
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
This work reports the isolation, purification, and anti-inflammatory evaluation of compounds from the plant species Ageratina pichinchesis. Using open-column chromatography, 11 known compounds were purified, whose chemical structures were elucidated by nuclear magnetic resonance techniques (1D and 2D). All compounds were evaluated in the in vitro model of RAW 264.7 mouse macrophage cells measuring nitric oxide inhibition as a determination of the anti-inflammatory effect. Outstandingly, the compound betuletol 3-O-β-glucoside (11) exhibited inhibition of nitric oxide with an IC50 of 75.08 ± 3.07% at 75 µM, additionally, it inhibited the secretion of interleukin 6 (IL-6) and activation of nuclear factor (NF-kβ). These results show that compound 11 could be considered as a potential anti-inflammatory agent in suppressing the expression of NF-kβ target genes such as the proinflammatory pathway and inducible nitric oxide synthase (iNOS). Consequently, the medicinal use attributed in traditional medicine to the A. pichinchensis species is due to the diversity of compounds that the plant biosynthesizes, demonstrating that its effect relates to anti-inflammatory effects associated with compound 11.
Keywords:
betuletol 3-O-β-glucoside
; A. pichinchensis
; anti-inflammatory activity
1. Introduction
The plant species A. pichinchensis (Kunth) R.M. King & Ho. Steal. (Asteraceae), is native to Mexico and grows in different states [1,2,3,4]. Particularly, in the state of Morelos it is popularly known, as “axihuitl” which is mainly used for treating stomach pain, and respiratory, gastrointestinal, and skin infections [5,6,7].
Scientific studies carried out in different biological evaluation models reveal that the aerial parts of A. pichinchensis exhibit activity against onychomycosis, tinea pedis, gastroprotective and healing effects, and even inhibit the in vitro proliferation of keratinocytes [8,9,10,11,12,13]. The main compounds associated with this type of biological effect are chromenes, furans, terpenes, essential oils, and glycosylated flavonoids [10,11,12,13,14,15]. However, the anti-inflammatory effect of the plant species has not been reported.
Inflammation is a response of the immune system whose stimulation can be caused by infections or stress and is associated with multiple diseases and triggers numerous biochemical, immunological, and cellular reactions [16,17,18,19]. During the inflammatory process, mediating substances are released in the nervous system that generate pain, among which are interleukins that induce genes that encode enzymes and other chemical species that contribute to the inflammatory response [20,21,22,23].
In particular, the enzyme nitric oxide synthase (iNOS) catalyzed the formation of the free radical nitric oxide (NO) in a gaseous state that stimulates vasodilation and cannot be stored [24,25,26]. It should be noted that the participation of NO plays a defensive role, acting as an immunoregulatory and antimicrobial agent due to its ability to generate DNA damage, inhibit energy metabolism enzymes, oxide proteins, and peroxidize membrane lipids of pathogenic agents, however, in high concentrations of NO exert the same effects on the cells of inflamed organs, exacerbating the damage and consequently the inflammatory process [27,28,29,30].
The iNOS enzyme can be elicited in different cell types, such as macrophages, hepatocytes, neutrophils, etc. [31,32,33], which generates a large amount of NO that can be toxic, which is why NO functions as a pro-inflammatory molecule [34,35,36]. In a chronic phase of inflammation, the inflammatory signal can be magnified by the production of molecules such as nuclear transcription factor kappa β (NF-kβ), TNF-α, and interferon-gamma (IFN-γ) which promote transcription of iNOS, generating large amounts of NO, greater vasodilation, edema, and plasma exudation that can even damage the DNA double helix [37,38,39,40]. High production of NO can be associated with the development of diseases such as Alzheimer’s, cardiovascular, rheumatoid arthritis, pulmonary fibrosis, diabetes, and cancer, among others [41,42,43,44,45,46]. This is why the iNOS enzyme is essential in the inflammatory process and allows us to understand the mechanism of action of active ingredients from medicinal plants [47,48,49].
Particularly, this study reports the anti-inflammatory activity of chemical compounds isolated from aerial parts (leaves and flowers) of A. pichinchensis on the inhibition of NO in RAW 264.7 mouse macrophage cells induced with lipopolysaccharides (LPS).
2. Materials and Methods
Compounds 1–11 were characterized using spectroscopic techniques and mass spectrometry; for compounds 1, 3, 4, 6, 7, and 8, a Varian Unity Inova 200 MHz equipment was used; Compounds 2, 5, 9, and 10 were analyzed on a Varian Mercury Plus 400 MHz—ID3 spectrometer, for these compounds CDCl3 was used, compound 11 was dissolved in DMSO-d6, the spectra of 1H, 13C, DEPT, COSY, HSQC, and HMBC were obtained on a Bruker AVANCE III HD 500 MHz Spectrometer (Billerica, MA, USA). FABMS spectra were obtained using a JEOL-AX 505HA mass spectrometer. Optical rotation was measured in CHCl3 on a Perkin Elmer 241 digital polarimeter at 25 °C. Melting points were determined on a Prendo apparatus [50,51,52]. Compounds 1-11 were purified using column chromatography (CC), silica gel 60 (70-230 and 230-400, mesh) as stationary phase, and thin layer chromatography (silica gel 60 F254, Merck) to monitor the separation of the compounds, which were visualized using a solution of Ce (SO4)2 (NH4)2SO4.2H2O.
2.1. Plant material
Complete plant A. pichinchensis was collected by M.B. Mariana Sánchez Ramos in the town of San Juan Tlacotenco, municipality of Tepoztlán, Morelos, Mexico, in April 2018 (19°00′43.88″ N, 99.05′38.66″ W). The plant was prepared (pressed) and taken to the HUMO Herbarium of the Autonomous University of the State of Morelos (UAEM). The Biol. Gabriel Flores Franco identified this species, registered with voucher number 33913 [53].
2.2. Extraction and isolation
Leaves were selected from the collected plants and dried at room temperature. This material (1.085 kg) was successfully extracted with AcOEt three times (each time with 4 L of the solvent).
The AcOEt extracts obtained were concentrated to dryness by distillation under reduced pressure using a rotary evaporator, obtaining 20.3 g of residue. Fractionation of the EtOAc extract by open CC (silica gel, 70–230 mesh; 10 cm i.d. × 60 cm) was performed with a gradient system of n-hexane-EtOAc 80:20 to 0:100, collecting 35 fractions of 300 mL each.
Based on TLC analysis, these fractions were grouped according to their chemical profile into four groups; AP-1A (fractions 1–5, n-hexane: EtOAc 80:20, 4.12 g), AP-1B (fractions 6–19, n-hexane: EtOAc 60:40 and 40:60, 5.23 g), AP-1C (fractions 20–32, n-hexane: EtOAc 20:80, 5.94 g) and AP-1D (fractions 33–35, EtOAc 100%, 3.52 g). AP-1A contains aliphatic esters, fatty acids, and O-methylenecalinol (1, 48 mg) as the main product, as well as AP-1D of sugars. The groups AP-1B and AP-1C were subjected to column chromatography using silica gel (70–230 mesh). The AP-1B fraction (5.23 g) was adsorbed on 4.9 g of silica gel and placed in a glass column (70 cm high and 3.5 cm in diameter) packed with 157 g of silica gel. Elution was carried out using a gradient system, n-hexane:Ethyl acetate (100:00→50:50) and 164 fractions were obtained in 50 mL. These were concentrated and monitored by TLC, grouped in 6 groups of fractions. The AP-1B-1 group (fractions 1–39, 0.306 g, n-hexane: ethyl acetate 95:05 to 85:15) was purified and identified as O-methylencecalinol (1, 32 mg) as the main product; from AP-1B-2 (fractions 40–69, 3.32 g, n-hexane:ethyl acetate 80:20 to 70:30) they were identified 7-hydroxiencecalin (3, 7 mg) and 8-hydroxiencalin (4, 6 mg); the compounds encecalin (2, 16 mg), 3,5-diprenyl-4-hydroxyacetophenone (5, 32 mg) and (+)-β-eudesmol (6, 8 mg) were isolated from the AP-1B-3 group (fractions 70–99, 0.391 g, n-hexane: ethyl acetate 65:35 to 55:45); The AP-1B-4 group (fractions 100–129, 0.821 g, n-hexane: ethyl acetate 50:50 to 40:60) led to the identification of the compounds (+)-β-eudesmol (6, 7 mg), dehydrospeletone (8, 12 mg) and speletone (7, 14 mg); through successive chromatography of the AP-1B-5 group ( fractions 130–149, 0.427 g, n-hexane: ethyl acetate 35:75 to 25:75) were isolated encecalinol (9, 28 mg) and 5-acetyl-3β-angeloxy-2β-(1-hydroxyisopropyl)-2,3-dihydrobenzofuran (10, 36 mg).
