
Submitted:
10 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
Constant exposure to a variety of environmental factors has become increasingly problematic. A variety of illnesses are initiated or aided by the presence of certain perturbing factors. In the case of autism spectrum disorder, the environmental component begins to play an important part in de-termining the overall picture. Moreover, the lack of therapies to relieve existing symptoms com-plicates the fight against this condition. As a result, the use of animal models has made biomedical research easier and more suited for disease investigations. The current study used zebrafish as an animal model to mimic a real-life scenario: acute exposure to an increased dose of pesticides, fol-lowed by prospective intervention-based therapy with vitamin B12 (vit. B12). It is known that vit. B12 is involved in brain function, nerve tissue and red blood cell formation. Aside from this, the role of vit. B12 in the redox processes was recognized for its help against free radicals. To investigate the vit. B12 effect, fish were divided in four different groups and exposed to a pesticide mixture (600 μg L-1 fipronil + 600 μg L-1 pyriproxyfen) and 0.24 μg L-1 vit. B12 for 14 days. The impact of the compound was assessed daily by EthoVision software for behavioral observations, especially for sociability, quantified by the social interaction test. In addition, at the end of the study, the activities of su-peroxide dismutase (SOD), glutathione peroxidase (GPx), and malondialdehyde (MDA) were measured. Results showed significant improvements for locomotor activity parameters and a pos-itive influence of vitamin on sociability. Regarding the state of oxidative stress, a high activity was found for SOD and GPx in the case of vit. B12, while the mixture of pesticides and vitamin had a lower level of MDA. In conclusion, the study brings new data about the effect of vit. B12 in zebrafish, highlighting the potential use of vitamin supplementation to maintain and help the organism function.
Keywords:
Danio rerio
; behavior
; autism spectrum disorder
; pesticide
; vitamin B12
; oxidative stress
1. Introduction
Being one of the eight vitamins of group B, vitamin B12 (vit. B12), also called cobalamin due to the presence of cobalt mineral in its structure, has been often mentioned to be essential for cellular metabolism [1]. It is known to be involved in the synthesis of DNA molecules and myelin but also intervenes in the mediation of oxidative stress [2]. Its deficiency in the body was linked to motor alterations, memory loss, irritability, poor balance, and cognitive impairment [1,2,3]. It has been frequently associated with increased anemia, alterations in the central or peripheric nervous system, and the onset of mechanisms prior to disorders such as autism spectrum disorder (ASD), schizophrenia, and epilepsy [3,4,5]. Vit. B12 serves as a cofactor for two enzymes [6]. Methionine synthase in the cytoplasm demands vit. B12 in the form of methylcobalamin to catalyze the conversion of homocysteine to methionine. If the process is impaired due to the lack in vit. B12, the intracellular homocysteine increases, homocysteine being implicated in the mediation of ROS buildup, e.g., homocysteine auto-oxidation [7]. Vit. B12 is also essential for the mitochondrial enzyme methylmalonyl CoA mutase, which converts methylmalonyl CoA to succinyl CoA, a step in the oxidation of odd-chain fatty acids and the catabolism of ketogenic amino acids [1].
Mentioned for the first time in 1985, the concept of ”oxidative stress” has been defined as the imbalance between oxidants and antioxidants, characterizing the capacity of an organism to maintain its physiological state [8,9,10]. Usually, the increased levels of reactive oxygen species (ROS) are linked with important changes in cell functioning that lead to biomolecule damage and, in the end, can determine cell apoptosis [11,12]. It was indicated that there is a strong correlation between ASD and oxidative stress [13].
Recognized through the presence of significant disruptions in speech, social skills, and repetitive behavior, ASD is a well-known neurodevelopmental illness that started to gain more attention due to its high incidence [14,15]. Recent data highlights the prevalence of ASD among US children, estimated at 1 in 36 [16]. The rate of prevalence depends on several variables. For instance, it was shown that the frequency of ASD is greater for Asian children, non-Hispanic Black children, children with higher Social Vulnerability Index scores, or for children that receive treatment in urban primary care locations [17]. Additionally, the sex ratio difference is another concern among specialists. There are reports that present a much higher incidence for boys than girls in detecting ASD, and this is due to the complicated way to discover the specific autistic features in girls [18]. In reality, according to the Diagnostic and Statistical Manual of Mental Disorders (DSM-V), ASD is diagnosed based on three main characteristics, such as difficulty in social communication, deficiencies in social relationships and specific, repetitive patterns of behavior, activities or interests [19,20,21]. Although its etiology is not fully understood, ASD may be seen from a complex perspective that combines genetic and environmental components [15,22,23,24]. The most common reason is genetic susceptibility, although prenatal stresses, infections, parental age, dysfunctional familiar relationships, and exposure to neurotoxic chemicals are all believed to be risk factors [25,26,27,28,29].
For instance, the lack of a sufficient amount of cobalamin in the bodies of autistic people is mainly due to poor nutrition poor in micronutrients, but this is not the only cause [4,30]. Data on the prevalence of clinical eating issues such as anorexia nervosa and bulimia nervosa in the autism population is limited, but the latest results indicate that these conditions are more common in people with autism and/or attention-deficit hyperactivity disorder (ADHD) compared to the general population [31,32,33,34]. Moreover, screening for vit. B12 in children diagnosed with ASD showed lower vitamin levels compared to the control group, according to several studies [35,36,37,38]. Methylcobalamin supplementation has been proven to be effective for participants in several studies carried out, leading to a relief of the autistic-related symptoms [39,40,41,42,43]. Even though not much research has been done on the effect of vit. B12 in ASD or its effectiveness in alleviating core or associated symptoms, it appears that the current evidence supports the vitamin's capacity to help and even to be regarded as a prospective treatment intervention [43].
The role of vit. B12 has long been acclaimed, starting with the preliminary data obtained from experimental research, whose outcome contributed to new data acquisition. For instance, vitamin activity has been investigated in several studies that used animals as model organisms (zebrafish, mice, rats) for a specific disorder. Although an animal model cannot 100% mimic human features, there are three rules for validating its suitability as a model; it should have similar causes or mechanisms of onset, similar symptoms, and similar responsiveness to treatment [44,45]. In this study, zebrafish (Danio rerio) was chosen due to its multiple advantages, and it was considered to be eligible for developing an animal model for ASD; a fact highlighted by numerous reports [46,47,48,49,50]. In addition, it possesses the full complement of cobalamin metabolic enzymes, and its deficiency can be caused by mutations in MMACHC, a gene responsible for vit. B12 activity regulation [6,51,52]. According to Sloan et al. [51], zebrafish with mmachc mutant alleles showed impaired growth and developmental delays, but also responded to established therapies, suggesting that this may be a suitable model for studying cobalamin deficiency. A 2012 study evaluated the effect of various vit. B12 levels in zebrafish and demonstrated that 5 µg vit. B12 kg-1 was insufficient to support whole-body vitamin storage, but there were also no signs of deficiency [53]. When vit. B12 was administered in a mixture with 100 µg L-1 malathion, an organophosphate insecticide, it conducted to an arousal of the motor functions by modulating acetylcholinesterase activity. These results were further sustained by the diminished oxidative stress status [54].
Aside from the genetic component, the environment has recently attracted more attention as a risk factor for ASD. Specialists started to point out the involvement of the environment in ASD and the negative impact [14,22,26,28]. An imbalance between excitatory and inhibitory neuronal activity in most cases of ASD has been hypothesized as a common underlying defect with many converging etiologies. For example, the impairment of gamma-aminobutyric acid (GABA) neurotransmission in autistic people is described by numerous reports [55,56,57]. The present findings led to the choice of a mixture of pesticides (fipronil and pyriproxyfen), whose synergistic effect was correlated with the induction of GABA malfunction. Fipronil (FIP) is a compound from the phenylpyrazole class of insecticides that acts on the insect nervous system [58]. The main mechanism of action is the inhibition of GABA receptors and glutamate gated chloride channels, and, depending of the dose, this could end in extreme neuronal excitation until the death of the organism [59,60]. Pyriproxyfen (PYR) is a compound that mimics the natural hormone for insect growth [61]. Both compounds, alone or in mixture, are linked to developmental and histological abnormalities, behavioral disturbances, and elevated levels of oxidative stress [62,63,64,65,66,67,68,69,70,71].
Due to the increased role of the environment as a risk factor for ASD and the need for new therapeutic interventions for autistic people, the present study aimed to evaluate the effects of vit. of B12 and a mixture of pesticides administered individually or in combination in a zebrafish animal model, mimicking a real-life scenario. This objective was approached in the following steps: (1) evaluation of the effect of the compounds on locomotor activity, (2) characterization of sociability after exposure to the compounds, and (3) measurement of specific parameters for oxidative stress.
2. Materials and Methods
2.1. Animals
The 200 zebrafish juveniles (Danio rerio, WT AB, 3-4 months, sex ratio 1:1) were obtained from an authorized local supplier and kept in the laboratory for three weeks as an accommodation period. The environment in the housing aquarium was changed every 48 hours with dechlorinated tap water and in the experimental tanks every single day. The water parameters were constantly measured and kept at normal values by filtration and an air pump (Table 1). Tanks were illuminated with LED bends (307.5 LUX) with a photoperiodic cycle of 14:10 hours (light:dark). The fish were fed twice a day with flakes from TetraMin Tropical Flakes (an average of 0.15 ± 0.05 g per portion per fish, from which vit. B12 was approximately 0.12 µg).
2.2. Chemicals
The pesticide mixture used in this study was purchased from a veterinary store in liquid form. The product purchased was mainly made up of the two pesticides (67.5 mg FIP and 67.5 mg PYR) but also other chemicals such as: 0.3 mg of butylatehydroxysisol, 60 mg of benzyl alcohol, and 0.15 mg of butylhydroxytoluene. The concentration for each pesticide compound was 600 μg L-1 which was acquired by dissolving a certain quantity of the previously prepared stock solution into the medium. Vit. B12 was bought as tablets from a local pharmacy, a recommended product for dietary supplementation. The concentration of the vitamin used was 0.24 μg L-1. Each solution was prepared daily by shredding the tablets and dissolving the fragments in a 100 ml volumetric flask. The plastic vials with a lid were filled with 40 ml of solution (prepared by dissolving an amount of the stock solution), and the fish were kept for 30 minutes according to Pena’s protocol [72]. Both chemicals were commercial compounds since it is more common to use it like that than in the pure state of the active ingredient (to avoid conflict of interest the brands of the products will not be mentioned). For the oxidative stress analysis, it was used the Superoxide Dismutase Determination Kit (SOD, 19160-1KT-F), Glutathione Peroxidase Cellular Activity Assay Kit (GPx, CGP1-1KT), and Total Protein Kit, Micro Lowry, Peterson’s Modification (TP0300-1KT) from Merck, Germany. In addition, malondialdehyde (MDA) levels were assessed through the thiobarbituric acid-reactive assay after the Balmus et al. protocol [73].
2.3. Experimental design
The fish were randomly transferred from the housing aquarium to the experimental tanks and left to get used to the new space. Zebrafish juveniles were exposed to a single dose of the pesticide mixture while vit. B12 was administered for a period of two weeks in order to simulate a real-life situation. Moreover, the vitamin treatment duration was chosen based on the existing literature data regarding the time required for vitamins to intervene in organism’s metabolism. Experimental groups were divided as it follows: control group (n= 15), vit. B12 group (0.24 μg, n= 15), mixture group (600 μg L-1 FIP + PYR, n= 15), and mixture & vit. group (0.24 μg L-1 vit. B12 + 600 μg L-1 FIP + PYR, n= 15). After the accommodation period ended, each fish was analyzed through two behavioral tests that aimed to record several specific parameters of the locomotor activity and time spent close to the group as an indicator of sociability, all of this phase being named: the pretreatment stage. A newly prepared pesticide solution was dissolved directly into the experimental tanks for the mixture and mixture & vit. groups. As regards vitamin administration, this was done by placing each fish in the plastic vials filled with vit. B12 solution and left in it for 30 minutes. The control and mixture groups simulated the exposure to the vitamin. Each fish was tested in the T-maze taking data on locomotor activity and sociability as in the initial assessment of behavior. These tests were repeated daily for two weeks. Data was acquired every day through the camera situated above the maze, connected to a computer with EthoVision XT software (Noldus, Holland), by which all the behavioral parameters were calculated. In the end, fish were killed by immersion in ice-cold water for a minimum of 5 minutes after opercular motion had stopped and then stored in the freezer at -80° C for oxidative stress analysis. The study had two more replicates. The schematic presentation of the entire study is represented in Figure 1.
2.3. Behavioral tests
2.3.1. Locomotor activity test
The specific locomotor activity parameters of the animals were measured before and after the treatment using the T-maze adapted for this test. The experimental apparatus is made from transparent Plexiglass and divided into three arms: left, right, and center (Figure 2). The starting point was established at the end of the center arm. To investigate the impact of the compounds on animals, a series of parameters were chosen to describe the locomotor activity. The total distance swam was the first one chosen, and it represents the total distance travelled by fish in the T-maze (cm) during a trial. Secondly, the average velocity parameter refers to how fast the fish is moving (cm s-1), while maximum acceleration was defined as the maximum fish speed of reaction (cm s-2). In addition, the time spent active or inactive describes the amount of time in which the fish was or was not moving (s). Each trial had a duration of 4 minutes, and the experimental session was carried out every day between 9 a.m. and 6 p.m.
2.3.2. Social interaction test
The social interaction test aims to assess the tendency of an individual to choose and/or spend time next to its conspecifics. For applying this test, it was used the same experimental apparatus described in the previous subsection, but with several adaptations. A transparent wall was added in the left arm to divide the area into two zones: the social stimulus and the tested zones (Figure 3). In the social stimulus zone was placed a group of three animals, while the tested zone corresponded to the experimental fish. Each fish was allowed to swim freely in all arms, except for the social stimulus zone, which was separated. The social behavior was quantified depending on the time spent by fish next to the social stimulus zone for a session of 4 minutes. Beside this, the time spent in the center and right arms was measured.
2.4. Oxidative stress measurement
At the end of the chronic exposure, all the experimental fish were killed by immersion in cold water (under 5° C) and kept in the freezer for the oxidative stress analysis. For measuring the oxidative stress markers, each fish was defrosted and homogenized in ice-cold saline (0.90% NaCl). After, all the samples were centrifuged at 5500 rpm for 15 minutes following Jin’s protocol [74]. The obtained supernatant was used to determine the SOD and GPx activity, MDA level, and protein concentrations. The enzyme activity was determined according to the suggested protocols from the kit packages and quantified by the spectrophotometer Specord 210 Plus from Analytik Jena, Germany, at the specific wavelengths. Protein measurement was made using the Bradford method [75].
