
Submitted:
12 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Although hydrogen gas (H2) treated soil improves crop biomass, this approach appears difficult for the field application due to the flammability of H2 gas. In this report, we investigated whether and how H2 applied in hydrogen nanobubble water (HNW) improves the yield and quality of cherry tomato (Lycopersicon esculentum var. cerasiforme) with/without fertilizers. Two-year field trials showed that compared to corresponding controls, HNW without/with fertilizers improved cherry tomato yield per plant by 39.7% and 26.5% in 2021 (Shanghai), and 39.4% and 28.2% in 2023 (Nanjing). Compared to surface water (SW), HNW increased soil available nitrogen (N), phosphorus (P), and potassium (K) consumption regardless of fertilizer application, which may be attributed to the increased NPK transport related genes in roots (LeAMT2, LePT2, LePT5, and SlHKT1,1). Also, HNW-irrigated cherry tomato displayed higher sugar-acid ratio (8.6%) and lycopene content (22.3%) than SW-irrigated plants without fertilizers. Importantly, the beneficial effects of HNW without fertilizers on yield per plant (9.1%), sugar-acid ratio (31.1%), volatiles (20.0%) and lycopene contents (54.3%) were stronger than those achieved by fertilizers alone. In short, this study clearly indicated that HNW-supplied H2 not only exhibited fertilization effect on enhancing tomato yield, but also improved fruit quality with a lower carbon footprint.
Keywords:
cherry tomato
; fertilizer
; hydrogen
; hydrogen nanobubble water
; quality
; yield
1. Introduction
Cherry tomato (Lycopersicon esculentum var. cerasiforme), a small fruited variety of tomato, is a popular and widely cultivated fruit vegetable in the world [1]. Since cherry tomato is rich in nutrients such as lycopene, vitamins, and minerals, it is favored by consumers for reducing risk of various diseases, such as cardiovascular disorders, hypercholesterolemic and hyperglycemic attributes, and cancer [2]. Although fertilizers can improve fruit yield, over-fertilization not only causes water pollution, but also results in flavor loss [3] and fruit nitrate and nitrite accumulation [4]. In addition, the large-scale application of fertilizers for crop production increases greenhouse gas emissions and accelerates global warming [5]. Therefore, it is a modern challenge to improve tomato yield and quality in a more fertilizer efficient and environmentally friendly way.
In the last decade, molecular hydrogen (H2) has been considered as a promising medical treatment for metabolic, digestive, respiratory, cardiovascular diseases, neurodegenerative disorders, and cancer [6]. In addition, H2 exhibits a variety of biological functions in plants, including alleviating oxidative damage caused by various abiotic stresses [7], promoting seed germination and root development [8], and improving the postharvest preservation of vegetables [9], fruits [10], and flowers [11]. It has been previously found that H2-exposed soils can promote the biomass of soybean, spring wheat, barley, and canola, suggesting that H2 has an effect of fertilizer utilization in soils [12,13]. However, H2 applied in a gaseous form for soil treatment is complicated and impractical in the field due to its low residency and flammable properties at higher concentrations.
Although the application of hydrogen-rich water (HRW) has been found to improve yield and prolong the shelf life of daylily buds [14], it has the disadvantages of the low solubility and short residence time of dissolved H2. Solid H2 storage materials, such as magnesium hydride (MgH2) [15], ammonia borane (AB) [16], and AB-loaded hollow mesoporous silica nanoparticle (AB@hMSN) [17] can improve the effective H2 residency of conventional HRW, and thus have positive effects on flower senescence, stress responses, and plant growth regulation. Nevertheless, the potential environmental risk of their by-products should be concerned, especially when they are extensively used in the field.
The nanobubble technology establishes a useful approach to accelerate gas dissolution and remain its stability in the liquids for longer times [18]. Hydrogen nanobubble water (HNW) has been reported to reduce the toxicity of copper to Daphnia magna by alleviating oxidative stress and inhibiting copper accumulation [19]. Besides, HNW can promote seed germination and concentrations of bioactive phytochemicals in sprouted black barley [20]. A solution of HNW was also shown to extend the vase life of cut carnation flowers [11]. The previous field trial showed that HNW increased size and quality of rice grain [21], and enhanced strawberry aroma [22].
In this study, two-year and multi-site trials were carried out to investigate whether and how preharvest HNW treatment improved cherry tomato yield and quality (including sugars, vitamin C, lycopene, phenols, and flavonoids contents), in the absence (especially) or presence of fertilizers. The changes in available nitrogen (N), phosphorus (P), and potassium (K) in soil and transcriptional profiles of genes associated with tomato nutrition absorption and quality were further investigated. The results thus provide a reference for the practical application of HNW in horticulture for better performance in both yield and quality, which might open a new window for the low carbon society.
2. Results
2.1. Preharvest Application of HNW Improves Cherry Tomato Yield
As shown in Figure 1A, the irrigation with HNW promoted the growth of tomato plants in Shanghai (2021). Similar with compound fertilizers, HNW increased yield per plant of cherry tomato (Figure 1B). Compared to SW irrigation, yield per plant of cherry tomatoes in the treatment of HNW without fertilizers was increased by 39.7% (P < 0.01). Meanwhile, HNW plus fertilizers treatment showed the obvious effect on cherry tomato yield, and increased yield per plant by 26.5% in comparison with SW plus fertilizers (P < 0.01). Especially, yield per plant in the treatment of HNW without fertilizers was even higher (9.1%, P < 0.05) than that in SW plus fertilizers treatment.
To further assess the reliability of the yield enhancement achieved by hydrogen-based irrigation in cherry tomato, we conducted another field trial test in Nanjing (2023). Consistently, the increase of yield per plant exhibited a similar trend as observed in the previous results (Figure 1). As shown in Figure 2A, under the conditions with/without fertilizers, HNW (0.50 ± 0.04 kg plant-1/0.46 ± 0.03 kg plant-1) remarkably resulted in the increase of the yield per plant by 28.2% (P < 0.01) and 39.4% (P < 0.01) compared with SW (0.39 ± 0.04 kg plant-1/0.33 ± 0.05 kg plant-1) in this year. In addition, the number of fruits per plant in HNW treatment was also higher than SW treatment regardless of fertilizers (Figure 2B). For example, compared with the SW alone and SW plus fertilizers group, HNW remarkably increased the number of fruits per plant by 39.10% (P < 0.01) and 27.91% (P < 0.01), respectively. Although HNW did not obviously influence the single fruits weight (Figure 2C), the increased numbers of fruits per plants might ultimately increase cherry tomato yield (Figure 2D) by 43.3% and 28.1%, in comparison with SW in the absence or presence of fertilizers. Comparatively, we clearly observed that the yield was higher in HNW-irrigated and no fertilizers addition group than SW-irrigated and fertilizers-added group (22.1%, P < 0.05), which could be partially explained by the obvious increases in yield per plant and number of fruits per plant, compared to fertilizers group (P < 0.05).
2.2. Effects of HNW on the Balance of Sugars and Acids in Cherry Tomatoes
The main factors affecting the flavor of cherry tomato are the content and ratio of sugars and acids, which are critical to its commercial value [23]. In the absence of fertilizers, HNW treatment significantly increased the soluble sugar content (13.9%, P < 0.01; Figure 3A), and slightly increased the titratable acid content (4.7%; Figure 3B), thus causing sugar-acid ratio increase (8.6%, P < 0.05; Figure 3C). Comparatively, fertilizers addition alone had no such effect on the soluble sugar content, but increased the titratable acid content, resulting in decreasing the sugar-acid ratio (-17.1%, P < 0.05; Figure 3C).
When HNW was applied without fertilizers, it clearly also showed the most obvious effect on increasing sugar-acid ratio, especially increased by 31.1% in comparison with fertilizers alone (P < 0.01). Meanwhile, HNW reversed the negative of fertilizers on the sugar-acid ratio when it was applied in combination with fertilizers.
Fructose, glucose, and sucrose are the three main soluble sugars in cherry tomatoes [24]. Compared with SW alone, HNW addition significantly increased fructose content (3.1%, P < 0.05), but not glucose and sucrose (Figure 3D–F). Fertilizer application decreased the sucrose content (-16.4%, P < 0.05), while it did not influence the other two sugars. Therefore, the treatment of HNW without fertilizers had better effects on fructose and sucrose contents compared with fertilizers alone (P < 0.05). In the presence of fertilizers, HNW only slightly increased fructose content. The observations above indicate that HNW might regulate the balance of sugars and acids in cherry tomato fruits.
2.3. Antioxidant Compounds Accumulation in Response to HNW
Cherry tomatoes contain numerous antioxidant ingredients such as ascorbic acid (vitamin C), phenols, and flavonoids, thus having antioxidative, anti-inflammatory, and anti-microbial effects [25]. Among them, lycopene has an outstanding contribution [26]. Further results showed that the application of HNW without fertilizers differentially increased the accumulation of lycopene (22.3%, P < 0.01; Figure 4A) and total phenols (8.1%; Figure 4B), except for vitamin C (Figure 4C) and flavonoids (Figure 4D). By contrast, under our experiment conditions, the application with fertilizers significantly decreased lycopene content, and obviously increased total phenols and flavonoids contents. Accordingly, in the absence of fertilizers, HNW significantly increased lycopene content (54.3%, P < 0.01), but decreased total flavonoid content (P < 0.01), in comparison with fertilizer alone. Meanwhile, when HNW was combined with fertilizers, only flavonoid content was significantly increased.
Phytoene synthase (PSY) and phytoene desaturase (PDS) are the key enzymes in determining the biosynthesis of lycopene [27]. The changes in expression levels of SlPSY1 and SlPDS displayed approximately similar trends compared to lycopene contents after the application of HNW with/without fertilizers (Figure 4E,F).
2.4. Modulation of the Aromatic Profiles in Cherry Tomatoes Achieved by HNW
In this experiment, a total of 49 major volatile compounds were identified, including aldehydes, alcohols, esters, phenols, etc. Among these, the proportion of aldehydes and alcohols were 77.1% and 12.3%, respectively (Supplementary Table S1). With or without fertilizers, HNW irrigation increased the contents of total volatile compounds by 8.0% and 4.4%, respectively (Figure 5A). The application of fertilizer alone reduced the content of total volatile compounds (-14.9%), compared with the SW treatment. Accordingly, the treatment of HNW without fertilizers could neutralize the negative effect of fertilizers alone on total volatile compounds (20.0%, P < 0.05).
