
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
One of the main families of antimicrobial peptides (AMPs) derived from the skin secretions of Hylidae frogs are dermaseptins. Among them, dermaseptin S4 (DS4) is characterized by its broad spectrum of activity against bacteria, protozoa and fungi. This study determined the physicochemical properties of the native peptide DS4(1-28) and its derivatives [DS4(1-28)a and DS4(1-26)a] isolated from the skin of the frog Phyllomedusa sauvagii and examined their antimicrobial properties against two marine pathogenic bacteria (Vibrio harveyi and V. anguillarum). The results indicate that DS4(1-26)a has high bacteriicdal activity against the tested strains and low hemolytic activity (> 30 % lysis at the highest tested concentration of 100 µg mL-1) compared to the other two peptides. In addition, all three peptides affected the membrane and cell wall integrity of both pathogenic bacteria, causing leakage of cellular contents, with DS4(1-26)a having the most severe impact. These properties were corroborated by tansmission electron microscopy and by the variation of cations in their binding sites due to the effect of AMPs. These results suggest that DS4 and its derivatives, particularly the truncated and amidated peptide DS4(1-26)a, could potentially be effective in the treatment of infections caused by these marine pathogenic bacteria. Future experiments are required to validate the use of DS4 in vivo in the prevention of fish bacterial diseases.
Keywords:
dermaseptin
; antimicrobial peptides
; amphibian
; aquaculture
; vibriosis
1. Introduction
In recent decades, the prevalence of drug-resistant pathogenic bacteria has surged, presenting a serious problem for the global health of both humans and livestock animals [1,2]. The Food and Agriculture Organization of the United Nations (FAO) reported that antibiotics were widely used in the aquaculture industry for their efficacy against bacterial infections. However, over the last decade, antibiotic residues (about 40-90%) have been detected in animals, due to their massive addition as growth promoters, surpassing their use as therapeutic agents for fish [3]. Consequently, this can have adverse effects on human health and the ecosystem in general [4]. Indeed, prolonged exposure of bacteria to antibiotics reinforced the selection of superbugs, facilitated by the horizontal transfer of their drug-resistant genes (horizontal gene transfer, HGT) between pathogenic and non-pathogenic bacteria [5]. In addition, these residues can have ecotoxic effects on humans through the consumption of agricultural products [6], exacerbating the critical state of fisheries [7]. Therefore, it is now urgent to search for alternative compounds to antibiotics that are effective against bacteria and safe for humans and the environment [8]. Currently, some most promising alternatives include acidifiers, probiotics, prebiotics and enzymes [9]. However, a promising strategy involves the use of antimicrobial peptides (AMPs) due to their potential in combating antibiotic resistance [10]. AMPs, also known as host defense peptides, are short and generally positively charged peptides, present in a wide variety of organisms, from microorganisms to plants and insects [11], fish [12], amphibians and humans [13]. Ranging from 10 to 50 residues, AMPs exhibit cationic and amphipathic structures. All these characteristics allow them to interact with the bacterial membrane through different mechanisms [14].
Among AMPs with a potent antimicrobial action, amphibian peptides are special due to their structural diversity provided by the lifestyle and habitat of these animals [15]. In particular, frog skin contains interesting AMPs, including dermaseptins [16]. Dermaseptins belong to the family of AMPs extracted from the skin of the frog Phyllomedusa sauvagii and typically range between 28 and 34 amino acids (aa) in length [17,18]. These sequences contain a conserved tryptophan (Trp) residue at position three, C-terminal amidation and a highly repeated AAA/GKAAL/G/NA pattern [16]. In an amphiphilic medium, they can form an α-helicoidal conformation, facilitating their interaction with bacterial lipid bilayers [19]. They are characterized by a positive charge, which plays a key role in the interaction with the negatively charged membrane [20]. This explains their broad microbicidal spectrum [21] since dermaseptins exhibit in vitro activity against bacteria, parasites, and fungi [22]. Consequently, changes in their positive charge can affect their antimicrobial activity. Similar to their charge, many studies have indicated that changes in the number of residues of AMPs can alter their activity and hydrophobicity [23]. Available studies illustrate the effective mechanism of action of dermaseptin S4 (DS4), characterized by destabilization of cell membranes, leakage of contents to the outside, depolarization and the possibility of causing bacterial death [24]. In addition to their high anticancer and antimicrobial activity, they possess low hemolytic activity for mammalian red blood cells [25,26]. To further understand the properties and functions of dermaseptins, DS4 peptide with 28aa, derived from the frog P. sauvagii, and its modified derivatives at the C-terminal were used in the present study [27]. This study aimed to explore the therapeutic use of DS4 peptide and its derivatives in fish. In this way, we examine their expected powerful mechanism of action against marine pathogenic bacteria (V. harveyi and V. anguillarum), and their hemolytic impact was assessed on red blood cells of gilthead seabream (Sparus aurata), one of the most important marine farmed fish in the Mediterranean area.
2. Results
2.1. Properties and structural analysis of the DS4 peptide and its derivatives
The physicochemical characteristics of the three DS4 peptides are presented in Table 1. In fact, compared to the native peptide named DS4(1-28), both analogs showed some modifications such as truncation and amidation on the C-terminal side. Moreover, both peptide analogs showed a positive charge (+5) higher than that of the parent molecule (+4), caused by this amidation. Furthermore, DS4(1-26)a has an aliphatic index (AI) of 154.32 which is superior to the other two, as it has a high hydrophobic index of 73% and has a positive value (1.17) for general average hydropathicity (GARVY). This is an indication of the thermal stability of this peptide in comparison to other peptides, such as DS4 (1-28) and DS4 (1-28)a. The predicted secondary structure of the DS4 peptide is an α-helical structure (78.57%) (Figure 1a).
The helical diagram of DS4 (1-28) and DS4(1-26)a respectively, indicated that the location of the positive charge, due to the presence of four lysines (K) (in blue) is located on the near N-terminal face, while hydrophobic residues such as alanine (A), leucine (L) and valine (V) are distributed near the C-terminal end (in yellow and grey) giving rise to hydrophobic spots (arrow) (Figure 1b, c).
2.2. Antibacterial activity
The three tested peptides exhibited strong antibacterial activity. With a MIC range of 3.03 to 3.84 µg mL-1 for V. harveyi and V. anguillarum, DS4(1-26)a was the most active, while DS4(1-28)a had 4.84 and 4.94 for V. harveyi and V. anguillarum respectively, and DS4(1-28) had 9 and 5.67 for V. harveyi and V. anguillarum, respectively. In addition, all peptides showed higher bactericidal activity when used at concentrations above 12 µg mL-1, for both bacteria. Finally, the antimicrobial activity of the three peptides was consistently higher against V. harveyi than against V. anguillarum, even at low concentrations (3 µg mL-1) (Figure 2).
2.3. Antibacterial mechanisms of DS4 and its derivatives
Cell membrane integrity was assessed by nucleic acid leakage. Nucleic acid optical density values of both bacteria (V. harveyi and V. anguillarum) augmented significantly (p < 0.01) with increasing AMPs concentration (from 1 to 2 MIC) used in the assays. Such leakage was dose-dependent, and it was higher when bacteria were incubated with the higher AMP concentration (Figure 3 and Figure 4).
The impact of AMPs on bacterial cell wall permeability was determined through AKPase activity. After 6 h of incubation of the bacteria with AMPs, the extracellular AKPase activity of both bacteria (V. harveyi and V. anguillarum) was significantly increased compared to the activity determined in control samples. Indeed, extracellular AKPase activity increased after 3h of incubation with DS4(1-26)a and DS4(1-28)a in both bacteria compared to the native DS4(1-28) peptide, which started to increase after 5h of incubation (Figure 5).
In addition, transmission electron microscopy was used to observe possible ultrastructural alterations caused by DS4 in the bacterial cells. In the control samples (bacteria not incubated with AMPs), the appearance of both bacteria was similar, showing a continuous cell wall with no evident alterations. Both bacteria had the typical bacillus shape and a flagellum (Figure 6).
However, both bacteria lost their structure and curiously, some of them also the flagellum, due to cationic AMPs. This caused ruptures in the cell wall of V. anguillarum. In the case of V. harveyi, cells underwent a significant change in shape as the formation of hollows after being incubated with AMPs (Figure 7 and Figure 8).
2.4. Effect of divalent cations on peptide–membrane interaction
The cation displacement assay also provides information on the mode of action of the peptides studied. The antimicrobial activity of peptides against V. harveyi and V. anguillarum was influenced by different salt ions (20 µg Ca2+/Mg2+), but they were always effective in physiological environments with salt. All peptides had a MIC value that was approximately fourfold and six-fold higher against V. harveyi and V. anguillarum, respectively. Among the peptides, DS4(1-26)a was particularly more resistant to physiological salts than other peptides in both bacteria, and the native peptide DS4(1-28) appeared to be the most susceptible to cations (Figure 9).
2.5. Hemolytic activity
The hemolytic activity of the three AMPs was studied against pig and fish red blood cells (Figure 10). DS4 (1-28)a, and DS4 (1-26)a exhibited less than 30% hemolysis for fish red blood cells, even at the maximum concentration of 100 µg mL-1. However, the native peptide showed more than 30% hemolysis at the same concentration. Notably, only 4-5% hemolysis was observed at 3 µg mL-1. However, for the pig's red blood cells, all peptides demonstrated around 60% hemolysis, with DS4 (1-26)a showing again the lower hemolytic activity of the three AMPs.
