
Submitted:
11 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
Angiogenesis is a critical process in various physiological and pathological conditions, and snake venom disintegrins (SVDs) have been shown to be important regulators of this process. In this review, we explore the dual roles of SVD in angiogenesis, both as antiangiogenic agents by inhibiting integrin binding and interfering with vascular endothelial growth factors and as proangiogenic agents by enhancing integrin binding, stimulating cell migration and proliferation, and inducing neoangiogenesis. Studies in vitro and in animal models have demonstrated these effects and offer significant therapeutic opportunities. The potential applications of SVD in diseases related to angiogenesis, such as cancer, ocular diseases, tissue regeneration, wound healing, and cardiovascular diseases, are also discussed. Overall, SVDs are promising potential therapeutics, and further advances in this field could lead to innovative treatments for diseases related to angiogenesis.
Keywords:
snake venom
; angiogenesis
; integrins
; metalloproteinases
; disintegrins
1. Introduction
1.1. Angiogenesis
Angiogenesis is a complex biological process involving the formation of new blood vessels from preexisting vessels [1]. It plays a crucial role in various physiological and pathological conditions, including embryogenesis, wound healing, and tumor growth [2]. The process of angiogenesis is tightly regulated by multiple signaling pathways and factors. Matrix metalloproteinases are primarily responsible for degrading the basement membrane surrounding existing blood vessels, allowing endothelial cells to migrate and proliferate toward the angiogenic stimulus [3]. One of the key proangiogenic factors is vascular endothelial growth factor (VEGF), which is essential for the formation of new blood vessels during embryonic development and is produced by various cell types, including tumors [4]. VEGF binds to specific receptors on endothelial cells, promoting their survival, migration, and differentiation [5,6]. Other factors involved in angiogenesis regulation include fibroblast growth factors (FGFs), platelet-derived growth factor (PDGF), and angiopoietins [3]. These factors act synergistically to ensure the proper formation and remodeling of blood vessels. The regulation of angiogenesis is maintained through a dynamic balance between proangiogenic and antiangiogenic factors. This balance can be disrupted in various diseases, leading to either excessive or insufficient blood vessel formation [1]. Insufficient angiogenesis, on the other hand, can lead to tissue ischemia, impaired wound healing, and various cardiovascular diseases [2]. Peripheral artery disease (PAD) is an example where the narrowing or blockage of blood vessels reduces blood flow to the legs or arms, resulting in pain, skin ulcers, and an increased risk of amputation [2]. Therapeutic angiogenesis, which aims to promote blood vessel formation, has been investigated as a potential treatment approach for PAD, with the administration of VEGF or the use of gene therapy [1].
It is important to emphasize that integrins, a group of transmembrane proteins that play a crucial role in cell adhesion and communication between cells and the extracellular environment, are the focus of interest, as they play an important role in angiogenesis [7].
1.2. Integrins
Integrins play an essential role in the regulation of biological processes such as cell migration, adhesion, proliferation, differentiation and signaling [8]. Integrins are heterodimeric glycoproteins consisting of alpha and beta subunits, which together form a complex transmembrane receptor [9]. A total of 18 alpha subunits and 8 beta subunits have been identified in mammalian cells, allowing the formation of 24 different heterodimers. The α subunit determines affinity to extracellular matrix component (ECM), while the β subunit associates with cytoplasmic structural and regulatory proteins. These subunits have a long transmembrane domain and a short cytoplasmic domain associated with cytoskeletal proteins [10]. Although integrins are constitutively expressed on the cell surface, they need to be activated to interact with their ligands [11]. Such activation can occur in the presence of chemokines and cytokines and is characterized by a conformational change at the extracellular integrin domain that exposes their binding sites on the α and β subunits, allowing them to interact with their ligands on the ECM or with proteins on the membranes of neighboring cells. This interaction is primarily controlled by a conserved tripeptide pattern of arginine, glycine, and aspartate, commonly known as the Arg-Gly-Asp motif (RGD) [12,13,14]. They are crucial for cell adhesion, migration, and signal transduction by mediating interactions between cells and ECM proteins. The main role of these molecules is to provide a link between the cytoskeleton of the cell and certain ECM components, such as fibronectin, vitronectin, laminin, and collagen. In addition, they are responsible for triggering intracellular signal transduction pathways upon interaction with the ECM. Integrins can undergo conformational changes that influence their ligand binding properties and downstream signaling events [15]. They possess a unique arrangement of cysteine residues that enables them to adopt a compact and stable structure consisting of a well-defined loop disulfide array [16]. This structural motif is critical for their high-affinity binding to integrins, as it allows them to bind to specific integrin subunits and block their ligand binding sites. In addition, integrins also interact with growth factor receptors to regulate cell migration, blood vessel development, and angiogenesis. Importantly, the understanding of the mechanism of action of integrins paralleled the discovery of proteins from snake venoms, called disintegrins, which act as potent inhibitors of platelet aggregation and integrin receptor-dependent cell adhesion [17,18].
1.3. Snake Venom Metalloproteinases and Snake Venom Disintegrins
Snake venom metalloproteinases (SVMPs) are a major component of most crotalid and viperid venoms [19]. SVMPs are known to be a class of key toxins involved in the pathophysiology associated with viperid venoms and are classified into classes and subclasses from P-I to P-III according to the organization of their domains, as shown in Figure 1 [20]. In general, class PI includes metalloproteinases that have only a catalytic domain containing zinc; class PII, metalloproteinases that have a catalytic domain followed by a disintegrin domain containing the tripeptide RGD (arginine-glycine-aspartic acid); class PIII, metalloproteinases that have a catalytic domain, a disintegrin-like domain, and a cysteine-rich domain [21].
Snake venom disintegrins (SVDs) are a class of proteins derived from SVMPs. The disintegrins present in snake venoms can be formed in the venom gland I n two distinct ways: (1) By proteolysis of SVMPs of class P-II, where cleavage occurs between the catalytic domain and the disintegrin domain, leaving only the disintegrin domain. These disintegrins are known as RGD-dependent disintegrins, which are abundant in viperid venoms and contain the sequence XGD (X-Gly-Asp), MLD (Met-Leu-Asp), or K/RTS (Lys/Arg-Thr-Ser) on the exposed surface of the loop that specifically binds to integrins on the surface of different cell types [22,23]. The amino acid sequences immediately adjacent to the RGD site of disintegrins could form an extended RGD locus that, in conjunction with the conformational representation of the RGD sequence, could be involved in determining integrin selectivity and affinity [13]. The family of disintegrins containing the RGD motif is widely recognized as the most extensive and well-studied group. Most of these units function as monomers (small, medium, or large), but a subgroup has the ability to combine into dimers and form homo or heterodimers [24]. (2) Proteolysis of class P-III SVMPs results in fragments that covalently link the disintegrin-like and cysteine-rich domains and are referred to as ECD-disintegrin-like/cysteine-rich domains (Figure 2). This disintegrin-like domain has a sequence of non-RGD tripeptides in its binding site [21,25].
Figure 1.
Disintegrins are categorized according to their structural composition. The number of disulfide bonds and the length of the polypeptide chain determine this categorization.
Figure 1.
Disintegrins are categorized according to their structural composition. The number of disulfide bonds and the length of the polypeptide chain determine this categorization.