2.5. Statiscal analysis
The OriginPro v.9.8 software (OriginLab Corporation, United States of America, 2021) was used to realize the statistical analysis for both experimental tests. The first step in analyzing the data consisted of verifying of the normality of the data distribution through the Shapiro-Wilk test. When this step was confirmed, Tukey's post hoc test was applied to demonstrate the significant differences between group parameters before and after the treatment period. The α value was established at 0.05 to indicate the mean differences between the group, with the data being expressed as the average ± standard error of the mean (S.E.M.). The graphic presentation of the results for the locomotor activity and social interaction tests was done using OriginPro software and Microsoft Package Excel files (Microsoft Office Professional Plus 2021) for oxidative stress results.
2.6. Ethical note
The guidelines for the accommodation and care of animals used for experimental and other scientific purposes, Directive 2010/63/EU of the European Parliament, and the Council of 22 September 2010 on the protection of animals used for scientific purposes were strictly followed and maintained [76,77]. Additionally, this experiment was also approved by the Ethical Commission from the Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine Iasi, with registration number 750/04.07.2019.
3. Results
3.1. Short-term changes in locomotor activity due to the presence of vitamin B12
The total distance swam by the control group did not reveal any changes during the experimental period (initial behavior: 886.3 ± 206.1 cm vs. the average of the study period: 886.6 ± 181.3 cm, p> 0.05, Tukey, ANOVA). Compared to the control group activity, the total distance for the 0.24 μg L-1 group registered increased values, with maximum peaks on D_5 (1518.3 ± 238.1 cm, p= 0.03, Tukey, ANOVA), D_7 (1333.4 ± 157.2 cm, p= 0.04, Tukey, ANOVA), and D_8 (1485.9 ± 152.8 cm, p = 0.04, Tukey, ANOVA) as opposed to the initial average: 755.1 ± 74.5 cm (Figure 4). A single exposure to the 600 μg L-1 FIP+PYR mixture triggered a decrease in the distance travelled in the first day (735.8 ± 166.4 cm, p> 0.05 Tukey, ANOVA) in comparison to the initial behavior (1090.1 ± 149.9 cm), but in the following days the values of this parameter exhibited an upward trend D_2 (1638.5 ± 421.8 cm), D_5 (1592.6 ± 404.8 cm), and D_8 (1666.4 ± 294.5 cm) without recording significant changes. Regarding the activity of the zebrafish exposed to the vitamin and pesticide mixture, the total distance value decreased on D_1 (707.2 ± 277.5 cm, p> 0.05, Tukey, ANOVA) versus pretreatment (913.6 ± 53.8 cm), with ups and downs during all the experimental period that recorded in the end a greater value (1203.4 ± 323.8 cm) than in the initial phase (Figure 4).
In regard to the parameter "swimming speed", it presented similar trends, as can be seen in Figure 5. The swimming speed registered high levels for the 0.24 μg L-1 (initial behavior: 3.14 ± 0.31 cm s-1 vs. end of the study: 5.07 ± 1.07 cm s-1, p> 0.05, Tukey, ANOVA) and 600 μg L-1 FIP+PYR (initial behavior: 4.54 ± 0.62 cm s-1 vs. end of the study: 5.44 ± 0.81 cm s-1, p> 0.05, Tukey, ANOVA) groups compared to the control (initial behavior: 4.09 ± 0.70 cm s-1 vs. end of the study: 4.50 ± 0.34 cm s-1, p> 0.05, Tukey, ANOVA) and the last group (initial behavior: 3.78 ± 0.17 cm s-1 vs. end of the study: 5.01 ± 1.35 cm s-1 (p> 0.05, Tukey, ANOVA).
Another quantified parameter for the locomotor activity characterization was "maximum acceleration". As it can be seen in Figure 6, this parameter did not record important modifications for any group during the experimental period. In addition, the single exposure to the pesticide mixture triggered a slight increase in maximum acceleration in the D_5: 235.8 ± 12.4 cm s-2 (p> 0.05, Tukey, ANOVA) compared to the pretreatment data: 211.9 ± 2.35 cm s-2. On the other hand, when vitamin was also present, this parameter showed reduced maximum acceleration for D_1 (192.6 ± 19.16 cm s-2, p> 0.05, Tukey, ANOVA) and D_13 (191.1 ± 15.6 cm s-2, p> 0.05, Tukey, ANOVA) versus the pretreatment data: 223.2 ± 11.06 cm s-2.
The “active swimming” parameter presented several changes in the activity of the fish. Although most zebrafish individuals did not exhibit important modifications in this parameter’s values, a fluctuating trend was observed during the experimental period. The vit. B12 group was the only one with constant activity. Even though the 0.24 μg L-1 group showed constant activity, there was a decrease in it on the D_2 (205.4 ± 8.54 s, p> 0.05, Tukey, ANOVA) in comparison to pretreatment data: 217.6 ± 3.14 s. After the first week of the single pesticide’s exposure, fish were able to regain similar values close to the initial behavior (223.3 ± 7.43 s). Thus, the lowest values of this parameter were recorded on the treatment day (176.9 ± 41.1 s), D_2 (191.5 ± 31.7 s), and D_5 (199 ± 37.9 s). When fish were exposed to vitamin and pesticides, the active swimming parameter revealed an unstable behavior. The highest value was recorded on D_12 (229.4 ± 3.91 s, p> 0.05, Tukey, ANOVA), and the lowest on D_1 (161.2 ± 32.7 s, p> 0.05, Tukey, ANOVA) compared to pretreatment (215.3 ± 10.2 s).
The ”inactive status” parameter revealed short-term changes in the experimental groups during the tested period. In the first week of exposure, as it can be seen from Figure 8, the group exposed to 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP + PIR presented a more pronounced time spent in inactivity compared to the initial behavior (average of the week: 62.36 ± 35.10 s vs. initial behavior: 24.64 ± 10.29 s). When the compounds were given alone, this trend changed. For instance, the vitamin group exhibited the lowest values for this parameter in comparison to the pesticide group, that had several elevated peaks recorded for the treatment day (63.01 ± 41.1 s), D_2 (48.44 ± 31.7 s), D_5 (40.9 ± 37.3 s) and D_7 (41.3 ± 41.1 s). The control group did not show any modifications to this parameter (p> 0.05, Tukey, ANOVA).
3.2. Impact of the presence of vitamin B12 on zebrafish social behavior
The sociability of zebrafish was evaluated through the social interaction test by measuring the time spent next to the stimulus zone. In Figure 9 the variations of this behavior during the experimental period are exhibited.
The control group revealed a normal and natural behavior that is specific to this organism, with more time spent next to the stimulus zone (124.5 ± 31.8 s), followed by the center arm (88.5 ± 27.3 s), and right arm (26.9 ± 15.7 s), with no significant difference between the experimental period and pretreatment (left: 135.04 ± 35.3 s vs. center: 92.4 ± 30.5 s, vs. right: 12.6 ± 8.84 s, p> 0.05, ANOVA) (Figure 10).
Regarding the 0.24 μg L-1 B12 group, the time spent in the maze arms reflected elevated values for the left and center arms (average of tested period: 111.3 ± 22.6 s vs. 106.1 ± 20.2 s) and lower for the right arm (22.6 ± 9.71 s) compared to initial behavior (left: 157.9 ± 16.1 s vs. center arm: 41.5 ± 8.5 s vs. right arm: 34.6 ± 14.6 s) (Figure 11).
Exposure to a single dose of 600 μg L-1 FIP+PIR did not trigger long-term effects on zebrafish sociability; the compound was capable of disturbing this behavior after the first days of treatment. The time spent in the left arm decreased on D_1 (70.4 ± 43.1 s) and D_3 (96.7 ± 27.7 s) compared to the pretreatment period (124.1 ± 29.7 s). Thus, after these days, the fish started to regain its normal behavior as can be observed in Figure 12. Also, the time spent in the right arm registered the lowest values (pretreatment: 20.2 ± 7.7 s vs. D_14: 14.1 ± 7.7 s) compared to the left arm (pretreatment: 95.7 ± 27.9 s vs. D_14: 75.1 ± 26.1 s) and center arm (pretreatment: 124.1 ± 29.7 s vs. D_14: 150.2 ± 33.1 s).
The group treated with 0.24 μg L-1 vit. B12 and 600 μg L-1 FIP+ PIR spent less time in the left arm on the treatment day (96.02 ± 42.1 s), D_1 (70.4 ± 43.1 s), and D_2 (108.1 ± 35.7 s) compared to the pretreatment period (120.4 ± 23.2 s). Starting with D_3, the time spent in the stimulus zone gradually increased, registering an average of 128.6 ± 30.8 s, almost like that from the initial period (Figure 13). Regarding the other areas of the maze, the time spent in the right and center arms by the fish presented ups and downs with no significant activity (p> 0.05, Tukey, ANOVA) (Figure 13).
3.3. Antioxidant boost after vitamin B12 supplementation
The biochemical activity is represented in Figure 14. Single exposure to the pesticide mixture did not trigger significant changes in SOD activity, neither did the group treated with vit. B12 (p> 0.05 Tukey, ANOVA). The same trend was observed for GPx activity (p> 0.05 Tukey, ANOVA). On the opposite, 14 days of treatment with 0.24 μg L-1 vit. B12 resulted in increased activities for SOD (p= 0.03 Tukey, ANOVA) and GPx (p= 0.02 Tukey, ANOVA). Regarding the lipid peroxidation process, the MDA marker did not show important variations among the experimental groups, excepting the last group (p= 0.03 Tukey, ANOVA) exposed to pesticide mixture and vitamin.
4. Discussion
The purpose of this study was to determine if vit. B12 can act as a therapeutic tool in an ASD animal model developed through environmental risk factor exposure. Modeling traits comparable to those stated in autistic people in an organism implies multiple phases. The first step in the construction of an animal model is the discovery of a suitable inductor, followed by the validation of the existence of ASD and the evaluation of an adequate response after trying a recognized therapeutic method. The effectiveness of vit. B12 in treating symptoms induced by exposure to FIP and PYR (already established to have potential to develop ASD impairments) was assessed in this trial. So far, no animal studies have been performed to evaluate the effects of vit. B12 on locomotor activity, on social features, or in association with ASD. Being the first study of its kind, these findings reveal that vit. B12 can assist zebrafish restore normal behavior after exposure to a mixture of pesticides by interacting with enzymes of the antioxidant system. As shown in the study results section, supplementing the zebrafish diet with vit. B12 led to improved levels of movement parameters describing fish locomotor activity.
Hyperactivity was one of the main findings recorded for the pesticide group that presented high values for distance, velocity, and maximum acceleration parameters in comparison to the other experimental groups. Even not significant, it could be seen the differences between pretreatment and treatment phases. Considering that there was a single high exposure to the mixture of pesticides, the impact could be seen over the whole study period, particularly during the first week. This may be explained by the time required for pesticides to be metabolized in fish. Besides, the excessive movement can also be a response to the interaction of fish with the two pesticides, that causes the appearance of stress, triggering an increase in cortisol secretion but also in ROS [78,79]. This observation has been made in many research studies after FIP and PYR exposure due to pesticide transformation into more toxic and lasting compounds. To give an example, the administration of a single oral dose of PYR (2 and 1000 mg kg-1) to rats showed not only that this pesticide has a higher excretion rate, but also a reduced depuration period, when the presence of tissue residues was determined after 7 days [80]. A more recent work, in which zebrafish were exposed to 10 and 100 μg L-1 PYR for 30 days, demonstrated the ability of the pesticide to accumulate after only one day of exposure, while the depuration period of 14 days indicated different half-life values: 2.3 days and 92.5% on day 7 for the lowest dose compared to 1.2 days and 94.6% on day 7 for the highest. In addition, compared with the other metabolites, both doses of PYR inhibited the activity of CAT and SOD in the liver [81]. Similar results as well as elevated levels of lipid peroxidation in the brain and kidney were obtained after oral administration of 2.5, 5, and 10 mg kg-1 body weight FIP to mice for 28 days [65]. The same observations were made in zebrafish after 96 hours of exposure to 0.5, 1 and 2 mg L-1 FIP [82]. Compared with the previous study, in the present work, no significant effects were recorded on the activities of SOD, GPx and MDA after pesticide exposure. This may be explained by variations in dose, exposure time and developmental stage that may trigger different consequences on zebrafish. Moreover, when this pesticide mixture was used in a 14-day treatment on zebrafish, it resulted in increased activities for SOD, GPx and MDA [83]. Consequently, it can be concluded that there is a link between the transformation of pesticides and the occurrence of oxidative stress, especially through the overproduction of free radicals, which can overwhelm the antioxidant system.
Although the effect of vit. B12 on the social behavior of zebrafish was not evidenced, oscillations between the left and central arms were observed after two weeks of treatment. On the other hand, the group exposed to 0.24 μg L-1 vit. B12 and 600 μg L-1 FIP+PIR, showed a clear preference for the region with social cues, suggesting that vit. B12 may participate in certain fish metabolism processes, allowing them to behave as prior to treatment. This observation could lead to the conclusion that the vitamin does not directly participate or act on social behavior, but its involvement in other metabolic processes could impact the behavior of individuals. For example, compared to the results of the previous group, a single dose of the pesticide mixture was capable to induce short- and medium-term changes in fish sociability in the first week of exposure. This behavioral alteration did not last until the end of experimental period, indicating that pesticides and their effects began to fade via the excretion process as soon as fish were transferred to system water. Even if the differences were not validated by a specific degree of significance, the graphical representations indicate certain trends for the behavioral parameters studied, which must be further investigated.
The potential antioxidant effect of vit. B12 was several times mentioned but it remains unclear, though its involvement as broad-spectrum micronutrient is known [84]. A recently published study discovered associations between subclinical vit. B12 deficiency and serum metabolic markers linked to neuron and mitochondria function, and increased oxidative stress [85]. Moreover, vit. B12 interacts with superoxide, a product of aerobic metabolism, at rates comparable to SOD, which highlighted the mimetic behavior of the vitamin towards the enzyme. [86,87]. Thereby, it can indicate a possible mechanism through which vitamin protects against chronic inflammation and controls redox homeostasis. In this study, exposure to 0.24 μg L-1 vit. B12 for 14 days conducted to elevated activities for SOD and GPx, while when the vitamin was given after the pesticide mixture, its activity was lower than for control group. Similar to these results, a 7-days treatment with 0.63 g kg-1 vit. B12 reduced hepatic enzymes levels (aminotransferase and aspartate aminotransferase), increased antioxidant activities, and diminished inflammatory cell infiltration and necrosis process in a male rat study for acetaminophen hepatotoxicity [88].
