
Submitted:
04 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
Crinoids (Echinodermata) exhibit unique characteristics that facilitate a wide range of symbiotic relationships with diverse organisms. Nonetheless, the comprehension of their interactions with microscopic copepod crustaceans is still in a nascent and fragmented state. Our analysis identifies 166 instances involving 35 copepod species representing in 6 families in association with 33 species of the Comatulida. The majority of these associations have been singularly reported, with their distribution covering five out of the 12 World Ocean ecoregions. A notable concentration of diversity is observed in the Central and Western Indo-Pacific regions, while documentation of Atlantic copepod-crinoid associations is markedly limited. Copepods are predominantly found in ectosymbiotic relationships, with a lesser incidence of endosymbiotic interactions. Among these, the genus Collocheres and Pseudanthessius emerge as particularly prominent, and the Comasteridae family is distinguished by its diverse copepod associations. While certain copepod families exhibit a specialization towards crinoids, others demonstrate specificity at the species level. The current scope of knowledge encompasses merely 5% of the potential crinoid host diversity, underscoring the critical need for more extensive research in this area.
Keywords:
Copepod Associations
; Symbiotic Relationships
; Crinoids
; Comatulida
; Marine Biodiversity
; Marine Ecology
; Marine Invertebrates
; Host-Symbiont Interactions
; Marine Parasitology
1. Introduction
The echinoderm class Crinoidea represents a diverse and enduring clade with a fossil record stretching back nearly half a billion years (Hess et al., 1999; Wright et al., 2017). Its modern-day diversity is predominantly found within the order Comatulida, which is documented to comprise a wealth of 671 distinct species (Pawson, 2007; Messing et al. 2023). These crinoids, characterized by their limited locomotive capabilities, rudimentary self-cleaning mechanisms, and the absence of saponin secretions, have provided a structural foundation for a diverse of vertebrate and invertebrate organisms (Burnell, ApSimon, 1983; Britayev, Mekhova, 2011). While direct predation on crinoids remains uncommon, their anatomical design—comprising mobile arms, pinnules, and cirri—and their unique filter-feeding method, wherein alimentary particles travel conspicuously along ambulacral grooves, serve as a habitat conducive to a plethora of symbionts (Fabricius, Dale, 1993; Deheyn, 2006; Britayev, Mekhova, 2011). The symbiotic taxa often found in association with crinoids range across gastropods, polychaetas, myzostomes, decapod and copepod crustaceans, ophiuroids, and fish (Clark, 1931; Fishelson, 1974; Humes, 2000; Mekhova, Britayev 2012).
Historical examinations within marine biology underscore a sustained interest in the relationships between crinoids and their symbiotic partners. The investigations have been conducted in various coastal ecosystems around the world, with notable studies including those conducted in the Bay of Bengal (Rao, Sowbhagyavathi, 1972), the Red Sea (Fishelson, 1974), the Marshall Islands (Zmarzly, 1984), the Maldives Archipelago (Tchesunov et al., 1989), Hong Kong (Morton and Mladenov, 1992), the Great Barrier Reef (Fabricius and Dale, 1993), Taiwan (Huang et al., 2005), New Guinea (Deheyn et al., 2006), South Africa (Hempson and Griffiths, 2008), Vietnam (Britayev and Mekhova, 2011; Britayev et al., 2016), and North Sulawesi (Virgili et al., 2020). Comprehensive investigations pertaining to this subject have consistently emphasized the prevalence of specialized fauna engaged in symbiotic associations with crinoids. However, it is imperative to exercise prudence in the interpretation of these findings, given the inherent discrepancies in the accuracy of species identification. Furthermore, a conspicuous gap in data persists regarding the diverse insufficiently studied microscopic symbionts (such as myzostomid polychaetes and copepod crustaceans) inhabiting crinoids (Humes, 1987; Summers et al. 2014). These relatively obscure organisms, despite their diminutive size, potentially exert a notable influence on the broader ecosystem dynamics intertwined with their host crinoids.
Microscopic copepods, a type of crustacean, play a significant, though still insufficiently explored, role in a wide range of ecological interactions within marine ecosystems (Humes, 1994; Ho, 2001; Bron et al., 2011; Bernot et al., 2021). Their establishment of symbiotic relationships with various echinoderm species across diverse marine environments underscores their remarkable ecological adaptability and highlights the intricate network of biotic interactions in aquatic ecosystems. Copepods from various families have been observed residing in association with echinoderm hosts representing Crinoidea (feather stars), Asteroidea (sea stars), Echinoidea (sea urchins), Holothuroidea (sea cucumbers), and Ophiuroidea (brittle stars) (Humes, 1986; Ivanenko et al., 2001; Boxshall and Halsey, 2004; Kim et al., 2007, Mahatma et al., 2008; Venmathi Maran et al. 2017; Yeom et al., 2018). This diverse range of symbiotic relationships underscores the pivotal role of copepods in marine ecosystems and provides valuable insights into the evolutionary intricacies governing these associations (Boxshall and Halsey, 2004; Bernot et al., 2021). The investigation of these interactions not only reveals the ecological significance of copepods but also contributes to a deeper understanding of the evolutionary mechanisms that underlie symbiosis within marine ecosystems.
This study is a component of a broader project aimed at elucidating patterns and assessing the depth of understanding pertaining to copepod symbionts found in various invertebrates, with a particular focus on echinoderms, sponges, and corals (Ivanenko et al. 2018; Korzhavina et al. 2019, 2021, 2023). Through an exhaustive analysis and synthesis, we aspire to offer a holistic view of these relationships, focusing on their ecological, evolutionary, and taxonomical dimensions, thereby enhancing our comprehensive understanding of marine symbiotic systems and the pivotal roles copepods play within them.
2. Materials and Methods
We have developed a comprehensive Microsoft Access database to meticulously analyze the symbiotic interactions between copepods and crinoids. This database comprises four intricately connected tables: 'Hosts,' 'Symbionts,' 'Sites,' and 'Publications,' which collectively merge into a comprehensive 'Literature Records' table (Table 1, Table A1, Table S1). Adhering to the standards set by the World Register of Marine Species (WoRMS 2022), this database provides a thorough record of the taxonomic classifications of hosts and symbionts. It also encompasses a broad spectrum of data, including detailed information on symbiotic relationships, geographical locations, sampling depths, and timestamps, as further elucidated in Table A2. Spatial data management involved extracting specific coordinates for each sampling location from the original literature and subsequently georeferencing them. All these data have been meticulously incorporated into the dataset entries, adhering to the Darwin Core standards (Wieczorek et al., 2012). To ensure consistency and accuracy in taxonomic nomenclature, we employed the 'Taxon Match' tool from WoRMS, a crucial step, especially given the evolving nomenclature for crinoid hosts. The classification of oceanic ecoregions adheres to the methodology advocated by Spalding et al. (2007). We do not endorse the recently proposed taxonomic status changes for the order Poecilostomatoida, as we believe they warrant further investigation (Mikhailov and Ivanenko, 2019, 2021). Visualization of these geographic data points was achieved using digital mapping platforms, including Google Maps and RStudio Version 1.2. To visualize and generate plots, we employed RStudio version 1.2.5001, harnessing the capabilities of various packages such as tidyverse [121], dplyr [122], ggplot2 [123], ggExtra [124], ggpubr [125], gridExtra [126], magrittr [127], maps [128], stringr [129], and RColorBrewer [130]. Additionally, all graphical representations were crafted using Adobe Photoshop CC.
3. Results and discussion
3.1. The history of research
Over the past century, the study of copepod symbionts associated with crinoids has culminated in the publication of 24 scientific articles, documenting a total of 166 symbiotic interactions between copepods and crinoids, as indicated in Figure 1. The research trajectory concerning crinoids can be divided into four significant stages, as identified by Wright et al. (2017). Notably, this trajectory demonstrates a notably greater taxonomic effort directed towards crinoids compared to copepod symbionts. This discrepancy can be attributed to the inherent challenges associated with collecting microscopic symbionts residing within galls, digestive systems, or on the surfaces of crinoids.
With the advent of methods for sampling shallow-water material using SCUBA diving, it became possible to gather copepods that are only weakly associated with shallow-water crinoids. However, over the past decade, there has been a noticeable decline in research activity. Few works have focused on morphological descriptions, with some providing brief comments on zoogeographical aspects and relationships between copepods and their hosts. This trend underscores the existence of numerous unexplored facets in the symbiotic relationship between copepods and crinoids, particularly concerning the nature of symbiosis and its implications for both partners.
3.2. Sampling methods and challenges
In the study of copepod-crinoid symbiosis, the prevailing methodology for copepod identification involves the use of a 5% ethanol solution to wash the crinoid hosts. This approach, although intricate and capable of recovering a significant diversity of microsymbionts, presents challenges when conducting quantitative assessments. The application of this methodology, which shapes our comprehension of copepod-host relationships, is applicable to the majority (155) of observations related to copepods on crinoids. Dissection has been relatively infrequent, limited to 11 instances (Table A1, Table S1).
The choice of methodology is inherently linked to the type of symbiotic interaction identified: specimens identified via washing are consistently categorized as ectosymbiotic, with descriptions typically lacking specific localization details on the host. Conversely, copepods identified through dissection are invariably classified as endosymbiotic, with precise intrahost localization described, such as within the intestinal tract (Giesbrecht in 1900 and 1950; Changeux and Delamare Deboutteville in 1956; Stock in 1959 and 1966), within galls (Ohtsuka, Kitazawa, Boxshall, 2010), or the coelom (Changeux, Delamare Deboutteville, 1956). The significant methodological influence on the types of copepods detected, as elucidated by Humes in 2000, suggests that the spectrum of endosymbiotic copepods associated with crinoids remains incompletely explored. Consequently, further research is imperative to ascertain the precise localization of the majority of ectosymbionts.
The research into copepod-crinoid symbiosis faces significant challenges, particularly the complexity of collecting loosely associated fauna from deep-sea specimens. This complexity is compounded by the current state of knowledge about microscopic copepods residing in or on crinoids, which is primarily characterized by a lack of an integrative approach, especially in the application of molecular methodologies. Additionally, much of the existing data is limited to faunistic or exploratory methodologies, which, while inevitable in the initial stages of researching any taxonomic group, constrain the breadth and depth of understanding. The absence of a comprehensive and multidisciplinary research approach thus represents a significant impediment to advancing the understanding of these intricate symbiotic associations.
3.3. Diversity and taxonomy of symbiotic copepods
Literature analysis revealed 166 instances involving 35 copepod species representing 6 families in association with 33 species of the Comatulida. The data analysis has unveiled intricate symbiotic associations among three distinct orders of copepods — Cyclopoida, Poecilostomatoida, and Siphonostomatoida — and the Comatulida order of crinoids. Consequently, this taxonomic rearrangement necessitates additional scrutiny and detailed research to substantiate its validity. This symbiotic interaction constitutes more than 5% of the known diversity within the Comatulida order.