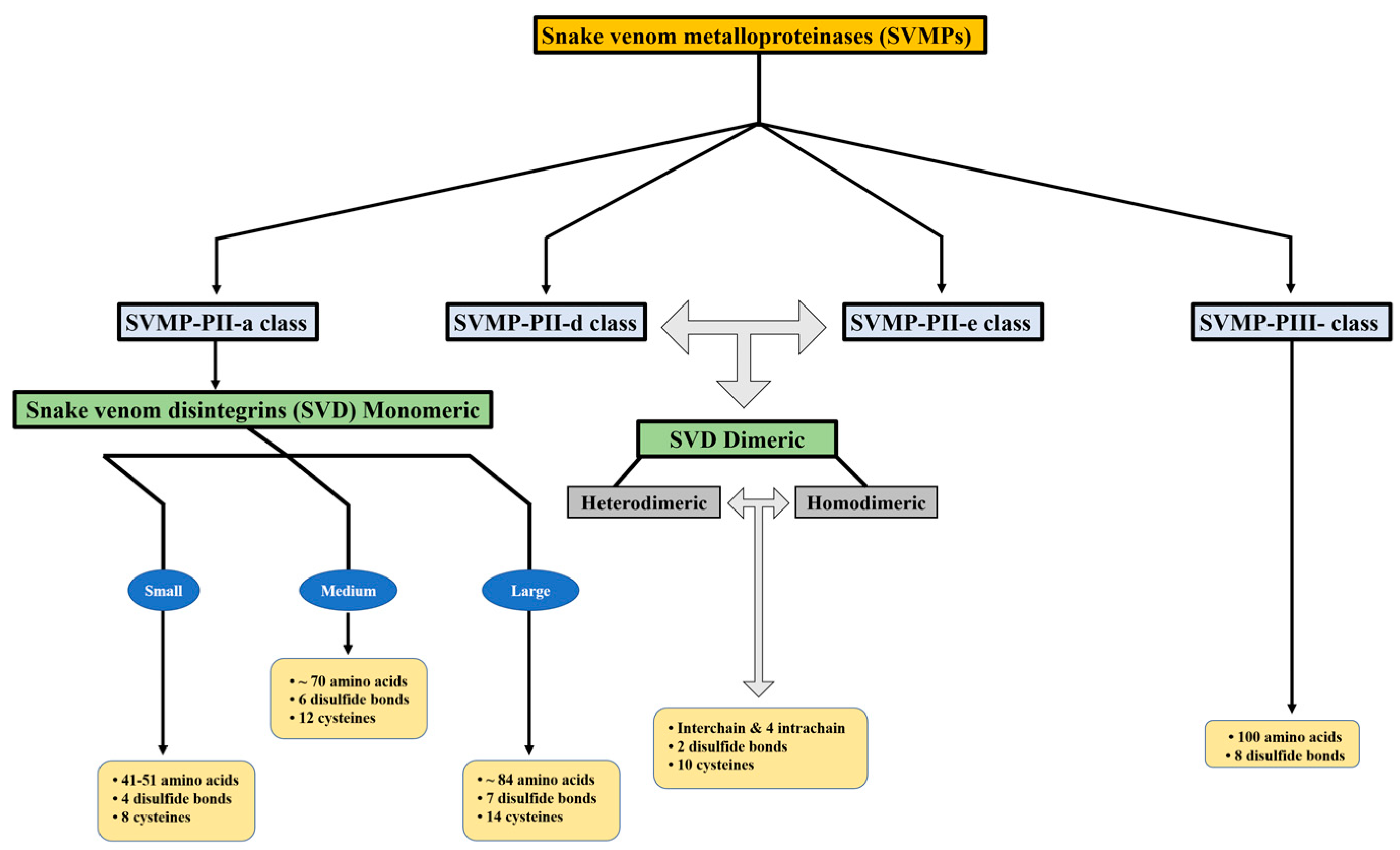
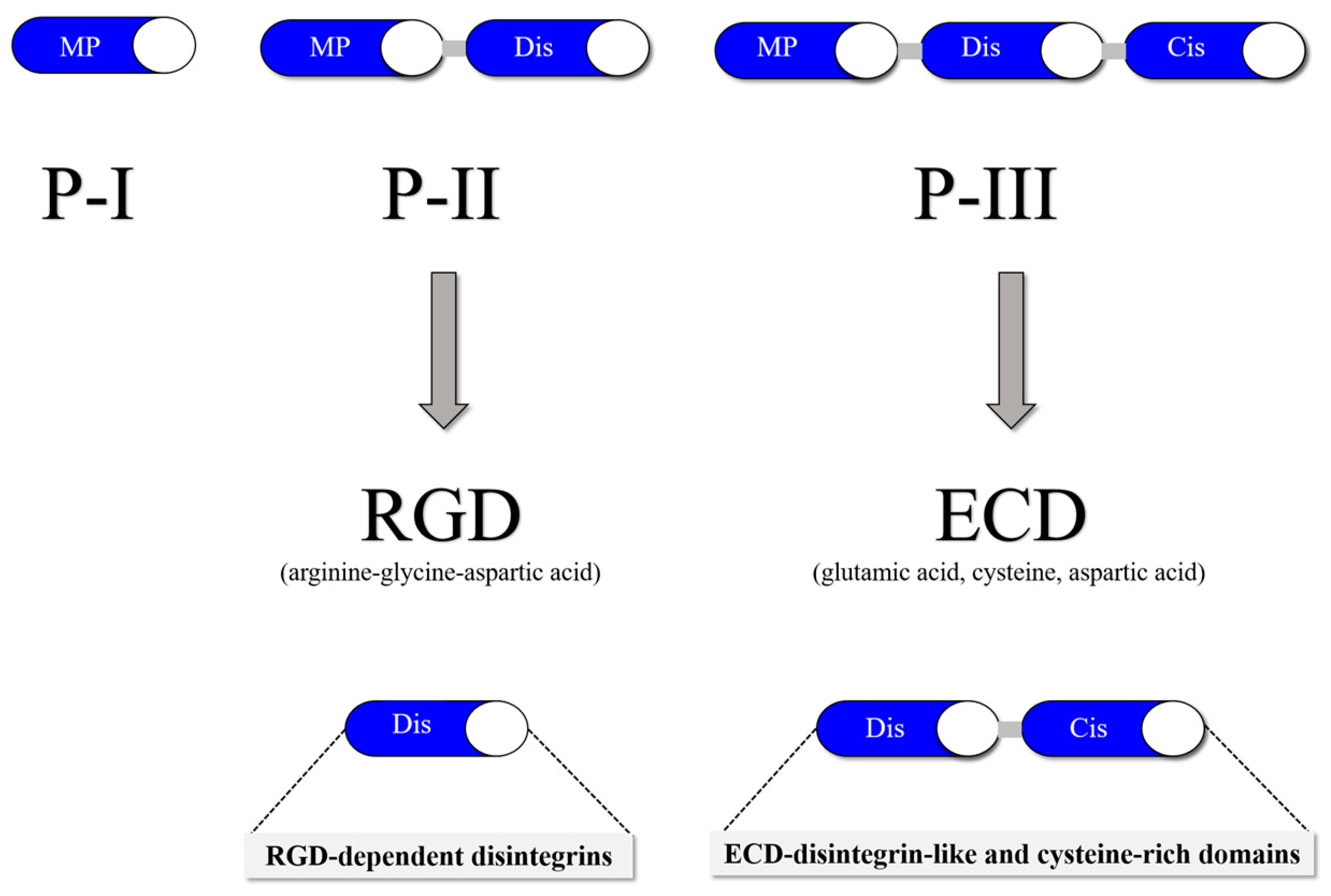
Figure 2.
SVMP classification into three classes: P–I (20–30 kDa), which in its mature form contains a metalloproteinase domain; P-II (30–60 kDa), which contains a disintegrin domain linked to the C-terminus of the metalloproteinase domain; and P-III (60–100 kDa), which consists of a metalloproteinase domain, a disintegrin-like domain and a cysteine-rich domain.
Figure 2.
SVMP classification into three classes: P–I (20–30 kDa), which in its mature form contains a metalloproteinase domain; P-II (30–60 kDa), which contains a disintegrin domain linked to the C-terminus of the metalloproteinase domain; and P-III (60–100 kDa), which consists of a metalloproteinase domain, a disintegrin-like domain and a cysteine-rich domain.
SVDs are known for their ability to bind to integrin receptors and modulate various cellular functions, such as inflammation, apoptosis in endothelial cells and inhibition of platelet aggregation [26]. Jararhagin-C, an SVD containing the ECD-disintegrin-like/cysteine-rich domains produced by the proteolytic cleavage of Jararhagin (SVMP P-III), is present in Bothrops jararaca venom and specifically interacts with α2β1 integrin, inhibiting collagen and ADP-induced platelet aggregation [8]. Jararhagin-C is also capable of triggering the local release of cytokines [27] and induces changes in leukocyte-endothelium interactions through the expression of the adhesion molecules ICAM-1, CD11a and CD11b [28,29]. Alternagin-C, a toxin isolated from B. alternatus venom, is a protein composed of the ECD-disintegrin-like/cysteine-rich domain, with 92% homology to Jar-C. It is able to interfere with α2β1 integrin functions and may contribute to apoptosis by interfering with cell adhesion [30].
One of the notable characteristics of SVDs is their ability to inhibit platelet aggregation, an important step in blood clot formation [31]. This property has led to the exploration of disintegrins as potential antithrombotic drugs for the prevention and treatment of conditions such as deep vein thrombosis and stroke. For example, Insularin, a monomeric RGD-disintegrin isolated from the venom of Bothrops insularis, strongly inhibits human platelet aggregation and fibrinogen-adhesion of endothelial cells [32]. Echistatin, an RGD disintegrin from the venom of the saw-scaled viper (Echis carinatus), effectively inhibits platelet aggregation by binding to integrin αIIbβ3 [33]. Upon binding to integrins, SVDs interfere with various cellular processes. Disintegrins have gained attention in the field of medicine due to their unique properties and potential therapeutic applications.
