spain). Vitamins (mg kg-1): vitamin A (retinyl acetate), 2,000,000 UI; vitamin D3 (DL-cholecalciferol), 200,000 UI; vitamin E (Lutavit E50), 10,000 mg; vitamin K3 (menadione sodium bisulphite), 2,500 mg; vitamin B1(thiamine hydrochloride), 3,000 mg; vitamin B2 (riboflavin), 3,000 mg; calcium pantothenate, 10,000 mg; nicotinic acid, 20,000 mg; vitamin B6 (pyridoxine hydrochloride), 2,000 mg; vitamin B9 (folic acid), 1,500 mg; vitamin B12 (cyanocobalamin), 10 mg vitamin H (biotin), 300 mg; inositol, 50,000 mg; betaine (Betafin S1), 50,000 mg. Minerals (mg kg-1): Co (cobalt carbonate), 65 mg; Cu (cupric sulphate), 900 mg; Fe (iron sulphate), 600 mg; I (potassium iodide), 50 mg; Mn (manganese oxide), 960 mg; Se (sodium selenite), 1 mg; Zn (zinc sulphate) 750 mg; Ca (calcium carbonate), 18.6%; (186,000 mg); KCl, 2.41%; (24,100 mg); NaCl, 4.0% (40,000 mg); 19 TECNOVIT, Spain; 20 EPSA, Spain.
Table 2.
Lipid classes (% dw) and FA (% total FA) composition of Senegalese sole experimental diets (CTRL, FM5, FM10, PP10, PP15) and T. molitor (TM) meal.
Table 2.
Lipid classes (% dw) and FA (% total FA) composition of Senegalese sole experimental diets (CTRL, FM5, FM10, PP10, PP15) and T. molitor (TM) meal.
| Diets | ||||||||
| TM | CTRL | FM5 | FM10 | PP10 | PP15 | |||
| Lipid classes (%TL) | ||||||||
| Lyso-phosphatidylcholine | 0.29 | 0.84 | 1.12 | 1.08 | 1.25 | 1.20 | ||
| Sphingomyelin | 0.41 | 0.43 | 0.26 | 0.50 | 0.39 | 0.65 | ||
| Phosphatidylcholine | 1.94 | 4.97 | 5.71 | 5.26 | 5.44 | 5.13 | ||
| Phosphatidylserine | 0.36 | 1.52 | 2.57 | 2.51 | 2.07 | 1.91 | ||
| Phosphatidylinositol | 0.38 | 1.93 | 1.35 | 1.62 | 1.49 | 1.83 | ||
| Phosphatidylethanolamine | 2.54 | 2.11 | 2.90 | 2.51 | 2.19 | 2.57 | ||
| Diacylglycerol | 3.01 | 3.19 | 3.23 | 3.85 | 3.48 | 3.11 | ||
| Sterols | 7.36 | 9.65 | 10.94 | 10.20 | 10.37 | 9.70 | ||
| Free fatty acids | 34.58 | 13.53 | 17.29 | 19.16 | 19.12 | 20.09 | ||
| Triacylglycerol | 43.45 | 47.08 | 42.77 | 41.90 | 43.24 | 42.59 | ||
| Sterol esters | 4.10 | 6.22 | 6.19 | 5.19 | 5.65 | 5.61 | ||
| Fatty acids (% TFA) | ||||||||
| 14:0 | 4.18 | 1.69 | 1.92 | 2.16 | 2.31 | 2.63 | ||
| 16:0 | 17.98 | 17.91 | 18.40 | 19.13 | 19.33 | 19.48 | ||
| 18:0 | 2.55 | 5.19 | 4.94 | 4.81 | 4.73 | 4.45 | ||
| Total Saturated FA | 24.72 | 26.19 | 26.45 | 27.25 | 27.44 | 27.56 | ||
| 16:1n-7 | 2.82 | 3.41 | 3.56 | 3.74 | 3.93 | 3.89 | ||
| 18:1n-9 | 51.75 | 17.96 | 22.46 | 26.90 | 26.64 | 30.44 | ||