Similarly, HNW irrigation increased the contents of aldehydes (by 15.4% and 10.8%; Figure 5B), including hexanal (by 150.2% and 41.5%, P < 0.05; Figure 5D) and (E)-2-hexanal (3.0%; Figure 5E), and trans-1,2-cyclopentanediol (26.1%; Figure 5F), respectively, with (except (E)-2-hexanal and trans-1,2-cyclopentanediol) or without fertilizers. The treatment of HNW without fertilizers showed greater effect than that of fertilizers alone on aldehydes (P < 0.01), including hexanal (P < 0.05) and (E)-2-hexanal (P < 0.05), and trans-1,2-cyclopentanediol (P < 0.05), respectively. However, the application of fertilizer alone had a negative effect on the above compounds other than 2,4-bis(1,1-dimethylethyl)-phenol (increased by 50.3%; Figure 5G), compared with the SW treatment. Meanwhile, no significant alteration was observed in the changes of alcohols (Figure 5C), and 2,4-bis(1,1-dimethylethyl)-phenol. We also noticed that in the presence of fertilizers, HNW addition could significantly increase hexanal level, but decrease contents of alcohols (P < 0.05).
2.5. The Absorption of Soil Elements was Influenced by HNW
Before planting, contents of soil available nitrogen (N), phosphorus (P), and potassium (K) in different treatments were basically at the same level (Figure S1). Subsequent results showed that without fertilizers, HNW treatment enhanced the decrease of soil available N, P, and K contents (especially N and P, P < 0.01) in comparison with surface water, and above-mentioned effects in soil available NP consumption achieved by HNW were pronounced than those treated with fertilizers alone (P < 0.05; Figure 6A–C). Meanwhile, in the presence of fertilizers, HNW irrigation exhibited similar effects on the reduction in available N (P < 0.01), P (P < 0.05), and K (P < 0.05) contents.
The expression levels of genes involved in plant N, P, and K accumulation in plants were further investigated (Figure 6D–G). They included a NH4+ transporter gene (LeAMT2), two phosphate transporters genes (LePT2 and LePT5), and a potassium transporter (SlHKT1,1) [28,29]. As expected, the changes in transcript levels of the four genes above were consistent with the reduction in soil available NPK contents, respectively. These results indicated that preharvest HNW application might positively improve NPK absorption in cherry tomatoes.
2.6. Principal Component Analysis
Principal component analysis (PCA) was performed to distinguish above mentioned four treatments. The first two components explained 84.4% of total variance (Figure 7A). Meanwhile, the four treatments were clearly separated; SW and HNW+F treatments were distinguished on PC 1, while PC 2 discriminated SW+F and HNW treatments, indicating that there were distinct differences. As shown in the biplot (Figure 7B), the quality characters (including sugars, volatiles, titratable acid, total phenols, flavonoids, vitamin C, and lycopene contents, and lycopene biosynthesis-related gene expression) were positively correlated with PC 1, whereas yield per plant, soil NPK reduction, and the related gene expression (including LeAMT2, SlHKT1,1, LePT2, and LePT5), and hexanal were negatively correlated. Moreover, there were further positive correlations among the quality characters. Similarly, results of yield showed positive correlations with NP absorption and expression of LeAMT2, SlHKT1,1, and LePT2. However, yield was negatively correlated with soluble sugars, sugar-acid ratio, volatiles (including alcohols, aldehydes, (E)-2-hexanal, and hexanal), and lycopene contents.
3. Discussion
As an environmentally benign gas, H2 plays a major role in promoting plant growth, improving crop yield and nutritional quality [30]. Both H2 gas-treated soil and conventional HRW irrigation have been proposed and observed to improve yield of crops [12,14]. However, H2 applied in gas form is not practical in the field (e.g., flammability), and H2 is effective in HRW for less than 6 h [11]. Under our trial conditions, the residence time of H2 as HNW was about 12 h, which was consistent with the previous study [19], and twice as long as that in HRW (Figure S2).
Although the mechanism of H2 fertilization using its gas in enhancing plant yield has yet to be fully understood, it was most probably attributed to the enhanced growth of H2-oxidizing bacteria in the soil. These microorganisms may improve the nutrient status of soil, enhance the plants growth regulator balance or disease resistance [12,31]. It was previously reported that H2 exposure can increase soil carbon deposition [32] and synthesis of soil enzymes such as catalase, dehydrogenase, and urease [33]. These results reflected the possibility that H2 improves soil fertility by inducing the metabolic activities of beneficial bacteria. Therefore, we chose the four greenhouses closely (Figure S3) and with the same crop rotation (tomato) to avoid the difference in climatic, illumination, and microorganism. Importantly, we tested the nutrition of soil samples, and the results (Figure S1) showed that the initial soil conditions of the four greenhouses were similar in terms of the key nutrition, including available nitrogen, phosphorus, and potassium.
Two-year field trials clearly showed that HNW-improved yield of greenhouse cherry tomatoes was more pronounced than that when cultured with fertilizers (Figure 1 and Figure 2). Moreover, an additional effect on cherry tomato yield was observed in the presence of HNW plus fertilizers.
NPK are the principal nutrients typically supplied plants, so that the absorption and utilization efficiency of these elements control crop yield [34]. It has been reported that HNW increased the transcription of the genes related to the absorption of NPK in rice, including NRT2.3, NiR, ARE1, NLP4, and AKT1 transcripts [21]. In this study, it was clearly observed that NPK transport related genes in plants (especially LeAMT2, LePT2, and SlHKT1,1) were positively correlated with soil NPK reduction and the yield of tomato fruits (Figure 7B).
It has been reported that the combined application of microbial consortium and fertilizer increased soil available NPK content, and promoted NPK absorption by sugarcane plants, thereby promoting plant growth and increasing sugarcane and sugar yields [35]. The previous study showed that high level of H2-oxidizing bacteria in H2-treated soil increased plant biomass and promoted plant growth [31]. Since HNW in this study was irrigated at the early growth stage of cherry tomatoes, we deduced that the beneficial roles of HNW in improving soil NPK absorption and cherry tomato yield may be partially associated with H2 impacting on soil microbes. This hypothesis is required further investigation on the interaction of plants and microbes responded to H2. Since in our experimental conditions, HNW may differentially increase the expression levels of above four genes and soil available NPK consumption (Figure 6), we further proposed that these changes may be also partially responsible for the promotion of cherry tomato yield in the absence/presence of fertilizers (Figure 1 and Figure 2).
Sugars, acids, and their ratio are major contributors to fruit taste [23,36]. High sugar-acid ratio enhances the desirable sweet perception. A previous study showed that HNW increased sugar-acid ratio as a result of the increased sugar content and decreased titratable acid content in strawberry [22]. In this study, without fertilizer addition, HNW increased sugar-acid ratio by increasing total soluble sugar content (especially fructose; Figure 3A,D). The previous study pointed out that nitrogen fertilization affected the activities of enzymes directly related to acid metabolism in fruit, thereby changing the acid content [37]. Consistently, the application of fertilizers increased titratable acid content, thus decreasing sugar-acid ratio (Figure 3B,C). However, HNW reversed above negative effect of fertilizers by regulating the balance of sugar and acid (Figure 3A–C), consistent with a previous study in strawberry [22].
Lycopene, vitamin C, total phenols and flavonoids are important antioxidants in fruits and vegetables [25]. A previous study observed that HRW enhanced tolerance against UV-B stress, associated with the improvement of flavonoids profiles in alfalfa seedlings [38]. In addition, HRW can also alleviate oxidative damage by increasing contents of vitamin C, total phenols, and flavonoids, resulting in prolonged shelf life of tomatoes [39], daylily buds [14], and lychee [40]. Consistently, our results showed that HNW alone increased lycopene content in tomato fruits (Figure 4A). This is a new finding. HNW control of lycopene increase was further supported by the up-regulation of SlPSY1 and SlPDS transcripts (Figure 4E,F), two lycopene synthesis genes [41], and results of the PCA (Figure 7B). Therefore, it was suggested that two genes above might be the target genes responsible for HNW-triggered lycopene accumulation.
Although having identified more than 400 volatile compounds in tomato fruit, current studies showed that the most important compounds aldehydes, such as hexanal, and (E)-2-hexenal, alcohols such as trans-1,2-cyclopentanediol, and phenols such as 2,4-bis (1,1-dimethylethyl)-phenol play key roles in tomato aroma [42]. Aldehydes are the most dominant, by giving off the ‘fresh green’ odor [43]. A recent study showed that the contents of total volatile compounds, and aldehydes such as hexanal and (E)-2-hexenal, were increased by preharvest HNW application in strawberries [22]. In this report, we discovered that HNW also increased hexanal content in cherry tomato with/without fertilizers (Figure 5D), reflecting the possible common mechanism.
Furthermore, a positive correlation between soluble sugars and volatile compounds contents in cherry tomatoes (Figure 7B) was consistent with the previous studies in strawberries [22,44]. Since the important volatile compounds such as esters, furanones, and terpenes, are present in the form of glycosides in cells, and the precursors of those were sugars [45,46], aromatic volatiles in tomato fruits may be really positively associated with sugars. In addition, sugars and volatile compounds are known to be the important factors influencing sweetness perception [47,48]. Consumer liking was associated with sweetness and aroma intensity [44]. Therefore, HNW-increased sugars and volatile compounds contents in cherry tomatoes should be more attractive for consumer.
Under the condition of limited fruit carbohydrates, plants preferentially utilize the carbohydrates transported into the fruits to form carbon skeletons, which may lead to lower fruit quality but higher yield [49]. Consistently, in this study, a negative correlation was observed between yield and quality characters, including soluble sugars, volatile compounds, and lycopene contents, and etc. (Figure 7B). As expected, compound fertilizers promoted yield of cherry tomatoes, but undesirably reduced fruit sugars, lycopene, and volatile contents [50].