The study indicates that the Poecilostomatoida and Siphonostomatoida orders display parallel trends in their frequency of occurrence and the spectrum of crinoid taxa they are associated with, as elaborated in Table 2. Significantly, the Poecilostomatoida order is characterized by a broad spectrum of families and genera involved with crinoids, suggesting more elaborate symbiotic connections with this marine class. The predominance of Poecilostomatoida, evidenced by 82 instances in the study, points towards an in-depth exploration of their symbiotic links in comparison to other copepod orders. In contrast, the Siphonostomatoida order, with a considerable 19 species linked to crinoids, demonstrates a heightened level of specialization within this group.
In the Cyclopoida order, encompassing 95 families, only the representatives from the Enterognathidae family have been identified in crinoids, comprising seven known copepod species. Of these, four species are associated with seven different further star species (Table 2). The average frequency of scientific mentions per copepod species stands at 2.75 (SE 0.85), reflecting variation in observations across different species. Half of the species in this family exhibit specialization for a single host, indicative of a selective adaptation towards specialization.
The Poecilostomatoida order is predominantly represented by the Pseudanthessiidae, with a high specialization, each of the three identified copepod species is exclusively linked to a singular further star species, demonstrating strict host specificity among these copepods. The Pseudanthessiidae family is noted for a considerable average of scientific mentions per species (13.57), with a standard error (SE) of 5.03, suggesting an extensive host range. The mean number of hosts per copepod species is three (SE 0.93), with approximately 28.57% of copepod species associated with a single further star species, indicative of a moderate degree of specialization in comparison to other families.
The Siphonostomatoida order has only one family associated with further stars (Table ). The Asterocheridae family, uniquely identified within Siphonostomatoida, displays an average research mention frequency of 3.8 per species (SD ± 1.17), emphasizing the variability in species encounters. A substantial proportion of copepods (63.16%) within this order are associated solely with a single further star species, suggesting a trend towards species-specific symbiosis.
Despite the rich species diversity within the Asterocheridae, Rhynchomolgidae, and Pseudanthessiidae families, only a small fraction of these species is found in association with crinoids – 7% of 271 species, 2.5% of 270, and 5% of 61, respectively. This observation implies a selective nature of the relationships between copepods and crinoids, with Asterocheridae and Rhynchomolgidae exhibiting a high level of host specialization, whereas Pseudanthessiidae demonstrates a broader variation in associations.
3.4. Specialization in Copepod-Crinoid Symbiosis
The examination of morphological adaptations in copepods, particularly those engaged in endosymbiotic relationships with crinoids, reveals significant deviations from typical crustacean morphology, as depicted in Figure 2. This phenomenon is notably apparent in the Cyclopoid family Enterognathidae, which predominantly associates with crinoids and is represented by genera such as Enterognathus and Parenterognathus. Descriptions of different endosymbiotic or gall inducing show that endosymbiosis leads to considerable morphological changes in the copepods. In the case of Enterognathidae, these alterations include a swollen, vermiform body structure, reduced segmentation and sclerotization, obscured prosome and urosome demarcations, and a potential diminution or complete absence of antennae and maxillipeds. Such changes are markedly more pronounced in Enterognathidae compared to other crinoid-dwelling copepods and exhibit a less extreme form than those observed in the obligate symbionts of octocorals from the Lamippidae family (Korzhavina et al. 2021).
The order Comatulida encompasses a diverse array of crinoid hosts for copepods, including eight families, 21 genera, and 33 species, as detailed in Table 3 and Supplement Table 2. The Comasteridae family emerges as a prominent symbiotic partner for copepods. Despite the Comatulidae family comprising 102 species, only 16% serve as hosts for copepods. Notably, this family is linked to half of all recorded copepod findings (81 out of 163) and half of the copepod species (20 out of 40) symbiotic with crinoids, as outlined in Tables 3 and Figure 3. The unique ambulacral architecture of the Comasteridae may provide specialized ecological niches conducive to symbiosis. Conversely, the Antedonidae family, despite its diversity (151 species), exhibits a minimal number of confirmed symbiotic relationships with copepods. These observations underscore the complexity and selectivity inherent in the symbiotic associations between copepods and crinoids, hinting at the potential influence of ecological and evolutionary factors that warrant further investigation. An alternative hypothesis might consider the uneven distribution of research efforts across different feather star species.
The degree of host specialization exhibited by various copepod species in their interactions with crinoids displays a spectrum ranging from highly specialized to more generalized associations, indicative of diverse evolutionary trajectories. Notably, species-specific symbionts constitute 61% (comprising 20 species), as opposed to 39% (encompassing 13 species) associated with a variety of hosts. This pattern highlights the significance of specialized host-symbiont relationships in the evolutionary ecology of copepods (Figure 4).
The range of host interactions among copepod species exhibits considerable heterogeneity. For example, Pseudanthessius major and P. minor are distinguished by their extensive host interactions, involving multiple host species (eight and four, respectively) and genera (seven and four, respectively). Similarly, Collocheres uncinatus associates with three distinct host families: Colobometridae, Comatulidae, and Himerometridae, presenting a broad spectrum of ecological associations.
A reduction in the number of host species typically leads to the evolution of specific adaptive traits. This is exemplified by P. angularis, P. comanthi, and P. madrasensis, characterized by the development of prominent egg sacs in females, a feature commonly observed in symbiotic copepod species. This trait distinguishes them from species such as P. major and P. minor. Additionally, Enterognathus comatulae and E. lateripes display significant morphological adaptations, including a vermiform body structure with inflated, rounded body segments, and fringed swimming appendages.
The species-specific Parenterognathus troglodytes exhibits a more pronounced degree of body modification compared to its Enterognathus counterparts. Distinct adaptations are also evident in P. planus and P. rostellatus, including broader and rounder thoracic segments in the former, and an abundance of long setae on the antennae and urosome in the latter. Kelleria gradata, another species-specific symbiont, demonstrates elongated setae, particularly on the swimming legs, and thoracic segment expansion, a feature also observed in Critomolgus fishelsoni and Doridicola patulus.
However, conducting a comprehensive comparative analysis presents considerable challenges, primarily due to the variability in research depth, taxonomic precision, and accuracy in species identification. Moreover, our understanding of host specificity is significantly constrained by the fragmentary nature of the available data and the absence of a systematic approach to this issue. This limitation is not exclusive to these copepods but also extends to the majority of copepods associated with invertebrates, underscoring a significant gap in our comprehensive understanding of symbiotic relationships in marine ecosystems, as noted by Ivanenko et al. (2018).
3.5. Distribution of Crinoid-Associated Copepods
Copepods engaged in symbiosis with crinoids are distributed across a wide range of ecosystems, extending from tropical to temperate latitudes in both Western and Eastern hemispheres, as depicted in Figure 5. Predominantly, the distribution of observational data is concentrated in the temperate zones of both hemispheres, with a comparatively reduced dataset originating from tropical regions. These symbiotic interactions are documented in five out of the twelve delineated marine ecoregions, according to Spalding et al. (2007), with regions such as the Central and Western Indo-Pacific epitomizing the richness in biodiversity, detailed in Table 4. Conversely, the temperate Northern Pacific and Atlantic regions are characterized by a notably lower diversity of these symbiotic associations.
The paucity of data in certain geographical areas can be attributed to either the absence of copepod populations or to the lack of extensive research in these locales. The distribution pattern of existing data may reflect the habitat preferences inherent to copepod species, as well as highlight areas of specific interest within the research community. This pattern underscores the critical need for enhanced research efforts in under-explored regions to attain a holistic understanding of the global distribution patterns of these marine symbionts.
Several taxa within the copepod community, namely Collocheres prionotus, C. uncinatus, Pseudanthessius madrasensis, and P. major, alongside crinoids such as Capillaster multiradiatus and Stephanometra indica, demonstrate a pan-Indo-Pacific distribution. Biodiversity hotspots for these organisms are identified in regions including Madagascar, Australia, and the Indo-West Pacific archipelagos, with notable biodiversity concentrations in Madagascar's northern area, the Moluccas of Indonesia, and New Caledonia. The heightened diversity observed in regions like Indonesia and Madagascar is likely a result of intensive sampling efforts, suggesting that further research could reveal additional, cryptic species. The Atlantic region, with rare findings like the parasitic Enterognathus comatulae, remains largely uncharted in this context.
The genus Scambicornus is observed in a limited number of localities, in contrast to the more widespread geographical presence of Collocheres and Enterognathus. Species such as Collocheres comanthiphilus, Pseudanthessius major, and Glyptocheres extrusus are of particular interest due to their extensive transmarine distribution. Additionally, the cosmopolitan presence of the scarcely documented Enterognathus genus, spanning from the northeast Atlantic to Japanese waters, presents a compelling case for further investigation. The morphological adaptations observed in these copepods, especially in terms of specialized oral structures and appendages, are indicative of their advanced symbiotic relationships. Yet, these taxa do not exhibit any distinct morphological characteristics that would differentiate them from their congeneric counterparts.
3.6. Bathymetric Distribution
The analysis of depth-related data offers an enhanced understanding of the habitat preferences in symbiotic relationships between copepods and their hosts. The concentration of research efforts within a depth range of 47 meters, contrasted with sporadic findings in deep-sea environments, points to two possible interpretations: a depth-specific specialization in these symbiotic relationships or a research bias towards sampling in more accessible, shallower waters. The discovery of the copepod species Parenterognathus troglodytes in deep-sea habitats underscores the remarkable adaptability of certain copepod taxa to diverse environmental conditions.
A significant correlation is observed between the bathymetric distribution of copepods and the preferred habitats of stalked crinoids, predominantly within the 0–200-meter depth range (Figure 5). This correlation may indicate a potential co-evolutionary relationship between these organisms, shaped by their long-term interactions in shared ecological niches. The depth range preference suggests that environmental factors associated with shallower marine zones play a crucial role in facilitating and maintaining these symbiotic relationships. The presence of copepods in deeper waters, albeit less frequently documented, opens avenues for further exploration into the adaptive capabilities and ecological breadth of these symbiotic copepods.