| 18:1n-7 | 0.10 | 2.37 | 2.08 | 1.87 | 1.90 | 1.66 | ||
| 20:1n-9 | 0.10 | 1.52 | 1.39 | 1.29 | 1.37 | 1.18 | ||
| 22:1n-11 | nd | 0.96 | 0.89 | 0.79 | 0.93 | 0.80 | ||
| Total Monounsaturated FA | 55.86 | 27.39 | 31.75 | 36.02 | 36.33 | 39.48 | ||
| 18:2n-6 | 18.00 | 19.54 | 16.65 | 13.89 | 12.13 | 12.89 | ||
| 20:4n-6 | 0.00 | 1.10 | 1.06 | 1.01 | 1.00 | 0.83 | ||
| 22:5n-6 | nd | 0.79 | 0.76 | 0.73 | 0.74 | 0.59 | ||
| Total n-6 Polyunsaturated FA | 18.00 | 22.05 | 19.13 | 16.25 | 14.41 | 14.82 | ||
| 18:3n-3 | 0.29 | 2.42 | 1.85 | 1.30 | 1.13 | 0.99 | ||
| 18:4n-3 | nd | 0.77 | 0.71 | 0.63 | 0.71 | 0.61 | ||
| 20:5n-3 | nd | 5.73 | 5.38 | 4.81 | 5.24 | 4.50 | ||
| 22:5n-3 | nd | 0.91 | 0.99 | 0.94 | 0.98 | 0.82 | ||
| 22:6n-3 | nd | 13.05 | 12.37 | 11.62 | 12.37 | 10.01 | ||
| Total n-3 Polyunsaturated FA | 0.29 | 23.61 | 22.00 | 19.94 | 21.12 | 17.49 | ||
TM: Tenebrio molitor; CTRL: control; FM5 and FM10: 5% and 10% of marine animal ingredients replaced by TM meal; PP10 and PP15: 10 and 15% of plant ingredients replaced by TM meal. nd: not detected.
Table 3.
Growth performance and nutrient utilization parameters of Senegalese sole fed with the experimental diets during the 98-day feeding trial.
Table 3.
Growth performance and nutrient utilization parameters of Senegalese sole fed with the experimental diets during the 98-day feeding trial.
| CTRL | FM5 | FM10 | PP10 | PP15 | p-value | |
| Initial body weight (IBW, g) | 219.2 ± 5.3 | 213.7 ± 4.7 | 215.1 ± 5.2 | 214.2 ± 5.3 | 210.8 ± 4.6 | 0.979 |
| Final body weight (FBW, g) | 288.6 ± 6.5 | 284.5 ± 5.8 | 288.6 ± 6.1 | 283.8 ± 6.2 | 288.3 ± 5.5 | 0.887 |
| Specific Growth Rate (SGR, % d-1) | 0.30 ± 0.01b | 0.32 ± 0.01ab | 0.33 ± 0.01ab | 0.31 ± 0.01b | 0.35 ± 0.01a | 0.012 |
| Feed Conversion Ratio (FCR) | 1.76 ± 0.20 | 1.58 ± 0.14 | 1.61 ± 0.02 | 1.71 ± 0.17 | 1.42 ± 0.23 | 0.426 |
| Protein efficiency ratio (PER) | 1.00 ± 0.08 | 1.11 ± 0.08 | 1.09 ± 0.08 | 1.04 ± 0.08 | 1.26 ± 0.08 | 0.299 |
Dietary treatment codes are CTRL: control; FM5 and FM10: 5% and 10% of marine-derived ingredients replaced by TM meal; PP10 and PP15: 10 and 15 % of plant ingredients replaced by TM meal. Values are mean ± SEM. Values in the same row with different superscript letter indicate significant differences among dietary treatments (p < 0.05).
Table 4.
Fatty acid composition (%TFA) in liver of Senegalese sole fed with experimental diets (mean ± SEM, n=6).
Table 4.