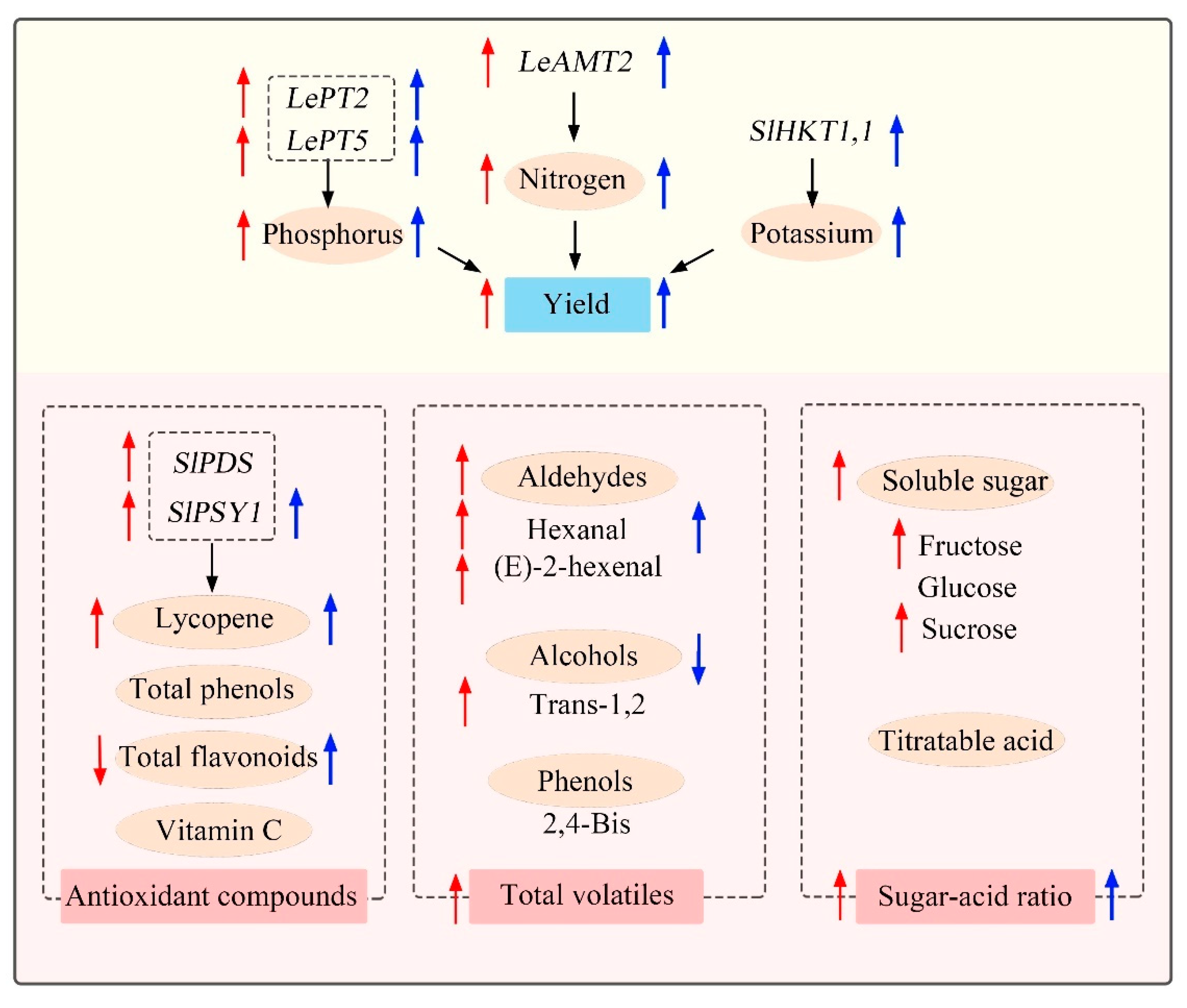
Together, as shown in the schematic model summarized the effects of yield and quality in response to HNW (Figure 8), it is worth noting that HNW may not only improve cherry tomato yield, which was better than grown with fertilizers alone to some extent, but also partly reverse the negative fertilizer effects on sugar-acid ratio, volatile compounds, and lycopene contents. The above effects of HNW may be attributable to the regulation of plant NPK absorption, carbohydrate and secondary metabolism.
4. Materials and Methods
4.1. Plant Materials and Experimental Design
The field experiment was carried out at the Qingpu Agriculture Base, Shanghai, China (longitude 121° 01' E and latitude 31° 02' N; 2021) and White Horse Agriculture Base, Lishui District, Nanjing, Jiangsu, China (longitude 119° 17' E and latitude 31° 58' N; 2023). The cherry tomato ‘Jintong’ (Lycopersicon esculentum var. cerasiforme ‘Jintong’) was planted on January 21, 2021 and Mar 7, 2023, respectively. Considering the fugitiveness of H2, four greenhouses were used for the experiment, and three plots (each 20 m long and 3 m wide) were randomly selected as replicates for each treatment in per greenhouse (80 m long and 6 m wide). Before fertilizing, contents of soil available nitrogen (N), phosphorus (P), and potassium (K) in four greenhouses were basically at the same level (Shanghai, 2021): available nitrogen content 215.8 ± 4.5 mg kg−1; available phosphorus content 410.7 ± 3.7 mg kg−1; available potassium content 406.4 ± 3.8 mg kg−1 (Figure S1). Also, the number of cherry tomato plants in one treatment per each greenhouse was 1288.
Four treatments were arranged, including (1) Irrigation with surface water (SW) and free of fertilization; (2) Irrigation with SW and normal fertilization; (3) Irrigation with HNW and free of fertilization; (4) Irrigation with HNW and normal fertilization.
Fertilizers were applied conventionally in the required treatments. A compound fertilizer (Nitrophoska® 15-15-15) was used as the base fertilizer on December 10, 2020 (Shanghai) and January 7, 2023 (Nanjing) without topdressing. The amount of fertilizers applied was 50 kg per greenhouse. Also, no pesticides were applied during the plant growth stages. Flood irrigation with HNW was carried out once on the day after planting at 10 t h-1 flow rate for 0.5 h per greenhouse for HNW treatment. Meanwhile, irrigation with the same amount of SW was set as the controls. Subsequent field operations were in accordance with the conventional agricultural managements.
4.2. Preparation of HNW
In this experiment, HNW (with ~ 300 nm nanobubbles) was prepared with a H2 nanobubbles generator (Air Liquide (China) R&D Co., Ltd., Shanghai, China). The dissolved H2 concentration was measured using the ENH-2000 portable dissolved H2 meter (TRUSTLEX, Osaka, Japan) that was calibrated with gas chromatography. The fresh HNW contained 1.0 mg L-1 H2, and it remained in HNW for ~ 12 h (Figure S2).
4.3. Determination of Cherry Tomato Yield
The cherry tomato fruits were harvested once a week from April 16 to June 15, 2021 and from June 1 to July 25, 2023, respectively when their color changed from green to yellow. Afterwards, the yield per plant was calculated.
The freshly picked tomatoes were placed in sampling bags during the fruiting stage and transported back to the laboratory in the dark at room temperature. The yield parameters were analyzed in triplicate, and at least 10 plants/treatment/repeat were used (except Figure 2D). For each treatment, ten or twenty fruits from different plants were mixed and ground using a mill (A11, IKA, Germany), and stored at -80 °C. Pooled samples were split into three replicates for the biochemical and molecular analysis.
4.4. Evaluation of Fructose, Glucose, Sucrose, Titratable Acid, and Soluble Sugar Contents
Following the previous method [51], twenty fruits from four greenhouses with different treatments were ground using a mill (A11, IKA, Germany), and stored at -80 °C, pooled samples were split into three replicates for the following analysis. Fructose and glucose contents were detected by a High Performance Liquid Chromatograph (HPLC; Infinity 1260; Agilent, Santa Clara, CA) with a 250 mm × 4.6 mm ZORBAX column (Agilent, USA) at 40 °C. Elution was used with 75% acetonitrile (v/v), with 20 μL injection and 1.3 mL min-1 for the flow rate. The amount of sucrose was assayed according to the Lane-Eynon method [52]. The test solution was titrated with Fehling’s solution, containing a methylene blue indicator.
The measurement of titratable acid was achieved by the sample being titrated with 0.1 M NaOH to an end-point pH of 8.2 [53]. The results were expressed as g kg-1 fresh weight.
The content of soluble sugars was determined with anthrone-sulfuric acid colorimetric method [54]. Firstly, the samples were mixed with 2% (w/v) anthrone regent/concentrated sulfuric acid (0.5:5, v/v) solution, and then incubated in boiling water for 10 min. After centrifugation, the absorbance of the supernatant was recorded at 620 nm. The soluble sugar content was expressed as g kg-1 fresh weight calculated using the standard curve of sucrose.
4.5. Determination of Lycopene, Vitamin C, Total Phenols, and Flavonoids Contents
Lycopene, vitamin C, total phenols, and flavonoids were averaged from three independent samples prepared by pooling 20 individual fruits from different plants for each treatment. Lycopene was exacted by using mixed solvent extraction method described previously [55]. The fruit samples (1.0 g) added with the solution of acetone/petroleum (5:5, v/v) was incubated in a 30 °C water bath for 15 min. Afterwards, the absorbance was determined at 472 nm. The standard curve was obtained by adding different concentrations of a lycopene standard solution.
Vitamin C content was detected with the HPLC system (D-2000, Hitachi, Ltd., Tokyo, Japan). According to the previous method [56], the sample homogenate was extracted with oxalic acid solution under dark conditions for 10 min. After centrifugation and filtration with a 0.45 μm water filtration membrane, the test solution was prepared for detection. Vitamin C content expressed as g kg-1 fresh weight was calculated from the standard curve of vitamin C.
The content of total phenols was estimated using the Folin-Ciocalteu reagent and the absorbance was measured at 765 nm [57]. The results were obtained from a standard curve for gallic acid, expressed as g kg-1 fresh weight.
The flavonoid content was determined using the aluminum chloride colorimetric method [58,59]. The samples (0.1 g) were dispersed in 1 mL of deionized water, and then mixed with the solution of 95% alcohol, 10% aluminum chloride hexahydrate, 1 M potassium acetate, and deionized water (1.5:0.1:0.1:2.8, v/v/v/v).
4.6. Extraction and Analyses of Aromatic Compounds
Headspace solid phase microextraction was used to sample volatiles of tomatoes according to the previous method [22]. Ten fruits were ground together using a mill (A11, IKA, Germany). These samples were split into three aliquots and all replicates of 5 g each were processed by following methods. The samples were added with 1 g of sodium chloride, containing 10 μL of 2-nonanone (internal standard; dilute 5 × 103 times; Macklin, China) in a 20 mL vial. Volatiles were collected using a 1cm DVB/CAR/PDMS Stable Flex fiber (50/30μm; Supelco, PA, USA) in a 50 °C water bath for 30 min after a 50 °C equilibration for 15 minutes. Afterwards, the analysis was performed on a 320-MS gas chromatography-mass spectrometer (GC-MS, Bruker, Germany) with a BR-5ms column (30 m × 0.25 mm ID × 0.25 μm). The program of GC-MS was set according to the previous method [22]. Chromatographic profiles of volatiles were identified by spectral library comparison (NIST standard library). The relative content of the compound referred to the internal standard and expressed as mg kg-1 fresh weight.
4.7. Determination of Soil Available Nitrogen (N), Potassium (K), and Phosphorus (P) Contents
Soil samples were collected on January 4, 2021 (before planting) and June 8, 2021 (later stage of fruiting), using the five-point sampling method. The soils were collected at 5-10 depth and mixed from 5 points in each treatment. Subsequently, the air-dried soil samples were filtered through a 2 mm sieve and divided into three replicates for the following analysis.
