
Submitted:
12 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
Perinatal brain injury is a major public health problem burdened with high morbidity, mortality and severe neurological sequelae. Despite advances in neonatal life-saving technologies, neonatal encephalopathy lacks specific and effective treatment. Therefore, mechanisms of central nervous system changes associated with perinatal brain injury have been a hot topic of neonatology research. In recent years, researchers have shifted more focus from neurons themselves to perineuronal cells and intercellular interactions. As a member of the nervous system network, astrocytes are one of the dominant glial cells that regulate ion homeostasis, blood flow, and antioxidant function in the brain. Their changes in molecular mechanisms (e.g., alterations in RNA regulatory networks) during brain injury storms deserve further attention. Non-coding RNAs participate in and promote reactive pathophysiologic changes in astrocytes. The review summarizes and discusses the role of non-coding RNAs in astrocytes in recent years in studies of neonatal brain injury. It focuses on the most studied disease-related ncRNAs: miRNAs, lncRNAs and circRNAs, and hopes to provide possible directions for pathophysiological studies and therapeutic approaches of neonatal brain injury.
Keywords:
astrocyte
; non-coding RNAs
; miRNAs
; lncRNAs
; circRNAs
; neonatal brain injury
; hypoxic-ischemic
; neuroinflammation
1. Introduction
Of the 530 million deaths among children younger than 5 years in 2000-19, the majority (46% 43.5-48.7%)) were neonates, and the leading causes of death among those children was preterm birth complications (17·7% [16.1–19.5])[1]. During the perinatal period, the immature brain undergoes several critical periods of neurodevelopment, and brain damage from internal and external insults impairs those crucial phases, followed by long-term consequences for neurological and mental health[2]. Hypoxia-ischemia (HI) and neuroinflammation, in particular, are the most prominent causes of neuronal loss, leading to diffuse white matter injury (dWMI) and gray matter injury in neonates[3]. Oxidative stress leads to apoptosis, necrosis, and autophagy pathway activation, determining the final infarct size. The rapid production of reactive oxygen species (ROS) after stroke further aggravates brain damage[4]. The inflammatory response has a bilateral contribution to the mechanism of ischemic brain injury[5]. Inflammatory mediators are upregulated from resident brain cells and infiltrating immune cells[6], which in turn propagate microvascular dysfunction, edema expansion, and ultimately worsen the clinical outcome[7].
Astrocytes are abundant glial cells in the central nervous system (CNS)[8]. In the neonatal brain, astrocytes collaborate with pericytes to form the neurovascular unit (NVU) that performs basic functions necessary for physiological processes in the CNS[9], including supporting neuronal structure, regulating blood flow, ion homeostasis, and stabilizing the extracellular environment[10]. The majority of synapse genesis and maturation, as well as lifelong functional regulation, are active in the astrocyte cradle[11]. In the context of brain injury induced by inflammation or neonatal hypoxia-ischemia, astrocytes activate and undergo morphological changes (cell foot swelling)[12], secrete cytokines that actively promote neuroinflammatory cascades[13]. Changes in astrocyte function or density lead to altered neurologic outcomes.
In about 95% of genes transcribed, coding-protein-RNAs account for only 1.5-2%[14]. Non-coding RNAs other than mRNAs, including microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), and other extracellular RNAs, have been further elucidated in recent years with the development of high-throughput profiling and comprehensive computational analysis techniques, which have been used to elaborate further the key mechanisms involved in disease regulation[15]. In the central nervous system, although ncRNAs do not encode proteins, they coordinate and control various developmental stages of neural stem cells, neurons, and glial cells at the post-transcriptional level[16], which makes them a novel and challenging class of potential drug targets.
In this review, we summarize and discuss the role of non-coding RNAs in astrocytes in recent years in studies of neonatal brain injury. We focus on the most studied disease-related ncRNAs: miRNAs, lncRNAs and circRNAs. Potential directions are provided for biomarkers and therapeutic targets of neonatal brain injury by drawing on the altered and therapeutic roles of relevant ncRNAs in astrocytes in adult neurological disorders.
The introduction should briefly place the study in a broad context and highlight why it is important. It should define the purpose of the work and its significance. The current state of the research field should be carefully reviewed and key publications cited. Please highlight controversial and diverging hypotheses when necessary. Finally, briefly mention the main aim of the work and highlight the principal conclusions. As far as possible, please keep the introduction comprehensible to scientists outside your particular field of research. References should be numbered in order of appearance and indicated by a numeral or numerals in square brackets—e.g., [1] or [2,3], or [4,5,6]. See the end of the document for further details on references.
2. Mechanism of ncRNA Action in Astrocytes under HI Conditions
Neonatal hypoxic-ischemia (HI), also known as hypoxic-ischemic encephalopathy (HIE),is a major public health problem with a high morbidity and mortality rate[17]. In the hours after glucose-oxygen deprivation, astrocyte gene expression is altered in response to hypoxia, neuronal cell death, neurotransmitter release, and cytotoxic substance production. Astrocyte activation and transition to type A2 in a transient middle cerebral artery occlusion stroke (tMACO) model, exerting neurotrophic or anti-inflammatory effects[18]. A growing number of studies in recent years have demonstrated that non-coding RNAs play an important role in molecular changes and phenotypic shifts in reactive astrocytes.
2.1. MiRNAs
Astrocyte swelling is one of the earliest responses to cerebral hypoxia-ischemia, which is regulated by water channel protein aquaporin 4 (AQP4). AQP4 is highly expressed in astrocytes and is enriched in astrocyte end-feet and gathered at cell synapses and edges[19]. It mediates water exchange through the blood-brain barrier (BBB), and controls cell volume, extracellular space volume, and astrocyte migration[20]. It was found that cellular volume during neonatal HI in AQP4−/− astrocytes was significantly less than that in AQP4+/+ astrocytes, and demonstrated that modulation of astrocyte AQP4 at different stages of HIE (HI and reoxygenation) may be beneficial for the treatment[21]. MiR-320a directly targets AQP4[22]. In a neonatal mouse model of epilepsy (intraperitoneal injection of lithium chloride-pilocarpine), brain miR-320a expression levels were decreased and negatively correlated with AQP4 expression[23]. Anti-miR-320a administration benefits ischemic rats and reduces cerebral infarct volume[22]. In cerebral ischemia/reperfusion injury (I/RI), miR-130b regulates AQP4 expression at the post-transcriptional level and produces neuroprotection[24]. Additionally, miR-145 protects astrocytes from post-OGD injury by inhibiting AQP4 expression[25]. With the established belief that AQP4 inhibitors may be a therapeutic option in the absence of cytotoxic edema after HIE, targeting relevant miRNAs provides possible new directions for drug development.
AQP4 co-localizes extensively with glutamate transporter 1 (GLT-1) and has a regulatory role for GLT-1 in astrocytes[26]. One of the basic functions of astrocytes is to encapsulate and take up most synaptically released glutamate, which optimizes neuronal function and prevents glutamate excitotoxicity[27]. The major transporters that function include the glutamate-aspartate transporter (GLAST, human homologs EAAT1) and glutamate transporter-1 (GLT-1, human homologs EAAT2)[28]. Interestingly, glutamate transport from neurons to astrocytes is inefficient in the neonatal brain, which may be related to the low expression of astrocytic glutamate transporters[29]. It has also demonstrated that GLAST predominates in the neonatal neocortex and hippocampus[30], which could explain the particular vulnerability to excitotoxicity of the neonatal brain[31]. Upregulation of astrocyte GLT-1 could be a potential therapeutic target for HIE. MiR-124 expression is significantly downregulated in a neonatal HIE model and an in vitro OGD model. It was found that overexpression of miR-124 was neuroprotective and anti-apoptotic and ameliorated long-term neurological deficits[32]. It also regulates GLT-1 expression in astrocytes after OGD/reperfusion via Akt and mTOR pathways[33]. In addition, of the neurologic disorders that cause brain injury in adults, miR-218 is abundantly expressed in motor neurons, and in studies of ALS, it was found that motor neuron-derived miR-218 can be taken up by peripheral astrocytes and is sufficient to down-regulate EAAT2 in astrocytes and affect glutamate uptake[34]. Interestingly, miR-218-5p promotes endovascular trophoblast differentiation and spiral artery remodeling and is down-regulated in preeclamptic (PE) placentas[35], which are associated with preterm labor, neonatal asphyxia and even neonatal death[36]. Targeting the specific mechanisms of miR-218 and its role in the placenta may increase mechanistic understanding of miR-218 within neonatal encephalopathy.
During cerebral ischemia, astrocytes undergo autophagy, and attempts to cope with stress in this way may protect astrocytes from apoptosis in response to ischemic injury[37]. Autophagy is a lysosomal degradation pathway in which the cell's own cytoplasmic proteins or organelles are phagocytosed and encapsulated into vesicles, which fuse with lysosomes and degrade their contents[38]. Melatonin administration attenuates brain damage in neonatal HI rats by inhibiting cellular autophagy and decreasing apoptosis, obtaining a neuroprotective effect[39]. The microRNA-30 family consists of six different miRNAs, including miR-30a, miR-30b, miR-30c-1, miR-30c-2, miR-30d, and miR-30e, which are important regulators of tissue and organ development as well as the pathogenesis of related diseases[40,41]. In related studies, miR-30d inhibited the expression of Beclin1 in rat primary astrocytes[42,43]. Beclin1 is closely associated with autophagosome formation and fusion of autophagosomes, and its interaction with Bcl-2 family proteins mediates cellular autophagy and apoptosis[44]. Thus miR-30d can promote apoptosis by inhibiting OGD-induced autophagy in astrocytes[43]. What’s more, miR-30d-5p was found to be involved in neuronal apoptosis and autophagy in neonatal HI rats and may also potentially target brain-derived neurotrophic factor (BDNF)[45]. Targeted modulation of the miR-30 family in neonatal encephalopathy could serve as a potential therapeutic strategy. In addition, astrocytes undergo pyroptosis during HIBD. It is found that miR-148-3p inhibits pyroptosis of rat hippocampal astrocytes by blocking STAT/JMJD3 axis, and dexmedetomidine (DEX), a potent α2-adrenoceptor agonist, attenuates brain injury in neonatal rats through upregulation of miR-148a-3p[46].
As one of the brain-specific microRNAs, miR-134 promotes the proliferation of neural precursor cells and embryonic neuronal maturation in neuronal development[47]. miR-134 localizes to the synaptic-dendritic spacer and regulates excitatory synaptic propagation at postsynaptic sites in rat hippocampal neurons thereby modulating synaptic development, maturation, and/or plasticity[48]. Overexpression of miR-134 negatively affects and impairs neuronal formation in the postnatal mouse brain[49]. The study demonstrates that miR-134-5 expression is increased and mediates hypoxic-ischemic brain injury and neuronal death via the C/EBPα/miR-134-5p/KPNA3 axis in pheochromocytoma (PC12) cells[50] and inhibition of NLRX1 expression[51] in a neonatal rat model of HIE. However, overexpression of miR-134 did not affect astrocyte survival but decreased extracellular glutamate concentration and promoted astrocyte maturation by increasing GLT-1[52]. This may be involved in the future synergistic treatment of neonatal HIE.