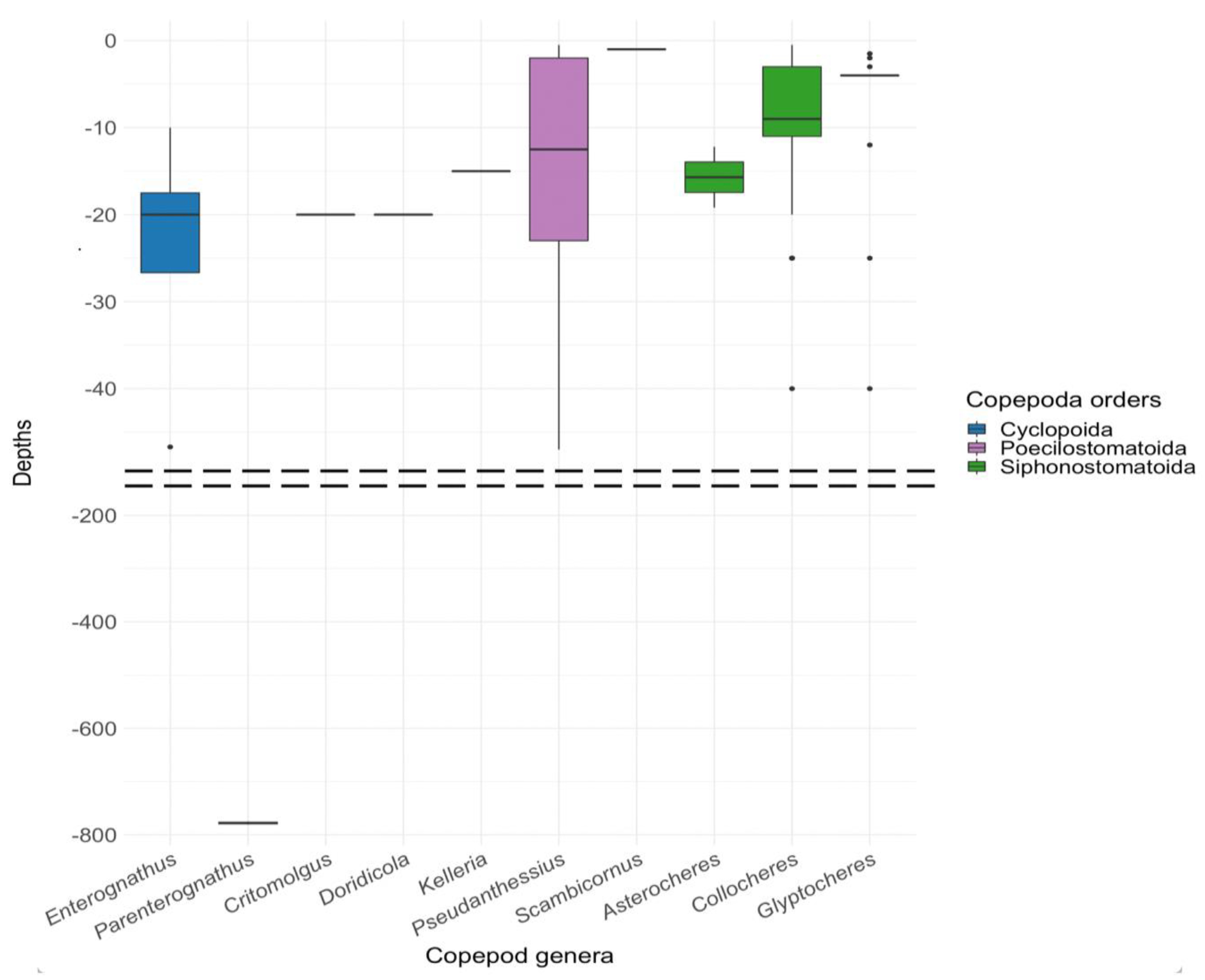
Figure 5.
Distribution of symbiotic copepods associated with crinoids by depth. This box plot illustrates the data distribution. The horizontal line within each box represents the median of the dataset. The box defines the interquartile range, covering the 25th to 75th percentiles. Whiskers extending from each box show the minimum and maximum data values. Data points appearing outside of these whiskers are identified as outliers.
Figure 5.
Distribution of symbiotic copepods associated with crinoids by depth. This box plot illustrates the data distribution. The horizontal line within each box represents the median of the dataset. The box defines the interquartile range, covering the 25th to 75th percentiles. Whiskers extending from each box show the minimum and maximum data values. Data points appearing outside of these whiskers are identified as outliers.
Differences in the bathymetric distribution of symbiotic copepods across various orders are observed. For instance, the Cyclopoida order is represented by a single genus favouring relatively shallow waters, predominantly around -20 meters. In contrast, the Poecilostomatoida order exhibits a more extensive range of habitat depths, ranging from shallow waters (-10 meters) to deeper locations (-40 meters and below), indicating a significant diversity in their living conditions. The copepods of the order Siphonostomatoida also shows adaptation to various depths, but with a narrower depth range than Poecilostomatoida, focusing around -10 meters. It is noteworthy that certain copepod genera, such as Collocheres, Enterognathus, and Scambicornus, inhabit a very narrow depth range. Exceptions in the data, particularly within the Poecilostomatoida order, may signal the presence of rare species that prefer significantly deeper waters compared to their counterparts. Overall, the data reflect a broad spectrum of adaptations among different copepod orders to living conditions at various depths, demonstrating their ecological flexibility and the diversity of ecological niches in the marine environment.
4. Conclusions
This investigation leads to a series of conclusions, tempered by the recognition of substantial gaps in the existing body of research. These gaps manifest as disparities in the depth and scope of studies, variations in taxonomic precision, and inconsistencies in the identification of specific copepod taxa. It is imperative to emphasize the need for the enhancement and standardization of research methodologies, specifically tailored to the study of microsymbiotic relationships within marine invertebrates. Such methodological advancements are crucial for addressing and bridging the conspicuous knowledge gaps in this field.
The exploration of copepod-crinoid symbiosis, representing a substantial yet largely uncharted domain within scientific inquiry, faces notable challenges. These challenges include a prevailing research bias towards macro-symbionts, the inherent complexities in the collection and analysis of microsymbiont data, and the nascent stage of marine invertebrate symbiosis research. To advance our understanding of these complex ecological interactions, concerted efforts aimed at overcoming these obstacles are imperative.
Current knowledge in the field of copepod-crinoid symbiosis represents only a fraction—approximately 5%—of the hypothesized diversity of crinoid hosts. Preliminary calculations, based on the observed diversity of copepods associated with crinoids and relying solely on morphological studies, suggest that a minimum of 600 copepod species remain to be described. The potential application of molecular methodologies, which have successfully identified previously unrecognized species diversity in other taxa at the morphological level, is expected to significantly and exponentially increase these preliminary estimates (Ivanenko et al., 2018; Martínez et al., 2020). This underscores the urgent need for more comprehensive and in-depth research efforts focused on microscopic crustaceans (Bron et al., 2011; Zeppilli et al., 2015). Future research initiatives in this area are encouraged to broaden their scope of investigation and to employ robust and innovative methodologies. Such approaches are essential for fully unraveling the complexities and nuances of symbiotic relationships within marine ecosystems. The advancement of this field critically hinges on the integration of interdisciplinary methods and a dedicated commitment to enhancing our understanding of these intricate ecological interactions. This collective effort will not only illuminate the intricate symbiotic dynamics but also make a substantial contribution to the broader understanding of marine biodiversity and ecosystem functioning.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Crinoids as hosts of copepod crustaceans.
Funding
This research was funded by the Russian Science Foundation Grant No. 22-24-00365.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in doi.org/10.15468/p2dsmy.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A

Table A1.
Copepod crustaceans recorded as associated with octocorals (see also Table S1. Octocorals as hosts of copepod crustaceans).
Table A1.
Copepod crustaceans recorded as associated with octocorals (see also Table S1. Octocorals as hosts of copepod crustaceans).
