
Submitted:
06 December 2023
Posted:
06 December 2023
You are already at the latest version
Abstract
An in-depth understanding of the Se uptake and metabolism in plants is necessary for developing Se biofortification strategies. Thus, hydroponic experiments were conducted to investigate the associated processes and mechanisms of organic Se [selenomethionine (SeMet) and selenomethionine-oxide (SeOMet)] uptake, translocation, transformation and their interaction in wheat, in comparison to inorganic Se. Results showed that Se uptake by roots and the root-to-shoot translocation factor under the SeMet treatment were higher than those under the selenite, selenate, and SeOMet treatments. The uptake and translocation of SeMet were higher than those of SeOMet within 72 h, although the differences gradually narrowed with time. The uptake of SeMet and SeOMet was also sensitive to the aquaporin inhibitor: AgNO3 addition resulted in 99.5% and 99.9% inhibition of Se in root in the SeMet and SeOMet treatments, respectively. Once absorbed by root, were rapidly assimilated to other Se forms, and SeMet and Se-methyl-selenocysteine (MeSeCys) were the dominant species in SeMet and SeOMet-treated plants, while notably, an unidentified Se form was also found in the root and xylem sap under the SeMet treatment. In addition, within 16 h, SeOMet inhibited the uptake and translocation of SeMet, while the interaction was weakened with longer treatment time. Taken together, the present study provides new insights for the uptake and transformation processes of organic Se within plants.
Keywords:
wheat
; selenomethionine
; selenomethionine-oxide
; uptake
; Se speciation
; interaction
1. Introduction
Selenium (Se) is an essential micronutrient for human and animal health [1], associated with multiple physiological functions such as antioxidation, anti-carcinogenesis and immunoregulation, while a deficiency in Se can lead to the risk of cancer and cardiovascular diseases [2,3]. The World Health Organization recommends a daily Se intake of 50–200 μg d-1 for adults, as the optimum level of Se in living organisms is quite narrow in range. However, due to the significant variances in soil Se contents worldwide and the distribution of Se-poor soils in some populous regions, Se deficiency has become a public health problem, and approximately one billion people suffer from insufficient Se intake [4,5]. Food fortification strategies should be developed and recommended to increase Se content in the human diet, especially in low-Se areas. Agronomic biofortification with Se-containing fertilizers supplemented in soil or substrate is the most effective way to increase Se levels, and it has also been widely applied [6,7,8].
The Se accumulation in plants mainly depends on the amount and availability of Se in soil, which is associated with the different chemical forms of Se, and the plant species [9]. Se naturally occurs as selenide, elemental Se, selenite, selenate and organic Se, and their distributions are governed by soil conditions, including pH, Eh, Fe/Mn/Al oxides, adsorbing surfaces, organic matter and microbial activity [10]. The uptake and biotransfomation mechanisms of selenate and selenite have been well established. Selenate is transported through the cell membrane into roots via sulfate transporters [11]. And it continued to be transported in the form of Se (VI) via the xylem to the shoots [12]. Nevertheless, selenite is taken up by root via phosphate and silicon transporters [13,14]. Then, it can easily be converted into organic Se and rarely transported to shoots [15,16]. Organic forms of Se, such as selenomethionine (SeMet), and selenomethionine Se-oxide (SeOMet), also make up an important fraction of available Se in soil, and absorbed by plant root readily [9,17,18]. However, the uptake and biotransformation mechanisms of organic Se in plants remain to be experimentally confirmed.
Previous reports consistently revealed that the uptake efficiency of SeMet was much higher than that of selenate, selenite and SeOMet [19,20,21]. Our latest result revealed that SeMet uptake was markedly inhibited by the aquaporin inhibitor (AgNO3), indicating that rice absorbs SeMet primarily via aquaporin. In comparison, SeOMet, a derivative obtained from the transformation of SeMet, it is taken up by an energy-dependent symport process and imported by rice roots via aquaporin and K+ channel When simultaneously supplied with both SeOMet and SeMet, SeOMet appeared to inhibit the uptake and translocation of SeMet [22]. Moreover, a recent report revealed that NRT1.1B was involved in SeMet translocation from root to shoot [23]. Taken together, the above researches about organic Se are primarily concentrated on rice, but the mechanism and application of organic Se in wheat is poorly understood.
In contrast with rice, wheat is a aerobic plant with significantly different structure of stem and root. Meanwhile, wheat can produce high-security organic Se by absorbing Se in soil or foliage and store them in grains, thus provides the major source of Se in dietary intake [24,25]. In this study, assimilation of SeMet and SeOMet by wheat in terms of uptake, translocation, biotransformation were reported through a series of hydroponic experiments, and the main purpose of the study was to investigate (1) the difference between the uptake and translocation of organic Se (SeMet and SeOMet) and inorganic Se (selenite and selenate) in wheat plants, (2) the distribution and speciation of Se in plants treated with organic Se, (3) the interaction between SeMet and SeOMet during the uptake and transformation processes, and provide a scientific basis for practical Se biofortification and some help for the application and development of Se resources.
2. Materials and Methods
2.1. Plant Culture and Experimental Condition
Seeds of wheat (Triticum aesticum L. cv. Luyuan502, a semi-winter middle-late cultivar) were surface-sterilized in 10% (v: v) NaClO solution for 15 min, rinsed thoroughly with deionized water and impregnated into saturated CaSO4 solution overnight, and then germinated on the pre-sterilized floating PVC screen in the solution of 0.5 mmol L−1 CaCl2. Seven days after germination, every four wheat seedlings were transplanted to a plastic pot containing 3 L 1/5 Hoagland nutrient solution. Its nutrient concentration was as follows (mmol L−1): 1.0 KNO3, 0.1 KH2PO4, 0.457 MgSO4·7H2O, 1.0 Ca(NO3)2·4H2O, 1×10−3 ZnSO4·7H2O, 0.060 Fe(III)-EDTA, 3×10−3 H3BO3, 0.2×10−3 CuSO4·5H2O, 1×10−3 (NH4)6Mo7O24·4H2O, 1×10−3 MnSO4·H2O. The pH of this solution was adjusted to 6.0 with 2 mmol L−1 MES (2-morpholinoethanesulphonic acid, pH was adjusted with 1 mmol L−1 KOH and HCl). The solution was aerated continuously and renewed every three days, throughout the experiment. Plants were grown in a greenhouse with controlled conditions as follows: 20℃/15℃ day/night temperatures, 14 h d−1 photoperiod with a light intensity of 240-350 μmol m−2 s−1 , and 60% of relative humidity . Seeds of wheat were provided by Shandong Academy of Agricultural Sciences, China.
2.2. Comparison of Uptake and Translocation Between Organic Se and Inorganic Se in Wheat Weedlings
Four-week-old wheat seedlings were transferred to pots (one plant per pot) containing 1 L of aerated uptake solutions, which composed of normal nutrients and different species of Se (5 μmol), including selenite, selenate, SeMet and SeOMet, respectively. Each treatment had three replicates. The pH of these uptake solutions was buffered to 6.0 with 2 mmol MES. After treated for 24 h, the seedling roots were rinsed with deionized water for three times, and then transferred to 150 mL ice-cold desorption solutions (1 mmol L−1 CaSO4 + 1 mmol L−1 K2HPO4, 2 mmol L−1 MES, pH 6.0) for 15 min to remove the Se and other ions adsorbed on the root surface. After desorption, the wheat plants were rinsed thoroughly with deionized water, and then separated into shoots and roots, weighed, frozen in liquid nitrogen, powdered, and analyzed for total Se contents. Selenite, selenate and Selenomethionine (SeMet) were obtained from Sigma-Aldrich (St Louis, MO, USA). Selenomethionine-oxide (SeOMet) was prepared according to Larsen et al. [26].
2.3. Effects of Aquaporin Inhibition on Uptake of SeMet and SeOMet
The aquaporin inhibition assay was performed according to the reported method with slight modification [27]. Four-week-old seedlings were transferred to absorption solutions containing 5 μmol L−1 organic Se (SeMet or SeOMet) and 0 or 100 μmol L−1AgNO3 (2 mmol L−1 MES, pH 6.0), and thus, there were four treatments in total:SeMet alone, SeMet+AgNO3, SeOMet alone, and SeOMet+AgNO3. Three replicates were used for each treatment. After 24 h, wheat roots were rinsed with deionized water and the Se adsorbed on the root surface was desorbed as described earlier. Then shoots and roots were separated and their total Se contents were analyzed.
