Submitted:
05 December 2023
Posted:
06 December 2023
You are already at the latest version
Abstract
The aim of our present study was to investigate the effects of Gln supplementation on liver inflammatory responses and protein synthesis and degradation in muscle of LPS-challenged broilers. A total of 120 1d-old male broilers (Arbor Acres Plus) were randomly arranged in a 2 × 2 factorial design with five replicates per treatment and six broilers per replicate, containing two main factors: immune challenge (injected with LPS in a dose of 0 or 500 µg/kg of body weight) and dietary treatments (supplemented with 1.22% alanine or 1% Gln). After feeding with alanine or Gln diet for 15 days, broilers were respectively administrated with LPS or saline injection at 16 and 21 d. The results showed that Gln supplementation significantly reversed the adverse effects of LPS administration on growth performance, as evidenced as the increased ADFI and ADG, as well as the lowered F/G. Furthermore, Gln supplementation alleviated the increased mRNA expression of interleukin-6, interleukin-1β and tumor necrosis factor-α in liver induced by LPS. Moreover, the increased activities of aspartate aminotransferase, combined with the decreased expression of glutaminase in muscle, were observed by Gln addition. In addition, in comparison with saline treatment, LPS challenge altered signaling molecules mRNA expressions associated with protein synthesis and degradation. However, Gln supplementation reversed the negative effects on protein synthesis and degradation in muscle of LPS-challenged broilers. Taken together, Gln supplementation had benefit effects on improving growth performance, alleviating inflammatory responses, promoting protein synthesis and inhibiting protein degradation of LPS-challenged broilers.
Keywords:
lipopolysaccharide challenge
; glutamine
; liver inflammatory
; protein synthesis
; protein degradation
1. Introduction
Under intense breeding conditions, broilers commonly suffered from immune stress induced by diverse pathogenic and nonpathogenic microorganisms, which caused adverse changes in body metabolism and immune system and ultimately resulted in the impaired growth performance [1]. Lipopolysaccharide (LPS), an active component of gram-negative bacterial cell membrane and an effective immunostimulatory component, was preferentially used to establish immune-stress model via intraperitoneal injection in broilers [2,3]. Previous studies suggested that an acute systemic inflammatory response such as the production of inflammatory factors and the altered expression of genes involved in the immune system occurred in response to LPS stimulation [4]. Our previous study showed that LPS challenge resulted in the impaired growth performance, the elevated inflammatory cytokines in serum and the destructed intestinal mucosal barrier in broilers [5]. In a previous study, it was demonstrated that LPS-induced immune stress led to a severe loss of muscle mass [6]. Moreover, LPS challenge also altered the molecular pathways regulating the catabolism of muscle mass, which contained the signaling molecules responsible for the increased protein degradation and the decreased protein synthesis [7].
Glutamine (Gln), the most abundant amino acid in plasma and skeletal muscle, falls dramatically in circulation and tissue during stress conditions [8]. Therefore, Gln becomes a conditionally essential amino acid during inflammatory conditions [9]. It has been demonstrated that Gln has a wide range of biological functions and serves important roles in promoting growth performance, maintaining intestine health and improving immune response [5,10,11,12]. Previous studies suggested that 1% Gln supplementation improved growth performance of broilers [8,13]. Besides, Gln addition contributed to alleviating the inflammatory reactions evidenced as the decreased concentrations of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) and interleukin-6 (IL-6) in plasma [5]. Additionally, Gln is considered to be one of the important amino acids in regulation of mTOR and autophagy [14,15], which are respectively responsible for protein synthesis and degradation. However, the regulation of Gln on protein synthesis and degradation of muscle in LPS-challenged broilers was still not well understood.
Therefore, the present study was to investigate the effects of Gln supplementation on liver inflammatory responses and protein synthesis and degradation in muscle of LPS-challenged broilers.
2. Materials and methods
2.1. Diets, Experimental design and Animal management
The protocols of this experiment were performed according to the guidelines of the Institutional Animal Care and Use Committee of Qingdao agricultural university (QAU1121010) and approved by the Animal Care and Use Committee.
The dosage of Gln (1.0%) supplementation and Ala (1.22%) as isonitrogenous control were according to our previous study [5]. Both Ala and Gln were purchased from Shanghai Feiya Technology Development Co., Ltd. (Shanghai, China).
120 1d-old male broilers (Arbor Acres Plus) were randomly arranged in a 2 × 2 factorial design, containing five replicates per treatment and six broilers per replicate. Broilers were fed with either Ala or Gln diet for 15 days. However, broilers were intraperitoneally injected with LPS solution at a dosage of 500 μg LPS/kg of body weight or the equal volume of 0.9% sterile saline on 8.00 a.m. of d 16 and d 21, respectively. Therefore, the four experimental treatments were as follows: (1) Ala-saline group, in which birds were fed with diet containing 1.22% Ala and received intraperitoneal administration of sterile saline; (2) Gln-saline group, in which birds were fed with diet containing 1% Gln and received intraperitoneal administration of sterile saline; (3) Ala-LPS group, in which birds were fed with diet containing 1.22% Ala and received intraperitoneal administration of LPS; (4) Gln-LPS group, in which birds were fed with diet containing 1% Gln and received intraperitoneal administration of LPS. The ingredients of diets and nutrition contents were presented in Table 1. During the entire experiment, broilers were ad libitum access to feed in mash form and fresh water.
2.2. Sample Collection
At 21 d of age after LPS injection for 2 h, 10 broilers (2 broilers per replicate) were randomly selected to obtain blood samples from the wing vein. The collected blood samples were centrifuged at 4 °C 3000 × g for 10 min to collect plasma and stored at -20°C for further analysis. Immediately after blood sampling, the selected broilers were sacrificed by cervical dislocation followed by exsanguination. About 10 g. pectoralis muscle were collected into sterile tube and stored in liquid nitrogen for subsequent analysis. All samples were collected within 10 minutes after killing.
2.3. Growth performance
At 16 and 21 d of age, broilers were weighed after a 12-h feed withdraw. The total feed consumption and the total weight of birds for each replicate were respectively recorded to calculate average feed daily intake (ADFI), average daily gain (ADG) and the ratio of feed to gain (F/G).