| Copepod | Host species: valid name (and as in original record) | Host abbreviation * | Symbiosis nature abbreviation ** | Site abbreviation ** | Depth (m) | Reference |
|---|---|---|---|---|---|---|
| Cyclopoida | ||||||
| Enterognathidae | ||||||
| Enterognathus comatulae Giesbrecht, 1900 | Antedon bifida (Pennant, 1777) | A | en | GB, IE | Grainger, 1950 | |
| Enterognathus comatulae Giesbrecht, 1901 | Antedon mediterranea (Lamarck, 1816) | A | en | FR, IT | Changeux, Delamare Deboutteville, 1956; Giesbrecht, 1900; Stock, 1959 | |
| Enterognathus inabai Ohtsuka, Shimomura, Kitazawa, 2012 | Lamprometra sp. | M | en | JP | 46.7-46.9 | Ohtsuka, Shimomura, Kitazawa, 2012 |
| Entherognathus lateripes Stock, 1966 | Decametra chadwicki (Clark, 1911) | Col | en | IL | 20 | Stock, 1966 |
| Entherognathus lateripes Stock, 1966 | Oligometra serripinna (Carpenter, 1811) | Col | en | IL | 20 | Stock, 1966 |
| Entherognathus lateripes Stock, 1966 | Heterometra savignii (Müller, 1841) (= Heterometra savignyi (Müller, 1841)) | H | en | IL | 10 | Stock, 1966 |
| Parenterognathus troglodytes Ohtsuka, Kitazawa, Boxshall, 2010 |
Glyptometra crassa (Clark, 1912) |
Ch | en | JP | 775, 780.8-787.1 | Ohtsuka, Kitazawa, Boxshall, 2010 |
| Poecilostomatoida | ||||||
| Kelleriidae | ||||||
| Kelleria gradata Stock, 1967 |
Heterometra savignii (Müller, 1841) (= Heterometra savignyi (Müller, 1841)) |
H | ec | IL | 15 | Stock, 1967 |
| Pseudanthessiidae |
Dichrometra flagellata (Müller, 1841) (= Dichrometra afra Clark, 1912) |
|||||
| Pseudanthessius angularis Humes, Ho, 1970 |
Stephanometra indica (Smith, 1876) (= Stephanometra spicata (Carpenter, 1881)) |
M | ec | MG | 1 | Humes, Ho, 1970 |
| Pseudanthessius angularis Humes, Ho, 1970 |
Anneissia bennetti (Müller, 1841) (= Comanthus bennetti (Müller, 1841)) |
M | ec | MG | 2, 6 | Humes, Ho, 1970 |
| Pseudanthessius comanthi Humes, 1972 | Comanthus wahlbergii (Müller, 1843) | Com | ec | MH | 4, 8 | Humes, 1972 |
| Pseudanthessius comanthi Humes, 1972 |
Oxycomanthus bennetti (Müller, 1841) (= Comanthus bennetti (Müller, 1841)) |
Com | ec | ID | 25 | Humes, 1987 |
| Pseudanthessius comanthi Humes, 1972 |
Heterometra savignii (Müller, 1841) (= Heterometra savignyi (Müller, 1841)) |
Com | ec | AU, ID, PH | 2, 3, 4, 10, 12, 40 | Humes, 1987 |
| Pseudanthessius madrasensis Reddiah, 1968 | Comatulida | ec | IN | Reddiah, 1968 | ||
| Pseudanthessius madrasensis Reddiah, 1968 | Tropiometra afra (Hartlaub, 1890) | T | ec | NC | 1.5, 2, 3 | Humes, 1977 |
| Pseudanthessius madrasensis Reddiah, 1968 | Tropiometra carinata (Lamarck, 1816) | T | ec | MG | 0.5, 1, 1.5, 2, 3, 15 | Humes, Ho, 1970 |
| Pseudanthessius major Stock, 1967 | Cenometra emendatrix (Bell, 1892) | Col | ec | MG | 10, 20 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 | Heterometra africana (Clark, 1911) | H | ec | MG | 17, 18, 25, 29, 34 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 |
Heterometra savignii (Müller, 1841) (= Heterometra savignyi (Müller, 1841)) |
H | ec | IL | 10, 15 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 |
Himerometra robustipinna (Carpenter, 1881) (= Himerometra magnipinna Clark, 1908) |
H | ec | NC | 1 | Humes, 1977 |
| Pseudanthessius major Stock, 1967 |
Dichrometra flagellata (Müller, 1841) (= Dichrometra afra Clark, 1912) |
M | ec | MG | 1, 2, 6 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 |
Lamprometra palmata (Müller, 1841) (= Lamprometra klunzingeri (Hartlaub, 1890)) |
M | ec | MG | 1, 13 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 | Liparometra sp. | M | ec | MG | 15, 23, 27, 35 | Stock, 1967 |
| Pseudanthessius major Stock, 1967 |
Stephanometra indica (Smith, 1876) (= Stephanometra spicata (Carpenter, 1881)) |
M | ec | MG, NC | 2, 3, 13, 17 | Stock, 1967; Humes, 1977 |
| Pseudanthessius minor Stock, 1967 | Heterometra africana (Clark, 1911) | H | ec | MG | 18 | Stock, 1967 |
| Pseudanthessius minor Stock, 1967 |
Dichrometra flagellata (Müller, 1841) (= Dichrometra afra Clark, 1912) |
M | ec | MG | 2 | Stock, 1967 |
| Pseudanthessius minor Stock, 1967 |
Lamprometra palmata (Müller, 1841) (= Lamprometra klunzingeri (Hartlaub, 1890)) |
M | ec | IL, MG | 0.5, 13 | Stock, 1967 |
| Pseudanthessius minor Stock, 1967 | Liparometra sp. | M | ec | MG | 15, 23, 27, 35 | Stock, 1967 |
| Pseudanthessius planus Kim, 2007 |
Himerometra robustipinna (Carpenter, 1881) (= Himerometra magnipinna Clark, 1908) |
H | ec | ID | 2 | Kim, 2007 |
| Pseudanthessius rostellatus Humes, Ho, 1970 |
Phanogenia distincta (Carpenter, 1888) (= Comaster distinctus (Carpenter, 1888)) |
Com | ec | MG | 47 | Humes, Ho, 1970 |
| Rhynchomolgidae | ||||||
| Critomolgus fishelsoni (Stock, 1967) | Oligometra serripinna (Carpenter, 1811) | Col | ec | IL | 20 | Stock, 1967 |
| Doridicola patulus (Humes, 1959) | Cenometra emendatrix (Bell, 1892) | Col | ec | MG | 20 | Humes, Stock, 1973 |
| Doridicola venustus (Humes, 1958) | Cenometra emendatrix (Bell, 1892) | Col | ec | MG | 20 | Humes, Stock, 1973 |
| Synapticolidae | ||||||
| Scambicornus pillaii Stock, 1983 |
Capillaster multiradiatus (Linnaeus, 1758) (= Capillaster multiradiata (Linnaeus, 1758)) |
Com | ec | IL | 1 | Stock, 1983 |
| Siphonostomatoida | ||||||
| Asterocheridae | ||||||
| Asterocheres crinoidicola Humes, 2000 | Comatulida | ec | JM | Kim, 2010 | ||
| Asterocheres crinoidicola Humes, 2000 | Davidaster rubiginosus (Pourtalès, 1869) | Com | ec | BZ | 12.2 | Humes, 2000 |
| Asterocheres crinoidicola Humes, 2000 | Nemaster grandis Clark, 1909 | Com | ec | BZ | 32.2 | Humes, 2000 |
| Asterocheres spinopaulus Johnsson, 1998 | Comatulida | ec | BR | Johnsson, 2002 | ||
| Collocheres amicus Kim, 2007 |
Comanthus briareus (Bell, 1882) (= Comantheria rotula Clark, 1912) |
Com | ec | ID | 17 | Kim, 2007 |
| Collocheres brevipes Shin, Kim, 2004 |
Anneissia solaster (Clark, 1907) (= Comanthus solaster Clark, 1907) |
Com | ec | KP | 25 | Shin, Kim, 2004 |
| Collocheres comanthiphilus Humes, 1987 | Comanthus parvicirrus (Müller, 1841) | Com | ec | NC | 1.5, 5 | Humes, 1987 |
| Collocheres comanthiphilus Humes, 1987 | Comanthus sp. | Com | ec | NC | 1, 3 | Humes, 1987 |
| Collocheres comanthiphilus Humes, 1987 | Comanthus wahlbergii (Müller, 1843) | Com | ec | ID, NC | 0.5, 2, 25 | Humes, 1987 |
| Collocheres comanthiphilus Humes, 1987 |
Oxycomanthus bennetti (Müller, 1841) (= Comanthus bennetti (Müller, 1841)) |
Com | ec | AU, ID, PH | 2, 3, 4, 12, 40 | Humes, 1987 |
| Collocheres humesi Kim, 2007 |
Comanthus briareus (Bell, 1882) (= Comantheria rotula Clark, 1912) |
Com | ec | ID | 17 | Kim, 2007 |
| Collocheres inaequalis Ho, 1982 |
Anneissia japonica (Müller, 1841) (= Comanthus japonica (Müller, 1841), Comanthus japonicus (Müller, 1841)) |
Com | ec | JP | Ho, 1982 | |
| Collocheres inflatiseta Humes, 1987 |
Phanogenia multibrachiata (Carpenter, 1888) (= Comaster multibrachiatus (Carpenter, 1888)) |
Com | ec | ID | 10 | Humes, 1987 |
| Collocheres marginatus Humes, 1987 |
Comaster multifidus (Müller, 1841) (= Comanthina variabilis (Bell, 1882)) |
Com | ec | AU | 9 | Humes, 1987 |
| Collocheres parvus Humes, 1987 | Davidaster rubiginosus (Pourtalès, 1869) | Com | ec | ID | 10 | Humes, 1987 |
| Collocheres prionotus Humes, 1990 | Nemaster grandis Clark, 1909 | Com | ec | ID, MG | 0.5, 1 | Humes, 1990 |
| Collocheres serrulatus Humes, 1987 | Comatulida | Com | ec | ID | 10 | Humes, 1987 |
| Collocheres solidus Shin, Kim, 2004 |
Comanthus briareus (Bell, 1882) (= Comantheria rotula Clark, 1912) |
Com | ec | KP | 25 | Shin, Kim, 2004 |
| Collocheres solidus Shin, Kim, 2004 |
Anneissia solaster (Clark, 1907) (= Comanthus solaster Clark, 1907) |
Com | ec | KP | 25 | Shin, Kim, 2004 |
| Collocheres tamladus Shin, Kim, 2004 | Comanthus parvicirrus (Müller, 1841) | Z | ec | KP | Shin, Kim, 2004 | |
| Collocheres thysanotus Humes, 1987 | Comanthus sp. | Com | ec | AU | 9 | Humes, 1987 |
| Collocheres thysanotus Humes, 1987 | Comanthus wahlbergii (Müller, 1843) | Com | ec | AU | 9 | Humes, 1987 |
| Collocheres titillator Humes, 1987 |
Oxycomanthus bennetti (Müller, 1841) (= Comanthus bennetti (Müller, 1841)) |
Com | ec | ID | 10 | Humes, 1987 |
| Collocheres uncinatus Stock, 1966 |
Comanthus briareus (Bell, 1882) (= Comantheria rotula Clark, 1912) |
Col | ec | IL | 20 | Stock, 1966 |
| Collocheres uncinatus Stock, 1966 |
Anneissia japonica (Müller, 1841) (= Comanthus japonica (Müller, 1841), Comanthus japonicus (Müller, 1841)) |
Com | ec | ID, MG | 0.5, 1, 3 | Humes, 1990 |
| Collocheres uncinatus Stock, 1966 |
Phanogenia multibrachiata (Carpenter, 1888) (= Comaster multibrachiatus (Carpenter, 1888)) |
H | ec | IL | 1, 15 | Stock, 1966 |
| Glyptocheres comanthinae Humes, 1987 |
Comaster multifidus (Müller, 1841) (= Comanthina variabilis (Bell, 1882)) |
Com | ec | ID | 4 | Humes, 1987 |
| Glyptocheres extrusus Humes, 1987 | Davidaster rubiginosus (Pourtalès, 1869) | Com | ec | NC | 1.5 | Humes, 1987 |
| Glyptocheres extrusus Humes, 1987 | Nemaster grandis Clark, 1909 | Com | ec | ID | 25 | Humes, 1987 |
| Glyptocheres extrusus Humes, 1987 | Comatulida | Com | ec | AU, ID, PH | 2, 3, 4, 12, 40 | Humes, 1987 |
* Host Abbreviations: A – Antedonidae, Ch – Charitometridae, Col – Colobometridae, Com – Comasteridae, H – Himerometridae, M – Mariametridae, T – Tropiometridae, Z – Zygometridae. ** Symbiosis nature abbreviation: ec – ectosymbiont, en – endosymbiont. *** Country abbreviation: AU – Australia, BR – Brazil, BZ – Belize, FR – France, GB – United Kingdom, ID – Indonesia, IE – Ireland, IL – Israel, IN – India, IT – Italy, JM – Jamaica, JP – Japan, KP – Korea, MG – Madagascar, MH – Marshall Islands, NC – New Caledonia, PH – Philippines.
Table A2.
Description of the dataset with specific information relative to column names, description, units, and attribute type.
Table A2.