SVDs have also shown promise in cancer research [34-36]. Integrins play a crucial role in tumor growth, invasion, and metastasis [37]. By targeting specific integrins expressed on cancer cells, SVD can potentially inhibit the proliferation and migration of these cells. Contortrostatin, a disintegrin from the venom of the Agkistrodon contortrix contortrix, has been found to have anticancer properties by interacting with integrins αvβ3 and α5β1 [38]. Vicrostatin, a recombinant disintegrin developed by fusing 62 N-terminal amino acids of the disintegrin Contortrostatin with 6 C-terminal amino acids of Echistatin, is the best-characterized and most preclinically advanced disintegrin shown to target multiple tumor-associated integrins and to exhibit potent antitumor and antiangiogenic activity in in vitro and in vivo models without appreciable toxicity [23].
Since the 1987 report on the isolation of trigramin, the first disintegrin isolated from the venom of Trimeresurus gramineus, approximately 100 other disintegrins have become known from snake venoms with potential applications in cancer research and therapy (see the list in [17]).
1.4. The role and properties of SVDs in angiogenesis
One of the most well-known effects of SVD is its ability to prevent cell adhesion to extracellular matrix proteins [16]. By occupying the ligand binding sites on integrins, disintegrins disrupt the attachment of cells to their surrounding microenvironment. This disruption can have profound effects on cell migration, invasion and angiogenesis, processes that are essential for tumor progression and metastasis [18]. Dysregulation of integrin activity has been implicated in numerous pathological conditions, such as cancer, inflammation, and thrombosis [39,40]. Consequently, inhibiting integrin binding has emerged as a promising therapeutic strategy [41,42].
Several studies highlight the dual role of SVD in angiogenesis, with some disintegrins acting as inhibitors and others acting as enhancers of the process [43,44,45,46]. The interaction between disintegrins and integrin receptors on endothelial cells plays a critical role in modulating angiogenesis. Therefore, inhibitors of α1β1 and α2β1 integrins alone or in combination with antagonists of other integrins involved in angiogenesis (eg. αvβ3, αvβ5, αIIbβ3 and α6β4) may prove beneficial in controlling neovascularization, making SVDs valuable tools for the study and potential manipulation of this complex biological process.
Some SVDs have been identified as potent inhibitors of angiogenesis (Table 1). For instance, obtustatin, a disintegrin isolated from the venom of Vipera lebetina obtusa, has been shown to selectively inhibit α1β1 integrin, leading to the suppression of angiogenesis in vitro and in vivo [47]. A similar effect was also observed with viperistatin isolated from the venom of Vipera paleastinae [48]. Furthermore, jerdostatin, isolated from the venom of Trimeresurus jerdonii [49], and lebestatin, isolated from the venom of Macrovipera lebetina [43], two other small monomeric disintegrins that antagonize the function of the α1β1 integrin, have also been described as inhibitors of angiogenesis. Rhodostomin, a medium disintegrin from the venom of Calloselasma rhodostoma, has been reported to inhibit angiogenesis by binding to integrins and inhibiting bFGF-induced proliferation of endothelial cells [50].

Table 1.
List of disintegrins found in snake venom that have pro- and/or anti-angiogenic properties.
Table 1.
List of disintegrins found in snake venom that have pro- and/or anti-angiogenic properties.
| Name (Source) | Recognizing Motif | Physiological Target | Angiogenic factors | References |
|---|---|---|---|---|
| Jararhagin-C | ECD-disintegrin-like/cysteine-rich domains | Interferes with α2β1 integrin functions | Pro-angiogenic | [8] |
| Alternagina-C | Exhibits both pro- and anti-angiogenic effects | [30,51,52] | ||
| Leberagin-C | Disintegrin-like | Interferes with αvβ3, αvβ6, and α5β1 integrins | Anti-angiogenesis | [53](63) |
| Echistatin | RGD-dependent disintegrins | Binds to integrin αIIbβ3, GPIIb/IIIa and interacts with αvβ3 integrin | Anti-angiogenesis | [14,33,54] |
| Rhodostomin | [50] | |||
| Contortrostatin | Interacts with the integrins αvβ3 and α5β1 | [38] | ||
| Vicrostatin | antagonize the function of the αIIbβ3, αvβ3, αvβ5 and α5β1 integrins | [23] | ||
| DisBa-01 | Binds to integrin αvβ3 | [55] | ||
| Aggretin | Binds to integrin α2β1 | Pro-angiogenic | [56] | |
| Trigramin | Binds to αIIbβ3, α8β1, αvβ3, αvβ5and/or α5β1 integrins | [57] | ||
| Obtustatin | KTS-disintegrin | Selectively inhibit α1β1-integrin | Anti-angiogenesis | [47] |
| Viperistatin | Inhibitory activity against collagen receptors, α1β1 and α2β1-integrins | [48] | ||
| Lebestatin | Inhibis binding of α1β1 integrin to type IV and type I collagen | [43] | ||
| Jerdostatin | RTS-disintegrin | Antagonizes the function of the α1β1 integrin | Anti-angiogenesis | [49] |
| Agkistin-s | InteractS with GPIB | Anti-angiogenesis | [58] |
On the other hand, certain SVDs have been found to promote angiogenesis. For instance, Jararhagin-C has been shown to promote angiogenesis by activating integrin receptors and stimulating endothelial cell migration, increasing the density of blood vessels and the synthesis of proangiogenic cytokines (VEGF and FGF) [45].
Alternagin-C exhibits both pro- and antiangiogenic effects depending on the concentration. Concentrations less than 50 nM were found to be proangiogenic, whereas concentrations greater than 100 nM were found to be antiangiogenic both in vitro and in vivo [51,52]. Alternagin-C inhibits VEGF/VEGFR2 signaling after binding to α2β1 integrin, resulting in impaired angiogenesis [51].
1.4.1. Interference of SVD with VEGF
VGEF is a key molecule for regulating and promoting the angiogenesis process through the stimulation of proliferation, migration and organization of endothelial cells, triggering signaling cascades such as Notch, angiopoietin/Tie, MAPK, FAK, PI3K/AKT, ERK1/2, Src and PLCγ. While VEGFs stimulate the proliferation of endothelial cells, integrins help anchor these cells to the extracellular matrix to ensure that vessel formation occurs in an organized manner. SVDs can affect vascular VEGF, which plays a crucial role in regulating blood vessel formation and vascular permeability [59,60,61].
There are several ways in which SVDs can interfere with VEGF function. Disintegrins can inhibit angiogenesis, the process of forming new blood vessels, which is essential for tumor growth and metastasis [7,62]. By interfering with VEGF-induced signaling, disintegrins hinder the formation of new blood vessels and thus impair tumor progression [63]. Moreover, disintegrins can interfere with the signal transduction pathways initiated by VEGF [64,65]. This interference can affect various cellular processes, including gene expression and protein synthesis, which are crucial for the regulation of blood vessel formation and permeability [59]. Overall, SVD exerts its effects on VEGFs through the inhibition of angiogenesis and interference with signal transduction pathways, ultimately influencing the regulation of blood vessel formation and vascular permeability.
1.5. Anti-angiogenic effects of SVDs
Recent studies have demonstrated the antiangiogenic effects of SVD, suggesting their potential as therapeutic agents for various human diseases, including cancer. In vitro experiments have shown that contortrostatin exhibits antiangiogenic activity [38]. Similarly, DisBa-01, a disintegrin from Bothrops alternatus snake venom, has been found to inhibit the proliferation, migration, and tube formation of human umbilical vein endothelial cells, which play crucial roles in angiogenesis [55]. Echistatin has also been shown to inhibit the proliferation and migration of human microvascular endothelial cells [64]. Animal model studies have further supported the antiangiogenic effects of SVD. For instance, contortrostatin has been found to inhibit tumor growth and angiogenesis in a mouse model of melanoma [38,63]. Similarly, Echistatin has demonstrated the ability to inhibit tumor growth and angiogenesis in a mouse model of glioma [66]. Agkistin is a P-II class SVMP containing a metalloproteinase and a disintegrin domain purified from crude venom of Formosan Agkistrodon acutus. Agkistin-s is the disintegrin domain of Agkistin and induces endothelial cell apoptosis, exhibiting profound antiangiogenic activity [58].
Additionally, Leberagin-C (Leb-C), a disintegrin from Macrovipera lebetina transmediterrannea snakes, has been shown to disrupt the adhesion, migration, and invasion capabilities of MDA-MB-231 breast cancer cells and its highly metastatic D3H2LN subpopulation [53].
