
Submitted:
08 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Transitions from seed to seedling represent a critical developmental step in the life cycle of higher plants, dramatically affecting plant ontogenesis and stress tolerance. The release from dormancy to acquiring the germination ability is defined by a balance of phytohormones, with a substantial contribution of abscisic acid (ABA), which inhibits germination. We studied the embryonic axes of Pisum sativum L. before and after radicle protrusion. Our previous work compared RNA sequencing-based transcriptomics in the embryonic axes isolated before and after radicle protrusion. The current study aims to analyze ABA-dependent gene regulation during the transition of embryonic axes from germination to post-germination stage. First, we determined the levels of abscisates (ABA, phaseic acid, dihydrophaseic acid, and neo-phaseic acid) using the ultra-high-performance liquid chromatography-tandem mass spectrometry. Second, we made a detailed annotation of ABA-associated genes using RNA sequencing-based transcriptome profiling. Finally, we analyzed the DNA methylation pattern in the promoters of the PsABI3, PsABI4, and PsABI5 genes. We showed that changes in the abscisate profile are characterized by the accumulation of ABA catabolites, and the ABA-related gene profile is accompanied by the upregulation of genes controlling the seedling development and the downregulation of genes controlling the water deprivation. The expression of ABI3, ABI4 and ABI5, which encode the crucial transcription factors during late maturation, was downregulated by more than 20-fold, and their promoters exhibited a high level of methylation already at the late germination stage. Thus, although ABA remains important, other regulators seems to be involved in the transition from seed to seedling.
Keywords:
abscisic acid
; DNA methylation
; embryonic axes
; ABA-associated genes
; Pisum sativum L.
; seed-to-seedling transition
1. Introduction
In higher plants, seed production is crucial to the species survival. Most seeds enter dormancy during late maturation and maintain this state until environmental conditions become favorable for germination [1,2]. The transition from dormancy to germination is influenced by a balance of phytohormones and significant environmental factors such as temperature, water availability, and light [1,3]. This transition, occurring at the end of germination, involves extensive transcriptome reprogramming and signaling pathway alterations, leading to the silencing of seed maturation genes and activation of those for vegetative growth [4,5,6,7,8,9].
Whether seeds acquire the ability to germinate or remain dormant depends on the phytohormones balance [10,11,12]. Notably, abscisic acid (ABA) promotes seed dormancy and inhibits germination, while gibberellins (GAs) breaks seed dormancy and induce germination [13,14,15,16,17,18]. During early embryogenesis, ABA prevents seed abortion and promotes embryo growth, initially provided by the maternal tissues and later produced by the seeds themselves [18,19]. Consequently, ABA levels rise sharply late in embryogenesis, counteracting GAs and suppressing embryo growth [19].
As the embryo develops, it enlarges through cell elongation and accumulates storage compounds. ABA regulates the transport of monosaccharides and amino acids from maternal tissues and their conversion into stored forms, like polysaccharides and proteins. In late maturation, metabolic processes slow down, seeds desiccate, and enter dormancy [20,21].
Numerous studies have shown that decreasing ABA levels is crucial for dormancy release and germination [7,12,19,22]. ABA degradation occurs through hydroxylation and conjugation, with ABA 8'-hydroxylases playing a key role in rapid ABA level decline during seed imbibition [17,23,24,25,26]. However, ABA's signaling role during the seed-to-seedling transition remains unclear.
Key player in the seed transition from dormancy to germination include the LAFL regulatory network, comprising LEAFY COTYLEDON1 (LEC1) and LEC1-LIKE (L1L) of the NF-YB family transcription factors (TFs), and ABSCISIC ACID INSENSITIVE3 (ABI3), FUSCA3 (FUS3), and LEC2 (LEAFY COTYLEDON2) of the B3-AFL gene family [27,28,29]. The LAFL network, originating in a common ancestor of bryophytes and vascular plants, acts as a positive regulator of seed maturation genes but suppresses germination [30,31,32]. This network allows orthodox seeds to maintain desiccation tolerance during dormancy and germination [33,34,35,36]. Radicle protrusion marks the transition to the post-germination stage, with seeds becoming seedlings and losing desiccation tolerance [9,33,37]. This stage is typically associated with LAFL network silencing [5,32,34,38,39].
Our previous transcriptomic profiling of P. sativum embryo axes before and after radicle protrusion revealed unexpected findings [4]. Although we anticipated the expression of LAFL network genes before radicle protrusion and their subsequent silencing, only PsABI3 showed significant expression in seed axes. We also observed the expression of other ABA-related genes (PsABI4 and PsABI5). Given that ABI3, ABI4, and ABI5 are central transcriptional factors in seed-specific events, including maturation, dormancy, longevity, germination, and post-germination growth [16,40,41], we propose that PsABI3, PsABI4, and PsABI5 also play a role in regulating the P. sativum seed-to-seedling transition [4,9].
Germination-related repression of the LAFL transcriptional network is due to epigenetic regulation of gene expression through DNA methylation and post-translational modifications of histones [5,8,32,42,43,44]. DNA methylation patterns change throughout seed development, germination and seedling establishment [8,45,46,47,48,49,50,51,52,53]. This study analyzes ABA metabolite profiles, ABA-associated gene expression, and DNA methylation in the promoters of PsABI3, PsABI4, and PsABI5 in embryonic axes of germinated pea seeds before and after radicle protrusion. We discuss these findings in the context of ABA-dependent gene regulation during the seed-to-seedling transition.
2. Materials and Methods
2.1. Plant Material
Pea seeds of the commercial cultivar “Prima” were sourced from the N.I. Vavilov All-Russian Institute of Plant Genetic Resources, St. Petersburg, Russia. Seeds were imbibed for 72 hours between layers of moist filter paper, then visually divided into two batches: (a) before embryonic root growth initiation (before radicle protrusion) and (b) post-initiation of root growth (after radicle protrusion). Seed axes from both batches were isolated, frozen in liquid nitrogen, homogenized, and stored at −80°C before use in biochemical experiments.
2.2. Quantitation of ABA and ABA-related metabolites
The selected plant hormones in the embryonic axes were quantified using ultra-high performance liquid chromatography coupled with tandem mass spectrometry (UHPLC-MS/MS). The sample preparation and the analysis was performed according to the modified protocol by Šimura and co-workers [54]. For the quantitation of ABA and ABA-related metabolites, 15 mg (fresh weight) of the homogenized plant material were extracted in 1 mL 60% (v/v) acetonitrile (ACN) with the addition of 5 pmol of [2H6]ABA as the internal standard. Four zirconium oxide 2.0 mm extraction beads (Next Advance, Troy, NY, USA) were added into the liquid sample. The sample was shaked in a Retsch MM400 bead mill (Retsch, Haan, Germany) at 27 Hz for 5 minutes, sonicated for 3 minutes and incubated for half an hour at 4 °C. Afterwards, the sample was centrifuged at 20,000 rpm for 10 minutes at 4 °C (Allegra 64R benchtop centrifuge, Beckman Coulter, USA). Supernatant was loaded onto Oasis® HLB 30 mg/1 cc extraction cartridge (Waters, Milford, USA). The cartridge was washed subsequently with 0.5 mL 60% (v/v) ACN and 0.5 mL 30% (v/v) ACN. All fractions (the flow-through and both washes) were collected and dried under the reduced pressure using SpeedVac concentrator (RC1010 Centrivap Jouan, ThermoFisher, USA). The sample was reconstructed in 40 µL of 25% (v/v) ACN and 5 µL of the sample were injected into Acquity UPLC CSH C18 RP 150 × 2.1 mm, 1.7 μm chromatographic column (Waters, Milford, USA). The UHPLC separation was performed using Acquity UPLC I-Class System (Waters, Milford, USA) coupled to a triple quadrupole tandem mass spectrometer Xevo TQ-XS equipped with an electrospray ionization (Waters, Manchester, UK). The gradient elution and the MS/MS working in the multiple reaction monitoring (MRM) mode followed the previously published conditions described in Šimura et al, 2018 [54]. The obtained chromatographic peaks were evaluated in MassLynx V4.2 software (Waters, Manchester, UK). The targeted compounds were quantified by isotope dilution method.
2.3. Annotation of ABA-Associated Genes
ABA-associated genes were annotated based on RNA sequencing-based transcriptome profiling [4]. Annotation utilized the Ensembl BioMart tool (https://plants.ensembl.org/biomart/martview) and the URGI database (https://urgi.versailles.inra.fr/Species/Pisum). Gene ontology (GO) terms, InterPro domains (https://www.ebi.ac.uk/interpro), and Arabidopsis thaliana orthologs were identified for each gene [55]. Clustering was performed using the k-means algorithm, and the optimal number of clusters was determined using the Elbow method.