Description of the dataset with specific information relative to column names, description, units, and attribute type.
| Attribute | Column_name | Description | Units | Attribute_Type |
|---|---|---|---|---|
| Record number | rID | Unique number corresponding to specific occurrence | Integer | |
| Record ID | recordID | A structured code incorporating a concise article reference, region and country observation identifiers, shorthand for the location coordinates, and specific abbreviations for the symbiont and host families, complemented by a distinct number. | Text | |
| Aphia ID of symbiont | aphiaID_Symbiont | Unique number for taxon from WoRMS database | Integer | |
| Kingdom of symbiont | kingdom_Symbiont | Taxonomic rank below Domain | Text | |
| Phylum of symbiont | phylum_Symbiont | Taxonomic rank below Kingdom | Text | |
| Class of symbiont | class_Symbiont | Taxonomic rank below Phylum | Text | |
| Order of symbiont | order_Symbiont | Taxonomic rank below Class | Text | |
| Family of symbiont | family_Symbiont | Taxonomic rank below Order | Text | |
| Genus of symbiont | genus_Symbiont | Taxonomic rank below Family and first element in the Latin binomial name | Text | |
| Specific epithet of symbiont | specificEpithet_Symbiont | Second element in the Latin binomial name | Text | |
| Scientific name authorship of symbiont | scientificNameAuthorship_Symbiont | Third element in the Latin binomial name | Text | |
| Symbiont ID | symbiontID | Reviewed species name | Text | |
| Taxon rank of symbiont | taxonRank_Symbiont | Taxonomic rank information (e.g., genus, species) | Text | |
| Taxonomic status of symbiont | taxonomicStatus_Symbiont | Taxonomic status information (e.g., accepted, unaccepted) | Text | |
| Link of symbiont | link_Symbiont | Link to taxon in WoRMS database | Text | |
| Female Body Length | femaleLength | The length of the female specimen, measured from head to tail | µm | Text |
| Female Body Weight | femaleWeight | The total weight of the female specimen | µm | Text |
| Male Body Length | maleLength | The length of the male specimen, measured from head to tail | µm | Text |
| Male Body Weight | maleWeight | The total weight of the male specimen | µm | Text |
| Aphia ID of host | aphiaID_Host | Unique number for taxon from WoRMS database | Integer | |
| Kingdom of host | kingdom_Host | Taxonomic rank below Domain | Text | |
| Phylum of host | phylum_Host | Taxonomic rank below Kingdom | Text | |
| Class of host | class_Host | Taxonomic rank below Phylum | Text | |
| Order of host | order_Host | Taxonomic rank below Class | Text | |
| Family of host | family_Host | Taxonomic rank below Order | Text | |
| Genus of host | genus_Host | Taxonomic rank below Family and first element in the Latin binomial name | Text | |
| Specific epithet of host | specificEpithet_Host | Second element in the Latin binomial name | Text | |
| Scientific name authorship of host | scientificNameAuthorship_Host | Third element in the Latin binomial name | Text | |
| Host ID | hostID | Reviewed species name | Text | |
| Taxon rank of host | taxonRank_Host | Taxonomic rank information (e.g., genus, species) | Text | |
| Taxonomic status of host | taxonomicStatus_Host | Taxonomic status information (e.g., accepted, unaccepted) | Text | |
| Link of host | link_Host | Link to taxon in WoRMS database | Text | |
| Site ID | siteID | Unique number for locality | Text | |
| Region code | regionCode | Unique number for region | Text | |
| Region | region | Division of the World Ocean (Spalding et al., 2007) | Text | |
| Ocean | ocean | The name of the ocean in which the locality occurs. | Text | |
| Water body | waterBody | The name of the water body in which the locality occurs. | Text | |
| Island | island | The name of the island near which the locality occurs. | Text | |
| Country | country | The name of the country in which the locality occurs. | Text | |
| Country code | countryCode | The standard code (ISO 3166-1-alpha-2) for the country in which the locality occurs. | Text | |
| Locality | locality | Particular area where the taxon was found | Text | |
| Exact Location Description | verbatimLocalition | A comprehensive description of the location from the original article | Text | |
| Geocoordinates | geocoordinates | A combined representation of both latitude and longitude | Degrees Minutes Seconds (DMS) | Text |
| Latitude | latitude | Coordinate that specifies the N–S position of a point on the Earth surface | Degrees Minutes Seconds (DMS) | Text |
| Longitude | longitude | Coordinate that specifies the E–W position of a point on the Earth surface | Degrees Minutes Seconds (DMS) | Text |
| Decimal geocoordinates | decimalGeocoordinates | A combined representation of both latitude and longitude | Decimal degrees, WGS84 | Numeric |
| Decimal latitude | decimalLatitude | Coordinate that specifies the N–S position of a point on the Earth surface | Decimal degrees, WGS84 | Numeric |
| Decimal longitude | decimalLongitude | Coordinate that specifies the E–W position of a point on the Earth surface | Decimal degrees, WGS84 | Numeric |
| Coordinate uncertainty | coordinateUncertaintyInMeters | The horizontal distance from the given decimal latitude and longitude describing the smallest circle containing the whole of the Location. | m | Integer |
| Minimum depth | minimumDepthInMeters | Vertical distance under sea level | m | Integer |
| Maximum depth | maximumDepthInMeters | Vertical distance under sea level | m | Integer |
| Collecting method | collectingMethod | The method of taking sample | Text | |
| Finding method | findingMethod | The method of finding copepods in sample | Text | |
| Type of association | note | Describes the nature of the interaction. | Text | |
| Host interaction site | locationAtHost | The general location or site on the host where the copepod interacts or resides. | Text | |
| Event date | eventDate | Date of sampling. | Date | |
| Year | year | The four-digit year in which the Occurence recorded. Format: yyyy. | Integer | |
| Month | month | The ordinal month in which the Occurence recorded. Format: mm. | Integer | |
| Article ID | articleID | Short reference | Text | |
| Reference | reference | Full reference to article | Text |
References
- Boxshall, G. A.; Halsey, S.H. An Introduction to Copepod Diversity, Part II. The Ray Society, London, 2004, pp. 422-966.
- Bron, J. E.; Frisch, D.; Goetze, E.; Johnson, S. C.; Lee, C. E.; Wyngaard, G. A. Observing copepods through a genomic lens. Front. Zool. 2011, 8(1), 1-15.
- Britayev, T.A.; Beksheneva, L.F.; Deart, Y.V. et al. Structure and variability of symbiotic assemblages associated with feather stars (Crinoidea: Comatulida) Himerometra robustipinna. Oceanology 2016, 56, 666–674.
- Britayev, T.A.; Mekhova, E.S. Assessment of hidden diversity of crinoids and their symbionts in the Bay of Nhatrang, Vietnam. Org. Divers. Evol. 2011, 11, 275–285.
- Burnell, D.J.; ApSimon, J.W. Echinoderm Saponins. In Marine Natural Products: Chemical and Biological Perspectives. Academic Press Inc., pp. 287–379.
- Changeux, J.P.; Delamare Deboutteville, C. Enterognathus comatulae Giesbrecht, 1900. Vie et Milieu 1956, 7(1), 106-107.
- Deheyn, D.D.; Lyskin, S.; Eeckhaut, I. Assemblages of symbionts in tropical shallow-water crinoids and assessment of symbionts' host-specificity. Symbiosis 2006, 42(3), 161-168.
- Fabricius, K. E.; Dale, M. B. Multispecies associations of symbionts on shallow water crinoids of the central Great Barrier Reef. Coenoses 1993, 8, 41–52.
- Fishelson, L. Ecology of northern Red Sea crinoids and their epi- and endozoic fauna. Mar. Biol. 1974, 26(2), 183-192.
- Grainger, J.N.R. Notes on parasitic Crustacea. Ann. Mag. Nat. Hist. 1950, (12)3, 635-638.
- Ho, J.S. Copepoda associated with echinoderms of the Sea of Japan. Rep. Sado Mar. Biol. Stn. Niigata Univ. 1982, 12, 33-61.
- Humes, A.G. Pseudanthessius comanthi n. sp. (Copepoda, Cyclopoida) associated with a crinoid at Eniwetok Atoll. Pac. Sci. 1972, 26(4), 373-380.
- Humes, A.G. Pseudanthessiid copepods (Cyclopoida) associated with crinoids and echinoids (Echinodermata) in the tropical western Pacific Ocean. Smithson. Contrib. Zool. 1977, 243, 1-43.
- Humes, A.G. Copepoda associated with crinoid echinoderms in the western Pacific. Publ. Seto Mar. Biol. Lab. 1987, 32(1-3), 63-108.
- Humes, A.G. Collocheres (Copepoda: Siphonostomatoida) associated with the crinoid Capillaster multiradiatus in the Indo-Pacific. Trans. Am. Microsc. Soc. 1990, 109(1), 61-68.
- Humes, A.G. Asterocheres crinoidicola n. sp., a copepod (Siphonostomatoida: Asterocheridae) parasitic on crinoids in Belize. Syst. Parasitol. 2000, 47(2), 103-110.
- Humes, A.G.; Stock, J.H. A revision of the family Lichomolgidae Kossman, 1877, cyclopoid copepods mainly associated with marine invertebrates. Smithson. Contrib. Zool. 1973, 127, 1-368.
- Humes, A.G.; Ho, J.S. Cyclopoid copepods of the genus Pseudanthessius associated with crinoids in Madagascar. Smithson. Contrib. Zool. 1970, 54, 1-20.
- Ivanenko, V.N.; Hoeksema, B.W.; Mudrova, S.V.; Nikitin, M.A.; Martínez, A.; Rimskaya-Korsakova, N.N.; Berumen, M.L.; Fontaneto, D. Lack of host specificity of copepod crustaceans associated with mushroom corals in the Red Sea. Mol. Phylogenet. Evol. 2018, 127, 770-780. doi: 10.1016/j.ympev.2018.06.024. [CrossRef]
- Huys, R.; Boxshall, G.A. Copepod Evolution. The Ray Society: London, 1991; pp. 468.
- Kim, I.-H.; Hendler, G. Larval Biology of Thaumatopsyllid Copepods Endoparasitic in Caribbean Ophiuroids. J. Crustacean Biol. 2010, 30(2), 206–224.
- Johnsson, R. Asterocherids (Copepoda; Siphonostomatoida) associated with invertebrates from California Reefs: Abrolhos (Brazil). Hydrobiologia 2002, 470, 247-266.
- Kim, I.H. Copepods (Crustacea) associated with marine invertebrates from the Moluccas. Korean J. Syst. Zool., Special Issue 2007, 6, 1-126.
- Kim, I.H. Siphonostomatoid Copepoda (Crustacea) associated with invertebrates from tropical waters. Korean J. Syst. Zool., Special issue 2010, 8, 1-176.
- Korzhavina, O.A.; Grishina, D.Y.; Chen, X.; Fontaneto, D.; Ivanenko, V.N. Diving into Diversity: Copepod Crustaceans in Octocoral Associations. Diversity 2023, 15(11), 1140.