2. Potential applications of SVD in angiogenesis-related diseases
Diseases characterized by abnormal angiogenesis, such as cancer and ocular diseases, represent a major public health challenge worldwide [67]. The search for effective therapeutic agents to modulate angiogenesis is of great interest to medical researchers [68]. SVDs have emerged as potential candidates for antiangiogenic and proangiogenic therapies [17,23,69]. Some of the diseases are characterized by abnormal angiogenesis.
2.1. Cancer
Cancer is a multifaceted group of diseases that poses a major health challenge worldwide and results in a significant burden of morbidity and mortality. The etiology of cancer involves a multifaceted interplay between genetic and epigenetic elements that lead to alterations in the genome and subsequently trigger uncontrolled cell proliferation in the tissues and organs of the body [70]. Cancer can be classified into different types based on the specific cell type from which the tumor originates. These include carcinoma, sarcoma, lymphoma and leukemia, germ cell tumor and blastoma [71]. Worldwide cancer statistics show that in 91 of 172 countries, cancer is the primary or secondary leading cause of death before age 70. In 22 other countries, cancer is the third or fourth leading cause of death [72]. Chemotherapy, radiotherapy, immunotherapy and surgery together form an integral part of modern cancer treatment. The pharmaceutical industry is currently conducting extensive research to develop innovative drugs of natural origin to mitigate the adverse effects associated with cancer treatments [73-75]. Tumor angiogenesis, the sprouting of new blood vessels into tumors, is vital for sustained tumor growth, progression, and metastasis [76,77]. Various growth factors and cytokines orchestrate the angiogenic process, including VEGF and FGF [78,79]. The abnormal angiogenesis associated with cancer provides an opportunity for therapeutic intervention by targeting angiogenic factors or inhibiting the endothelial cell response [80,81]. Changes in the structure of tumor vessels toward a more mature phenotype could also promote resistance. Therefore, the development of drug resistance is important in the development of new antiangiogenic therapies [7].
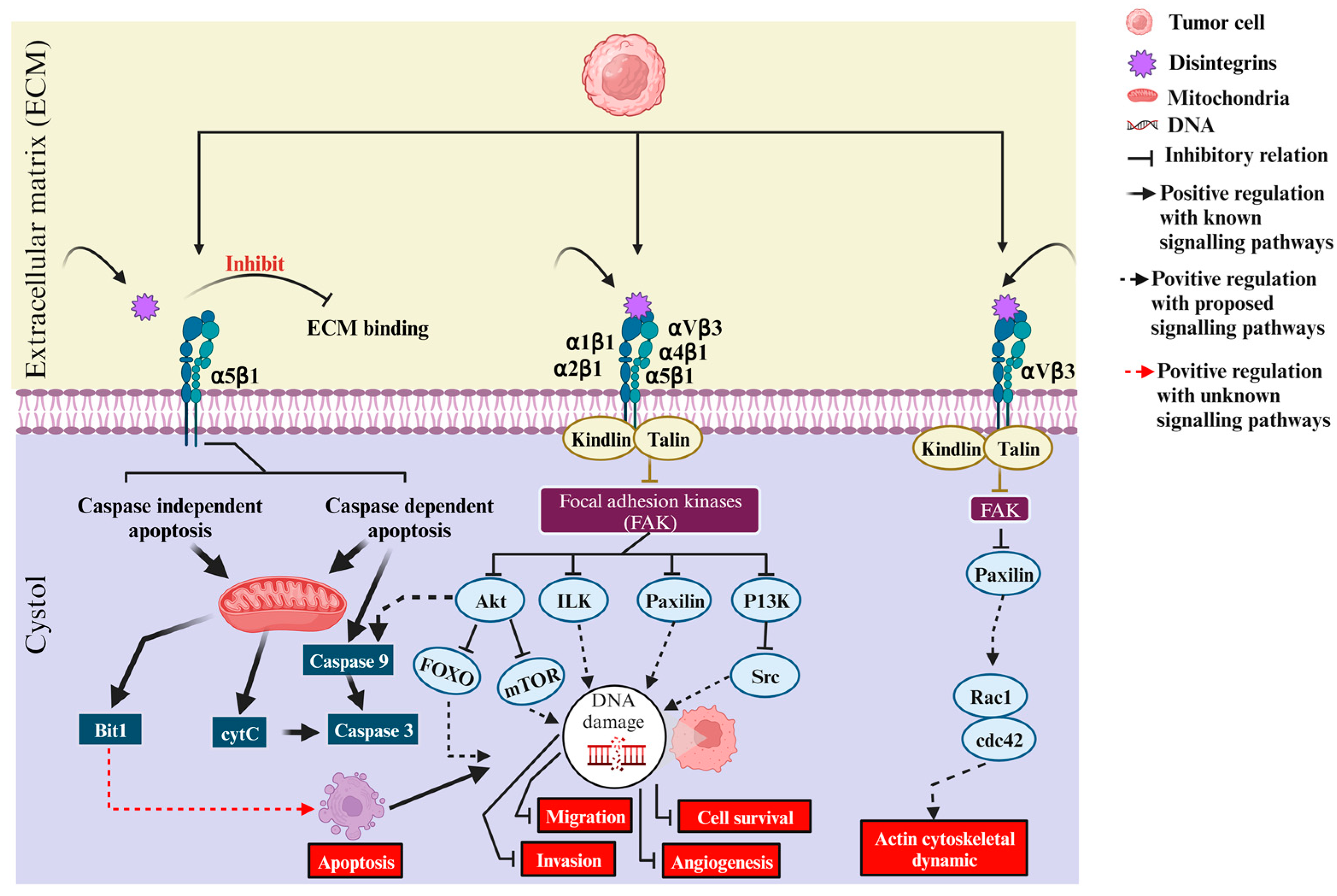
A chimeric recombinant disintegrin called Vicrostatin (VCN) has been used in a number of preclinical studies in various tumor models. VCN is a monomer prepared by recombinant modification of the primary Contortrostatin sequence by replacing the C-terminal tail with 6 aa derived from the C-terminus of Echistatin. It retained the targeting of Contortrostatin to αIIbβ3-, α5β1, αvβ3, and αvβ5 integrins but additionally showed more than 10-fold higher affinity for α5β1-integrin. Compared with the homodimeric structure of contortrostatin, the novel chimeric disintegrin VCN was active as a monomer, which allowed its production in larger amounts than several other cloned and expressed disintegrins. Like Contortrostatin, VCN has been shown to inhibit tumor cell adhesion, endothelial and tumor cell invasion, and angiogenesis. This chimeric recombinant toxin has shown promising results in preclinical experimental models of breast cancer, prostate cancer, ovarian cancer and glioblastoma [23]. Figure 3 shows the intracellular reactions that take place when disintegrins bind to integrin targets, as illustrated in the current literature. The involvement of α5β1-, αVβ3-, α1β1-, α2β1-, α4β1-, and α5β1-integrins in survival, proliferation, infiltration, motility, cytoskeletal reorganization, angiogenesis, apoptosis, and interaction with snake venom disintegrins triggers the onset of these mechanisms in cancer cells [82].
Figure 3.
This figure shows how disintegrins act on different integrins and initiate intracellular signaling in cancer cells.
Figure 3.
This figure shows how disintegrins act on different integrins and initiate intracellular signaling in cancer cells.
2.2. Ocular Diseases, e.g., Diabetic Retinopathy
Diabetic retinopathy (DR), a common microangiopathic sequela of diabetes mellitus [83], has a significant impact worldwide. It affects more than 100 million people and is a major contributor to visual impairment and blindness in industrialized countries [84]. DR is characterized by abnormal neovascularization in the retinal tissue of diabetic patients, leading to impaired visual function and eventual loss of vision [85]. The etiology of DR is primarily defined by the simultaneous occurrence of neurovascular unit dysfunction, blood‒retinal barrier disruption, inflammatory processes, capillary nonperfusion or ischemia, and neoangiogenesis [86]. The disease is primarily driven by increased VEGF levels in response to ischemic or hypoxic stimuli, causing several alterations at different levels and subsequent neovascularization [87,88]. Pathological angiogenesis in DR disrupts retinal function and causes visual complications. It has been reported that various fractions in snake venoms markedly increase insulin secretion without causing harmful effects. For example, a number of disintegrin isoforms were identified in Crotalus vegrandis venom and confirmed through partial sequencing as being homologous to various other snake disintegrins that share the active RGD motif near the C-terminus [89]. These snake venom disintegrins were shown to significantly enhance insulin secretion from BRIN-BD11 cells, suggesting disruption of the cellular signaling pathways activated by integrins, including receptor tyrosine kinases. These fractions have the potential to positively influence insulin secretion [89].