Available N content was measured according to the previous methods [60]. The soil samples were hydrolyzed with ferrous sulfate under alkaline conditions and then the hydrolyzed and nitrate N was converted to ammonium nitrogen. The diffusion ammonium nitrogen was absorbed by boric acid solution and titrated with standard acid to obtain the content of soil available nitrogen.
Available K content was estimated by Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES; Optima 8000, Perkin Elmer, USA) mentioned previously [61]. The soil sample (5 g) was added with 50 mL of ammonium acetate solution (1 mol/L, pH 7.0). After filtering, the test solution was detected by ICP-OES.
Available P was extracted with the solution of ammonium fluoride-hydrochloric acid and incubated at 25 °C with shaking for 30 min, and finally estimated by ICP-OES. The amounts of P and K were calculated from standard curves of P and K standard solution, respectively.
4.8. Real-time Fluorescence Quantitative PCR Analysis
Total RNA was extracted from the fruits and roots of the plants, respectively, during the fruiting stage (4 May 2021). Root samples were immediately frozen in liquid nitrogen after they were taken. The samples were mixed and ground using a mill, and stored at -80 °C, pooled samples were split into three replicates for the following analysis. The exaction of total RNA was used by a TransZol Plant kit (TransGen Biotech,Beijing, China). Afterwards, the concentration and purity of RNA were measured using a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, USA). cDNA was synthesized based on the manufacturer’s instruction of HiScript III-RT SuperMix kit (+gDNA wiper) for qPCR (Vazyme, Nanjing, China). The TransStart® Top Green qPCR SuperMix kit from TransGen was used for qPCR, which was conducted on a Mastercycler® EP Realplex Real-time PCR system from Eppendorf. The 18S rRNA and Actin (SlACT) were used as reference genes. The primer sequences are showed in Supplementary Table S2. Relative genes expression levels were determined with the 2−∆∆CT method [62].
4.9. Statistical Analysis
The results were analyzed by SPSS 24.0 software to express results by mean ± standard deviation (SD) for three independent experiments. One-way analysis of variance (ANOVA), Duncan’s multiple range test, and t-test were used for data analysis. Differences were considered significant at * P < 0.05 and ** P < 0.01.
The data was normalized and scaled (sum-based normalization, square root transformation, and auto scaling procedures). Principal component analysis (PCA) was carried out using MetaboAnalyst 5.0 (https://www.metaboanalyst.ca, accessed on 30 June 2022).
5. Conclusions
Sustained improvement of horticultural yield requires NPK fertilizers, which could be offset by healthier and cleaner alternatives to maintain fruit consumer quality. Compared to solid H2 storage materials such as MgH2, AB, and other nanoparticles, HNW will not bring additional elements. Recently, genetic and molecular evidence showed that molecular hydrogen not only influenced root organogenesis [63,64], but also increased nitrogen use efficiency (NUE) by targeting nitrate reductase [65]. Consistently, in this report, we revealed that the preharvest application of HNW exhibited fertilization effect on cherry tomato yield and improved its quality, and even to some extent as an alternative for the conventional fertilizers. Besides, production costs of renewable H2 are reducing to USD 0.7-1.6/kg H2 before 2050 on a global scale [66]. Consequently, HNW may provide an easy, affordable, and environment friendly solution for the reduction in fertilizer use, thus improving agricultural sustainability.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Profiles of volatiles in cherry tomatoes (2021); Table S2: Primers of qPCR used in this study. Figure S1: The contents of available nitrogen, available phosphorus, available potassium in soil of four greenhouses before fertilizing (Shanghai, 2021); Figure S2: Changes in H2 content of fresh HRW and HNW; Figure S3: The design and location information of the four greenhouses (Shanghai, 2021).
Author Contributions
Conceptualization, W.S.; methodology, S.W., M.L. and Y.L.; formal analysis, M.L. and Z.L.; investigation, S.W. and Y.L.; writing—original draft preparation, M.L., G.Z. and Z.L.; writing—review and editing, Z.L., L.L. and W.S.; visualization, G.Z.; W.L, Y.Z., X.C. and W.S.; project administration, Y.Z., X.C. and W.S.; funding acquisition, W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Air Liquide (China) R&D Co., Ltd., and Shanghai Key Laboratory of Hydrogen Science, Center of Hydrogen Science, Shanghai Jiao Tong University, China.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Evan Evans (University of Tasmania) for the English editing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest. Yan Zeng and Xu Cheng are employees and hold ownership interest (including patents) in Air Liquide (China) R&D Co., Ltd.
References
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Perveen, R.; Suleria, H.A.R.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Ahmad, S. Tomato (Solanum lycopersicum) carotenoids and lycopenes chemistry; metabolism, absorption, nutrition, and allied health claims—a comprehensive review. Crit. Rev. Food Sci. 2015, 55, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Weber, N.C.; Koron, D.; Jakopič, J.; Veberič, R.; Hudina, M.; Česnik, H.B. Influence of nitrogen, calcium and nano-fertilizer on strawberry (Fragaria × ananassa Duch.) fruit inner and outer quality. Agronomy-Basel. [CrossRef]
- Zhong, W.; Hu, C.; Wang, M. Nitrate and nitrite in vegetables from north China: Content and intake. Food Addit. Contam. 2002, 19, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.F.; Dou, Z.X.; He, P.; Ju, X.T.; Powlson, D.; Chadwick, D.; Norse, D.; Lu, Y.L; Zhang, Y; Wu, L; Chen, X.P; Cassman, K.G; Zhang, F.S. New technologies reduce greenhouse gas emissions from nitrogenous fertilizer in China. Proc. Natl Acad. Sci. USA 2013, 110, 8375–8380. [Google Scholar] [CrossRef] [PubMed]
- Matei, N.; Camara, R.; Zhang, J.H. Emerging mechanisms and novel applications of hydrogen gas therapy. Med. Gas Res. 2018, 8, 98–102. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signaling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Duan, X.; Yao, P.; Cui, W.; Cheng, D.; Zhang, J.; Jin, Q.; Chen, J.; Dai, T.; Shen, W. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Kuang, Y.; Feng, L.; Liu, Y.; Wang, S.; Du, H.; Shen, W. Molecular hydrogen maintains the storage quality of Chinese chive through improving antioxidant capacity. Plants 2021, 10, 1095. [Google Scholar] [CrossRef]
- Hu, H.; Zhao, S.; Li, P.; Shen, W. Hydrogen gas prolongs the shelf life of kiwifruit by decreasing ethylene biosynthesis. Postharvest Biol. Technol. 2018, 135, 123–130. [Google Scholar] [CrossRef]
- Li, L.; Yin, Q.; Zhang, T.; Cheng, P.; Xu, S.; Shen, W. Hydrogen nanobubble water delays petal senescence and prolongs the vase life of cut carnation (Dianthus caryophyllus L.) flowers. Plants 2021, 10, 1662. [CrossRef]
- Dong, Z.; Wu, L.; Kettlewell, B.; Caldwell, C.D.; Layzell, D.B. Hydrogen fertilization of soils-is this a benefit of legumes in rotation? Plant Cell Environ. 2003, 26, 1875–1879. [Google Scholar] [CrossRef]
- Golding, A.L.; Dong, Z. Hydrogen production by nitrogenase as a potential crop rotation benefit. Environ. Chem. Lett. 2010, 8, 101–121. [Google Scholar] [CrossRef]
- Hu, H.; Li, P.; Shen, W. Preharvest application of hydrogen-rich water not only affects daylily bud yield but also contributes to the alleviation of bud browning. Sci. Hortic. 2021, 287, 110267. [Google Scholar] [CrossRef]
- Xia, G.; Zhang, L.; Chen, X.; Huang, Y.; Sun, D.; Fang, F.; Guo, Z.; Yu, X. Carbon hollow nanobubbles on porous carbon nanofibers: An ideal host for high-performance sodium-sulfur batteries and hydrogen storage. Energy Storage Mater. 2018, 14, 314–323. [Google Scholar] [CrossRef]
- Zhao, G.; Cheng, P.; Zhang, T.; Abdalmegeed, D.; Xu, S.; Shen, W. Hydrogen-rich water prepared by ammonia borane can enhance rapeseed (Brassica napus L.) seedlings tolerance against salinity, drought or cadmium. Ecotox. Environ. Safe. 2021, 224, 112640. [CrossRef]
- Wang, Y.; Lv, P.; Kong, L.; Shen, W.; He, Q. Nanomaterial-mediated sustainable hydrogen supply induces lateral root formation via nitrate reductase-dependent nitric oxide. Chem. Eng. J. 2021, 405, 126905. [Google Scholar] [CrossRef]
- Temesgen, T.; Bui, T.T.; Han, M.; Kim, T.I.; Park, H. Micro and nanobubble technologies as a new horizon for water-treatment techniques: A review. Adv. Colloid Interfac. 2017, 246, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Zhang, Y.; Liu, S.; Li, X.; Li, J. Alleviation of copper toxicity in Daphnia magna by hydrogen nanobubble water. J. Hazard. Mater. 2020, 389, 122155. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Ding, X.W.; Jiang, R.; Ouyang, P.L.; Gui, J.; Feng, L.; Yang, L.; Song, L.H. Effects of hydrogen-rich water on the nutrient composition and antioxidative characteristics of sprouted black barley. Food Chem. 2019, 299, 125095. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Wang, J.; Zhao, Z.; Kong, L.; Lou, W.; Zhang, T.; Jing, D.; Yu, J.; Shu, Z.; Huang, L.; Zhu, W.; Yang, Q.; Shen, W. Molecular hydrogen increases quantitative and qualitative traits of rice grain in field trials. Plants 2021, 10, 2331. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, J.; Jiang, K.; Kuang, Y.; Zeng, Y.; Cheng, X.; Liu, Y.; Wang, S.; Shen, W. Preharvest application of hydrogen nanobubble water enhances strawberry flavor and consumer preferences. Food Chem. 2022, 377, 131953. [Google Scholar] [CrossRef]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [CrossRef]