A possible explanation for this finding could be the ability of the vitamin administered in a non-stressful environment to promote and enhance the activity of the enzymes of the antioxidant system. On the other hand, in a disturbed environment (exposure to a mixture of pesticides) the vitamin participated together with the antioxidant system in counteracting the effects induced by FIP and PYR even if it was a single exposure. At the same time, the rate of accumulation in the zebrafish body and the products resulting from the metabolism of FIP and PYR may cause variations in the response to contact with the compounds. The ability of FIP to accumulate and the rate of metabolite formation in rainbow trout (Oncorhynchus mykiss) was measured after 32 days exposure followed by 96 days depuration. It concluded that FIP is rapidly converted to its metabolite, fipronil sulfone; known to have a longer half-life [89]. For instance, the elimination half-life of FIP was 8.5 hours compared to 208 hours for fipronil sulfone after administration of 4 mg kg-1 FIP in rats [60,90]. Also, the persistence of fipronil sulfone was confirmed after a 7 days depuration process in European sea bass (Dicentrarchus labrax) juveniles previously fed with 10 mg kg-1 FIP for 14 days [91].
Another parameter measured for identifying the existence of oxidative stress was MDA, a popular marker for lipid peroxidation. The exposure to a single dose of pesticides followed by 14 days vit. B12 led to a decrease in the MDA level compared to the other groups. This result indicates a possible intervention of the vitamin to regulate the balance between ROS products and antioxidants, which is also supported by the decreased activities of SOD and GPx. Dietary supplementation with vit. B12 had a positive influence on the methylation process, GSH activity, and oxidative stress in autistic children, after an 8-week therapy [41]. Similar results were previously reported by Bertoglio et al. following a 3-month therapy with 0.06 mg kg-1 vit. B12 [40]. A substantial decreased level of vit. B12 was recently found by two studies that assessed the vitamin's activity in children with ASD [37,92]. Perturbation in vitamin activity can also occur due to genetic mutations, as demonstrated in zebrafish by mutations in the abcd4 gene, whose deficiency caused anemia, or the mmachc gene responsible for vitamin processing and transporting to the cellular level [51,93]. Vitamin deficiency was also proven in a mouse model of ischemic stroke, where both females and males had impaired balance and coordination, as well as higher homocysteine levels compared to the 0.025 mg kg-1 group [94]. Neurologic alterations such as anxiety, deficits in learning and memory, and changes in brain mass were obtained in a knockout mouse in which the transcobalamin receptor (TCblR) gene (CD320) was ablated [95].
Finally, the interaction between vit. B12, antioxidants and ROS should be further studied and developed using animal models in an environmental setting, especially due to the high incidence of side effects, but also in a neuropsychiatric context.
5. Conclusions
The usefulness of the vitamin will need to be examined by developing animal models with similar characteristics to ASD, considering the new research that has come to light that indicates a lack of vitamin B12 in people with autism, whether it is genetic or environmental. Furthermore, the present findings revealed the vitamin participation in the oxidative stress environment, beside its contribution on zebrafish behavior. This study requires further attention to find out the optimal route of exposure, dose, treatment duration; all of these are considered limitations to the current study.
Author Contributions
Conceptualization, M.A.R.; methodology, M.A.R. and O.D.I.; software, M.A.R.; validation, A.C.; formal analysis, M.A.R. and O.D.I.; resources, M.N.N., L.E.R., D.U. and A.C.; data curation, M.A.R. and O.D.I.; writing—original draft preparation, M.A.R.; writing—review and editing, M.N.N., G.S. and A.C.; supervision, A.C.; project administration, M.N.N. and A.C.; funding acquisition, M.N.N. and D.U. All authors have read and agreed to the published version of the manuscript.
Funding
M.A.R. was supported by the Project POCU/993/6/13/153322, project title << Educational and training support for PhD students and young researchers in preparation for insertion into the labour market >> co-funded by the European Social Fund, through Operational Programme Human Capital 2014-2020. The other authors are thankful to the Romanian Ministry of Research, Innovation and Digitization, within Program 1—Development of the national RD system, Subprogram 1.2—Institutional Performance—RDI excellence funding projects, Contract no. 11PFE/30.12.2021, for financial support.
Institutional Review Board Statement
The animal study protocol was approved by the Ethical Commission from the Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine Iasi, with registration number 750/04.07.2019.
Data Availability Statement
Data supporting this study cannot be made available due to ethical and commercial reasons.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Green, R.; Allen, L. H.; Bjorke-Monsen, A.-L.; Brito, A.; Gueant, J.-L.; Miller, J. W.; Molloy, A. M.; Nexo, E.; Stabler, S.; Toh, B.-H.; Ueland, P. M.; Yajnik, C. Vitamin B12 Deficiency. Nat. Rev. Dis. Prim. 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Ospina, C. A.; Nava-Mesa, M. O. B Vitamins in the Nervous System: Current Knowledge of the Biochemical Modes of Action and Synergies of Thiamine, Pyridoxine, and Cobalamin. CNS Neurosci. Ther. 2020, 26(1), 5–13. [Google Scholar] [CrossRef] [PubMed]
- Briani, C.; Dalla Torre, C.; Citton, V.; Manara, R.; Pompanin, S.; Binotto, G.; Adami, F. Cobalamin Deficiency: Clinical Picture and Radiological Findings. Nutrients 2013, 5(11), 4521–4539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hodgson, N. W.; Trivedi, M. S.; Abdolmaleky, H. M.; Fournier, M.; Cuenod, M.; Do, K. Q.; Deth, R. C. Decreased Brain Levels of Vitamin B12 in Aging, Autism and Schizophrenia. PLoS One 2016, 11(1), e0146797–e0146797. [Google Scholar] [CrossRef]
- Serin, H. M.; Arslan, E. A. Neurological Symptoms of Vitamin B12 Deficiency: Analysis of Pediatric Patients. Acta Clin. Croat. 2019, 58(2), 295–302. [Google Scholar] [CrossRef] [PubMed]
- Froese, D. S.; Kopec, J.; Fitzpatrick, F.; Schuller, M.; McCorvie, T. J.; Chalk, R.; Plessl, T.; Fettelschoss, V.; Fowler, B.; Baumgartner, M. R.; Yue, W. W. Structural Insights into the MMACHC-MMADHC Protein Complex Involved in Vitamin B12 Trafficking. J. Biol. Chem. 2015, 290(49), 29167–29177. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M. R.; Tyagi, S. C. Homocysteine and Reactive Oxygen Species in Metabolic Syndrome, Type 2 Diabetes Mellitus, and Atheroscleropathy: The Pleiotropic Effects of Folate Supplementation. Nutr. J. 2004, 3, 4. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D. P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86(1), 715–748. [Google Scholar] [CrossRef] [PubMed]
- Manivasagam, T.; Arunadevi, S.; Essa, M. M.; SaravanaBabu, C.; Borah, A.; Thenmozhi, A. J.; Qoronfleh, M. W. Role of Oxidative Stress and Antioxidants in Autism. Adv. Neurobiol. 2020, 24, 193–206. [Google Scholar] [CrossRef]
- Liu, X.; Lin, J.; Zhang, H.; Khan, N. U.; Zhang, J.; Tang, X.; Cao, X.; Shen, L. Oxidative Stress in Autism Spectrum Disorder-Current Progress of Mechanisms and Biomarkers. Front. psychiatry 2022, 13, 813304. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Concept and Some Practical Aspects. Antioxidants (Basel, Switzerland) 2020, 9(9). [Google Scholar] [CrossRef] [PubMed]
- Jelinek, M.; Jurajda, M.; Duris, K. Oxidative Stress in the Brain: Basic Concepts and Treatment Strategies in Stroke. Antioxidants (Basel, Switzerland) 2021, 10(12). [Google Scholar] [CrossRef] [PubMed]
- Pangrazzi, L.; Balasco, L.; Bozzi, Y. Oxidative Stress and Immune System Dysfunction in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21(9), 3293. [Google Scholar] [CrossRef] [PubMed]
- Fakhoury, M. Autistic Spectrum Disorders: A Review of Clinical Features, Theories and Diagnosis. Int. J. Dev. Neurosci. 2015, 43, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism Spectrum Disorder. Lancet (London, England) 2018, 392(10146), 508–520. [Google Scholar] [CrossRef]
- Wormwood, K. L.; Charette, L.; Ryan, J. P.; Darie, C. C.; Woods, A. G. A Proteomics Investigation of Salivary Profiles as Potential Biomarkers for Autism Spectrum Disorder (ASD). Protein J. 2023, 42(5), 607–620. [Google Scholar] [CrossRef] [PubMed]
- Wallis, K. E.; Adebajo, T.; Bennett, A. E.; Drye, M.; Gerdes, M.; Miller, J. S.; Guthrie, W. Prevalence of Autism Spectrum Disorder in a Large Pediatric Primary Care Network. Autism 2023, 27(6), 1840–1846. [Google Scholar] [CrossRef] [PubMed]
- Salari, N.; Rasoulpoor, S.; Rasoulpoor, S.; Shohaimi, S.; Jafarpour, S.; Abdoli, N.; Khaledi-Paveh, B.; Mohammadi, M. The Global Prevalence of Autism Spectrum Disorder: A Comprehensive Systematic Review and Meta-Analysis. Ital. J. Pediatr. 2022, 48(1), 112. [Google Scholar] [CrossRef] [PubMed]
- Association, A. P. Diagnostic and Statistical Manual of Mental Disorders, fifth.; American Psychiatric Association: London, England, 2013. [Google Scholar] [CrossRef]
- Zwaigenbaum, L.; Penner, M. Autism Spectrum Disorder: Advances in Diagnosis and Evaluation. BMJ 2018, 361, k1674. [Google Scholar] [CrossRef]
- Al-Dewik, N.; Al-Jurf, R.; Styles, M.; Tahtamouni, S.; Alsharshani, D.; Alsharshani, M.; Ahmad, A. I.; Khattab, A.; Al Rifai, H.; Walid Qoronfleh, M. Overview and Introduction to Autism Spectrum Disorder (ASD). Adv. Neurobiol. 2020, 24, 3–42. [Google Scholar] [CrossRef]
- Sealey, L. A.; Hughes, B. W.; Sriskanda, A. N.; Guest, J. R.; Gibson, A. D.; Johnson-Williams, L.; Pace, D. G.; Bagasra, O. Environmental Factors in the Development of Autism Spectrum Disorders. Environ. Int. 2016, 88, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E. H.; Leventhal, B. L.; DiLavore, P. C.; Pickles, A.; Rutter, M. The Autism Diagnostic Observation Schedule—Generic: A Standard Measure of Social and Communication Deficits Associated with the Spectrum of Autism. J. Autism Dev. Disord. 2000, 30(3), 205–223. [Google Scholar] [CrossRef]
- Scheffer, I. E.; Nabbout, R. SCN1A-Related Phenotypes: Epilepsy and Beyond. Epilepsia 2019, 60 Suppl 3, S17–S24. [Google Scholar] [CrossRef]
- Moh, T. A.; Magiati, I. Factors Associated with Parental Stress and Satisfaction during the Process of Diagnosis of Children with Autism Spectrum Disorders. Res. Autism Spectr. Disord. 2012, 6(1), 293–303. [Google Scholar] [CrossRef]
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental Risk Factors for Autism: An Evidence-Based Review of Systematic Reviews and Meta-Analyses. Mol. Autism 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, G.; Geschwind, D. H. Genetics of Autism Spectrum Disorder. Handb. Clin. Neurol. 2018, 147, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Ongono, J. S.; Béranger, R.; Baghdadli, A.; Mortamais, M. Pesticides Used in Europe and Autism Spectrum Disorder Risk: Can Novel Exposure Hypotheses Be Formulated beyond Organophosphates, Organochlorines, Pyrethroids and Carbamates? - A Systematic Review. Environ. Res. 2020, 187, 109646. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, B.; Wu, C.; Wang, J.; Sun, M. Autism Spectrum Disorder: Neurodevelopmental Risk Factors, Biological Mechanism, and Precision Therapy. International Journal of Molecular Sciences. 2023. [CrossRef] [PubMed]
- Vissoker, R. E.; Latzer, Y.; Gal, E. Eating and Feeding Problems and Gastrointestinal Dysfunction in Autism Spectrum Disorders. Res. Autism Spectr. Disord. 2015, 12, 10–21. [Google Scholar] [CrossRef]
- Schreck, K. A.; Williams, K. Food Preferences and Factors Influencing Food Selectivity for Children with Autism Spectrum Disorders. Res. Dev. Disabil. 2006, 27(4), 353–363. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.; Xiong, X.; Yang, T.; Hou, N.; Liang, X.; Chen, J.; Cheng, Q.; Li, T. Correlation between Nutrition and Symptoms: Nutritional Survey of Children with Autism Spectrum Disorder in Chongqing, China. Nutrients 2016, 8(5). [Google Scholar] [CrossRef] [PubMed]
- Sharp, W. G.; Postorino, V.; McCracken, C. E.; Berry, R. C.; Criado, K. K.; Burrell, T. L.; Scahill, L. Dietary Intake, Nutrient Status, and Growth Parameters in Children with Autism Spectrum Disorder and Severe Food Selectivity: An Electronic Medical Record Review. J. Acad. Nutr. Diet. 2018, 118(10), 1943–1950. [Google Scholar] [CrossRef]
- Lundin Remnélius, K.; Neufeld, J.; Isaksson, J.; Bölte, S. Eating Problems in Autistic Females and Males: A Co-Twin Control Study. J. Autism Dev. Disord. 2022, 52(7), 3153–3168. [Google Scholar] [CrossRef]
- Al-Farsi, Y. M.; Waly, M. I.; Deth, R. C.; Al-Sharbati, M. M.; Al-Shafaee, M.; Al-Farsi, O.; Al-Khaduri, M. M.; Gupta, I.; Ali, A.; Al-Khalili, M.; Al-Adawi, S.; Hodgson, N. W.; Ouhtit, A. Low Folate and Vitamin B12 Nourishment Is Common in Omani Children with Newly Diagnosed Autism. Nutrition 2013, 29(3), 537–541. [Google Scholar] [CrossRef] [PubMed]
- Yektaş, Ç.; Alpay, M.; Tufan, A. E. Comparison of Serum B12, Folate and Homocysteine Concentrations in Children with Autism Spectrum Disorder or Attention Deficit Hyperactivity Disorder and Healthy Controls. Neuropsychiatr. Dis. Treat. 2019, 15, 2213–2219. [Google Scholar] [CrossRef] [PubMed]
- Mahruba, S.; Begum, S.; Shahjadi, S.; Afroz, S.; Siddiqi, U.; Parvin, J. Serum Vitamin B12 and Folic Acid Status in Autism Spectrum Disorder Children. J. Bangladesh Soc. Physiol. 2020, 14(2), 43–47. [Google Scholar] [CrossRef]
- Li, B.; Xu, Y.; Pang, D.; Zhao, Q.; Zhang, L.; Li, M.; Li, W.; Duan, G.; Zhu, C. Interrelation between Homocysteine Metabolism and the Development of Autism Spectrum Disorder in Children. Front. Mol. Neurosci. 2022, 15, 947513. [Google Scholar] [CrossRef]
- Laud, R. B.; Girolami, P. A.; Boscoe, J. H.; Gulotta, C. S. Treatment Outcomes for Severe Feeding Problems in Children with Autism Spectrum Disorder. Behav. Modif. 2009, 33(5), 520–536. [Google Scholar] [CrossRef] [PubMed]