2.4. DNA Extraction and Sodium Bisulfite Treatment
Total genomic DNA from seeds at two developmental stages (before and after radicle protrusion) was extracted using the DNeasy Plant Mini Kit (QIAGEN, Germany) according to the manufacturer’s instructions (www.qiagen.com). Sodium bisulfite treatment of 1 μg genomic DNA from each sample was conducted with the EpiTect Fast Bisulfite Kit (QIAGEN, Germany).
2.5. Primer Design and in silico Analysis
Primers for amplifying bisulfite-treated DNA were designed against cytosine-converted sequences using SnapGene 6.1.2 (https://www.snapgene.com). Prediction of CpG islands in PsABI3, PsABI4, and PsABI5 promoter sequences utilized Meth-Primer 2.0 (https://www.urogene.org/methprimer2) and PlantPAN 3.0 (http://plantpan.itps.ncku.edu.tw/index.html). Promoter mapping for transcription factor binding sites was performed using PlantPAN 3.0 and PCBase (http://pcbase.itps.ncku.edu.tw/index), followed by filtering for stress and hormone response motifs at Similar Score = 1.
2.6. PCR, Electrophoretic Analysis, Extraction, and Purification
To amplify genomic and bisulfite-treated DNA, the PCR was performed in a 50 μL mixture containing 70 ng of DNA template, 10 pM of each primer, and BioMaster HS-Taq PCR kit (2×) (BioLabMix, Russia) or Tersus Plus PCR kit (Evrogen, Russia) according to the manufacturer's instructions. The PCR conditions included an initial denaturation step at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing 50 °C for 1 min, and extension at 72 °C for 2 min, and a final elongation step at 72 °C for 5 min. The PCR screening of colonies was performed in a 25 μL mixture containing 10 pM of M13F and M13R primers (Evrogen, Russia), 0.25 mM of each dNTP, 1x reaction buffer (67 mM TrisHCl, pH 8.8; 2 mM MgCl2; 18 mM (NH4)2SO4; 0.01% Tween 20) and 0,5 U Taq polymerase (Syntol, Russia). After an initial denaturation at 95°C for 15 minutes, 35 cycles were performed at 94°C for 20 seconds, 55°C for 30 seconds, and 72°C for 1 minute, followed by a final elongation at 72°C for 5 minutes. Electrophoretic analysis was performed on a 1% agarose gel (Helicon, Russia) prepared on a TAE buffer (Sigma-Aldrich, USA) with ethidium bromide (VWR (Amresco), USA). The amplified fragments were extracted from the gel using the MinElute Gel Extraction Kit (QIAGEN, Germany).
2.7. Cloning and Sequencing the Amplified PCR Fragments
Freshly prepared PCR products were ligated with a vector using the Quick-TA kit (Evrogen, Russia), which included the pAL2-T vector, Quick-TA T4 DNA Ligase, buffer, M13 Forward primer, and M13 Reverse primer according to the manufacturer's instructions. Chemical transformation of competent Escherichia coli (Migula 1895) Castellani and Chalmers 1919 DH10B cells was then performed. Transformed colonies carrying inserts of the expected size were selected on selective LB medium (DIA-M, Russia) with 100 µg/ml of ampicillin (BioChemica, PanReac Applichem, Spain). The purified amplified fragments were sequenced in both directions using M13 primers and the BigDye™ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems™, USA) on a 3500 Applied Biosystems Genetic Analyzer. For DNA methylation analysis, at least 10 clones were sequenced for each amplicon. The alignment of sequences was carried out using SnapGene 6.1.2 (https://www.snapgene.com/).
2.8. Statistical Analyses
Two-tailed t-tests (alpha = 0.05) compared means of ABA-related metabolites. Analysis was performed using MS Excel add-in, with data representing mean ± standard error of nine biological replicates.
3. Results
3.1. Quantitation of ABA and ABA-related metabolites in Pea Embryonic Axes Before and After Radicle Protrusion
To delve deeper into ABA homeostasis, we examined the levels of ABA and its metabolites in embryonic axes from germinated pea seeds, both before and after radicle protrusion. These axes encompass the first true leaves, epicotyl, hypocotyl, and root (Figure 1).
We analyzed levels of abscisic acid (ABA), phaseic acid (PA), dihydrophaseic acid (DPA), neo-phaseic acid (neoPA), and 7′-hydroxy ABA (7′-OH-ABA). Notably, we observed an accumulation of PA and DPA, which are key products of ABA catabolism, against a backdrop of decreasing ABA levels (Figure 2). Intriguingly, 7′-OH-ABA was not detected in embryonic axes before or after radicle protrusion.
3.2. Categorization and Functional Annotation of ABA-Associated DEGs in Pea Embryonic Axes Before and After Radicle Protrusion
In our previous work we performed RNA sequencing of isolated embryonic axes before and after radicle protrusion [4]. Here, we provide a more detail profiling of ABA-associated differentially expressed genes (DEGs) annotated using the Pea Genome Assembly v1a from the UGRI server as an main annotation source [56]. A total of 70 ABA-associated DEGs were annotated in embryonic axes. Among these, 46 genes showed higher expression and 24 genes lower expression after radicle protrusion by more than 4-fold (|logFC| > 2) (Figure 3, Figure S1, Table S1).
ABA-dependent DEGs up-regulated in seed axes after radicle protrusion included those related to cellular signaling and stress resistance, membrane transporters and TFs regulating developmental programs (Table S1). These comprised genes encoding serine-threonine/tyrosine protein kinases CRK29 (Psat6g212040) and CIPK17 (Psat0s2012g0280), protein phosphatase 2C family member (Psat7g017080), α-subunit of G-protein (Psat6g097080), and inositol polyphosphate-related phosphatase (Psat4g078320). The expression of genes associated with water deprivation response, antifungal proteins, and calcium-signaling also increased significantly. The expression of Psat6g199400 encoding a protein RD29B/LTI65 increased 4.5-fold, Psat5g266320 encoding an antifungal protein ginkbilobin-2 – 5-8-fold, and Psat4g146960 encoding a calcium-signaling protein ANNEXIN4 – 9-fold. Genes responsible for the synthesis of membrane transporters included Psat4g117800 (encoding P-ATPase) and Psat4g184760 (encoding potassium channel AKT2/3). The expression of Psat2g121520 (encoding the TCP15 protein) increased 8-fold.
Conversely, downregulated DEGs included key ABA-response genes like ABI5 (Psat3g033680), ABI3 (Psat3g142040), ABI4 (Psat2g031240), LTI65 (Psat0s2227g0040), LTP4 (Psat7g227120), HVA22E (Psat5g052360), and RD22 (Psat6g033920 and Psat6g033960) (Table S1). These genes are highly conserved across functional domains, with ABI4, ABI5, and HVA22E exhibiting sequence homology in various drought-tolerant species [4]. These findings suggest these genes play a crucial role in dehydration tolerance during the transition from seed germination to seedling establishment.
3.3. DNA Methylation in the Promoters of the PsABI3, PsABI4, and PsABI5 Genes
We selected the PsABI3 gene along with newly identified drought-responsive genes PsABI4 and PsABI5 for epigenetic analysis. These genes were identified in the P. sativum genome and sequenced from the commercial cultivar “Prima” (Table S2).
In silico analysis of the promoters and first exons (including 5'-UTR) of PsABI3, PsABI4, and PsABI5 revealed low GC composition (29%, 34%, and 23% respectively), with only individual CpG sites predicted and no CpG islands detected (Figure S2). Considering that plant DNA methylation can occur at CpG, CpHpG, and CpHpH sites, we designed primers for bisulfite sequencing (with conversion of unmethylated C to T) of both CpG and non-CpG sites (Table S2).
To analyze the methylation profile of the promoter and the beginning of the first exon of the PsABI3, PsABI4 and PsABI5 genes, we performed the amplification of bisulfite-treated DNA using designed primers (Table S3). Bisulfite-treated DNA amplification and subsequent cloning revealed methylation in the promoters of PsABI3, PsABI4, and PsABI5 already before radicle protrusion (Figure 4).
Additionally, we mapped the promoters of these genes to compare potential methylation sites and binding sites for TFs (Table S4). Notably, the PsABI4 promoter had the lowest number of TFs binding sites, while PsABI5's promoter contained numerous potential LAFL protein binding sites, along with motifs associated with responses to cold and water deprivation.