- Yelle, S.; Hewitt, J.D.; Robinson, N.L.; Damon, S.; Bennett, A.B. Sink metabolism in tomato fruit: III. Analysis of carbohydrate assimilation in a wild species. Plant Physiol. 1988, 87, 737–740. [Google Scholar] [CrossRef]
- Navarro-González, I.; García-Valverde, V.; García-Alonso, J.; Periago, M.J. Chemical profile, functional and antioxidant properties of tomato peel fiber. Food Res. Int. 2011, 44, 1528–1535. [Google Scholar] [CrossRef]
- Arathi, B.P.; Sowmya, P.R.R.; Kuriakose, G.C.; Shilpa, S.; Shwetha, H.J.; Kumar, S.; Raju, M.; Vallikannan, B.; Lakshminarayana, R. Fractionation and characterization of lycopene oxidation products by LC-MSMS (ESI)+: Elucidation of chemoprevention potency of oxidized lycopene in breast cancer cell lines. J. Agric. Food Chem. 2018, 66, 11362–11371. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 815, 6. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Raddatz, N.; Pardo, J.M.; Yun, D.J. HKT sodium and potassium transporters in Arabidopsis thaliana and related halophyte species. Physiol. Plantarum 2021, 171, 546–558. [Google Scholar] [CrossRef]
- Li, L.; Lou, W.; Kong, L.; Shen, W. Hydrogen commonly applicable from medicine to agriculture: From molecular mechanisms to the field. Curr. Pharm. Design 2021, 27, 747–759. [Google Scholar] [CrossRef]
- Weldegebriel, R.; Araya, T.; Egziabher, Y.G. Effect of NPK and blended fertilizer application on nutrient uptake and use efficiency of selected sorghum (Sorghum bicolor (L.) Moench) varieties under rain-fed condition in Sheraro District, Northern Ethiopia. Momona Ethiop. J. Sci. 2018, 10, 140156. [CrossRef]
- Maimaiti, J.; Zhang, Y.; Yang, J.; Cen, Y.P.; Layzell, D.B.; Peoples, M.; Dong, Z. Isolation and characterization of hydrogen-oxidizing bacteria induced following exposure of soil to hydrogen gas and their impact on plant growth. Environ. Microbiol. 2007, 9, 435–444. [Google Scholar] [CrossRef]
- Dong, Z.; Layzell, D.B. H2 oxidation, O2 uptake and CO2 fixation in hydrogen treated soils. Plant Soil 2001, 229, 1–12. [Google Scholar] [CrossRef]
- Liu, H.; Wang, W.; Cao, G.; Tang, M. Effect of hydrogen on microbial population and enzyme activity in Robinia pseudoacacia rhizosphere soil. Chin. J. Appl. Environ. Biol. 2010, 16, 515–518. [Google Scholar] [CrossRef]
- Shukla, S.K.; Sharma, L.; Jaiswal, V.P.; Pathak, A.D.; Tiwari, R.; Awasthi, S.K.; Gaur, A. Soil quality parameters vis-a-vis growth and yield attributes of sugarcane as influenced by integration of microbial consortium with NPK fertilizers. Sci. Rep-UK. 2020, 10, 1-17. [CrossRef]
- Zhu, F.; Wen, W.; Cheng, Y.; Fernie, A.R. The metabolic changes that effect fruit quality during tomato fruit ripening. Mol. Hortic. 2022, 2, 1–19. [Google Scholar] [CrossRef]
- Liao, L.; Dong, T.; Qiu, X.; Rong, Y.; Wang, Z.; Zhu, J. Nitrogen nutrition is a key modulator of the sugar and organic acid content in citrus fruit. Plos One 2019, 14, e0223356. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhang, W.; Duan, X.; Dai, C.; Zhang, Y.; Cui, W.; Wang, R.; Shen, W. Hydrogen-rich water-alleviated ultraviolet-B-triggered oxidative damage is partially associated with the manipulation of the metabolism of (iso)flavonoids and antioxidant defence in Medicago sativa. Funct. Plant Biol. 2015, 42, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, G.; Cheng, P.; Yan, X.; Li, Y.; Cheng, D.; Wang, R.; Chen, J.; Shen, W. Nitrite accumulation during storage of tomato fruit as prevented by hydrogen gas. Int. J. Food Prop. 2019, 22, 1425–1438. [Google Scholar] [CrossRef]
- Yun, Z.; Gao, H.; Chen, X.; Chen, Z.; Zhang, Z.; Li, T.; Qu, H.; Jiang, Y. Effects of hydrogen water treatment on antioxidant system of litchi fruit during the pericarp browning. Food Chem. 2021, 336, 127618. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Kai, W.; Zhao, B.; Sun, Y.; Yuan, B.; Dai, S.; Li, Q.; Chen, P.; Wang, Y.; Pei, Y.; Wang, H.; Guo, Y.; Leng, P. SlNCED1 and SlCYP707A2: key genes involved in ABA metabolism during tomato fruit ripening. J. Exp. Bot. 2014, 65, 5243–5255. [Google Scholar] [CrossRef]
- Lomelí-Martín, A.; Martínez, L.M.; Welti-Chanes, J.; Escobedo-Avellaneda, Z. Induced changes in aroma compounds of foods treated with high hydrostatic pressure: A review. Foods 2021, 10, 878. [Google Scholar] [CrossRef] [PubMed]
- Tandon, K.S.; Baldwin, E.A.; Shewfelt, R.L. Aroma perception of individual volatile compounds in fresh tomatoes (Lycopersicon esculentum, Mill.) as affected by the medium of evaluation. Postharvest Biol. Technol. 2000, 20, 261–268. [CrossRef]
- Fan, Z.; Hasing, T.; Johnson, T.S.; Garner, D.M.; Schwieterman, M.L.; Barbey, C.R.; Colquhoun, T.A.; Sims, C.A.; Resende, M.F.R.; Whitaker, V.M. Strawberry sweetness and consumer preference are enhanced by specific volatile compounds. Hortic. Res. 2021, 8, 66. [CrossRef]
- Bood, K.G.; Zabetakis, I. The biosynthesis of strawberry flavor (II): Biosynthetic and molecular biology studies. J. Food Sci. 2002, 67, 2–8. [Google Scholar] [CrossRef]
- Ubeda, C.; San-Juan, F.; Concejero, B.; Callejón, R.M.; Troncoso, A.M.; Morales, M. L.; Ferreira, V.; Hernández-Orte, P. Glycosidically bound aroma compounds and impact odorants of four strawberry varieties. J. Agric. Food Chem. 2012, 60, 6095–6102. [Google Scholar] [CrossRef]
- Booth, D.A.; Kendal-Reed, M.S.; Freeman, R.P.J. A strawberry by any other name would smell as sweet, green, fruity and buttery. Multisensory cognition of a food aroma. Appetite 2010, 55, 738–741. [Google Scholar] [CrossRef]
- Yan, J.W.; Ban, Z.J.; Lu, H.Y.; Li, D.; Poverenov, E.; Luo, Z.S.; Li, L. The aroma volatile repertoire in strawberry fruit: A review. J. Sci. Food Agric. 2018, 98, 4395–4402. [Google Scholar] [CrossRef]
- Choi, S.T.; Park, D.S.; Kang, S.M.; Kang, S.K. Influence of leaf-fruit ratio and nitrogen rate on fruit characteristics, nitrogenous compounds, and nonstructural carbohydrates in young persimmon trees. Hortscience 2012, 47, 410–413. [Google Scholar] [CrossRef]
- Davis, D.R. Declining fruit and vegetable nutrient composition: What is the evidence? Hortic. Sci. 2009, 44, 15–19. [Google Scholar] [CrossRef]
- Liu, L.; Ji, M.L.; Chen, M.; Sun, M.Y.; Fu, X.L.; Li, L.; Gao, D.S.; Zhu, C.Y. The flavor and nutritional characteristic of four strawberry varieties cultured in soilless system. Food Sci. Nutr. 2016, 4, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Buckman, E.S.; Oduro, I.; Plahar, W.A.; Tortoe, C. Determination of the chemical and functional properties of yam bean (Pachyrhizus erosus (L.) Urban) flour for food systems. Food Sci. Nutr 2018, 6, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Gao, J.; Gu, L.; Wang, S.; Zeng, R. Effects of agaro-oligosaccharide treatment on postharvest quality of cherry tomatoes during cold storage. J. Food Process. Preserv. 2015, 39, 949–955. [Google Scholar] [CrossRef]
- Liu, Y.J.; Wang, G.L.; Ma, J.; Xu, Z.S.; Wang, F.; Xiong, A.S. Transcript profiling of sucrose synthase genes involved in sucrose metabolism among four carrot (Daucus carota L.) cultivars reveals distinct patterns. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogy. 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Giovanelli, G.; Zanoni, B.; Lavelli, V.; Nani, R. Water sorption, drying and antioxidant properties of dried tomato products. J. Food Eng. 2002, 52, 135–141. [Google Scholar] [CrossRef]
- Musa, K.H.; Abdullah, A.; Jusoh, K.; Subramaniam, V. Antioxidant activity of pink-flesh guava (Psidium guajava L.): Effect of extraction techniques and solvents. Food Anal. Method. 2011, 4, 100–107. [Google Scholar] [CrossRef]
- Hasperué, J.H.; Rodoni, L.M.; Guardianelli, L.M.; Chaves, A.R.; Martínez, G.A. Use of LED light for Brussels sprouts postharvest conservation. Sci. Hortic. 2016, 213, 281–286. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, J.; Tian, J.; Li, N.; Jia, L.; Shen, W.; Cui, J. Enhanced anthocyanin accumulation of immature radish microgreens by hydrogen-rich water under short wavelength light. Sci. Hortic. 2019, 247, 75–85. [Google Scholar] [CrossRef]
- Lu, B.; Liu, N.; Wang, X.; Hu, C.; Tang, X. A feasibility quantitative analysis of NIR spectroscopy coupled Si-PLS to predict coco-peat available nitrogen from rapid measurements. Comput. Electron. Agr. 2020, 173, 105410. [Google Scholar] [CrossRef]
- Alexander, D.; Ellerby, R.; Hernandez, A.; Wu, F.; Amarasiriwardena, D. Investigation of simultaneous adsorption properties of Cd, Cu, Pb and Zn by pristine rice husks using ICP-AES and LA-ICP-MS analysis. Microchem. J. 2017, 135, 129–139. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, Y.; Zhang, G.; Li, L.; Shen, W. Molecular hydrogen positively influences lateral root formation by regulating hydrogen peroxide signaling. Plant Sci. 2022, 325, 111500. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, S.; Liu, Z.; Chen, G.; Cheng, P.; Li, L.; Xu, S.; Shen, W. H2 supplied via ammonia borane stimulates lateral root branching via phytomelatonin signaling. Plant Physiol. 2023. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, Y.; Cai, C.; Li, L.; Zeng, Y.; Cheng, X.; Shen, W. Molecular hydrogen positively regulates nitrate uptake and seed size by targeting nitrate reductase. Plant Physiol. 2023, 193, 2734–2749. [Google Scholar] [CrossRef]
- BloombergNEF. Hydrogen economy outlook. 2020. Available online: https://data.bloomberglp.com/professional/sites/24/BNEF-Hydrogen-Economy-Outlook-Key-Messages-30-Mar-2020.pdf (accessed on 12 Nov 2023).