- Bertoglio, K.; Jill James, S.; Deprey, L.; Brule, N.; Hendren, R. L. Pilot Study of the Effect of Methyl B12 Treatment on Behavioral and Biomarker Measures in Children with Autism. J. Altern. Complement. Med. 2010, 16(5), 555–560. [Google Scholar] [CrossRef]
- Hendren, R. L.; James, S. J.; Widjaja, F.; Lawton, B.; Rosenblatt, A.; Bent, S. Randomized, Placebo-Controlled Trial of Methyl B12 for Children with Autism. J. Child Adolesc. Psychopharmacol. 2016, 26(9), 774–783. [Google Scholar] [CrossRef]
- Allen, L. H.; Miller, J. W.; de Groot, L.; Rosenberg, I. H.; Smith, A. D.; Refsum, H.; Raiten, D. J. Biomarkers of Nutrition for Development (BOND): Vitamin B-12 Review. J. Nutr. 2018, 148 (suppl_4), 1995S–2027S. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D. A.; Frye, R. E. The Effectiveness of Cobalamin (B12) Treatment for Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. J. Pers. Med. 2021, 11(8). [Google Scholar] [CrossRef]
- Held, J. R. Appropriate Animal Models. Ann. N. Y. Acad. Sci. 1983, 406, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M. K.; Lindsey, J. R.; Davis, J. K. Requirements and Selection of an Animal Model. Isr. J. Med. Sci. 1987, 23(6), 551–555. [Google Scholar]
- Kalueff, A. V; Stewart, A. M.; Gerlai, R. Zebrafish as an Emerging Model for Studying Complex Brain Disorders. Trends Pharmacol. Sci. 2014, 35(2), 63–75. [Google Scholar] [CrossRef]
- Stewart, A. M.; Nguyen, M.; Wong, K.; Poudel, M. K.; Kalueff, A. V. Developing Zebrafish Models of Autism Spectrum Disorder (ASD). Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 50, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Meshalkina, D. A.; N. Kizlyk, M.; V. Kysil, E.; Collier, A. D.; Echevarria, D. J.; Abreu, M. S.; Barcellos, L. J. G.; Song, C.; Warnick, J. E.; Kyzar, E. J.; Kalueff, A. V. Zebrafish Models of Autism Spectrum Disorder. Exp. Neurol. 2018, 299, 207–216. [Google Scholar] [CrossRef]
- Shams, S.; Rihel, J.; Ortiz, J. G.; Gerlai, R. The Zebrafish as a Promising Tool for Modeling Human Brain Disorders: A Review Based upon an IBNS Symposium. Neurosci. Biobehav. Rev. 2018, 85, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Tayanloo-Beik, A.; Hamidpour, S. K.; Abedi, M.; Shojaei, H.; Tavirani, M. R.; Namazi, N.; Larijani, B.; Arjmand, B. Zebrafish Modeling of Autism Spectrum Disorders, Current Status and Future Prospective. Front. Psychiatry 2022, 13. [Google Scholar] [CrossRef]
- Sloan, J. L.; Achilly, N. P.; Arnold, M. L.; Catlett, J. L.; Blake, T.; Bishop, K.; Jones, M.; Harper, U.; English, M. A.; Anderson, S.; Trivedi, N. S.; Elkahloun, A.; Hoffmann, V.; Brooks, B. P.; Sood, R.; Venditti, C. P. The Vitamin B12 Processing Enzyme, Mmachc, Is Essential for Zebrafish Survival, Growth and Retinal Morphology. Hum. Mol. Genet. 2020, 29(13), 2109–2123. [Google Scholar] [CrossRef]
- Hannibal, L.; Jacobsen, D. W. Intracellular Processing of Vitamin B(12) by MMACHC (CblC). Vitam. Horm. 2022, 119, 275–298. [Google Scholar] [CrossRef]
- Hansen, A.-C.; Olsvik, P. A.; Hemre, G.-I. Effect of Different Dietary Vitamin B12 Levels on Their Retention in the Body of Zebrafish Danio rerio and on the Gene Expression of Vitamin B12 Binding Proteins. Aquac. Nutr. 2013, 19(3), 413–420. [Google Scholar] [CrossRef]
- Karmakar, S.; Sen Gupta, P.; Bhattacharya, S.; Sarkar, A.; Rahaman, A.; Mandal, D. P.; Bhattacharjee, S. Vitamin B12 Alleviates Malathion-Induced Toxicity in Zebrafish by Regulating Cytochrome P450 and PgP Expressions. Toxicol. Mech. Methods 2023, 33(5), 364–377. [Google Scholar] [CrossRef]
- Coghlan, S.; Horder, J.; Inkster, B.; Mendez, M. A.; Murphy, D. G.; Nutt, D. J. GABA System Dysfunction in Autism and Related Disorders: From Synapse to Symptoms. Neurosci. Biobehav. Rev. 2012, 36(9), 2044–2055. [Google Scholar] [CrossRef]
- Purkayastha, P.; Malapati, A.; Yogeeswari, P.; Sriram, D. A Review on GABA/Glutamate Pathway for Therapeutic Intervention of ASD and ADHD. Curr. Med. Chem. 2015, 22(15), 1850–1859. [Google Scholar] [CrossRef]
- Umesawa, Y.; Matsushima, K.; Atsumi, T.; Kato, T.; Fukatsu, R.; Wada, M.; Ide, M. Altered GABA Concentration in Brain Motor Area Is Associated with the Severity of Motor Disabilities in Individuals with Autism Spectrum Disorder. J. Autism Dev. Disord. 2020, 50(8), 2710–2722. [Google Scholar] [CrossRef]
- Gupta, R. C.; Miller Mukherjee, I. R.; Malik, J. K.; Doss, R. B.; Dettbarn, W.-D.; Milatovic, D. Chapter 26 - Insecticides; Gupta, R. C. B. T.-B. in T. (Second E., Ed.; Academic Press, 2019; pp 455–475. [CrossRef]
- Gupta, R. C.; Anadón, A. Chapter 42 - Fipronil; Gupta, R. C. B. T.-V. T. (Third E., Ed.; Academic Press, 2018; pp 533–538. [CrossRef]
- Singh, N. S.; Sharma, R.; Singh, S. K.; Singh, D. K. A Comprehensive Review of Environmental Fate and Degradation of Fipronil and Its Toxic Metabolites. Environ. Res. 2021, 199, 111316. [Google Scholar] [CrossRef]
- Sullivan, J. J.; Goh, K. S. Environmental Fate and Properties of Pyriproxyfen. J. Pestic. Sci. 2008, 33, 339–350. [Google Scholar] [CrossRef]
- Bedient, P.; Horsak, R.; Schlenk, D.; Hovinga, R.; Pierson, J. Environmental Impact of Fipronil to the Louisiana Crawfish Industry. Environ. Forensics - Env. FORENSICS 2005, 6, 289–299. [Google Scholar] [CrossRef]
- Mehrnoush, G.; Mehrdad, S.; Saeid, K. H. A. Effect Of Pyriproxyfen On Function And Tissue Of Testis In Adult Rat. Int. J. Curr. Res. Rev. 2013, 5, 66–74. [Google Scholar]
- Wu, H.; Gao, C.; Guo, Y.; Zhang, Y.; Zhang, J.; Ma, E. Acute Toxicity and Sublethal Effects of Fipronil on Detoxification Enzymes in Juvenile Zebrafish (Danio rerio). Pestic. Biochem. Physiol. 2014, 115, 9–14. [Google Scholar] [CrossRef]
- Badgujar, P. C.; Chandratre, G. A.; Pawar, N. N.; Telang, A. G.; Kurade, N. P. Fipronil Induced Oxidative Stress Involves Alterations in SOD1 and Catalase Gene Expression in Male Mice Liver: Protection by Vitamins E and C. Environ. Toxicol. 2016, 31(9), 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martínez, M. A.; Wu, Q.; Ares, I.; Martínez-Larrañaga, M. R.; Anadón, A.; Yuan, Z. Fipronil Insecticide Toxicology: Oxidative Stress and Metabolism. Crit. Rev. Toxicol. 2016, 46(10), 876–899. [Google Scholar] [CrossRef]
- Araújo, C. V. M.; Roque, D.; Blasco, J.; Ribeiro, R.; Moreira-Santos, M.; Toribio, A.; Aguirre, E.; Barro, S. Stress-Driven Emigration in Complex Field Scenarios of Habitat Disturbance: The Heterogeneous Multi-Habitat Assay System (HeMHAS). Sci. Total Environ. 2018, 644, 31–36. [Google Scholar] [CrossRef]
- Ghazanfar, M.; Shahid, S.; Qureshi, I. Z. Vitamin C Attenuates Biochemical and Genotoxic Damage in Common Carp (Cyprinus carpio) upon Joint Exposure to Combined Toxic Doses of Fipronil and Buprofezin Insecticides. Aquat. Toxicol. 2018, 196, 43–52. [Google Scholar] [CrossRef]
- Gusso, D.; Reolon, G. K.; Gonzalez, J. B.; Altenhofen, S.; Kist, L. W.; Bogo, M. R.; Bonan, C. D. Pyriproxyfen Exposure Impairs Cognitive Parameters and Alters Cortisol Levels in Zebrafish. Front. Behav. Neurosci. 2020, 14, 103. [Google Scholar] [CrossRef]
- Bownik, A.; Szabelak, A. Short-Term Effects of Pesticide Fipronil on Behavioral and Physiological Endpoints of Daphnia magna. Environ. Sci. Pollut. Res. 2021, 28(25), 33254–33264. [Google Scholar] [CrossRef]
- Staldoni de Oliveira, V.; Gomes Castro, A. J.; Marins, K.; Bittencourt Mendes, A. K.; Araújo Leite, G. A.; Zamoner, A.; Van Der Kraak, G.; Mena Barreto Silva, F. R. Pyriproxyfen Induces Intracellular Calcium Overload and Alters Antioxidant Defenses in Danio rerio Testis That May Influence Ongoing Spermatogenesis. Environ. Pollut. 2021, 270, 116055. [Google Scholar] [CrossRef]
- Pena, I. A.; Roussel, Y.; Daniel, K.; Mongeon, K.; Johnstone, D.; Weinschutz Mendes, H.; Bosma, M.; Saxena, V.; Lepage, N.; Chakraborty, P.; Dyment, D. A.; van Karnebeek, C. D. M.; Verhoeven-Duif, N.; Bui, T. V.; Boycott, K. M.; Ekker, M.; MacKenzie, A. Pyridoxine-Dependent Epilepsy in Zebrafish Caused by Aldh7a1 Deficiency. Genetics 2017, 207(4), 1501–1518. [Google Scholar] [CrossRef]
- Balmus, I.-M.; Lefter, R.; Ciobica, A.; Cojocaru, S.; Guenne, S.; Timofte, D.; Stanciu, C.; Trifan, A.; Hritcu, L. Preliminary Biochemical Description of Brain Oxidative Stress Status in Irritable Bowel Syndrome Contention-Stress Rat Model. Medicina (Kaunas). 2019, 55(12). [Google Scholar] [CrossRef]
- Jin, Y.; Liu, Z.; Liu, F.; Ye, Y.; Peng, T.; Fu, Z. Embryonic Exposure to Cadmium (II) and Chromium (VI) Induce Behavioral Alterations, Oxidative Stress and Immunotoxicity in Zebrafish (Danio rerio). Neurotoxicol. Teratol. 2015, 48, 9–17. [Google Scholar] [CrossRef]
- Bradford, M. M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Commission Recommendation. Guidelines for the Accommodation and Care of Animals Used for Experimental and Other Scientific Purposes (Notified under Document Number C(2007) 2525). Off. J. Eur. Union 2007, 50. [Google Scholar]
- Directive 63. The Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, 53. [Google Scholar]
- Stehr, C. M.; Linbo, T. L.; Incardona, J. P.; Scholz, N. L. The Developmental Neurotoxicity of Fipronil: Notochord Degeneration and Locomotor Defects in Zebrafish Embryos and Larvae. Toxicol. Sci. 2006, 92(1), 270–278. [Google Scholar] [CrossRef]
- Wang, C.; Qian, Y.; Zhang, X.; Chen, F.; Zhang, Q.; Li, Z.; Zhao, M. A Metabolomic Study of Fipronil for the Anxiety-like Behavior in Zebrafish Larvae at Environmentally Relevant Levels. Environ. Pollut. 2016, 211, 252–258. [Google Scholar] [CrossRef]
- Matsunaga, H.; Yoshino, H.; Isobe, N.; Kaneko, H.; Nakatsuka, I.; Yamada, H. Metabolism of Pyriproxyfen in Rats. 1. Absorption, Disposition, Excretion, and Biotransformation Studies with [Phenoxyphenyl-14C]Pyriproxyfen. J. Agric. Food Chem. 1995, 43(1), 235–240. [Google Scholar] [CrossRef]
- Wei, Y.; Cui, J.; Zhai, W.; Liu, X.; Zhou, Z.; Wang, P.; Liu, D. Toxicity and Fate of Chiral Insecticide Pyriproxyfen and Its Metabolites in Zebrafish (Danio rerio). Environ. Pollut. 2021, 280, 116894. [Google Scholar] [CrossRef]
- Wu, C.-H.; Lu, C.-W.; Hsu, T.-H.; Wu, W.-J.; Wang, S.-E. Neurotoxicity of Fipronil Affects Sensory and Motor Systems in Zebrafish. Pestic. Biochem. Physiol. 2021, 177, 104896. [Google Scholar] [CrossRef] [PubMed]
- Robea, M. A.; Jijie, R.; Nicoara, M.; Plavan, G.; Ciobica, A. S.; Solcan, C.; Audira, G.; Hsiao, C.-D.; Strungaru, S.-A. Vitamin C Attenuates Oxidative Stress and Behavioral Abnormalities Triggered by Fipronil and Pyriproxyfen Insecticide Chronic Exposure on Zebrafish Juvenile. Antioxidants (Basel, Switzerland) 2020, 9(10). [Google Scholar] [CrossRef]
- Manzanares, W.; Hardy, G. Vitamin B12: The Forgotten Micronutrient for Critical Care. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13(6), 662–8. [Google Scholar] [CrossRef]
- Brito, A.; Grapov, D.; Fahrmann, J.; Harvey, D.; Green, R.; Miller, J. W.; Fedosov, S. N.; Shahab-Ferdows, S.; Hampel, D.; Pedersen, T. L.; Fiehn, O.; Newman, J. W.; Uauy, R.; Allen, L. H. The Human Serum Metabolome of Vitamin B-12 Deficiency and Repletion, and Associations with Neurological Function in Elderly Adults. J. Nutr. 2017, 147(10), 1839–1849. [Google Scholar] [CrossRef]
- Suarez-Moreira, E.; Yun, J.; Birch, C. S.; Williams, J. H. H.; McCaddon, A.; Brasch, N. E. Vitamin B(12) and Redox Homeostasis: Cob(II)Alamin Reacts with Superoxide at Rates Approaching Superoxide Dismutase (SOD). J. Am. Chem. Soc. 2009, 131(42), 15078–15079. [Google Scholar] [CrossRef]
- Ayesha, A.; Bahnson, E. M.; Kayashima, Y.; Wilder, J.; Huynh, P. K.; Hiller, S.; Maeda-Smithies, N.; Li, F. Vitamin B12 Does Not Increase Cell Viability after Hydrogen Peroxide Induced Damage in Mouse Kidney Proximal Tubular Cells and Brain Endothelial Cells. Adv. redox Res. an Off. J. Soc. Redox Biol. Med. Soc. Free Radic. Res. 2022, 4. [Google Scholar] [CrossRef]
- Ahmed Mohammed, R.; Fadheel, Q. J. Hepatoprotective Effect of Vitamin B12 in Acetaminophen Induce Hepatotoxicity in Male Rats. Arch. Razi Inst. 2023, 78(1), 419–425. [Google Scholar] [CrossRef]
- Konwick, B. J.; Garrison, A. W.; Black, M. C.; Avants, J. K.; Fisk, A. T. Bioaccumulation, Biotransformation, and Metabolite Formation of Fipronil and Chiral Legacy Pesticides in Rainbow Trout. Environ. Sci. Technol. 2006, 40(9), 2930–2936. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.; Senarathna, L.; Percy, A.; Abeyewardene, M.; Eaglesham, G.; Cheng, R.; Azher, S.; Hittarage, A.; Dissanayake, W.; Sheriff, M. H. R.; Davies, W.; Buckley, N. A.; Eddleston, M. Acute Human Self-Poisoning with the N-Phenylpyrazole Insecticide Fipronil—a GABAA-Gated Chloride Channel Blocker. J. Toxicol. Clin. Toxicol. 2004, 42(7), 955–963. [Google Scholar] [CrossRef]
- Dallarés, S.; Dourado, P.; Sanahuja, I.; Solovyev, M.; Gisbert, E.; Montemurro, N.; Torreblanca, A.; Blázquez, M.; Solé, M. Multibiomarker Approach to Fipronil Exposure in the Fish Dicentrarchus labrax under Two Temperature Regimes. Aquat. Toxicol. 2020, 219, 105378. [Google Scholar] [CrossRef]
- Metyas, M. M.; Abdelhakim, A. S.; Ghandour, H. H. Screening of Vitamin B12 in Children Diagnosed as Autism Spectrum Disorder. QJM An Int. J. Med. 2020, 113 (Supplement_1). [Google Scholar] [CrossRef]
- Choi, Y. M.; Kim, Y.-I.; Choi, J.-H.; Bhandari, S.; Nam, I.-K.; Hong, K.; Kwak, S.; So, H.-S.; Park, D.-S.; Kim, C.-H.; Choi, T.-Y.; Choe, S.-K. Loss of Abcd4 in Zebrafish Leads to Vitamin B12-Deficiency Anemia. Biochem. Biophys. Res. Commun. 2019, 514(4), 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Yahn, G.; Wasek, B.; Bottiglieri, T.; Jadavji, N. A Dietary Vitamin B12 Deficiency Impairs Balance and Coordination After Ischemic Injury to the Sensorimotor Cortex in Adult Male and Female Mice. Curr. Dev. Nutr. 2021, 5, 932. [Google Scholar] [CrossRef]
- Arora, K.; Sequeira, J. M.; Hernández, A. I.; Alarcon, J. M.; Quadros, E. V. Behavioral Alterations Are Associated with Vitamin B12 Deficiency in the Transcobalamin Receptor/CD320 KO Mouse. PLoS One 2017, 12(5), e0177156. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The experimental design of the study for evaluating the impact of vitamin B12 and pesticide mixture on zebrafish.