2.2. LncRNAs and circRNAs
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is a long noncoding RNA that was initially identified to be associated with non-small cell lung cancer as well as lung cancer metastasis[53]. Recent studies have revealed that lncRNA MALAT1 is also involved in the regulation of endothelial cell function and vascular growth[54]. In vivo studies confirmed that lncRNA MALAT1 is significantly increased under hypoxia and controls phenotypic switch in endothelial cells. Genetic ablation of lncRNA MALAT1 inhibited endothelial cell proliferation and reduced neonatal retinal vascularization[55]. Of attention is the fact that lncRNA MALAT1 also exerts its regulatory function in astrocytes. It was demonstrated that lncRNA MALAT1 affects AQP4 expression in astrocytes by competitively binding miR-145, increases the number of apoptotic cells and infarct size, and promotes cerebral ischemia-reperfusion injury[56]. Knockdown or silencing of lncRNA MALAT1 protects neonatal bronchopulmonary dysplasia (BPD) and ischemic cardiomyopathy[57,58]. Correspondingly, the role of lncRNA MALAT1 in ischemic stroke provides a new therapeutic strategy for neonatal ischemic encephalopathy.
LncRNA H19 (H19) was originally identified as a fetal transcript, which is widely expressed in mouse fetal liver, intestine, cardiac muscle, and skeletal muscle[59]. At present, this transcript has been recognized as an oncogenic lncRNA, which is involved in human cancer development through different mechanisms[60]. H19 was upregulated in HIE newborns and rat models. It was proved that H19 alleviated nerve damage of neonatal HIE rats by upregulating the Akt3/mTOR pathway as a miR-29b sponge[61]. It is confirmed that H19 is transported from neurons to astrocytes by exosomes to induce downregulation of insulin-like growth factor-1 (IGF-1) through the H19/let-7a/IGF-1 receptor axis[62]. A further study elucidated that H19 could sponge miR-130a-3p and competitively regulate the downstream target DAPK1, which in turn worsen brain injury in neonatal HIE[63]. Inhibition of H19 may benefit neonatal brain injury, but further studies are needed regarding the specific role played by H19 in astrocytes after neonatal HI.
A study comparing three-month-old and eight-year-old pigs by cerebral cortex transcriptomic analysis found that circRNA expression in the brain depended on developmental age, while >80% of mRNAs, miRNAs, and lncRNAs but <20% of circRNAs were expressed at both ages[64]. Interestingly, circRNAs can determine developmental and aging processes by altering the expression of specific mRNAs[65]. The study identified an altered expression pattern of circRNAs in neonatal hypoxic-ischemic brain damage (HIBD) rats by high-throughput sequencing analysis and predicted circRNA/ miRNA interactions[66], further demonstrating that dysregulated circRNAs mechanistically behave similarly to those in adults during the physiopathological process of HIBD.
Additionally, circRNAs play a role in astrocytes in adult stroke, for example: circRNA HECTD1 (circHectd1) expression level is significantly increased in tMCAO mice model. Mechanistically, circHectd1 can function as an endogenous miR-142 sponge to inhibit the activity of miR-142/TIPARP axis, which in turn inhibits astrocyte activation by promoting cell autophagy[67]. Furthermore, circ_0025984 acts as a sponge for miR-143-3p to counteract its effects on autophagy and apoptosis in astrocytes under ischemic conditions to achieve protection of astrocytes[68]. (Table 1)
3. Non-Coding RNAs Modulate Astrocytes in Neuroinflammation
Inflammation is usually central to the progression of brain injury disease as a secondary injury factor[81]. Inflammation is also a trigger for brain injury in neonates, especially preterm infants. Chorioamnionitis is the most common cause of preterm birth[82]. Fetal inflammatory response syndrome (FIRS) can be induced when the fetus is directly exposed to intrauterine infection/inflammation through direct skin contact or intrauterine swallowing and respiration[83], while the immune system releases pro-inflammatory factors such as IL-6, which is associated with an increased risk of intraventricular hemorrhage (IVH)[84]. During neonatal neuroinflammation, astrocytes that are tightly coupled to cerebral vasculature have altered morphology (astrocytic feet swelling) and decreased vascular coverage in the white matter[85], while actively contributing to the neuroinflammatory cascade response. Inflammation-induced reactive astrocytes, mostly A1 type, secrete pro- and anti-inflammatory cytokines and matrix metalloproteinases (MMPs)[86], increase glial fibrillary acidic protein (GFAP) expression, and proliferate to form glial scars after insult[87,88].
3.1. MiRNAs
The reactive proliferation of astrocytes is an important step in their ability to inhibit the spread of neuroinflammation[88], but hyperproliferation of astrocytes can lead to scarring and thus long-term neurodevelopmental effects. MicroRNAs can be potential markers and targeted therapeutic factors by regulating astrocyte activation and proliferation. For instance, hsa-miR-145-5p expression is significantly increased in neonates with poor neurodevelopmental outcomes after asphyxia[77]. What’s more, it was found to be a negative regulator of astrocyte proliferation, and its down-regulation promoted smad3 activity, which in turn promoted astrocyte proliferation[89]. In vitro experiments showed that the potent inflammatory agent lipopolysaccharide (LPS) down-regulated miR-145 expression in astrocytes isolated from neonatal spinal cord tissues, and it was also suggested that miR-145 potentially targets GFAP and c-myc to inhibit inflammation-induced astrocyte proliferation[90]. In combination, miR-145/miR-145-5p may be partially neuroprotective by inhibiting reactive astrogliosis after HIE.
The transcription factor NF-κB is a central regulator of inflammation[91]. Proinflammatory mediators such as reactive oxygen species (ROS), cytokines (TNF-α, IL-1β, IL-17[92]), LPS, Toll-like receptors ligand (CD40L), and altered environmental elements activate NF-κB signaling in astrocytes[93] and may promote neuroinflammation and neurodegeneration[94]. Neuroinflammation of neonatal brain structures, including the forebrain cortex, hippocampus, and cerebellum, can be directly linked to increased levels of NF-kB expression[95]. NF-κB downregulation in reactive astrocytes exerts an inhibitory effect on CNS inflammation[96]. MiRNA-155 can promote hypoxic-ischemic brain injury in neonatal rats by regulating the NOD1/NF-κB signaling pathway. Its mechanism may be related to the activation of the NOD1 signaling pathway by miRNA-155, phosphorylation of downstream NF-κB, and promotion of brain inflammation in neonatal HIE rats[97]. In spinal cord injury (SCI) rat model, miR-136-5p was found to up-regulates p-NF-κB by down-regulating the expression of A20 protein, which promotes the production of inflammatory factors and chemokines by astrocytes, thereby increasing inflammatory cell infiltration and injury in the spinal cord[98]. Reactive changes in microglia in neonatal brain injury are often the first step in response to neuroinflammation. MiRNAs such as miR-210 induce microglial M1 activation and increase NF-kB signaling pathway activity in neonatal HIE and promote microglia-mediated neuroinflammation and brain injury[99]. Activation of A1 astrocytes is influenced by microglia activity. Regulating both phenotypic transitions by targeting ncRNAs may be an effective therapeutic strategy. For example, In vitro administration of miR-124-3pOE-Exos inhibits activation of M1 microglia and A1 astrocyte through activation of PI3K/AKT and hence inhibit the NF-κB signaling cascade[100], thereby providing protection against neuroinflammatory injury and offering a possible therapeutic strategy for neonatal inflammatory encephalopathy.
MicroRNA-124 has two mature forms: miR-124-3p (known as miR-124 or miR-124a) and miR-124-5p (known as miR-124*)[101]. They are abundantly and specifically expressed in the human and rodent nervous system. MiR-124 has been reported to regulate neurodevelopment[102], and up-regulation of miR-124 expression level promotes microglial cells to switch from pro-inflammatory M1 type to anti-inflammatory M2 type[103,104]. In neonatal HIE rat model, miR-124-3p directly targets and regulates Bax, one of the pro-apoptotic members of the Bcl-2 family, and subsequently protects neurons from ischemic and hypoxic damage[105]. As previously described, anti-inflammatory miR-124 was found in young microglia and might be able to indirectly affect the proliferation and gliosis of reactive astrocytes, playing a potential therapeutic role in immune-inflammatory neonatal brain injury.
Notch can be split by γ-secretase and release intracellular domains (NICD) into the nucleus to mediate target gene functioning[106]. The study found that the γ-secretase inhibitor DAPT rescued neonatal rats suffering from severe global hypoperfusion or focal ischemia and protected neurons from apoptosis by blocking the Notch signaling pathway[107]. The Notch1 signaling pathway is activated in astrocytes after brain injury. It regulates astrocyte proliferation and activation by inhibiting VEGF or NF-κB/p65 signaling pathways[108]. miR-146a shows aberrant expression or function in neuroinflammatory diseases[109] and indirectly down-regulates pro-inflammatory factors (e.g., IL-1β, IL-6, and TNF-α[110]) as well as promotes oligodendrocyte precursor cells (OPCs) differentiation[111]. In addition, a study transferred miR-146a-5p to astrocyte via extracellular vesicles (EVs) and revealed its ability to inhibit the Notch signaling pathway[112]. It suggests that miR-146a may play a critical role in neurons and glial cells in the immature brain.
3.2. LncRNAs and circRNAs
Increasing evidences suggested that lncRNA H19 is upregulated and promotes inflammatory responses in hypoxic-ischemic brain injury. It is upregulated in the brains of ischemia/reperfusion (I/R) injury rat model and acts as a ceRNA to competitively regulate the miR-138-5p/p65 axis to promote inflammatory responses[113]. Abnormal upregulation of lncRNA H19 was found in an in vitro model of LPS-induced inflammatory response in astrocytes, and H19 knockdown inhibited astrocyte activation and proliferation while decreasing the secretion of pro-inflammatory factors CCL2, IL-6, and TNF-α by regulating the H19/miR-1-3p/CCL2 axis[114]. Aberrant expression of H19 is also closely related to embryonic development and fetal growth[115]. Furthermore, H19 mediates proliferation and autophagy in astrocytes and glioma cells by regulating the mTOR/ULK1 signaling pathway[116]. LncRNA colorectal neoplasia differentially expressed (lncRNA CRNDE) is one of the highest expressed lncRNAs during neuronal differentiation. It was found upregulated in an intrauterine infection-induced neonatal rat brain injury model. Intraperitoneal injection of CRNDE shRNA inhibited LPS-induced activation of astrocytes and microglia, and an improvement in spatial learning memory ability was observed in neonatal rats, demonstrating the neuroprotective effect of lncRNA CRNDE inhibition[117].