2.4. Uptake, transformation and interaction of SeMet and SeOMet in wheat seedlings at different times
This experiment was conducted to investigate the transformation regulation of Se speciation , and whether SeMet and SeOMet interact during uptake and assimilation by wheat plants. Four-week-old wheat seedlings were exposed to 1 L pots (one plant per pot) containing aerated uptake solutions for 4, 16, 32, and 72 h, respectively. The uptake solutions were composed of normal nutrients and 10 μmol L−1 SeMet, 10 μmol L−1 SeOMet or 10 μmol L−1 SeMet + 10 μmol L−1 SeOMet (2 mmol L−1 MES, pH 6.0). Each treatment was replicated in three pots. After the organic Se absorption, the roots were rinsed with deionized water and desorbed as described above. The fresh shoots and roots were weighed, frozen in liquid nitrogen, powdered, and stored at –80 °C for subsequent determine of total Se and Se speciation. In addition, when treated for 16 h, wheat shoots were cut at 2 cm above the roots, and the xylem saps were collected using a 1-mL pipette for 2 h. After collection, the xylem saps were stored at –80 °C for the analysis of Se speciation.
2.5. Analysis of Total Se and Se Speciation in Plant Tissues
For the analysis of total Se content, powdered plant samples were digested with HNO3 (GR) using microwave sample preparation system (CEM, MARS5, CEM Corp., Matthews, NC, USA). Total Se content in wheat tissues were determined based on the method of Fujii et al. [28].During the whole analysis process, a certified reference material (GBW10014) and blanks were included for quality assurance. The recovery for GBW10014 was 85%-110%.
Powdered fresh plant samples (0.4000 g) were extracted with 5 mL 8 mg mL−1 protease XIV (Sigma Aldrich, USA) in an oscillation box (37 °C, 125 rpm) for 24 h. The extracts were subsequently centrifuged at 12,000 rpm for 15 min, and then filtered through 0.22-μm filters (Millipore, Billerica, MA, USA). The xylem saps were also filtered through 0.22-μm filters. Selenium speciation in plant extracts and xylem sap was determined using HPLC-ICP-MS (Agilent LC1260 series and Agilent ICP-MS 7700, Agilent Technologies, Santa Clara, CA, USA).
Selenium species were separated by an anion exchange chromatography column (Hamilton PRP-X100), and the outlet of the separation column was connected to an ICP-MS detection system. The mobile phase was 40 mmol L−1(NH4)2HPO4 (pH 6.0). The Se standards, namely, Se (IV) (Na2SeO3), Se (VI) (Na2SeO4), SeMet (mono-methylarsonic acid), SeCys2 (selenocysteine) and MeSeCys (Se-methyl-selenocysteine), were obtained from the National Research Center for Certified Reference Materials, Beijing, China. SeOMet (Selenomethionine Se-oxide) was prepared as Larsen et al. [26]. The chemical species of Se in samples were identified and quantified by retention times and peak areas, respectively.
2.6. Data Analysis
Total Se concentration (TShoot, TRoot and TSe) and the transfer factor (TF) of Se were calculated using Equations (1–4):
where CShoot−Se and CRoot−Se represent the Se content in wheat shoot and root, respectively.
TShoot−Se = CShoot−Se × BiomassShoot
TRoot−Se = CRoot−Se × BiomassRoot
TSe = TShoot−Se + TRoot−Se
TF = CShoot−Se / CRoot−Se
The proportion of Se distribution in wheat tissues was calculated using Equations (5–6):
Shoot − Se% = TShoot−Se / TSe× 100%
Root − Se% = TRoot−Se / TSe× 100%
The proportion of each Se species (Proi) in wheat was calculated using Equation (7):
where Ci and CShoot/Root−Serepresent the content of a certain Se species and the sum of the contents of the four Se species in the wheat tissues, respectively.
Proi = Ci / × 100%
All data are presented as means ± standard errors (SEs; n = 3). Analysis of variance was performed using SAS 9.3 (least significant difference, p < 0.05).
3. Results
3.1. Uptake and Translocation of Different Se Treatments in Wheat Seedlings
To evaluate the uptake and translocation of organic Se versus inorganic Se, Se (IV), Se (VI), SeMet and SeOMet were supplied in this experiment. Results showed that Se contents in wheat shoots and roots were significantly affected by Se forms, and the level of Se in root was obviously higher than that in shoot (Figure 1). In wheat root, the Se content in SeMet treatment was 65.4%, 69.0% and 7.09 times higher than that in SeOMet, Se (IV) and Se (VI) treatments, respectively; while the differences in shoot were even higher (10.2-, 37.5- and 28.1 times).
The translocation and distribution of Se in wheat plants were also significantly affected by Se forms (Table 1). In the Se (IV) treatment, most of Se distributed to root, accounting for 94.0% of total Se uptake by wheat seedlings. In contrast, more the half of Se distributed to shoot (57.6%) in SeMet treatment. No significant difference was obtained between the Se (VI) and SeOMet treatments, and the former was slightly higher in the Se distribution in shoot. The translocation factor (TF) from root to shoot in SeMet treatment was 2.61-, 4.22- and 22.0 times higher that of the Se (VI), SeOMet and Se (IV) treatment, respectively.
3.2. Effects of aquaporin inhibition on uptake of SeMet and SeOMet
To examine the uptake of SeMet and SeOMet by wheat through water channels, 0.1 mmol L-1 AgNO3 was used as a water channel blocker [29]. The Se contents in wheat plants were measured after 24 h exposure in aquaporin inhibition and two organic Se forms. Generally, the Se contents in wheat root and shoot in SeMet treatment were higher than those in SeOMet treatment, and the former were 20.4% and 1.63 times higher than the latter, respectively, in the Se alone treatment. Compared with the control (Se alone treatment), AgNO3 addition dramatically reduced Se contents in root and shoot by 99.5% and 96.4% in SeMet treatment, respectively; while the differences in SeOMet treatment were 99.9% and 92.2%, respectively.
3.3. Uptake, translocation and interaction of SeMet and SeOMet in wheat seedlings in different time
The Se contents in wheat plants were significantly affected by Se forms, exposure time and the interactions between these two factors (Figure 3, p < 0.05). Irrespective of the Se treatments, root and shoot Se increased gradually with increasing exposure time. When the treatment time was ≤32 h, the Se in wheat root treated with SeMet was significantly higher than that treated with SeOMet, while the difference decreased with time, from 81.3% to 22.5% (p < 0.05); when treated for 72 h, there was no significant difference between the two Se forms (Figure 3a). Similarly, shoot Se content in SeMet treatment was higher than that in SeOMet treatment at all Se-treated time-points, while the difference gap narrowed with time (Figure 3b). When treated for 4 h, only a small proportion of Se was transferred to shoot, with the prolonged exposure time, the proportions of Se distributed to shoot were 18.1%-56.8%. Besides, the transfer factor and shoot-Se% in SeMet treatment were higher than those in SeOMet treatment at all exposure time (Figure 4).
In the experiment, both SeMet and SeOMet were applied to the nutrient solution to investigate the interaction on Se uptake and translocation. When the exposure time was ≤ 16 h, root and shoot Se in SeMet/SeOMet treatment were lower than those in single-SeMet treatment, by 3.0%-45.2% and 31.0%-95.5%, respectively; while higher than those when treated over 32 h (except for the root at 72 h). No significant differences of Se in plants were found between SeMet/SeOMet and single-SeOMet treatments at 4 h; and after 16 h, the former was higher than the latter (except for the root at 72 h) (Figure 3). The transfer factor and shoot-Se% of plants treated with SeMet/SeOMet were lower than those in single-SeMet treatment when the exposure time was ≤32 h, while higher than those at 72 h. And the transfer factor and shoot-Se% of mixed Se treatment were higher than those of single-SeOMet treatment at 32 h and 72 h (Figure 4).
3.4. Transformation of SeMet and SeOMet in Wheat Seedlings
Under the current working conditions, MeSeCys, Se (IV), SeMet and Se (VI) were ideally separated through HPLC-ICP-MS, and the retention times were 186 s, 274 s (RT274), 374 s (RT374) and 756 s (RT756), respectively. However, the retention times for SeOMet and SeCys2 were close at 141 s (RT141) and 144 s (RT144), respectively. Therefore, the standard curve for SeOMet was tested and plotted solely (Figure S1). Besides, an unidentified Se compound (the first peak on the chromatogram, RT121) was found in plants treated with SeMet and SeMet/SeOMet, especially in wheat root (Figure 5).