Figure 1.
The experimental design of the study for evaluating the impact of vitamin B12 and pesticide mixture on zebrafish.
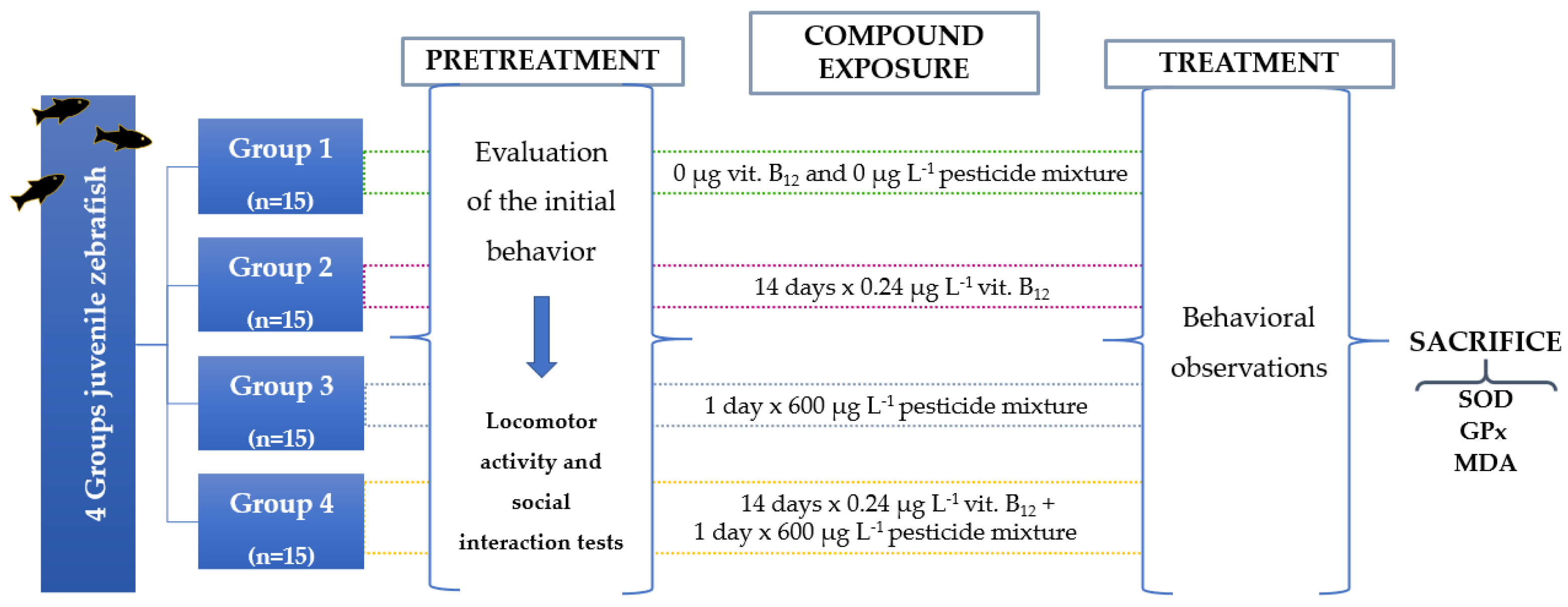
Figure 2.
The T-maze adaptation for performing the locomotor activity test; C: center arm, L: left arm, R: right arm and SP: start point.
Figure 2.
The T-maze adaptation for performing the locomotor activity test; C: center arm, L: left arm, R: right arm and SP: start point.
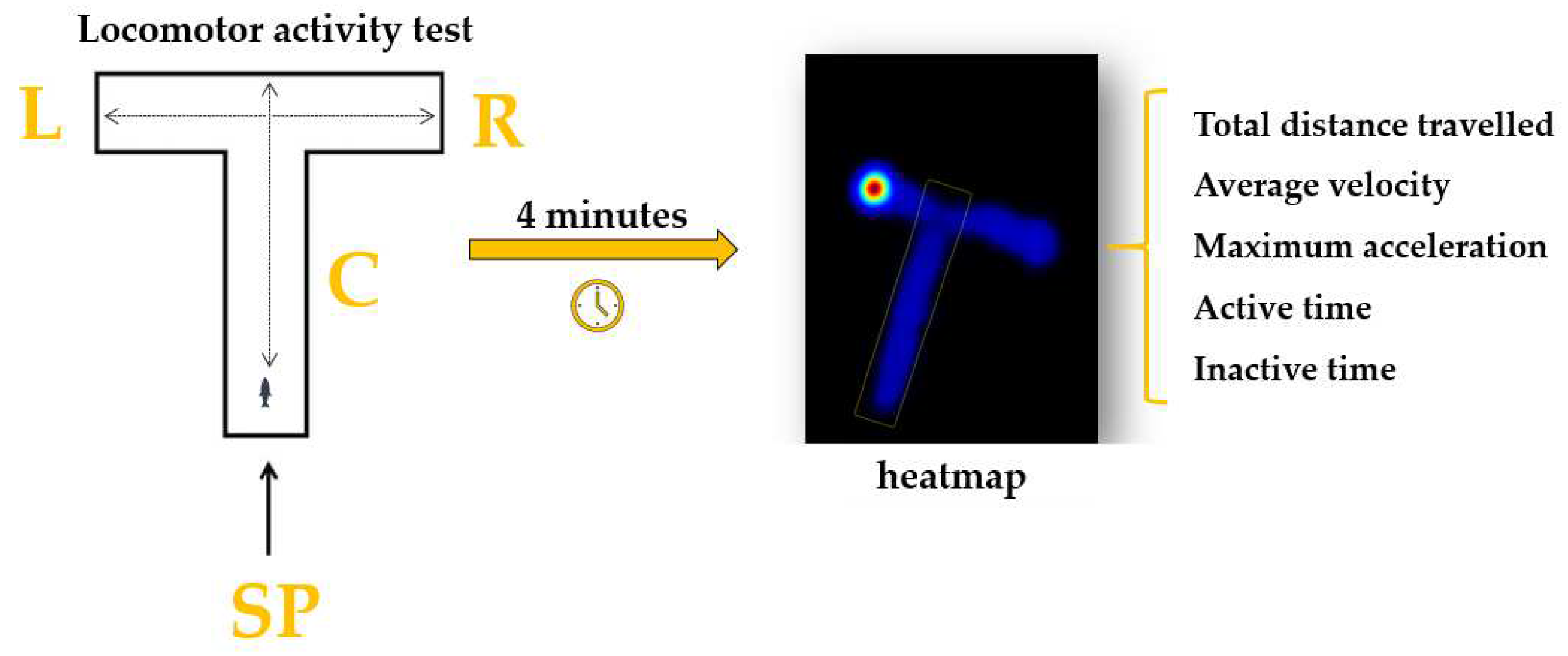
Figure 3.
The T-maze adaptation for performing the social interaction test; C: center arm, L: left arm, R: right arm, SP: start point, SSZ: social stimulus zone, TZ: tested zone.
Figure 3.
The T-maze adaptation for performing the social interaction test; C: center arm, L: left arm, R: right arm, SP: start point, SSZ: social stimulus zone, TZ: tested zone.
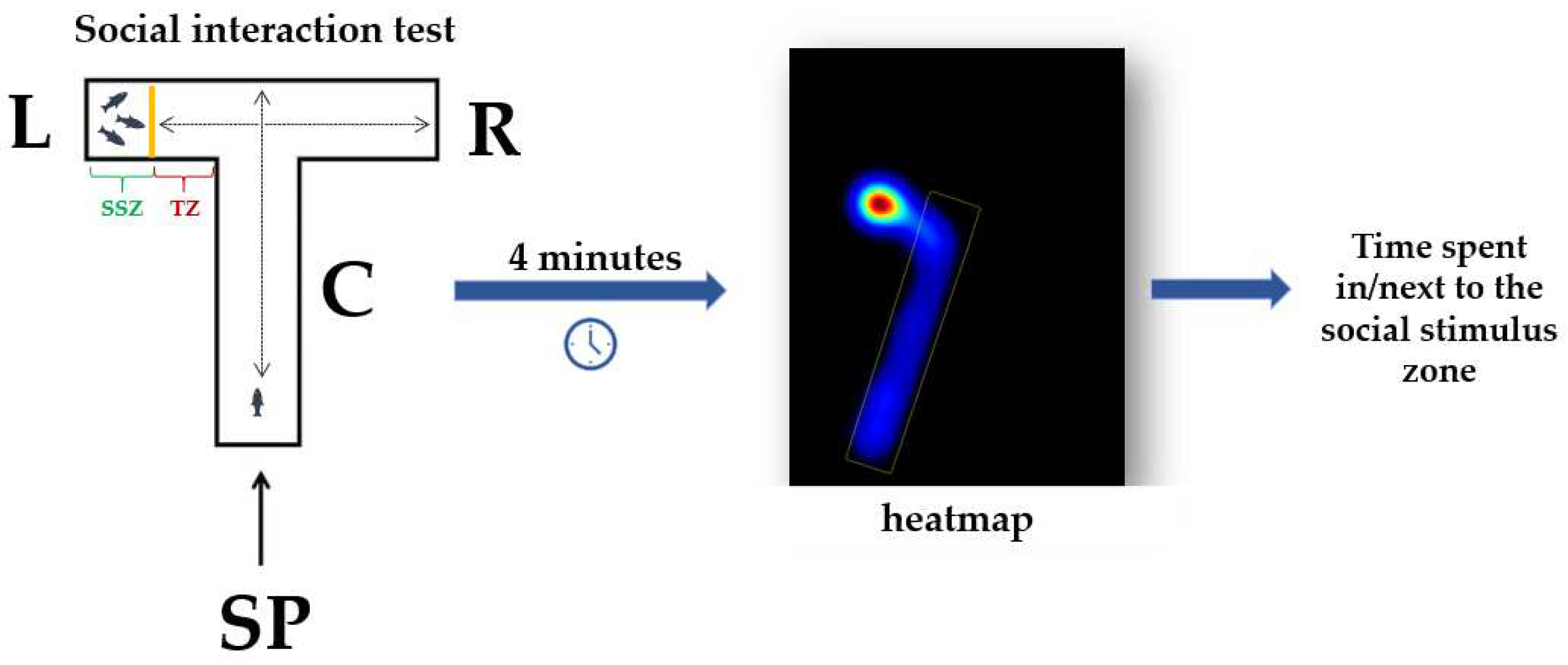
Figure 4.
The total distance travelled by juveniles from experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); *p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 4.