3. Discussion
In the present study, we evaluated the antibacterial properties against V. harveyi and V. anguillarum of three antimicrobial peptides obtained from DS4. These bacteria were selected because Vibrio is an opportunistic and ubiquitous bacterium in the marine environment [39,40] causing serious diseases in fish [41,42]. Outbreaks of vibriosis can directly cause fish kills or can slow the growth of farmed marine fish. Both situations lead to decreased production and significant economic losses [43,44]. In addition, another problem related to these bacteria is that it is necessary to use antibiotics to combat vibriosis. The prolonged and sometimes excessive use of antibiotics has led to the emergence and development of resistance phenomena in many bacteria [45]. Currently, the discovery of new antibacterial agents can help to alleviate or solve this serious problem of antibiotic resistance, which poses a great threat to human health and the health of ecosystems. At present, the discovery of new antibacterial agents can solve this resistance problem [46]. According to previous research, the dermaseptin family is an important source of AMPs [47], characterized by a broad spectrum of activity against various microorganisms including bacteria [48], protozoa, viruses and fungi [49]. In this work, we have focused on three DS4 peptides [DS4(1-28), DS4(1-28)a and DS4(1-26)a]. These peptides belong to an AMP family isolated from the skin of P. sauvagii frog [50] and could be considered potential therapeutic agents. This makes our research significant in exploring new avenues for drug development. It has been suggested that the size, net charge, and amphipathic structure adjust the biological activity of the AMPs by allowing them to bind, insert and destabilize the membrane of pathogenic bacteria [51]. Due to the inclusion of certain residues (arg / lys / hist), in the current work, both analog peptides DS4(1-28)a and DS4(1-26)a have a positive charge of +5 compared to the original molecule, which has a +4 charge [52]. In fact, the C-terminal amidation was responsible for increasing the charge of the peptide [53].Additionally, all AMPs have an aliphatic index greater than 100 as proteins from thermophilic organisms, ensuring the stability of AMPs over a wide temperature spectrum. In particular, DS4(1-26)a exhibited the highest aliphatic index of 154.23 compared to other peptides [54]. Moreover, the instability index is less than 40, which means that it is a stable AMP [55,56]. All of these physicochemical characteristics promote electrostatic interactions between the AMPs and the surface of bacteria [24]. To better understand these interactions, in the present study, the antimicrobial activity of the AMPs was directly measured calculating the MIC of each AMPs. We found that an increase in the AMPs concentration is accompanied by an increase in the antimicrobial activity against the two pathogenic bacteria assayed (lower MIC; minimum inhibitory concentrations). Therefore, all AMPs showed potent antibacterial activity in vitro against the strains tested. Among them, DS4(1-26)a was found to be much more active than the native molecule. This suggests that cationicity favors the initial electrostatic interaction between AMPs and the anionic bacterial membrane [57]. Furthermore, a low ability to destabilize erythrocyte membranes of gilthead seabream was detected after being incubated with the peptides compared to pigs red blood cells. This is not surprising because it is known that there is a similar structure between certain fish peptides and the peptide dermaseptin. One of such peptide is the pleurocidin, extracted from the skin mucous secretions of the winter flounder Pleuronectes americanus, which shares homology with frog dermaseptin [58]. The results suggest that it could be beneficial to use DS4(1-26)a in marine fish aquaculture. Furthermore, these results are in agreement with those found for cathelicidin (HR-CATH), and dermaseptin-PS3 (in which a double lysine residue was introduced at positions 5 and 17), peptides that also have high antimicrobial activity against Gram-positive bacteria and V. parahaemolyticus, with low levels of hemolysis activity [59,60,61]. These results confirm the property more extended and characteristic of the AMPs, which is their bactericidal activity [62].
Previous studies have indicated that AMPs act on the membrane of bacteria causing nucleic acid leakage through the cell wall, which ultimately destroys the bacteria [63,64]. This is consistent with our results, which show that AMPs caused a transformation of the bacterial membrane permeability and integrity. It is known that one of the main mechanisms of antimicrobial drugs is to damage cell membrane integrity and inhibit nucleic acid synthesis [65]. To better understand the antimicrobial action of DS4 and demonstrate the permeability of bacteria's cell wall after incubation with AMPs, the study of AKPase was conducted. The AKPase results show that the cell membrane was damaged, which leads to an increase in cell wall permeability. After incubating both bacteria with AMPs, the extracellular content of AKPase rapidly increased compared to the values obtained for control bacteria (not incubated with AMPs). These results are consistent with those obtained with the peptide Brevinin-1 from the frog Hydrophylax bahuvistara, which caused bacterial death by altering the permeability of the membrane of V. cholerae [66]. It has been already confirmed for many frog AMPs that they cause degradation of the bacterial cell membrane and changes in the cell morphology [67,68]. For this reason, the transmission electron microscopy technique was used to further analyze the effects of AMPs on the bacterial membrane and wall. Typically, natural AMPs concentrate their primary aim on the cell membrane [69]. Gram-negative bacteria possess an outer membrane that contains negatively charged phospholipids (phosphatidylserine and phosphatidylglycerol), which help to attach cationic AMPs [70]. The peptide is then introduced into the lipid bilayer of the cytoplasmic membrane, causing disruption of membrane integrity and permeability or pore formation [69,71]. Furthermore, our results indicate that all cationic peptides exhibit the ability to damage membranes and alter their morphology by formation of bulge and/or pore depending on the bacteria used [72,73]. Therefore, these results supported previous research and demonstrated the varying effects of dermaseptin on different types of Vibrio-bacteria. Likewise, to better understand the mode of action of the AMPs studied, we performed the cation displacement assay. Mg2+ and Ca2+ cations have binding sites on the membrane of lipopolysaccharide located in Gram-negative bacteria [74]. The potential negative impact of these cations on AMPs was estimated in both bacteria. Indeed, the activity of all peptides was decreased for the two tested bacteria when incubated with the highest concentration of both cations. Similarly, it has been shown that peptides, s-thanatin and β-defensins HD-5 are salt sensitive and lose their activity in the presence of cations [37,75]. The above results show that these antagonisms are the result of competitive blockade. Bacterial membrane permeability and fluidity decrease due to degradation of the membrane structure (inner and outer) by the movement of these ions (such as Ca 2+ and Mg 2+) from their binding sites on LPS. The same results were observed with the peptide brevinin1, which is considered to be salt-resistant and could be a potent molecule for drug development [66]. Many previous studies have shown that the cationic peptide caerin1 (from frog skin) is more effective, not only against pathogenic bacteria, but also against fish viruses, than a fish-derived AMP, dicentracin [76]. Furthermore, the human peptide LL-37 and the insect peptide LSB-37 tested in vitro and in vivo showed higher activity against fish pathogenic bacteria [10,77]. Based on our results, we found that all peptides were capable of killing the two tested marine pathogenic bacteria. The present results agree with previous research and opens the door to their possible use against virus. Among the tested peptides, DS4(1-26)a displayed the highest activity level. All this makes this peptide a good candidate for future in vivo studies that support these results.
4. Materials and Methods
4.1. Peptides synthesis and physicochemical properties
Dermaseptin S4 (DS4) peptide and its derivatives [DS4(1-28), DS4(1-28)a, and DS4(1-26)a] were obtained using the previously reported method [28]. Briefly, the box strategy and teabag methodology were used to perform the solid-phase peptide synthesis in parallel. First, 100 mg of p-methylbenzdrylamine (pMBHA) resin per peptide was sealed in a mesh tea bag, neutralized with 5 % diisopropylethylamine (DIEA) in dichloromethane (DCM), and then swollen with additional DCM washes. In the presence of diisopropylcarbodiimide (DIC, 6 eq.), for 60 min the first amino acid was introduced by coupling the amino acid Boc (6 eq) in dimethylformamide (DMF 0.1 M). After 30 min the Boc protecting group was removed with 55 % TFA/DCM and then neutralized with 5 % DIEA/DCM (3x). The final products were cleaved for 1.5 h from the resin support in the presence of HF/anisole (95:5) at 0 °C. The products were extracted with 95 % acetic acid for 1 h, transferred to a vial and lyophilized after freezing and removal of HF. Freezing and lyophilization were repeated three times, after which the samples were transferred to a 50/50 mixture of acetonitrile and water in pre-weighed vials. Finally, the samples were reweighed, and their crude masses were recorded after drying.
The physicochemical properties of DS4 and its derivatives, such as molecular weight, net charge, hydrophobicity, aliphatic number (AI), and instability index (II) were determined with the APD3 antimicrobial peptide database (http://aps.unmc.edu/AP/). Furthermore, the organization of amino acids in the alpha-helix was performed with HELIQUEST (http://heliquest.ipmc.cnrs.fr/) [29]. The secondary structure of DS AMP was predicted with PEP-FOLD3 (http://bioserv.rpbs.univ-paris-diderot.fr/services/PEP-FOLD3) [30], and with the self-optimized prediction method with alignment (SOPMA) (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_server.html) [31].
4.2. Antibacterial assay
Antimicrobial activity was evaluated against V. harveyi (isolated from infected European seabass) and V. anguillarum (ATCC 19264). Bacteria were grown on Tryptic Soy Agar (TSA, Difco Laboratories) at 25 ºC, and then inoculated in liquid Tryptic Soy Broth (TSB, Difco Laboratories). Both media were supplemented with NaCl to a final concentration of 1.5 % (w/v). The number of bacteria was adjusted to 106 colonies forming units (c.f.u.) mL-1 after estimation by optical density at 620 nm against a standard curve [32].
The minimum inhibitory concentration (MIC) was defined as the lowest peptide concentration that inhibits bacterial growth and was determined by microdilution methods. Briefly, in a 96-well microplate, 50 µL aliquots of bacterial suspension were previously adjusted to 106 c.f.u. mL-1 were added per well, followed by 50 µL of different peptides [DS4(1-28), DS4(1-28)a, and DS4(1-26)a] added at different concentrations (3, 25, 50 and 100 µg mL-1 in TSB medium). The medium with bacteria was used as a positive control and the medium without peptides as a negative control. The plate was incubated at 25 ºC for 18 h. MIC was determined by measuring OD at 620 nm (BMG, SPECTROstarNano). Three runs were performed on three different days as replicates [33]. Percent inhibition (PI) was determined by the following equation:
PI=1-(DO test/DO control) *100
4.3. Effects of DS4 and its derivatives on cell membrane integrity and cell wall permeability
When cellular contents leak, it indicates that the cell membrane has been damaged. Nucleic acid leakage was used to estimate the cell membrane integrity of V. harveyi and V. anguillarum using the time-varying 260 nm UV absorption method (from 1, 3, 5 and 7 h). For this purpose, native peptide and its derivatives at 0, MIC, 1.5 MIC, and 2 MIC, were added to a final bacterial suspension adjusted to 106 c.f.u. mL-1, with PBS alone or with PBS plus 3 µM bovine serum albumin (BSA), for negative and positive controls, respectively [34].