Overall, the proportions of SeMet, MeSeCys, Se (IV) and SeCys2 or SeOMet in the total selenium content in wheat roots were 10.3%~25.8%, 3.65%~21.7%, 0.10%-0.66%, 0.19%-1.44%, respectively. Similarly, the corresponding proportions in shoot were 31.1%-87.8%, 5.22%-11.4%, 0.86%-1.75% and 0.55%-2.19% respectively. In wheat roots, the contents of MeSeCys and SeMet in the single-SeMet treatment were 66.7% and 89.1% higher than those in the single-SeOMet treatment, respectively; while the Se (IV) content in the former was lower than that of latter, although not significantly. In addition, the contents of MeSeCys, Se (IV) and SeCys2 or SeOMet in SeMet/SeOMet treatment were higher than those in single-SeMet or single-SeOMet treatment: MeSeCys in SeMet/SeOMet treatment was 2.27- and 4.45- times higher than that in single-SeMet and single-SeOMet treatment, respectively (p < 0.05); while SeMet content in SeMet/SeOMet treatment was similar to that in single-SeMet treatment. In wheat shoot, the content of SeMet in single-SeMet treatment was 2.06 times higher than that in single-SeOMet treatment (p < 0.05); while the other Se forms [MeSeCys, Se (IV), SeCys2 or SeOMet] in the former was lower than those of latter. Under SeMet/SeOMet treatment, Se forms of MeSeCys and Se (IV) were higher than those in single-SeMet or single-SeOMet treatment; while SeMet and SeOMet or SeCys2 were similar to those in single-SeMet treatment.
3.5. Se Concentration and Speciation in Xylem Saps
After 16 h of exposure, Se speciation in the xylem sap was determined, and the unidentified Se compound (the first peak on the chromatogram, RT121) was also found therein (Figure 6). In the SeMet alone treatment, MeSeCys, Se (IV), SeMet and the unknown Se form were detected, and the concentrations were as follows: unknown Se form>MeSeCys>Se (IV)>SeMet. When SeOMet added alone, however, only an unknown Se form was detected in xylem sap, which concentration was much lower than that of SeMet treatment. Under SeMet/SeOMet treatment, the Se forms in xylem sap were same as those under SeMet treatment, while the concentration of MeSeCys in the former was 90.3% lower than that in the latter (Table 3).
4. Discussion
4.1. Difference of The Uptake and Translocation between Organic Se and Inorganic Se in Wheat Seedlings
In this study, the uptake and translocation characteristics of organic Se (SeMet and SeOMet) by wheat plants were various from those of inorganic Se (selenite and selenate). The order of the SeMet uptake ability by wheat root was much higher than those of SeOMet, selenite and selenate (Figure 1 and Table 1). Huang et al. observed that rice roots could absorb more selenite than selenate applied at the same dosage [30]. Some reports have indicated that the uptake of SeMet by maize, wheat, and rice roots was much faster than that of selenite or selenate [31,32,33]. The differences in the uptake could be attributed to differences in the activities of their respective transporters, none of which, however, are Se-specific. Selenate is taken up by roots via sulfate transporters [11], and selenite is absorbed in an active process mediated by phosphate and silicon transporters [13,14,15]. On the other hand, SeMet, as a seleno amino acid, might be absorbed via root amino acid transporters [24,34]. Our study showed that the uptake of SeMet and SeOMet was sensitive to the aquaporin inhibitor, such as AgNO3 that is reported to partially inhibit the uptake of selenite and Se nanoparticles in crops [27,35,36]. In this study, the AgNO3 addition resulted in 99.5% and 99.9% inhibition of Se in root in SeMet and SeOMet treatment, respectively (Figure 2), indicating that the influx of SeMet and SeOMet might be mediated via aquaporins. The phenomenon that plant root could absorb more organic Se than inorganic Se might be attributed to the following mechanisms: (1) transporters mediated SeMet and SeOMet intake are more active than those of inorganic Se [19]; (2) the uptake of inorganic forms might be inhibited by sulfate and phosphate in the culture solution [15,37]; (3) inorganic forms such as selenate and selenite are more toxic to plants than SeMet and SeOMet, and resulting in less intake due to the defensive responses of plants [32,38].
In rice seedlings exposed to different sources of Se for 24 h, we found that the proportions of Se distributed in the wheat shoot (and transfer factor from root to shoot) decreased in the order SeMet > selenate > SeOMet > selenite (Table 1). Sulfate transporters (such as Sultr2;1, Sultr3;5 and Sultr1;3) and phosphorus transporters (such as OsPT8) are the main transporters involved in the translocation of selenate and selenite from root to shoot, respectively [11,39,40]. In addition, Zhang et al. found that NRT1.1B, which is associated with nitrate uptake and transport, mediates the transport activity of SeMet in rice [23]. Upon uptake, most selenite is quickly metabolized into organic Se compounds and retained in the roots, whereas selenate can be translocated to shoots rapidly [15,41,42], thus the transport capacity of Se in plants treated with selenate was greater than that treated with selenite [30,43,44]. In the study of Xu et al., the translocation of Se from rice root to shoot in SeMet treatment was higher than that in the selenate or selenite treatment [33]; and Kowalska et al. reported that the TF of Se in lettuce treated with SeMet was 3.56 times higher than that treated with selenite, which were consistent with our study [20]. However, in another study, Se in shoot originating from SeMet was higher than that originating from selenate in garlic (Allium sativum), while tended to be lower than that originating from selenate in Indian mustard (Brassica juncea) [45]. Wang et al. reported that the order of Se-shoot% in maize seedling was selenate treatment> selenite treatment> SeMet treatment, and selenate treatment> SeMet treatment> selenite treatment when 0.01 mg L-1 or 0.1 mg L-1 Se supplied, respectively [32]. Therefore, the transport capacity of in plants not only depends on Se forms, but also on plant species and Se concentrations.
As for the two organic Se forms, the uptake and transport of SeMet were greater than those of SeOMet within 72 h (Figure 1, Figure 2 and Figure 3), while the difference gradually narrowed with time (Figure 3). It’s probably because the uptake of SeMet is gradually approaching to saturation during the treatment time, and the uptake rate is decreasing. Besides, the effect of exposure time on organic Se absorption might be attributed to the transformation between SeOMet and SeMet in plant root or rhizosphere. It has been demonstrated that SeOMet was detected in the root of lettuce (Lactuca sativa L.) exposed to SeMet [20]. And in this study, SeMet was detected in wheat root in SeOMet, while SeOMet or SeCys2 was also detected in root treated by SeMet (Figure 5), thereby indicating the occurrence of the oxidative and reductive transformation of Se in plants.
4.2. Transformation of SeMet and SeOMet in Wheat Seedlings
In the present study, MeSeCys, Se (IV), SeMet and Se (VI) were quantitatively and qualitatively measured by HPLC-ICP-MS; while SeCys2 and SeOMet cannot be identified clearly as the retention times of them overlapped (Figure S1), which was also found in previous studies [46,47]. Although it was hard to identify, the chromatograms showed that the contents of these two Se forms were low in wheat plant (Figure 5 and Figure 6). In addition, there was a difference between the sum of the identified peaks and the total Se in plant (Table 2), which might be due to the limitation of standard Se compounds (unknown compound) and the low extraction efficiency of protease XIV.
Plants accumulate Se in different chemical forms, and the speciation of Se in plants depends on the plant species and the Se forms in their surroundings [48]. It has been proved that plants can absorb inorganic Se (i.e., selenate and selenite) and nano-Se, and convert to organic Se [26,35,49]; for example, SeMet, MeSeCys and SeCys2 were all detected in wheat root and shoot treated with selenite [35]. As for organic Se, SeMet and MeSeCys were the dominant forms in wheat in SeMet or SeOMet treatment in our study (Figure 5 and Table 2). SeMet and MeSeCys are well assimilated by humans and animals and advocated to be used for nutritional selenium supplement [50]. Furthermore, it has been reported that MeSeCys has anticarcino-genic and antitumor activities [51], and the results showed that the pharmaceutically valuable MeSeCys can be efficiently biosynthesized in plants fortified with SeMet (or SeOMet).