4. Discussion
4.1. ABA catabolism
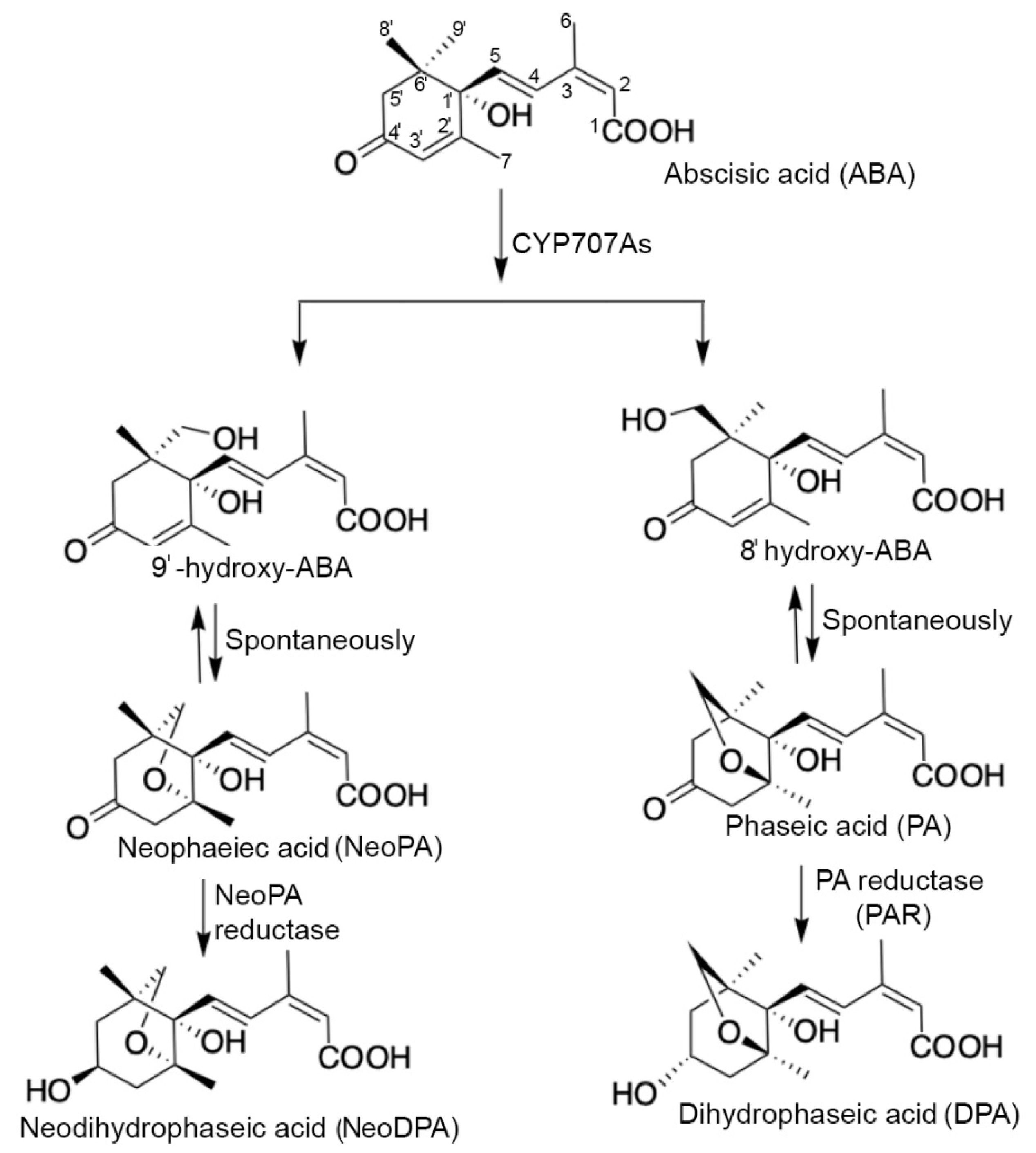
ABA plays vital roles in seed development and maturation, encompassing the accumulation of storage compounds, acquisition of desiccation tolerance, induction of dormancy, and suppression of precocious germination [12,17,18,19,57,58,59]. However, to break dormancy and initiate germination, ABA needs to be catabolized, primarily through hydroxylation and conjugation. The primary ABA hydroxylation route is the ABA catabolic pathway (Figure 5), which relies on the activities of CYP707A cytochromes P450, notably ABA 8'-hydroxylases [60].
Initially, ABA is catalyzed by 8'-hydroxylase, converting it to 8'-hydroxy ABA (8'-OH ABA), an unstable intermediate [61,62]. This intermediate is then spontaneously rearranged into PA and subsequently reduced by PA reductase (PAR) to DPA [24,63]. The 9'-hydroxylation pathway, similar to 8'-hydroxylation, involves CYP707As enzymes and converts 9'-hydroxy ABA (9'-OH ABA) to neoPA with both 8'-C and 9'-C hydroxylations catalyzed by the same enzyme [16]. Recently, Bai et al. (2022) [24] identified a downstream catabolite of neoPA in the 9'-hydroxylation pathway as epi-neodihydrophaseic acid (epi-neoDPA) and discovered the responsible enzyme, neoPA reductase 1 (NeoPAR1) (Figure 5).
Our study examined ABA and ABA-related catabolites in embryonic axes of P. sativum seeds before and after radicle protrusion. We found a decline in ABA content with a concurrent rise in its catabolites (PA, DPA, and neoPA) (Figure 2). Intriguingly, PA, similar to ABA, can regulate stomatal closure and suppress seed germination [64,65]. Weng et al. (2016) Weng et al. (2016) demonstrated that PA functions as a signaling molecule through ABA receptors. Similar ABA-like hormonal activity was observed for neoPA, but not for epi-neoDPA [24]. Additionally, altered seed germination patterns were noted in neo-PAR1 mutant and overexpression lines, implicating the ABA catabolic pathway as a critical regulatory mechanism during the seed-to-seedling transition [24]. Despite reduced ABA levels, the accumulation of its catabolic products (PA, neoDPA) in embryonic axes suggests a continued regulatory influence via ABA receptors.
4.2. Annotation of ABA-associated DEGs
In our prior RNA sequencing-based transcriptomic analysis of pea embryonic axes isolated from seeds before and after radicle protrusion [4], we identified 24,184 DEGs, with 2,101 showing notably higher expression. This work extends that analysis by focusing on ABA-associated DEGs (ABA-DEGs). Of the 70 ABA-DEGs annotated, 46 genes up-regulated and 24 genes down-regulated more than 4-fold after radicle protrusion (Figure 3).
The up-regulated ABA-DEGs predominantly pertained to cellular signaling, stress resistance, membrane transporters, and transcription factors that regulate seedling development. For instance, Psat6g199400, encoding RD29B/LTI65 which responds to water deprivation, was up-regulated 4.5-fold. This gene's promoter region contains two ABA-responsive elements (ABREs) that required as cis-acting elements for the dehydration-responsive expression of RD29B/LTI65 [66,67] Similarly, Psat4g146960, encoding ANNEXIN4, a calcium-binding protein involved in drought and other stress responses [68,69], showed a 9-fold increase in expression (Table S1).
Among the up-regulated genes were those coding for membrane transporters like Psat4g117800 (P-ATPase) and Psat4g184760 (potassium channel AKT2/3). P-type ATPases play a role in ion transport across membranes, utilizing ATP for transmembrane conformational changes [70,71]. Additionally, Psat2g121520, encoding TCP15, a transcription factor implicated in cell expansion and proliferation [72,73] was up-regulated 8-fold. The TCP proteins, known as TEOSINTE BRANCHED 1 (TB1) in maize, CYCLOIDEA (CYC) in Anthirrinum majus, and PCF in rice [74] have been linked to various developmental processes, including light-induced cotyledon opening in Arabidopsis [75].
Conversely, down-regulated ABA-DEGs included genes central to ABA signaling (ABI3, ABI4, ABI5) and those involved in water deprivation response (LEA14, RD22, HVA22, PER1, and LTI65) (Table S1). Seed germination is governed by the antagonistic balance of ABA/GA, with ABA catabolism preceding GA synthesis and activation [5,7,17]. Key ABA signaling genes ABI3, ABI4, and ABI5 encode the TFs featuring B3, AP2, and bZIP domains which control the expression of ABA-responsive genes crucial for seed maturation, dormancy, longevity, germination, and post-germination growth ([12,16,76,77,78]. .
ABI5 encodes a member of the basic leucine zipper TF family and involved in ABA signaling in seeds by acting as a signal integrator between ABA and other hormones [41,79,80]. The Arabidopsis abi5 mutants have pleiotropic defects in ABA response, including reduced sensitivity to ABA, inhibition of germination, and altered expression of some ABA-regulated genes [81,82]. Notably, Psat3g033680, encoding ABI5, exhibited a 22-fold downregulation after radicle protrusion.
ABI4 was shown to be a key integration node for multiple signals, participating in critical transition steps during plant ontogenesis [83,84,85]. In dormant seeds, ABI4 acts as a repressor of ABA catabolism by binding to the promoter of CYP707A, being the main enzyme of ABA catabolism [86]. Thereby, ABA and GAs can antagonistically modify the expression and stability of ABI4, suggesting the existence of regulatory loops [83]. In germinating seeds, ABI4 can regulate both ABA synthesis and catabolism. Some authors suggest that ABI4 plays as a key regulator of the balance between ABA and GAs in seeds at post-germination stages [83,85]. In our study, the level of the Psat2g031240 gene encoding ABI4 was decreased 21-fold.