3. Utilization of SVD as Pro-Angiogenic Agents
While disintegrins are usually known for their antiangiogenic properties, recent research has also revealed their potential as proangiogenic agents. Alternagin-C (ALT-C) is an ECD-containing disintegrin-like/cysteine-rich disintegrin isolated from the venom of the snake Rhinocerophis alternatus that induces endothelial cell proliferation and angiogenesis both in vitro and in vivo by upregulating the expression of VEGF and its receptors [90]. ALT-C binds to the major collagen receptor α2β1 integrin, inhibiting cell adhesion to collagen, triggering downstream signaling molecules, and inducing a significant increase in several genes related to cell cycle control (VEGF, inducible early growth response, interleukin 11, early growth response 2 and 3, and the insulin-induced gene) [52]. ALT-C also induced significant cytoskeleton dynamic changes with the polymerization of F-actin, focal adhesion kinase (FAK), and phosphoinositol 3-kinase (PI3K) activation, as well as erk-2 translocation [91]. ALT-C induced the formation of new vessels, and the expression of VEGF in the injured tissue indicated the usefulness and effectiveness of ALT-C as a proangiogenic disintegrin-like protein [46].
3.1. Tissue regeneration and wound healing
Wound healing and regeneration are multifaceted biological phenomena that occur throughout the human lifespan. After an injury, various cellular processes are immediately initiated and coordinated to initiate a response [92]. Improper repair procedures can cause this process to be delayed, with immediate consequences for the individual, including physical discomfort, impaired rehabilitation progress, limb amputation, and in the most severe cases, death from septicemia [93]. Like other natural healing processes, the repair mechanism also relies on the coordination of different cell activities vital for recovery. These activities encompass cell survival, growth, movement, and the creation of new cells. They are orchestrated through cellular interactions with both the extracellular matrix (ECM) that surrounds them and with other neighboring cells. These interactions are made possible by specialized receptors found on cell membranes, which belong to the integrin family [94]. A recent study led by Ferreira and colleagues [95] used an in vivo model with subcutaneous sponge implants to investigate the potential of jararhagin-C (Jar-C). Their goal was to understand how Jar-C might stimulate collagen deposition and the production of important soluble substances, including VEGF and transforming growth factor beta-1 (TGFβ-1). These substances play important roles in processes such as angiogenesis and fibrogenesis, which are closely associated with tissue repair. The results of the study suggest that Jar-C may have positive effects on tissue repair by promoting these natural responses in the body. Moreover, Alternagin-C is also the subject of research. Within 7 days, this cysteine-rich, disintegrin-like protein accelerates wound healing in rats by increasing type I collagen deposition and fibroblast density and reducing inflammation [96]. In another study, Rabelo et al [97] revealed that ALT-C increased collagen synthesis in mouse fibrovascular tissue. From the above results, it can be concluded that SVD has the ability to promote angiogenesis by modulating endothelial cell behavior, facilitating cell migration, and upregulating proangiogenic factors. However, further research is needed to gain a comprehensive understanding of the underlying mechanisms and to improve the therapeutic application of disintegrins in the context of tissue regeneration and wound healing.
3.2. Cardiovascular Diseases
Integrins have a considerable influence on the development of cardiac fibrosis. The diseased heart exhibits altered expression and function of integrins [98]. Targeting integrins and their associated proteins can be a potential therapeutic target for myocardial fibrosis. Certain disintegrins have been extensively researched and subsequently approved by the Food and Drug Administration (FDA), making them viable pharmaceutical agents in modern medicine. Tirofiban, marketed as Aggrastat®, is a synthetic pharmaceutical agent developed by Medicure International, Inc. of Winnipeg, Manitoba, Canada. This drug is derived from the RGD domain found in Echistatin [99]. In addition, this compound undergoes a chemical change that increases its affinity for platelet glycoproteins, especially GPIIb/IIIa receptors [33]. Therefore, this drug is able to prevent platelet aggregation and other thrombotic activities by competing with fibrinogen for the RGD domain recognition site in the GPIIb/IIIa receptor [99-101]. In 1998, the Food and Drug Administration (FDA) approved tirofiban as a therapeutic intervention for acute coronary syndrome [102]. Additionally, in 1998, the FDA approved eptifibatide (Integrilin®, Millennium Pharmaceuticals, Inc.), an alternative molecule to inhibit platelet aggregation. Schering-Plow subsequently acquired license rights to this drug in 2005 [103]. The development of this active substance took place in parallel with research into synthetic peptide analogs of barbourin, a disintegrin from Sistrurus miliarius barbouri [104].
Cardiovascular diseases, including acute myocardial infarction, coronary artery disease, endothelial dysfunction, and chronic ischemia, are considered one of the leading causes of death worldwide [105]. Therefore, there is great interest in pharmaceutical agents that offer a new and effective therapeutic approach to improve the functionality of the myocardium and/or facilitate its regeneration after injury. Platelets have been studied extensively because of their crucial role in primary hemostasis, the body's initial response to arterial injury. Only recently, however, have we begun to study their contribution to immunological processes, cardiovascular disease, cancer, and various other pathological conditions [106-108]. The glycoprotein receptor GPIIbIIIa has been extensively studied as a primary receptor on platelets for the functional effects of snake venom-derived disintegrins. Each individual platelet is equipped with approximately 80,000 GPIIbIIIa receptors located both on the plasma membrane and in the α-granules [109]. After activation, the number of these receptors increases significantly, facilitating the formation of a permanent hemostatic plug [82]. Certain SVDs, including those of Agkistrodon piscivorus piscivorus and Echis carinatus sochureki, namely, Applagin and Echistatin, show remarkable affinity for the RGD motif on resting platelets. This specific binding results in potent inhibition of platelet aggregation. Consequently, SVDs have emerged as promising candidates for drug development to antagonize platelet integrins, demonstrating their pharmacological potential as both platelet aggregation inhibitors and antithrombotic agents. From a clinical perspective, these drugs can effectively reduce the likelihood of acute ischemic episodes and serve as preventive measures against thrombotic sequelae. RGD disintegrins have been extensively studied and are considered the largest family within this category [17,110]. A number of disintegrins have been extracted from snake venoms, particularly viper venom, and characterized as agents with antithrombotic properties [111]. For example, trigramin inhibits platelet aggregation in platelet-rich plasma triggered by adenosine diphosphate (ADP), collagen, or epinephrine [57]. The same disintegrin has been shown to inhibit platelet aggregation both in vitro and in vivo by preventing the binding of fibrinogen to platelets induced by agonists of aggregation, such as ADP-activated platelets [112]. By promoting angiogenesis, SVDs have shown promise in preclinical models of cardiac ischemia. Previous studies have shown that administration of ALT-C in a single dose after a period of 7-9 days resulted in an increase in cardiac muscle contractile force in fish [46,113]. This intervention also led to an upregulation of the expression of important proteins involved in calcium processing and an increase in the level of VEGF in the myocardium. In addition, administration of ALT-C stimulated angiogenesis and thus protected cardiomyocytes from the deleterious effects of negative tropism caused by hypoxia/reoxygenation. Evaluation of the safety and efficacy of disintegrins in promoting angiogenesis and improving cardiac function in patients with cardiovascular disease requires the conduct of clinical trials.
4. Conclusion
SVDs play a critical role in modulating angiogenesis by either inhibiting or promoting this process. Their mechanisms of action offer valuable insights into potential therapeutic applications for angiogenesis-related diseases. These SVDs show promise as potent antiangiogenic agents for the treatment of cancer and ocular diseases and as proangiogenic agents for tissue regeneration and wound healing. However, further research is needed to fully understand their molecular mechanisms and optimize their therapeutic potential. Further exploration of SVDs could lead to the discovery of new molecules or the development of synthetic analogs with improved stability and specificity. These advances have the potential to revolutionize the treatment of diseases related to angiogenesis and open new avenues for personalized medicine approaches targeting angiogenesis.
Author Contributions
P.B.C. and S.S.S. conceptualized the idea; P.B.C. and S.S.S. prepared the initial draft, and M.S.D.C. and B.C.Z. critically revised the manuscript. All authors have read and agreed to this version of the manuscript.