2.4. Real-Time PCR analysis
Total RNA extraction from pectoralis muscle samples isolated using RNAiso Plus reagent (catalogue no. 9108, TaKaRa Biotechnology (Dalian) CO., LTD. Dalian China). were reverse transcribed into cDNA using PrimeScript TM RT Master Mix (catalogue no. RR037A, TaKaRa), followed by a Real time RT-PCR analysis with TB Green Premix Ex Taq (catalogue no. RR420A) according to the instructions of manufacture. PCR program consisted of one cycle at 95°C for 30s, 40 cycles of denaturation at 95°C for 5s, followed by a 60°C annealing step for 30s. The expression of selected genes relative to the housekeeping gene (β-actin) were analyzed according to the method by Livak and Schmittgen [16]. The primer sequences for the target genes toll-like receptor 4 (TLR4), muscle atrophy F-box (MAFbx), muscle ring finger 1 (MuRF1), forkhead Box O1 (FOXO1), forkhead Box O4 (FOXO4), protein kinase B (Akt), mammalian target of rapamycin (mTOR), eIF-4E binding protein-1 (4E-BP1), ribosomal protein S6 kinase (S6K1) and β-actin are shown in Table 2.
2.5. Statistical analysis
Data analysis was conducted by a 2×2 factorial arrangement (SPSS version 20.0). The statistical model contained diet (1.2% Ala or 1.0% Gln) and intraperitoneal injection challenge (LPS or saline) and their interactions. P<0.05 was considered significant. All values are shown as means and standard error of the mean.
3. Results
3.1. The growth performance of broilers challenged with LPS
As reported in our previous study [5], there were no differences in the control group and Gln addition group before LPS-challenge treatment (P<0.05). However, lower body weight, ADFI and ADG, but higher F/G were observed in broilers received LPS challenge in comparison with those receiving saline-injection treatment (P<0.05). Moreover, compared with the control group, Gln addition significantly increased body weight of 21 d, ADFI and ADG, as well as the lowered F/G (P<0.05). There were no interactions for body weight of 16 d and 21d, ADG, ADFI and F/G of broilers between Gln addition and LPS challenge (P>0.05).
3.2. mRNA expression of TNF-α、IL-6、IL-1β in liver
As illustrated in Figure 1, in comparison with those received saline treatment, LPS challenge significantly increased mRNA expression of TNF-α、IL-6 and IL-1β in liver (P<0.05). However, Gln supplementation reversed the increased expression of TNF-α、IL-6 and IL-1β induced by LPS stimulation (P<0.05). Significant interactions for IL-6 and IL-1β (P<0.05) except for TNF-α (P>0.05) were observed between Gln addition and LPS challenge.
3.3. The activities of ALT and AST in muscle of broilers
The activity of AST in Pecloralis Muscle of broilers was decreased by LPS administration in comparison with those treated with saline injection (P<0.05, Table 3). However, no difference in the activity of ALT was observed in LPS treatment group (P>0.05). In contrast, dietary Gln supplementation significantly elevated the activity of AST of LPS-challenged broilers (P<0.05). But Gln supplementation did not affect the activity of ALT(P>0.05). In addition, no interaction for the activities of AST or ALT was observed between LPS treatment and Gln supplementation (P>0.05).
3.4. GS activity and mRNA expression of GA in Pecloralis Muscles of broilers
As presented by Figure 2, in compared with broilers received saline injection, LPS challenge significantly increased the activity of GS (P<0.05). Meanwhile, mRNA expressions of GA were also upregulated by LPS treatment (P<0.05). However, Gln supplementation downregulated the increased mRNA expression of GA induced by LPS stimulation (P<0.05). There were no interactions for GS activity and GA mRNA expression between LPS challenge and Gln supplementation (P>0.05).
GS, glutamine synthetase; GA, glutaminase; Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means ± SEM, n=10. Different superscripts above the column means differences, P<0.05.
3.5. mTOR signaling molecules in muscle
In comparison with saline treatment, LPS treatment significantly lowered mRNA expressions of mTOR, 4E-BP1 and S6K1 in Pecloralis Muscles of broilers (P<0.05, Figure 3). However, mRNA expressions of mTOR, 4E-BP1 and S6K1 were all elevated by Gln supplementation (P<0.05). A significant interaction for 4E-BP1 was observed between LPS challenge and Gln supplementation (P<0.05). But there were no interactions for mTOR and S6K1 between LPS challenge and Gln supplementation (P>0.05).
mTOR, mammalian target of rapamycin; S6K1, ribosomal protein S6 kinase; 4E-BP1, eIF-4E binding protein-1; Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means ± SEM, n=10. Different superscripts above the column means differences, P<0.05.
3.6. mRNA expression of Akt/FOXO signals mediated by TLR4
As shown in Table 4, when compared with broilers in saline treatment, LPS challenge significantly decreased Akt mRNA expression, but increased mRNA expressions of TLR4, FOXO4, FOXO1, MAFbx and MuRF1 (P<0.05). In contrast, Gln supplementation upregulated Akt mRNA expression and downregulated mRNA expressions of TLR4, FOXO4, FOXO1, MAFbx and MuRF1 (P<0.05). There were significant interactions for TLR4, FOXO1 and MuRF1 between LPS challenge and Gln supplementation (P<0.05), however, no interactions for Akt, FOXO4 or MAFbx was observed between LPS challenge and Gln supplementation (P>0.05).
4. Discussion
Under the condition of LPS exposure, animals generally were subjected to the reduced appetite [17], the destroyed integrity of intestinal mucosal barrier [18] and nutrients were redistributed from anabolic and body protein deposition towards utilization for immune system functioning [19], followed by the reduced ADFI and ADG combined with the increased F/G. Accordingly, compared with those received saline administration, LPS challenge reduced the retardation of the growth performance, evidenced as the decreased in ADFI and ADG as well as the increased F/G in our present study. Similar with this, it was reported that LPS challenge with intra-peritoneal injection significantly decreased the growth performance of broilers [20,21]. However, Gln supplementation reversed the negative effects on the growth performance of broilers indued by LPS challenge. Moreover, Gln, a neutral and multifunctional essential amino acid, is proven to be particularly prominent in the anti-stress response [9,22]. In addition, in previous studies, it was demonstrated that 1% Gln supplementation improved the growth performance of broilers under stress, accompanied by the increased in ADFI and ADG and the decreased F/G [8,13], which indicated that Gln administration contributed to ameliorating the adverse effects of LPS stimulation on the growth performance of broilers.