In protein synthesis in plants, SeMet can non-specifically replace Met to form Se-containing proteins, as the methionine-tRNA cannot distinguish between Met and SeMet [52,53]. During Se assimilation, a variety of intermediates are produced by organisms, such as selenohomocysteine (SeHCys), selenocystathionine (SeCysTH) Se-adenosyl-selenomethionine (SeAM) [9,54]. As shown in Figure 5 and Figure 6, an obvious peak (RT121) corresponding to an unknown Se compound was detected in wheat root and xylem sap in SeMet or SeMet/SeOMet treatment, but not in SeOMet treatment (except for xylem sap). Therefore, we speculated that SeMet transformed into an intermediate after absorbed by root, and the intermediate could be both easily transformed into other forms [such as MeSeCys and Se (IV)] in root and transfer to shoot, and then converted to other Se forms in shoot. As for SeOMet, however, once absorbed by root, it is rapidly converted to SeMet, which become the main rate-limiting step of Se assimilation. The underlying mechanisms need further investigation. Similarly, we also found the aforementioned unknown Se species in plant root treated with selenite in our previous studies [35,36]. Due to the technical condition restriction, the possible molecular formula of this unknown species needs to be determined using a higher sensitive instrument in the future, such as UHPLC-ESI-Orbitrap MS [19].
4.3. Interaction between SeMet and SeOMet in Wheat Seedlings
Results showed that Se contents in wheat root and shoot treated with SeMet/SeOMet were lower than the single SeMet treatment (within 16 h) (Figure 3 and Figure 4), and at 16 h, the MeSeCys content in SeMet/SeOMet treatment was lower than that in SeMet treatment (Figure 6), indicating an interaction between SeMet and SeOMet occurred. Previous studies have reported non-additive effects in the uptake and translocation of different Se forms, for example, a certain interaction exited between selenate and selenite. In previous studies, selenite inhibited the uptake of selenate by wheat, when both Se forms were supplied [15,43]; consistent phenomenon was also observed in tomato (S. lycopersicum L.) [12]. Similarly, in the present study, we found that the presence of SeOMet appeared to suppress the accumulation of SeMet in wheat. The interactions between different Se forms might be caused by the specific absorption of plants: an optimal absorption scheme is adopted based on intrinsic synergistic action in response to mixed supplement of different Se forms, thus conserving the enzymes and energy required for subsequent Se assimilation [55].
In addition, the interactions between SeMet and SeOMet in wheat depended on the exposure time. Within 16 h treatment, SeOMet inhibited the uptake and translocation of SeMet; while the treatment time was prolonged (32 and 72 h), the interaction was weakened (Figure 3, Figure 4, Figure 5 and Figure 6). This might be explained by the metabolism of Se in plant: in short time (≤ 16 h), the saturation threshold of the corresponding enzyme was not reached, so the rate of SeMet uptake was high; in this case, the SeOMet addition would have a negative effect on root SeMet absorption. After 32 h, the SeMet uptake rate was gradually declined; and the transformation between SeMet and SeOMet in plant root or rhizosphere might also account for the decrease of interaction. Furthermore, at 32 h, the Se content in root treated with SeMet/SeOMet was significantly higher than that of SeMet treatment; while at 72 h, Se in shoot of the former was higher than the latter (Figure 3). As we know, Se has a dual effect on plant, excessive accumulation of Se would disrupt the structure and function, and cause cytotoxicity, thus organisms might alleviate this damage by accumulate more Se in root and regulating Se speciation [56]. Structural Se-containing amino acids (such as SeMet and SeCys) will rapidly convert to non-structural amino acids (such as MeSeCys), thereby improving plant tolerance [57]. The higher content of MeSeCys in SeMet/SeOMet treatment also proved this, to a certain degree (Figure 5). However, this needs to be verified in further studies.
5. Conclusions
The present study has revealed differences between SeMet and SeOMet with respect to Se uptake, translocation and speciation in wheat seedlings. Compared with SeOMet and inorganic Se forms (selenite and selenate), SeMet exhibited a greater uptake and transport capacity, though the differences between the two organic Se forms (SeMet and SeOMet) narrowed with time. And the influx of SeMet and SeOMet might be mediated via aquaporins. Speciation analysis showed that SeMet and MeSeCys were the dominant species in all wheat plants, while an unidentified Se species was also found in the root and xylem sap of plants treated with SeMet. In addition, we also found that when simultaneously supplied with both SeOMet and SeMet, SeOMet appeared to inhibit the uptake and translocation of SeMet, while the interaction was weakened with time. We anticipated that the findings of this study would provide a theoretical basis for developing efficient Se-enriched foods and resolving Se-deficiency in humans and animals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Additional Supporting Information may be found in the online version of this article, Figure S1. Analytical HPLC-ICP-MS chromatogram of six standard Se species (100 µg L-1): (a) SeCys2, MeSeCys, Se (IV), SeMet and Se (VI); (b) SeOMet.
Funding
This work was supported by the National Natural Science Foundation of China (41907146) and the Special Fund for Agro-scientific Research in the Public Interest of China (201303106).
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [CrossRef]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Sign. 2011, 14, 1337–1383. [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [CrossRef]
- Haug, A.; Graham, R.D.; Christophersen, O.A; Lyons, G.H. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228.
- Lv, Y.Y; Yu, T.; Yang, Z.F.; Zhao, W.F.; Zhang, M.; Wang, Q. Constraint on selenium bioavailability caused by its geochemical behavior in typical KaschinBeck disease areas in Aba, Sichuan Province of China. Sci. Total Environ. 2014, 493, 737–749. [CrossRef]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminen, P.; Hietaniemi, V.; Aspila, P.; Aro, A. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: from deficiency to optimal selenium status of the population. J. Trace Elem. Med. Bio. 2015, 31, 142–147. [CrossRef]
- de Oliveira, A.P.; Naozuka, J.; Landero-Figueroa, J.Á. Effects of Se (IV) or Se (VI) enrichment on proteins and protein-bound Se distribution and Se bioaccessibility in oyster mushrooms. Food Chem. 2022, 383, 132582. [CrossRef]
- Galinha, C.; Sánchez-Martínez, M.; Pacheco, A.M.G.; Freitas, M.D.; Coutinho, J.; Maças, B.; Almeida, A.S.; Pérez-Corona, M.T.; Madrid, Y.; Wolterbeek, H.T. Characterization of selenium-enriched wheat by agronomic biofortification. J. Food Sci. Tech. Mys. 2015, 52, 4236–4245. [CrossRef]
- Trippe III, R.C.; Pilon-Smits, E.A.H. Selenium transport and metabolism in plants: phytoremediation and biofortification implications. J. Hazard. Mater. 2021, 404, 124178. [CrossRef]
- Neal, R.H.; Sposito, G.; Holtzclaw, K.; Traina, S. Selenite adsorption on alluvial soils: soil composition and pH effects. Soil Sci. Soc. Am. J. 1987, 51, 1161–1165. [CrossRef]
- El Mehdawi, A.F.; Jiang, Y.; Guignardi, Z.S.; Esmat, A.; Pilon, M.; Pilon-Smits, E.A.; Schiavon, M. Influence of sulfate supply on selenium uptake dynamics and expression of sulfate/selenate transporters in selenium hyperaccumulator and nonhyperaccumulator Brassicaceae. New Phytol. 2018, 217, 194–205. [CrossRef]
- Wang, M.K.; Peng, Q.; Zhou, F.; Yang, W.X.; Dinh, Q.T.; Liang, D.L. Uptake kinetics and interaction of selenium species in tomato (Solanum lycopersicum L.) seedlings. Environ. Sci. Pollut. R. 2019, 26, 9730–9738. [CrossRef]
- Zhao, X.Q.; Mitani, N.; Yamaji, N.; Shen, R.F.; Ma, J.F. Involvement of silicon influx transporter OsNIP2;1 in selenite uptake in rice. Plant Physiol. 2010, 153, 1871–1877. [CrossRef]
- Zhang, L.H.; Hu, B.; Li, W.; Che, R.H.; Deng, K.; Li, H.; Yu, F.Y.; Ling, H.Q.; Li, Y.J.; Chu, C.C. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice. New Phytol. 2014, 201, 1183–1191. [CrossRef]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [CrossRef]
- Longchamp, M.; Castrec-Rouelle, M.; Biron, P.; Bariac, T. Variations in the accumulation, localization and rate of metabolization of selenium in mature Zea mays plants supplied with selenite or selenate. Food Chem. 2015, 182, 128-135. [CrossRef]
- Abrams, M.; Burau, R.G.;Zasoski, R.J. Organic selenium distribution in selected California soils. Soil Sci. Soc. Am. J. 1990, 54, 979–982. [CrossRef]
- Li, H.F.; Lombi, E.; Stroud, J.L.; Mcgrath, S.P.; Zhao, F.J. Selenium speciation in soil and rice: influence of water management and Se fertilization. J. Agr. Food Chem. 2010, 58, 11837–11843. [CrossRef]
- Kieliszek, M.; Bła˙zejak, S. Selenium: significance, and outlook for supplementation. Nutriton 2013, 29, 713–718. [CrossRef]
- Kowalska, I.; Smole’n, S.; Czernicka, M.; Halka, M.; Keska, K.; Pitala, J. Effect of selenium form and Salicylic acid on the accumulation of selenium speciation forms in hydroponically grown lettuce. Agriculture 2020, 10, 584. [CrossRef]
- Hu, C.X.; Nie, Z.J.; Shi, H.Z.; Peng, H.Y.; Li, G.X.; Liu, H.Y.; Li, C.; Liu, H.E. Selenium uptake, translocation, subcellular distribution and speciation in winter wheat in response to phosphorus application combined with three types of selenium fertilizer. BMC Plant Biol. 2023, 23, 224. [CrossRef]
- Wang, Q.; Kong, L.X.; Huang, Q.Q.; Li, H.F.; Wan, Y.A. Uptake and translocation mechanisms of different forms of organic selenium in rice (Oryza sativa L.). Front. Plant Sci. 2022, 13, 970480. [CrossRef]
- Zhang, L.H.; Hu, B.; Deng, K.; Gao, X.K.; Sun, G.X.; Zhang, Z.L.; Li, P.; Wang, W.; Li, H.; Zhang, Z.H.; Fu, Z.H; Yang, J.Y; Gao, S.P.; Li, L.G; Yu, F.Y.; Li, Y.J.; Ling, H.Q.; Chu, C.C. NRT1.1B improves selenium concentrations in rice grains by facilitating selenomethinone translocation. Plant Biotechnol. J. 2019, 17, 1058–1068. [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant Soil 2008, 302 , 1–17. [CrossRef]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.J.; Breward, N.; Harriman, M.; Tucker, M. Biofortification of UK food crops with selenium. P. Nutr. Soc. 2011, 65, 169–181. [CrossRef]
- Larsen, E.H.; Hansen, M.; Paulin, H.; Sven, M.; Mary, R.; Margaret, R. Speciation and bioavailability of selenium in yeast-based intervention agents used in cancer chemoprevention studies. J. Aoac Int. 2004, 87, 225–232.