CircRNAs perform important roles in regulating central nervous system inflammation. The study observes many brain-enriched circRNAs associated with neurotransmitter function, neuronal maturation, and synaptogenesis[65]. Sigma receptors (σR) are unique drug-binding proteins found in the central nervous system[118], which are located in brain regions involved in motor function, including dopaminergic projections from the substantia nigra to the striatum[119]. The σR 1 agonist DTG reverses behavioral and neuromorphological changes induced by ventral hippocampal lesions in neonatal rats[120]. It was found that circRNA HIPK2 (circ HIPK2) acts as a sponge of miR124-2HG targeting to regulate σR 1 expression and astrocyte activation, which has been shown to attenuate neuroinflammation through the cooperation of cellular autophagy and ER stress[121]. In addition, circ HIPK3 can also function as a sponge for miR124-2HG to regulate cell growth[122].
Similar to neonatal brain injury, both disruption of BBB integrity and astrocyte activation modulating neuroinflammation are major pathologic processes in the secondary response after traumatic brain injury (TBI)[123]. Upregulation of circRNA chr8_87,859,283-87,904,548 (circRNA-chr-8) in the cerebral cortex of a mice TBI model promotes neuroinflammation and disrupts neuronal functional recovery by sponging the miRNA mmu-let-7a-5p to increase the expression of CXCR2[124]. A study found that circRNA METTL9 directly binds to and increases SND1 expression in astrocytes, leading to upregulation of chemokines (CCL2, CXCL1, CCL3, CXCL3, CXCL10) and ultimately enhancing neuroinflammation[125]. (Table 2)
So far, we have exemplified and discussed ncRNAs involved in astrocytes activation and their possible or validated roles in neonates (Figure 1), They assist astrocytes in hypoxia, inflammatory cascades, cell proliferation, autophagy apoptosis, and differentiation fate. Of interest, some ncRNAs are highly conserved whereas the action of a subset of ncRNAs is age-dependent. Their unstable expression becomes a noteworthy difficulty in further research.
4. Regulatory Effect of Non-Coding RNAs in Astrocytes for Treatment Strategy
As mentioned earlier, reactive astrocytes activated under stress conditions have a dual function that requires a high degree of control: On the one hand, it protects the nervous system after injury in terms of intercellular communication[133], homeostatic regulation[134], water transport[19], maintenance of BBB integrity, release of antioxidant substances[135], etc. On the other hand, it exacerbates its heterogeneous function[136], inhibits axonal regeneration by excessive formation of glial scarring[137], and promotes an increase in the levels of pro-inflammatory factors[138], which inevitably leads to neuroinflammatory toxicity. The dual role of astrocytes can bring about different therapeutic strategies. The question is how to specifically and effectively target and regulate astrocytes in a treatment-favorable direction, and how to ensure the safety of the approach.
4.1. Cell Therapy: Potential in Treatment Targeting Astrocyte
As early as the end of the last century, researchers inserted Millipore implants coated with embryonic astrocytes into neuromas of young acallosal animals and found that the implants supported the migration of immature astrocytes and, in turn, the de novo growth of commissural axons between the hemispheres. In addition, implantation after a critical period when the glial response no longer promotes axonal growth reduces host glial scarring, inhibits massive hemorrhage and secondary necrosis, and promotes axonal regeneration[139,140]. Transplantation of astrocytes after in vitro intervention (silencing, gene correction, gene overexpression, etc.) has become a new therapeutic strategy. In global cerebral ischemic rat model, Cyclin-dependent kinase 5 (CDK5) microRNA knock-down (CDK5-KD) was employed into astrocytes, and transplantation of CDK5-KD astrocytes induce neuroprotection and rescue cognitive and motor impairment after ischemia[141,142]. The combination of the Mokola-G envelope pseudotype, glutamine synthetase promoter, and two distinct microRNA target sequences also provides a powerful tool for CNS astrocyte-specific silencing, e.g., efficient astrocyte-specific gene silencing can be performed by the combined effect of miR9*T-miR124T de-targeting[143].
It is found that transplantation of astrocyte precursors or stem cells to derive astrocytes was able to maximize neuroprotection and functional recovery by promoting the growth of broken axons through regeneration, modulating host immune responses, and providing neurotrophic factors[144]. Stem cell transplantation is a novel and long-lasting treatment for newborns. The type of stem cell, the infusion dose, the mode of administration, and the duration of treatment all require further study. It was found that the intranasal administration of umbilical cord blood stem cells or amniotic fluid stem cells during the acute phase of hypoxia, in combination with hypothermia treatment[145,146], which had a long-lasting neuroprotective effect in HIE mice. Cell therapy still has limitations in maintaining long-term cellular recovery and regenerative capacity as well as safe use in humans, but partial successes in adults and models may provide a profile for astrocyte-targeted therapy in neonatal brain injury.
4.2. The Use of miRNAs in Nanotherapeutics
EVs of various sizes, namely microvesicles (ranging from 100–1000 nm) or exosomes (with sizes between 30–150 nm) are released by most eukaryotic cells[147]. Evidences suggested that therapeutic effectiveness of pluripotent stem cell might be a consequence of the paracrine action of EVs[148]. EVs are proved capable of encapsulating mRNAs, miRNAs, proteins, and liposomes, which regulate gene expression and cellular function in recipient cells[149]. EVs have a therapeutic advantage over intact stem cells due to their small size and ability to infiltrate the blood-brain barrier [150].
The study showed that mesenchymal stem cell-derived exosomes (MSC-Exo) could reverse hippocampal astrocyte oxidation and inflammation phenotypes, and attenuate the inflammation-induced astrocytic activation by regulating the Nrf2-NF-κB signaling pathway[151]. Transformation of cellular phenotypes by EVs can act through transported and delivered miRNAs. MiR-21a-5p, highly expressed in MSCs-EVs[152], was found to improve sensory and motor functions after cerebral hemorrhage in rats by affecting the expression of the target gene Trpm7[153]. It alleviates HI brain injury in neonatal mice by targeting STAT3 to induce microglia M2 polarization[154]. In addition, miR-125b-5p delivered by MSC-EVs plays an indispensable role in suppressing astrocyte activation and associated inflammation and maintaining BBB integrity by targeting TLR4 and inhibiting NF-κB signaling in astrocytes[155]. MiR-124 was shown to be neuroprotective and to induce astrocyte-to-neuron transition. MiR-124-3p in astrocyte-derived extracellular vesicles (ADEVs) inhibits aberrant activation of the immune system to rescue neonatal HIBD[156]. This provides a powerful and reliable strategy for the treatment of neonatal brain injury.
4.3. The Neurogenic Potential of Astrocytes Contributes to Nerve after Brain Injury
There are growing evidences that parenchymal astrocytes have potential neurogenic capacity, implying that they may be able to be reprogrammed into functional neurons[157,158]. In inflammatory conditions or injury, astrocytes isolated from the cortex produce multipotent neurospheres[159]. Reactive astrocytes regain more neural progenitor cells (NPC)-like immature properties[160]. This manifestation in neonatal immature astrocytes is early proof of their neurogenic potential: Astrocytes from embryonic or neonatal brains can form multipotent neurospheres[160], forced expression of neurogenic transcription factors (i.e., Ngn2, Pax6, Mash1) are able to convert postnatal astrocytes into functional neurogenic[161,162]. Transformation of astrocytes into functional induced neurons (iNs) has been well established in vitro[163], microRNAs regulate neurogenic facilitators post-transcriptionally and can be used to complement or replace transcription factors (TFs) in neuronal reprogramming[164]. Neurogenic miRNA-124 has potent potency to direct astrocyte cell fate switches[165]. MiR-124 directly targets ZFP family member Zfp36L1 and unlocks the endogenous cortical neurogenesis pathway. In vivo, miR-124 induces reprogramming of reactive astrocytes into immature iNs capable of long-term survival, especially in coordination with ISX9[166]. The ability of miRNAs to trigger the reprogramming of reactive astrocytes also provides a therapeutic strategy for neural repair after neonatal and adult brain injury—to improve functional maturation of iNs after injury, inflammation condition, or neurodegeneration
5. Conclusions
Non-coding RNAs and astrocytes no longer serve as “margins” for cellular physiopathological processes for the past few years. As neurological research advances,It becomes promising for the development/immature brain to therapeutic use ncRNAs to transform the state of astrocytes. This review summarizes and discusses the mechanisms and therapeutic role of non-coding RNAs in reactive astrocytes of nervous system disorders, which may provide further research directions and new potential therapeutic strategies for neonatal brain injury.
However, due to the “gulf” between the mature or aging brain and the immature brain in response to injury, whether these molecular mechanisms actually work in neonatal brain injury is still open to debate and further research. Points are worth noting, 1) The nervous system is an intricate network of regulatory relationships. As current research indicates, non-coding RNAs are involved in multiple complex regulatory networks, making it difficult to specifically regulate particular cells or molecules in the nervous system, which poses a challenge for targeted therapeutic strategies that utilize these agents. 2) High heterogeneity of astrocytes[167], complexity and spatiotemporal specificity of ncRNA regulatory networks[168], and uncertainty about the critical period of treatment of neonatal brain injury constitute formidable obstacles to the targeting astrocytes for participation in neuroprotective therapies using ncRNAs. 3) The reliance on EVs carrying regulatory factors (e.g., MSC-EVs) to enter the nervous system to exert immunomodulatory, neuroprotective, and regenerative effects is well established in rodent models and holds promise as an emerging therapeutic tool for neonatal brain injury[169]. How to conjugate ncRNAs with targeted regulatory effects and translate them into therapeutic products that are safe for newborns deserves further research and exploration.