- Martínez, A.; Eckert, E.M.; Artois, T. et al. Human access impacts biodiversity of microscopic animals in sandy beaches. Commun Biol 2020, 3, 175.
- Mikhailov, K.V.; Ivanenko, V.N. Lack of reproducibility of molecular phylogenetic analysis of Cyclopoida. Mol. Phylogenet. Evol. 2019, 139, 106574.
- Mikhailov, K.V.; Ivanenko, V.N. Low support values and lack of reproducibility of molecular phylogenetic analysis of Copepoda orders. Arthrop. Sel. 2021, 30, 39–42.
- Mekhova, E.S.; Britayev, T.A. Feather stars (Crinoidea, Comatulida) of Nhatrang Bay, Vietnam; fauna, habitat and symbionts. In Benthic fauna of the Bay of Nhatrang, Southern Vietnam, vol. 2; Britayev, T.A., Pavlov, D.S., Eds.; KMK Scientific Press: Moscow, 2012; pp. 447–478.
- Messing, C.; Gondim, A.I.; Taylor, K. World List of Crinoidea. Crinoidea. Accessed through: World Register of Marine Species at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=123081 on 2023-08-05.
- Ohtsuka, S.; Kitazawa, K.; Boxshall, G.A. A new genus of endoparasitic copepods (Cyclopoida: Enterognathidae), forming a gall in the calyx of deep-sea crinoids. Zool. Sci. 2010, 27(8), 689-696.
- Ohtsuka, S.; Shimomura, M.; Kitazawa, K. A new species of Enterognathus (Copepoda, Cyclopoida, Enterognathidae) collected from the Seto Inland Sea, western Japan. ZooKeys 2012, 180, 1-8.
- Reddiah, K. Two new Pseudanthessius species (Copepoda - Lichomolgidae) from the Madras harbour. J. Mar. Biol. Assoc. India 1968, (1-2), 320-328.
- Shin, S.; Kim, I.H. Three new species of Collocheres (Copepoda, Siphonostomatoida, Asterocheridae) associated with crinoids and ophiuroids from Korea. Korean J. Biol. Sci. 2004, 8(4), 267-280.
- Stock, J.H. Copepoda associated with Neapolitan invertebrates. Pubblicazione della Stazione Zoologica di Napoli 1959, 31(1), 59-75.
- Stock, J.H. Copepoda associated with invertebrates from the Gulf of Aqaba. 2. Enterognathus n. sp., a new endoparasite of Crinoida (Cyclopoida, Ascidicolidae). Proc. Kon. Ned. Akad. Wet. Amsterdam (C) 1966, 69(2), 211-216.
- Stock, J.H. On Collocheres Canu, 1893, and Leptomyzon Sars, 1915, two synonymous genera of Copepoda. Beaufortia 1966, 13(163), 221-239.
- Stock, J.H. Copepoda associated with invertebrates from the Gulf of Aqaba. 3. The genus Pseudanthessius Claus, 1889 (Cyclopoida, Lichomolgidae). Proc. K. ned. Akad. Wet. (C) 1967, 70(2), 232-248.
- Stock, J.H. Copepoda associated with invertebrates from the Gulf of Aqaba. 4. Two new Lichomolgidae associated with Crinoida. Proc. Kon. Ned. Akad. Wet. Amsterdam, Ser. C, Biol. Sci. 1967, 70(5), 569-578.
- Stock, J.H. A new species of Scambicornus (Copepoda, Cyclopoidea, Sabelliphilidae), associated with an unusual host, the crinoid Capillaster in the Red Sea. In Selected papers on Crustacea. Prof. (Dr.) N. Krishna Pillai felicitation volume; John, P.A., Ed.; Prof. N. Krishna Pillai Farewell Committee: Trivandrum, 1983; pp. 1-207.
- Virgili, R.; Cerrano, C.; Ponti, M. et al. Crinoid diversity and their symbiotic communities at Bangka Island (North Sulawesi, Indonesia). Mar. Biodivers. 2020, 50, 90.
- Walter, T.C.; Boxshall, G. World of Copepods database. Accessed at http://www.marinespecies.org/copepoda on 2022-04-05.
- Wright, J.S.; Fu, R.; Worden, J.R.; Chakraborty, S.; Clinton, N.E.; Risi, C.; Sun, Y.; Yin, L. Rainforest-initiated wet season onset over the southern Amazon. Proc. Natl. Acad. Sci. U.S.A. 2017, 114(32), 8481-8486.
- Zeppilli, D.; Sarrazin, J.; Leduc, D. et al. Is the meiofauna a good indicator for climate change and anthropogenic impacts? Mar Biodiv 2015, 45, 505–535. https://doi.org/10.1007/s12526-015-0359-z. [CrossRef]
- World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org/ (accessed on 15 December 2022).
Figure 1.
Numbers of new species and cumulative percentage (green line) of known species of (A) crinoids and associated with them (B) symbiotic copepods described published over time. Based on the WoRMS database (WoRMS 2022).
Figure 1.
Numbers of new species and cumulative percentage (green line) of known species of (A) crinoids and associated with them (B) symbiotic copepods described published over time. Based on the WoRMS database (WoRMS 2022).
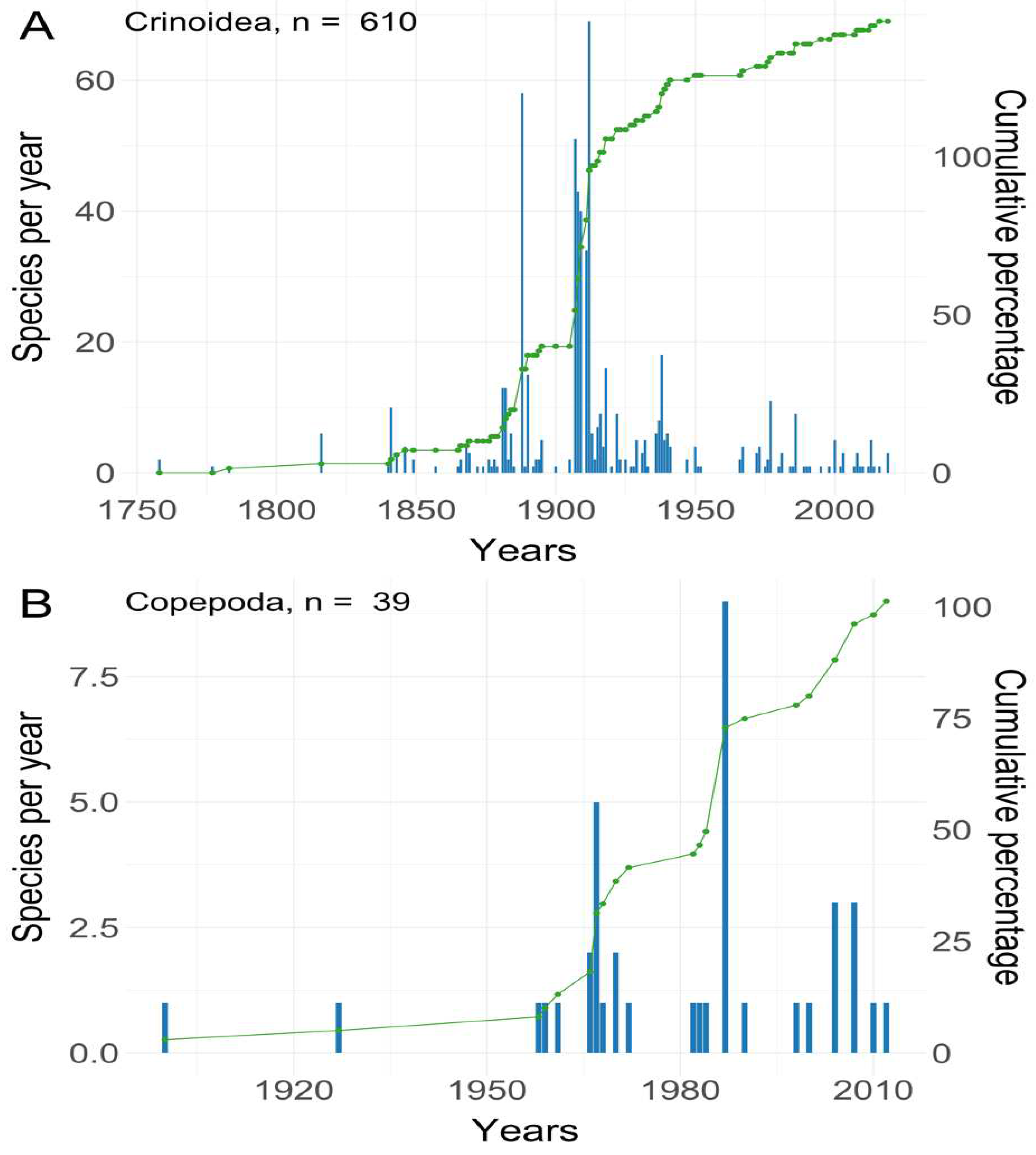
Figure 2.
Habitus of сopepod crustaceans living on crinoids: a – Enterognathus inabai, dorsal view, scale bar 1 mm; b – Parenterognathus troglodytes, dorsal view, scale bar 0.5 mm; c – Critomolgus fishelsoni, dorsal view, scale bar 0.5 mm; d – Dordicola patulus, dorsal view; e – Kelleria gradata, dorsal view, 0.2 mm; f – Pseudanthessius comanthi, dorsal view, 0.5 mm; g – Scambicornus pillaii, dorsal view, 0.1 mm; h – Asterocheres crinoidicola, dorsal view, 0.3 mm; i – Collocheres brevipes, dorsal view, 0.1 mm; (a-b – Cyclopoida, c-g – Poecilostomatoida, h-i – Siphonostomatoida). After Ohtsuka, Shimomura, Kitazawa, 2012 (a), Ohtsuka, Kitazawa, Boxshall, 2010 (b), Stock, 1967 (c, e), Humes, 1958 (d), Humes, 1972 (f), Stock, 1983 (g), Humes, 2000 (h), Shin, Kim, 2004 (i).
Figure 2.
Habitus of сopepod crustaceans living on crinoids: a – Enterognathus inabai, dorsal view, scale bar 1 mm; b – Parenterognathus troglodytes, dorsal view, scale bar 0.5 mm; c – Critomolgus fishelsoni, dorsal view, scale bar 0.5 mm; d – Dordicola patulus, dorsal view; e – Kelleria gradata, dorsal view, 0.2 mm; f – Pseudanthessius comanthi, dorsal view, 0.5 mm; g – Scambicornus pillaii, dorsal view, 0.1 mm; h – Asterocheres crinoidicola, dorsal view, 0.3 mm; i – Collocheres brevipes, dorsal view, 0.1 mm; (a-b – Cyclopoida, c-g – Poecilostomatoida, h-i – Siphonostomatoida). After Ohtsuka, Shimomura, Kitazawa, 2012 (a), Ohtsuka, Kitazawa, Boxshall, 2010 (b), Stock, 1967 (c, e), Humes, 1958 (d), Humes, 1972 (f), Stock, 1983 (g), Humes, 2000 (h), Shin, Kim, 2004 (i).