It had been demonstrated that LPS is a potential stimulation triggering the release of proinflammatory cytokines [23,24]. The secretion of proinflammatory cytokines are crucial for activating the innate host defense system and subsequently regulating the adaptive immune response, such as IL-6, TNF-α and IL-1β [21]. TNF-α, IL-6 and IL-1β, originated from macrophages, are the major regulators in diverse inflammatory responses [5]. The results of our present study showed that LPS challenge increased mRNA expressions of TNF-α, IL-6 and IL-1β in liver, indicating that LPS challenge induced an acute inflammatory response. It was demonstrated that LPS challenge resulted in higher mRNA expression TNF-α, IL-6 and IL-1β in liver of broilers [25]. In similar with this, the increased mRNA expressions of IL-6 and IL-1β were observed in LPS-challenged broilers [21]. The results of our previous study suggested that Gln supplementation decreased the contents of TNF-α, IL-6 and IL-1β in plasma of LPS-challenged broilers [5]. Moreover, it was proven that Gln deprivation exacerbates the production of proinflammatory cytokines, whereas Gln supplementation limits the inflammatory response in vitro [26]. The results aforementioned indicated that Gln supplementation might help to alleviate the inflammatory responses induced by LPS challenge.
Because of the physiological demand of Gln exceeded the synthesis capacity under catabolic stresses, it became the conditionally essential amino acid [27]. Skeletal muscle plays important roles in Gln metabolism and is quantitatively the most relevant site of Gln stock, synthesis and release site [28]. The two intracellular enzymes are glutamine synthetase (GS) and phosphate-dependent glutaminase (GA), which respectively are responsible for Gln synthesis and Gln hydrolysis [29]. It has been proven that there are a concomitant increase in GA expression under catabolic conditions, such as sepsis and infections [30,31]. Similarly, in our present study, LPS administration significantly increased mRNA expression of GA in muscle of broilers suggesting that Gln consumption accelerated induced by LPS challenge. In addition, the increased GS enzyme activity in skeletal muscle was observed during severe catabolic states [29]. In according with this, our results also showed that LPS challenge elevated the activity of GS in muscle. Moreover, Gln metabolism is also influenced by glutamine aminotransferase. In this study, Gln supplementation increased AST activity in muscle. Consistent with the result of ours, it was also reported that 1% Gln supplementation increased AST activity in muscle associated with Gln metabolism [32]. However, Gln addition significantly decreased GA mRNA expression in muscle, indicating that Gln supplementation might compensated for the decline of Gln in muscle under stress and partly contributed to inhibiting the catabolism of Gln.
It is well documented that mTOR signaling pathway is an evolutionally conserved protein kinase and is important in regulating protein synthesis. The active of mTOR and its downstream regulators 4E-BP1 and S6K1 synergistically leads to the initiation of polypeptide formation [33]. But mTOR signaling is inhibited by sepsis and endotoxin-related inflammation [34]. It was found that LPS challenge downregulated mRNA expressions of mTOR, 4E-BP1 and S6K1 in muscle in our present study. It was also reported that the expressions of mTOR, 4E-BP1 and S6K1 were inhibited in septic rats or animals treated with LPS [34], indicating that protein synthesis was inhibited by LPS challenge. Independent of LPS administration, the increased in mRNA expressions of mTOR, combined with 4E-BP1 and S6K1 in muscle, were observed by Gln addition. It has been demonstrated that Gln is required for the activation of mTOR signaling [35]. In similar with this, in a previous study, Gln supplementation significantly elevated the protein expressions of mTOR, 4E-BP1 and S6K1 in skeletal muscle [27]. In addition, Gln addition to the medium stimulated protein synthesis through mTOR signaling pathway [36]. The results mentioned above suggested that Gln contributed to promoting protein synthesis associated with the activation of mTOR signaling.
The major of intra-cellular proteins are degraded by the ubiquitin-proteasome pathway in all tissues [37], which contributes to 75% protein degradation during skeletal muscle atrophy [38]. The activation of Akt and inactivation of FOXO transcriptionally upregulated of FOXO gene targets MAFbx and MuRF1, subsequently induced muscle protein degradation [39]. TLR4 was also demonstrated to be a master regulator of muscle wasting induced by endotoxemia [40]. The present study found that LPS challenge significantly decreased mRNA expression of Akt and increased mRNA expression of TLR4, FOXO1, FOXO4, MAFbx and MuRF1 in muscle. It has been demonstrated that the decreased in Akt expression and the increased expressions of MAFbx and MuRF1 occurred in response to LPS-induced endotoxaemia [41]. Besides, in a previous study, it was demonstrated that increased mRNA expression of FOXO1 and FOXO4, combined with the increased MAFbx and MuRF1 mRNA expression were induced by LPS challenge [7]. In similar, in a previous study about LPS-administrated rat, it was demonstrated that LPS challenge downregulated Akt expression and upregulated MAFbx and MuRF1 in skeletal muscle [39]. Currently, Gln has been shown to be an amino acid involved in the regulation of autophagy [42]. It was proven that Gln administration inhibited protein degradation of intestinal epithelial cells [36]. Our results also showed that Gln addition increased Akt mRNA expression and lowered mRNA expression of MAFbx and MuRF1 associated with protein degradation. Moreover, in our previous study in piglets, alanyl-glutamine (a dipeptide of Gln) supplementation decreased mRNA expressions of both MAFbx and MuRF1 in skeletal muscle under normal and LPS condition [7,27]. In addition, Gln administration induced the reduced mRNA expression of MAFbx and MuRF1, and the loss in the skeletal muscle mass was alleviated partially by Gln supplementation [43]. Therefore, based on the mentioned above, we could speculate that Gln supplementation could contribute to inhibiting protein degradation of skeletal muscle via TLR4/Akt/UPP signaling pathway.
5. Conclusion
In conclusion, Gln supplementation improved growth performance of broilers administrated with LPS and alleviated inflammatory response in liver. Moreover, Gln supplementation increased the expressions of signaling molecules in mTOR and Akt/FOXO/UPP pathway, indicating that Gln addition might contributed to promoting protein synthesis and inhibiting protein degradation of LPS-challenged broilers.
Author Contributions
B. L. Zhang mainly wrote this manuscript and experimental administration. N. Liu assisted in data analysis and writing the manuscript. Q. Yang conducted in animal trial. Q.Z. Zhong and Z.W. Sun assisted in revising the manuscript. All authors reviewed the final manuscript and approved the submitted version. All authors have no conflict on the interest of this paper.
Funding
This work was supported by the Basic Project of Guizhou Provincial Natural Science Foundation [Qian Kehe jichu-ZK (2022) Yiban 578] and Characteristic Laboratory of Animal Resources Conservation and Utilization of Chishui River Basin (Qianjiaohe KY [2013]111-03).