Figure 1.
Hydrogen nanobubble water (HNW)-promoted cherry tomato growth (A) and yield per plant (B) with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 1.
Hydrogen nanobubble water (HNW)-promoted cherry tomato growth (A) and yield per plant (B) with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
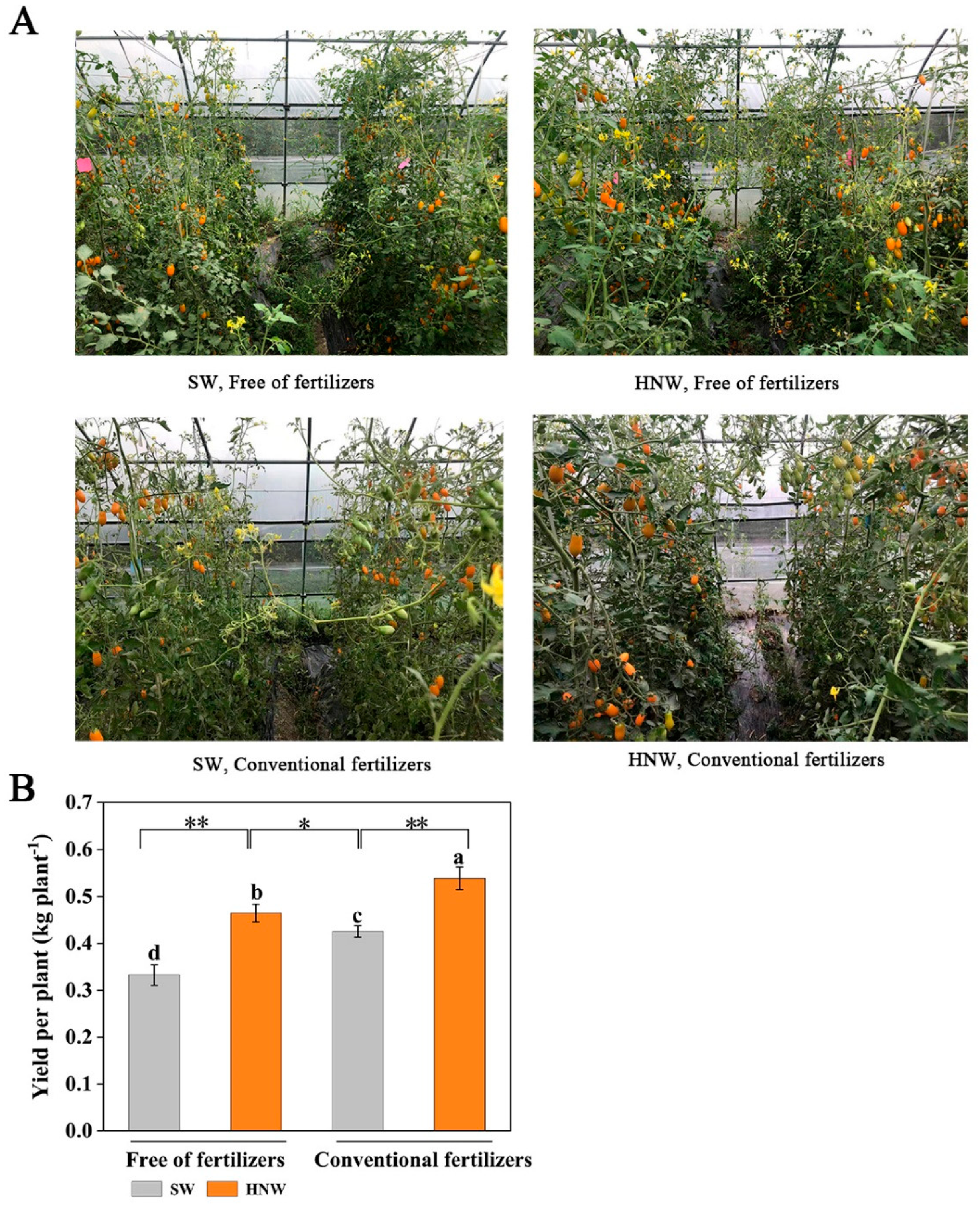
Figure 2.
Effects of hydrogen nanobubble water on the yield per plant (A), number of fruits per plant (B), single fruit weight (C) and yield (D) of cherry tomatoes with/without fertilizers (Nanjing, 2023). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 2.
Effects of hydrogen nanobubble water on the yield per plant (A), number of fruits per plant (B), single fruit weight (C) and yield (D) of cherry tomatoes with/without fertilizers (Nanjing, 2023). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
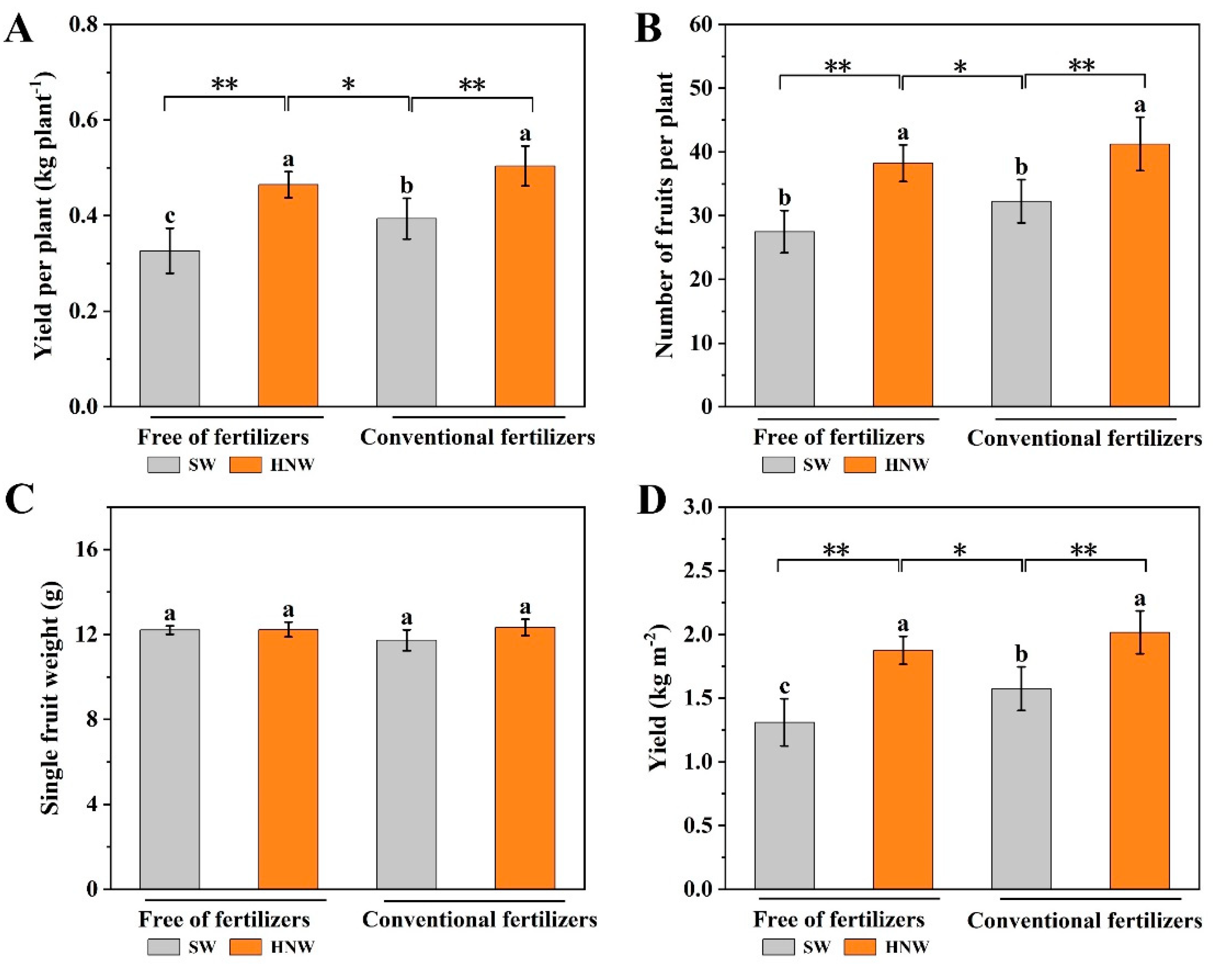
Figure 3.
Effects of hydrogen nanobubble water on the contents of soluble sugar (A), titratable acid (B), sugar-acid ratio (C), fructose (D), glucose (E), and sucrose (F) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 3.