O-methylencecalinol (1)
Colorless oil, 1H-NMR (200 MHz, CDCl3), δH: 6.95 (1H, s, H-5), 6.32 (1H, s, H-8), 6.26 (1H, d, J = 9.6 Hz, H-4), 5.43 (1H, d, J = 10 Hz, H-3), 4.61 (1H, q, J= 12.8, 6.8 Hz, H-11), 3.75 (3H, s,-OMe), 3.21 (3H, , OMe), 1.40 (3H, s, CH3-13), 1.39 (3H, s, CH3-14), 1.34 (3H, d, J=6.8 Hz, CH3-12). 13C-NMR (50 MHz, CDCl3). δC: 157.70 (C-7), 153.25 (C-10), 127.76 (C-3), 124.06 (C-5), 122.30 (C-4), 120.02 (C-6), 114.19 (C-9), 99.43 (C-8), 76.55 (C-2), 72.97 (C-11), 56.59 (-OMe-C11), 56.03 (OMe-C-7), 28.32 (C-13), 28.26 (C-14), 22.65 (C-12), these data match those in the literature [12,54]. Spectra of 1H and 13C-NMR are in Figures S1 and S2.
Encecalin (2)
Yellow oil; 1H-NMR (400 MHz, CDCl3), δH: 7.23 (1H, s, H-5), 6.27 (1H, s, H-8), 6.12 (1H, d, J = 9.8 Hz, H-4), 5.44 (1H, d, J = 9.7 Hz, H-3), 3.55 (3H, s,-OMe), 2.10 (3H, s, CH3-12), 1.20 (6H, s, CH3-13 and CH3-14). 13C-NMR (100 MHz, CDCl3). δC: 197.68 (C-11), 161.37 (C-7), 158.28 (C-10), 128.94 (C-5), 128.36 (C-3), 121.36 (C-4), 120.42 (C-6), 114.22 (C-9), 99.76 (C-8), 77.21 (C-2), 55.72 (MeO-), 32.26 (C-12), 26.68 (C-13), 28.32 (C-14), these data match those in the literature [55,56]. Spectra of 1H and 13C-NMR are in Figures S3 and S4.
Euparoriochromene (3)
Yellow needles; mp: 78–80 ◦C; 1H-NMR (200 MHz, CDCl3), δH: 7.21 (1H, s, H-5), 6.24 (1H, s, H-8), 6.21 (1H, d, J = 10 Hz, H-4), 5.41 (1H, d, J = 10 Hz, H-3), 2.49 (3H, s, CH3-12), 1.39 (6H, s, CH3-13 and CH3-14). 13C-NMR (50 MHz, CDCl3). δC: 198.13 (C-11), 162.18 (C-7), 159.18 (C-10), 128.68 (C-5), 128.53 (C-3), 126.14 (C-6), 122.08 (C-4), 114.38 (C-9), 104.26 (C-8), 77.43 (C-2), 32.12(C-12), 28.86 (C-13) and 28.78 (C-14), these data match those in the literature [57]. Spectra of 1H and 13C-NMR are in Figures S5 and 62.
6-acethyl-8-hydroxy-2,2-dimethylchromene (4)
White powder; mp: 98 °C; 1H-NMR (200 MHz, CDCl3), δH: 7.62 (1H, d, J= 1.2 Hz, H-7), 7.43 (1H, d, J= 1.2 Hz, H-5), 6.34 (1H, d, J = 10.2 Hz, H-4), 5.71 (1H, d, J = 10 Hz, H-3), 2.56 (3H, s, CH3-12) and 1.43 (6H, s, CH3-13 and CH3-14). 13C-NMR (50 MHz, CDCl3). δC: 202.98 (C-11), 165.23 (C-8), 160.68 (C-10), 128.61 (C-6), 128.32 (C-3), 125.34 (C-9), 122.24 (C-4), 118.38 (C-5), 114.31 (C-7), 78.26 (C-2), 28.58 (C-13 and C-14) and 26.38 (C-12), these data match those in the literature [58,59,60]. Spectra of 1H and 13C-NMR are in Figures S7 and S8.
3.5-diprenyl-4-hydroxyacetophenone (5)
Crystalline solid; mp: 93-95 °C; 1H-NMR (400 MHz, CDCl3), δH: 7.44 (2H, s, H-2 y H-6), 5.98 (1H, s, OH), 5.31(2H, m, H-2′ y H-2″), 3.37 (2H, d, J = 7.1 Hz, H-1′y H-1″), 2.48 (3H, d, J = 19.8 Hz, CH3-8), 1.74 (12H, d, J = 11.1 Hz, CH3-4′, CH3-4″y CH3-5′, CH3-5″). 13C-NMR (100 MHz, CDCl3), δC: 197.52 (C-7), 157.59 (C-4), 135.28 (C-1), 130.15 (C-31, C-3″), 129.02 (C-2, C-6), 127.30 (C-3, C-5), 121.58 (C-2′, C-2″), 29.82 (C-1′, C-1″), 26.51 (C-8), 25.99 (C-5″, C-4′) y 18.10 (C-4″, C-5′), these data match those in the literature [55,61]. Spectra of 1H and 13C-NMR are in Figures S9 and S10.
β-eudesmol (6)
White amorphous solid; mp = 78-79 °C; 1H-NMR (200 MHz, CDCl3), δH: 4.70 (1H, d, J = 2 Hz, H-15b), 4.43 (1H, d, J = 1.6 Hz, H-15a), 2.38 (2H, m, Hs-3), 1.98 (1H, m, H-10), 1.25-1.62 (6H, m, CH2-5, CH2-7 and CH2-8), 1.00–1.36 (5H, m, CH2-1, CH2-2 and CH-6), 1.39 (3H, s, CH3-11), 1.40 (3H, s, CH3-12), 0.69 (3H, s, CH3-14). 13C-NMR (50 MHz, CDCl3), δC: 152.41 (C-4), 105.14(C-15), 72.23 (C-11), 49.58 (C-10), 49.42 (C-6),42.12 (C-3), 41.86 (C-1), 41.12 (C-8), 36.89 (C-9), 26.76 (C-12 and C-13), 24.73 (C-5), 23.45 (C-2), 22.76 (C-7) and 16.24 (C-14), these data match those in the literature [55,62,63]. Spectra of 1H and 13C-NMR are in Figures S11 and S12.
Speletone (7)
Colorless oil; 1H-NMR (200 MHz, CDCl3), δH: 8.09 (1H, d, J = 2.4 Hz, H-2), 7.78 (1H, dd, J = 2.2, 7.8 Hz, H6), 6.84 (1H, d, J = 8.4 Hz, H-5), 3.60 (3H, s, OMe), 2.67 (2H, d, J = 7.8 Hz, H-10), 2.33 (3H, s, CH3-8), 221 (1H, m, H-11), 0.78 (6H, d, J = 6.8 Hz, CH3-12 and CH3-13), these data match those in the literature [55,64]. Spectra of 1H-NMR is in Figure S13.
Dehydrospeletone (8)
Colorless oil; 1H-NMR (200 MHz, CDCl3), δH: 8.16 (1H, d, J = 2.2 Hz, H-2), 8.06 (1H, dd, J = 2.2, 7.8 Hz, H6), 6.98 (1H, d, J = 8.2 Hz, H-5), 6.56 (1H, q, H-10), 3.92 (3H, s, OMe), 2.55 (3H, s, CH3-8), 2.22 (3H, s, CH3-12) and 1.95 (3H, s, CH3-13), these data match those in the literature [64,65]. Spectra of 1H-NMR is in Figure S14.