References
- Liu K. X., Zhen W. R., Bai D. Y., Tan H. Q., He X. L., Li Y. Q., Liu Y. H., Zhang Y., Ito K., Zhang B. K., Ma Y. B. Lipopolysaccharide-induced immune stress negatively regulates broiler chicken growth the COX-2-PGE-EP4 signaling pathway. Frontiers in immunology 2023, 14, 1193798. [CrossRef] [PubMed]
- Bi S., Shao J. J., Qu Y. W., Hu W. D., Ma Y., Cao L. T. Hepatic transcriptomics and metabolomics indicated pathways associated with immune stress of broilers induced by lipopolysaccharide. Poultry science 2022, 101, 102199. [CrossRef] [PubMed]
- Wu Y. P., Li Q., Liu J. S., Liu Y. L., Xu Y. L., Zhang R. Q., Yu Y., Wang Y. X., Yang C. M. Integrating Serum Metabolome and Gut Microbiome to Evaluate the Benefits of Lauric Acid on Lipopolysaccharide- Challenged Broilers. Frontiers in immunology 2021, 12, 759323. [CrossRef]
- Lee Y., Lee S. H., Gadde U. D., Oh S. T., Lee S. J., Lillehoj H. S. Dietary Allium hookeri reduces inflammatory response and increases expression of intestinal tight junction proteins in LPS-induced young broiler chicken. Research in veterinary science 2017, 112, 149–55. [CrossRef] [PubMed]
- Zhang B. L.., Zhong Q Z., Liu N., Song P. Y., Zhu P., Zhang C. C., Sun Z. W. Dietary Glutamine Supplementation Alleviated Inflammation Responses and Improved Intestinal Mucosa Barrier of LPS-Challenged Broilers. Animals 2022, 12, 1729. [CrossRef]
- Orellana R. A., Suryawan A., Wilson F. A., Gazzaneo M. C., Fiorotto M. L., Nguyen H. V., Davis T. A. Development aggravates the severity of skeletal muscle catabolism induced by endotoxemia in neonatal pigs. American journal of physiology. Regulatory, integrative comparative physiology 2012, 302, R682-90. [CrossRef]
- Zhang B., Yu C., Lin M., Fu Y., Zhang L., Meng M., Xing S., Li J., Sun H., Gao F. Regulation of skeletal muscle protein synthetic and degradative signaling by alanyl-glutamine in piglets challenged with Escherichia coli lipopolysaccharide. Nutrition 2015, 31, 749–56. [CrossRef] [PubMed]
- Dai S. F., Wang L. K., Wen A. Y., Wang L. X., Jin G. M. Dietary glutamine supplementation improves growth performance, meat quality and colour stability of broilers under heat stress. British poultry science 2009, 50, 333–40. [CrossRef] [PubMed]
- Bai X., Wang K., Khan R., U, Zhang C., Hu H. Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers. Animals 2023, 13, 652. [CrossRef]
- Wu Q. J.., Zhu D. D., Wang D. D.., Zhang B B., Ren A., Zhang Z. B. Effects of dietary supplementation with glutamine on the lymphocyte proliferation and intestinal immune gene expression in broiler chickens infected with Salmonella Enteritidis. Research in veterinary science 2021, 139, 18–24. [CrossRef] [PubMed]
- Zhang B. L., Liu N., Hao M. L., Xie Y. X., Song P. Y. Effects of substitution of soybean meal with rapeseed meal and glutamine supplementation on growth performance, intestinal morphology, and intestinal mucosa barrier of Qiandongnan Xiaoxiang Chicken. Animal bioscience 2022, 35, 1711–24. [CrossRef] [PubMed]
- Xing S., Zhang B. L., Lin M., Zhou P., Li J. L., Zhang L., Gao F., Zhou G. H. Effects of alanyl-glutamine supplementation on the small intestinal mucosa barrier in weaned piglets. Asian-Australasian journal of animal sciences 2017, 30, 236–45. [CrossRef]
- Xue G. D., Barekatain R., Wu S. B., Choct M., Swick R. A. Dietary L-glutamine supplementation improves growth performance, gut morphology, and serum biochemical indices of broiler chickens during necrotic enteritis challenge. Poultry science 2018, 97, 1334–41. [CrossRef] [PubMed]
- Zhou J., Chen H. H., Du J. T., Tai H. R., Han X. J., Huang N., Wang X. B., Gong H., Yang M. Y., Xiao H. Y. Glutamine Availability Regulates the Development of Aging Mediated by mTOR Signaling and Autophagy. Frontiers in pharmacology 2022, 13, 924081. [CrossRef]
- Yi D., Hou Y. Q., Wang L.., Ouyang W. J., Long M. H., Zhao D., Ding B. Y., Liu Y. L., Wu G. Y. L-Glutamine enhances enterocyte growth via activation of the mTOR signaling pathway independently of AMPK. Amino acids 2015, 47, 65–78. [CrossRef] [PubMed]
- Livak K. J., Schmittgen T. D.. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-△△Ct Method. Methods 2001, 25, 402–8. [CrossRef]
- Tang J., Xu Lq., Zeng Y. W., Gong F. Effect of gut microbiota on LPS-induced acute lung injury by regulating the TLR4/NF-kB signaling pathway. International immunopharmacology 2021, 91, 107272. [CrossRef]
- Ruan D., Wu S. W., Fouad A. M., Zhu Y. W., Huang W. J., Chen Z. L., Gou Z. Y., Wang Y. B., Han Y. Q., Yan S. J.., Zheng C. T., Jiang S. Q.. Curcumin alleviates LPS-induced intestinal homeostatic imbalance through reshaping gut microbiota structure and regulating group 3 innate lymphoid cells in chickens. Food function 2022, 13, 11811–24. [CrossRef]
- Kampman-van de Hoek E., Sakkas P., Gerrits W. J., van den Borne J. J., van der Peet-Schwering C. M., Jansman A. J. Induced lung inflammation and dietary protein supply affect nitrogen retention and amino acid metabolism in growing pigs. British journal of nutrition 2015, 113, 414–25. [CrossRef]
- Zhang W. H., Jiang Y., Zhu Q. F., Gao F., Dai S. F., Chen J., Zhou G. H. Sodium butyrate maintains growth performance by regulating the immune response in broiler chickens. British poultry science 2011, 52, 292–301. [CrossRef] [PubMed]
- Yang L., Liu G., Zhu X. Q., Luo Y., Shang Y. X., Gu X. L. The anti-inflammatory and antioxidant effects of leonurine hydrochloride after lipopolysaccharide challenge in broiler chicks. Poultry science 2019, 98, 1648–57. [CrossRef] [PubMed]