Author Contributions
Conceptualization, Zhiyi Y. and Tianming Y.; writing—original draft preparation, Zhiyi Y.; writing—review and editing, Tianming Y.; supervision, Tianming Y.; funding acquisition, Tianming Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grants from the National Natural Science Foundation of China, grant number 81571466
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perin, J.; Mulick, A.; Yeung, D.; Villavicencio, F.; Lopez, G.; Strong, K.L.; Prieto-Merino, D.; Cousens, S.; Black, R.E.; Liu, L. Global, regional, and national causes of under-5 mortality in 2000-19: an updated systematic analysis with implications for the Sustainable Development Goals. Lancet Child Adolesc. Health 2022, 6, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Amer, R.; Moddemann, D.; Seshia, M.; Alvaro, R.; Synnes, A.; Lee, K.-S.; Lee, S.K.; Shah, P.S.; Canadian Neonatal Network, C. Neurodevelopmental Outcomes of Infants Born at < 29 Weeks of Gestation Admitted to Canadian Neonatal Intensive Care Units Based on Location of Birth. J. Pediatr. 2018, 196, 31–+. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, X.; Chen, X.; Wei, Y. Neuronal injuries in cerebral infarction and ischemic stroke: From mechanisms to treatment (Review). Int. J. Mol. Med. 2022, 49. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Urzua, S.; Rojas, I.; Libano, L.; Rodrigo, R. Pathophysiology of Ischemic Stroke: Role of Oxidative Stress. Curr. Pharm. Des. 2020, 26, 4246–4260. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.D.; McCullough, L.D. Inflammatory responses in hypoxic ischemic encephalopathy. Acta Pharmacol. Sin. 2013, 34, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Kulesh, A.; Drobakha, V.; Kuklina, E.; Nekrasova, I.; Shestakov, V. Cytokine Response, Tract-Specific Fractional Anisotropy, and Brain Morphometry in Post-Stroke Cognitive Impairment. J. Stroke Cerebrovasc. Dis. 2018, 27, 1752–1759. [Google Scholar] [CrossRef]
- Urday, S.; Kimberly, W.T.; Beslow, L.A.; Vortmeyer, A.O.; Selim, M.H.; Rosand, J.; Simard, J.M.; Sheth, K.N. Targeting secondary injury in intracerebral haemorrhage-perihaematomal oedema. Nat. Rev. Neurol. 2015, 11, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.R. Specification and Morphogenesis of Astrocytes. Science 2010, 330, 774–778. [Google Scholar] [CrossRef]
- Disdier, C.; Stonestreet, B.S. Hypoxic-ischemic-related cerebrovascular changes and potential therapeutic strategies in the neonatal brain. J. Neurosci. Res. 2020, 98, 1468–1484. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the neurovascular unit: key functions and signaling pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Astroglial cradle in the life of the synapse. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Mari, C.; Odorcyk, F.K.; Sanches, E.F.; Wartchow, K.M.; Martini, A.P.; Nicola, F.; Zanotto, C.; Wyse, A.T.; Gonçalves, C.A.; Netto, C.A. Arundic acid administration protects astrocytes, recovers histological damage and memory deficits induced by neonatal hypoxia ischemia in rats. Int. J. Dev. Neurosci. 2019, 76, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.; Farina, C. Astrocytes: Key Regulators of Neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.P.; Fang, G.; Rozowsky, J.; Snyder, M.; Gerstein, M.B. Annotating non-coding regions of the genome. Nat. Rev. Genet. 2010, 11, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Marangon, D.; Castro, E.S.J.H.; Lecca, D. Neuronal and Glial Communication via Non-Coding RNAs: Messages in Extracellular Vesicles. Int. J. Mol. Sci. 2022, 24. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Escobar, M.; Weiss, M.D. Hypoxic-ischemic encephalopathy: a review for the clinician. JAMA Pediatr. 2015, 169, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Huo, J. A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem. Int. 2021, 148, 105080. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; MacDonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784. [Google Scholar] [CrossRef]
- Salman, M.M.; Kitchen, P.; Halsey, A.; Wang, M.X.; Tornroth-Horsefield, S.; Conner, A.C.; Badaut, J.; Iliff, J.J.; Bill, R.M. Emerging roles for dynamic aquaporin-4 subcellular relocalization in CNS water homeostasis. Brain 2022, 145, 64–75. [Google Scholar] [CrossRef]
- Fu, X.; Li, Q.; Feng, Z.; Mu, D. The roles of aquaporin-4 in brain edema following neonatal hypoxia ischemia and reoxygenation in a cultured rat astrocyte model. Glia 2007, 55, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Sepramaniam, S.; Armugam, A.; Lim, K.Y.; Karolina, D.S.; Swaminathan, P.; Tan, J.R.; Jeyaseelan, K. MicroRNA 320a Functions as a Novel Endogenous Modulator of Aquaporins 1 and 4 as Well as a Potential Therapeutic Target in Cerebral Ischemia. J. Biol. Chem. 2010, 285, 29223–29230. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-C.; Li, W.-J.; Li, L.-Z. Regulatory effect of miRNA 320a on expression of aquaporin 4 in brain tissue of epileptic rats. Asian Pac. J. Trop. Med. 2015, 8, 791–796. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, L.; Chen, M.; Pei, A.; Xie, L.; Zhu, S. Upregulation of miR-130b protects against cerebral ischemic injury by targeting water channel protein aquaporin 4 (AQP4). Am. J. Transl. Res. 2017, 9, 3452–3461. [Google Scholar] [PubMed]
- Zheng, L.F.; Cheng, W.; Wang, X.J.; Yang, Z.G.; Zhou, X.L.; Pan, C.L. Overexpression of MicroRNA-145 Ameliorates Astrocyte Injury by Targeting Aquaporin 4 in Cerebral Ischemic Stroke. Biomed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.-N.; Sun, X.-L.; Gao, L.; Fan, Y.; Ding, J.-H.; Hu, G. Aquaporin-4 deficiency down-regulates glutamate uptake and GLT-1 expression in astrocytes. Mol. Cell. Neurosci. 2007, 34, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8. [Google Scholar] [CrossRef]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The role of astrocytic glutamate transporters GLT-1 and GLAST in neurological disorders: Potential targets for neurotherapeutics. Neuropharmacology 2019, 161. [Google Scholar] [CrossRef]
- Morken, T.S.; Brekke, E.; Håberg, A.; Widerøe, M.; Brubakk, A.M.; Sonnewald, U. Altered astrocyte-neuronal interactions after hypoxia-ischemia in the neonatal brain in female and male rats. Stroke 2014, 45, 2777–2785. [Google Scholar] [CrossRef]
- Benediktsson, A.M.; Marrs, G.S.; Tu, J.C.; Worley, P.F.; Roth-Stein, J.D.; Bergles, D.E.; Dailey, M.E. Neuronal activity regulates glutamate transporter dynamics in developing astrocytes. Glia 2012, 60, 175–188. [Google Scholar] [CrossRef]
- Johnston, M.V.; Fatemi, A.; Wilson, M.A.; Northington, F. Treatment advances in neonatal neuroprotection and neurointensive care. Lancet Neurol. 2011, 10, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhou, H.; Zhao, Q.; Xue, L.; Al-Hawwas, M.; He, J.; Wu, M.; Zou, Y.; Yang, M.; Dai, J.; et al. Overexpression of miR-124 Protects Against Neurological Dysfunction Induced by Neonatal Hypoxic-Ischemic Brain Injury. Cell. Mol. Neurobiol. 2020, 40, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-Y.; Jiang, C.; Ye, H.-B.; Jiao, J.-T.; Cheng, C.; Huang, J.; Liu, J.; Zhang, R.; Shao, J.-F. miR-124 upregulates astrocytic glutamate transporter-1 via the Akt and mTOR signaling pathway post ischemic stroke. Brain Res. Bull. 2019, 149, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Hoye, M.L.; Regan, M.R.; Jensen, L.A.; Lake, A.M.; Reddy, L.V.; Vidensky, S.; Richard, J.P.; Maragakis, N.J.; Rothstein, J.D.; Dougherty, J.D.; et al. Motor neuron-derived microRNAs cause astrocyte dysfunction in amyotrophic lateral sclerosis. Brain 2018, 141, 2561–2575. [Google Scholar] [CrossRef] [PubMed]
- Brkic, J.; Dunk, C.; O'Brien, J.; Fu, G.; Nadeem, L.; Wang, Y.-l.; Rosman, D.; Salem, M.; Shynlova, O.; Yougbare, I.; et al. MicroRNA-218-5p Promotes Endovascular Trophoblast Differentiation and Spiral Artery Remodeling. Mol. Ther. 2018, 26, 2189–2205. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Mei, L.; Xie, L. The Clinical Features of 101 Cases with Early-Onset Severe Preeclampsia. J. Pract. Obstet. Gynecol. 2013, 29, 63–66. [Google Scholar]
- Mo, Y.; Sun, Y.-Y.; Liu, K.-Y. Autophagy and inflammation in ischemic stroke. Neural Regen. Res. 2020, 15, 1388–1396. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Z.; Liu, Y.; Pan, S.; Zhang, H.; Fang, M.; Jiang, H.; Yin, J.; Zou, S.; Li, Z.; et al. Melatonin reduces hypoxic-ischaemic (HI) induced autophagy and apoptosis: An in vivo and in vitro investigation in experimental models of neonatal HI brain injury. Neurosci. Lett. 2017, 653, 105–112. [Google Scholar] [CrossRef]
- Mao, L.; Liu, S.; Hu, L.; Jia, L.; Wang, H.; Guo, M.; Chen, C.; Liu, Y.; Xu, L. miR-30 Family: A Promising Regulator in Development and Disease. Biomed Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Chang, T.-C.; Yu, D.; Lee, Y.-S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microRNA repression by Myc contributes to tumorigenesis. Nat. Genet. 2008, 40, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, W.Q.; Zhu, H.; Qian, Y.Y.; Zhou, L.; Ren, Y.J.; Ren, X.C.; Zhang, L.; Liu, X.P.; Liu, C.G.; et al. Regulation of autophagy by miR-30d impacts sensitivity of anaplastic thyroid carcinoma to cisplatin. Biochem. Pharmacol. 2014, 87, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Qu, Y.; Wang, H.; Huang, L.; Zhu, J.; Li, S.; Tong, Y.; Zhang, L.; Li, J.; Mu, D. The effect of miR-30d on apoptosis and autophagy in cultured astrocytes under oxygen-glucose deprivation. Brain Res. 2017, 1671, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Prerna, K.; Dubey, V.K. Beclin1-mediated interplay between autophagy and apoptosis: New understanding. Int. J. Biol. Macromol. 2022, 204, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Qu, Y.; Zhu, J.; Zhang, L.; Huang, L.; Liu, H.; Li, S.; Mu, D. miR-30d-5p Plays an Important Role in Autophagy and Apoptosis in Developing Rat Brains After Hypoxic-Ischemic Injury. J. Neuropathol. Exp. Neurol. 2017, 76, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Wang, S.; Yin, Y.; Yu, J.; Liu, Y.; Gao, H. Dexmedetomidine suppresses hippocampal astrocyte pyroptosis in cerebral hypoxic-ischemic neonatal rats by upregulating microRNA-148a-3p to inactivate the STAT/JMJD3 axis. Int. Immunopharmacol. 2023, 121, 110440. [Google Scholar] [CrossRef] [PubMed]