To assess the permeability of the bacterial cell wall, the bacteria were adjusted to 106 c.f.u. mL-1 and then added to each peptide solution at a final concentration of MIC, 1.5 MIC, or 2 MIC. After incubation at different times (from 1 to 6 h), the samples were centrifuged (12000 rpm, 5 min). Bacterial supernatants were collected each time to measure extracellular alkaline phosphatase (AKPase) activity with the AKPase detection kit. Samples with bacteria but without peptides were used as negative control [35].
The morphology of bacteria with peptides was examined using transmission electron microscopy. V. harveyi and V. anguillarum (106 c.f.u. mL-1) were incubated with each peptide at MIC for 120 min at 25 ºC. The samples were then centrifuged (400 x g, 10 min, 4 ºC), fixed with 2.5% glutaraldehyde in a 0.1 M cacodylate buffer, pH 7.2-7.4 (4 ºC, 1h), post-fixed in 1 % OsO4 for 2 h and embedded in Epon-812 [36]. Ultrathin sections were counterstained with uranyl acetate and lead citrate and examined under a transmission electron microscope (JEOL 1011).
4.4. Effects of divalent cations on peptide–membrane interaction
The ability of DS4 AMPs to bind divalent cations, localized on the bacterial cell membrane, was measured by determining the MIC of each peptide in the TSB supplemented with 20 µg of Mg2 + and Ca2+. The bacterial cells were incubated with all of the peptides at the MIC (at 106 c.f.u. mL-1 in TSB containing the cations) for 24 h at 25 °C. Standard samples without ions were used as controls [37].
4.5. Ethics statement
Ethical approval for all experimental procedures realized with fish was obtained from the Ethics Committee of the University of Murcia (protocol code A13150104) following the European Union guidelines for animal handling (2010/63/EU).
4.6. Hemolytic activity of DS4 and its derivatives
Fresh blood samples were obtained from pigs (D. gaditano) at a local butchery (Murcia, Spain) for the hemolysis test. Plasma was separated after centrifugation (500 x g, 5 min, 4 °C), and erythrocytes were washed three times with PBS. Similarly, gilthead seabreams (S. aurata L.), with a mean weight of 250 g and a mean length of 21 cm were obtained from a local farm (Murcia, Spain). These specimens were placed in recirculating seawater aquaria (450 L) at the Marine Fish Facility of the University of Murcia for a quarantine period of one month. Environmental conditions included a water temperature of 20 ± 2ºC, a flow rate of 900 L h-1, a salinity of 28 ‰, a photoperiod of 12 h light to 12 h dark and continuous aeration. The tank water was carefully maintained to keep ammonium and nitrite levels below species-specific limits (0.1 mg L-1 and 0.2 mg L-1, respectively). Fish were fed a commercial diet (Skretting, Spain) at a rate of 2% of their body weight per day.
For erythrocyte isolation, six fish were randomly selected and anesthetized with clove oil (20 mg L-1, Guinama®). Subsequently, 1 mL of blood samples were drawn from the tail vein using a heparinized syringe. These samples were immediately placed in 7 mL of PBS (phosphate buffered saline) containing 0.35% sodium chloride to adjust the osmolarity of the medium, along with 10 mM glucose (termed PBS-glu). The fish were then returned to the tank. The blood was layered on a 51 % Percoll density gradient (Pharmacia) and centrifuged (400 x g, 30 min, 4 °C) to separate leucocytes from erythrocytes.
Erythrocyte suspensions (from pigs and gilthead seabream) at 1% were incubated with various concentrations of different peptides (3, 25, 50 and 100 µg mL-1) for 1 h at 28 °C and absorbance was then measured at 540 nm. PBS and 1 % Triton X-100 were used as negative and positive controls, respectively. Each assay and condition were repeated in triplicate [38]. The percent lysis was calculated using the following formula:
% of lysis = (As− APBS)/ (ATriton− APBS) × 100
4.7. Statistical study
All results are presented as mean ± standard error of the mean (SEM). Differences between each group were obtained by Student's t-test, and two-way ANOVA (followed by Tukey's post-hoc analysis). Shapiro-Wilk and Levene's tests were used to determine normality and homogeneity of variance, respectively. Non-normally distributed data were log-transformed before analysis. The U-Mann-Whitney test was used when data did not meet parametric assumptions. Statistical tests were performed with SPSS 24 software. For all statistical analyses, the significance level used was p < 0.05.
5. Conclusions
To conclude, this study examines the mode of action of three types of dermaseptin S4, AMPs identified from the skin secretions of P. sauvagii. The interaction between AMPs and bacteria was found to show potent antibacterial activity through the disruption of cell membranes and cell walls. The divalent cations Mg 2+ and Ca 2+ lose their binding sites in Gram-negative bacteria after peptide administration. Even at high concentrations, these peptides have little hemolytic effect on fish erythrocytes. Moreover, dermaseptin and other AMPs can be utilized to prevent infections in aquaculture species. Nonetheless, efficient methods for their administration are needed to be addressed. Before using this therapy strategy, it is necessary to research how the peptides affect other environmental bacteria.
Author Contributions
A.B. and J.A.S.D. performed the experiments and wrote the draft of the manuscript. A.N. synthesized all the peptides, supervise the assays. S.A. data curation, validation and supervised the work of A.B. M.A.E. conceived and designed the study, got the funding and was responsible for the overall study coordination of this manuscript. All authors read and approved the final manuscript.
Funding
This work was funding by the proyecto PID2020- 113637RB-C21 de investigación financiado por MCIN/AEI/10.13039/ 501100011033.
Institutional Review Board Statement
Ethical approval for all experimental procedures realized with fish was obtained from the Ethics Committee of the University of Murcia (protocol code A13150104) following the European Union guidelines for animal handling (2010/63/EU).
Informed Consent Statement
Not applicable.
Acknowledgments
JASD has a PhD grant (FPU19/02192) and it forms part of the ThinkInAzul program and was supported by MCIN with funding from European Union Next Generation EU (PRTR-C17.I01) and by Comunidad Autónoma de la Región de Murcia, Fundación Séneca.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, J.; Buhe, C.; Yu, D.; Zhong, H.; Wei, Y. Ammonia Stress Reduces Antibiotic Efflux but Enriches Horizontal Gene Transfer of Antibiotic Resistance Genes in Anaerobic Digestion. Bioresource Technology 2020, 295, 122191. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. The Drug-Resistant Bacteria That Pose the Greatest Health Threats. Nature 2017, 543, 15–15. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current Status of the Use of Antibiotics and the Antimicrobial Resistance in the Chilean Salmon Farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wang, Y.; Yuan, Y.; Xie, Y. A Systematic Review on Antibiotics Misuse in Livestock and Aquaculture and Regulation Implications in China. Science of The Total Environment 2021, 798, 149205. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.; Domingues, S.; Da Silva, G.J. Manure as a Potential Hotspot for Antibiotic Resistance Dissemination by Horizontal Gene Transfer Events. Veterinary Sciences 2020, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Rasul, M.G.; Majumdar, B.C. Abuse of Antibiotics in Aquaculture and It’s Effects on Human, Aquatic Animal and Environment. The Saudi Journal of Life Sciences 2017, 2, 81–88. [Google Scholar]
- Rossi, B.; Esteban, M.A.; García-Beltran, J.M.; Giovagnoni, G.; Cuesta, A.; Piva, A.; Grilli, E. Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio Spp.: An In Vitro Study. Microorganisms 2021, 9, 966. [Google Scholar] [CrossRef] [PubMed]
- Manniello, M.D.; Moretta, A.; Salvia, R.; Scieuzo, C.; Lucchetti, D.; Vogel, H.; Sgambato, A.; Falabella, P. Insect Antimicrobial Peptides: Potential Weapons to Counteract the Antibiotic Resistance. Cell. Mol. Life Sci. 2021, 78, 4259–4282. [Google Scholar] [CrossRef]
- Thacker, P.A. Alternatives to Antibiotics as Growth Promoters for Use in Swine Production: A Review. J Animal Sci Biotechnol 2013, 4, 35. [Google Scholar] [CrossRef]