- Dai S. F., Bai X., Zhang D., Hu H., Wu X. Z.., Wen A. Y., He S. J., Zhao L. Dietary glutamine improves meat quality, skeletal muscle antioxidant capacity and glutamine metabolism in broilers under acute heat stress. Journal of applied animal research 2018, 46, 1412–7. [CrossRef]
- Wu Q. J., Wang Y. Q., Qi Y. X. Influence of procyanidin supplementation on the immune responses of broilers challenged with lipopolysaccharide. Animal science journal 2017, 88, 983–90. [CrossRef] [PubMed]
- Ramires C. C., Balbinot D. T., Cidral-Filho F. J., Dias D. V., Dos Santos A. R., da Silva M. D. Acupuncture reduces peripheral and brainstem cytokines in rats subjected to lipopolysaccharide-induced inflammation. Acupuncture in medicine 2021, 39, 376–84. [CrossRef] [PubMed]
- Yang L., Liu G., Liang X. R., Wang M. M., Zhu X. Q., Luo Y., Shang Y. X., Yang J. Q., Zhou P., Gu X. L. Effects of berberine on the growth performance, antioxidative capacity and immune response to lipopolysaccharide challenge in broilers. Animal science journal 2019, 90, 1229–38. [CrossRef] [PubMed]
- Achamrah N., Déchelotte P., Coëffier M. Glutamine and the regulation of intestinal permeability: from bench to bedside. Current opinion in clinical nutrition metabolic care 2017, 20, 86–91. [CrossRef] [PubMed]
- Zhang B. L., Lin M., Yu C. N., Li J. L., Zhang L., Zhou P., Yang W. W., Gao F., Zhou G. H. Alanyl-glutamine supplementation regulates mTOR and ubiquitin proteasome proteolysis signaling pathways in piglets. Nutrition 2016, 32, 1123–31. [CrossRef] [PubMed]
- Newsholme E. A., Parry-Billings M. Properties of glutamine release from muscle and its importance for the immune system. Journal of parenteral enteral nutrition 1990, 14, 63S–7S. [CrossRef]
- Cruzat V., Macedo Rogero M. M., Noel Keane K. N., Curi R. C., Newsholme P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10, 1564. [CrossRef]
- Labow B. I., Souba W. W., Abcouwer S. F.. Mechanisms governing the expression of the enzymes of glutamine metabolism--glutaminase and glutamine synthetase. Journal of nutrition 2001, 131, 2467S–74S. [CrossRef]
- Karinch A. M., Pan M., Lin C. M., Strange R., Souba W. W. Glutamine metabolism in sepsis and infection. Journal of Nutrition 2001, 131, 2531S–50S. [CrossRef]
- Xiao Y. P., Li X. Y., Wu T. X., Yang L., Hong Q. H., Yang C. M., Chen A. G. Effects of dietary glutamine supplementation on nutrient absorption and activity of enzymes involved in glutamine metabolism and energy production in the jejunum of weaned piglets. Journal of animal and vetrinary advances 2012, 11, 1441–9. [CrossRef]
- Cao Y., Liu S., Liu K. X., Abbasi I. H. R., Cai C., Yao J. Molecular mechanisms relating to amino acid regulation of protein synthesis. Nutrition research reviews 2019, 32, 183–91. [CrossRef] [PubMed]
- Frost R. A., Lang C. H. mTOR signaling in skeletal muscle during sepsis and inflammation: where does it all go wrong? Physiology 2011, 26, 83–96. [CrossRef] [PubMed]
- Nicklin P., Bergman P., Zhang B. L., Triantafellow E., Wang H., Nyfeler B., Yang H. D., Hild M., Kung C., Wilson C.., Myer V. E., MacKeigan J. P., Porter J. A., Wang Y. K., Cantley L. C., Finan P. M., Murphy L. O.. Bidirectional transport of amino acids regulates mTOR and autophagy. Cell 2009, 136, 521–34. [CrossRef]
- Xi P. B., Jiang Z. Y., Dai Z. L., Li X. L., Yao K., Zheng C. T., Lin Y. C., Wang J. J., Wu G. Y. Regulation of protein turnover by L-glutamine in porcine intestinal epithelial cells. Journal of nutritional biochemistry 2012, 23, 1012–7. [CrossRef] [PubMed]
- Lecker S. H., Goldberg A. L., Mitch W. E. Protein degradation by the ubiquitin-proteasome pathway in normal and disease states. Journal of the American Society of Nephrology 2006, 17, 1807–19. [CrossRef]
- Franch H. A., Price S. R. Molecular signaling pathways regulating muscle proteolysis during atrophy. Current opinion in clinical nutrition metabolic care 2005, 8, 271–5. [CrossRef]
- Crossland H., Constantin-Teodosiu D., Gardiner S. M., Constantin D., Greenhaff P. L. A potential role for Akt/FOXO signalling in both protein loss and the impairment of muscle carbohydrate oxidation during sepsis in rodent skeletal muscle. Journal of physiology 2008, 586, 5589–600. [CrossRef] [PubMed]
- Doyle A., Zhang G.., Abdel Fattah E. A., Eissa N. T., Li Y. P. Toll-like receptor 4 mediates lipopolysaccharide-induced muscle catabolism via coordinate activation of ubiquitin-proteasome and autophagy-lysosome pathways. FASEB 2011, 25, 99–110. [CrossRef]
- Crossland H., Constantin-Teodosiu D., Greenhaff P. L., Gardiner S. M. Low-dose dexamethasone prevents endotoxaemia-induced muscle protein loss and impairment of carbohydrate oxidation in rat skeletal muscle. Journal of physiology 2010, 588, 1333–47. [CrossRef] [PubMed]
- Tan H. W. S.., Sim A. Y. L., Long Y. C. Glutamine metabolism regulates autophagy-dependent mTORC1 reactivation during amino acid starvation. Nature communications 2017, 8, 338. [CrossRef] [PubMed]
- Lambertucci A. C.., Lambertucci R. H., Hirabara S. M., Curi R., Moriscot A. S., Alba-Loureiro T. C., Guimarães-Ferreira L., Levada-Pires A. C., Vasconcelos D. A., Sellitti D. F., Pithon-Curi T. C. Glutamine supplementation stimulates protein-synthetic and inhibits protein-degradative signaling pathways in skeletal muscle of diabetic rats. PloS one 2012, 7, e50390. [CrossRef]
Figure 1.