- Gaughwin, P.; Ciesla, M.; Yang, H.; Lim, B.; Brundin, P. Stage-Specific Modulation of Cortical Neuronal Development by Mmu-miR-134. Cereb. Cortex 2011, 21, 1857–1869. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, S.; Kadkhoda, S.; Ghafouri-Fard, S. Synaptic plasticity and depression: the role of miRNAs dysregulation. Mol. Biol. Rep. 2022, 49, 9759–9765. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.; Larsen, L.A.; Kauppinen, S.; Schratt, G. Recombinant adeno-associated virus-mediated microRNA delivery into the postnatal mouse brain reveals a role for miR-134 in dendritogenesis in vivo. Front. Neural Circuits 2010, 3. [Google Scholar] [CrossRef]
- Chen, W.B.; Zhang, L.X.; Zhao, Y.K.; Li, J.; Jiao, Y. C/EBP alpha-mediated transcriptional activation of miR-134-5p entails KPNA3 inhibition and modulates focal hypoxic-ischemic brain damage in neonatal rats. Brain Res. Bull. 2020, 164, 350–360. [Google Scholar] [CrossRef]
- Guan, X.H.; Zhou, W.; Li, L.; Peng, Q.X. Dexmedetomidine Alleviates Hypoxic-Ischemic Brain Damage in Neonatal Rats Through Reducing MicroRNA-134-5p-Mediated NLRX1 Downregulation. J. Stroke Cerebrovasc. Dis. 2022, 31. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Nakajima, S.; Yamamoto, N.; Ooshima, Y.; Odaka, H.; Hashido, K.; Adachi, N.; Kunugi, H. Basic fibroblast growth factor induces miR-134 upregulation in astrocyte for cell maturation. Biochem. Biophys. Res. Commun. 2015, 456, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Haemmerle, M.; Eissmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Gross, M.; et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zoernig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long Noncoding RNA MALAT1 Regulates Endothelial Cell Function and Vessel Growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, X.; Jin, J.; Zheng, L.; Guan, T.; Huo, Y.; Xie, S.; Wu, Y.; Chen, W. LncRNA MALAT1 silencing protects against cerebral ischemia-reperfusion injury through miR-145 to regulate AQP4. J. Biomed. Sci. 2020, 27. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Wang, Q.; Mao, Y.; Cao, Q.; Li, S.; Qiao, C.; Zhang, D.; Zhou, G. Knockdown of LncRNA MALAT1 contributes to cell apoptosis via regulating NF-kappa B/CD80 axis in neonatal respiratory distress syndrome. Int. J. Biochem. Cell Biol. 2018, 104, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wu, J.; Yu, X.; Zhou, J.; Yu, H.; Ma, L. Long non-coding RNA MALAT1 enhances the apoptosis of cardiomyocytes through autophagy inhibition by regulating TSC2-mTOR signaling. Biol. Res. 2019, 52. [Google Scholar] [CrossRef] [PubMed]
- Pachnis, V.; Belayew, A.; Tilghman, S.M. LOCUS UNLINKED TO ALPHA-FETOPROTEIN UNDER THE CONTROL OF THE MURINE RAF AND RIF GENES. Proc. Natl. Acad. Sci. USA 1984, 81, 5523–5527. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Esmaeili, M.; Taheri, M. H19 lncRNA: Roles in tumorigenesis. Biomed. Pharmacother. 2020, 123. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, L.; Chen, J.; Shen, H.; Chen, Z. Mechanisms Underlying Abnormal Expression of lncRNA H19 in Neonatal Hypoxic-Ischemic Encephalopathy. Am. J. Perinatol. 2022, 39, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, B.; Gao, Y.; Chen, Y.H.; Feng, J. Exosome-transported lncRNA H19 regulates insulin-like growth factor-1 via the H19/let-7a/insulin-like growth factor-1 receptor axis in ischemic stroke. Neural Regen Res. 2023, 18, 1316–1320. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Zhu, X.; Zhuo, C. H19/miR-130a-3p/DAPK1 axis regulates the pathophysiology of neonatal hypoxic-ischemia encephalopathy. Neurosci. Res. 2021, 163, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zou, Q.; Lv, D.; Raza, M.A.; Wang, X.; Li, P.; Chen, Y.; Xi, X.; Wen, A.; Zhu, L.; et al. Comprehensive transcriptional profiling of porcine brain aging. Gene 2019, 693, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, E.; Cairns, M.J. Circular RNAs are temporospatially regulated throughout development and ageing in the rat. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, H.J.; Fan, Z.M.; Zhao, R.B.; Xia, Z.K. Circular RNA expression profiles in neonatal rats following hypoxic-ischemic brain damage. Int. J. Mol. Med. 2019, 43, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Zhang, Y.; Zhang, Y.; Bai, Y.; Chen, X.; Huang, R.; Wu, F.; Leng, S.; Chao, J.; Zhang, J.H.; et al. Novel insight into circular RNA <i>HECTD1</i> in astrocyte activation via autophagy by targeting <i>MIR142</i>-TIPARP: implications for cerebral ischemic stroke. Autophagy 2018, 14, 1164–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Huang, Z.; Zhu, X.; Hong, T.; Zhao, Y. Circular RNA 0025984 Ameliorates Ischemic Stroke Injury and Protects Astrocytes Through miR-143-3p/TET1/ORP150 Pathway. Mol. Neurobiol. 2021, 58, 5937–5953. [Google Scholar] [CrossRef]
- Guo, X.X.; He, Q.Z.; Li, W.; Long, D.X.; Pan, X.Y.; Chen, C.; Zeng, H.C. Brain-Derived Neurotrophic Factor Mediated Perfluorooctane Sulfonate Induced-Neurotoxicity via Epigenetics Regulation in SK-N-SH Cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Meng, X.; Zhong, J.; Zeng, C.; Yung, K.K.L.; Zhang, X.; Wu, X.; Qu, S. MiR-30a-5p Regulates GLT-1 Function via a PKCα-Mediated Ubiquitin Degradation Pathway in a Mouse Model of Parkinson's Disease. ACS Chem. Neurosci. 2021, 12, 1578–1592. [Google Scholar] [CrossRef]
- Reichenstein, I.; Eitan, C.; Diaz-Garcia, S.; Haim, G.; Magen, I.; Siany, A.; Hoye, M.L.; Rivkin, N.; Olender, T.; Toth, B.; et al. Human genetics and neuropathology suggest a link between miR-218 and amyotrophic lateral sclerosis pathophysiology. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, M. Astrocyte-derived extracellular vesicles inhibit the abnormal activation of immune function in neonatal mice with hypoxic-ischemic brain damage by carrying miR-124-3p. Neurol. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, B.; Sun, S.; Wang, B. Overexpression of long non-coding RNA H19 relieves hypoxia-induced injury by down-regulating microRNA-107 in neural stem cells. Neurosci. Lett. 2021, 753. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, G.; Sabato, C.; Russo, M.; Rosa, A.; Abballe, L.; Besharat, Z.M.; Po, A.; Miele, E.; Bellavia, D.; Chiacchiarini, M.; et al. Loss of miR-107, miR-181c and miR-29a-3p Promote Activation of Notch2 Signaling in Pediatric High-Grade Gliomas (pHGGs). Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-B.; Luo, X.-J.; Ren, K.-D.; Peng, J.-J.; Tan, B.; Liu, B.; Lou, Z.; Xiong, X.-M.; Zhang, X.-J.; Ren, X.; et al. Beneficial effect of magnesium lithospermate B on cerebral ischemia-reperfusion injury in rats involves the regulation of miR-107/glutamate transporter 1 pathway. Eur. J. Pharmacol. 2015, 766, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Y.; Wang, L.Q.; Chen, M.L.; Pei, A.J.; Xie, L.W.; Zhu, S.M. Upregulation of miR-130b protects against cerebral ischemic injury by targeting water channel protein aquaporin 4 (AQP4). Am. J. Transl. Res. 2017, 9, 3452–3461. [Google Scholar]
- Winkler, I.; Heisinger, T.; Hammerl, M.; Huber, E.; Urbanek, M.; Kiechl-Kohlendorfer, U.; Griesmaier, E.; Posod, A. MicroRNA Expression Profiles as Diagnostic and Prognostic Biomarkers of Perinatal Asphyxia and Hypoxic-Ischaemic Encephalopathy. Neonatology 2022, 119, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimy, N.; Gasterich, N.; Behrens, V.; Amini, J.; Fragoulis, A.; Beyer, C.; Zhao, W.Y.; Sanadgol, N.; Zendedel, A. Neuroprotective effect of the Nrf2/ARE/miRNA145-5p signaling pathway in the early phase of spinal cord injury. Life Sci. 2022, 304. [Google Scholar] [CrossRef]
- Zhao, H.P.; Li, G.W.; Wang, R.L.; Tao, Z.; Zhang, S.J.; Li, F.F.; Han, Z.P.; Li, L.Z.; Liu, P.; Luo, Y.M. MiR-424 prevents astrogliosis after cerebral ischemia/reperfusion in elderly mice by enhancing repressive H3K27me3 via NFIA/DNMT1 signaling. Febs J. 2019, 286, 4926–4936. [Google Scholar] [CrossRef]
- Zhang, L.; Bai, X.; Yan, W. LncRNA-MALAT1, as a biomarker of neonatal BPD, exacerbates the pathogenesis of BPD by targeting miR-206. Am. J. Transl. Res. 2021, 13, 462–479. [Google Scholar]
- Jin, R.; Yang, G.; Li, G. Inflammatory mechanisms in ischemic stroke: role of inflammatory cells. J. Leukoc. Biol. 2010, 87, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Yap, V.; Perlman, J.M. Mechanisms of brain injury in newborn infants associated with the fetal inflammatory response syndrome. Semin. Fetal Neonatal Med. 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Galinsky, R.; Polglase, G.R.; Hooper, S.B.; Black, M.J.; Moss, T.J.M. The consequences of chorioamnionitis: preterm birth and effects on development. J. Pregnancy 2013, 2013, 412831–412831. [Google Scholar] [CrossRef] [PubMed]
- Arayici, S.; Simsek, G.K.; Oncel, M.Y.; Eras, Z.; Canpolat, F.E.; Oguz, S.S.; Uras, N.; Zergeroglu, S.; Dilmen, U. The effect of histological chorioamnionitis on the short-term outcome of preterm infants <= 32 weeks: a single-center study. J. Matern. -Fetal Neonatal Med. 2014, 27, 1129–1133. [Google Scholar] [CrossRef]
- Disdier, C.; Awa, F.; Chen, X.; Dhillon, S.K.; Galinsky, R.; Davidson, J.O.; Lear, C.A.; Bennet, L.; Gunn, A.J.; Stonestreet, B.S. Lipopolysaccharide-induced changes in the neurovascular unit in the preterm fetal sheep brain. J. Neuroinflammation 2020, 17. [Google Scholar] [CrossRef]
- Leonardo, C.C.; Eakin, A.K.; Ajmo, J.M.; Collier, L.A.; Pennypacker, K.R.; Strongin, A.Y.; Gottschall, P.E. Delayed administration of a matrix metalloproteinase inhibitor limits progressive brain injury after hypoxia-ischemia in the neonatal rat. J. Neuroinflammation 2008, 5. [Google Scholar] [CrossRef]