Effects of hydrogen nanobubble water on the contents of soluble sugar (A), titratable acid (B), sugar-acid ratio (C), fructose (D), glucose (E), and sucrose (F) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
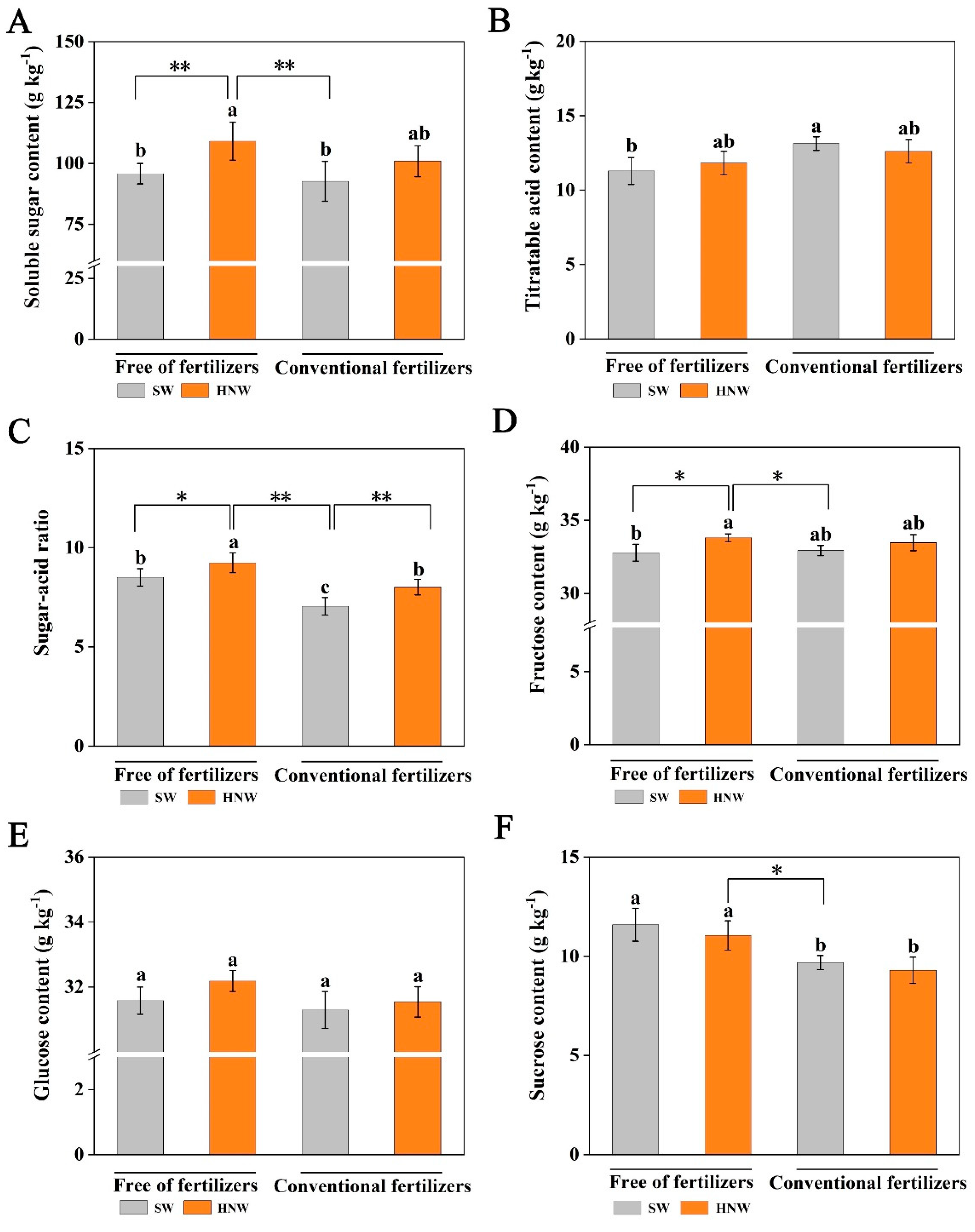
Figure 4.
Effects of HNW on the accumulation of lycopene (A), total phenols (B), vitamin C (C), and flavonoids (D) and the expression level of SlPSY1 (E) and SlPDS (F) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 4.
Effects of HNW on the accumulation of lycopene (A), total phenols (B), vitamin C (C), and flavonoids (D) and the expression level of SlPSY1 (E) and SlPDS (F) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
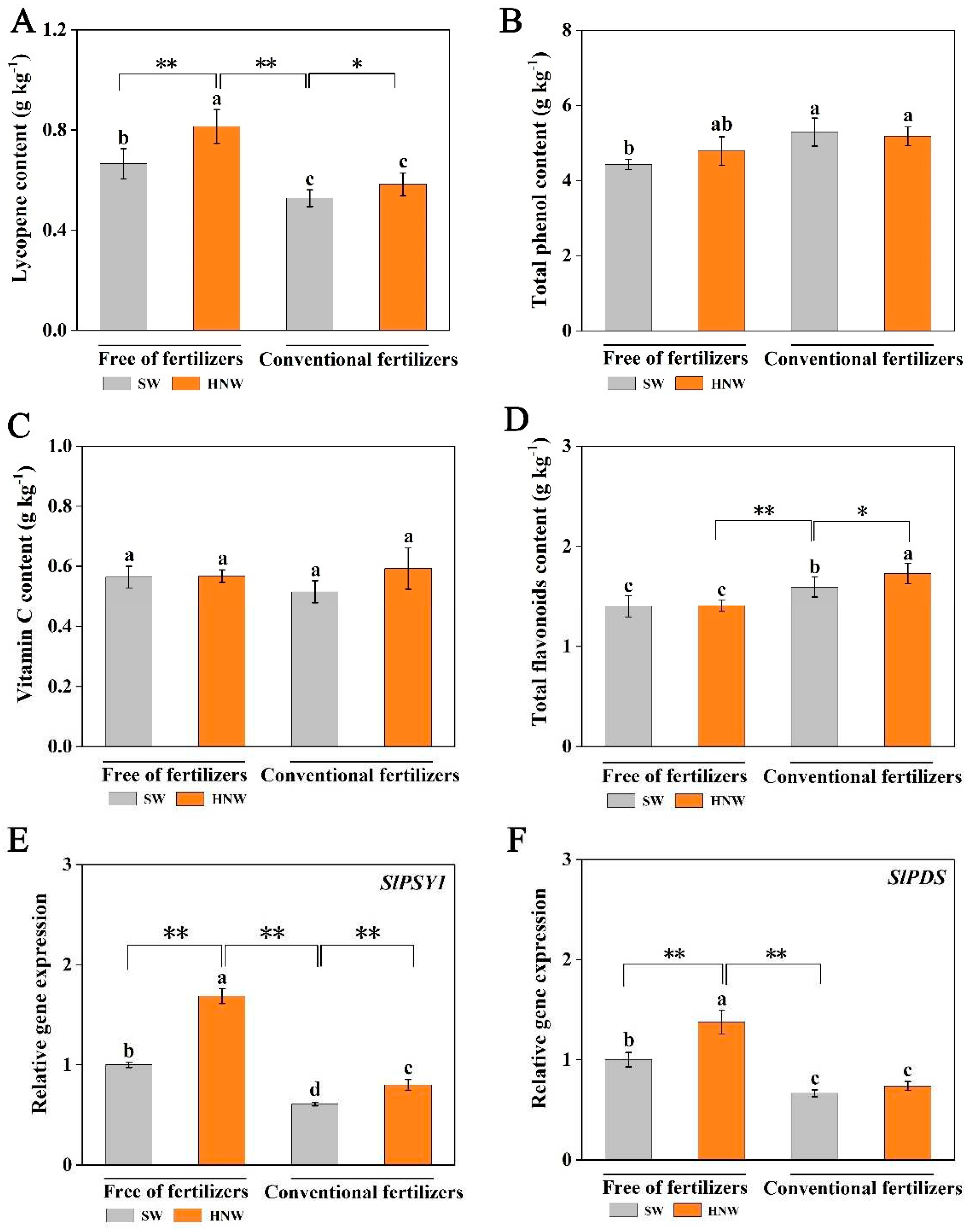
Figure 5.
Effects of HNW on the concentrations of the volatile compounds (A), aldehydes (B), alcohols (C), hexanal (D), E-2-hexenal (E), trans-1,2-Cyclopentanediol (F) and 2,4-Bis(1,1-dimethylethyl)-phenol (G) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 5.
Effects of HNW on the concentrations of the volatile compounds (A), aldehydes (B), alcohols (C), hexanal (D), E-2-hexenal (E), trans-1,2-Cyclopentanediol (F) and 2,4-Bis(1,1-dimethylethyl)-phenol (G) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
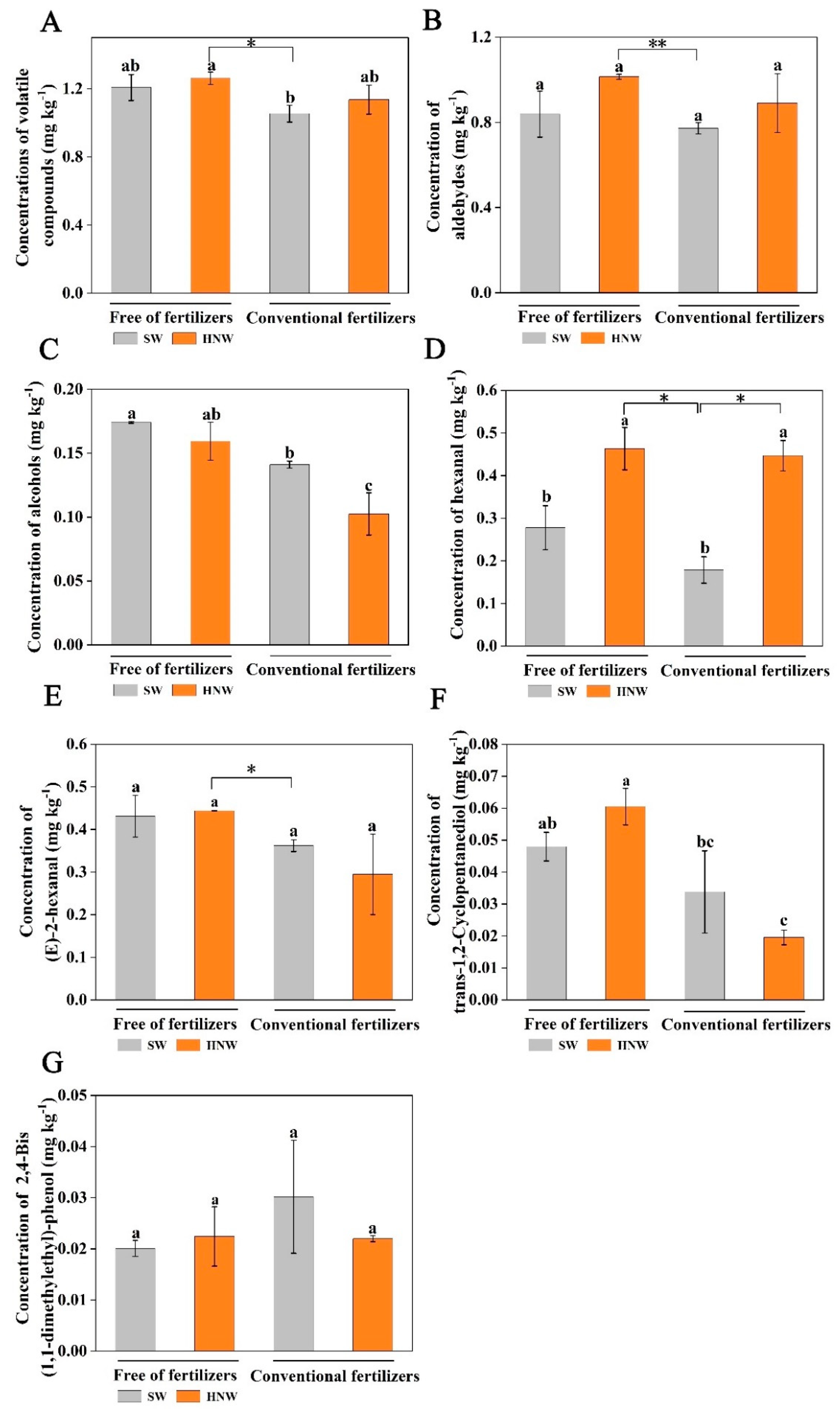
Figure 6.