Encecalinol (9)
Yellow oil; :-77° (c 0.92, CHCl3); 1H-NMR (400 MHz, CDCl3), δH: 6.95 (s, H-5), 6.37 (s, H-8), 6.27 (d, J = 9.7 Hz, H-4), 5.47 (d, J = 9.8 Hz, H-3), 5.02 (q, J = 6.5 Hz, H-13), 3.81 (s, MeO-), 1.47 (d, J = 6.5 Hz, CH3-12), 1.42 (6H, s, CH3-13 and CH3-14). 13C-NMR (100 MHz, CDCl3) δC: 157.26 C-7), 153.18 (C-10), 127.64 (C-3), 125.76 (C-6),123.83 (C-5), 122.02 (C-4), 113.75 (C-9), 99.53 (C-8), 77.06 (C-2), 65.53 (C-11), 55.45 (MeO-7), 28.04 (C-13), 27.96 (C-14), 22.86 (CH3-12), these data match those in the literature [55,56,66]. Spectra of 1H and 13C-NMR are in Figures S15 and S16.
5-acetyl-3β--angeloyloxy-2β-(1-hydroxyisopropyl)-2,3-dihydrobenzofurane (10)
Yellow oil; = +47 (c= 0.8, CHCl3); 1H NMR (400 MHz, CDCl3), δH: 7.86 (d, J = 2.1 Hz, H-4), 7.82 (dd, J = 8.6, 2.2 Hz, H-6), 6.88 (d, J = 8.6 Hz, H-7), 6.24 (m, H-3′), 5.96 (d, J = 7.5 Hz, H-3), 3.88 (d, J = 7.5 Hz, H-2), 2.52 (s, CH3-14), 2.04 (dq, J = 7.3, 1.5 Hz, CH3-4′), 1.93 (p, J = 1.5 Hz, CH3-5′), 1.51 (s, CH3-11), 1.35 (s, CH3-12). 13C-NMR (100 MHz, CDCl3), δC: 196.49 (C-13), 169.43 (C-1′), 157.12 (C-9), 141.17 (C-3′), 132.48 (C-5), 130.42 (C-4), 129.76 (C-6), 126.81(C-2′), 119.53 (C-8), 117.49 (C-7), 79.60 (C-10), 74.03 (C-2), 71.73 (C-3), 26.29 (C-12), 25.88 (C-14), 20.58 (C-5′), 19.73 (C-13) and 16.12 (C-4′), these data match those in the literature [12,55]. Spectra of 1H and 13C-NMR are in Figures S17 and S18.
Selected flowers from the collected plants and were dried in the shade at room temperature. The plant material (256.7 g) was successively extracted with MeOH: H2O (95:05 v/v) three times (each time with 4 L of the solvent). The hydroalcoholic extracts obtained were concentrated to dryness by distillation under reduced pressure using a rotary evaporator, obtaining 6.7 g of residue. Fractionation of the hydroalcoholic extract by open CC (silica gel, 70–230 mesh; 10 cm i.d. × 60 cm) was performed with a gradient system of n-hexane-CH2Cl2: MeOH 90:10:00 to 100% MeOH. Fractions of 100 mL were obtained (53 fractions). Based on TLC analysis, these fractions were grouped according to their chemical profile into two main groups: AP-M-1A (1-32, 2.02 g) and AP-M-1B (33-53, 3.98 g). The groups were subjected to column chromatography using silica gel (70–230 mesh). The AP-M-1A fraction was adsorbed on 3 g of silica gel and placed in a glass column (80 cm high and 3.5 cm in diameter) packed with 70 g of silica gel; elution was carried out using a gradient system, n-hexane: CH2Cl2 (100:00→80:20), 37 fractions were obtained 100 mL. These were concentrated and monitored by TLC and grouped into three groups of fractions. The AP-M-1A-1 group (fractions 1–17, 0.87 g), AP-M-1A-2 (fractions 18-26, 0.606 g) and AP-M-1A-3 (fractions 27-32, 0.87 g). The three groups of fractions were subjected to successive purification processes, obtaining from the AP-M-1A-1 fraction as the main product O-methylencecalinol (1, 32.4 mg), from the AP-M-1A-2 group the encecalin (2, 24 mg) and 3, 5-diprenyl-4-hydroxyacetophenone (5, 16 mg) and the AP-M-1A-3 group speletone (7, 14 mg), dehydrospeletone (8, 11 mg) and encecalinol (9, 17 mg) were purified. Likewise, through successive chromatographers using a gradient system (CH2Cl2: MeOH 95:05 to 80:20), betuletol 3-O-β-glucoside (11, 64 mg) was purified.
Betuletol 3-O-β-glucoside (11)
1H-NMR (500 MHz, DMSO-d6); yellow amorphous solid; mp: 152-154 °C; δH: 12.58 (1H, s, OH), 8.00 (2H, d, J = 8.9 Hz, H-2′ H-6′), 6.90 (1H, s, H-8), 6.82 (2H, d, J = 8.9 Hz, H-3′ H-5′), 5.25 (1H, d, J = 7.7 Hz, H-1″), 3.85 (3H, s, OMe-H-4′), 3.70 (3H, s, OMe-H-6), 3.58-3.56 (1H, ddd, J = 9.7, 7.7, 4.7 Hz, H-4″), 3.32-3.31 (1H, m, H-5″), 3.37-3.36 (1H, m, H-3″), 3.47-3.45 (1H, m, H-2″), 3.33-3.29 (1H, m, H-6″). 13C-NMR (125 MHz, DMSO-d6), δC: 177.86, (C-4), 160.13, (C-4′), 158.80 (C-7), 156.78, (C-2), 151.81 (C-5), 151.68 (C-9), 133.29 (C-3), 131.76 (C-6), 128.57 (C-2′, C-6′), 120.65 (C-1′), 115.56 (C-3′, C-5′), 105.36 (C-10), 101.99 (C-1″), 91.82 (C-8), 73.95 (C-5″), 73.54 (C-3″), 74.13 (C-2″), 70.59 (C-4″), 60.63 (C-6″), 60.64 (C-OMe-6), 56.97 (C-OMe-4′), these data match those in the literature [67,68]. Spectra of 1H and 13C, DEPT and HMBC NMR are in Figures S19 to S22. C23H25O12 (MSFAB+ m/z = 493) MS is in Figure S23.
2.3. In vitro anti-inflammatory activities
For the biological assay, the following reagents were purchased: Murine macrophage cell line RAW 264.7 of ATCC were purchased (Georgetown, Washington, DC, USA ), Fetal bovine serum (FBS), Advanced DMEM/F12 medium, GlutaMAX (Gibco, Waltham, MA USA), GlutaMAX), DMSO, indomethacin, etoposide, LPS (Escherichia coli serotype 055:B5), NaNO2, H3PO4, ursolic acid (UA) and sulfanilamide (Sigma Aldrich, St. Louis, MO, USA), salt MTS (Promega Co), Kit IL-6 (Pharmingen, USA).and MycoZapMT Plus-CL antibiotic (Lonza).
2.3.1. Cell Viability Assay (RAW 264.7)
Murine macrophage cell line RAW 264.7 (ATCC) was grown in ADVANCED DMEM/F12 medium supplemented with 2 mM glutamine and 3.5% heat-inactivated FBS, without antibiotics. Cells were at 37 °C in a humidified incubator in an atmosphere of 5% CO2. RAW 264.7 cells were grown in 24-well plates at a density of approximately 1X105 cells per well. The compounds were dissolved in dimethyl sulfoxide (DMSO) and filtered through 0.45 μm cellulose membranes [53].
Cell viability was determined by MTS (Promega) assay. Cells were seeded in 96-well plates (2 x 105 cells/well) in 100 µL of medium and incubated for 24 h. The cells were treated with various concentrations (0−75 μM) of test compounds (1-11), vehicle (DMSO, 0.5%, v/v) or etoposide (positive control, 68 μM) and incubated for 20 h. After treatment, 20 μL of MTS was added to each well, and the cells were incubated for another 4 h. The optical density was measured at 490 nm on a microplate reader (Epoch microplate spectrophotometer, Bio-Tek) [69,70].