- Revuelta, M.; Elicegui, A.; Moreno-Cugnon, L.; Buehrer, C.; Matheu, A.; Schmitz, T. Ischemic stroke in neonatal and adult astrocytes. Mech. Ageing Dev. 2019, 183. [Google Scholar] [CrossRef]
- Colombo, E.; Farina, C. Astrocytes: Key Regulators of Neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef]
- Ye, Y.; Hao, J.; Hong, Z.; Wu, T.; Ge, X.; Qian, B.; Chen, X.; Zhang, F. Downregulation of MicroRNA-145-5p in Activated Microglial Exosomes Promotes Astrocyte Proliferation by Removal of Smad3 Inhibition. Neurochem. Res. 2022, 47, 382–393. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Yang, S.-H.; Tzeng, S.-F. MicroRNA-145 as One Negative Regulator of Astrogliosis. Glia 2015, 63, 194–205. [Google Scholar] [CrossRef]
- Capece, D.; Verzella, D.; Flati, I.; Arboretto, P.; Cornice, J.; Franzoso, G. NF-κB: blending metabolism, immunity, and inflammation. Trends Immunol 2022, 43, 757–775. [Google Scholar] [CrossRef]
- Kang, Z.; Altuntas, C.Z.; Gulen, M.F.; Liu, C.; Giltiay, N.; Qin, H.; Liu, L.; Qian, W.; Ransohoff, R.M.; Bergmann, C.; et al. Astrocyte-Restricted Ablation of Interleukin-17-Induced Act1-Mediated Signaling Ameliorates Autoimmune Encephalomyelitis. Immunity 2010, 32, 414–425. [Google Scholar] [CrossRef]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-kappaB Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, R.; Bracchi-Ricard, V.; Hu, W.H.; Frydel, B.; Bramwell, A.; Karmally, S.; Green, E.J.; Bethea, J.R. Inhibition of astroglial nuclear factor kappa B reduces inflammation and improves functional recovery after spinal cord injury. J. Exp. Med. 2005, 202, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Chibowska, K.; Korbecki, J.; Gutowska, I.; Metryka, E.; Tarnowski, M.; Goschorska, M.; Barczak, K.; Chlubek, D.; Baranowska-Bosiacka, I. Pre- and Neonatal Exposure to Lead (Pb) Induces Neuroinflammation in the Forebrain Cortex, Hippocampus and Cerebellum of Rat Pups. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Lee, H.-G.; Wheeler, M.A.; Quintana, F.J. Function and therapeutic value of astrocytes in neurological diseases. Nat. Rev. Drug Discov. 2022, 21, 339–358. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Li, Q.; Kang, L.; Jia, W.; Yan, Y. MicroRNA-155 intensifies hypoxic-ischemic brain injury in neonatal rats by regulating nucleatide-binding oligomerizaton domains 1 / nuclear facter kappaB signaling pathway. Acta Anat. Sin. 2020, 51, 848–854. [Google Scholar]
- He, J.; Zhao, J.; Peng, X.; Shi, X.; Zong, S.; Zeng, G. Molecular Mechanism of MiR-136-5p Targeting NF-kappa B/A20 in the IL-17-Mediated Inflammatory Response after Spinal Cord Injury. Cell. Physiol. Biochem. 2017, 44, 1224–1241. [Google Scholar] [CrossRef]
- Ma, Q.; Dasgupta, C.; Shen, G.; Li, Y.; Zhang, L. MicroRNA-210 downregulates TET2 and contributes to inflammatory response in neonatal hypoxic-ischemic brain injury. J. Neuroinflammation 2021, 18, 6. [Google Scholar] [CrossRef]
- Jiang, D.; Gong, F.; Ge, X.; Lv, C.; Huang, C.; Feng, S.; Zhou, Z.; Rong, Y.; Wang, J.; Ji, C.; et al. Neuron-derived exosomes-transmitted miR-124-3p protect traumatically injured spinal cord by suppressing the activation of neurotoxic microglia and astrocytes. J. Nanobiotechnology 2020, 18. [Google Scholar] [CrossRef]
- Tian, Y.; Tian, Y.; Tu, Y.; Zhang, G.; Zeng, X.; Lin, J.; Ai, M.; Mao, Z.; Zheng, R.; Yuan, Y. microRNA-124 inhibits stem-like properties and enhances radiosensitivity in nasopharyngeal carcinoma cells via direct repression of expression of JAMA. J. Cell. Mol. Med. 2020, 24, 9533–9544. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.-C.; Pastrana, E.; Tavazoie, M.; Doetsch, F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat. Neurosci. 2009, 12, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Zhang, T.; Duan, H.; Pan, Y.; Zhang, X.; Yang, G.; Wang, J.; Deng, Y.; Yang, Z. MiR-124 contributes to M2 polarization of microglia and confers brain inflammatory protection via the C/EBP-alpha, pathway in intracerebral hemorrhage. Immunol. Lett. 2017, 182, 1–11. [Google Scholar] [CrossRef]
- Periyasamy, P.; Liao, K.; Kook, Y.H.; Niu, F.; Callen, S.E.; Guo, M.-L.; Buch, S. Cocaine-Mediated Downregulation of miR-124 Activates Microglia by Targeting KLF4 and TLR4 Signaling. Mol. Neurobiol. 2018, 55, 3196–3210. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Wang, J.; Luo, F.; He, Y. Role of microRNA-124-3p/Bax axis in neonatal hypoxic-ischaemic encephalopathy. Biotechnol. Biotechnol. Equip. 2020, 34, 163–170. [Google Scholar] [CrossRef]
- Hur, J.Y. gamma-Secretase in Alzheimer's disease. Exp. Mol. Med. 2022, 54, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, J.; Zhao, C.; Ren, K.; Xia, Z.; Yu, H.; Jiang, K. Acute Blockage of Notch Signaling by DAPT Induces Neuroprotection and Neurogenesis in the Neonatal Rat Brain After Stroke. Transl. Stroke Res. 2016, 7, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; He, K.; Wang, F.W.; Li, X.; Liu, D.J. Notch-1 signaling regulates astrocytic proliferation and activation after hypoxia exposure. Neurosci. Lett. 2015, 603, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Aslani, M.; Mortazavi-Jahromi, S.S.; Mirshafiey, A. Efficient roles of miR-146a in cellular and molecular mechanisms of neuroinflammatory disorders: An effectual review in neuroimmunology. Immunol. Lett. 2021, 238, 1–20. [Google Scholar] [CrossRef]
- Zhong, X.; Jiang, Y.Z.; Liu, P.; He, W.; Xiong, Z.; Chang, W.; Zhu, J.; Cui, Q. Toll-like 4 receptor /NFκB inflammatory/miR-146a pathway contributes to the ART-correlated preterm birth outcome. Oncotarget 2016, 7, 72475–72485. [Google Scholar] [CrossRef]
- Liu, X.S.; Chopp, M.; Pan, W.L.; Wang, X.L.; Fan, B.Y.; Zhang, Y.; Kassis, H.; Zhang, R.L.; Zhang, X.M.; Zhang, Z.G. MicroRNA-146a Promotes Oligodendrogenesis in Stroke. Mol. Neurobiol. 2017, 54, 227–237. [Google Scholar] [CrossRef]
- Rigg, E.; Wang, J.; Xue, Z.; Lunavat, T.R.; Liu, G.; Hoang, T.; Parajuli, H.; Han, M.; Bjerkvig, R.; Nazarov, P.V.; et al. Inhibition of extracellular vesicle-derived miR-146a-5p decreases progression of melanoma brain metastasis via Notch pathway dysregulation in astrocytes. J. Extracell Vesicles 2023, 12, e12363. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Wang, D. LncRNA H19 promotes inflammatory response induced by cerebral ischemia-reperfusion injury through regulating the miR-138-5p-p65 axis. Biochem Cell Biol. 2020, 98, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.; Dai, Y.; Wang, B.; Li, L.; Jiang, B.; Wu, P.; Xu, J. The LncRNA H19/miR-1-3p/CCL2 axis modulates lipopolysaccharide (LPS) stimulation-induced normal human astrocyte proliferation and activation. Cytokine 2020, 131. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z. Igf2-H19, an imprinted tandem gene, is an important regulator of embryonic development, a guardian of proliferation of adult pluripotent stem cells, a regulator of longevity, and a 'passkey' to cancerogenesis. Folia Histochem. Et Cytobiol. 2012, 50, 171–179. [Google Scholar] [CrossRef]
- Zhao, W.; Lin, X.Y.; Han, H.; Zhang, H.X.; Li, X.L.; Jiang, C.M.; Feng, M. Long noncoding RNA H19 contributes to the proliferation and autophagy of glioma cells through mTOR/ULK1 pathway. Neuroreport 2021, 32, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.H.; Zhang, B.H.; Fang, C.Z.; Yan, C.X.; Lai, F.F.; Chen, S.; Wang, G.H. Long non-coding RNA CRNDE deteriorates intrauterine infection-induced neonatal brain injury. Mol. Cell Probes. 2020, 52, 101565. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.L.; Hiranita, T.; Hong, W.C.; Job, M.O.; McCurdy, C.R. A Role for Sigma Receptors in Stimulant Self-Administration and Addiction. In Sigma Proteins: Evolution of the Concept of Sigma Receptors; Handbook of Experimental Pharmacology; Kim, F.J., Pasternak, G.W., Eds.; 2017; Volume 244, pp. 177–218. [Google Scholar]
- Martin, P.; Reeder, T.; Sourbron, J.; de Witte, P.A.M.; Gammaitoni, A.R.; Galer, B.S. An Emerging Role for Sigma-1 Receptors in the Treatment of Developmental and Epileptic Encephalopathies. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo-Loranca, B.E.; Garces-Ramirez, L.; Rosales, A.A.M.; Ramirez, C.L.; Hernandez, G.V.; Morales-Dionisio, O.; Gonzalez-Elizalde, K.; Flores, G.; Zamudio, S.; De la Cruz-Lopez, F. The Sigma Agonist 1,3-Di-o-tolyl-guanidine Reduces the Morphological and Behavioral Changes Induced by Neonatal Ventral Hippocampus Lesion in Rats. Synapse 2015, 69, 213–225. [Google Scholar] [CrossRef]
- Huang, R.R.; Zhang, Y.; Han, B.; Bai, Y.; Zhou, R.B.; Gan, G.M.; Chao, J.; Hu, G.; Yao, H.H. Circular RNA HIPK2 regulates astrocyte activation via cooperation of autophagy and ER stress by targeting MIR124-2HG. Autophagy 2017, 13, 1722–1741. [Google Scholar] [CrossRef]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yan, X.F.; Si, Y.; Chen, X.Z. CTGF Triggers Rat Astrocyte Activation and Astrocyte-Mediated Inflammatory Response in Culture Conditions. Inflammation 2019, 42, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, H.; Zhong, J.; Yang, J.; Darwazeh, R.; Tian, X.; Huang, Z.; Jiang, L.; Cheng, C.; Wu, Y.; et al. Significant changes in circular RNA in the mouse cerebral cortex around an injury site after traumatic brain injury. Exp. Neurol. 2019, 313, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.L.; Sun, L.L.; Xiao, C.Y.; You, W.J.; Sun, L.; Wang, S.Y.; Zhang, Z.J.; Liu, S. Circular RNA METTL9 contributes to neuroinflammation following traumatic brain injury by complexing with astrocytic SND1. J. Neuroinflammation 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Zurolo, E.; Prabowo, A.; Fluiter, K.; Spliet, W.G.M.; van Rijen, P.C.; Gorter, J.A.; Aronica, E. MicroRNA-146a: A Key Regulator of Astrocyte-Mediated Inflammatory Response. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Gomes, C.; Sequeira, C.; Gonçalves-Ribeiro, J.; Pina, C.C.; Carvalho, L.A.; Moreira, R.; Vaz, S.H.; Vaz, A.R.; Brites, D. Recovery of Depleted miR-146a in ALS Cortical Astrocytes Reverts Cell Aberrancies and Prevents Paracrine Pathogenicity on Microglia and Motor Neurons. Front. Cell Dev. Biol. 2021, 09. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Chang, Q.; Zhang, Y.; Guan, X.; Ma, Z.; Chen, X.; Liu, W.; Li, Y.; Feng, H. MiR-155-5p Aggravated Astrocyte Activation and Glial Scarring in a Spinal Cord Injury Model by Inhibiting Ndfip1 Expression and PTEN Nuclear Translocation. Neurochem. Res. 2023, 48, 1912–1924. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.Y.; Gao, Y.B.; Cen, Z.X.; He, J.C.; Cao, B.C.; Zeng, G.F.; Zong, S.H. miR-136-5p Regulates the Inflammatory Response by Targeting the IKK/NF-B/A20 Pathway After Spinal Cord Injury. Cell. Physiol. Biochem. 2018, 50, 512–524. [Google Scholar] [CrossRef]