The total distance travelled by juveniles from experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); *p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
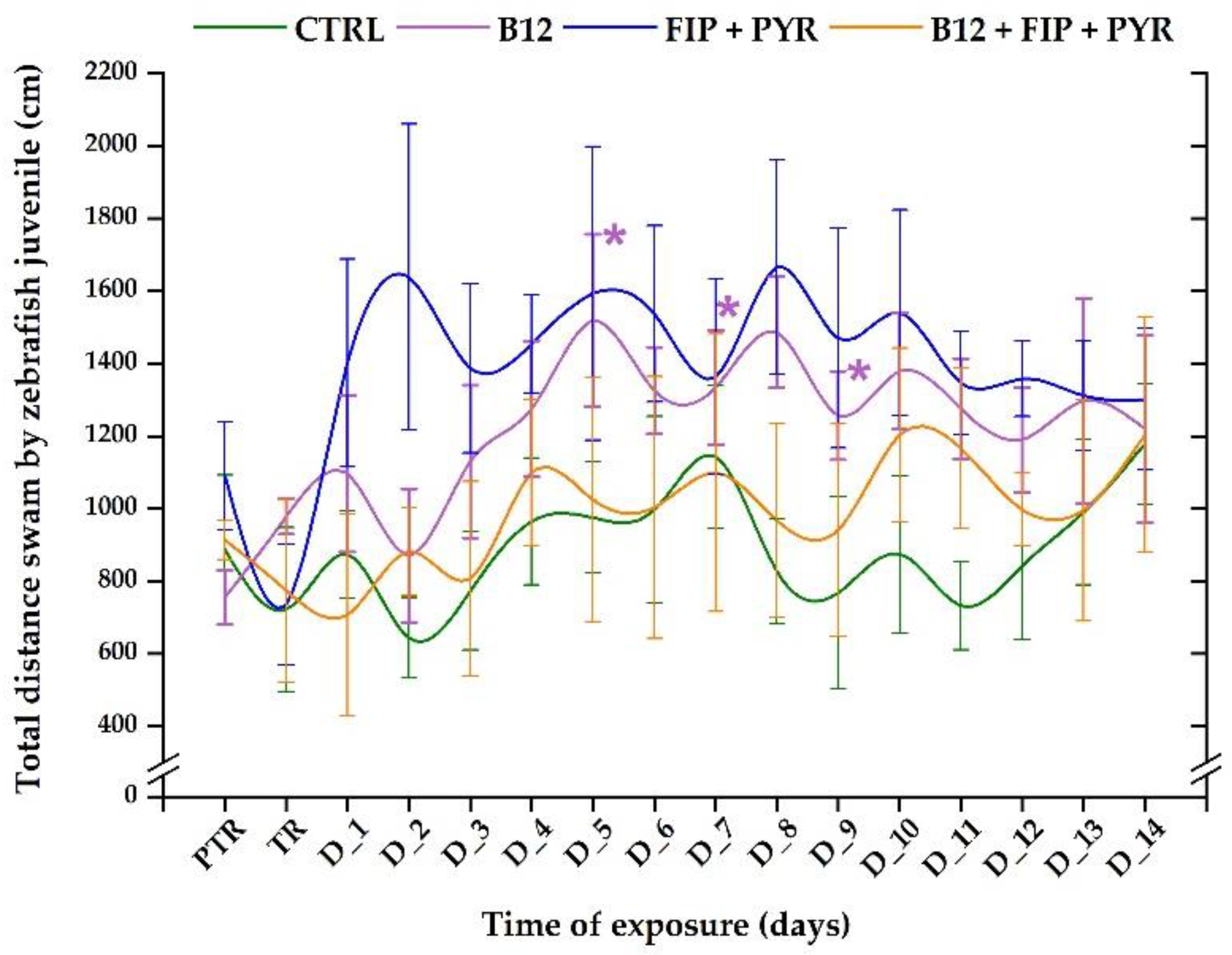
Figure 5.
The average swimming speed of the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 5.
The average swimming speed of the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
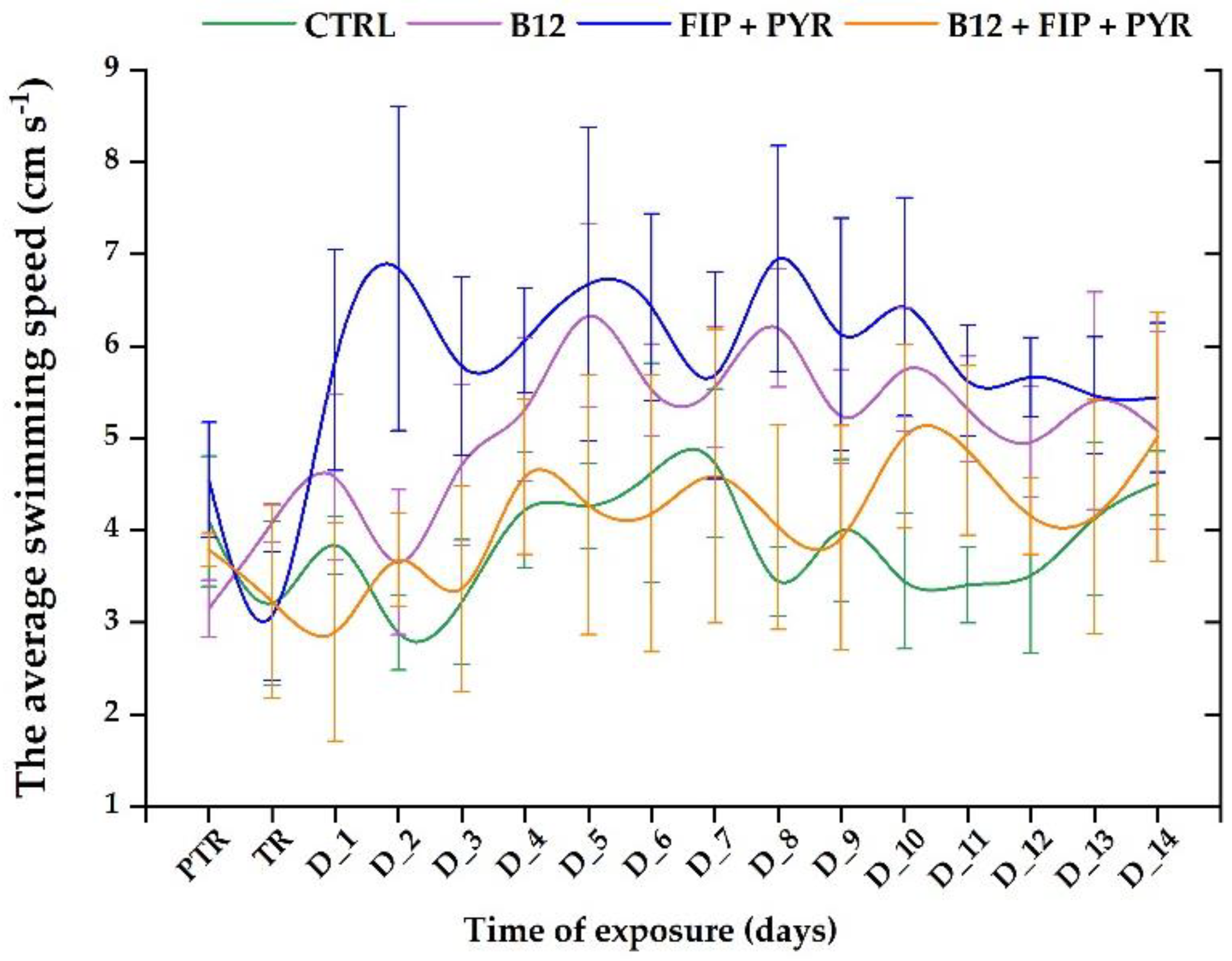
Figure 6.
The maximum acceleration of the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 6.
The maximum acceleration of the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
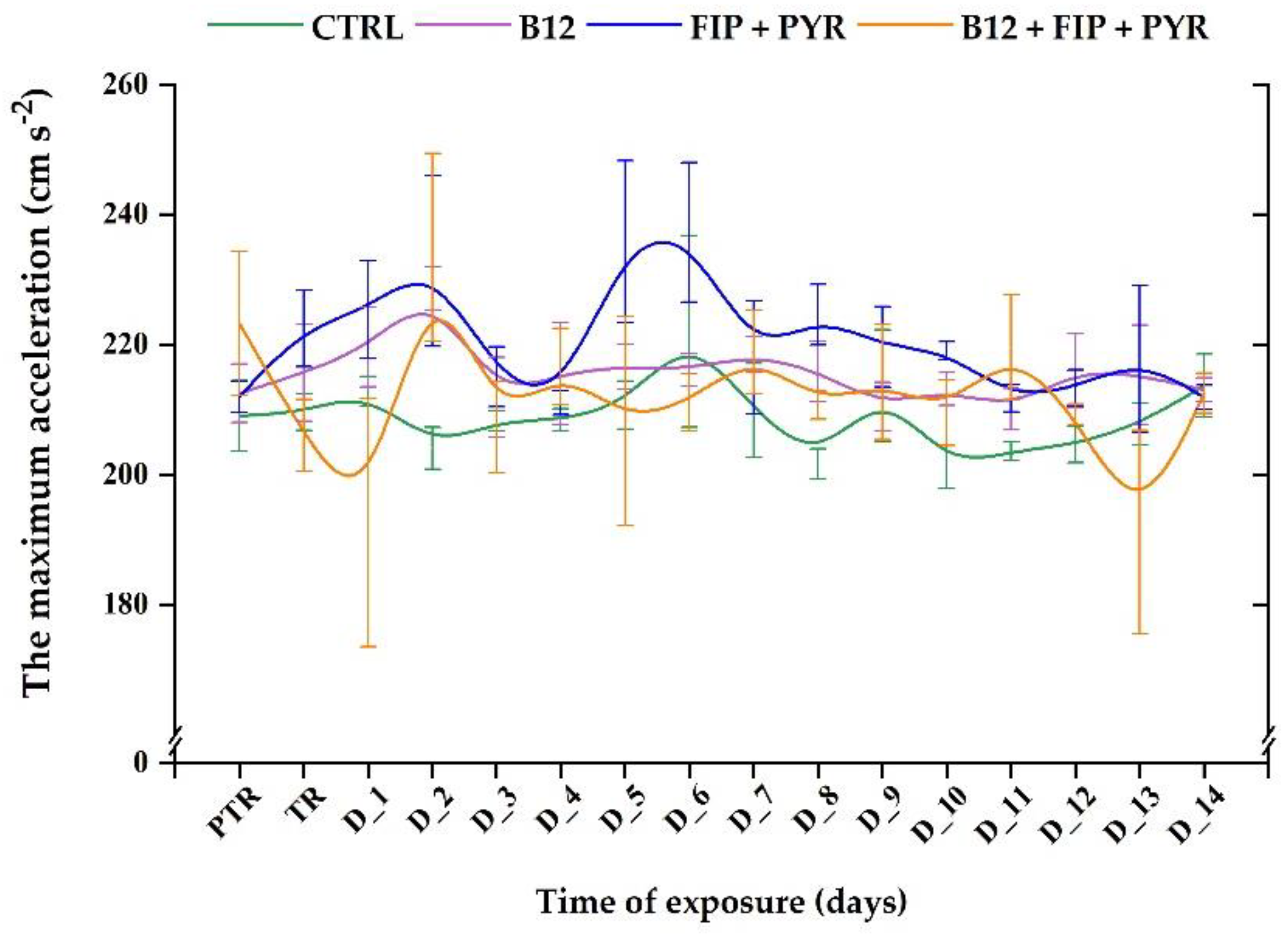
Figure 7.
The active swimming parameter recorded for all the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 7.
The active swimming parameter recorded for all the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
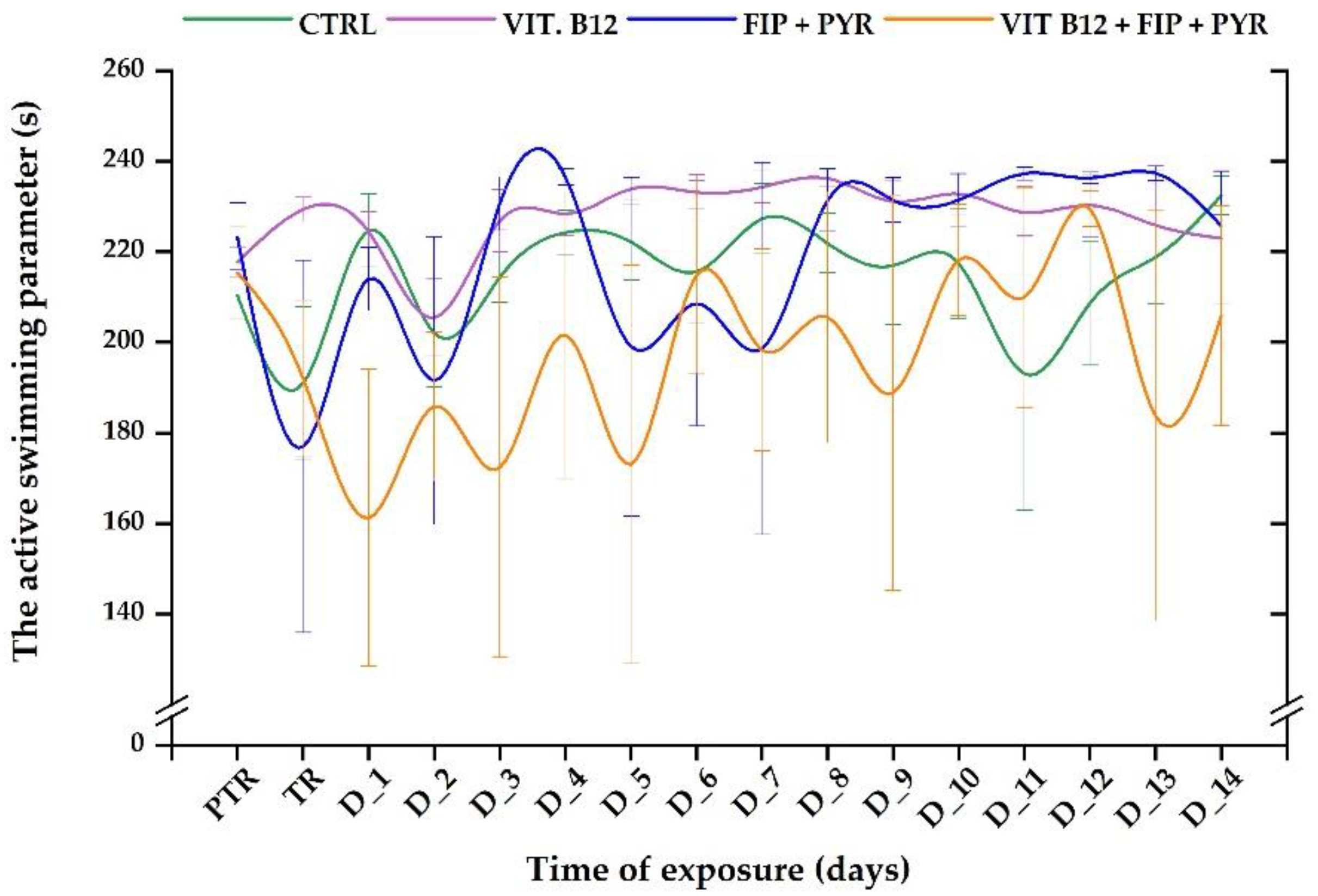
Figure 8.