2.3.2. Stimulation of RAW 264.7 Cells with LPS
RAW 264.7 macrophages were cultured at 2 X 105 cells/mL density in 96-well plates for 24 h. The cells were then pre-treated with compounds (1-5, 7-11) at noncytotoxic concentrations (0-75 µM), or vehicle (DMSO, 0.5%, v/v), or indomethacin (84 μM) for 2 h, and after two h, were incubated with LPS (10 μg/mL) for 20 h to stimulate NO production. Cell-free supernatants were collected and were kept at-20 °C until NO quantification. The suppressive effect of compounds on NO production was assessed using the Griess reagent [71,72,73].
2.3.3. Determination of NO concentration
Nitrite released in the culture medium was measured according to Griess reaction. Briefly, 50 μL of each cell culture supernatant was mixed with 100 μL of Griess reagent (50 μL of 1% sulfanilamide and 50 μL of 0.1% N-(1-naphtyl) ethylenediamine dihydrochloride in 2.5% phosphoric acid), for 10 min at room temperature. The optical density at 540 nm (OD540) was measured with a microplate reader and nitrite concentration in the samples was calculated by comparison with the OD540 of a standard curve of NaNO2 prepared in fresh culture medium [45,74,75].
2.3.4. Analysis of Pro-Inflammatory Cytokine IL-6
The anti-inflammatory effect of compound 11 was analyzed in RAW 264.7 cells induced by LPS. Cells were seeded in 96-well plates at a density of 3 X 104 cells/well incubated for 24 h. The cells were expressed with 1 microgram/ml of LPS and compound 11 at concentrations 37.5, 75, and 150 microgram/mL for 24 h. IL-6 levels in the cell culture medium were measured using an Elisa kit following the manufacturer’s instructions (Pharmingen, USA) [76,77].
2.4. Treatment of RAW-Blue Cells with LPS
RAW-Blue Cells (3 × 104 cells/well) were plated in 96-well plates in 0.1 mL of DMEM/F12 medium supplemented with 10% FBS, 1% MycoZap, and 200 μg/mL Zeocin, and incubated for 24 h at 37 °C with 5% CO2. Macrophages were then incubated with the test compounds for 2 hours at the maximum noncytotoxic concentration that showed an inhibitory effect on NO production to subsequently be incubated with LPS at 10 μg/mL (for wells with compounds and 100% control). stimulus) as a pro-inflammatory stimulus and without LPS (negative control) at 37 °C for 20 h to stimulate NF-κβ activation. Finally, cell-free supernatants were collected and used fresh to determine NF-κβ activation.
2.5. Statistical analysis
The results shown were obtained at least by three independent experiments and are presented as means ± SDs. Statistical analyses were performed by one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test. All statistical analyses were performed using GraphPad Prism, version 6.0 software. p values < 0.05 were considered to indicate statistical significance [80].
3. Results and discussion
3.1. Chemical Composition
The ethyl acetate (leaves) and hydroalcoholic (flowers) extracts obtained from the species A. pichinchensis were subjected to successive purification processes using the column chromatography technique and normal phase silica gel, a gradient elution system.-hexane: ethyl acetate (100:00→50:50 v/v) and n-hexane:CH2Cl2:CH3OH (90:10:00→00:00:100 v/v). After several successive purification processes, pure compounds were obtained and identified by analysis of their spectroscopic data and by comparison with literature data. From the ethyl acetate extract (leaves) were identified: O-methylencecalinol (1) [55], encecalin (2) [12,55], eupatoriochromene (3) [81], 6-acetyl-8-hydroxy-2,2-dimethylchromene (4) [82,83,84], 3, 5-diprenyl-4-hydroxyacetophenone (5) [53,55], (+)-β-eudesmol (6) [50,51,55], espeletone (7) [52,55], dehydrospeletone (8) [52,53], encecalinol (9) [54,55,86], 5-acetyl-3β-angeloyloxy-2β-(1-hydroxyisopropyl)-2,3-dihydrobenzofurane (10) [12] and of the hydroalcoholic extract (flowers), in addition to the compounds (1, 2, 5 and 6), betuletol 3-O-β-glucoside (11) was identified [55,69].
Compounds (3, 4, 7, and 11) were isolated from this plant for the first time and investigated the abilities of compounds 1–11 (Figure 1) to inhibit NO production.
Since there is little evidence on the chemical characterization of compound 11, we will provide more information on its structural elucidation in this study. Compound (11) was obtained as a yellow-colored amorphous solid; the analysis of the data obtained by 1H, 13C, and DEPT NMR (S1, S2, and S3), the two-dimensional heteronuclear HMBC experiment (S4) as well as in FAB+ mode mass spectrometry (spectrum 5) gave evidence on the chemical structure of the obtained solid. In the hydrogen NMR spectrum, two double signals are observed at (δ 6.88 and δ 8.12), which show ortho-type coupling constants (J = 8.7 Hz and 8.9 Hz) which are characteristic of the symmetric hydrogens that are in a 1,4.-disubstituted aromatic system, in addition to these signals, a single signal is observed at δ 6.9 and that is assigned to a pentasubstituted benzene. At δ 5.44 another double signal appears (J= 7.7 Hz), as well as several signals between δ 3.5 and δ 4.5 characteristic of a glucopyranose; These signals are evidence in the 13C NMR spectrum, indicating that the aglycone is glycosylated. Once these signals were assigned, it was established that it was a glycosylated flavonol and that, based on the HMBC experiment (S5), the sugar molecule is connected to the oxygen in the 3-position of the aglycone. Once the structural identity was established, the yellow solid was identified as betuletol 3-O-β-glucoside.
3.2. Inhibition of LPS-induced NO production by compounds 1-11
The evaluation of cytotoxicity in RAW 264.7 cells by MTS indicated that up to 75 microg/mL for 48 h of incubation, compounds 1-5 and 7-11 did not affect cell viability. After treatment with LPS (1 μg/mL) for 24 h, the concentration of nitrite in the medium increased markedly. When RAW 264.7 cells were treated with different concentrations of compounds 1 to 5 and 7 to 11, a significant inhibition by compound 11 on nitrite production was detected as a concentration-dependent response. The IC50 values of compounds 1-11 to inhibit LPS-induced NO production are presented in Table 1.
The cell viability assay showed that, except for compound 6 (61.14%), all the compounds tested at concentrations of 9.4, 18.8, 37.5 and 75 µM did not affect the viability of macrophages and showed survival rates greater than 90%. Based on these results, except compound 6 (37.5 µM), the compounds were tested at 75 µM to evaluate their effect on nitric oxide production in RAW 264.7 cells stimulated with LPS. Results showed that only compound 11 inhibited the production of NO in a dose-dependent manner (Figure 2a), with an IC50 value of 20.55 ± 0.27 µM. This compound turned out to be more active than indomethacin, since the maximum evaluated concentration (75 µM) inhibited by 75.08 ± 3.07% NO production, and indomethacin inhibited by 65.93 ± 6.03 at the concentration of 84 µMThis effect was not due to cytotoxicity, since it did not affect cell viability of the RAW 264.7 cells up to a concentration of 75 µM. Inflammation is a defense mechanism against external stimuli such as infections (bacteria) or internal stimuli inherent to living organisms (stress); however, excessive inflammation can cause chronic inflammatory problems. Some researchers indicate that resident macrophages are key in the Inflammation cascade, which is why the inhibition of inflammatory mediators or the NF-kB signaling pathway in macrophages can be useful in the development of therapies aimed at specific pro-inflammatory [61]. Also studied was the effect of compound 11 on the expression of cytokine IL6 y factor nuclear kappa B (KF-kB). Per the previous results, the effect of compound 11 on the secretion of the proinflammatory cytokines IL-6 and NF-kβ are shown in Figures 2b and 2c. As mentioned before, it is well known that in the inflammatory process, the anti- and pro-inflammatory cytokines play an important role in the activation, maintenance, and regulation of inflammation [40,41,42,43].