ABI3 encoded AP2/B3-like transcriptional factor family protein [87]. ABI3 belongs to the LAFL regulatory network where interacts with LEAFY COTYLEDON1 (LEC1), ABSCISIC ACID INSENSITIVE3 (ABI3), FUSCA3 (FUS3), and LEC2 [31,32]. The LAFL network is a positive regulator of seed dormancy and need to be suppressed for seed germination. Together, ABI3, FUS3, and LEC1 are involved in sensitivity of seeds to ABA, and regulate the expression of the 12S storage protein gene family [88]. In addition, both FUS3 and LEC1 positively regulate the ABI3 protein abundance in the seeds [89]. The expression of Psat3g142040 encoded ABI3 was decreased 21-fold.
We also found the downregulation of ABA-dependent genes involved in response to water deprivation (LEA14, RD22, HVA22, PER1 and LTI65) (Table S1). In accordance with our findings, Psat7g085840 encoding peroxiredoxin1 (PER1), Psat0s2227g0040 encoding protein LTI65/78, and Psat0s2780g0040 encoding late embryogenesis abundant (LEA) protein were down-regulated 20-30-fold. Peroxiredoxins are thiol-dependent antioxidants containing one (1-Cys) or two (2-Cys) conserved Cys residues [90]. PER1 encodes a 1-Cys peroxiredoxin (PER1) protein that accumulates during seed development but rapidly disappears upon germination [91]. PER1 is involved in the quenching reactive oxygen species (ROS) during late maturation, dormancy, and early germination, thereby maintaining seed viability [91,92,93]. The low-temperature-induced (LTI) protein family is associated with responses to abiotic stresses. In Arabidopsis, the homologous genes RD29A (LTI78) and RD29B (LTI65) are induced by cold, drought, salt, and abscisic acid [66]. Most LEA genes have ABA response elements in promoters and their expression can be induced not only by ABA, but also by cold or drought. The desiccation-related protein LEA14 belongs to the group II LEA proteins, also known as dehydrins [94]. LEA14 was induced in response to salt and low temperature [95].
4.3. Epigenetic regulation of PsABI3, PsABI4, and PsABI5 genes based on DNA promoter methylation.
Major transitions in the plant life cycle require fine-tuned regulation at the molecular and cellular levels. Epigenetic regulation, particularly DNA methylation, is crucial for maintaining genome stability in plants by inhibiting transposable element movement and modulating gene expression during development and stress responses [8,96]. DNA methylation patterns in seeds undergo significant changes during development and germination [8,46,47,50,52,97,98]. DNA methylation (mC) occurs in three sequence contexts (CG, CHG, and CHH) and refers as addition a methyl group to the C5 position of cytosine to form 5-methylcytosine [96]. Methylation of CHH sites notably increases from early to late stages of seed development, then decreases during germination [8,47,48]. Two DNA methylases, RdDM (RNA directed DNA methylation) and CMT2 (DOMAINS REARRANGED METHYLTRANSFERASE 2), responsible for methylating CHH sites in developing seeds, are inactivated during germination [51,52]. In contrast, CG and CHG methylation patterns are relatively stable throughout seed development [46,98,99]. Therefore, monitoring the level of 5-methylcytosine (m5C) is considered as a universal marker for seeds at the different stages of their ontogenesis [97].
Our study reveals that during the transition from germination to post-germination, expression of key ABA signaling pathway genes (ABI3, ABI4, and ABI5) is markedly suppressed. We analyzed the DNA promoter methylation profiles of PsABI3, PsABI4, and PsABI5 to understand their epigenetic regulation. Contrary to our expectations of low promoter methylation levels based on their expression before radicle protrusion [4], we observed high methylation levels both before and after this developmental stage (Figure 4). Notably, approximately one-third of the PsABI3 gene promoter region showed reduced methylation. However, this region might belong to the 5'-UTR as per the Pea Genome International Consortium version 1a (Figure S3, pink).
We further investigated the coincidence of epigenetic marks with transcription factor binding sites in the promoters of these genes, using PlantPAN 3.0 and PCBase, focusing on stress and hormone response motifs. PsABI5 showed numerous potential binding sites for LAFL network proteins, along with motifs associated with cold and water deprivation responses (Figure S4). This finding aligns with the role of ABI5 as a major regulator of seed maturation and longevity in legumes [41]. Our results suggest that epigenetic modifications, impacting the binding ability of ABI3, ABI4, and ABI5 to DNA promoters, occur prior to the initiation of the seed transition from germination to post-germination.
Thus, our study provided an insight into the involvement of ABA in the transition in P. sativum from germination to the post-germination stage when seeds turn into seedlings. The initiation of embryonic axis growth corresponds with changes in the abscisate profile: a decrease in ABA levels and an accumulation of its catabolites (PA, DPA, and neoPA), which possess hormonal activity similar to ABA [24,100]. Our in-depth analysis of ABA-DEGs revealed 46 up-regulated and 24 down-regulated genes with more than 4-fold changes. Most up-regulated ABA-DEGs were related to the regulation of seedling development. Most notably, the expression of ABI3, ABI4, and ABI5 was significantly downregulated, and their promoters exhibited a high level of methylation both before and after radicle protrusion. While ABA continues to be important, other regulators appear to be involved in the seed-to-seedling transition.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Expression heatmap of ABA-associated DEGs in embryonic axes of P. sativum before and after radicle protrusion (RP); Figure S2: Predicted methylation sites for CpG motifs in the sequences of the seed resistance to dehydration genes in the P. sativum genome; Figure S3: Mapping of the PsABI3 gene promoter; Figure S4: Mapping of the PsABI4 gene promoter; Figure S5: Mapping of the PsABI5 gene promoter; Table S1: Annotation of the ABA-associated genes in the transcriptome of P. sativum embryonic axes; Table S2: The ABA-dependent genes of P. sativum seeds selected for analysis of DNA methylation in the promoters; Table S3: Primers used for amplification of gene promoter regions from bisulfite-treated DNA isolated from embryonic axes of P. sativum; Table S4: Functions of transcription factors, binding sites of which were identified in gene promoter regions PsABI3, PsABI4 и PsABI5.