The inactive status recorded for all the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 8.
The inactive status recorded for all the experimental groups. D stands for day, PTR: pretreatment, TR: treatment; Green: control, purple: 0.24 μg L-1 vit. B12, blue: 600 μg L-1 FIP+PIR, and orange: 0.24 μg L-1 vit. B12 + 600 μg L-1 FIP+PIR. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
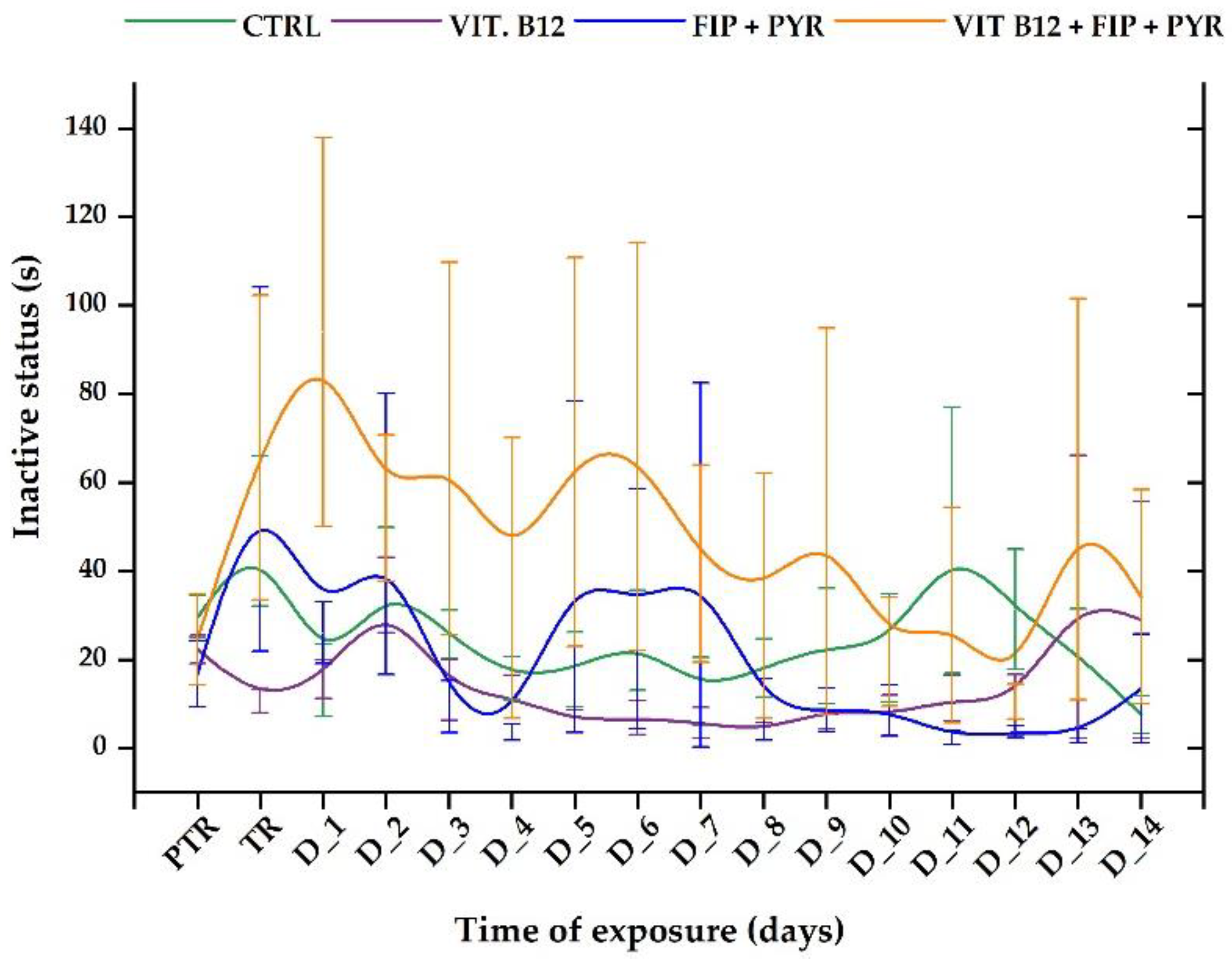
Figure 9.
The time spent in the T-maze areas during the social interaction test presented through heatmaps. D stands for day, PTR: pretreatment, TR: treatment.
Figure 9.
The time spent in the T-maze areas during the social interaction test presented through heatmaps. D stands for day, PTR: pretreatment, TR: treatment.
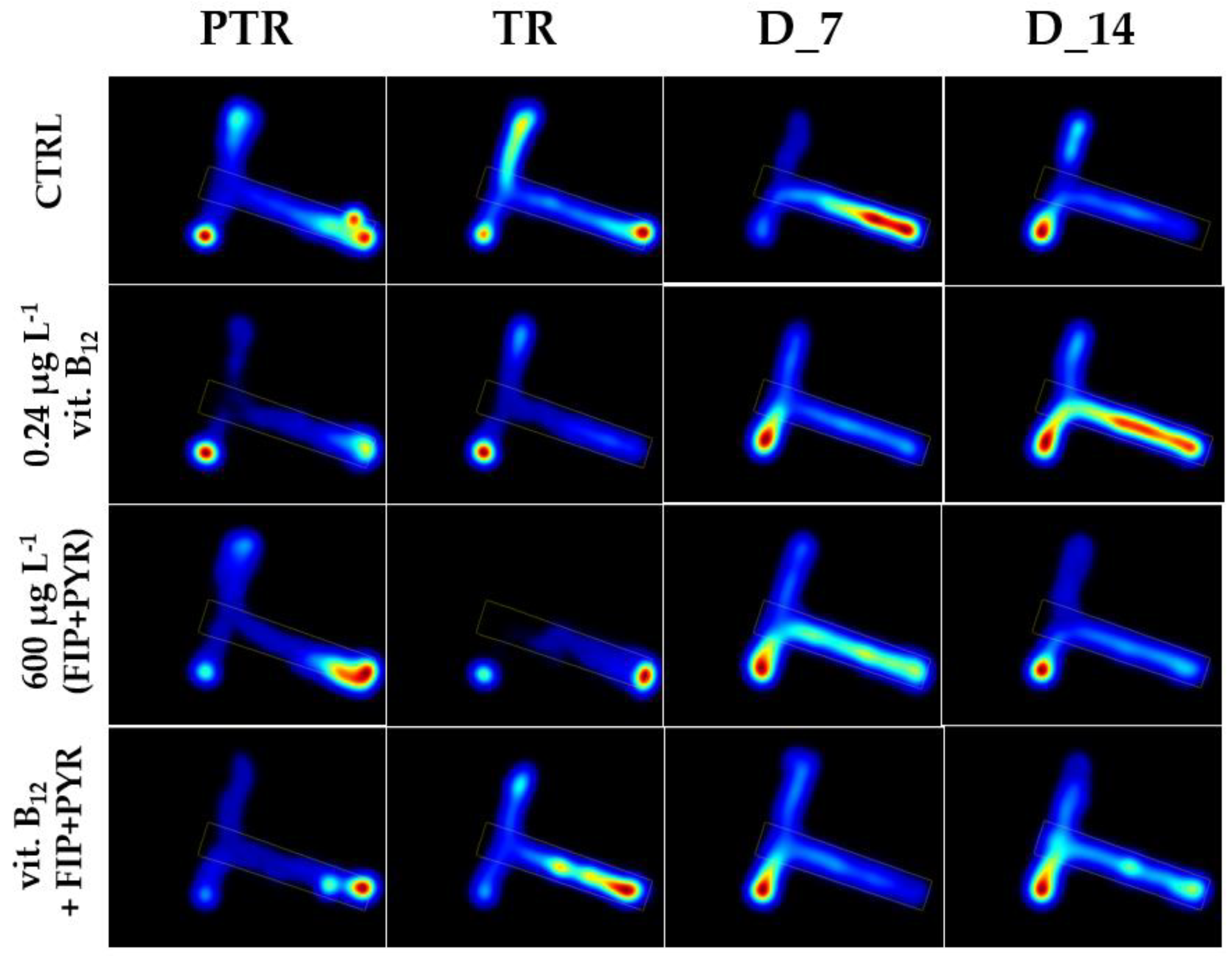
Figure 10.
The time spent in the T-maze areas, recorded for the control group. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 10.
The time spent in the T-maze areas, recorded for the control group. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
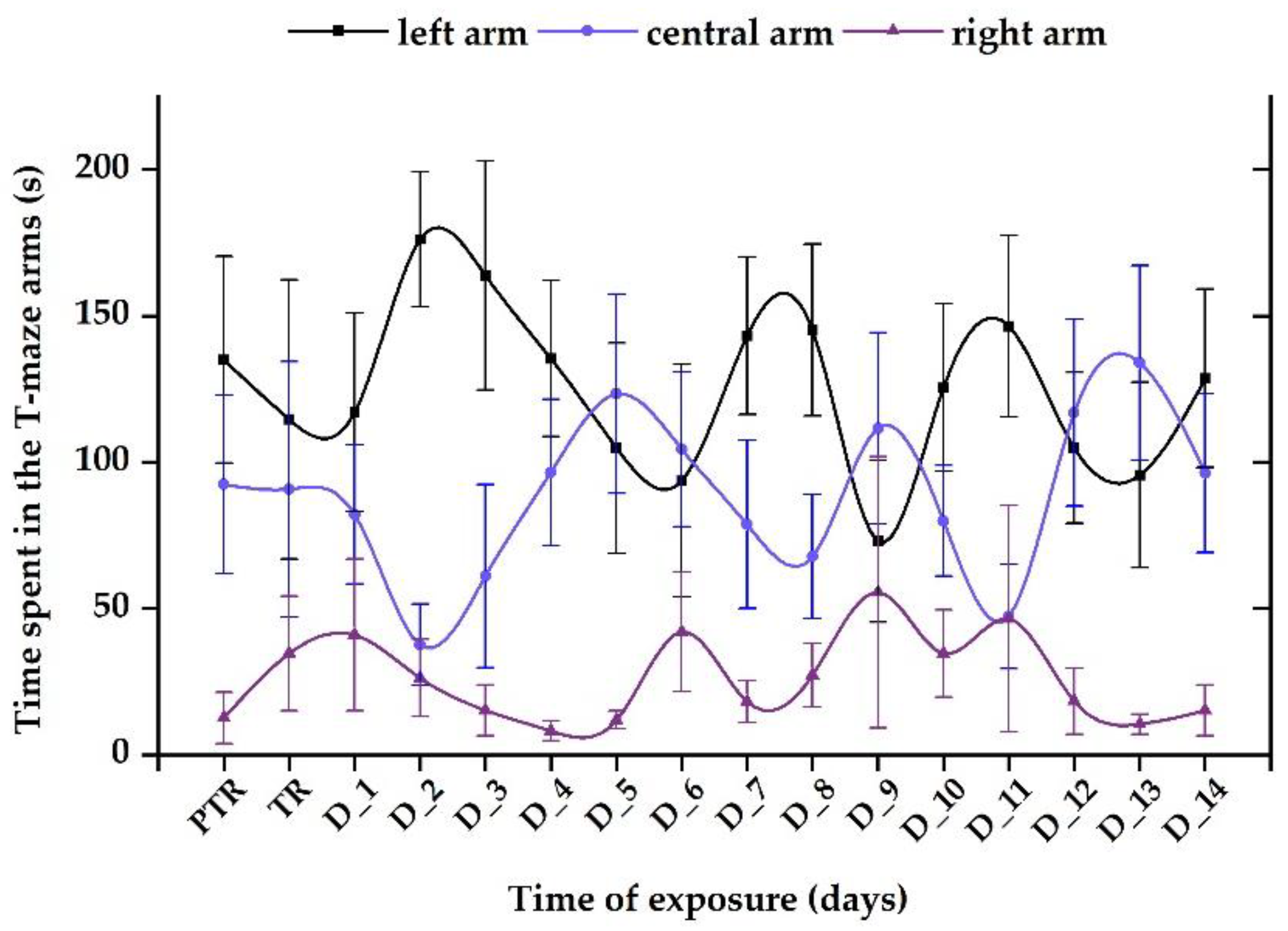
Figure 11.
The time spent in the T-maze areas, recorded for the group exposed to 0.24 μg L-1 vit. B12. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 11.
The time spent in the T-maze areas, recorded for the group exposed to 0.24 μg L-1 vit. B12. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
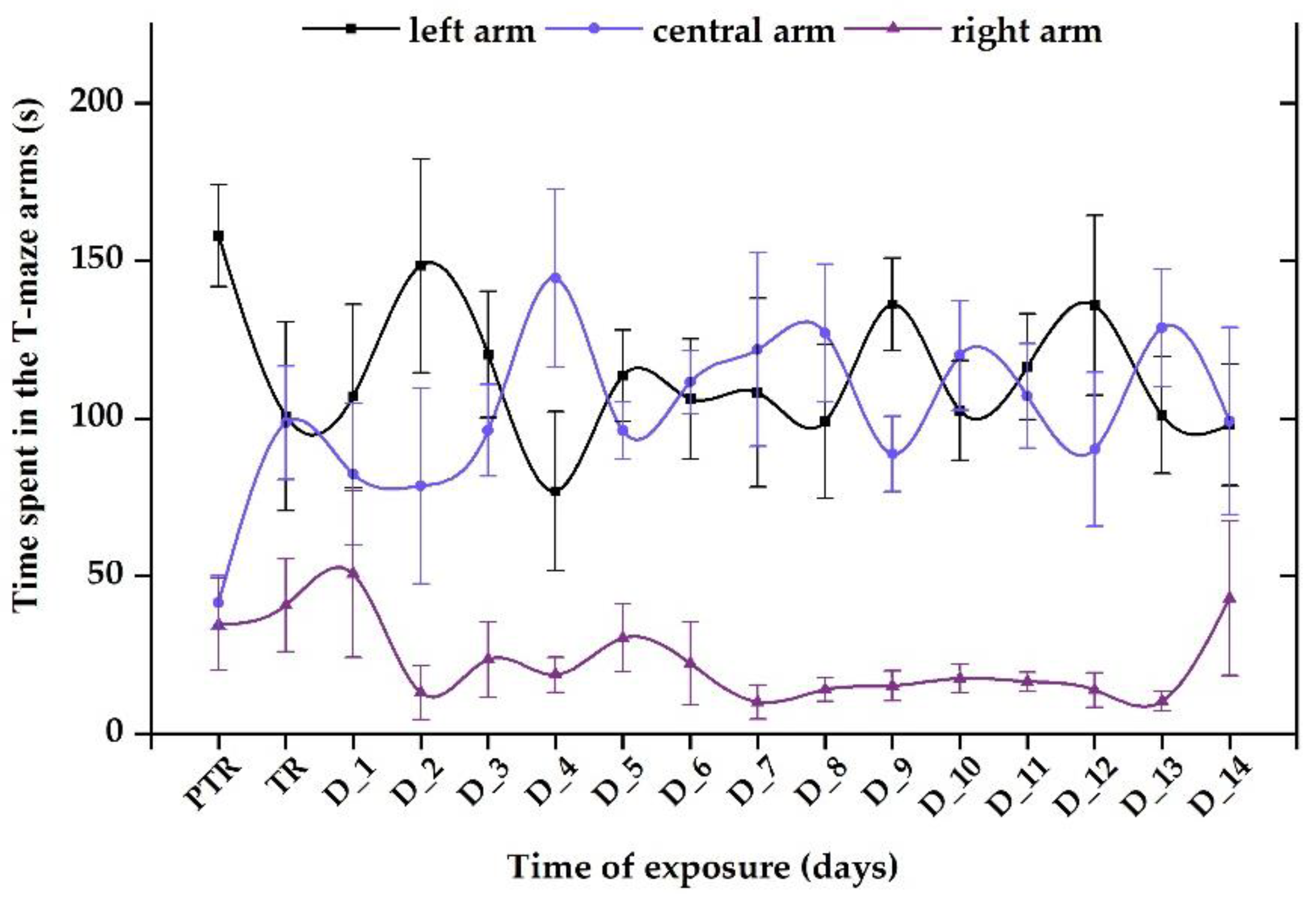
Figure 12.