The compound 11, like some essential oils, showed an anti-inflammatory effect demonstrated by the inhibition of nitric oxide associated with regulating the expression of pro-inflammatory cytokines [63,64,65]. Other compounds have shown similar effects to compound (11) in the same inflammation model, such as the case of the compound 4-methoxycinnamyl p-coumarate isolated from the species Etlingera pavieana, which revealed an IC50 of 15.0 ± 1.4 µM [66]. On the other hand, the triterpene methyl lucidenate L isolated from the species Ganoderma lucidum exhibited an IC50 of 36.8 ± 1.0 µM [67]. It should be noted that compound (11) has not been reported in Ageratina species, although it has been reported in species of the Asteraceae family, for example, in the species Arnica montana and Arnica chamissonis, both plants have been used in traditional medicine as a healing and anti-inflammatory agent at the extract level [55,69]. These species are characterized by containing sesquiterpene lactones such as helenalin, 11α,13-dihydrohelenalin, and chamissonolide, which inhibit activation of transcription factor NF-κβ [70].
Regarding compound (6), it was not evaluated to determine its effect on nitric oxide secretion, this due to the cell viability assay having a survival rate of RAW 264.7 macrophages of less than 90% (61.14 ± 6.31%) at the highest concentration evaluated, however, at a concentration of 37 mM it does not inhibit cell viability (104.00 ± 3.64 µM). β-Eudesmol is a sesquiterpene that has already been isolated from several species, to mention a few, there are found Atracrylodes lancea [57], Zingiber Zerumbet [71], Guttería friesiana [72] and Murraya tetramera [73]. Likewise, several biological effects have been demonstrated for this secondary metabolite, of which we can highlight its anti-inflammatory effect, as a tumor suppressor, and anticancer [71,72,73]. The above agrees with the inhibitory effect that compound (6) showed on cell viability.
4. Conclusions
The inhibitory effect on NO production in an in vitro inflammatory model was evaluated of compounds (1-5, 7-11) isolated from leaves and flowers of A. pichinchensis. Among these, the betuletol 3-O-β-glucoside (11) inhibited the production of NO in a dose-dependent manner, in particular, treatment with 75 µM significantly decreased NO production by 75.08 ± 3.07% (IC50 = 20.55 ± 0.27). The effect demonstrated by the inhibition of nitric oxide associated with the regulation of the expression of proinflammatory cytokines (IL-6) and inhibition of NF-kβ activation indicates that compound 11 may be useful as a therapeutic agent in the treatment of inflammation-related diseases caused by the overactivation of macrophages.
Supplementary Materials
Effect of compounds (1-11) on cell viability and NO production in RAW 264.7 cells, at different concentrations, tables 1 and 2. 1H and 13C NMR spectra of compounds (1-6 and 9-10) Figures S1-S12 and S15-S18, 1H NMR spectra of compounds (7-8) Figure S13-S14, 1D and 2D NMR spectra of compound (11), Figures S19-S22 and Mass-Mass Spectrum (FAB+), Figure S23.
Author Contributions
Funding acquisition, Author Contributions: Conceptualization, S.M.-B.; Formal analysis, A.R.-E., A.R.-G. and J. G.- C.; Funding acquisition, F.C.-S., and L.A.; Investigation, M.S.-R. and S.M.-B.; Methodology, A.R.-E., A.G.-A., J. G.- C. and M.S.-R.; Resources, F.C.-S., and L.A.; Supervision, L. A., A.R.-G., F. C.-S., and M.S.-R.; Writing: S. M. B. and M. S. R. original draft; Writing—review & editing, S. M.- B.,M. S.- R and F. C.-S.
Acknowledgments
This research was supported in part by Conahcyt (Grant CB 240801). The authors thank Laboratorio Nacional de Estructura de Macromoléculas (Conahcyt 279905) for the spectroscopic and mass analyses. The authors thank MSc Gabriel Flores curator of the HUMO Herbarium for his support in the taxonomic identification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McVaugh, R. Compositae. Flora Novo-Galiciana. A Descriptive Account of the Vascular Plants of Western Mexico; The University of Michigan Press: Ann Arbor, MI, USA, 1984; Volume 12.
- GBFI. Ageratina pichinchensis. Available online: https://www.gbif.org/es/species/5400249 (accessed on 01 December 2023).
- Villaseñor, R.; Espinosa, G.F.J. Catálogo de Malezas de México; Universidad Nacional Autónoma de México: México City, México, 1998; pp. 1-448.
- Villaseñor, J.L. Diversidad y distribución de la familia Asteraceae en México. Bot. Sci. 2018, 96, 332-358.
- INAH. Jardín Etnobotánico y Museo de la Medicina Tradicional. 2023. Available online: https://lugares.inah.gob.mx/es/museos-inah/colecciones/piezas/12915-12915-axihuitl.html?lugar_id=389 (accessed on 1 December 2023).
- Argueta, A.; Cano, L.; Rodarte, M. Atlas de la Medicina Tradicional Mexicana, Tomo 1-3; Instituto Nacional Indigenista: Mexico City; Mexico, 1994; p. 1786.
- Avilés, M.; Suárez, G. Catálogo de Plantas Medicinales. Jardín Etnobotánico; Centro INAH: Cuernavaca, Mexico, 1994; p. 47.
- Ríos, M.Y.; Aguilar-Guadarrama, B.; Navarro, V. Two new benzofurans from Eupatorium aschenbornianum and their antimicrobial activity. Planta Med. 2003, 69, 967-970. [CrossRef]
- Navarro-García, V.M.; Gonzalez, A.; Fuentes, M.; Aviles, M.; Ríos, M.Y.; Zepeda, G.; Rojas, M.G. Antifungal activities of nine traditional Mexican medicinal plants. J. Ethnopharmacol. 2003, 87, 85-88. [CrossRef]
- Sánchez-Mendoza, M.E.; Reyes-Trejo, B.; Sánchez-Gómez, P.; Rodriguez-Silverio, J.; Castillo-Henkel, C.; Cervantes-Cuevas, H.; Arrieta, J. Bioassay-guided isolation of an anti-ulcer chromene from Eupatorium aschembornianum: Role of nitric oxide, prostaglandins and sulfydryls. Fitoterapia 2010, 81, 66-71. [CrossRef]
- Sánchez-Mendoza, M.; Rodriguez-Silverio, J.; Rivero-Cruz, J.F. Rocha-González, H.; Pineda-Farías, J.; Arrieta, J. Antinociceptive effect and gastroprotective mechanisms of 3,5-diprenyl-4-hydroxyacetophenone from Ageratina pichinchensis. Fitoterapia 2013, 87, 11-19. [CrossRef]
- Aguilar-Guadarrama, B.; Navarro, V.; León-Rivera, I.; Ríos, M.Y.; Active compounds against tinea pedis dermatophytes from Ageratina pichinchensis var. bustamenta. Nat. Prod. Res. 2009, 23, 1559-1565. [CrossRef]
- Romero-Cerecero, O.; Zamilpa, A.; Jiménez, E.; Tortoriello, J. Effect on the wound healing process and in vitro cell proliferation by the medical Mexican plant Ageratina pichinchensis. Planta Med. 2011, 77, 979-983. [CrossRef]
- Torres-Barajas, L.; Rojas-Vera, J.; Morales-Méndez, A.; Rojas-Fermín, L.; Lucena, M.; Buitrago, A. Chemical composition and evaluations of antibacterial activity of oils of Ageratina jahnii and Ageratina pichinchensis collected in Mérida, Venezuela. Bol. Latinioam. Caribe Plant. Med. Aromat. 2013, 12, 92-98.