Author Contributions
G.S. and S.M. conceived the project. E.K., P.V. and I.P. performed the experiments. K.S., E.K. and A.V. analyzed data. G.S., K.S. and S.M. wrote the manuscript. M.S., A.F. and E.Kh. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the infrastructural support provided by the Timiryazev Institute of Plant Physiology, Russian Academy of Sciences (theme No. 1021052706080-4-1.6.11). The authors are grateful to Ms. Tatiana Leonova and Ms. Alena Kusnetsova for help in preparing plant material for phytohormone analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chahtane, H.; Kim, W.; Lopez-Molina, L. Primary seed dormancy: a temporally multilayered riddle waiting to be unlocked. J. Exp. Bot. 2017, 68, 857–869. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. Arab. B. 2008, 6, e0119. [Google Scholar] [CrossRef]
- Smolikova, G.; Strygina, K.; Krylova, E.; Vikhorev, A.; Bilova, T.; Frolov, A.; Khlestkina, E.; Medvedev, S. Seed-to-seedling transition in Pisum sativum L.: a transcriptomic approach. Plants 2022, 11, 1686. [Google Scholar] [CrossRef]
- Smolikova, G.; Strygina, K.; Krylova, E.; Leonova, T.; Frolov, A.; Khlestkina, E.; Medvedev, S. Transition from seeds to seedlings: hormonal and epigenetic aspects. Plants 2021, 10, 1884. [Google Scholar] [CrossRef]
- Silva, A.T.; Ligterink, W.; Hilhorst, H.W.M. Metabolite profiling and associated gene expression reveal two metabolic shifts during the seed-to-seedling transition in Arabidopsis thaliana. Plant Mol. Biol. 2017, 95, 481–496. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An updated overview on the regulation of seed germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Luján-Soto, E.; Dinkova, T.D. Time to wake up: Epigenetic and small-RNA-mediated regulation during seed germination. Plants 2021, 10, 236. [Google Scholar] [CrossRef]
- Smolikova, G.; Medvedev, S. Seed-to-seedling transition : Novel aspects. Plants 2022, 11, 1988. [Google Scholar] [CrossRef]
- Finkelstein, R.R. The role of hormones during seed development and germination. In Plant Hormones; Davies, P.J., Ed.; Springer Netherlands: Dordrecht, 2010; pp. 549–573. ISBN 978-1-4020-2684-3. [Google Scholar]
- Shu, K.; Meng, Y.J.; Shuai, H.W.; Liu, W.G.; Du, J.B.; Liu, J.; Yang, W.Y. Dormancy and germination: How does the crop seed decide? Plant Biol. 2015, 17, 1104–1112. [Google Scholar] [CrossRef]
- Ali, F.; Qanmber, G.; Li, F.; Wang, Z. Updated role of ABA in seed maturation, dormancy, and germination. J. Adv. Res. 2022, 35, 199–214. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.; Xie, Q.; He, Z. Two faces of one seed: hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The pivotal role of abscisic acid signaling during transition from seed maturation to germination. Plant Cell Rep. 2017, 36, 689–703. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Steber, C.M. GA signaling is essential for the embryo-to-seedling transition during Arabidopsis seed germination, a ghost story. Plant Signal. Behav. 2020, 15, 1705028. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.; Bressan, R.A.; Song, C.; Zhu, J.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Sano, N.; Marion-Poll, A. ABA metabolism and homeostasis in seed dormancy and germination. Int. J. Mol. Sci. 2021, 22, 5069. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. B. 2013, 11, e0166. [Google Scholar] [CrossRef]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic acid and the control of seed dormancy and germination. Seed Sci. Res. 2010, 20, 55. [Google Scholar] [CrossRef]
- Gutierrez, L.; Van Wuytswinkel, O.; Castelain, M.; Bellini, C. Combined networks regulating seed maturation. Trends Plant Sci. 2007, 12, 294–300. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Bai, Y.L.; Yin, X.; Xiong, C.F.; Cai, B.D.; Wu, Y.; Zhang, X.Y.; Wei, Z.; Ye, T.; Feng, Y.Q. Neophaseic acid catabolism in the 9′-hydroxylation pathway of abscisic acid in Arabidopsis thaliana. Plant Commun. 2022, 3, 100340. [Google Scholar] [CrossRef]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K.; Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8′-hydroxylases: key enzymes in ABA catabolism. EMBO J. 2004, 23, 1647–1656. [Google Scholar] [CrossRef]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol. 2006, 141, 97–107. [Google Scholar] [CrossRef]
- Carbonero, P.; Iglesias-Fernández, R.; Vicente-Carbajosa, J. The AFL subfamily of B3 transcription factors: evolution and function in angiosperm seeds. J. Exp. Bot. 2016, 68, erw458. [Google Scholar] [CrossRef]
- Han, J.-D.; Li, X.; Jiang, C.-K.; Wong, G.K.-S.; Rothfels, C.J.; Rao, G.-Y. Evolutionary analysis of the LAFL genes involved in the land plant seed maturation program. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Roscoe, T.T.; Guilleminot, J.; Bessoule, J.J.J.-J.; Berger, F.; Devic, M. Complementation of seed maturation phenotypes by ectopic expression of ABSCISIC ACID INSENSITIVE3, FUSCA3 and LEAFY COTYLEDON2 in arabidopsis. Plant Cell Physiol. 2015, 56, 1215–1228. [Google Scholar] [CrossRef]
- Cagliari, A.; Turchetto-Zolet, A.C.; Korbes, A.P.; Maraschin, F. dos S.; Margis, R.; Margis-Pinheiro, M. New insights on the evolution of Leafy cotyledon1 (LEC1) type genes in vascular plants. Genomics 2014, 103, 380–387. [CrossRef]
- Jia, H.; Suzuki, M.; McCarty, D.R. Regulation of the seed to seedling developmental phase transition by the <scp>LAFL</scp> and <scp>VAL</scp> transcription factor networks. WIREs Dev. Biol. 2014, 3, 135–145. [Google Scholar] [CrossRef]
- Lepiniec, L.; Devic, M.; Roscoe, T.J.; Bouyer, D.; Zhou, D.-X.; Boulard, C.; Baud, S.; Dubreucq, B. Molecular and epigenetic regulations and functions of the LAFL transcriptional regulators that control seed development. Plant Reprod. 2018, 31, 291–307. [Google Scholar] [CrossRef]
- Faria, J.M.R.; Buitink, J.; van Lammeren, A.A.M.; Hilhorst, H.W.M. Changes in DNA and microtubules during loss and re-establishment of desiccation tolerance in germinating Medicago truncatula seeds. J. Exp. Bot. 2005, 56, 2119–2130. [Google Scholar] [CrossRef]
- Dekkers, B.J.W.; Costa, M.C.D.; Maia, J.; Bentsink, L.; Ligterink, W.; Hilhorst, H.W.M. Acquisition and loss of desiccation tolerance in seeds: from experimental model to biological relevance. Planta 2015, 241, 563–577. [Google Scholar] [CrossRef]
- Smolikova, G.; Leonova, T.; Vashurina, N.; Frolov, A.; Medvedev, S. Desiccation tolerance as the basis of long-term seed viability. Int. J. Mol. Sci. 2021, 22, 101. [Google Scholar] [CrossRef]
- Sano, N.; Lounifi, I.; Cueff, G.; Collet, B.; Clément, G.; Balzergue, S.; Huguet, S.; Valot, B.; Galland, M.; Rajjou, L. Multi-omics approaches unravel specific features of embryo and endosperm in rice seed germination. Front. Plant Sci. 2022, 13, 1–20. [Google Scholar] [CrossRef]
- Buitink, J.; Ly Vu, B.; Satour, P.; Leprince, O. The re-establishment of desiccation tolerance in germinated radicles of Medicago truncatula Gaertn. seeds. Seed Sci. Res. 2003, 13, 273–286. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Saijo, T.; Shibata, D.; Morikami, A.; Nakamura, K. Analysis of a sugar response mutant of Arabidopsis identified a novel B3 domain protein that functions as an active transcriptional repressor. Plant Physiol. 2005, 138, 675–685. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Morikami, A.; Nakamura, K. Two B3 domain transcriptional repressors prevent sugar-inducible expression of seed maturation genes in Arabidopsis seedlings. Proc. Natl. Acad. Sci. 2007, 104, 2543–2547. [Google Scholar] [CrossRef]