- Zhang, L.H.; Shi, W.M.; Wang, X.C. Difference in selenite absorption between high- and low-selenium rice cultivars and its mechanism. Plant Soil 2006, 282, 183–193. [CrossRef]
- Fujii, R.; Deverel, S.J.; Hatfield, D.B. Distribution of selenium in soils of agricultural fields, western San Joaquin Valley, California. Soil Sci. Soc. Am. J. 1988, 52, 1274–1283. [CrossRef]
- Niemietz, C.M.; Tyerman, S.D. New potent inhibitors of aquaporins: silver and gold compounds inhibit aquaporins of plant and human origin. Febs Lett. 2002, 531, 443–447. [CrossRef]
- Huang, Q.Q.; Yu, Y.; Wang, Q.; Luo, Z.; Jiang, R.F.; Li, H.F. Uptake kinetics and translocation of selenite and selenate as affected by iron plaque on root surfaces of rice seedlings. Planta 2015, 241, 907–916. [CrossRef]
- Kikkert, J.; Berkelaar, E. Plant uptake and translocation of inorganic and organic forms of selenium. Arch. Environ. Con. Tox. 2013, 65, 458–465. [CrossRef]
- Wang, M.K.; Dinh, Q.T.; Qi, M.X.; Wang, M.; Yang, W.X.; Zhou, F., Liang, D.L. Radicular and foliar uptake, and xylem- and phloem-mediated transport of selenium in maize (Zea mays L.): a comparison of five Se exogenous species. Plant Soil 2020, 446, 111–123. [CrossRef]
- Xu, H.Z.; Yan, J.P.; Qin, Y.; Xu, J. M.; Shohag, M.J.I.; Wei, Y.Y.; Gu, M.H. Effect of different forms of selenium on the physiological response and the cadmium uptake by rice under cadmium stress. Int. J. Env. Res. Pub. He. 2020, 17, 6991. [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular aminoacid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [CrossRef]
- Hu, T.; Li, H.F.; Li, J.X.; Zhao, G.S.; Wu, W.L.; Liu, L.P.; Wang, Q.; Guo, Y.B. Absorption and bio-transformation of selenium nanoparticles by wheat seedlings (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 597. [CrossRef]
- Wang, K.; Wang, Y.Q.; Li, K.; Wan, Y.N.; Wang, Q.; Zhuang, Z.; Guo, Y.B.; Li, H.F. Uptake, translocation and biotransformation of selenium nanoparticles in rice seedlings (Oryza sativa L.). J. Nanobiotechnol. 2020, 18, 103. [CrossRef]
- Deng, X.F.; Zhao, Z.Q.; Lv, C.H.; Zhang, Z.Z.; Yuan, L.X.; Liu, X.W. Effects of sulfur application on selenium uptake and seed selenium speciation in soybean (Glycine max L.) grown in different soil types. Ecotoxicol. Environ. Saf. 2021, 209, 111790. [CrossRef]
- Zhou, F.; Yang, W.X.; Wang, M.K.; Miao, Y.X.; Li, Z.; Liang, D.L. Effects of selenium application on Se content and speciation in Lentinula edodes. Food Chem. 2018, 265, 182–188. [CrossRef]
- Maruyama-Nakashita, A. Metabolic changes sustain the plant life in low-sulfur environments. Curr. Opin. Plant Biol., 2017, 39, 144–151. [CrossRef]
- Song, Z.; Shao, H.; Huang, H.; Shen, Y.; Wang, L.; Wu, F.; Han, D.; Song, J.Y.; Jia, H.F. Overexpression of the phosphate transportergene OsPT8 improves the Pi and selenium contents in Nicotiana tabacum. Environ. Exp. Bot. 2017, 137, 158–165. [CrossRef]
- Gong, R.; Ai, C.; Zhang, B.; Cheng, X. Effect of selenite on organic selenium speciation and selenium bioaccessibility in rice grains of two Se-enriched rice cultivars. Food Chem. 2018, 264, 443–448. [CrossRef]
- da Silva, D.F.; Cipriano, P.E.; de Souza, R.R.; Júnior, M.S.; da Silva, R.F.; Faquin, V.; Silva, M.L.D.; Guilherme, L.R.G. Anatomical and physiological characteristics of Raphanus sativus L. submitted to different selenium sources and forms application. Sci. Hortic-Amsterdam 2020, 260, 108839. [CrossRef]
- Xiao, T.T.; Boada, R.; Marini, C.; Mercè, L.; Valiente, M. Influence of a plant biostimulant on the uptake, distribution and speciation of se in se-enriched wheat (Triticum aestivum L. cv. Pinzón). Plant Soil 2020, 455, 409–423. [CrossRef]
- Wang, M.; Ali, F.; Qi, M.X.; Peng, Q.; Wang, M.K.; Ba˜nuelos, G.;Miao, S.Y.; Li, Z.; Dinh, Q.T.; Liang, D.L. Insights into uptake, accumulation, and subcellular distribution of selenium among eight wheat (Triticum aestivum L.) cultivars supplied with selenite and selenate. Ecotoxicol. Environ. Saf. 2021, 207, 111544. [CrossRef]
- Ogra, Y.; Ogihara, Y.; Anan, Y. Comparison of the metabolism of inorganic and organic selenium species between two selenium accumulator plants, garlic and Indian mustard. Metallomics 2017, 9, 61–68. [CrossRef]
- Mazej. D.; Falnoga, I.; Veber, M.; Stibilj, V. Determination of selenium species in plant leaves by HPLC-UV-HG-AFS. Talanta 2006, 68, 558–568. [CrossRef]
- Mazej, D.; Osvald, J.; Stibilj, V. Selenium species in leaves of chicory, dandelion, lamb’s lettuce and parsley. Food Chem. 2008, 107, 75–83. [CrossRef]
- Chen, N.; Zhao, C.H.; Zhang, T.H. Selenium transformation and selenium-rich foods. Food Biosci. 2021, 40, 100875. [CrossRef]
- Ari, B.; Oz, E.; Can, S.Z.; Bakirdere, S. Bioaccessibility and bioavailability of selenium species in Se-enriched leeks (Allium Porrum) cultivated by hydroponically. Food Chem. 2022, 372, 131314. [CrossRef]
- Zhang, K.; Guo, X.; Zhao, Q.; Han, Y.; Zhan, T.; Li, Y.; Tang, C.H.; Zhang, J.M. Development and application of a HPLC-ICP-MS method to determine selenium speciation in muscle of pigs treated with different selenium supplements. Food Chem. 2020, 302, 125371. [CrossRef]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: spotlight on speciation. Brit. J. Nutr. 2008, 100, 238–253. [CrossRef]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: current knowledgeand future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484s−1491s. [CrossRef]
- Zhang, L.H.; Chu, C.C. Selenium uptake, transport, metabolism, reutilization, and biofortification in rice. Rice 2022, 15, 30. [CrossRef]
- Kieliszek, M.; Bła˙zejak, S.; Gientka, I.; Bzducha-Wróbel, A. Accumulation and metabolism of selenium by yeast cells. Appl. Microbiol. 2015, 99, 5373–5382. [CrossRef]
- Versini, A.; Di Tullo, P.; Aubry, E.; Bueno, M.; Thiry, Y.; Pannier, F.; Castrec-Rouelle, M. Influence of Se concentrations and species in hydroponic cultures on Se uptake, translocation and assimilation in non-accumulator ryegrass. Plant Physiol. Bioch. 2016, 108, 372–380. [CrossRef]
- Kieliszek, M.; Bła˙zejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on growth and antioxidative system of yeast cells. Mol. Biol. Rep. 2019, 46, 1797–1808. [CrossRef]
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium speciation from food source to metabolites: a critical review. Anal. Bional. Chem. 2006, 385, 1304–1323. https://doi.org/ 10.1007/s00216-006-0529-8.