- Romero-Cerecero, O.; Zamilpa, A.; González-Cortazar, M.; Alonso-Cortés, D.; Jiménez-Ferrer, E.; Nicasio-Torres, P.; Aguilar-Santamaría, L.; Tortoriello, J. Pharmacological and chemical study to identify wound-healing active compounds in Ageratina pichinchensis. Planta Med. 2013, 79, 622-627. [CrossRef]
- White, M.M.D. Mediators of inflammation and the inflammatory process. Journal of Allergy and Clinical Immunology. 1999, 103, S978-S981. [CrossRef]
- Duleba, M.D.A.J.; Dokras, M.D.A. Is PCOS an inflammatory process? Fertility and Sterility. 2012, 97, 7-12. [CrossRef]
- Lugrin, J.; Rosenblantt-Velin, N.; Parapanov, R.; Liaudet, L. The role of oxidative stress during inflammatory processes. Biological Chemistry 2014, 395, 203-230. [CrossRef]
- Adbulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: a review. Veterinary World 2018, 11, 627-635. [CrossRef]
- Sugimoto, M.A.; Vago, J.P.; Perretti, M.; Teixeira M.M. Mediators of the resolutions of the inflammatory response. Trends in Immunology 2019, 1545, 1-16. [CrossRef]
- Veríssimo, F.J.; Oliveira, C.R.N.; Nunes, J.C.A.; Araújo, M.F.T.A.; Veríssimo, A.J.W.; Galvao, A.J.M. The role of the mediators of inflammation in cancer development. Pathol. Onco. Res. 2015, 21, 527-534. [CrossRef]
- Salgado, A.; Bóveda, J.L.; Monasterio, J.; Segura, R.M.; Mourelle, M.; Gómez-Jiménez, J.; Peracaula, R. Inflammatory mediators and their influence on haemostasis. Pathophysiology of Haemostasis and Trombosis 1994, 24, 132-138. [CrossRef]
- Schlag, G.; Redl, H. Mediators of injury and inflammation. World J. Sug. 1996, 20, 406-410. [CrossRef]
- Bradley, T.S.; Geller, D.A. Molecular regulation of the human inducible nictric oxidase synthase (iNOS) gene. Shock 2000, 13, 413-424. [CrossRef]
- Krishna, R.K.M. Molecular mechanisms regulating iNOS expression in various cell types. J Toxicol Environ Health B Crit Rev 2000, 3, 27-58. [CrossRef]
- Chiou, W.-F.; Chen, C.-F.; Lin, J.-J. Mechanisms of suppression of inducible nitric oxide synthase (iNOS) expression in RAW 264.7 cells by andrographolide. Br. J. Pharmacol. 2000, 129, 1553-1560. [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639-653. [CrossRef]
- Panaro, M.A.; Brandonisio, O.; Acquafredda, A.; Sisto, M.; Mitolo, V. Evidences for iNOS Expression and Nitric Oxide Production in the Human Macrophages. Curr Drug Targ Immune, Endocrine & Metabolic Disorders 2003, 3, 210-221. [CrossRef]
- Poteser, M.; Wakabayashi, I. Serum albumin induces iNOS expression and NO production in RAW 264.7 macrophages. Br. J. Pharmacol. 2004, 143, 143-151. [CrossRef]
- Gosgnach, W.; Messika-Zeitoun, D.; Gonzalez, W.; Philipe, M.; Michel, J.B. Shear stress induces iNOS expression in cultured smooth muscle cells: role of oxidative stress. Am J Physiol Cell Physiol 2000, 279, C1880-C1888. [CrossRef]
- Anavi, S.; Tirosh, O. iNOS as a metabolic enzyme under stress conditions. Free Radic. Biol. Med. 2020, 146, 16-35. [CrossRef]
- Weinberg, J.B.; Misukonis, M.A.; Shami, P.J.; Mason, S.N.; Sauls, D.L.; Dittman, W.A.; Wood, E.R.; Smith, G.K.; McDonald, B.; Bachus, K.E. Human molecular phagocyte inducible nitric oxide synthase (iNOS): analysis of iNOS mRNA, iNOS protein, biopterin, and nitric oxide production by blood monocytes and peritoneal macrophages. Blood 1995, 83, 1184-1195.
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of iNOS on immune cells and its role in diseases. Int. J. Mol. Sci. 2018, 19, 3805. [CrossRef]
- Xu, W.; Liu, L.Z.; Loizidou, M.; Ahmed, M.; Charles, I.G. The role of nitric oxide in cancer. Cell Research 2002, 12, 311-320. [CrossRef]
- Xie, Q.-W.; Nathan, C. The high-output nitric oxide pathway: role and regulation. J. Leuk. Biol. 1994, 56, 576-582. [CrossRef]
- Bruckdorfer, R. The basics about nitric oxide. Mol. Asp. Med. 2005, 26, 3-31. [CrossRef]
- Nagy, G.; Clark, J.M.; Buzás, E.I.; Gorman, C.L.; Cope, A.P. Nitric oxide, chronic inflammation and autoimmune. Immunol Let 2007, 111, 1-5. [CrossRef]
- Laroux, F.S.; Pavlick, K.P.; Hines, I.N.; Kawachi, S.; Harada, H.; Bharwani, S.; Hoffman, J.M.; Grisham, M.B. Role of nitric oxide in inflammation. Acta Physiol Scand 2001, 173, 113-118. [CrossRef]
- Papi, S.; Ahmadizar, F.; Hasanvand, A. The role of nitric oxide in inflammation and oxidative stress. Immunopathol Persa 2019, 5, e08. [CrossRef]
- Tripathi, P.; Tripathi, P.; Kashyap, L.; Singh, V. The role of nitric oxide in inflammatory reactions. Pathog. Dis. 2007, 51, 443-452. [CrossRef]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible nitric oxide synthase and inflammatory diseases. Molecular Medicine 2000, 6, 347-373. [CrossRef]
- Maniscalco, M.; Sofia, M.; Pelaia, G. Nitric oxide in upper airways inflammatory diseases. Inflamm. Res. 2007, 56, 58-69. [CrossRef]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immune-pathogenesis of inflammatory bowel diseases. World J Gatrointest Pharmacol Ther 2016, 7, 353-360. [CrossRef]
- Rask-Madsen, P. Review article: the potential role of nitric oxide in chronic inflammatory bowel disorders. AP&T 2001, 13, 135-144. [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of the oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252-259. [CrossRef]
- Blantz, R.C.; Munger, K. Role of nitric oxide in inflammatory conditions. Nephron 2002, 90, 373-378. [CrossRef]
- Amirghofran, Z.; Malek-Hosseini, S.; Golmoghaddam, H.; Kalantar, F.; Shabani, M. Inhibition of nitric oxide production and proinflammatory cytokines by several medicinal plants. Iran. J. Immunol. 2011, 8, 159-169.
- Perera, H.D. S. M.; Samarasekera, J.K.R.R.; Handunnetti, S.M.; Weerasena, D.S.J.; Weeratunga, H.D.; Jabeen, A.; Choudhary, M.I. In vitro pro-inflammatory enzyme inhibition and anti-oxidant potential of selected Sri Lankan medicinal plants. BMC 2018, 18, 271. [CrossRef]
- Jae-Ha, R.; Han-Na, A.; Hwa-Jin, L.; Li, F.; Wen-He, Q.; Yong-Nam, H.; Byung-Hoon, H. Inhibitory activity of Chinese medicinal plants on nitric oxide synthesis in lipopolysaccharide activated macrophages. Biomolecules & Therapeutics 2001, 9, 183-187.
- Achenbach, H.; Waibel, R.; Addae-Mensah, I. Constituents of West African medicinal plants. Part 17. Sesquiterpenes from Carissa edulis. Phytochemistry 1985, 24, 2325-8.
- You, C.X.; Yang, K.; Wang, C.F.; Zhang, W.J.; Wang, Y.; Han, J.; Fan, L.; Du, S.S.; Geng, Z.F.; Deng, Z.W. Cytotoxic compounds isolated from Murraya tetramera huang. Mol. 2014, 19, 13225-13234. [CrossRef]
- Dupre, S.; Bohlmann, F.; Knox, E. Prenylated p-hydroxyacetophenone derivatives from the giant Senecio johnstonii. Biochem. Syst. Ecol. 1990, 18, 149-50. [CrossRef]
- Bohlmann, F.; Rao, N. New hydroxyacetophenone derivatives from Espeletia Schultzii. Chem. Ber. 1973, 106, 3035-8.
- Becerra, J.; Silva, M.; Delle-Monache, G.; Delle-Monache, F.; Botta, M. Two new chromenes from Eupatorium glechonophyllum Less. Rev. Lat. Quím. 1983, 14, 92-4.