Effects of glutamine supplementation on the mRNA expression of TNF-α (A), IL-6 (B) and IL-1β (C) in liver of LPS-challenged broilers.Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means ± SEM, n=10. Different superscripts above the column means differences, P<0.05.
Figure 1.
Effects of glutamine supplementation on the mRNA expression of TNF-α (A), IL-6 (B) and IL-1β (C) in liver of LPS-challenged broilers.Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means ± SEM, n=10. Different superscripts above the column means differences, P<0.05.
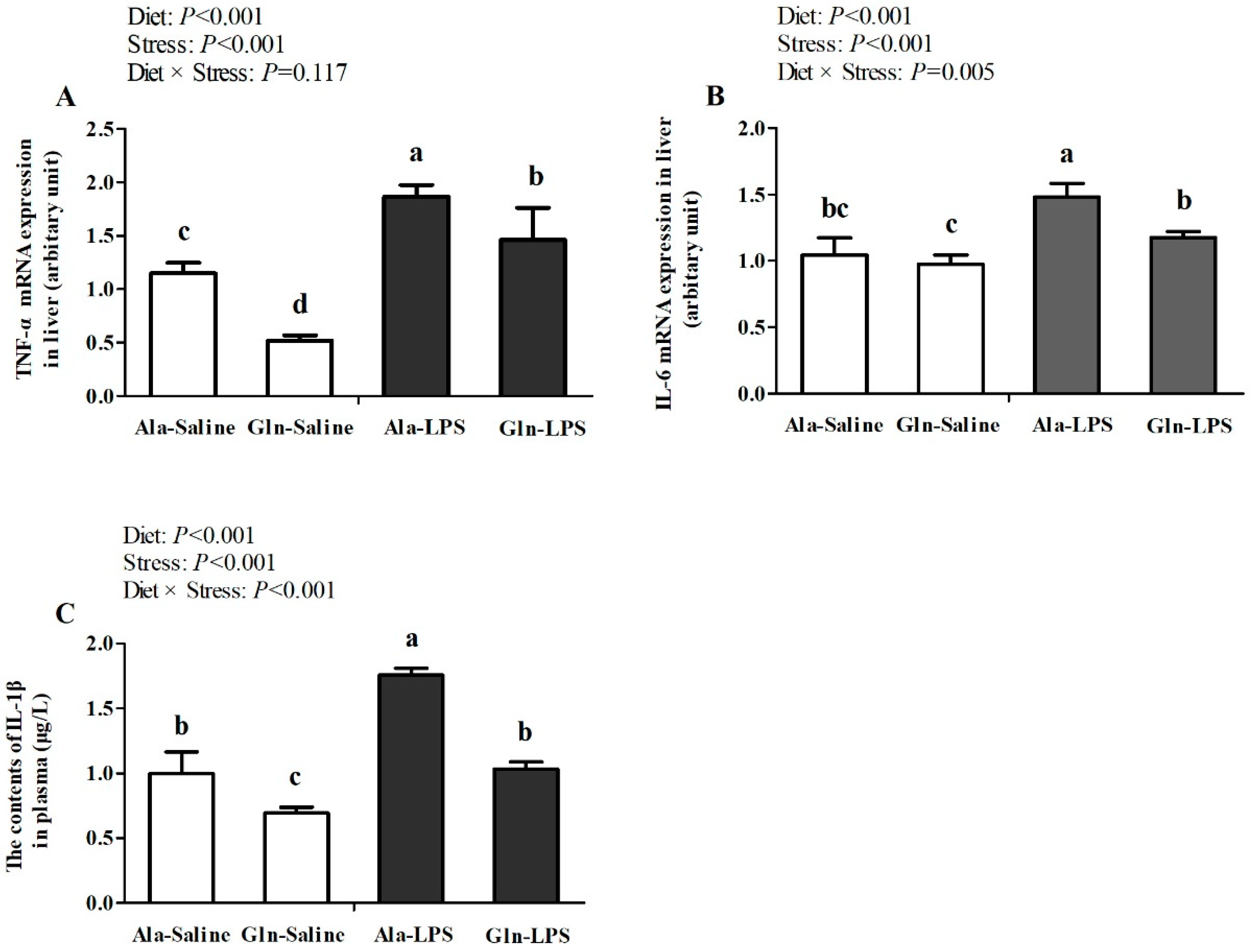
Figure 2.
Effects of Gln supplementation on the GS activity (A) and GA mRNA expression (B) in muscle of LPS-challenged broilers.
Figure 2.
Effects of Gln supplementation on the GS activity (A) and GA mRNA expression (B) in muscle of LPS-challenged broilers.
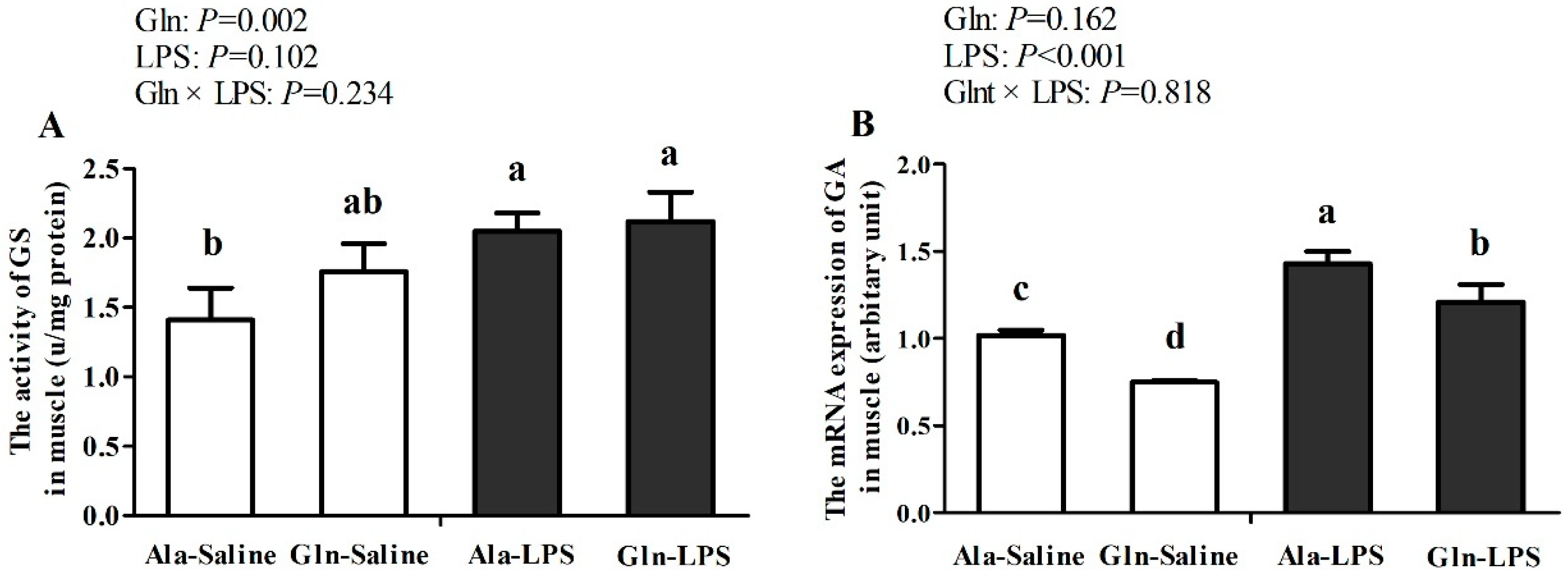
Figure 3.