Funding
This research received no external funding. S.S.S. is supported by grant 2022/09354-9 from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249-257. [CrossRef]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nature medicine 1995, 1, 27-31. [CrossRef]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873-887. [CrossRef]
- Matsumoto, T.; Claesson-Welsh, L. VEGF receptor signal transduction. Science's STKE : signal transduction knowledge environment 2001, 2001, re21. [CrossRef]
- Zhang, F.; Tang, Z.; Hou, X.; Lennartsson, J.; Li, Y.; Koch, A.W.; Scotney, P.; Lee, C.; Arjunan, P.; Dong, L.; et al. VEGF-B is dispensable for blood vessel growth but critical for their survival, and VEGF-B targeting inhibits pathological angiogenesis. Proceedings of the National Academy of Sciences of the United States of America 2009, 106, 6152-6157. [CrossRef]
- Dvorak, H.F. Vascular permeability factor/vascular endothelial growth factor: a critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 2002, 20, 4368-4380. [CrossRef]
- Swenson, S.; Ramu, S.; Markland, F.S. Anti-angiogenesis and RGD-containing snake venom disintegrins. Current pharmaceutical design 2007, 13, 2860-2871. [CrossRef]
- Usami, Y.; Fujimura, Y.; Miura, S.; Shima, H.; Yoshida, E.; Yoshioka, A.; Hirano, K.; Suzuki, M.; Titani, K. A 28 kDa-protein with disintegrin-like structure (jararhagin-C) purified from Bothrops jararaca venom inhibits collagen- and ADP-induced platelet aggregation. Biochemical and biophysical research communications 1994, 201, 331-339. [CrossRef]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome biology 2007, 8, 215. [CrossRef]
- Rose, D.M.; Alon, R.; Ginsberg, M.H. Integrin modulation and signaling in leukocyte adhesion and migration. Immunological reviews 2007, 218, 126-134. [CrossRef]
- Hynes, R.O. Integrins: bidirectional, allosteric signaling machines. Cell 2002, 110, 673-687. [CrossRef]
- Ruoslahti, E.; Pierschbacher, M.D. New perspectives in cell adhesion: RGD and integrins. Science 1987, 238, 491-497. [CrossRef]
- Lu, X.; Lu, D.; Scully, M.F.; Kakkar, V.V. Integrins in drug targeting-RGD templates in toxins. Current pharmaceutical design 2006, 12, 2749-2769. [CrossRef]
- Marcinkiewicz, C.; Vijay-Kumar, S.; McLane, M.A.; Niewiarowski, S. Significance of RGD loop and C-terminal domain of echistatin for recognition of alphaIIb beta3 and alpha(v) beta3 integrins and expression of ligand-induced binding site. Blood 1997, 90, 1565-1575.
- Kamata, T.; Handa, M.; Sato, Y.; Ikeda, Y.; Aiso, S. Membrane-proximal alpha/beta stalk interactions differentially regulate integrin activation. The Journal of biological chemistry 2005, 280, 24775-24783. [CrossRef]
- Calvete, J.J. Snake venomics: from the inventory of toxins to biology. Toxicon : official journal of the International Society on Toxinology 2013, 75, 44-62. [CrossRef]
- Arruda Macedo, J.K.; Fox, J.W.; de Souza Castro, M. Disintegrins from snake venoms and their applications in cancer research and therapy. Current protein & peptide science 2015, 16, 532-548. [CrossRef]
- Marcinkiewicz, C. Applications of snake venom components to modulate integrin activities in cell-matrix interactions. The international journal of biochemistry & cell biology 2013, 45, 1974-1986. [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9. [CrossRef]
- Fox, J.W.; Serrano, S.M. Timeline of key events in snake venom metalloproteinase research. Journal of proteomics 2009, 72, 200-209. [CrossRef]
- Fox, J.W.; Serrano, S.M. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. The FEBS journal 2008, 275, 3016-3030. [CrossRef]
- Calvete, J.J.; Marcinkiewicz, C.; Monleon, D.; Esteve, V.; Celda, B.; Juarez, P.; Sanz, L. Snake venom disintegrins: evolution of structure and function. Toxicon : official journal of the International Society on Toxinology 2005, 45, 1063-1074. [CrossRef]
- Schonthal, A.H.; Swenson, S.D.; Chen, T.C.; Markland, F.S. Preclinical studies of a novel snake venom-derived recombinant disintegrin with antitumor activity: A review. Biochemical pharmacology 2020, 181, 114149. [CrossRef]
- Calvete, J.J.; Moreno-Murciano, M.P.; Theakston, R.D.; Kisiel, D.G.; Marcinkiewicz, C. Snake venom disintegrins: novel dimeric disintegrins and structural diversification by disulphide bond engineering. The Biochemical journal 2003, 372, 725-734. [CrossRef]
- Marcinkiewicz, C. Functional characteristic of snake venom disintegrins: potential therapeutic implication. Current pharmaceutical design 2005, 11, 815-827. [CrossRef]
- !!! INVALID CITATION !!!
- Clissa, P.B.; Lopes-Ferreira, M.; Della-Casa, M.S.; Farsky, S.H.; Moura-da-Silva, A.M. Importance of jararhagin disintegrin-like and cysteine-rich domains in the early events of local inflammatory response. Toxicon : official journal of the International Society on Toxinology 2006, 47, 591-596. [CrossRef]
- Zychar, B.C.; Clissa, P.B.; Carvalho, E.; Baldo, C.; Goncalves, L.R.C. Leukocyte recruitment induced by snake venom metalloproteinases: Role of the catalytic domain. Biochemical and biophysical research communications 2020, 521, 402-407. [CrossRef]
- Zychar, B.C.; Clissa, P.B.; Carvalho, E.; Alves, A.S.; Baldo, C.; Faquim-Mauro, E.L.; Goncalves, L.R.C. Modulation of Adhesion Molecules Expression by Different Metalloproteases Isolated from Bothrops Snakes. Toxins 2021, 13. [CrossRef]
- Souza, D.H.; Iemma, M.R.; Ferreira, L.L.; Faria, J.P.; Oliva, M.L.; Zingali, R.B.; Niewiarowski, S.; Selistre-de-Araujo, H.S. The disintegrin-like domain of the snake venom metalloprotease alternagin inhibits alpha2beta1 integrin-mediated cell adhesion. Archives of biochemistry and biophysics 2000, 384, 341-350. [CrossRef]
- Mebs, D. Myotoxic activity of phospholipases A2 isolated from cobra venoms: neutralization by polyvalent antivenoms. Toxicon : official journal of the International Society on Toxinology 1986, 24, 1001-1008. [CrossRef]
- Della-Casa, M.S.; Junqueira-de-Azevedo, I.; Butera, D.; Clissa, P.B.; Lopes, D.S.; Serrano, S.M.; Pimenta, D.C.; Magalhaes, G.S.; Ho, P.L.; Moura-da-Silva, A.M. "Insularin, a disintegrin from Bothrops insularis venom: inhibition of platelet aggregation and endothelial cell adhesion by the native and recombinant GST-insularin proteins". Toxicon : official journal of the International Society on Toxinology 2011, 57, 125-133. [CrossRef]
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. Echistatin. A potent platelet aggregation inhibitor from the venom of the viper, Echis carinatus. The Journal of biological chemistry 1988, 263, 19827-19832.
- Kamiguti, A.S.; Zuzel, M.; Theakston, R.D. Snake venom metalloproteinases and disintegrins: interactions with cells. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas 1998, 31, 853-862. [CrossRef]
- Tang, C.H.; Yang, R.S.; Liu, C.Z.; Huang, T.F.; Fu, W.M. Differential susceptibility of osteosarcoma cells and primary osteoblasts to cell detachment caused by snake venom metalloproteinase protein. Toxicon : official journal of the International Society on Toxinology 2004, 43, 11-20. [CrossRef]
- Brooks, S.A.; Lomax-Browne, H.J.; Carter, T.M.; Kinch, C.E.; Hall, D.M. Molecular interactions in cancer cell metastasis. Acta histochemica 2010, 112, 3-25. [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: biological implications and therapeutic opportunities. Nature reviews. Cancer 2010, 10, 9-22. [CrossRef]
- Golubkov, V.; Hawes, D.; Markland, F.S. Anti-angiogenic activity of contortrostatin, a disintegrin from Agkistrodon contortrix contortrix snake venom. Angiogenesis 2003, 6, 213-224. [CrossRef]
- Maiguel, D.; Faridi, M.H.; Wei, C.; Kuwano, Y.; Balla, K.M.; Hernandez, D.; Barth, C.J.; Lugo, G.; Donnelly, M.; Nayer, A.; et al. Small molecule-mediated activation of the integrin CD11b/CD18 reduces inflammatory disease. Science signaling 2011, 4, ra57. [CrossRef]
- Petpiroon, N.; Sritularak, B.; Chanvorachote, P. Correction: Phoyunnanin E inhibits migration of non-small cell lung cancer cells via suppression of epithelial-to-mesenchymal transition and integrin alphav and integrin beta3. BMC complementary medicine and therapies 2023, 23, 196. [CrossRef]
- Jahangiri, A.; Aghi, M.K.; Carbonell, W.S. beta1 integrin: Critical path to antiangiogenic therapy resistance and beyond. Cancer research 2014, 74, 3-7. [CrossRef]
- Liu, F.; Wu, Q.; Dong, Z.; Liu, K. Integrins in cancer: Emerging mechanisms and therapeutic opportunities. Pharmacology & therapeutics 2023, 247, 108458. [CrossRef]
- Olfa, K.Z.; Jose, L.; Salma, D.; Amine, B.; Najet, S.A.; Nicolas, A.; Maxime, L.; Raoudha, Z.; Kamel, M.; Jacques, M.; et al. Lebestatin, a disintegrin from Macrovipera venom, inhibits integrin-mediated cell adhesion, migration and angiogenesis. Laboratory investigation; a journal of technical methods and pathology 2005, 85, 1507-1516. [CrossRef]
- Yeh, C.H.; Peng, H.C.; Huang, T.F. Accutin, a new disintegrin, inhibits angiogenesis in vitro and in vivo by acting as integrin alphavbeta3 antagonist and inducing apoptosis. Blood 1998, 92, 3268-3276.