- De Sousa, E.L.; Assane, I.M.; Santos-Filho, N.A.; Cilli, E.M.; De Jesus, R.B.; Pilarski, F. Haematological, Biochemical and Immunological Biomarkers, Antibacterial Activity, and Survival in Nile Tilapia Oreochromis Niloticus after Treatment Using Antimicrobial Peptide LL-37 against Streptococcus Agalactiae. Aquaculture 2021, 533, 736181. [Google Scholar] [CrossRef]
- Mudalungu, C.M.; Tanga, C.M.; Kelemu, S.; Torto, B. An Overview of Antimicrobial Compounds from African Edible Insects and Their Associated Microbiota. Antibiotics 2021, 10, 621. [Google Scholar] [CrossRef] [PubMed]
- Raju, S.V.; Sarkar, P.; Kumar, P.; Arockiaraj, J. Piscidin, Fish Antimicrobial Peptide: Structure, Classification, Properties, Mechanism, Gene Regulation and Therapeutical Importance. Int J Pept Res Ther 2021, 27, 91–107. [Google Scholar] [CrossRef]
- Fabisiak, A.; Murawska, N.; Fichna, J. LL-37: Cathelicidin-Related Antimicrobial Peptide with Pleiotropic Activity. Pharmacological Reports 2016, 68, 802–808. [Google Scholar] [CrossRef]
- Aronica, P.G.A.; Reid, L.M.; Desai, N.; Li, J.; Fox, S.J.; Yadahalli, S.; Essex, J.W.; Verma, C.S. Computational Methods and Tools in Antimicrobial Peptide Research. J. Chem. Inf. Model. 2021, 61, 3172–3196. [Google Scholar] [CrossRef] [PubMed]
- König, E.; Bininda-Emonds, O.R.P.; Shaw, C. The Diversity and Evolution of Anuran Skin Peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Structural Diversity and Species Distribution of Host-Defense Peptides in Frog Skin Secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xi, X.; Lu, Y.; Hu, H.; Dong, Z.; Ma, C.; Wang, L.; Zhou, M.; Chen, T.; Du, S.; et al. In Vitro Activities of a Novel Antimicrobial Peptide Isolated from Phyllomedusa Tomopterna. Microbial Pathogenesis 2021, 153, 104795. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Fang, H.; Chen, X.; Wang, L.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Exploration of the Structure–Function Relationships of a Novel Frog Skin Secretion-Derived Bioactive Peptide, t-DPH1, through Use of Rational Design, Cationicity Enhancement and In Vitro Studies. Antibiotics 2021, 10, 1529. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ding, X.; Li, W.; Lu, T.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burden, R.; Chen, T. Discovery of Two Skin-Derived Dermaseptins and Design of a TAT-Fusion Analogue with Broad-Spectrum Antimicrobial Activity and Low Cytotoxicity on Healthy Cells. PeerJ 2018, 6, e5635. [Google Scholar] [CrossRef]
- Henderson, J.M.; Iyengar, N.S.; Lam, K.L.H.; Maldonado, E.; Suwatthee, T.; Roy, I.; Waring, A.J.; Lee, K.Y.C. Beyond Electrostatics: Antimicrobial Peptide Selectivity and the Influence of Cholesterol-Mediated Fluidity and Lipid Chain Length on Protegrin-1 Activity. Biochimica et Biophysica Acta (BBA) - Biomembranes 2019, 1861, 182977. [Google Scholar] [CrossRef]
- Li; Xi; Ma; Chen; Zhou; Burrows; Chen; Wang A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa Tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities. Biomolecules 2019, 9, 628. [CrossRef]
- Dos Santos, C.; Hamadat, S.; Le Saux, K.; Newton, C.; Mazouni, M.; Zargarian, L.; Miro-Padovani, M.; Zadigue, P.; Delbé, J.; Hamma-Kourbali, Y.; et al. Studies of the Antitumor Mechanism of Action of Dermaseptin B2, a Multifunctional Cationic Antimicrobial Peptide, Reveal a Partial Implication of Cell Surface Glycosaminoglycans. PLoS ONE 2017, 12, e0182926. [Google Scholar] [CrossRef] [PubMed]
- Schmidtchen, A.; Pasupuleti, M.; Malmsten, M. Effect of Hydrophobic Modifications in Antimicrobial Peptides. Advances in Colloid and Interface Science 2014, 205, 265–274. [Google Scholar] [CrossRef]
- Schmidt, N.W.; Wong, G.C.L. Antimicrobial Peptides and Induced Membrane Curvature: Geometry, Coordination Chemistry, and Molecular Engineering. Current Opinion in Solid State and Materials Science 2013, 17, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Hou, X.; Wang, L.; Gao, Y.; Wu, D.; Xi, X.; Zhou, M.; Kwok, H.; Duan, J.; Chen, T.; et al. Two Novel Dermaseptin-Like Antimicrobial Peptides with Anticancer Activities from the Skin Secretion of Pachymedusa Dacnicolor. Toxins 2016, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A. Antimicrobial Peptides from Frog Skin Biodiversity and Therapeutic Promises. Front Biosci 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed]
- Lorin, C.; Saidi, H.; Belaid, A.; Zairi, A.; Baleux, F.; Hocini, H.; Bélec, L.; Hani, K.; Tangy, F. The Antimicrobial Peptide Dermaseptin S4 Inhibits HIV-1 Infectivity in vitro. Virology 2005, 334, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Belaid, A.; Braiek, A.; Alibi, S.; Hassen, W.; Beltifa, A.; Nefzi, A.; Mansour, H.B. Evaluating the Effect of Dermaseptin S4 and Its Derivatives on Multidrug-Resistant Bacterial Strains and on the Colon Cancer Cell Line SW620. Environ Sci Pollut Res 2021, 28, 40908–40916. [Google Scholar] [CrossRef] [PubMed]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A Web Server to Screen Sequences with Specific α-Helical Properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Liscano, Y.; Medina, L.; Oñate-Garzón, J.; Gúzman, F.; Pickholz, M.; Delgado, J.P. In Silico Selection and Evaluation of Pugnins with Antibacterial and Anticancer Activity Using Skin Transcriptome of Treefrog (Boana Pugnax). Pharmaceutics 2021, 13, 578. [Google Scholar] [CrossRef]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network Protein Sequence Analysis. Trends in Biochemical Sciences 2000, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Cámara-Ruiz, M.; Cerezo, I.M.; Guardiola, F.A.; García-Beltrán, J.M.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á. Alteration of the Immune Response and the Microbiota of the Skin during a Natural Infection by Vibrio harveyi in European Seabass (Dicentrarchus Labrax). Microorganisms 2021, 9, 964. [Google Scholar] [CrossRef]
- Cheng, A.-C.; Lin, H.-L.; Shiu, Y.-L.; Tyan, Y.-C.; Liu, C.-H. Isolation and Characterization of Antimicrobial Peptides Derived from Bacillus Subtilis E20-Fermented Soybean Meal and Its Use for Preventing Vibrio Infection in Shrimp Aquaculture. Fish & Shellfish Immunology 2017, 67, 270–279. [Google Scholar] [CrossRef]
- Han, Y.; Chen, W.; Sun, Z. Antimicrobial Activity and Mechanism of Limonene against Staphylococcus Aureus. Journal of Food Safety 2021, 41, e12918. [Google Scholar] [CrossRef]
- Lv, J.; Da, R.; Cheng, Y.; Tuo, X.; Wei, J.; Jiang, K.; Monisayo, A.O.; Han, B. Mechanism of Antibacterial Activity of Bacillus Amyloliquefaciens C-1 Lipopeptide toward Anaerobic Clostridium Difficile. BioMed Research International 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.Á.; Muñoz, J.; Meseguer, J. Blood Cells of Sea Bass (Dicentrarchus Labrax l.). Flow Cytometric and Microscopic Studies. Anat. Rec. 2000, 258, 80–89. [Google Scholar] [CrossRef]
- Wu, G.; Ding, J.; Li, H.; Li, L.; Zhao, R.; Fan, X.; Shen, Z. Effects of Cations and PH on Antimicrobial Activity of Thanatin and S-Thanatin against Escherichia coli ATCC25922 and B. Subtilis ATCC 21332. Nat Prec 2008. [Google Scholar] [CrossRef]
- Wang, H.; He, H.; Chen, X.; Zhou, M.; Wei, M.; Xi, X.; Ma, C.; Du, Q.; Chen, T.; Shaw, C.; et al. A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina Senegalensis. Biology 2020, 9, 148. [Google Scholar] [CrossRef]
- Zeng, P.; Cheng, Q.; Xu, J.; Xu, Q.; Xu, Y.; Gao, W.; Wong, K.-Y.; Chan, K.-F.; Chen, S.; Yi, L. Membrane-Disruptive Engineered Peptide Amphiphiles Restrain the Proliferation of Penicillins and Cephalosporins Resistant Vibrio alginolyticus and Vibrio parahaemolyticus in Instant Jellyfish. Food Control 2022, 135, 108827. [Google Scholar] [CrossRef]
- Ji, Q.; Wang, S.; Ma, J.; Liu, Q. A Review: Progress in the Development of Fish Vibrio Spp. Vaccines. Immunology Letters 2020, 226, 46–54. [Google Scholar] [CrossRef]
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J Aqua Anim Hlth 2019, 31, 3–22. [Google Scholar] [CrossRef]
- Li, J.; Zhou, L.; Woo, N.Y.S. Invasion Route and Pathogenic Mechanisms of Vibrio Alginolyticus to Silver Sea Bream Sparus Sarba. J Aqua Anim Hlth 2003, 15, 302–313. [Google Scholar] [CrossRef]
- Destoumieux-Garzón, D.; Duperthuy, M.; Vanhove, A.; Schmitt, P.; Wai, S. Resistance to Antimicrobial Peptides in Vibrios. Antibiotics 2014, 3, 540–563. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.; Amal, M.N.A.; Yasin, I.S.M.; Zamri Saad, M.; Nasruddin, N.S.; Al-saari, N.; Mino, S.; Sawabe, T. Vibriosis in Cultured Marine Fishes: A Review. Aquaculture 2019, 512, 734289. [Google Scholar] [CrossRef]