Effects of Gln supplementation on mRNA expressions of mTOR (A), 4E-BP1 (B) and S6K1 (C) in muscle of LPS-challenged broilers.
Figure 3.
Effects of Gln supplementation on mRNA expressions of mTOR (A), 4E-BP1 (B) and S6K1 (C) in muscle of LPS-challenged broilers.
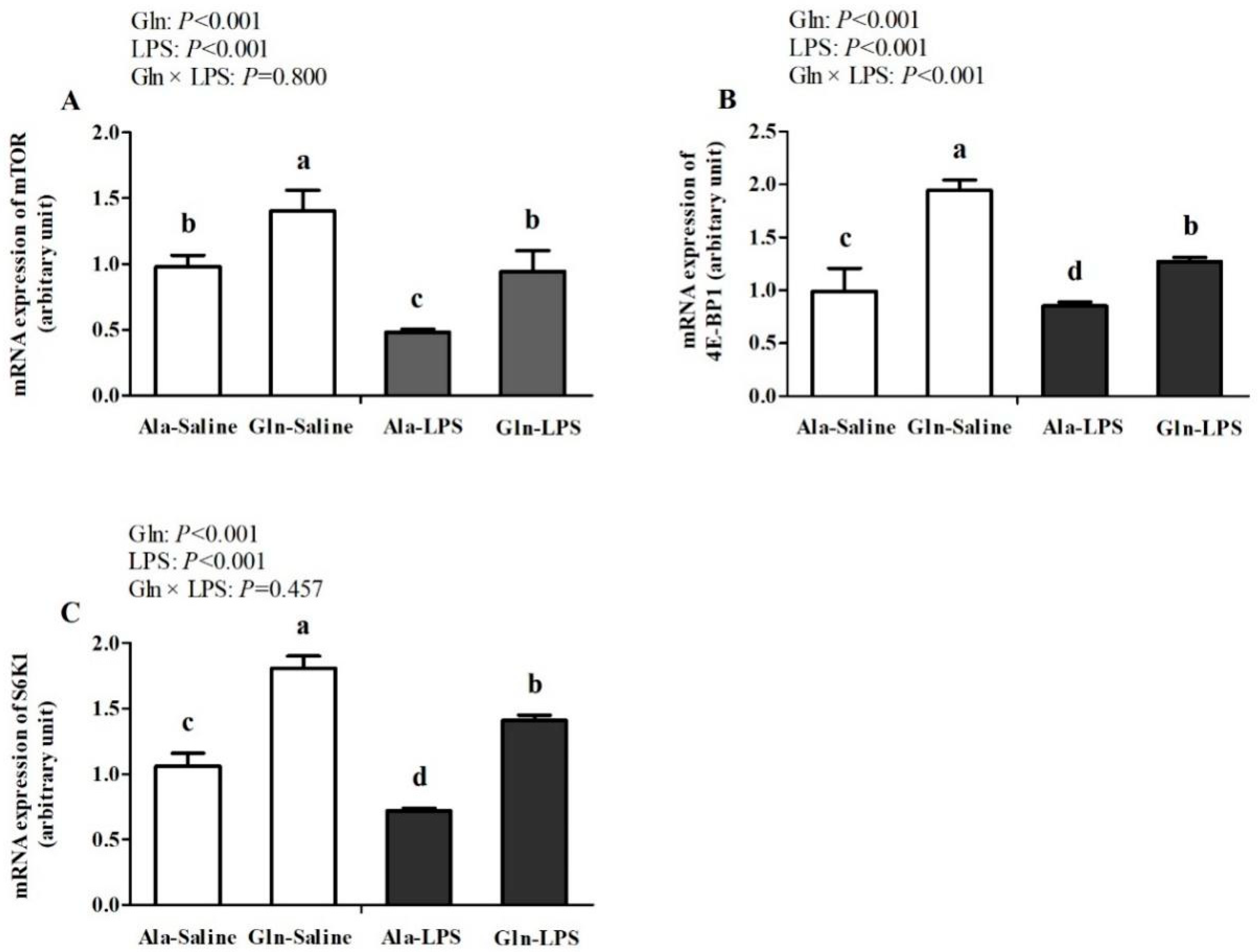

Table 1.
Ingredients and nutrient content of the basal diets.
| Ingredients (g/kg diet) | Nutrient content (g/kg diet) | ||
|---|---|---|---|
| Maize | 563.0 | Crude protein‡ | 210.8 |
| Wheat bran | 51.30 | Metabolism energy(MJ/kg) | 121.2 |
| Soybean meal | 285.0 | Calcium(%) | 10.00 |
| Corn gluten meal | 43.0 | Phosphorus(%) | 4.50 |
| DL-Methionine | 1.50 | DL-Methionine(%) | 8.60 |
| Phytase | 0.40 | L-Lysine(%) | 10.60 |
| Choline | 1.50 | Threonine(%) | 8.0 |
| Dicalcium phosphate | 18.70 | ||
| Limestone | 12.60 | ||
| Salt | 1.50 | ||
| Soybean oil | 16.50 | ||
| Vitamin and mineral premix† | 5.00 |
†Premix per kg diet provided: Vitamin A 12 000IU; Vitamin D3 2 500IU; Vitamin E 30 mg; menadione 2.8 mg; thiamin 2.21 mg; riboflavin 7.8 mg; nicotinamide 40 mg; Calcium pantothenate 10 mg; pyridoxine·HCl 4 mg; biotin 0.04 mg; folic acid 1.2 mg; Vitamin B12 0.015 mg; Fe 80 mg; Cu 8 mg; Mn 110 mg; Zn 65 mg; I 0.35 mg; Se 0.3 mg. ‡Nutrient contenst of the diets were the value of measurement.