- Ferreira, B.A.; De Moura, F.B.R.; Tomiosso, T.C.; Correa, N.C.R.; Goulart, L.R.; Barcelos, L.S.; Clissa, P.B.; Araujo, F.A. Jararhagin-C, a disintegrin-like protein, improves wound healing in mice through stimulation of M2-like macrophage, angiogenesis and collagen deposition. International immunopharmacology 2021, 101, 108224. [CrossRef]
- Monteiro, D.A.; Kalinin, A.L.; Selistre-de-Araujo, H.S.; Nogueira, L.A.N.; Beletti, M.E.; Fernandes, M.N.; Rantin, F.T. Cardioprotective effects of alternagin-C (ALT-C), a disintegrin-like protein from Rhinocerophis alternatus snake venom, on hypoxia-reoxygenation-induced injury in fish. Comparative biochemistry and physiology. Toxicology & pharmacology : CBP 2019, 215, 67-75. [CrossRef]
- Marcinkiewicz, C.; Weinreb, P.H.; Calvete, J.J.; Kisiel, D.G.; Mousa, S.A.; Tuszynski, G.P.; Lobb, R.R. Obtustatin: a potent selective inhibitor of alpha1beta1 integrin in vitro and angiogenesis in vivo. Cancer research 2003, 63, 2020-2023.
- Staniszewska, I.; Walsh, E.M.; Rothman, V.L.; Gaathon, A.; Tuszynski, G.P.; Calvete, J.J.; Lazarovici, P.; Marcinkiewicz, C. Effect of VP12 and viperistatin on inhibition of collagen-receptor-dependent melanoma metastasis. Cancer biology & therapy 2009, 8, 1507-1516. [CrossRef]
- Sanz, L.; Chen, R.Q.; Perez, A.; Hilario, R.; Juarez, P.; Marcinkiewicz, C.; Monleon, D.; Celda, B.; Xiong, Y.L.; Perez-Paya, E.; et al. cDNA cloning and functional expression of jerdostatin, a novel RTS-disintegrin from Trimeresurus jerdonii and a specific antagonist of the alpha1beta1 integrin. The Journal of biological chemistry 2005, 280, 40714-40722. [CrossRef]
- Yeh, C.H.; Peng, H.C.; Yang, R.S.; Huang, T.F. Rhodostomin, a snake venom disintegrin, inhibits angiogenesis elicited by basic fibroblast growth factor and suppresses tumor growth by a selective alpha(v)beta(3) blockade of endothelial cells. Molecular pharmacology 2001, 59, 1333-1342. [CrossRef]
- Dos Santos, P.K.; Altei, W.F.; Danilucci, T.M.; Lino, R.L.B.; Pachane, B.C.; Nunes, A.C.C.; Selistre-de-Araujo, H.S. Alternagin-C (ALT-C), a disintegrin-like protein, attenuates alpha2beta1 integrin and VEGF receptor 2 signaling resulting in angiogenesis inhibition. Biochimie 2020, 174, 144-158. [CrossRef]
- Selistre-de-Araujo, H.S.; Cominetti, M.R.; Terruggi, C.H.; Mariano-Oliveira, A.; De Freitas, M.S.; Crepin, M.; Figueiredo, C.C.; Morandi, V. Alternagin-C, a disintegrin-like protein from the venom of Bothrops alternatus, modulates alpha2beta1 integrin-mediated cell adhesion, migration and proliferation. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas 2005, 38, 1505-1511. [CrossRef]
- Limam, I.; Abdelkarim, M.; El Ayeb, M.; Crepin, M.; Marrakchi, N.; Di Benedetto, M. Disintegrin-like Protein Strategy to Inhibit Aggressive Triple-Negative Breast Cancer. International journal of molecular sciences 2023, 24. [CrossRef]
- Zhou, Q.; Nakada, M.T.; Arnold, C.; Shieh, K.Y.; Markland, F.S., Jr. Contortrostatin, a dimeric disintegrin from Agkistrodon contortrix contortrix, inhibits angiogenesis. Angiogenesis 1999, 3, 259-269. [CrossRef]
- Ramos, O.H.; Kauskot, A.; Cominetti, M.R.; Bechyne, I.; Salla Pontes, C.L.; Chareyre, F.; Manent, J.; Vassy, R.; Giovannini, M.; Legrand, C.; et al. A novel alpha(v)beta (3)-blocking disintegrin containing the RGD motive, DisBa-01, inhibits bFGF-induced angiogenesis and melanoma metastasis. Clinical & experimental metastasis 2008, 25, 53-64. [CrossRef]
- Chang, C.H.; Chung, C.H.; Hsu, C.C.; Peng, H.C.; Huang, T.F. Inhibitory effects of polypeptides derived from a snake venom C-type lectin, aggretin, on tumor cell-induced platelet aggregation. Journal of thrombosis and haemostasis : JTH 2014, 12, 540-549. [CrossRef]
- Huang, T.F.; Ouyang, C. Action mechanism of the potent platelet aggregation inhibitor from Trimeresurus gramineus snake venom. Thrombosis research 1984, 33, 125-138. [CrossRef]
- Ren, A.; Wang, S.; Cai, W.; Yang, G.; Zhu, Y.; Wu, X.; Zhang, Y. Agkistin-s, a disintegrin domain, inhibits angiogenesis and induces BAECs apoptosis. Journal of cellular biochemistry 2006, 99, 1517-1523. [CrossRef]
- Takahashi, H.; Hattori, S.; Iwamatsu, A.; Takizawa, H.; Shibuya, M. A novel snake venom vascular endothelial growth factor (VEGF) predominantly induces vascular permeability through preferential signaling via VEGF receptor-1. The Journal of biological chemistry 2004, 279, 46304-46314. [CrossRef]
- Yamazaki, Y.; Matsunaga, Y.; Tokunaga, Y.; Obayashi, S.; Saito, M.; Morita, T. Snake venom Vascular Endothelial Growth Factors (VEGF-Fs) exclusively vary their structures and functions among species. The Journal of biological chemistry 2009, 284, 9885-9891. [CrossRef]
- Ferrara, N. VEGF-A: a critical regulator of blood vessel growth. European cytokine network 2009, 20, 158-163. [CrossRef]
- Chakrabarty, D.; Chanda, C. Snake Venom Disintegrins. In Snake Venoms, Inagaki, H., Vogel, C.-W., Mukherjee, A.K., Rahmy, T.R., Gopalakrishnakone, P., Eds.; Springer Netherlands: Dordrecht, 2017; pp. 437-449.