- Han, H.; Li, T.; Wang, Z.; Teng, D.; Mao, R.; Hao, Y.; Yang, N.; Wang, X.; Wang, J. Improved Stability and Activity of a Marine Peptide-N6NH2 against Edwardsiella Tarda and Its Preliminary Application in Fish. Marine Drugs 2020, 18, 650. [Google Scholar] [CrossRef]
- Nicolas, P.; El Amri, C. The Dermaseptin Superfamily: A Gene-Based Combinatorial Library of Antimicrobial Peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A. How Do Bacteria Resist Human Antimicrobial Peptides? Trends in Microbiology 2002, 10, 179–186. [Google Scholar] [CrossRef]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef]
- Bartels, E.J.H.; Dekker, D.; Amiche, M. Dermaseptins, Multifunctional Antimicrobial Peptides: A Review of Their Pharmacology, Effectivity, Mechanism of Action, and Possible Future Directions. Front. Pharmacol. 2019, 10, 1421. [Google Scholar] [CrossRef]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Nanomedicine-Based Antimicrobial Peptide Delivery for Bacterial Infections: Recent Advances and Future Prospects. J. Pharm. Investig. 2021, 51, 377–398. [Google Scholar] [CrossRef]
- Kang, S.-J.; Kim, D.-H.; Mishig-Ochir, T.; Lee, B.-J. Antimicrobial Peptides: Their Physicochemical Properties and Therapeutic Application. Arch. Pharm. Res. 2012, 35, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Oren, Z.; Shai, Y. A Class of Highly Potent Antibacterial Peptides Derived from Pardaxin, A Pore-Forming Peptide Isolated from Moses Sole Fish Pardachirus Marmoratus. European Journal of Biochemistry 1996, 237, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Thermostability and Aliphatic Index of Globular Proteins. The Journal of Biochemistry 1980. [CrossRef]
- Jhong, J.-H.; Chi, Y.-H.; Li, W.-C.; Lin, T.-H.; Huang, K.-Y.; Lee, T.-Y. dbAMP: An Integrated Resource for Exploring Antimicrobial Peptides with Functional Activities and Physicochemical Properties on Transcriptome and Proteome Data. Nucleic Acids Research 2019, 47, D285–D297. [Google Scholar] [CrossRef] [PubMed]
- Guruprasad, K.; Reddy, B.V.B.; Pandit, M.W. Correlation between Stability of a Protein and Its Dipeptide Composition: A Novel Approach for Predicting in Vivo Stability of a Protein from Its Primary Sequence. Protein Eng Des Sel 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural Determinants of Host Defense Peptides for Antimicrobial Activity and Target Cell Selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ríos, D.; Ramírez-Malule, H. Bibliometric Analysis of Recent Research on Multidrug and Antibiotics Resistance (2017–2018). Journal of Applied Pharmaceutical Science 2019, 9, 112–116. [Google Scholar]
- McMillan, K.A.M.; Coombs, M.R.P. Investigating Potential Applications of the Fish Anti-Microbial Peptide Pleurocidin: A Systematic Review. Pharmaceuticals 2021, 14, 687. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, Y.-F.; Chen, J.-H.; Chen, X.; Lin, Z.-H. Molecular Characterization of Cathelicidin in Tiger Frog (Hoplobatrachus Rugulosus): Antimicrobial Activity and Immunomodulatory Activity. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2021, 247, 109072. [Google Scholar] [CrossRef]
- Sánchez-Acosta, Y.A.; Castillo Vargas, J.A.; Ramírez Quintero, K.J.; Orduz Peralta, S.; Camargo Rodríguez, D.O. Peptide Derivatives of Dermaseptin S4 in Fresh Bovine Semen for Bacterial Contamination Control: Physicochemical and Structural Characterization, Antibacterial Potency, and Effects on Red Blood and Sperm Cells. Reprod Domestic Animals 2020, 55, 905–914. [Google Scholar] [CrossRef]
- Tan, Y.; Chen, X.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.; Kwok, H.; Chen, T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa Sauvagii. Toxins 2018, 10, 320. [Google Scholar] [CrossRef]
- Lee, T.-H.; Hofferek, V.; Separovic, F.; Reid, G.E.; Aguilar, M.-I. The Role of Bacterial Lipid Diversity and Membrane Properties in Modulating Antimicrobial Peptide Activity and Drug Resistance. Current Opinion in Chemical Biology 2019, 52, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Makovitzki, A.; Avrahami, D.; Shai, Y. Ultrashort Antibacterial and Antifungal Lipopeptides. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 15997–16002. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Barra, D.; Simmaco, M.; Bozzi, A.; Di Giulio, A.; Rinaldi, A.C. Effects of the Antimicrobial Peptide Temporin L on Cell Morphology, Membrane Permeability and Viability of Escherichia coli. Biochemical Journal 2004, 380, 859–865. [Google Scholar] [CrossRef]
- Tenover, F.C. Mechanisms of Antimicrobial Resistance in Bacteria. The American Journal of Medicine 2006, 119, S3–S10. [Google Scholar] [CrossRef]
- Vineethkumar, T.V.; Asha, R.; Shyla, G.; George, S. Studies on the Mode of Membrane Interaction of C-Terminally Amidated Brevinin1 HYba1 and 2 Peptides Against Bacteria. Int J Pept Res Ther 2018, 24, 117–129. [Google Scholar] [CrossRef]
- Sato, H.; Feix, J.B. Peptide–Membrane Interactions and Mechanisms of Membrane Destruction by Amphipathic α-Helical Antimicrobial Peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes 2006, 1758, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Zhou, L.; Yang, J.; Zhuang, L.; Tang, J.; Liu, T.; Wu, J.; Yang, H. The First Identified Cathelicidin from Tree Frogs Possesses Anti-Inflammatory and Partial LPS Neutralization Activities. Amino Acids 2017, 49, 1571–1585. [Google Scholar] [CrossRef]
- Wei, L.; Yang, J.; He, X.; Mo, G.; Hong, J.; Yan, X.; Lin, D.; Lai, R. Structure and Function of a Potent Lipopolysaccharide-Binding Antimicrobial and Anti-Inflammatory Peptide. J. Med. Chem. 2013, 56, 3546–3556. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of Lipids in the Interaction of Antimicrobial Peptides with Membranes. Progress in Lipid Research 2012, 51, 149–177. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing Antimicrobial Peptides: Form Follows Function. Nat Rev Drug Discov 2012, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Mukhopadhyay, R.; Wen, B.; Gitai, Z.; Wingreen, N.S. Cell Shape and Cell-Wall Organization in Gram-Negative Bacteria. Proceedings of the National Academy of Sciences 2008, 105, 19282–19287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ouyang, J.; Fu, L.; Xu, C.; Ge, Y.; Sun, S.; Li, X.; Lai, S.; Ke, H.; Yuan, B.; et al. Hydrophobicity Determines the Bacterial Killing Rate of α-Helical Antimicrobial Peptides and Influences the Bacterial Resistance Development. J. Med. Chem. 2022, 65, 14701–14720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dhillon, P.; Yan, H.; Farmer, S.; Hancock, R.E.W. Interactions of Bacterial Cationic Peptide Antibiotics with Outer and Cytoplasmic Membranes of Pseudomonas Aeruginosa. Antimicrob Agents Chemother 2000, 44, 3317–3321. [Google Scholar] [CrossRef]
- Tomita, T.; Hitomi, S.; Nagase, T.; Matsui, H.; Matsuse, T.; Kimura, S.; Ouchi, Y. Effect of Ions on Antibacterial Activity of Human Beta Defensin 2. Microbiology and Immunology 2000, 44, 749–754. [Google Scholar] [CrossRef] [PubMed]
- León, R.; Ruiz, M.; Valero, Y.; Cárdenas, C.; Guzman, F.; Vila, M.; Cuesta, A. Exploring Small Cationic Peptides of Different Origin as Potential Antimicrobial Agents in Aquaculture. Fish & Shellfish Immunology 2020, 98, 720–727. [Google Scholar] [CrossRef]
- Kelly, D.G.; Wolters, W.R.; Jaynes, J.M.; Newton, J.C. Enhanced Disease Resistance to Enteric Speticemia in Channel Catfish, Ictalurus Punctatus, Administered Lytic Peptide. Journal of Applied Aquaculture 1994, 3, 25–34. [Google Scholar] [CrossRef]
Figure 1.
Secondary structure of the dermaseptin S4 antimicrobial peptides predicted by the PEP-FOLD server, N-t and C-t are shown in blue and red respectively (a). Diagrams of helical wheel projection of DS4 (1-28) (b) and DS4(1-26) a (c) peptides. The blue and red circles represent the positively and the negatively charged amino acids, respectively. Hydrophilic residues are colored purple, hydrophobic residues are colored yellow, small residues are colored gray and amides are colored pink. Hydrophobic moment is indicated by arrows.
Figure 1.
Secondary structure of the dermaseptin S4 antimicrobial peptides predicted by the PEP-FOLD server, N-t and C-t are shown in blue and red respectively (a). Diagrams of helical wheel projection of DS4 (1-28) (b) and DS4(1-26) a (c) peptides. The blue and red circles represent the positively and the negatively charged amino acids, respectively. Hydrophilic residues are colored purple, hydrophobic residues are colored yellow, small residues are colored gray and amides are colored pink. Hydrophobic moment is indicated by arrows.