- Zinsmeister, J.; Lalanne, D.; Ly Vu, B.; Schoefs, B.; Marchand, J.; Dang, T.T.; Buitink, J.; Leprince, O. ABSCISIC ACID INSENSITIVE4 coordinates eoplast formation to ensure acquisition of seed longevity during maturation in Medicago truncatula. Plant J. 2023, 113, 934–953. [Google Scholar] [CrossRef]
- Zinsmeister, J.; Lalanne, D.; Terrasson, E.; Chatelain, E.; Vandecasteele, C.; Vu, B.L.; Dubois-Laurent, C.; Geoffriau, E.; Signor, C. Le; Dalmais, M.; et al. ABI5 is a regulator of seed maturation and longevity in legumes. Plant Cell 2016, 28, 2735–2754. [Google Scholar] [CrossRef]
- Molitor, A.M.; Bu, Z.; Yu, Y.; Shen, W.-H. Arabidopsis AL PHD-PRC1 complexes promote seed germination through H3K4me3-to-H3K27me3 chromatin state switch in repression of seed developmental genes. PLoS Genet. 2014, 10, e1004091. [Google Scholar] [CrossRef]
- Mozgova, I.; Köhler, C.; Hennig, L. Keeping the gate closed: functions of the polycomb repressive complex PRC2 in development. Plant J. 2015, 83, 121–132. [Google Scholar] [CrossRef]
- Ding, X.; Jia, X.; Xiang, Y.; Jiang, W. Histone modification and chromatin remodeling during the seed life cycle. Front. Plant Sci. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Xing, M.Q.; Zhang, Y.J.; Zhou, S.R.; Hu, W.Y.; Wu, X.T.; Ye, Y.J.; Wu, X.X.; Xiao, Y.P.; Li, X.; Xue, H.W. Global analysis reveals the crucial roles of DNA methylation during rice seed development. Plant Physiol. 2015, 168, 1417–1432. [Google Scholar] [CrossRef]
- Narsai, R.; Gouil, Q.; Secco, D.; Srivastava, A.; Karpievitch, Y. V.; Liew, L.C.; Lister, R.; Lewsey, M.G.; Whelan, J. Extensive transcriptomic and epigenomic remodelling occurs during Arabidopsis thaliana germination. Genome Biol. 2017, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, D.; Kramdi, A.; Kassam, M.; Heese, M.; Schnittger, A.; Roudier, F.; Colot, V. DNA methylation dynamics during early plant life. Genome Biol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Park, K.; Shin, S.-Y.; Frost, J.M.; Hsieh, P.-H.; Shin, C.; Fischer, R.L.; Hsieh, T.-F.; Choi, Y. Distinct regulatory pathways contribute to dynamic CHH methylation patterns in transposable elements throughout Arabidopsis embryogenesis. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Li, W.-Y.; Chen, B.-X.; Chen, Z.-J.; Gao, Y.-T.; Chen, Z.; Liu, J. Reactive oxygen species generated by NADPHoxidases promote radicle protrusion and root elongation during rice seed germination. Int. J. Mol. Sci. 2017, 18, 110. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cabellos, S.; Toorop, P.E.; Cañal, M.J.; Iannetta, P.P.M.; Fernández-Pascual, E.; Pritchard, H.W.; Visscher, A.M. Global DNA methylation and cellular 5-methylcytosine and H4 acetylated patterns in primary and secondary dormant seeds of Capsella bursa-pastoris (L.) Medik. (shepherd’s purse). Protoplasma 2022, 259, 595–614. [Google Scholar] [CrossRef]
- Dew-Budd, K.J.; Chow, H.T.; Kendall, T.; David, B.C.; Rozelle, J.A.; Mosher, R.A.; Beilstein, M.A. RNA-directed DNA Methylation is required for seed development in outbreeding species. bioRxiv 2023, 1–20. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Nery, J.R.; Castanon, R.; Ecker, J.R. Dynamic DNA methylation reconfiguration during seed development and germination. Genome Biol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bartels, A.; Han, Q.; Nair, P.; Stacey, L.; Gaynier, H.; Mosley, M.; Huang, Q.; Pearson, J.; Hsieh, T.-F.; An, Y.-Q.; et al. Dynamic DNA methylation in plant growth and development. Int. J. Mol. Sci. 2018, 19, 2144. [Google Scholar] [CrossRef] [PubMed]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant hormonomics: multiple phytohormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: an integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinformatics 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed]
- Kreplak, J.; Madoui, M.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, F.; Luo, X.; Dai, Y.; Yang, Y.; Zheng, C.; Yang, W.; Shu, K. A matter of life and death: Molecular, physiological, and environmental regulation of seed longevity. Plant Cell Environ. 2020, 43, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant. Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Jikumaru, Y.; Hanada, A.; Nambara, E.; Abrams, S.R.; Kamiya, Y.; Seo, M. Comprehensive hormone profiling in developing Arabidopsis seeds: examination of the site of ABA biosynthesis, ABA transport and hormone interactions. Plant Cell Physiol. 2010, 51, 1988–2001. [Google Scholar] [CrossRef]
- Saito, S.; Hirai, N.; Matsumoto, C.; Ohigashi, H.; Ohta, D.; Sakata, K.; Mizutani, M. Arabidopsis CYP707A s encode (+)-abscisic acid 8′-hydroxylase, a key enzyme in the oOxidative catabolism of abscisic acid. Plant Physiol. 2004, 134, 1439–1449. [Google Scholar] [CrossRef]
- Milborrow, B. V; Vaughan, G.T. Characterization of dihydrophaseic acid 4′-O-β-D-glucopyranoside as a major metabolite of abscisic acid. Funct. Plant Biol. 1982, 9, 361–372. [Google Scholar] [CrossRef]
- Cai, W.-J.; Zeng, C.; Zhang, X.-Y.; Ye, T.; Feng, Y.-Q. A structure–guided screening strategy for the discovery and identification of potential gibberellins from plant samples using liquid chromatography–mass spectrometry assisted by chemical isotope labeling. Anal. Chim. Acta 2021, 1163, 338505. [Google Scholar] [CrossRef]
- Zeevaart, J.A.D.; Milborrow, B. V Metabolism of abscisic acid and the occurrence of epi-dihydrophaseic acid in Phaseolus vulgaris. Phytochemistry 1976, 15, 493–500. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Raschke, K. Effects of phaseic acid and dihydrophaseic acid on stomata and the photosynthetic apparatus. Plant Physiol. 1980, 65, 291–297. [Google Scholar] [CrossRef]
- Walker-Simmons, M.K.; Holappa, L.D.; Abrams, G.D.; Abrams, S.R. ABA metabolites induce group 3 LEA mRNA and inhibit germination in wheat. Physiol. Plant. 1997, 100, 474–480. [Google Scholar] [CrossRef]
- Msanne, J.; Lin, J.; Stone, J.M.; Awada, T. Characterization of abiotic stress-responsive Arabidopsis thaliana RD29A and RD29B genes and evaluation of transgenes. Planta 2011, 234, 97–107. [Google Scholar] [CrossRef]
- Liu, W.; Thapa, P.; Park, S.-W. RD29A and RD29B rearrange genetic and epigenetic markers in priming systemic defense responses against drought and salinity. Plant Sci. 2023, 337, 111895. [Google Scholar] [CrossRef]
- Medvedev, S.S. Principles of calcium signal generation and transduction in plant cells. Russ. J. Plant Physiol. 2018, 65, 771–783. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y. How do plants maintain pH and ion homeostasis under saline-alkali stress? Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Dyla, M.; Basse Hansen, S.; Nissen, P.; Kjaergaard, M. Structural dynamics of P-type ATPase ion pumps. Biochem. Soc. Trans. 2019, 47, 1247–1257. [Google Scholar] [CrossRef]
- Sim, S.I.; Park, E. P5-ATPases: Structure, substrate specificities, and transport mechanisms. Curr. Opin. Struct. Biol. 2023, 79, 102531. [Google Scholar] [CrossRef]