Fatty acid composition (%TFA) in liver of Senegalese sole fed with experimental diets (mean ± SEM, n=6).
| CTRL | FM5 | FM10 | PP10 | PP15 | P(diet) | |||||
| 14:0 | 3.14±0.21 | 3.46±0.19 | 3.42±0.07 | 3.40±0.16 | 3.69±0.09 | 0.122 | ||||
| 15:0 | 0.45±0.018 | 0.38±0.026 | 0.41±0.03 | 0.46±0.03 | 0.40±0.01 | 0.092 | ||||
| 16:0 | 17.29±0.29 | 17.44±0.32 | 16.88±0.31 | 16.69±0.30 | 16.89±0.15 | 0.351 | ||||
| 18:0 | 3.93±0.38 | 3.92±0.27 | 3.72±0.37 | 3.39±0.24 | 3.25±0.19 | 0.347 | ||||
| 20:0 | 0.20±0.02 | 0.19±0.01 | 0.19±0.01 | 0.19±0.01 | 0.19±0.00 | 0.974 | ||||
| 22:0 | 0.18±0.012 | 0.16±0.01 | 0.14±0.01 | 0.14±0.01 | 0.16±0.01 | 0.095 | ||||
| 24:0 | 0.10±0.02 | 0.15±0.039 | 0.11±0.02 | 0.13±0.02 | 0.11±0.03 | 0.640 | ||||
| Total Saturated FA | 25.29±0.75 | 25.70±0.46 | 24.89±0.56 | 24.41±0.55 | 24.69±0.32 | 0.396 | ||||
| 16:1n-9 | 0.78±0.03b | 0.90±0.05ab | 1.02±0.07a | 1.09±0.05a | 1.05±0.03a | 0.002 | ||||
| 16:1n-7 | 4.94±0.18 | 5.54±0.33 | 5.23±0.12 | 5.33±0.17 | 5.48±0.11 | 0.306 | ||||
| 18:1n-9 | 28.09±0.37c | 31.44±1.11b | 32.28±0.54ab | 31.18±0.80b | 34.24±0.59a | 0.000 | ||||
| 18:1n-7 | 3.34±0.06 | 3.19±0.09 | 3.11±0.05 | 3.19±0.08 | 3.01±0.10 | 0.103 | ||||
| 20:1n-11 | 0.15±0.01 | 0.13±0.01 | 0.15±0.00 | 0.15±0.00 | 0.15±0.01 | 0.069 | ||||
| 20:1n-9 | 1.38±0.06 | 1.44±0.05 | 1.44±0.05 | 1.435±0.07 | 1.68±0.19 | 0.172 | ||||
| 20:1n-7 | 0.20±0.01 | 0.20±0.00 | 0.19±0.01 | 0.19±0.01 | 0.18±0.01 | 0.566 | ||||
| 22:1n-11 | 0.51±0.042 | 0.46±0.03 | 0.49±0.03 | 0.54±0.02 | 0.54±0.04 | 0.391 | ||||
| 22:1n-9cis | 0.27±0.015 | 0.27±0.02 | 0.27±0.01 | 0.28±0.01 | 0.27±0.02 | 0.986 | ||||
| 24:1n-9 | 0.31±0.061 | 0.33±0.03 | 0.34±0.03 | 0.35±0.02 | 0.26±0.06 | 0.658 | ||||
| Total Monounsaturated FA | 40.00±0.56c | 43.92±1.37ab | 44.52±0.48ab | 43.74±0.87b | 46.85±0.47a | 0.000 | ||||
| 18:2n-6 | 15.47±0.57a | 13.16±0.76b | 12.57±0.73b | 12.26±0.66b | 12.12±0.13b | 0.001 | ||||
| 18:3n-6 | 0.10±0.01 | 0.10±0.01 | 0.08±0.00 | 0.10±0.01 | 0.09±0.00 | 0.252 | ||||
| 20:2n-6 | 1.18±0.065ab | 1.21±0.06a | 1.03±0.03ab | 0.94±0.04b | 0.94±0.02b | 0.000 | ||||
| 20:3n-6 | 0.16±0.007 | 0.15±0.01 | 0.16±0.01 | 0.16±0.01 | 0.14±0.01 | 0.480 | ||||
| 20:4n-6 | 0.65±0.07 | 0.66±0.09 | 0.62±0.06 | 0.76±0.10 | 0.61±0.05 | 0.698 | ||||
| 22:4n-6 | 0.27±0.014 | 0.26±0.03 | 0.25±0.01 | 0.26±0.02 | 0.21±0.01 | 0.269 | ||||