Table 2.
Primer sequences for RT-PCR analysis.
| Genes | ID | Primers sequences(5’-3’) | Product size (bp) |
|---|---|---|---|
| MAFbx | NM_001030956.1 | F: GCCAGTACCACTTCACAGACAGAC R: GCGTGTCACCATACTGCTCCTTC |
132 |
| MuRF1 | XM_424369.4 | F: GAACGACCGCATCCAGACCATC R: TCCGTCTTCTTCTCCTCCAGCAG |
138 |
| FOXO1 | NM_204328.1 | F: GACCTCATCACCAAGGCCATCG R: GCACGCTCTTGACCATCCACTA |
85 |
| Akt | NM_205055.1 | F: GGCTACAAGGAACGACCGCAAG R: TACTGTGGTCCACTGGAGGCATC |
141 |
| TLR4 | NM_001030693.1 | F: TTCGGTTGGTGGACCTGAATCTTG R: ACAGCTTCTCAGCAGGCAATTCC |
114 |
| GA | NM_001031248.1 | F: TCCTCGCAGAGAAGGTGGTGATC R: TACGTGCAATGCTGTTCGTGAGTC |
154 |
| S6K1 | NM_001030721.1 | F: GTTCAGGCTCACCCGTTCTTCAG R: TGGCTCACATCCTCTTCAGATTGC |
107 |
| FOXO4 | XM_015278657.2 | F: CAACGTTCCACCACCCGTGA R: TGGAGGCAGATTGCTGGGTA |
101 |
| TNF-α | NM_204267.1 | F: TGTGTATGTGCAGCAACCCG R: AACAACCAGCTATGCACCCC |
178 |
| mTOR | XM_417614.6 | F: AACCACTGCTCGCCACAATGC R: CATAGGATCGCCACACGGATTAGC |
120 |
| 4E-BP1 | XM_424384.6 | F: GACCGTAAGTTCCTGATGGAGTGC R: ATTGGGCTGGTAACACCTGGAATG |
92 |
| IL-1β | NM_204524.1 | F: AAGCCTCGCCTGGATTCTAG R: TCAGGTCGCTGTCAGCAAAG |
90 |
| IL-6 | NM_204628.1 | F: TCCCTCCTCGCCAATCTGAA R: AAATAGCGAACGGCCCTCAC |
80 |
| β-actin | NM_205518.1 | F: ATTGTCCACCGCAAATGCTTC R: AAATAAAGCCATGCCAATCTCGTC |
113 |
MAFbx, muscle atrophy F-box; MuRF1, muscle Ring finger 1; FOXO1, Forkhead Box O1; Akt, protein kinase B, also named PKB; TLR4, Toll-like receptor 4; GA, glutaminase; S6K1, ribosomal protein S6 kinase; FOXO4, Forkhead Box O4; TNF-α: Tumor necrosis factor-α; mTOR, mammalian target of rapamycin; 4E-BP1, eIF-4E binding protein-1; IL-1β: Interleukin-1β; IL-6: Interleukin-6.
Table 3.
Effects of Gln supplementation on the activities of ALT and AST in muscle of LPS-challenged broilers.
Table 3.
Effects of Gln supplementation on the activities of ALT and AST in muscle of LPS-challenged broilers.
| Treatment | ALT(u/g of protein) | AST(u/g of protein) |
|---|---|---|
| Ala-Saline | 2.93 | 22.97 |
| Ala-LPS | 2.43 | 16.49 |
| Gln-Saline | 3.05 | 27.79 |
| Gln-LPS | 2.68 | 24.63 |
| SEM Main Effect Diet Ala Gln Stress Saline LPS P Value |
0.542 2.68 2.87 2.99 2.56 |
4.587 19.73b 26.21a 25.38a 20.56b |
| Gln | 0.603 | <0.001 |
| LPS | 0.236 | 0.002 |
| Gln × LPS | 0.848 | 0.160 |
ALT, alanine aminotransferase; AST, aspartate aminotransferase; Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means and SEM, n=10. Different superscripts in an array means differences, P<0.05.
Table 4.
Effects of Gln supplementation on mRNA expressions of protein degradation related genes in muscle of LPS-challenged broilers.
Table 4.
Effects of Gln supplementation on mRNA expressions of protein degradation related genes in muscle of LPS-challenged broilers.
| Treatment | TLR4 | Akt | FOXO4 | FOXO1 | MAFbx | MuRF1 |
|---|---|---|---|---|---|---|
| Ala-Saline | 1.00c | 1.00 | 0.99 | 1.04b | 1.0 | 1.07c |
| Ala-LPS | 1.43a | 0.59 | 1.42 | 2.93a | 1.4 | 2.81a |
| Gln-Saline | 0.86d | 1.39 | 0.92 | 0.94b | 0.69 | 1.03c |
| Gln-LPS | 1.01b | 0.96 | 1.11 | 1.28b | 1.14 | 1.47b |
| SEM Main Effect Diet Ala Gln Stress Saline LPS P Value |
0.234 1.22a 0.94b 0.93b 1.22a |
0.301 0.80b 1.18a 1.20a 0.78b |
0.249 1.21a 1.02b 0.96b 1.27a |
0.868 1.99a 1.11b 0.99b 2.11a |
0.292 1.29a 0.92b 0.89b 1.32a |
0.762 1.94a 1.25b 1.05b 2.14a |
| Gln | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| LPS | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Gln × LPS | 0.002 | 0.932 | 0.091 | <0.001 | 0.266 | <0.001 |
TLR4: Toll-like receptors 4; Akt: protein kinase B; FOXO4: Forkhead Box O 4; FOXO1: Forkhead Box O 1; MAFbx: Muscle atrophy F-box; MuRF1: Muscle Ring finger 1; Ala, alanine; Gln, glutamine; LPS, lipopolysaccharide. Values are means ± SEM, n=10. Different superscripts in an array means differences, P<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