- Merfort, I.; Wendisch, D. Flavonoid glycosides from Arnica montana and Arnica chamissonis. Planta Med. 1987, 53, 434-7. [CrossRef]
- Juergens, U.R. Anti-inflammatory properties of the monoterpene 1.8-cineole: Current evidence for co-medication in inflammatory airway diseases. Drug Res. 2014, 64, 638–646. [CrossRef]
- Schwartz, M.A. Syntheses of (+)-α- and (+)-β-eudesmol and their diastereomers by intramolecular nitrone-olefin cycloaddition. J. Org. Chem. 1985, 50, 1359–1365. [CrossRef]
- Ismail, M.; Javed, S.; Kazim, M.; Razaq, A.; Hussain, E.; Attia-tul-Wahab; Ali, S.; Choudhary, Muhammad Iqbal. Phenylpropanoids from Tanacetum baltistanicum with Nematocidal and Insecticidal Activities. Chem. Nat. Compd. 2022, 58, 637-643. [CrossRef]
- Bohlmann, F.; Vorwerk, E. Synthesis of naturally occurring p-hydroxyacetophenone derivatives. II. Chem. Ber. 1980, 113, 261-6. [CrossRef]
- Bjeldanes, L.; Geissman, T. Euparinoid constituents of Encelia californica. Phytochemistry. 1969, 8, 1293–1296. [CrossRef]
- Sfikakis P.P. The first decade of biologic TNF antagonists in clinical practice: lessons learned, unresolved issues and future directions. Curr Dir Autoimmun. 2010, 11, 180–210. [CrossRef]
- Owona, B.A.; Njayou, N.F.; Laufer, S.; Moundipa, P.F.; Schluesener, H.J. A fraction of stem bark extract of Entada africana suppresses lipopolysaccharide-induced inflammation in RAW 264.7 cells. J. Ethnopharmacol. 2013, 149, 162–168. [CrossRef]
- Almeida, J.R.G.S.; Souza, G.R.; Silva, J.C.; Saraiva, S.R.G.D.; Junior, R.G.O.; Quintans, J.S.S.; Barreto, R.S.S.; Bonjardim, L.R.; Cavalcanti, S.C.H.; Junior, L.J.Q. Borneol, a bicyclic monoterpene alcohol, reduces nociceptive behavior and inflammatory response in mice. Sci. World J. 2013, 808460. [CrossRef]
- Choi, Y.K.; Cho, G.S.; Hwang, S.; Kim, B.W.; Lim, J.H.; Lee, J.C.; Kim, H.C.; Kim, W.K.; Kim, Y.S. Methyleugenol reduces cerebral ischemic injury by suppression of oxidative injury and inflammation. Free Radical. Res. 2010, 44, 925–935. [CrossRef]
- Kim, D.S.; Lee, H.J.; Jeon, Y.D.; Han, Y.H.; Kee, J.Y.; Kim, H.J.; Shin, H.J.; Kang, J.W.; Lee, B.S.; Kim, S.H.; Kim, S.J.; Park, S.H.; Choi, B.M.; Park, S.J.; Um, J.Y.; Hong, S.H. Alpha-pinene exhibits anti-inflammatory activity through the suppression of MAPKs and the NF-kB pathway in mouse peritoneal macrophages. Am. J. Chin. Med. 2015, 43, 731–742. [CrossRef]
- Mankhong, S.; Iawsipo, P.; Srisook, E.; Srisook, K. 4-methoxycinnamyl p-coumarate isolated from Etlingera pavieana rhizomes inhibits inflammatory response via suppression of NF-κB, Akt and AP-1 signaling in LPS-stimulated RAW 264.7 macrophages. Phytomedicine. 2019, 54, 89-97. [CrossRef]
- Wu, Y.L.; Han, F.; Luan, S.S.; Ai, R.; Zhang, P.; Li, H.; Chen, L.X. Triterpenoids from Ganoderma lucidum and Their Potential Anti-inflammatory Effect. J. Agric. Food Chem. 2019, 67, 5147-5158. [CrossRef]
- Iloki, S.B.; Gil-Salido, A.A.; Lewis, L.M.; Rosas-Durazo, A.; Acosta-Silva, A.L.; Rivera-Castañeda, E.G.; Rubio-Pino, J.L. Cell growth curves for different cell lines and their relationship with biological activities. Int. J. Biotechnol. Mol. Biol. Res. 2013, 4, 60−70.
- Merfort, I.; Wendisch, D. New flavonoid glycosides from Arnicae flos DAB 91. Planta Med. 1992, 58, 355-7. [CrossRef]
- Lyss, G.; Schmidt, T.J.; Merfort, I.; Pahl, H.L. Helenalin, an anti-inflammatory sesquiterpene lactone from Arnica, selectively inhibits transcription factor NF-κB. Biol. Chem. 1997, 378, 951-961. [CrossRef]
- Nam, S.Y.; Kim, H.Y.; Kim, H.M.; Jeong, H.J. Βeta-eudesmol reduces stem cell factor-induced mast cell migration. Int. Immunopharmacol. 2017, 48, 1–7. [CrossRef]
- Ma, E.L.; Li, Y.C.; Tsuneki, H.; Xiao, J.F.; Xia, M.; Wang, M.W.; Kimura, I. β-Eudesmol suppresses tumour growth through inhibition of tumour neovascularisation and tumour cell proliferatio. J. Asian Nat. Prod. Res. 2008, 10, 159–167. [CrossRef]
- Seo, M.J.; Kim, S.J.; Kang, T.H.; Rim, H.K.; Jeong, H.J.; Um, J.Y.; Hong, S.H.; Kim, H.M. The regulatory mechanism of β-eudesmol is through the suppression of caspase-1 activation in mast cell–mediated inflammatory response. Immunopharmacol Immunotoxicol. 2011, 33, 178–185. [CrossRef]
- Yang G, Lee K, Lee M, Ham I, Choi H-Y. Inhibition of lipopolysaccharide induced nitric oxide and prostaglandin E2 production by chloroform fraction of Cudrania tricuspidata in RAW 264.7 macrophages. BMC 2012, 12, 250. [CrossRef]
- Brice Ayissi Owona, B.; Frederic Njayou, N.; Laufer, S.; Fewou Moundip, P.; Schluesener, H.J. A fraction of stem bark extract of Entada africana suppresses lipopolysaccharide-induced inflammation in RAW264.7cells. J. Ethnopharmacol 2013, 149, 162–168. [CrossRef]
- An, X.; Gil Lee, S.; Kang, H.; Jin Heo, H.; Cho, Y.S: and Ok Kim, D. Antioxidant and Anti-Inflammatory Effects of Various Cultivars of Kiwi Berry (Actinidia arguta) on Lipopolysaccharide-Stimulated RAW 264.7 Cells. J. Microbiol. Biotechnol. 2016, 26, 1367–1374. [CrossRef]
- Lim JY, Won TJ, Hwang BY, Kim HR, Hwang KW, Sul D, et al. The new diterpene isodojaponin D inhibited LPS- induced microglial activation through NF-kβ and MAPK signaling pathways. Eur J Pharmacol 2010, 642, 10–8. [CrossRef]
- Checker, R.; Sandur, S.K.; Sharma, D.; Patwardhan, R.S.; Jayakumar, S.; Kohli, V.; Sethi, G.; Aggarwal, B.B.; Sainis, K.B. Potent anti-inflamatory activity of ursolic acid, a triterpenoid antioxidant, is mediated throgh suppresion of NF-kB, AP-1 and NF-AT. PLoS ONE 2012, 7, e31318. [CrossRef]
- Shishodia, S.; Majumdar, S.; Banerjee, S.; Aggarwal, B.B. Ursolic acid inhibits nuclear factor-kB activation induced by carcinogenic agents through suppression of IƙBα kinase and p65 phosphorylation: Correlation with down-regulation of cyclooxygenase 2, matrix metalloproteinase 9, and cyclin D1. Cancer Res. 2003, 63, 4375–4383.