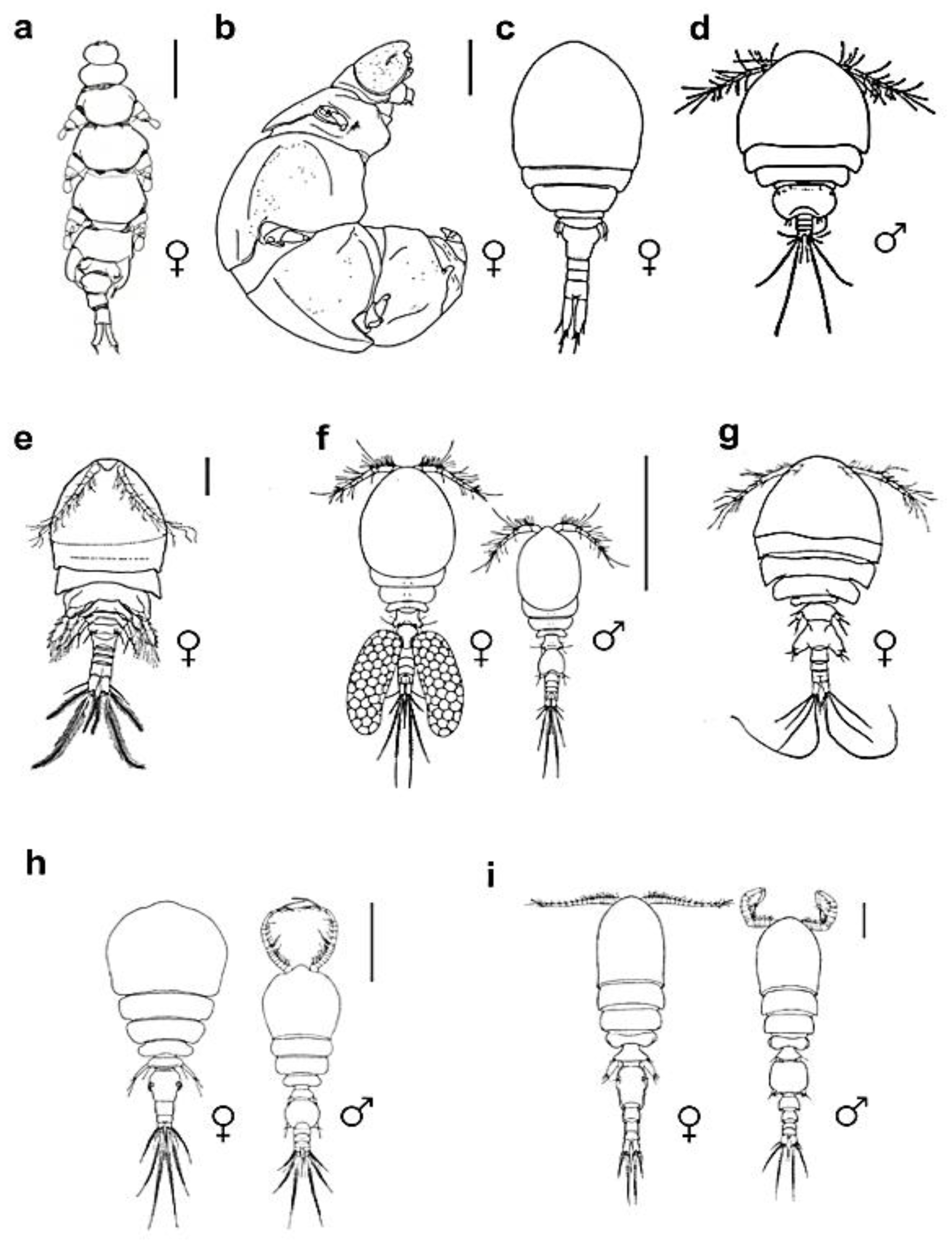
Figure 3.
Distribution of copepod associations across different crinoid families.
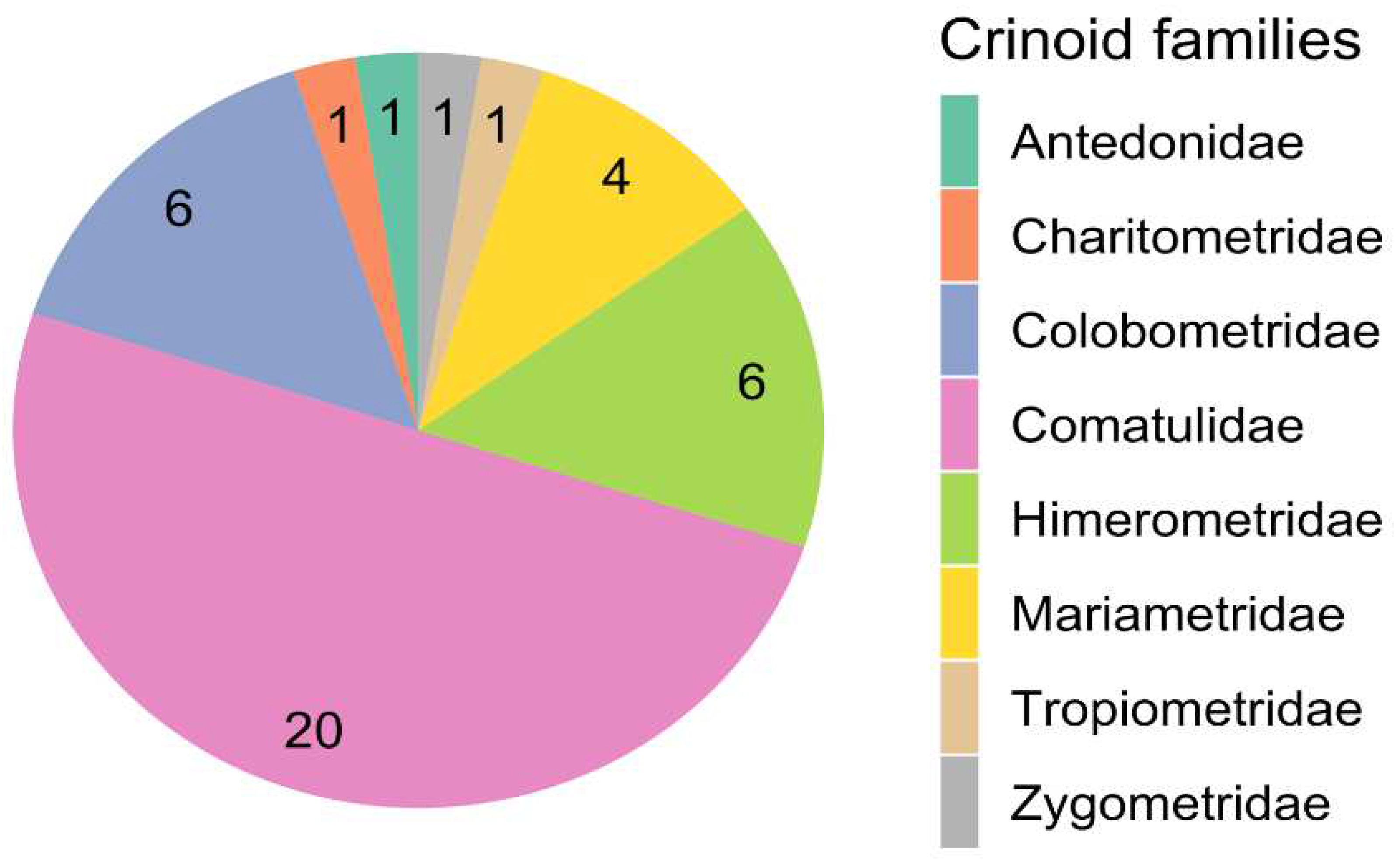
Figure 4.
Number of records per association of symbiotic copepod genera with crinoid families. Size of figure means number of records.
Figure 4.
Number of records per association of symbiotic copepod genera with crinoid families. Size of figure means number of records.
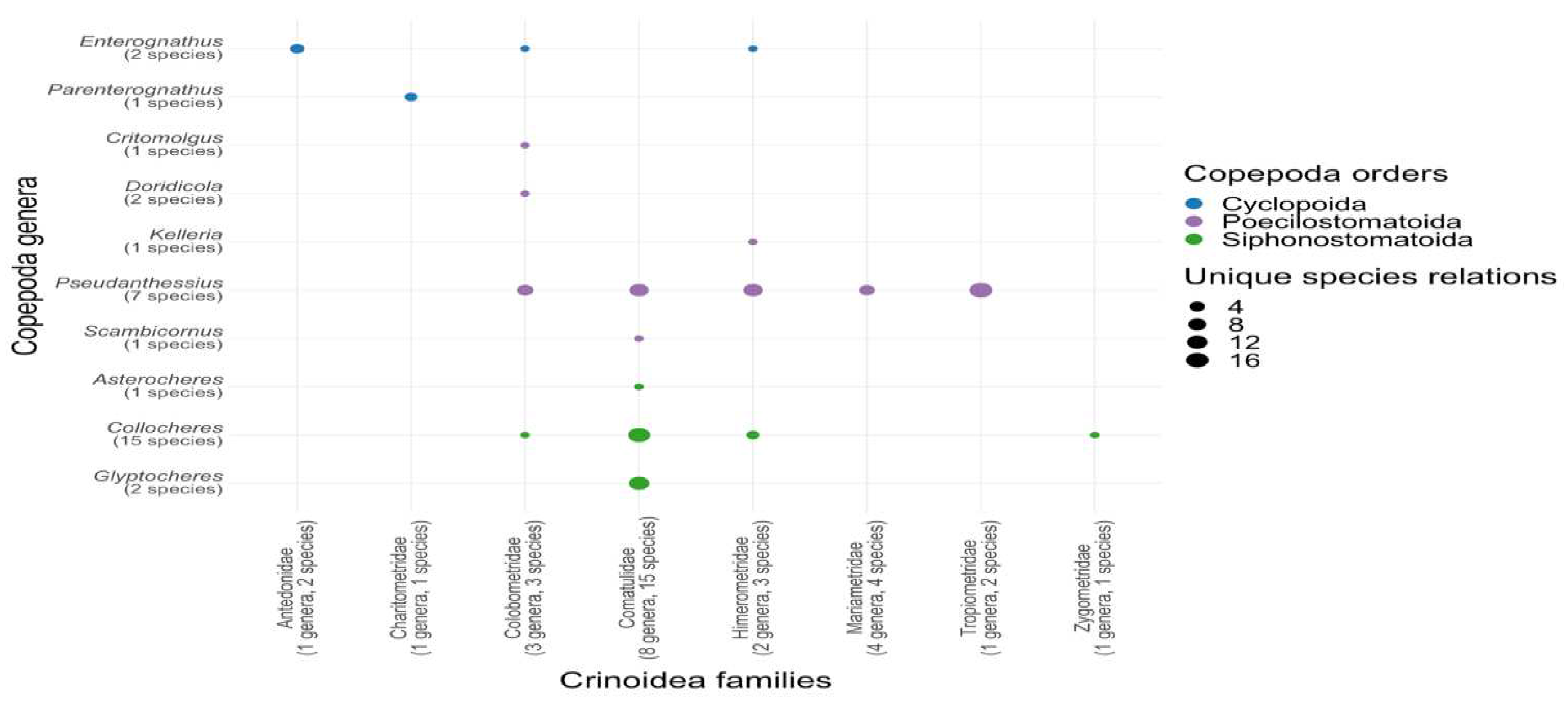
Figure 5.