- Aguilar-Martínez, J.A.; Sinha, N. Analysis of the role of Arabidopsis class I TCP genes AtTCP7, AtTCP8, AtTCP22, and AtTCP23 in leaf development. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Ferrero, L. V.; Gastaldi, V.; Ariel, F.D.; Viola, I.L.; Gonzalez, D.H. Class I TCP proteins TCP14 and TCP15 are required for elongation and gene expression responses to auxin. Plant Mol. Biol. 2021, 105, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: a motif found in proteins regulating plant growth and development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Alem, A.L.; Ariel, F.D.; Cho, Y.; Hong, J.C.; Gonzalez, D.H.; Viola, I.L. TCP15 interacts with GOLDEN2-LIKE1 to control cotyledon opening in Arabidopsis. Plant J. 2022, 110, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.M.; Lynch, T.J.; Mobin, R.; Finkelstein, R.R. Direct targets of the transcription factors ABA-Insensitive(ABI)4 and ABI5 reveal synergistic action by ABI4 and several bZIP ABA response factors. Plant Mol. Biol. 2011, 75, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Chen, Y.; Wang, C.; Kong, Y.; Wu, W.; Chen, Y. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J. 2014, 80, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhong, M.; Qu, L.; Yang, J.; Liu, X.; Zhao, Q.; Liu, X.; Zhao, X. AtMYB32 regulates the ABA response by targeting ABI3, ABI4 and ABI5 and the drought response by targeting CBF4 in Arabidopsis. Plant Sci. 2021, 310, 110983. [Google Scholar] [CrossRef] [PubMed]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-insensitive 5) in plant development, abiotic stress responses and phytohormone crosstalk. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Han, X.; Yang, M.; Zhang, M.; Pan, J.; Yu, D. The transcription factor INDUCER OF CBF EXPRESSION1 interacts with ABSCISIC ACID INSENSITIVE5 and DELLA proteins to fine-tune abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2019, 31, 1520–1538. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, K.; Zhou, H.; Yan, X.; Zhan, Q.; Zheng, Y.; Song, C. ABI5 modulates seed germination via feedback regulation of the expression of the PYR/PYL/RCAR ABA receptor genes. New Phytol. 2020, 228, 596–608. [Google Scholar] [CrossRef]
- Wei, J.; Li, X.; Song, P.; Wang, Y.; Ma, J. Studies on the interactions of AFPs and bZIP transcription factor ABI5. Biochem. Biophys. Res. Commun. 2022, 590, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, P.; Li, Y.; Wang, S.; Tang, S.; Liu, C.; et al. ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J. 2016, 85, 348–361. [Google Scholar] [CrossRef]
- Maymon, T.; Eisner, N.; Bar-Zvi, D. The ABCISIC ACID INSENSITIVE (ABI) 4 transcription factor is stabilized by stress, ABA and phosphorylation. Plants 2022, 11, 2179. [Google Scholar] [CrossRef]
- Gregorio, J.; Hernández-Bernal, A.F.; Cordoba, E.; León, P. Characterization of evolutionarily conserved motifs involved in activity and regulation of the ABA-INSENSITIVE (ABI) 4 transcription factor. Mol. Plant 2014, 7, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhang, H.; Wang, S.; Chen, M.; Wu, Y.; Tang, S.; Liu, C.; Feng, Y.; Cao, X.; Xie, Q. ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Wang, F.; Zheng, Q.; Niza, V.M.A.G.E.; Downie, A.B.; Perry, S.E. Direct and indirect targets of the arabidopsis seed transcription factor ABSCISIC ACID INSENSITIVE3. Plant J. 2020, 103, 1679–1694. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, X.; Wang, K.; Li, Z.; Jia, Q.; Zhao, C.; Zhang, M. ABA-INSENSITIVE 3 with or without FUSCA3 highly up-regulates lipid droplet proteins and activates oil accumulation. J. Exp. Bot. 2022, 73, 2077–2092. [Google Scholar] [CrossRef]
- Parcy, F.; Valon, C.; Kohara, A.; Misera, S.; Giraudat, J. The ABSCISIC ACID-INSENSITIVE3, FUSCA3, AND LEAFY COTYLEDON1 loci act in concert to control multiple aspects of Arabidopsis seed development. Plant Cell 1997, 9, 1265–1277. [Google Scholar]
- Dietz, K.J. Peroxiredoxins in plants and cyanobacteria. Antioxidants Redox Signal. 2011. [CrossRef] [PubMed]
- Chen, H.; Chu, P.; Zhou, Y.; Ding, Y.; Li, Y.; Liu, J.; Jiang, L.; Huang, S. Ectopic expression of NnPER1, a Nelumbo nucifera 1-cysteine peroxiredoxin antioxidant, enhances seed longevity and stress tolerance in Arabidopsis. Plant J. 2016, 88, 608–619. [Google Scholar] [CrossRef]
- Chen, H.; Ruan, J.; Chu, P.; Fu, W.; Liang, Z.; Li, Y.; Tong, J.; Xiao, L.; Liu, J.; Li, C.; et al. AtPER1 enhances primary seed dormancy and reduces seed germination by suppressing the ABA catabolism and GA biosynthesis in Arabidopsis seeds. Plant J. 2020, 101, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Haslekås, C.; Viken, M.K.; Grini, P.E.; Nygaard, V.; Nordgard, S.H.; Meza, T.J.; Aalen, R.B. Seed 1-cysteine peroxiredoxin antioxidants are not involved in dormancy, but contribute to inhibition of germination during stress. Plant Physiol. 2003, 133, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Abdul Aziz, M.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant group II LEA proteins: intrinsically disordered structure for multiple functions in response to environmental stresses. Biomolecules 2021, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Qi, S.; Li, H.; Liu, P.; Li, P.; Wu, C.; Zheng, C.; Huang, J. Overexpression of Late Embryogenesis Abundant 14 enhances Arabidopsis salt stress tolerance. Biochem. Biophys. Res. Commun. 2014, 454, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Ecker, J.R. Diversity and dynamics of dna methylation: Epigenomic resources and tools for crop breeding. Breed. Sci. 2019, 69, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Plitta-Michalak, B.P.; Suszka, J.; Naskręt-Barciszewska, M.Z.; Kotlarski, S.; Barciszewski, J.; Chmielarz, P. Identification of DNA methylation changes in European beech seeds during desiccation and storage. Int. J. Mol. Sci. 2023, 24, 3557. [Google Scholar] [CrossRef] [PubMed]
- An, Y.Q.C.; Goettel, W.; Han, Q.; Bartels, A.; Liu, Z.; Xiao, W. Dynamic changes of genome-wide DNA methylation during soybean seed development. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-Y.; Le, B.H.; Chen, M.; Henry, K.F.; Hur, J.; Hsieh, T.-F.; Chen, P.-Y.; Pelletier, J.M.; Pellegrini, M.; Fischer, R.L.; et al. Similarity between soybean and Arabidopsis seed methylomes and loss of non-CG methylation does not affect seed development. Proc. Natl. Acad. Sci. 2017, 114, E9730–E9739. [Google Scholar] [CrossRef]
- Weng, J.-K.; Ye, M.; Li, B.; Noel, J.P. Co-evolution of hormone metabolism and signaling networks expands plant adaptive plasticity. Cell 2016, 166, 881–893. [Google Scholar] [CrossRef]
Figure 1.
Image of P. sativum mature embryo. Embryo includes cotyledons (Cot), first true leaves (Lf), epicotyl (Ep), hypocotyl (Hy), and root (R). Embryonic axe includes Lf, Ep, Hy and R.
Figure 1.
Image of P. sativum mature embryo. Embryo includes cotyledons (Cot), first true leaves (Lf), epicotyl (Ep), hypocotyl (Hy), and root (R). Embryonic axe includes Lf, Ep, Hy and R.
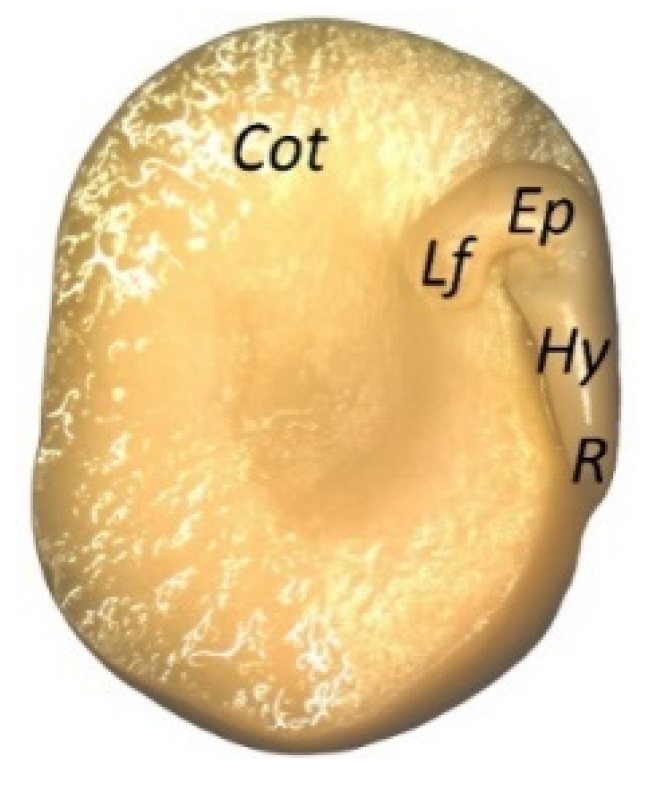
Figure 2.
The contents of abscisic acid (ABA), phaseic acid (PA), dihydrophaseic acid (DPA), and neo-phaseic acid (neoPA) observed in embryonic axes of P. sativum before and after radicle protrusion (RP). The data represent the mean ± standard error of nine biological replicates. The statistical analysis relied on two-tailed t-test with a critical alpha value of 0.05. Significant differences between the mean values are indicated (*** p ≤ 0.001, ** p ≤ 0.005).
Figure 2.
The contents of abscisic acid (ABA), phaseic acid (PA), dihydrophaseic acid (DPA), and neo-phaseic acid (neoPA) observed in embryonic axes of P. sativum before and after radicle protrusion (RP). The data represent the mean ± standard error of nine biological replicates. The statistical analysis relied on two-tailed t-test with a critical alpha value of 0.05. Significant differences between the mean values are indicated (*** p ≤ 0.001, ** p ≤ 0.005).
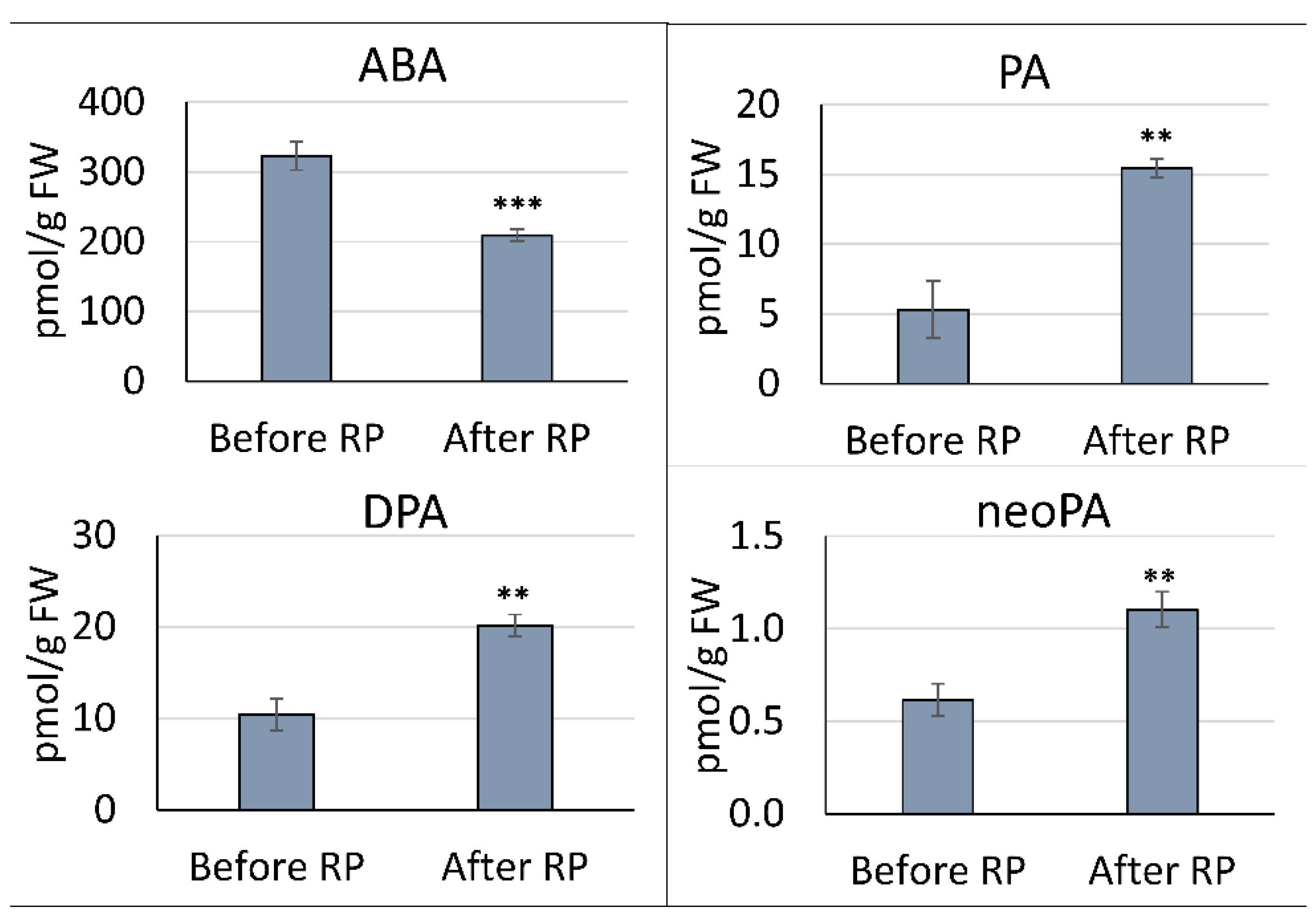
Figure 3.