- Fu, C.H.; Lai, F.F.; Chen, S.; Yan, C.X.; Zhang, B.H.; Fang, C.Z.; Wang, G.H. Silencing of long non-coding RNA CRNDE promotes autophagy and alleviates neonatal hypoxic-ischemic brain damage in rats. Mol. Cell Biochem. 2020, 472, 1–8. [Google Scholar] [CrossRef]
- Huang, R.; Cai, L.; Ma, X.; Shen, K. Autophagy-mediated circHIPK2 promotes lipopolysaccharide-induced astrocytic inflammation via SIGMAR1. Int. Immunopharmacol. 2023, 117. [Google Scholar] [CrossRef]
- Chen, Z.H.; Wang, H.; Zhong, J.J.; Yang, J.Q.; Darwazeh, R.; Tian, X.C.; Huang, Z.J.; Jiang, L.; Cheng, C.J.; Wu, Y.; et al. Significant changes in circular RNA in the mouse cerebral cortex around an injury site after traumatic brain injury. Exp. Neurol. 2019, 313, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Dallerac, G.; Chever, O.; Rouach, N. How do astrocytes shape synaptic transmission? Insights from electrophysiology. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Xu, D.; Luo, Y.; Li, X.; Gu, Y.; Wang, L. Homeostatic Regulation of Astrocytes by Visual Experience in the Developing Primary Visual Cortex. Cereb Cortex 2022, 32, 970–986. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, X.; Chen, L.; Lenahan, C.; Fu, Z.; Fang, Y.; Yu, W. Crosstalk Between the Oxidative Stress and Glia Cells After Stroke: From Mechanism to Therapies. Front. Immunol. 2022, 13, 852416. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; Ikegaya, Y.; Koyama, R. The effects of microglia- and astrocyte-derived factors on neurogenesis in health and disease. Eur. J. Neurosci. 2021, 54, 5880–5901. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.M.; Miller, R.H.; Silver, J. CHANGING-ROLE OF FOREBRAIN ASTROCYTES DURING DEVELOPMENT, REGENERATIVE FAILURE, AND INDUCED REGENERATION UPON TRANSPLANTATION. J. Comp. Neurol. 1986, 251, 23–43. [Google Scholar] [CrossRef]
- Kliot, M.; Smith, G.M.; Siegal, J.D.; Silver, J. ASTROCYTE POLYMER IMPLANTS PROMOTE REGENERATION OF DORSAL-ROOT FIBERS INTO THE ADULT MAMMALIAN SPINAL-CORD. Exp. Neurol. 1990, 109, 57–69. [Google Scholar] [CrossRef]
- Becerra-Calixto, A.; Cardona-Gómez, G.P. Neuroprotection Induced by Transplanted CDK5 Knockdown Astrocytes in Global Cerebral Ischemic Rats. Mol. Neurobiol. 2017, 54, 6681–6696. [Google Scholar] [CrossRef]
- Becerra-Calixto, A.; Posada-Duque, R.; Cardona-Gómez, G.P. Recovery of Neurovascular Unit Integrity by CDK5-KD Astrocyte Transplantation in a Global Cerebral Ischemia Model. Mol. Neurobiol. 2018, 55, 8563–8585. [Google Scholar] [CrossRef] [PubMed]
- Merienne, N.; Delzor, A.; Viret, A.; Dufour, N.; Rey, M.; Hantraye, P.; Deglon, N. Gene transfer engineering for astrocyte-specific silencing in the CNS. Gene Ther. 2015, 22, 830–839. [Google Scholar] [CrossRef]
- Nicaise, C.; Mitrecic, D.; Falnikar, A.; Lepore, A.C. Transplantation of stem cell-derived astrocytes for the treatment of amyotrophic lateral sclerosis and spinal cord injury. World J. Stem Cells 2015, 7, 380–398. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.Y.; Chang, Y.S.; Sung, D.K.; Sung, S.I.; Park, W.S. Hypothermia broadens the therapeutic time window of mesenchymal stem cell transplantation for severe neonatal hypoxic ischemic encephalopathy. Sci. Rep. 2018, 8, 7665. [Google Scholar] [CrossRef]
- Corcelli, M.; Hawkins, K.; Vlahova, F.; Hunjan, A.; Dowding, K.; De Coppi, P.; David, A.L.; Peebles, D.; Gressens, P.; Hagberg, H.; et al. Neuroprotection of the hypoxic-ischemic mouse brain by human CD117(+)CD90(+)CD105(+) amniotic fluid stem cells. Sci. Rep. 2018, 8, 2425. [Google Scholar] [CrossRef]
- Mora, S.; Muniz-Garcia, A.; Zorzano, A. Molecular Mechanisms of Hypoxia-Induced Extracellular Vesicle Release. Faseb J. 2020, 34. [Google Scholar] [CrossRef]
- Urabe, F.; Kosaka, N.; Ito, K.; Kimura, T.; Egawa, S.; Ochiya, T. Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am. J. Physiol. Cell Physiol. 2020, 318, C29–c39. [Google Scholar] [CrossRef]
- Li, Y.; Tan, J.; Miao, Y.; Zhang, Q. MicroRNA in extracellular vesicles regulates inflammation through macrophages under hypoxia. Cell Death Discov. 2021, 7. [Google Scholar] [CrossRef]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Xian, P.; Hei, Y.; Wang, R.; Wang, T.; Yang, J.; Li, J.; Di, Z.; Liu, Z.; Baskys, A.; Liu, W.; et al. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice. Theranostics 2019, 9, 5956–5975. [Google Scholar] [CrossRef]
- Buller, B.; Liu, X.; Wang, X.; Zhang, R.L.; Zhang, L.; Hozeska-Solgot, A.; Chopp, M.; Zhang, Z.G. MicroRNA-21 protects neurons from ischemic death. Febs J. 2010, 277, 4299–4307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Y.; Lv, Q.; Gao, J.; Hu, L.; He, Z. MicroRNA-21 Overexpression Promotes the Neuroprotective Efficacy of Mesenchymal Stem Cells for Treatment of Intracerebral Hemorrhage. Front. Neurol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Xin, D.-Q.; Zhao, Y.-J.; Li, T.-T.; Ke, H.-F.; Gai, C.-C.; Guo, X.-F.; Chen, W.-Q.; Liu, D.-X.; Wang, Z. The delivery of miR-21a-5p by extracellular vesicles induces microglial polarization via the STAT3 pathway following hypoxia-ischemia in neonatal mice. Neural Regen. Res. 2022, 17, 2238–2246. [Google Scholar] [CrossRef]
- Qiu, L.; Cai, Y.; Geng, Y.; Yao, X.; Wang, L.; Cao, H.; Zhang, X.; Wu, Q.; Kong, D.; Ding, D.; et al. Mesenchymal stem cell-derived extracellular vesicles attenuate tPA-induced blood-brain barrier disruption in murine ischemic stroke models. Acta Biomater. 2022, 154, 424–442. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Li, M.C. Astrocyte-derived extracellular vesicles inhibit the abnormal activation of immune function in neonatal mice with hypoxic-ischemic brain damage by carrying miR-124-3p. Neurol. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.-Y.D.; Shetty, A.K. Treating Parkinson's disease by astrocyte reprogramming: Progress and challenges. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.A.; Jaronen, M.; Covacu, R.; Zandee, S.E.J.; Scalisi, G.; Rothhammer, V.; Tjon, E.C.; Chao, C.C.; Kenison, J.E.; Blain, M.; et al. Environmental Control of Astrocyte Pathogenic Activities in CNS Inflammation. Cell 2019, 176, 581. [Google Scholar] [CrossRef] [PubMed]
- Buffo, A.; Rite, I.; Tripathi, P.; Lepier, A.; Colak, D.; Horn, A.-P.; Mori, T.; Goetz, M. Origin and progeny of reactive gliosis: A source of multipotent cells in the injured brain. Proc. Natl. Acad. Sci. USA 2008, 105, 3581–3586. [Google Scholar] [CrossRef] [PubMed]
- Robel, S.; Berninger, B.; Goetz, M. The stem cell potential of glia: lessons from reactive gliosis. Nat. Rev. Neurosci. 2011, 12, 88–104. [Google Scholar] [CrossRef]
- Blum, R.; Heinrich, C.; Sanchez, R.; Lepier, A.; Gundelfinger, E.D.; Berninger, B.; Goetz, M. Neuronal Network Formation from Reprogrammed Early Postnatal Rat Cortical Glial Cells. Cerebral Cortex 2011, 21, 413–424. [Google Scholar] [CrossRef]
- Berninger, B.; Costa, M.R.; Koch, U.; Schroeder, T.; Sutor, B.; Grothe, B.; Gotz, M. Functional properties of neurons derived from in vitro reprogrammed postnatal astroglia. J. Neurosci. 2007, 27, 8654–8664. [Google Scholar] [CrossRef]
- Aravantinou-Fatorou, K.; Ortega, F.; Chroni-Tzartou, D.; Antoniou, N.; Poulopoulou, C.; Politis, P.K.; Berninger, B.; Matsas, R.; Thomaidou, D. CEND1 and NEUROGENIN2 Reprogram Mouse Astrocytes and Embryonic Fibroblasts to Induced Neural Precursors and Differentiated Neurons. Stem Cell Rep. 2015, 5, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Cates, K.; McCoy, M.J.; Kwon, J.-S.; Liu, Y.; Abernathy, D.G.; Zhang, B.; Liu, S.; Gontarz, P.; Kim, W.K.; Chen, S.; et al. Deconstructing Stepwise Fate Conversion of Human Fibroblasts to Neurons by MicroRNAs. Cell Stem Cell 2021, 28, 127. [Google Scholar] [CrossRef] [PubMed]
- Drouin-Ouellet, J.; Legault, E.M.; Nilsson, F.; Pircs, K.; Bouquety, J.; Petit, F.; Shrigley, S.; Birtele, M.; Pereira, M.; Storm, P.; et al. Age-related pathological impairments in directly reprogrammed dopaminergic neurons derived from patients with idiopathic Parkinson's disease. Stem Cell Rep. 2022, 17, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, E.; Koutsoudaki, P.N.; Thanou, I.; Karagkouni, D.; Karamitros, T.; Chroni-Tzartou, D.; Gaitanou, M.; Gkemisis, C.; Margariti, M.; Xingi, E.; et al. A miR-124-mediated post-transcriptional mechanism controlling the cell fate switch of astrocytes to induced neurons. Stem Cell Rep. 2023, 18, 915–935. [Google Scholar] [CrossRef]
- Diaz-Castro, B.; Bernstein, A.M.; Coppola, G.; Sofroniew, M.V.; Khakh, B.S. Molecular and functional properties of cortical astrocytes during peripherally induced neuroinflammation. Cell Rep. 2021, 36. [Google Scholar] [CrossRef]
- Zhang, P.J.; Wu, W.Y.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16. [Google Scholar] [CrossRef]
- Drommelschmidt, K.; Serdar, M.; Bendix, I.; Herz, J.; Bertling, F.; Prager, S.; Keller, M.; Ludwig, A.K.; Duhan, V.; Radtke, S.; et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain Behav. Immun. 2017, 60, 220–232. [Google Scholar] [CrossRef]
Figure 1.