Effects of HNW on the absorption of soil available nitrogen (A), phosphorus (B), and potassium (C) and the expression level of LeAMT2 (D), LePT5 (E), LePT2 (F), and SlHKT1,1 (G) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
Figure 6.
Effects of HNW on the absorption of soil available nitrogen (A), phosphorus (B), and potassium (C) and the expression level of LeAMT2 (D), LePT5 (E), LePT2 (F), and SlHKT1,1 (G) in cherry tomatoes with/without fertilizers (Shanghai, 2021). Values are mean ± SD of three independent experiments. The asterisks *, ** indicate significant differences at P < 0.05 and P < 0.01, respectively (t-test). The different letters indicate significant differences at P < 0.05 (one-way ANOVA; Duncan's multiple range tests). SW: surface water; HNW: hydrogen nanobubble water.
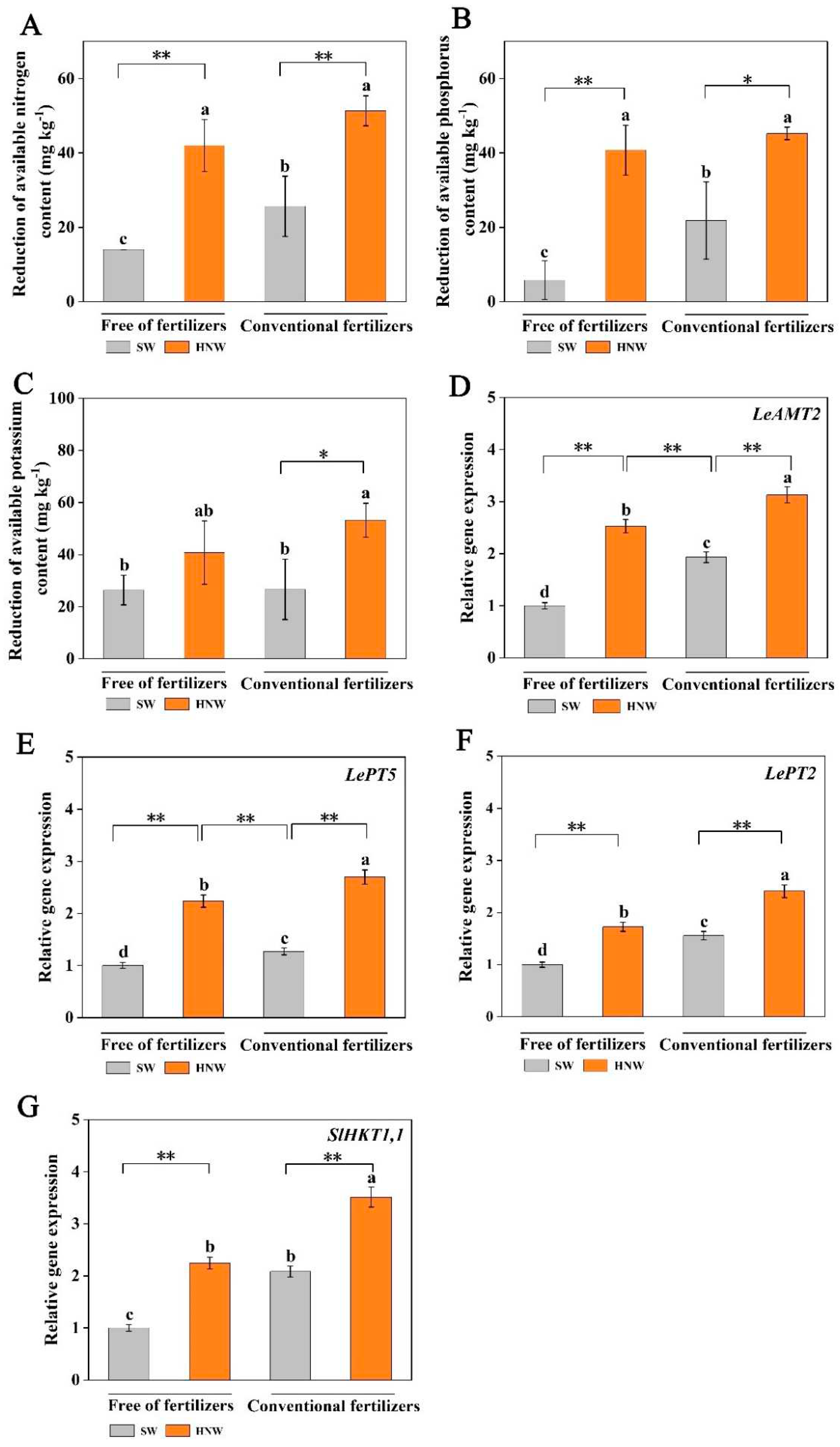
Figure 7.
Scores plot (A) and biplot (B) of pr incipal component analysis (PCA) for effects of HNW on yield and quality in cherry tomatoes with/without fertilizers (Shanghai, 2021). HNW: hydrogen nanobubble water; 2,4-Bis: 2,4-Bis(1,1-dimethylethyl)-phenol; HNW + F: hydrogen nanobubble water plus fertilizers; SW: surface water; SW + F: surface water plus fertilizers; Trans-1,2: trans-1,2-cyclopentanediol.
Figure 7.
Scores plot (A) and biplot (B) of pr incipal component analysis (PCA) for effects of HNW on yield and quality in cherry tomatoes with/without fertilizers (Shanghai, 2021). HNW: hydrogen nanobubble water; 2,4-Bis: 2,4-Bis(1,1-dimethylethyl)-phenol; HNW + F: hydrogen nanobubble water plus fertilizers; SW: surface water; SW + F: surface water plus fertilizers; Trans-1,2: trans-1,2-cyclopentanediol.
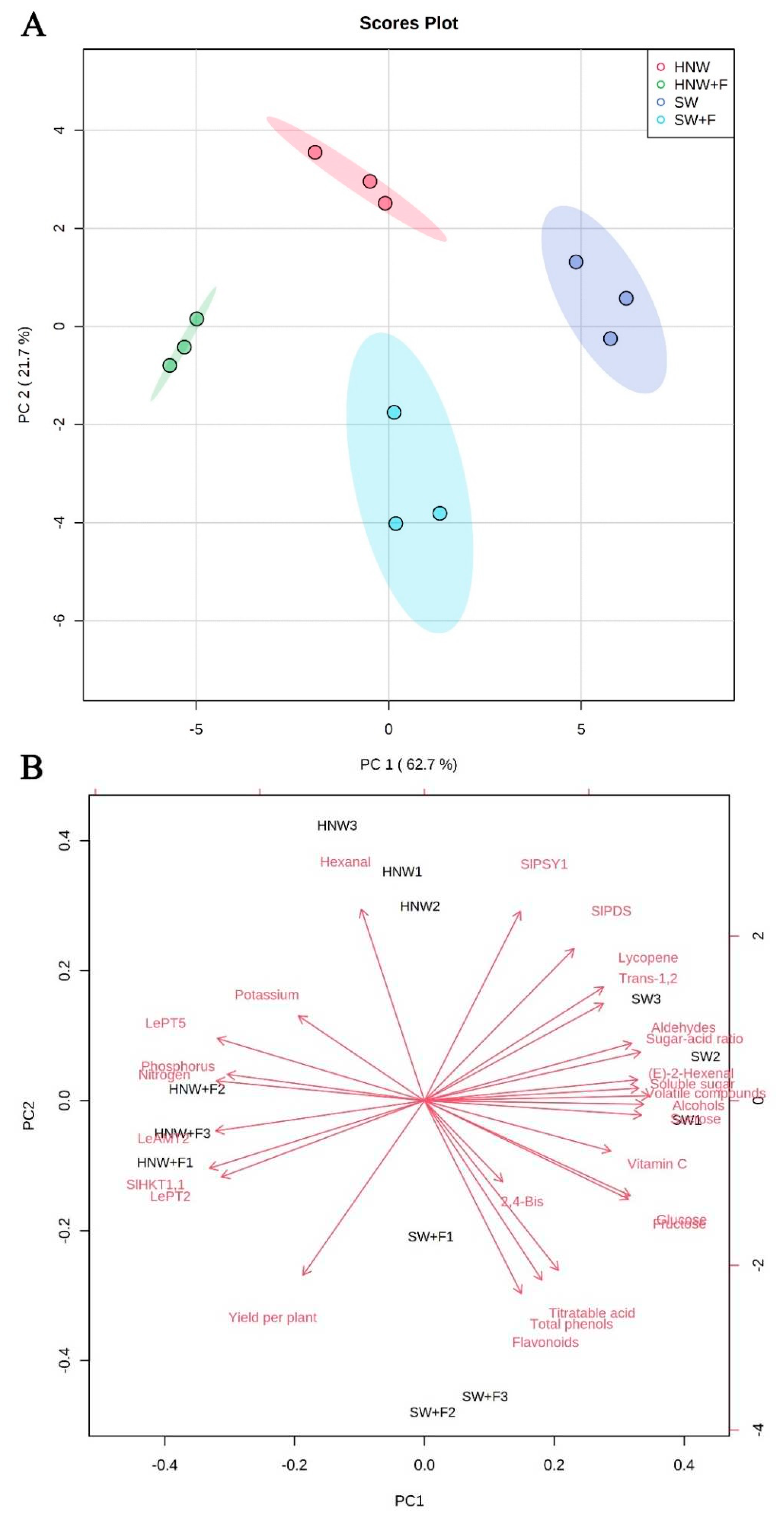
Figure 8.
Proposed schematic model of HNW-improved yield and quality in cherry tomatoes. Compared with surface water plus fertilizers, red arrows (left) indicate the effects achieved by HNW without fertilizers, and blue arrows (right) indicate the effects achieved by HNW plus fertilizers. AMT2: ammonium transporter 2; 2,4-Bis: 2,4-Bis(1,1-dimethylethyl)-phenol; HKT1,1: high-affinity K+ channel transporter; PT2/5: phosphate transporter 2/5; PDS: phytoene desaturase; PSY: phytoene synthase; Trans-1,2: trans-1,2-cyclopentanediol.
Figure 8.
Proposed schematic model of HNW-improved yield and quality in cherry tomatoes. Compared with surface water plus fertilizers, red arrows (left) indicate the effects achieved by HNW without fertilizers, and blue arrows (right) indicate the effects achieved by HNW plus fertilizers. AMT2: ammonium transporter 2; 2,4-Bis: 2,4-Bis(1,1-dimethylethyl)-phenol; HKT1,1: high-affinity K+ channel transporter; PT2/5: phosphate transporter 2/5; PDS: phytoene desaturase; PSY: phytoene synthase; Trans-1,2: trans-1,2-cyclopentanediol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