Volcano plot representing 70 differentially expressed genes (DEGs). The X-axis indicates the log2-transformed gene expression fold changes between seed axes before and after radicle protrusion. The Y-axis indicates the log10-transformed p-value. Significant DEGs with lower expression are highlighted in blue (№ 1-24). Significant DEGs with higher expression are highlighted in red (№ 25-70). See Table S1 for the full description of the down-regulated and up-regulated genes.
Figure 3.
Volcano plot representing 70 differentially expressed genes (DEGs). The X-axis indicates the log2-transformed gene expression fold changes between seed axes before and after radicle protrusion. The Y-axis indicates the log10-transformed p-value. Significant DEGs with lower expression are highlighted in blue (№ 1-24). Significant DEGs with higher expression are highlighted in red (№ 25-70). See Table S1 for the full description of the down-regulated and up-regulated genes.
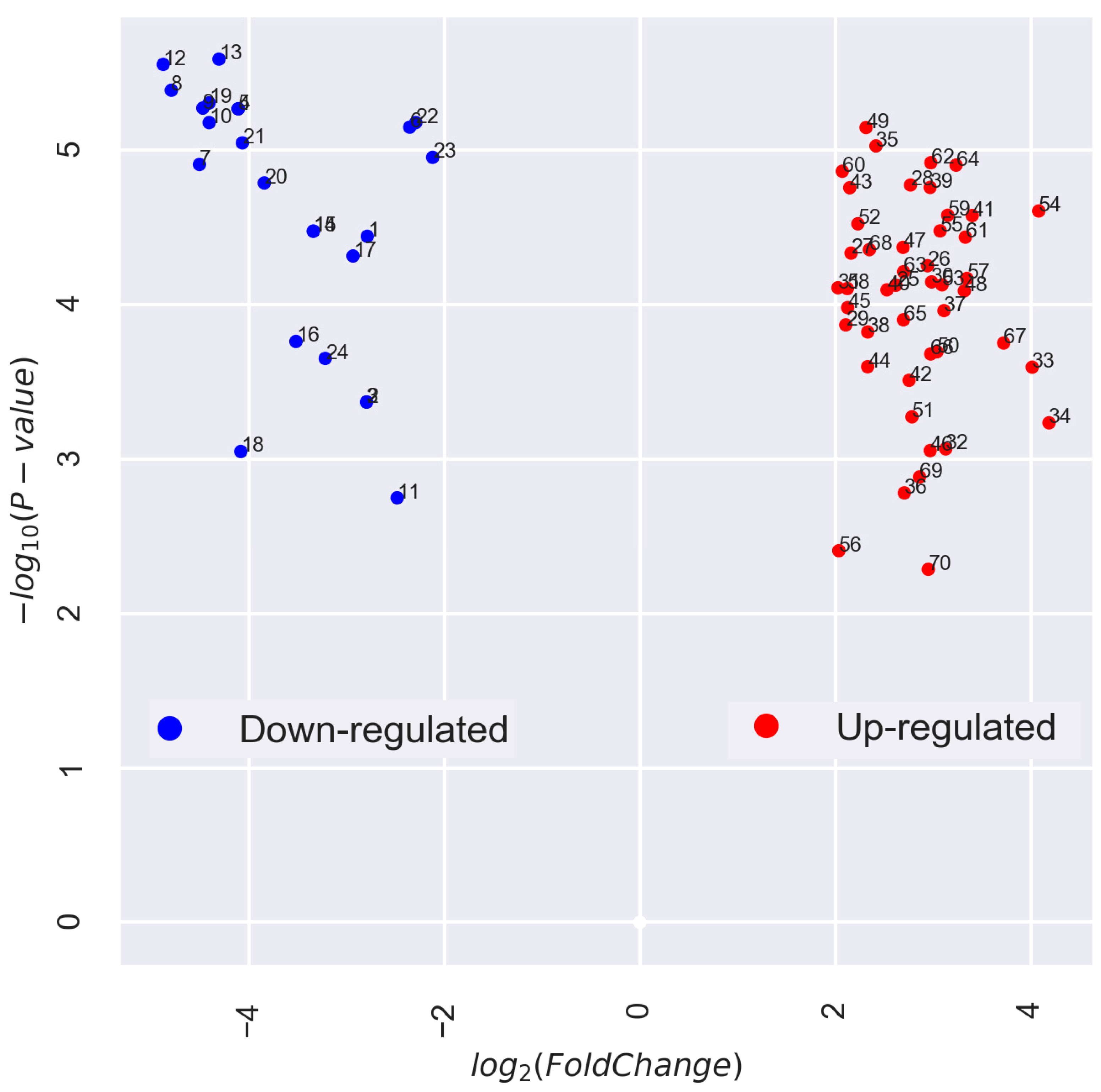
Figure 4.
Methylation of the ABA-related gene promoters in embryonic axes of germinated P. sativum seeds before and after radicle protrusion (RP). (a) The PsABI3 gene promoter before RP. (b) The PsABI3 gene promoter after RP. (c) The PsABI4 gene promoter before RP. (d) The PsABI4 gene promoter after RP. (e) The PsABI5 gene promoter before RP. (f) The PsABI5 gene promoter after RP. The length of the analyzed segment PsABI3 promoter is 1057 bp with the number of cytosines is 160, for the PsABI4 promoter is 721 bp with 181 cytosines, and for the PsABI5 promoter is 1231 bp with 142 cytosines. Circles represent cytosines, with methylated bases shown in black and unmethylated bases in white. See Figure S3, S4 and S5 for mapping of the PsABI3, PsABI4 and PsABI5, accordingly.
Figure 4.
Methylation of the ABA-related gene promoters in embryonic axes of germinated P. sativum seeds before and after radicle protrusion (RP). (a) The PsABI3 gene promoter before RP. (b) The PsABI3 gene promoter after RP. (c) The PsABI4 gene promoter before RP. (d) The PsABI4 gene promoter after RP. (e) The PsABI5 gene promoter before RP. (f) The PsABI5 gene promoter after RP. The length of the analyzed segment PsABI3 promoter is 1057 bp with the number of cytosines is 160, for the PsABI4 promoter is 721 bp with 181 cytosines, and for the PsABI5 promoter is 1231 bp with 142 cytosines. Circles represent cytosines, with methylated bases shown in black and unmethylated bases in white. See Figure S3, S4 and S5 for mapping of the PsABI3, PsABI4 and PsABI5, accordingly.

Figure 5.
Oxidative pathways of ABA catabolism: CYP707s – – cytochrome P450 monooxygenases; PA – phaseic acid; neoPA – neophaseic acid; DPA – dihydrophaseic acid; neoDPA – epi-neodihydrophaseic acid; PAR – PA reductase.
Figure 5.
Oxidative pathways of ABA catabolism: CYP707s – – cytochrome P450 monooxygenases; PA – phaseic acid; neoPA – neophaseic acid; DPA – dihydrophaseic acid; neoDPA – epi-neodihydrophaseic acid; PAR – PA reductase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