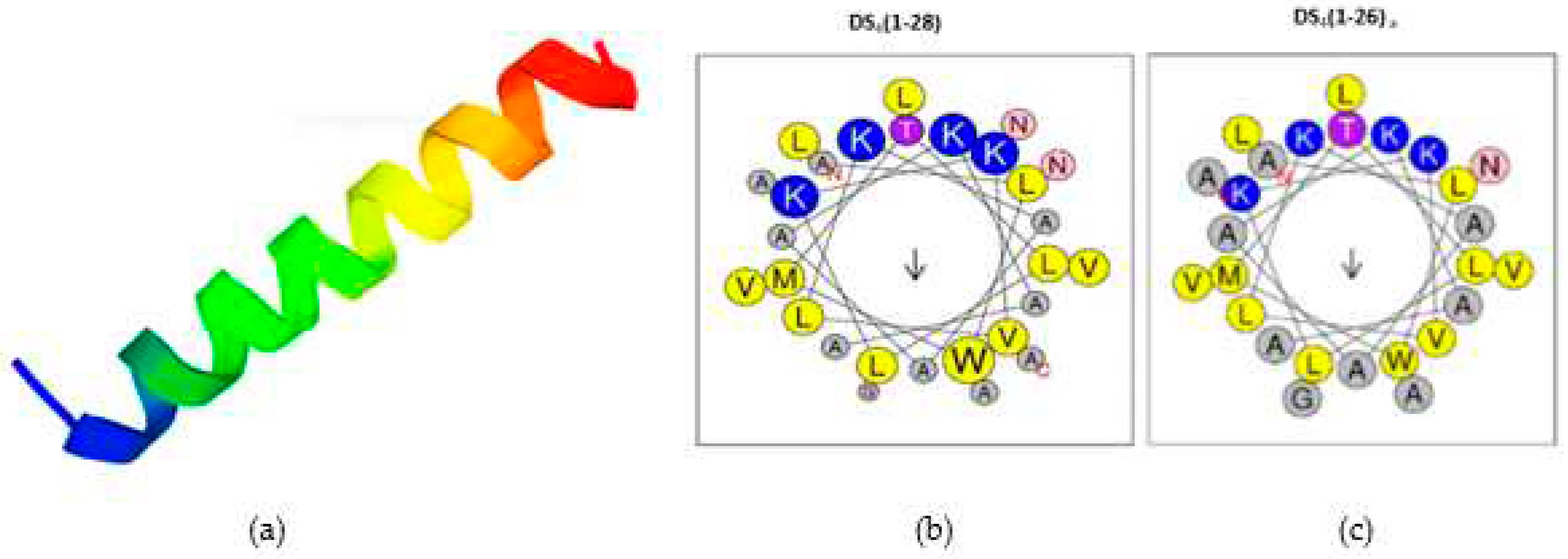
Figure 2.
Antibacterial activity determined by the percentage (%) of inhibition of the three dermaseptin S4 peptides tested against Vibrio harveyi and V. anguillarum. Data represent the mean ± standard error of the mean (SEM) (n = 3). P*<0.1; P**<0.01 and P***<0.001.
Figure 2.
Antibacterial activity determined by the percentage (%) of inhibition of the three dermaseptin S4 peptides tested against Vibrio harveyi and V. anguillarum. Data represent the mean ± standard error of the mean (SEM) (n = 3). P*<0.1; P**<0.01 and P***<0.001.
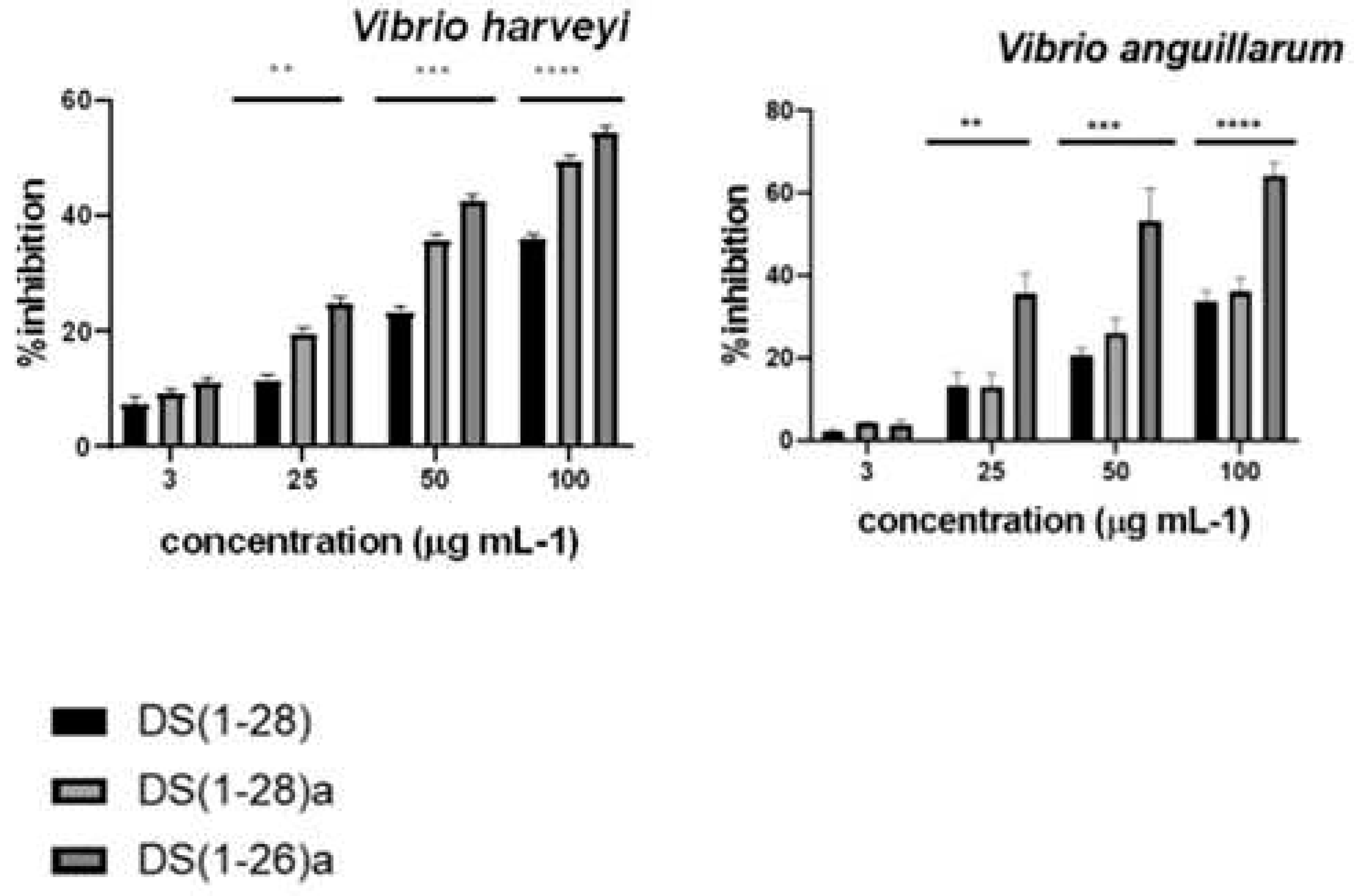
Figure 3.
Release of intracellular nucleic acids (determined by optical density, O.D.) from V. harveyi after being incubated with the three dermaseptin S4 peptides. Values shown as the mean ± SEM, (n = 3). P-values were calculated by two-way ANOVA; *** P < 0.001 vs. control group.
Figure 3.
Release of intracellular nucleic acids (determined by optical density, O.D.) from V. harveyi after being incubated with the three dermaseptin S4 peptides. Values shown as the mean ± SEM, (n = 3). P-values were calculated by two-way ANOVA; *** P < 0.001 vs. control group.
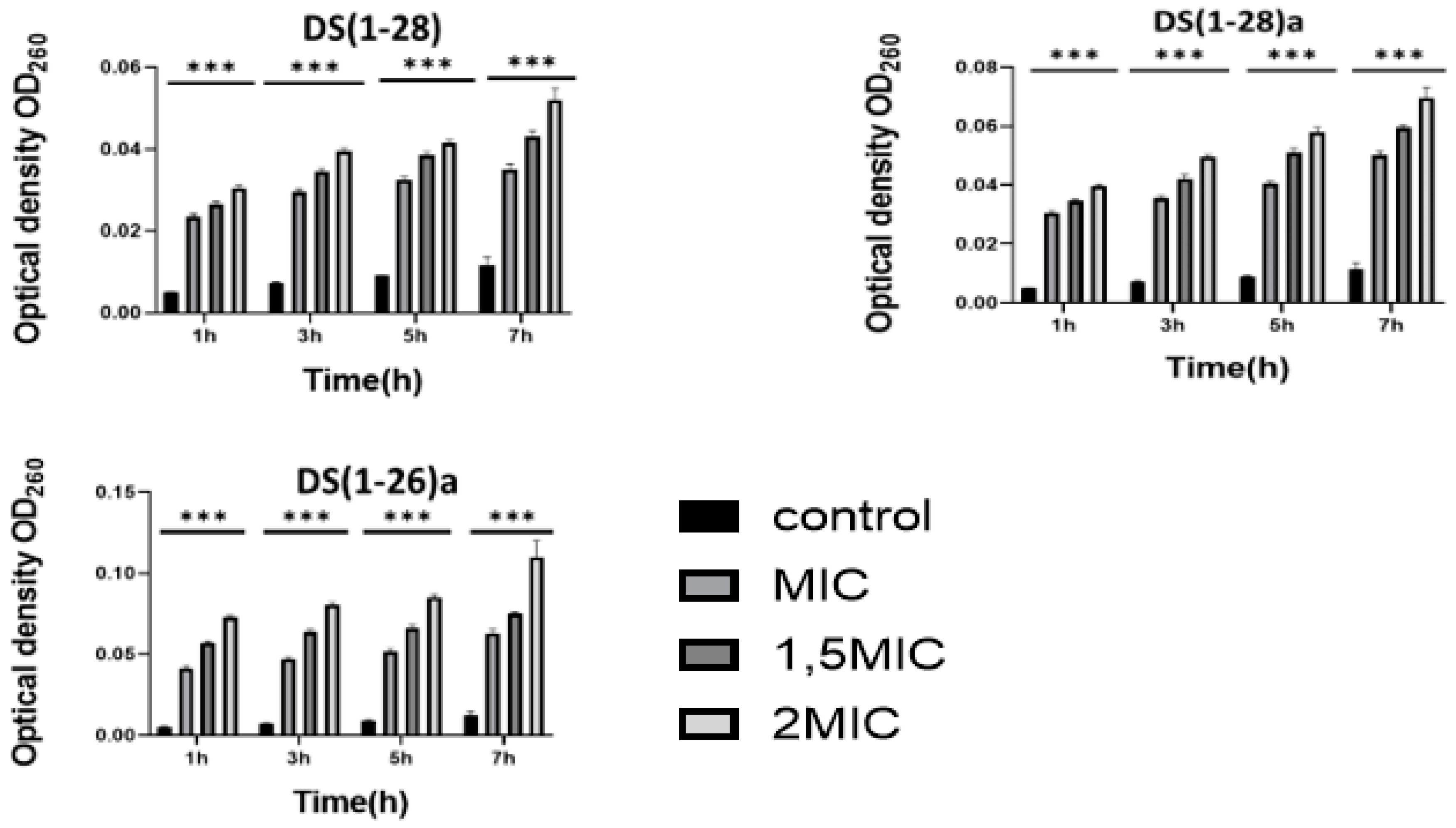
Figure 4.