| 22:5n-6 | 0.74±0.03 | 0.70±0.07 | 0.71±0.02 | 0.74±0.05 | 0.61±0.02 | 0.226 | ||||
| Total n-6 Polyunsaturated FA | 18.57±0.57a | 16.24±1.00ab | 15.42±0.81b | 15.22±0.72b | 14.71±0.14b | 0.001 | ||||
| 18:3n-3 | 1.39±0.06a | 1.09±0.05b | 1.03±0.07b | 1.01±0.08b | 0.91±0.05b | 0.000 | ||||
| 18:4n-3 | 0.24±0.02ab | 0.18±0.01b | 0.22±0.02ab | 0.25±0.02a | 0.21±0.01ab | 0.035 | ||||
| 20:3n-3 | 0.56±0.021a | 0.55±0.02a | 0.45±0.01b | 0.41±0.02b | 0.39±0.02b | 0.000 | ||||
| 20:4n-3 | 0.35±0.02ab | 0.29±0.01b | 0.33±0.02ab | 0.36±0.03a | 0.31±0.01ab | 0.034 | ||||
| 20:5n-3 | 0.36±0.05ab | 0.25±0.03b | 0.30±0.03ab | 0.40±0.06a | 0.31±0.02ab | 0.024 | ||||
| 22:5n-3 | 2.34±0.31 | 1.83±0.15 | 2.24±0.06 | 2.34±0.32 | 1.96±0.12 | 0.260 | ||||
| 22:6n-3 | 10.13±0.41 | 9.13±1.00 | 9.73±0.29 | 11.05±0.58 | 8.91±0.33 | 0.140 | ||||
| Total n-3 Polyunsaturated FA | 15.41±0.80 | 13.33±1.16 | 14.32±0.28 | 15.83±0.89 | 13.00±0.48 | 0.061 | ||||
| n-3PUFA/n-6PUFA | 0.83±0.04 | 0.82±0.06 | 0.94±0.05 | 1.05±0.07 | 0.88±0.03 | 0.048 |
Table 5.
Fatty acid composition (%TFA) in muscle of Senegalese sole fed with experimental diets (mean ± SEM, n=6).
Table 5.
Fatty acid composition (%TFA) in muscle of Senegalese sole fed with experimental diets (mean ± SEM, n=6).
| CTRL | FM5 | FM10 | PP10 | PP15 | P(diet) | |||||
| 14:0 | 2.30±0.16 | 2.36±0.16 | 1.86±0.16 | 1.99±0.16 | 2.16±0.16 | 0.190 | ||||
| 15:0 | 0.48±0.0a | 0.47±0.02ab | 0.40±0.02b | 0.40±0.02ab | 0.40±0.02b | 0.009 | ||||
| 16:0 | 17.79±0.19 | 18.11±0.19 | 17.97±0.19 | 18.08±0.19 | 18.43±0.19 | 0.223 | ||||
| 18:0 | 3.54±0.12b | 3.63±0.12ab | 4.10±0.12a | 3.82±0.12ab | 3.90±0.12ab | 0.035 | ||||
| 20:0 | 0.25±0.01 | 0.25±0.01 | 0.25±0.01 | 0.26±0.01 | 0.26±0.01 | 0.457 | ||||
| 22:0 | 0.19±0.01b | 0.19±0.01ab | 0.18±0.01ab | 0.17±0.01b | 0.18±0.01ab | 0.035 | ||||
| 24:0 | 0.08±0.02 | 0.10±0.02 | 0.12±0.02 | 0.10±0.02 | 0.07±0.02 | 0.657 | ||||
| Total Saturated FA | 24.65±0.33 | 25.11±0.33 | 24.88±0.33 | 24.83±0.33 | 25.39±0.33 | 0.568 | ||||
| 16:1n-9 | 0.47±0.01b | 0.54±0.01a | 0.55±0.01a | 0.56±0.01a | 0.58±0.01a | 0.000 | ||||
| 16:1n-7 | 4.79±0.15 | 4.76±0.15 | 4.18±0.15 | 4.40±0.15 | 4.35±0.15 | 0.031 | ||||
| 18:1n-9 | 25.33±0.36c | 26.88±0.36b | 27.29±0.36ab | 27.31±0.36ab | 28.62±0.36a | 0.000 | ||||