Distribution of the copepods associated with crinoids in the World Ocean. The marginal histogram illustrates the latitudinal and longitudinal distribution of the reports of copepods.
Figure 5.
Distribution of the copepods associated with crinoids in the World Ocean. The marginal histogram illustrates the latitudinal and longitudinal distribution of the reports of copepods.
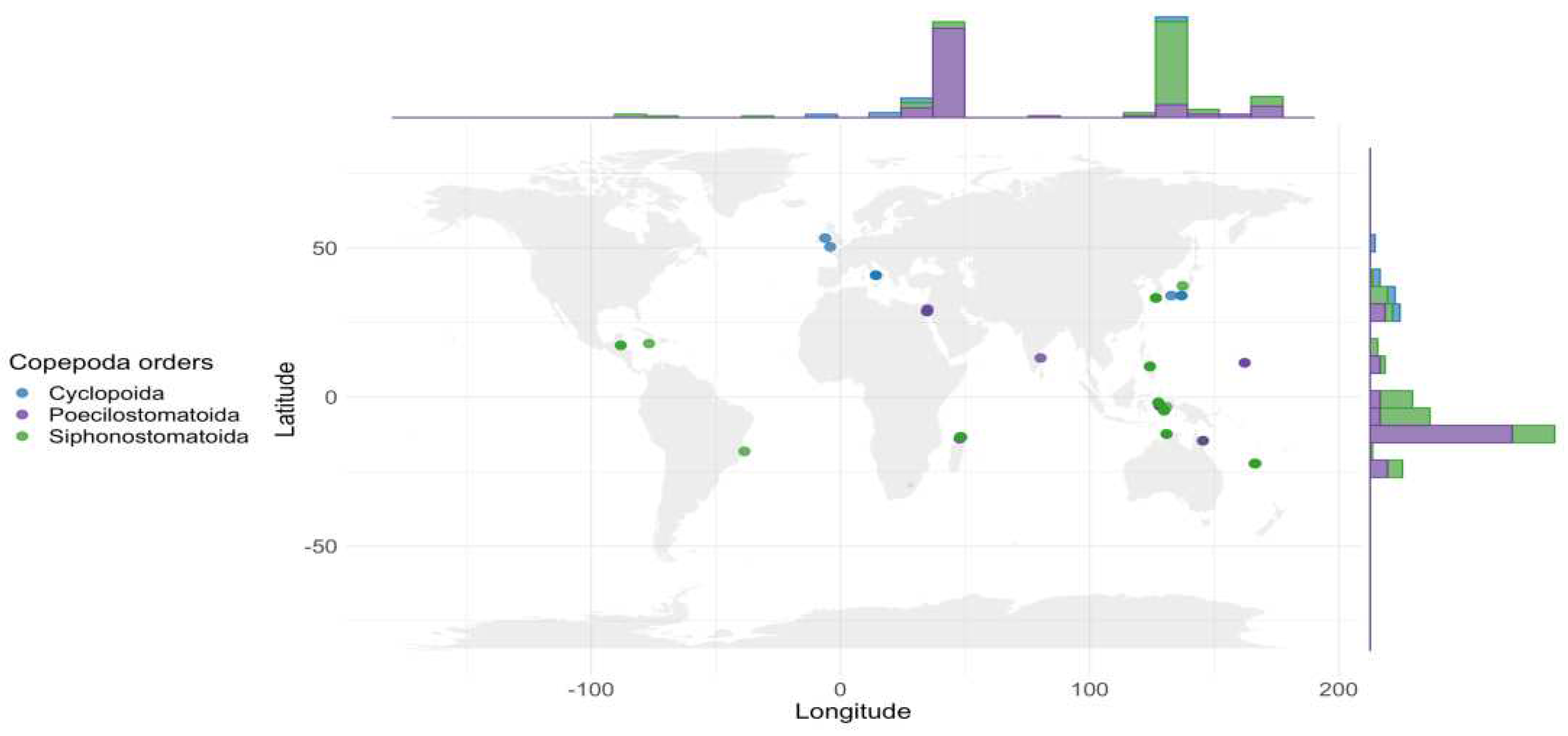
Table 1.
List of references reporting records of copepods, divided by world ocean regions and countries (for more details see Tables A1, S1).
Table 1.
List of references reporting records of copepods, divided by world ocean regions and countries (for more details see Tables A1, S1).
| Region | Country | Reference |
|---|---|---|
| Central Indo-Pacific | Australia | Humes, 1987 |
| Indonesia | Humes, 1987; Humes, 1990; Kim, 2007 | |
| Marshall Islands | Humes, 1972 | |
| New Caledonia | Humes, 1977; Humes, 1987 | |
| Philippines | Humes, 1987 | |
| Temperate Northern Atlantic | France | Changeux, Delamare Deboutteville, 1956 |
| Ireland | Grainger, 1950 | |
| Italy | Giesbrecht, 1900; Stock, 1959 | |
| United Kingdom | Grainger, 1950 | |
| Temperate Northern Pacific | Japan | Ho, 1982; Ohtsuka, Kitazawa, Boxshall, 2010; Ohtsuka, Shimomura, Kitazawa, 2012 |
| Korea | Shin, Kim, 2004 | |
| Tropical Atlantic | Belize | Humes, 2000 |
| Brazil | Johnsson, 2002 | |
| Jamaica | Kim, 2010 | |
| Western Indo-Pacific | India | Reddiah, 1968 |
| Israel | Stock, 1966; Stock, 1967; Stock, 1983 | |
| Madagascar | Humes, 1990; Humes, Ho, 1970; Humes, Stock, 1973; Stock, 1967 |
Table 2.
The families of Copepoda in relation to Octocorallia
| Taxa | # of known copepod species | # copepod species | # of copepod records found on crinoids | # crinoid families | # crinoid genera | # crinoid species | Mean of records per copepod species + SE | Mean of host species per copepod species + SE | % copepod species with a single crinoid host |
|---|---|---|---|---|---|---|---|---|---|
| Cyclopoida | |||||||||
| Enterognathidae | 7 | 4 | 11 | 5 | 6 | 7 | 2.75 + 0.85 | 1.75 + 0.48 | 50 |
| Poecilostomatoida | |||||||||
| Kelleriidae | 19 | 1 | 1 | 1 | 1 | 1 | 1 + NA | 1 + NA | 100 |
| Pseudanthessiidae | 61 | 7 | 77 | 6 | 13 | 14 | 13.57 + 5.03 | 3 + 0.93 | 28,57 |
| Rhynchomolgidae | 270 | 3 | 3 | 1 | 2 | 2 | 1 + 0 | 1 + 0 | 100 |
| Synapticolidae | 50 | 1 | 1 | 1 | 1 | 1 | 1 + NA | 1 + NA | 100 |
| Siphonostomatoida | |||||||||
| Asterocheridae | 271 | 19 | 73 | 5 | 12 | 17 | 3.84 + 1.17 | 2 + NA | 63,16 |
| Total | 678 | 35 | 166 | 19 | 35 | 42 |
* WoRMS database [33].
Table 3.
Crinoidea families in relation to copepods
| Host taxa | # of known crinoid genera | # host crinoid genera (%) | # of known species | # host crinoid species (%) | # records | # of copepod species found on crinoids | # of host species with | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||||||
| copepod species | ||||||||||
| Comatulida | ||||||||||
| Antedonidae | 50 | 1 (2%) | 151 | 2 (1.32%) | 5 | 1 | 2 | |||
| Charitometridae | 8 | 1 (12.5%) | 33 | 1 (3.03%) | 2 | 1 | 1 | |||
| Colobometridae | 18 | 3 (16.67%) | 47 | 3 (6.38%) | 11 | 6 | 1 | 2 | ||
| Comatulidae | 23 | 8 (34.78%) | 102 | 16 (15.69%) | 81 | 20 | 7 | 5 | 3 | 1 |
| Himerometridae | 5 | 2 (40%) | 39 | 3 (7.69%) | 18 | 6 | 2 | 1 | ||
| Mariametridae | 7 | 4 (57.14%) | 22 | 5 (22.73%) | 24 | 4 | 1 | 3 | 1 | |
| Tropiometridae | 1 | 1 (100%) | 4 | 2 (50%) | 21 | 1 | 2 | |||
| Zygometridae | 2 | 1 (50%) | 10 | 1 (10%) | 1 | 1 | 1 | |||
| Total | 114 | 21 | 408 | 33 | 163 | 40 | 15 | 10 | 6 | 2 |
* WoRMS database [33]
Table 4.
The distribution of symbiotic copepods and their hosts in the ecoregions *
| Region | # of localities | # of records | # of symbiontorders | # of symbiontfamilies | # of symbiontgenera | # of symbiontspecies | # of host families | # of host genera | # of host species |
|---|---|---|---|---|---|---|---|---|---|
| Central Indo-Pacific | 21 | 74 | 2 | 2 | 3 | 17 | 4 | 9 | 14 |
| Temperate Northern Atlantic | 4 | 5 | 1 | 1 | 1 | 1 | 1 | 1 | 2 |
| Temperate Northern Pacific | 5 | 11 | 2 | 2 | 3 | 6 | 4 | 4 | 5 |
| Tropical Atlantic | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| Western Indo-Pacific | 23 | 71 | 3 | 6 | 7 | 13 | 5 | 11 | 12 |
* WoRMS database
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