Release of intracellular nucleic acids (determined by optical density, O.D.) from V. anguillarum after being incubated with the three dermaseptin S4 peptides. Values shown as the mean ± SEM, (n = 3). P-values were calculated by two-way ANOVA; *** P < 0.001 vs. control group.
Figure 4.
Release of intracellular nucleic acids (determined by optical density, O.D.) from V. anguillarum after being incubated with the three dermaseptin S4 peptides. Values shown as the mean ± SEM, (n = 3). P-values were calculated by two-way ANOVA; *** P < 0.001 vs. control group.
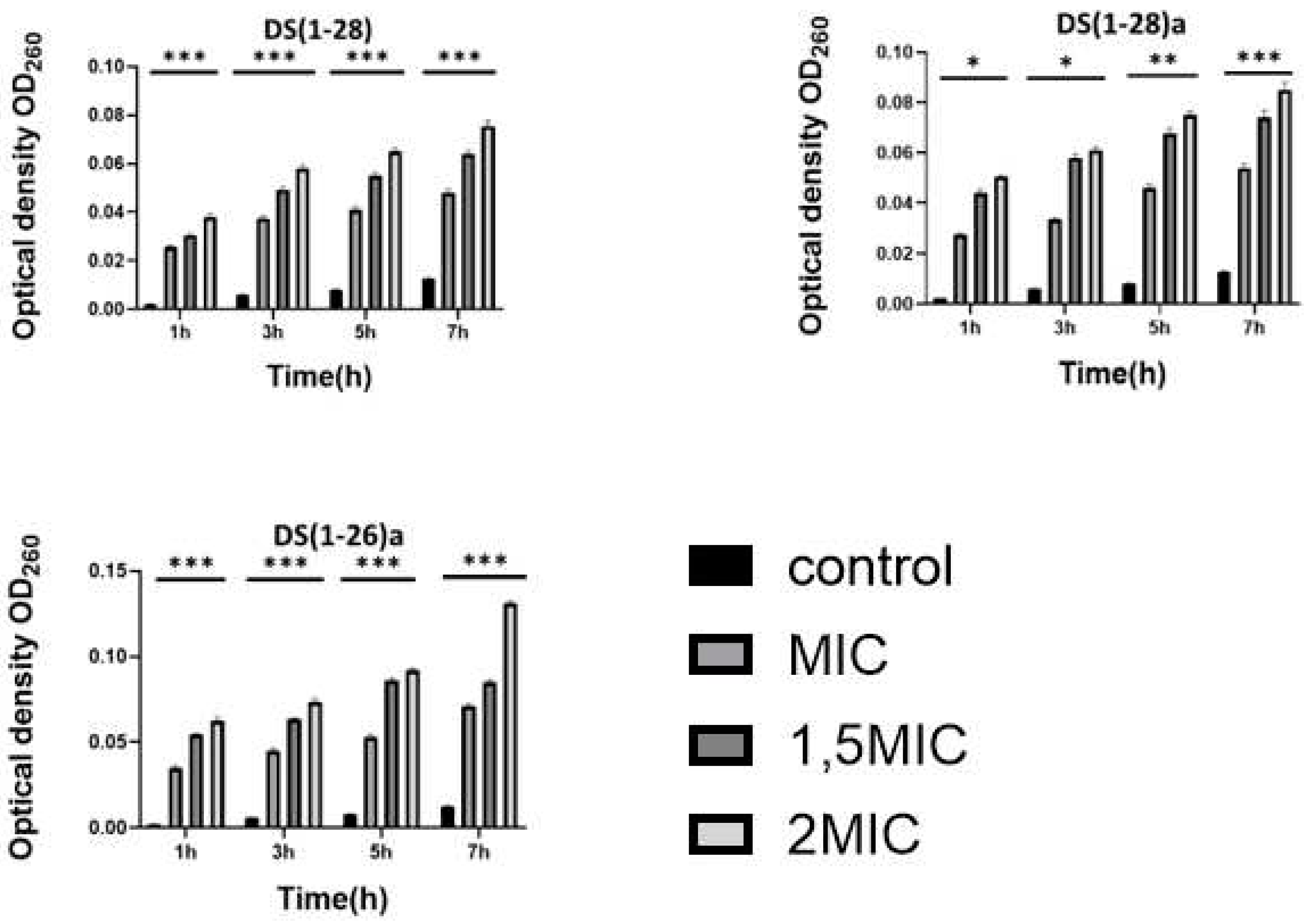
Figure 5.
Extracellular AKPase activity (units mL-1) for V. harveyi (a, b, c) and V. anguillarum (d, e, f) after being incubated with the three dermaseptin S4 peptides. ***p < 0.001 indicated statistically significant differences of AMP treatment vs. negative control. Data represent the mean ± standard error of the mean SEM (n=3).
Figure 5.
Extracellular AKPase activity (units mL-1) for V. harveyi (a, b, c) and V. anguillarum (d, e, f) after being incubated with the three dermaseptin S4 peptides. ***p < 0.001 indicated statistically significant differences of AMP treatment vs. negative control. Data represent the mean ± standard error of the mean SEM (n=3).
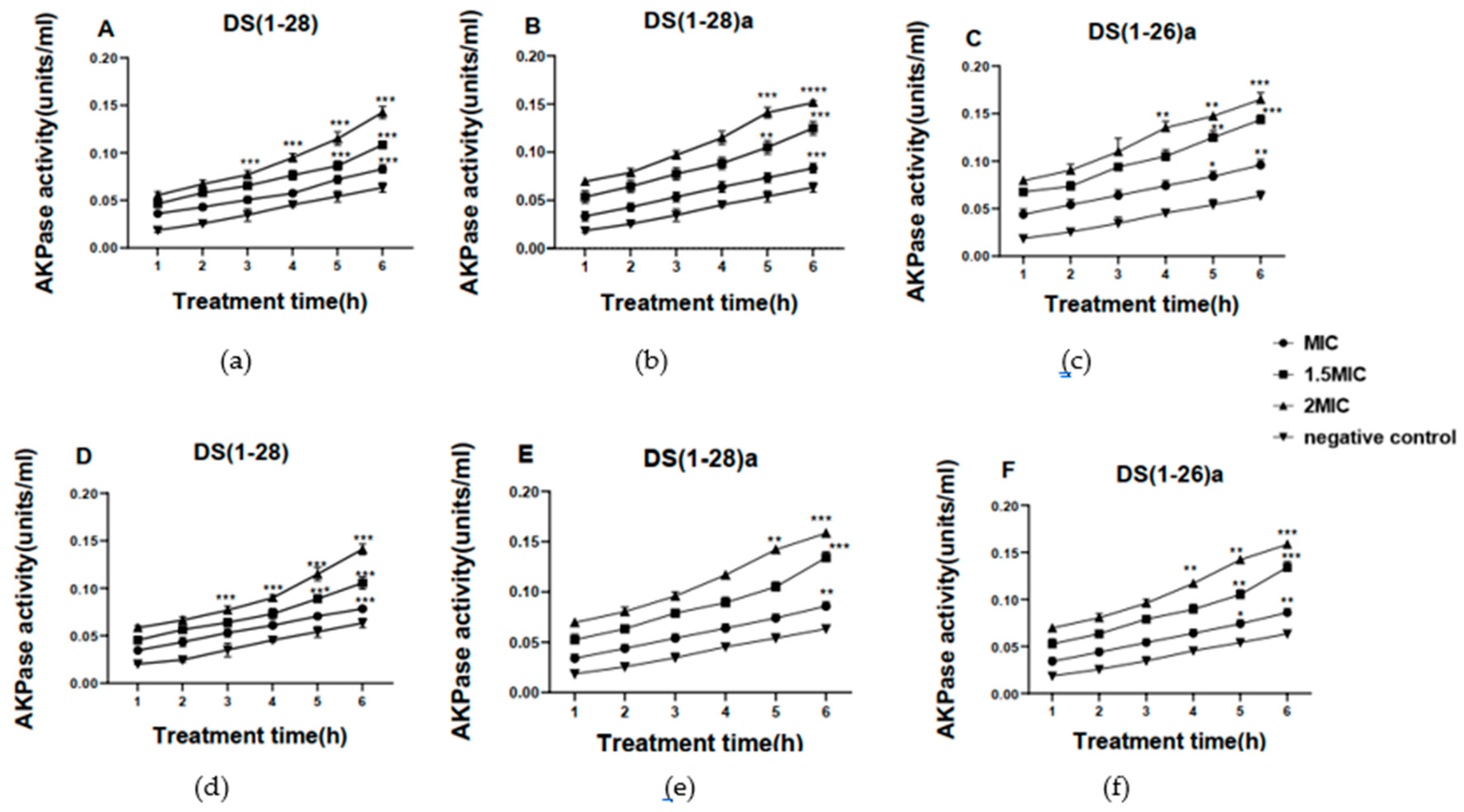
Figure 6.
Representative transmission electron micrographs of V. anguillarum (A, B) and V. harveyi (C, D), after incubation for 120 min without (0 µg mL-1, control) AMPs. Bar = 2 μm.
Figure 6.
Representative transmission electron micrographs of V. anguillarum (A, B) and V. harveyi (C, D), after incubation for 120 min without (0 µg mL-1, control) AMPs. Bar = 2 μm.

Figure 7.