Figure 1.
The Se content in root and shoot of wheat seedlings under different Se treatments. Data are means + SE (n = 3). The different lowercase letters indicate significant differences between Se treatment at p < 0.05, under Se (IV), Se (VI), SeMet and SeOMet treatment, respectively.
Figure 1.
The Se content in root and shoot of wheat seedlings under different Se treatments. Data are means + SE (n = 3). The different lowercase letters indicate significant differences between Se treatment at p < 0.05, under Se (IV), Se (VI), SeMet and SeOMet treatment, respectively.
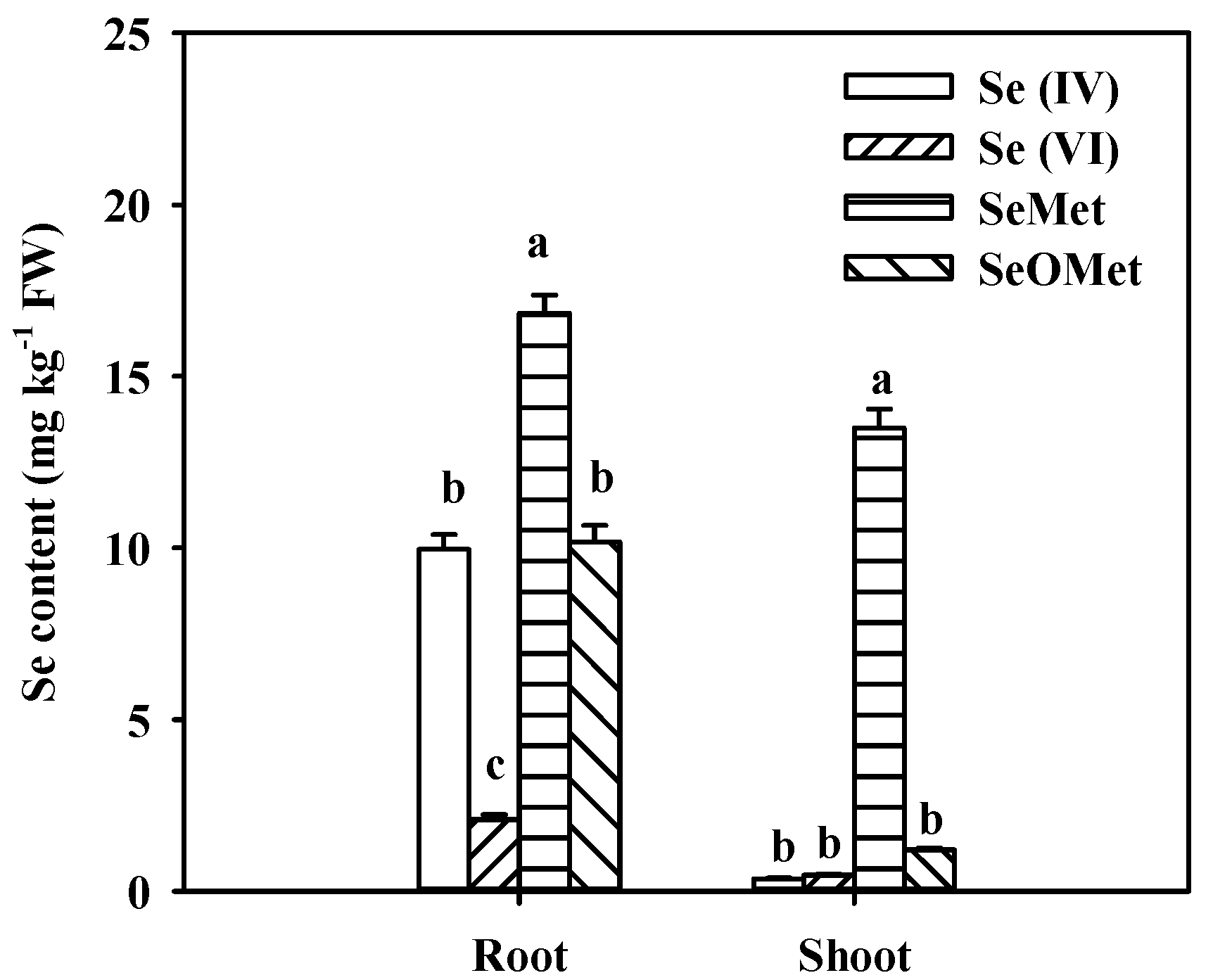
Figure 2.
Effects of aquaporin inhibition AgNO3 on Se contents in the wheat root (a) and shoot (b) under SeMet and SeOMet treatments. Data are means + SE (n = 3). The different letters indicate significant differences among the Se treatments at p < 0.05.
Figure 2.
Effects of aquaporin inhibition AgNO3 on Se contents in the wheat root (a) and shoot (b) under SeMet and SeOMet treatments. Data are means + SE (n = 3). The different letters indicate significant differences among the Se treatments at p < 0.05.
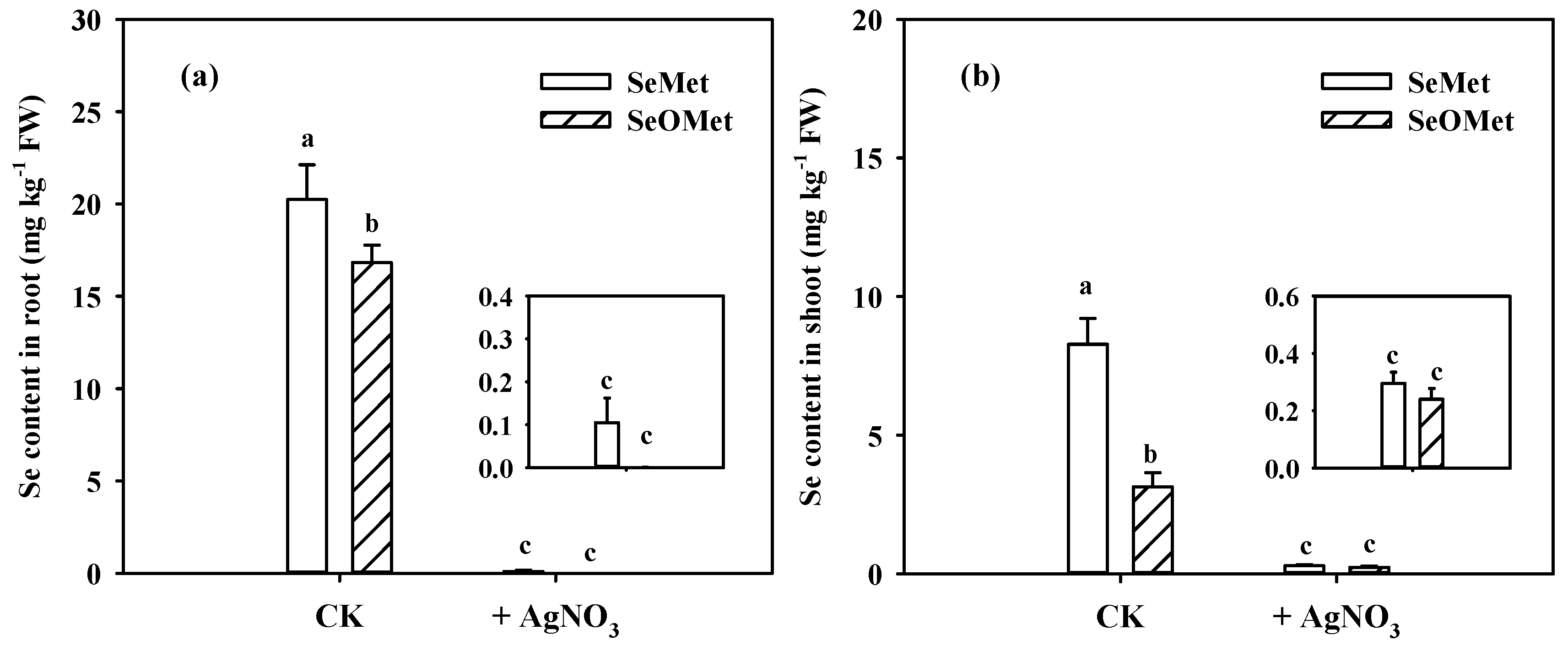
Figure 3.