- Sánchez-Ramos, M.; Marquina, B.S.; Romero-Estrada, A.; Bernabé-Antonio, B.; Cruz-Sosa, F.; González-Christen, J.; Acevedo-Fernández, J.J.; Perea-Arango, I.; Alvarez, L. Establishment and phytochemical analysis of a callus cultures from Ageratina pichinchensis (Asteraceae) and its anti-inflammatory activity. Molecules 2018, 23, 1258. [CrossRef]
- Schwartz, M.A. Syntheses of (+)-α- and (+)-β-eudesmol and their diastereomers by intramolecular nitrone-olefin cycloaddition. J. Org. Chem. 1985, 50, 1359–1365. [CrossRef]
- Shamsuddin, K.M.; Musharraf, M.A.; Zobairi, M.O.; Ali, N. Demethylacetovanillochromene from Tithonia diversifolia (Hemes1.) A. Gray. Indian J. Chem.- Sect. B Org. Med. Chem. 2001, 8, 751-752.
- Zhai, H.L.; Zhao, G.J.; Yang, G.J.; Sun, H.; Yi, B.; Sun, L.N.; Chen, W.S.; Zheng, S.Q. A new chromene glycoside from Tithonia diversifolia. Chem. Nat. Compd. 2010, 46, 198-200. [CrossRef]
- Aguilar, M.I.; Delgado, Guillermo; Bye, R.; Linares, E. Bisabolenes, polycyclic diterpenoids and other constituents from the roots of Iostephane heterophylla. Phytochemistry. 1993, 33, 1161-3. [CrossRef]
- Bohlmann, F.; Zdero, C.; Franke, H. Naturally occurring coumarin derivatives. IX. Constituents of the genus Gerbera. Chem. Ber. 1973, 106, 382-7. [CrossRef]
- Castañeda, P.; Gómez, L.; Mata, R.; Lotina-Hennsen, B.; Anaya, A.L.; Bye, R. Phytogrowth-Inhibitory and Antifungal Constituents of Helianthella quinquenervis. J. Nat. Prod. 1996, 59, 323–326. [CrossRef]
Figure 1.
Compounds isolated from aerial parts of A. pichinchensis.
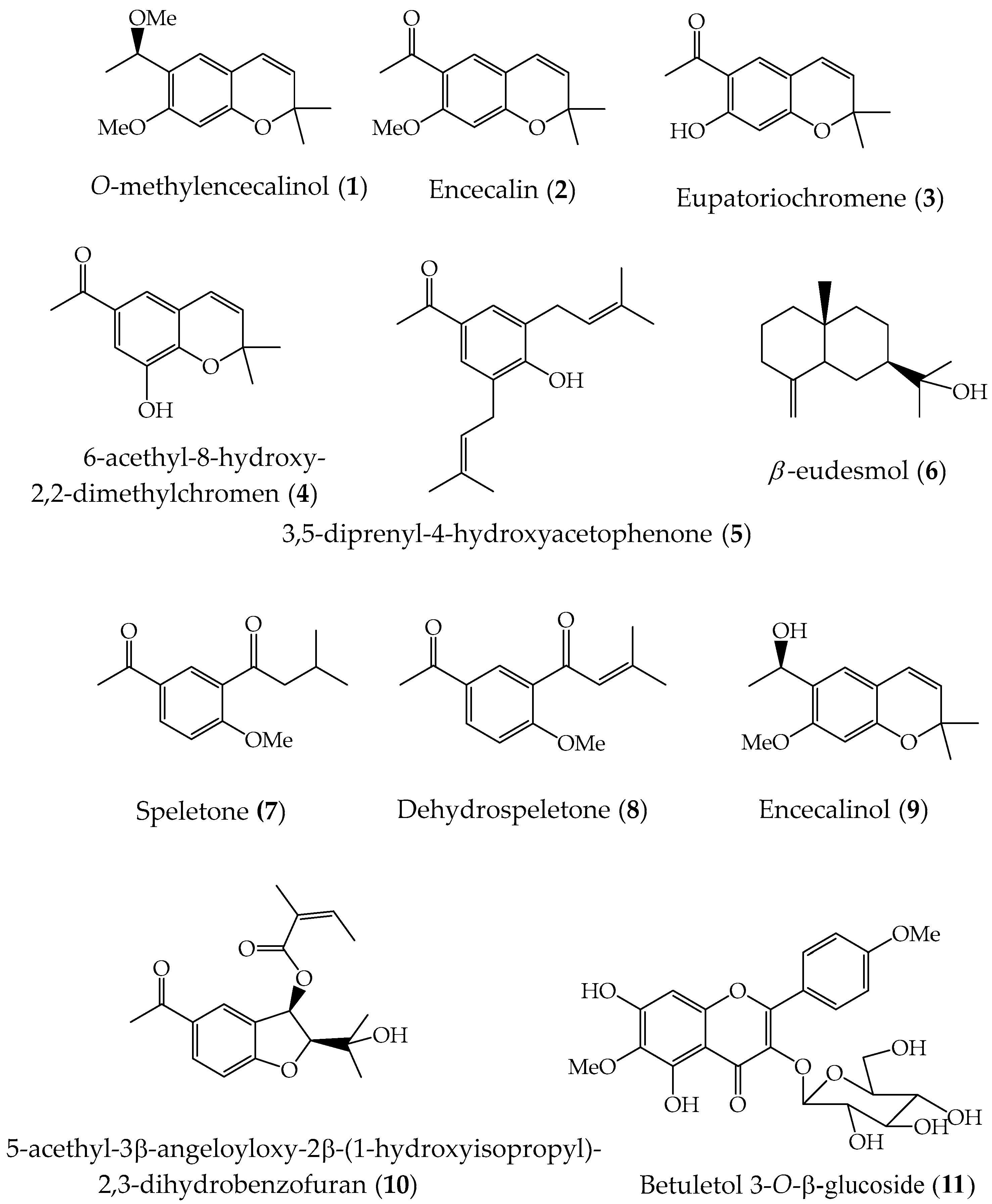
Figure 2.
Effect of compound 11 on a) NO, b) IL-6 and c) NF-kβ of RAW 264.7 macrophages activated with LPS. Data are expressed as mean ± SD values of experiments in triplicate independent assays. ** p < 0.01, ***p < 0.001 vs LPS-treated cells; ####p < 0.001 vs vehicle control.
Figure 2.
Effect of compound 11 on a) NO, b) IL-6 and c) NF-kβ of RAW 264.7 macrophages activated with LPS. Data are expressed as mean ± SD values of experiments in triplicate independent assays. ** p < 0.01, ***p < 0.001 vs LPS-treated cells; ####p < 0.001 vs vehicle control.


Table 1.
Percentage of inhibition of NO production and cell viability in RAW 264.7 macrophages at a concentration of 75 µM of compounds 1-11.
Table 1.
Percentage of inhibition of NO production and cell viability in RAW 264.7 macrophages at a concentration of 75 µM of compounds 1-11.
| Compounds | Cell viability (%)a | NO Inhibition (%) | NO inhibition (IC50, µM) |
|---|---|---|---|
| 1 | 109.9 ± 7.16 | 0.95 ± 1.35 | > 75 |
| 2 | 112.4 ± 24.08 | 16.75 ± 5.36 | > 75 |
| 3 | 115.6 ± 1.58 | 11.98 ± 7.85 | > 75 |
| 4 | 99.33 ± 12.39 | 22.63 ± 10.38 | > 75 |
| 5 | 100.20 ± 2.95 | 29.77 ± 18.27 | ˃ 75 |
| 6 | 61.14 ± 6.31 | ----- | ----- |
| 7 | 104.7 ± 1.82 | 5.90 ± 8.35 | > 75 |
| 8 | 103.9 ± 3.83 | 36.73 ± 16.93 | > 75 |
| 9 | 110.9 ± 8.3 | 29.77 ± 9.37 | ˃ 75 |
| 10 | 121.2 ± 10.20 | 5.98 ± 5.22 | > 75 |
| 11 | 101.3 ± 1.62 | 75.08 ± 3.07 | 20.55 ± 0.27 |
| DMSOb | ----- | ----- | ----- |
| Indomethacinc (84 µM) | ----- | 65.93 ± 6.03 | 54.69 ± 10.34 |
| Etoposided (68 µM) | 42.02 ± 4.23 | ----- | ----- |
a Cell viability at 75 µM. b Blank control. c Positive control for NO production assay. d Positive control for cytotoxicity against RAW 264.7 cell.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