| 18:1n-7 | 2.80±0.05a | 2.70±0.05ab | 2.56±0.05bc | 2.58±0.05bc | 2.43±0.05c | 0.000 | ||||
| 20:1n-11 | 0.21±0.00a | 0.20±0.00ab | 0.19±0.00ab | 0.201±0.005ab | 0.19±0.00b | 0.023 | ||||
| 20:1n-9 | 1.51±0.03 | 1.45±0.03 | 1.40±0.03 | 1.43±0.03 | 1.36±0.03 | 0.084 | ||||
| 20:1n-7 | 0.16±0.01 | 0.16±0.01 | 0.15±0.01 | 0.15±0.01 | 0.15±0.01 | 0.860 | ||||
| 22:1n-11 | 0.76±0.02a | 0.71±0.02ab | 0.62±0.02b | 0.69±0.02ab | 0.64±0.022b | 0.002 | ||||
| 22:1n-9cis | 0.29±0.01 | 0.29±0.01 | 0.26±0.01 | 0.28±0.01 | 0.27±0.01 | 0.157 | ||||
| 24:1n-9 | 0.40±0.05 | 0.22±0.05 | 0.41±0.05 | 0.43±0.05 | 0.41±0.05 | 0.046 | ||||
| Total Monounsaturated FA | 36.72±0.55 | 37.89±0.55 | 37.61±0.55 | 38.05±0.55 | 39.01±0.55 | 0.100 | ||||
| 18:2n-6 | 13.50±0.26a | 12.86±0.26a | 11.69±0.26b | 11.15±0.26b | 11.25±0.26b | 0.000 | ||||
| 18:3n-6 | 0.14±0.01 | 0.14±0.01 | 0.14±0.01 | 0.13±0.01 | 0.12±0.01 | 0.117 | ||||
| 20:2n-6 | 0.66±0.01abc | 0.68±0.01a | 0.67±0.01ab | 0.62±0.01bc | 0.62±0.01c | 0.001 | ||||
| 20:3n-6 | 0.17±0.01 | 0.16±0.01 | 0.17±0.01 | 0.18±0.01 | 0.16±0.01 | 0.341 | ||||
| 20:4n-6 | 0.82±0.04b | 0.83±0.04b | 1.01±0.04a | 0.98±0.04ab | 0.96±0.04ab | 0.008 | ||||
| 22:4n-6 | 0.28±0.01ab | 0.29±0.01b | 0.30±0.01a | 0.30±0.01ab | 0.27±0.01b | 0.009 | ||||
| 22:5n-6 | 0.61±0.02 | 0.63±0.02 | 0.70±0.02 | 0.69±0.02 | 0.66±0.02 | 0.061 | ||||
| Total n-6 Polyunsaturated FA | 16.19±0.25a | 15.60±0.25b | 14.68±0.25c | 14.05±0.25c | 14.03±0.25c | 0.000 | ||||
| 18:3n-3 | 1.85±0.05a | 1.60±0.05b | 1.35±0.05c | 1.32±0.05c | 1.20±0.05c | 0.000 | ||||
| 18:4n-3 | 0.64±0.03a | 0.57±0.03ab | 0.49±0.03b | 0.52±0.03b | 0.46±0.03b | 0.001 | ||||
| 20:3n-3 | 0.36±0.01a | 0.35±0.01a | 0.32±0.01ab | 0.29±0.01b | 0.28±0.01b | 0.000 | ||||
| 20:4n-3 | 0.44±0.01a | 0.41±0.01ab | 0.39±0.01b | 0.39±0.01b | 0.37±0.01b | 0.001 | ||||
| 20:5n-3 | 1.69±0.05a | 1.43±0.05ab | 1.46±0.05b | 1.57±0.05b | 1.40±0.05b | 0.001 | ||||
| 22:5n-3 | 3.76±0.10a | 3.50±0.10ab | 3.54±0.10ab | 3.54±0.10ab | 3.28±0.10b | 0.043 | ||||
| 22:6n-3 | 13.04±0.45ab | 12.93±0.45b | 14.77±0.45ab | 14.92±0.45a | 14.05±0.45ab | 0.012 | ||||
| Total n-3 Polyunsaturated FA | 21.79±0.44 | 20.80±0.44 | 22.32±0.44 | 22.56±0.44 | 21.07±0.44 | 0.045 | ||||
| n-3PUFA/n-6PUFA | 1.35±0.04bc | 1.33±0.040c | 1.52±0.04a | 1.61±0.04a | 1.50±0.04ab | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