Se contents in root (a) and shoot (b) of wheat seedlings treated with different time and organic Se.Data are means + SE (n = 3). The different letters in individual treatment times indicate significant differences among the organic Se treatments at the p < 0.05.
Figure 3.
Se contents in root (a) and shoot (b) of wheat seedlings treated with different time and organic Se.Data are means + SE (n = 3). The different letters in individual treatment times indicate significant differences among the organic Se treatments at the p < 0.05.
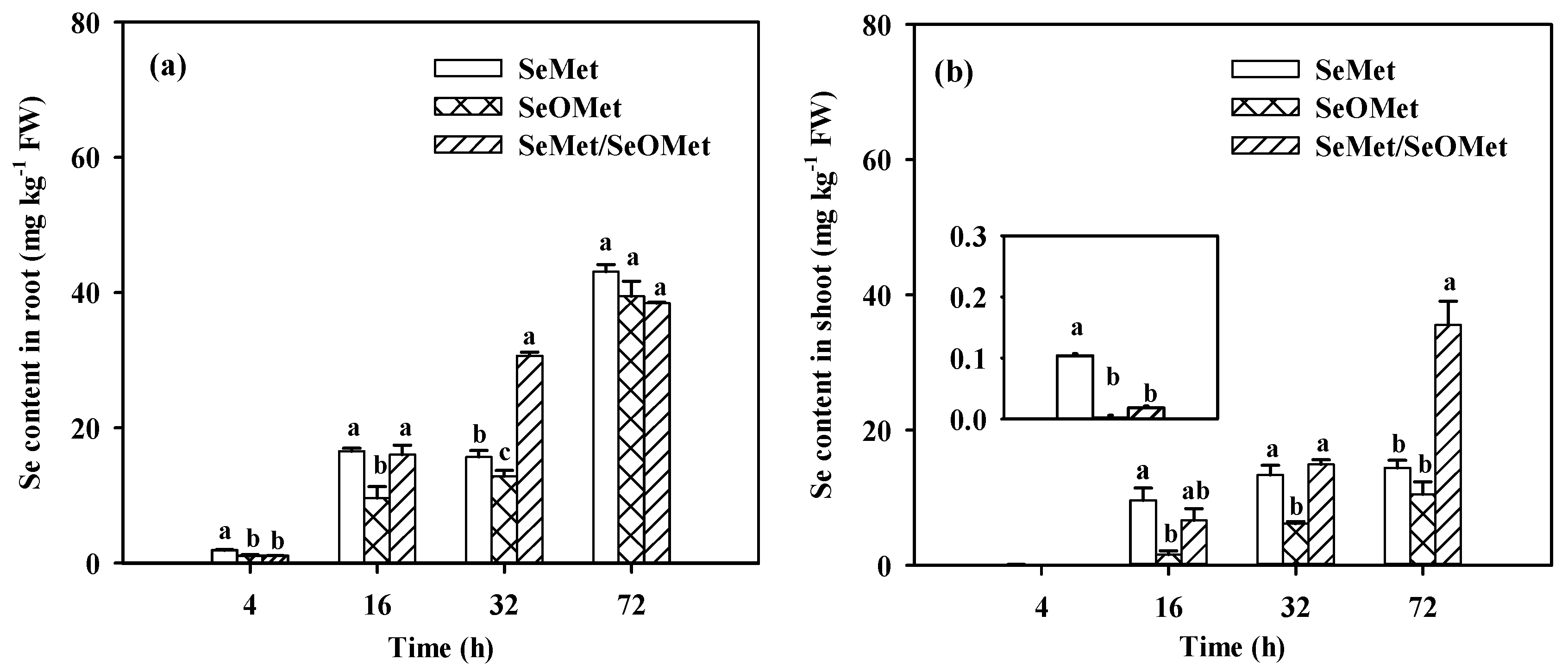
Figure 4.
Proportion of Se distributed to shoot (a) and transfer factor of Se (b) in wheat seedlings treated with different time and organic Se. Data are means + SE (n = 3). The different letters in individual treatment times indicate significant differences among the organic Se treatments at the p < 0.05.
Figure 4.
Proportion of Se distributed to shoot (a) and transfer factor of Se (b) in wheat seedlings treated with different time and organic Se. Data are means + SE (n = 3). The different letters in individual treatment times indicate significant differences among the organic Se treatments at the p < 0.05.
Figure 5.
Examples of HPLC-ICP-MS chromatograms of Se speciation in wheat roots and shoots treated for 72 h.
Figure 5.
Examples of HPLC-ICP-MS chromatograms of Se speciation in wheat roots and shoots treated for 72 h.
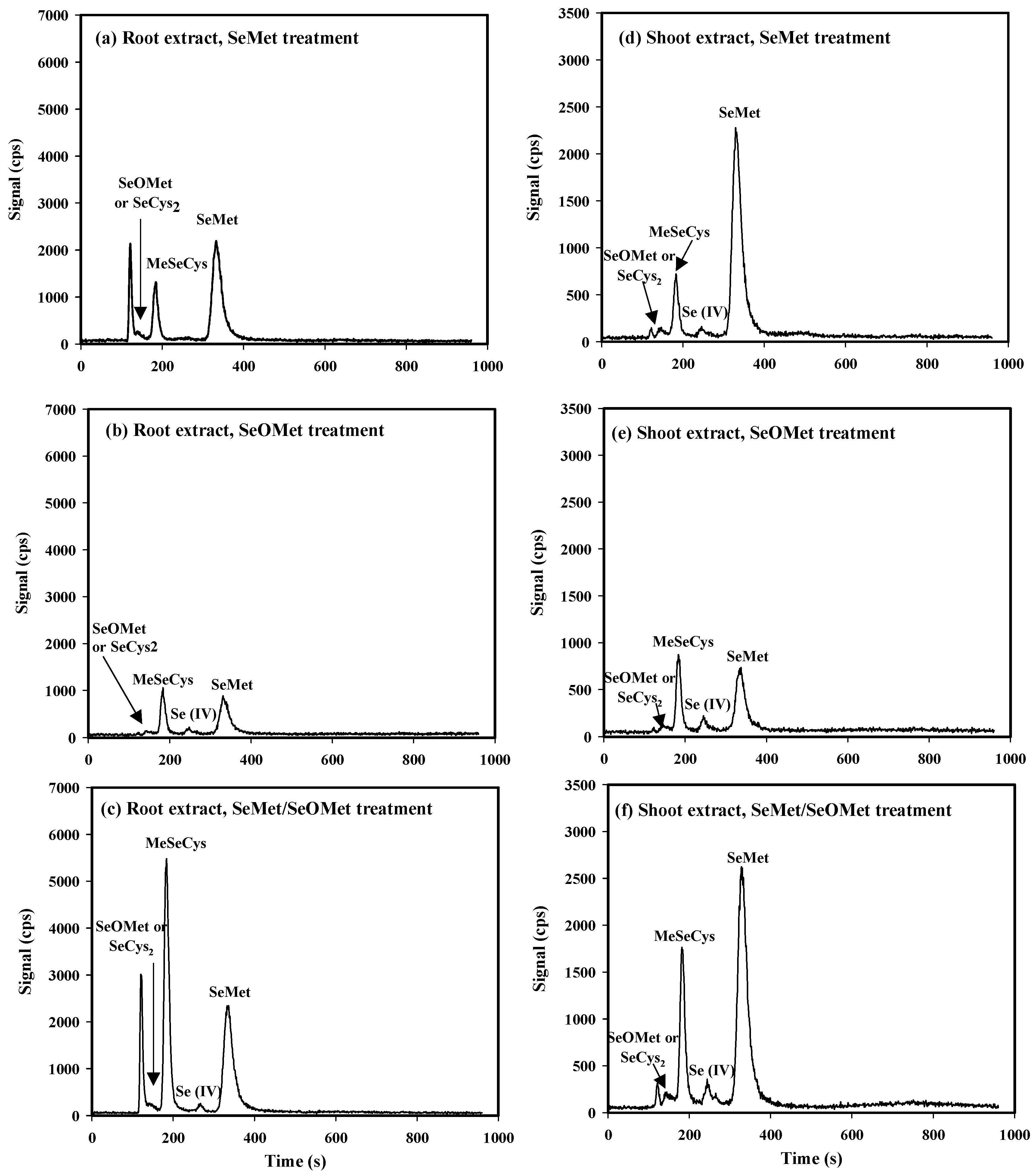
Figure 6.