Non-coding RNAs involved in processes related to apoptosis autophagy (A), hypoxia-ischemia (B), inflammatory response (C), and cell proliferation (D) interact with other molecules in astrocytes. (Image Source: By Figdraw.).
Figure 1.
Non-coding RNAs involved in processes related to apoptosis autophagy (A), hypoxia-ischemia (B), inflammatory response (C), and cell proliferation (D) interact with other molecules in astrocytes. (Image Source: By Figdraw.).
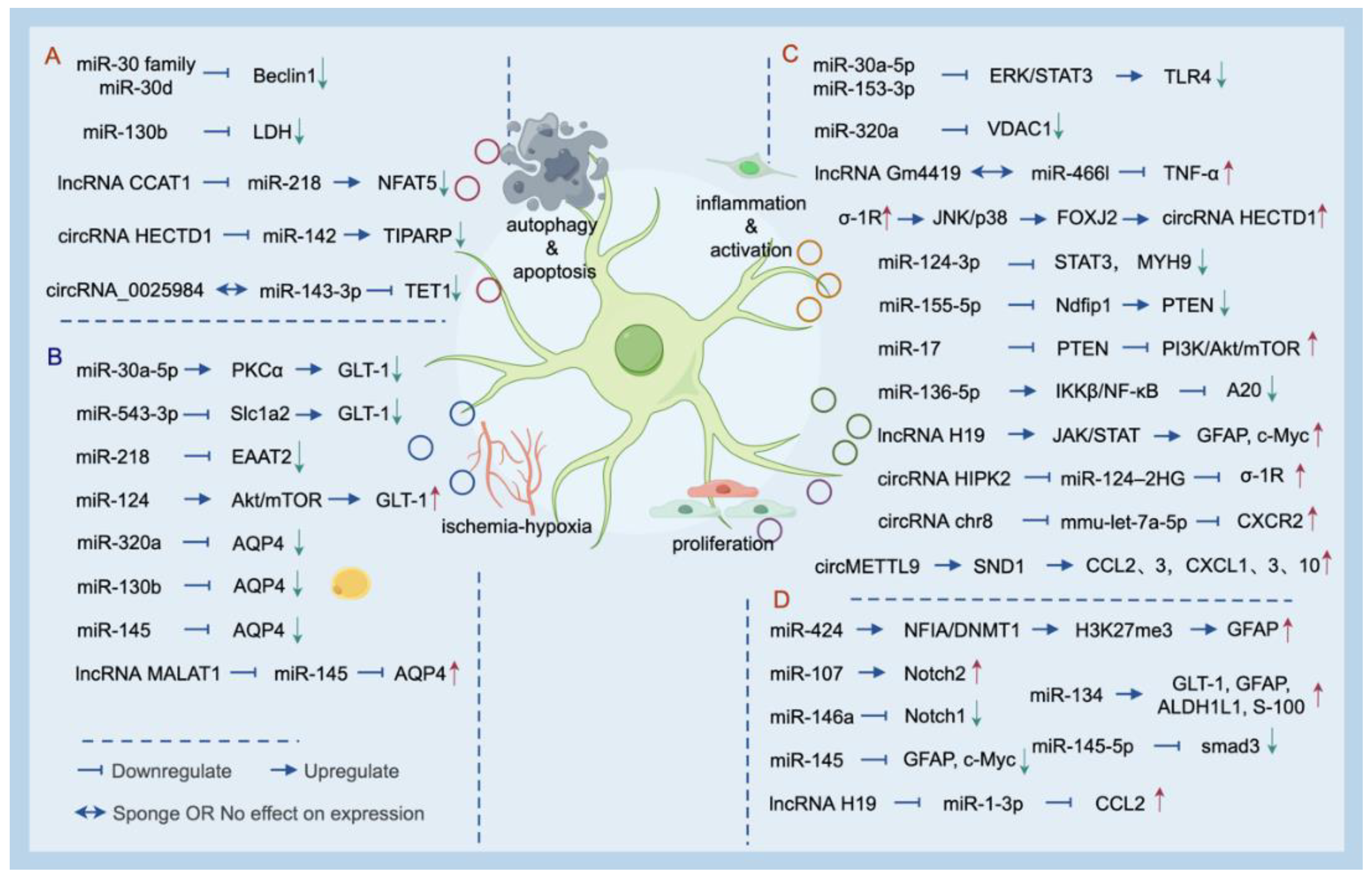

Table 1.
The functions of relevant non-coding RNAs associated with neonatal HIE.
| miR-30 family | miR-30a-5p | Regulation of reactive astrocyte or microglia proliferation | Neonatal exposure[69]; PD[70]; |
| miR-30d | Promotion of autophagy in astrocytes | Neonatal HIE[45]; OGD-induced autophagy[43] |
|
| miR-148a-3p | Inhibition of pyroptosis in astrocytes | Neonatal HIE[46] | |
| miR-218 | miR-218-5p | Inhibition of GLT-1 expression in astrocyte; Promotion of vascular remodeling | PE[35]; ALS[71]; |
| miR-124 | Inhibit of GLT-1 expression; Biomarkers of cerebral ischemia |
Perinatal Asphyxia and HIE[72]; Ischemic Stroke[33]; |
|
| miR-107 | Inhibit of GLT-1 expression; promotion of tumor cell proliferation | Neonatal HIE[73]; Pediatric High-grade glioma[74]; Ischemic Stroke[75]; |
|
| miR-320a | Reduction of cellular edema | Ischemic Injury; Neonatal Epilepsy[23] |
|
| miR-130b | Protection against astrocytes and nerve damage | OGD- induced brain injury; I/RI[76] |
|
| miR-145 | hsa-miR-145-5p | Protection against astrocytes damage; Inhibition of astrocyte proliferation | Neonatal HIE[77]; SCI[78] |
| miR-424 | Inhibition of reactive astrocyte proliferation | I/RI[79] | |
| miR-134 | miR-134-5p | Promotion of astrocyte maturation | Ischemic Injury; Neonatal HIE[52] |
| lncRNA MALAT1 | Influence of AQP4 expression in astrocytes | Neonatal BPD[80]; I/RI[56]; |
|
| lncRNA H19 | Downregulation of IGF-1 in astrocyte | Neonatal HIE[62] | |
| circRNA HECTD1 | Inhibition of astrocyte activation | Ischemic Stroke[67] | |
| circRNA 0025984 | Inhibition of astrocyte autophagy and apoptosis | Ischemic Stroke[68] | |
Table 2.
Regulatory functions played by non-coding RNA in astrocytes under neuroinflammatory conditions.
Table 2.
Regulatory functions played by non-coding RNA in astrocytes under neuroinflammatory conditions.
| Non-Coding RNA | Function | Disease | |
|---|---|---|---|
| miR-145 | miR-145-5p | Influence of inflammation-induced astrocyte proliferation | Neonatal HIE[90]; Ischemic Stroke; |
| miR-146a | Negative regulators of astrocyte-mediated inflammatory responses | Epilepsy[126]; ALS[127] | |
| miR-124 | miR-124-3p | Inhibition of astrocyte activation | Neonatal HIE[105]; SCI[100] |
| miR-155 | miR-155-5p | Promotion of astrocyte activation | Neonatal HIE[97]; SCI[128] |
| miR-136 | miR-136-5p | Promotion of astrocyte activation | SCI[129] |
| lncRNA H19 | Promotion of astrocyte activation and proliferation | I/R[113] | |
| lncRNA CRNDE | Inhibition of astrocyte activation | intrauterine infection-induced injury[117]; HIBD[130] |
|
| circRNA HIPK2 | Promotion of astrocyte activation by autophagy and ER stress | TBI[131] | |
| circRNA_chr8 | Promotion of astrocyte activation and neuroinflammation | TBI[132] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