Representative transmission electron micrographs of V. anguillarum (a, b, c) after incubation for 120 min with dermaseptin S4 peptides, DS4(1-28), DS4(1-28)a and DS4(1-26)a, respectively, at 1*MIC. Bar = 2µm.
Figure 7.
Representative transmission electron micrographs of V. anguillarum (a, b, c) after incubation for 120 min with dermaseptin S4 peptides, DS4(1-28), DS4(1-28)a and DS4(1-26)a, respectively, at 1*MIC. Bar = 2µm.

Figure 8.
Representative transmission electron micrographs of V. harveyi (a, b, c) respectively after incubation for 120 min with dermaseptin S4 peptides, DS4(1-28), DS4(1-28)a, and DS4(1-26)a at 1 MIC. Bar = 2µm.
Figure 8.
Representative transmission electron micrographs of V. harveyi (a, b, c) respectively after incubation for 120 min with dermaseptin S4 peptides, DS4(1-28), DS4(1-28)a, and DS4(1-26)a at 1 MIC. Bar = 2µm.

Figure 9.
Effects of divalent cations Mg2+, Ca2+ (0, control and 20 µg mL-1) on the antimicrobial activity of the three dermaseptin S4 peptides tested against Vibrio harveyi (a, b) and V. anguillarum (c, d).
Figure 9.
Effects of divalent cations Mg2+, Ca2+ (0, control and 20 µg mL-1) on the antimicrobial activity of the three dermaseptin S4 peptides tested against Vibrio harveyi (a, b) and V. anguillarum (c, d).
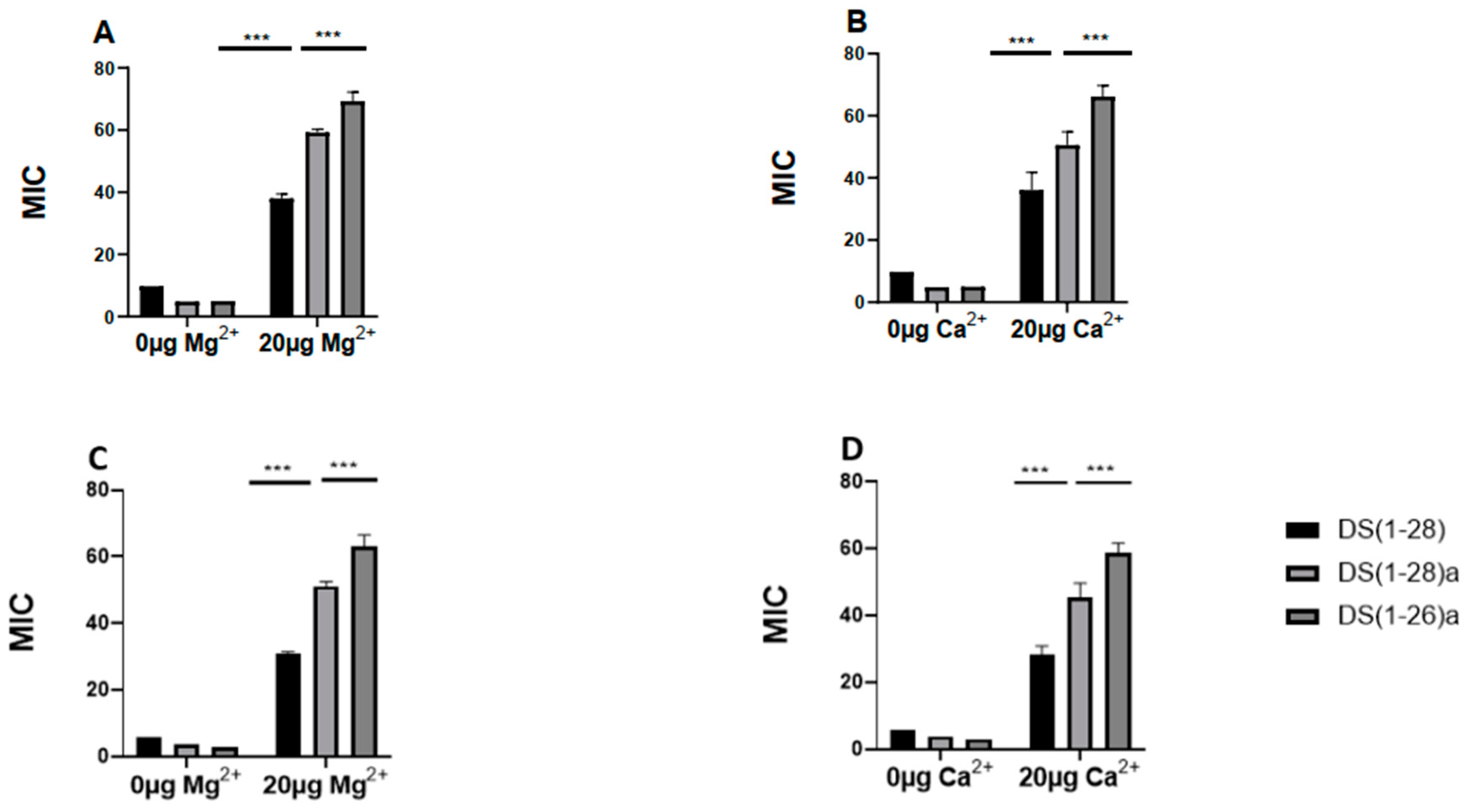
Figure 10.
Hemolytic activity of the three dermaseptin S4 peptides tested against red of pig (a) or gilthead seabream (b) blood cells. Data represent the mean ± SEM (n=3). P*<0.1; P**<0.01.
Figure 10.
Hemolytic activity of the three dermaseptin S4 peptides tested against red of pig (a) or gilthead seabream (b) blood cells. Data represent the mean ± SEM (n=3). P*<0.1; P**<0.01.
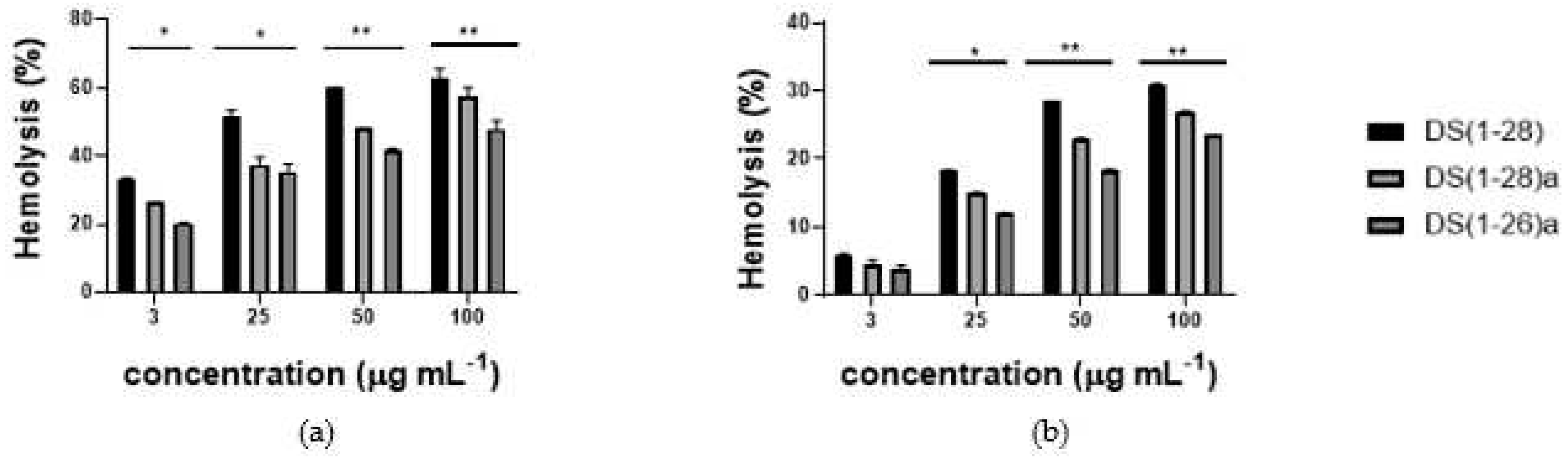

Table 1.
Sequence and main characteristics of the native antimicrobial peptide DS4 and its derivatives used in this study. MW: Molecular Weight; PI: Isoelectric Point; GRAVY: Grand Average of Hydropathicity; II: Instability Index; HR: Hydrophobic Ratio; AI: Aliphatic Index.
Table 1.
Sequence and main characteristics of the native antimicrobial peptide DS4 and its derivatives used in this study. MW: Molecular Weight; PI: Isoelectric Point; GRAVY: Grand Average of Hydropathicity; II: Instability Index; HR: Hydrophobic Ratio; AI: Aliphatic Index.
| Peptide | sequence | MW | Charge | PI | GRAVY | II | HR | AI | Structure | |
|---|---|---|---|---|---|---|---|---|---|---|
| α-helix | β-turn | |||||||||
| DS4(1-28) | NH2-A-L-W-M-T-L-L-K-K-V-L-K-A-A-A-K-A-A-L-N-A-V-L-V-G-A-N-A-COOH | 2850.55 | +4 | 10.48 | 1.032 | 10.37 | 72% | 146.79 | 78.57% | 0.00% |
| DS4(1-28)a | NH2-A-L-W-M-T-L-L-K-K-V-L-K-A-A-A-K-A-A-L-N-A-V-L-V-G-A-N-A-CONH2 | 2850.55 | +5 | 10.48 | 1.032 | 10.37 | 72% | 146.79 | 78.57% | 0.00% |
| DS4(1-26)a | NH2-A-L-W-M-T-L-L-K-K-V-L-K-A-A-A-K-A-A-L-N-A-V-L-V-G-A-CONH2 | 2665.36 |
+5 | 10.04 | 1.17 | 10.40 | 73% | 154.23 | 84.62% | 0.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