Examples of HPLC-ICP-MS chromatograms of Se speciation in the xylem sap from wheat seedlings treated for 16 h.
Figure 6.
Examples of HPLC-ICP-MS chromatograms of Se speciation in the xylem sap from wheat seedlings treated for 16 h.
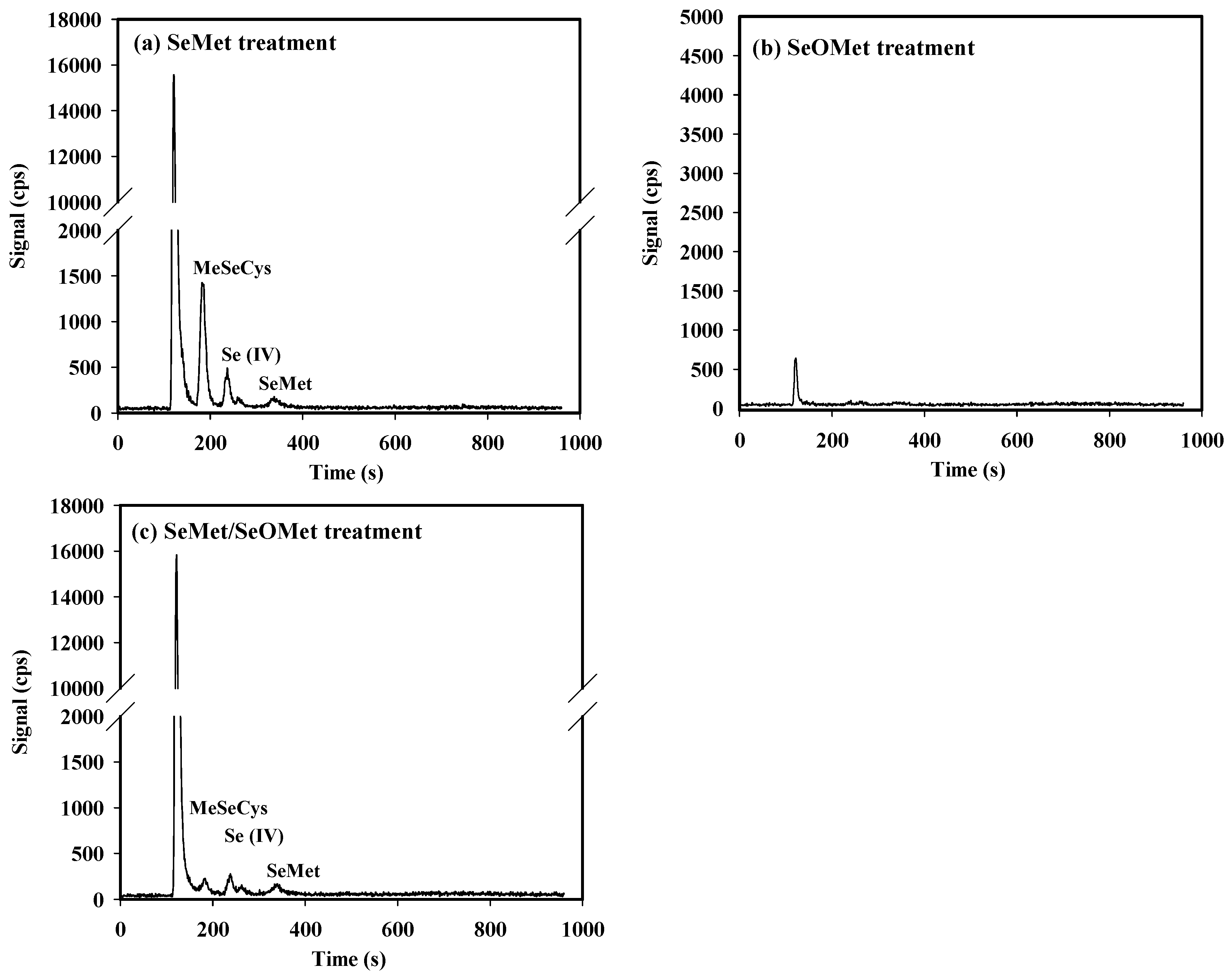

Table 1.
Effect of different Se forms on Se translocation and distribution in wheat seedlings.
| Se form | Se distribution (%) | Transfer factor | |
|---|---|---|---|
| Root | Shoot | ||
| Se (IV) | 94.0 ± 0.55 a | 6.00 ± 0.63 c | 0.028 ± 0.004 c |
| Se (VI) | 71.3 ± 0.29 b | 28.7 ± 0.34 b | 0.178 ± 0.004 b |
| SeMet | 42.4 ± 1.03 c | 57.6 ± 1.19 a | 0.643 ± 0.038 a |
| SeOMet | 77.1 ± 0.83 b | 22.9 ± 0.96 b | 0.123 ± 0.008 b |
Data are means ± SE (n = 3). The different letters in the same column indicate significant differences among the Se treatments at p < 0.05.
Table 2.
Effect of organic Se species supplied on Se speciation in the protease XIV extract from wheat root and shoot treated for 72 h.
Table 2.
Effect of organic Se species supplied on Se speciation in the protease XIV extract from wheat root and shoot treated for 72 h.
| Treatment | Se species (mg kg-1 FW) | ||||
|---|---|---|---|---|---|
| MeSeCys | Se (IV) | SeMet | SeOMet or SeCys2 | ||
| Root | SeMet | 2.55±0.21 b (6.84%) | 0.04±0.01 a (0.10%) | 8.19±1.14 a (22.0%) | 0.10±0.03 a (0.19%) |
| SeOMet | 1.53±0.16 b (3.65%) | 0.10±0.06 a (0.24%) | 4.33±0.09 a (10.3%) | 0.13±0.03 a (0.30%) | |
| SeMet+SeOMet | 8.34±1.82 a (21.7%) | 0.25±0.01 a (0.66%) | 9.89±0.46 a (25.8%) | 0.56±0.25 a (1.44%) | |
| Shoot | SeMet | 0.68±0.54 a (5.22%) | 0.11±0.07 a (0.86%) | 11.6±1.58 a (87.8%) | 0.29±0.16 a (2.19%) |
| SeOMet | 1.41±0.15 a (11.4%) | 0.22±0.18 a (1.75%) | 3.79±0.81 b (31.2%) | 0.07±0.03 a (0.55%) | |
| SeMet+SeOMet | 2.65±0.41 a (7.60%) | 0.50±0.07 a (1.43%) | 10.6±0.43 a (31.1%) | 0.20±0.01 a (0.58%) | |
Values in parentheses represent Se species percentage in wheat shoots or roots calculated using the expression: Se species content / total tissue content × 100%. Data are means ± SEs (n = 3). Different letters of the same Se species indicate significant differences among the organic Se treatments at the p < 0.05.
Table 3.
Se speciation in the xylem sap from wheat seedlings treated for 16 h.
| Treatment | Se concentration in xylem sap (mg L-1) | ||
|---|---|---|---|
| MeSeCys | Se (IV) | SeMet | |
| SeMet | 1.29 | 0.27 | 0.16 |
| SeOMet | 0.00 | 0.00 | 0.00 |
| SeMet/SeOMet | 0.13 | 0.15 | 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