The time spent in the T-maze areas, recorded for the group exposed to 600 μg L-1 FIP+PIR mixture. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 12.
The time spent in the T-maze areas, recorded for the group exposed to 600 μg L-1 FIP+PIR mixture. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
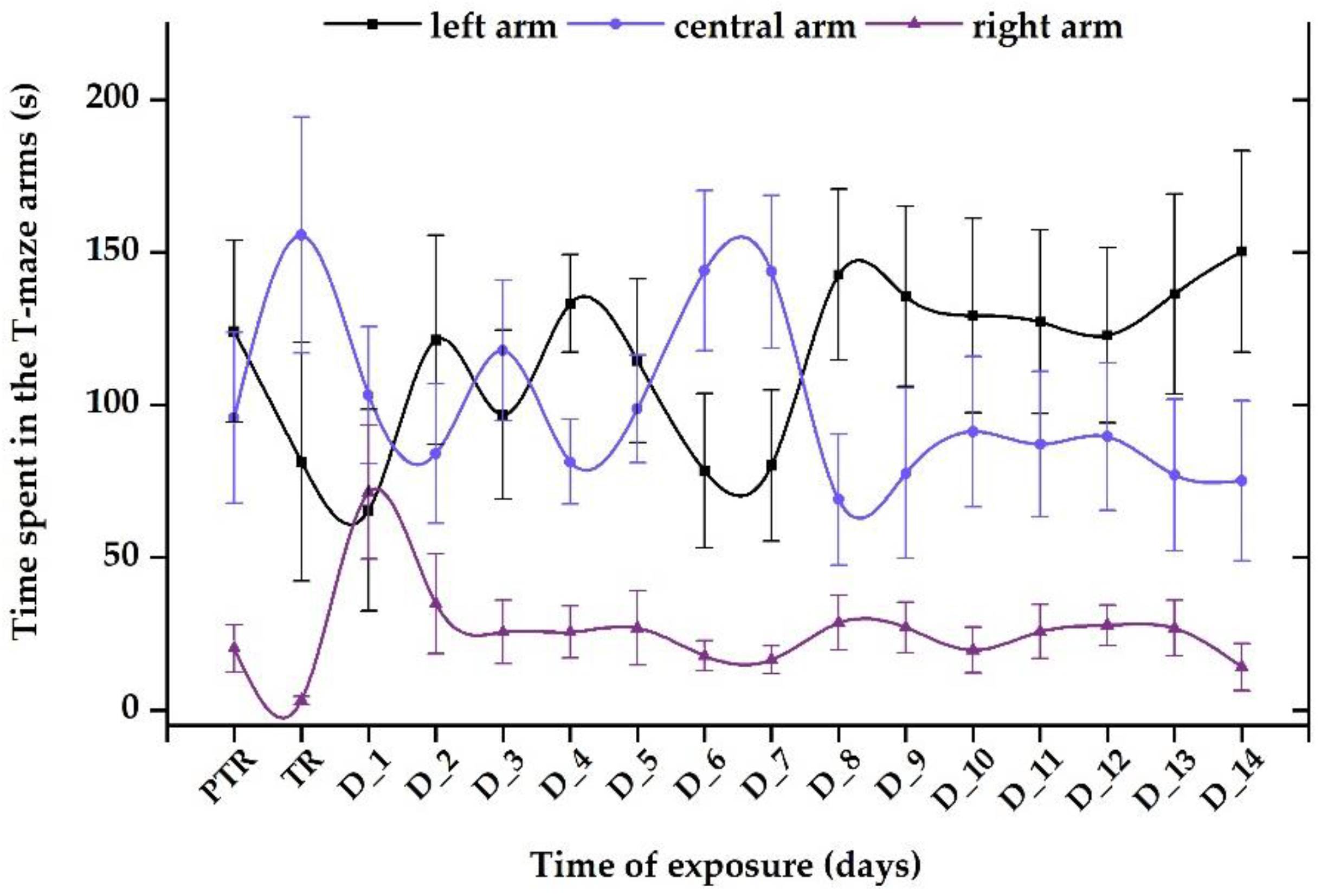
Figure 13.
The time spent in the T-maze areas, recorded for the group exposed to 0.24 μg L-1 vit. B12 and 600 μg L-1 FIP+ PIR mixture. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
Figure 13.
The time spent in the T-maze areas, recorded for the group exposed to 0.24 μg L-1 vit. B12 and 600 μg L-1 FIP+ PIR mixture. D stands for day, PTR: pretreatment, TR: treatment. The data is expressed as average ± SEM (n= 15); p< 0.05 ANOVA, Tukey is significant compared to the results from PTR stage.
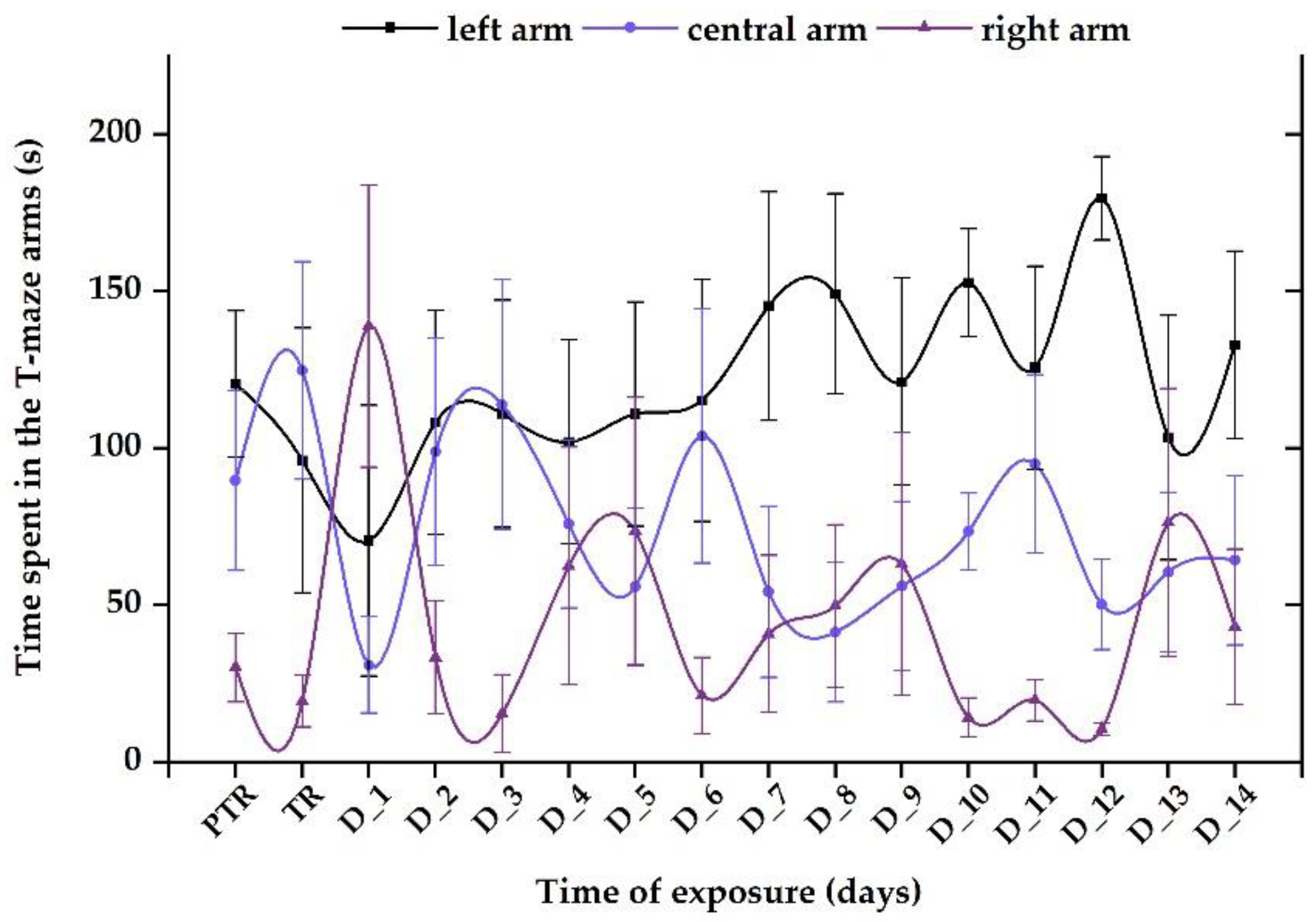
Figure 14.
The graphical representation of the activity of superoxide dismutase (SOD), glutathione peroxidase (GPx), and malondialdehyde (MDA) for the experimental groups. The data is expressed as average ± SEM; *p< 0.05 Tukey compared to the control.
Figure 14.
The graphical representation of the activity of superoxide dismutase (SOD), glutathione peroxidase (GPx), and malondialdehyde (MDA) for the experimental groups. The data is expressed as average ± SEM; *p< 0.05 Tukey compared to the control.
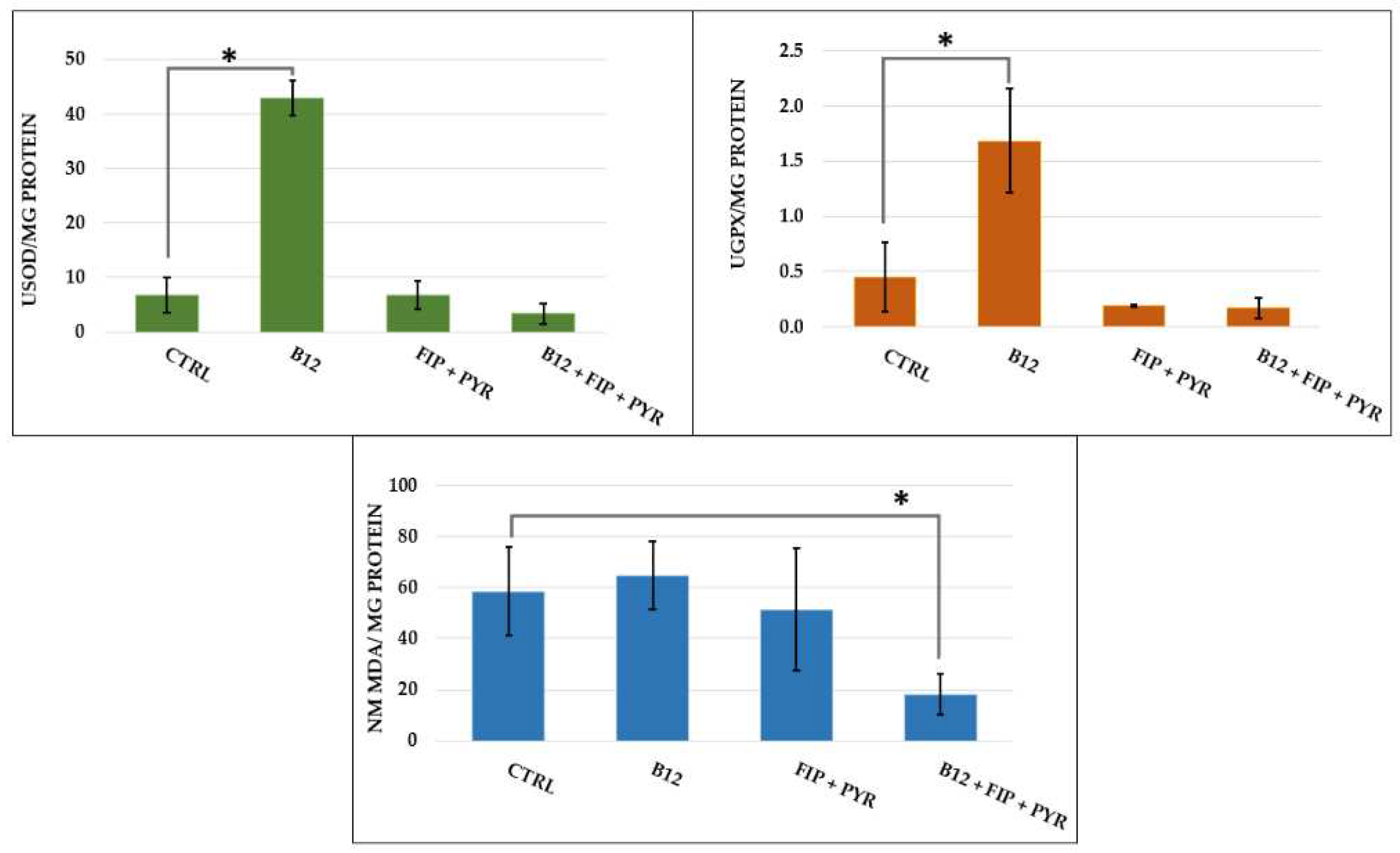

Table 1.
Environmental conditions from the housing and experimental tanks observed during the experimental period.
Table 1.
Environmental conditions from the housing and experimental tanks observed during the experimental period.
| Type of tank | Temperature (° C) |
pH | Conductivity (µS cm-1) | Salinity | Ammonia (mg L-1) |
|---|---|---|---|---|---|
| Housing tank | 26 ± 0.5 | 7.6 | 551 | 0.26 | 0.05 |
| Experimental tanks | 25 ± 0.5 | 7.5 | 553 | 0.24 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