- Swenson, S.; Costa, F.; Ernst, W.; Fujii, G.; Markland, F.S. Contortrostatin, a snake venom disintegrin with anti-angiogenic and anti-tumor activity. Pathophysiology of haemostasis and thrombosis 2005, 34, 169-176. [CrossRef]
- Cesar, P.H.S.; Braga, M.A.; Trento, M.V.C.; Menaldo, D.L.; Marcussi, S. Snake Venom Disintegrins: An Overview of their Interaction with Integrins. Current drug targets 2019, 20, 465-477. [CrossRef]
- Sanchez, E.E.; Rodriguez-Acosta, A.; Palomar, R.; Lucena, S.E.; Bashir, S.; Soto, J.G.; Perez, J.C. Colombistatin: a disintegrin isolated from the venom of the South American snake (Bothrops colombiensis) that effectively inhibits platelet aggregation and SK-Mel-28 cell adhesion. Archives of toxicology 2009, 83, 271-279. [CrossRef]
- Kumar, C.C.; Malkowski, M.; Yin, Z.; Tanghetti, E.; Yaremko, B.; Nechuta, T.; Varner, J.; Liu, M.; Smith, E.M.; Neustadt, B.; et al. Inhibition of angiogenesis and tumor growth by SCH221153, a dual alpha(v)beta3 and alpha(v)beta5 integrin receptor antagonist. Cancer research 2001, 61, 2232-2238.
- Al-Ostoot, F.H.; Salah, S.; Khamees, H.A.; Khanum, S.A. Tumor angiogenesis: Current challenges and therapeutic opportunities. Cancer treatment and research communications 2021, 28, 100422. [CrossRef]
- Weis, S.M.; Cheresh, D.A. Tumor angiogenesis: molecular pathways and therapeutic targets. Nature medicine 2011, 17, 1359-1370. [CrossRef]
- Akhtar, B.; Muhammad, F.; Sharif, A.; Anwar, M.I. Mechanistic insights of snake venom disintegrins in cancer treatment. European journal of pharmacology 2021, 899, 174022. [CrossRef]
- Kalita, B.; Saviola, A.J.; Mukherjee, A.K. From venom to drugs: a review and critical analysis of Indian snake venom toxins envisaged as anticancer drug prototypes. Drug discovery today 2021, 26, 993-1005. [CrossRef]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta neuropathologica 2007, 114, 97-109. [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a cancer journal for clinicians 2018, 68, 394-424. [CrossRef]
- Dehelean, C.A.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O.M.; Pinzaru, I. Plant-Derived Anticancer Compounds as New Perspectives in Drug Discovery and Alternative Therapy. Molecules 2021, 26. [CrossRef]
- Huang, M.; Lu, J.J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Natural products and bioprospecting 2021, 11, 5-13. [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. Journal of natural products 2020, 83, 770-803. [CrossRef]
- Zhang, Q.Z.; Zhu, Y.P.; Rahat, M.A.; Kzhyshkowska, J. Editorial: Angiogenesis and tumor metastasis. Frontiers in oncology 2022, 12, 1129736. [CrossRef]
- Bielenberg, D.R.; Zetter, B.R. The Contribution of Angiogenesis to the Process of Metastasis. Cancer journal 2015, 21, 267-273. [CrossRef]
- Cross, M.J.; Claesson-Welsh, L. FGF and VEGF function in angiogenesis: signalling pathways, biological responses and therapeutic inhibition. Trends in pharmacological sciences 2001, 22, 201-207. [CrossRef]
- Laddha, A.P.; Kulkarni, Y.A. VEGF and FGF-2: Promising targets for the treatment of respiratory disorders. Respiratory medicine 2019, 156, 33-46. [CrossRef]
- Zhao, Y.; Adjei, A.A. Targeting Angiogenesis in Cancer Therapy: Moving Beyond Vascular Endothelial Growth Factor. The oncologist 2015, 20, 660-673. [CrossRef]
- Ansari, M.J.; Bokov, D.; Markov, A.; Jalil, A.T.; Shalaby, M.N.; Suksatan, W.; Chupradit, S.; Al-Ghamdi, H.S.; Shomali, N.; Zamani, A.; et al. Cancer combination therapies by angiogenesis inhibitors; a comprehensive review. Cell communication and signaling : CCS 2022, 20, 49. [CrossRef]
- Kolvekar, N.; Bhattacharya, N.; Sarkar, A.; Chakrabarty, D. How snake venom disintegrins affect platelet aggregation and cancer proliferation. Toxicon : official journal of the International Society on Toxinology 2023, 221, 106982. [CrossRef]
- Leasher, J.L.; Bourne, R.R.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Naidoo, K.; Pesudovs, K.; Price, H.; White, R.A.; Wong, T.Y.; et al. Global Estimates on the Number of People Blind or Visually Impaired by Diabetic Retinopathy: A Meta-analysis From 1990 to 2010. Diabetes care 2016, 39, 1643-1649. [CrossRef]
- Lee, R.; Wong, T.Y.; Sabanayagam, C. Epidemiology of diabetic retinopathy, diabetic macular edema and related vision loss. Eye and vision 2015, 2, 17. [CrossRef]
- Toh, H.; Smolentsev, A.; Bozadjian, R.V.; Keeley, P.W.; Lockwood, M.D.; Sadjadi, R.; Clegg, D.O.; Blodi, B.A.; Coffey, P.J.; Reese, B.E.; et al. Vascular changes in diabetic retinopathy-a longitudinal study in the Nile rat. Laboratory investigation; a journal of technical methods and pathology 2019, 99, 1547-1560. [CrossRef]
- Stitt, A.W.; Curtis, T.M.; Chen, M.; Medina, R.J.; McKay, G.J.; Jenkins, A.; Gardiner, T.A.; Lyons, T.J.; Hammes, H.P.; Simo, R.; et al. The progress in understanding and treatment of diabetic retinopathy. Progress in retinal and eye research 2016, 51, 156-186. [CrossRef]
- Gupta, N.; Mansoor, S.; Sharma, A.; Sapkal, A.; Sheth, J.; Falatoonzadeh, P.; Kuppermann, B.; Kenney, M. Diabetic retinopathy and VEGF. The open ophthalmology journal 2013, 7, 4-10. [CrossRef]
- Simo, R.; Sundstrom, J.M.; Antonetti, D.A. Ocular Anti-VEGF therapy for diabetic retinopathy: the role of VEGF in the pathogenesis of diabetic retinopathy. Diabetes care 2014, 37, 893-899. [CrossRef]
- Moore, S.W.; Bhat, V.K.; Flatt, P.R.; Gault, V.A.; McClean, S. Isolation and characterisation of insulin-releasing compounds from Crotalus adamanteus, Crotalus vegrandis and Bitis nasicornis venom. Toxicon : official journal of the International Society on Toxinology 2015, 101, 48-54. [CrossRef]
- Ramos, O.H.; Terruggi, C.H.; Ribeiro, J.U.; Cominetti, M.R.; Figueiredo, C.C.; Berard, M.; Crepin, M.; Morandi, V.; Selistre-de-Araujo, H.S. Modulation of in vitro and in vivo angiogenesis by alternagin-C, a disintegrin-like protein from Bothrops alternatus snake venom and by a peptide derived from its sequence. Archives of biochemistry and biophysics 2007, 461, 1-6. [CrossRef]
- Selistre-de-Araujo, H.S.; Pontes, C.L.; Montenegro, C.F.; Martin, A.C. Snake venom disintegrins and cell migration. Toxins 2010, 2, 2606-2621. [CrossRef]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314-321. [CrossRef]
- Zhao, R.; Liang, H.; Clarke, E.; Jackson, C.; Xue, M. Inflammation in Chronic Wounds. International journal of molecular sciences 2016, 17. [CrossRef]
- Schnittert, J.; Bansal, R.; Storm, G.; Prakash, J. Integrins in wound healing, fibrosis and tumor stroma: High potential targets for therapeutics and drug delivery. Advanced drug delivery reviews 2018, 129, 37-53. [CrossRef]
- Ferreira, B.A.; Deconte, S.R.; de Moura, F.B.R.; Tomiosso, T.C.; Clissa, P.B.; Andrade, S.P.; Araujo, F.A. Inflammation, angiogenesis and fibrogenesis are differentially modulated by distinct domains of the snake venom metalloproteinase jararhagin. International journal of biological macromolecules 2018, 119, 1179-1187. [CrossRef]
- Sant'Ana, E.M.; Gouvea, C.M.; Durigan, J.L.; Cominetti, M.R.; Pimentel, E.R.; Selistre-de-Araujo, H.S. Rat skin wound healing induced by alternagin-C, a disintegrin-like, Cys-rich protein from Bothrops alternatus venom. International wound journal 2011, 8, 245-252. [CrossRef]
- Rabelo, L.F.G.; Ferreira, B.A.; Deconte, S.R.; Tomiosso, T.C.; Dos Santos, P.K.; Andrade, S.P.; Selistre de Araujo, H.S.; Araujo, F.A. Alternagin-C, a disintegrin-like protein from Bothrops alternatus venom, attenuates inflammation and angiogenesis and stimulates collagen deposition of sponge-induced fibrovascular tissue in mice. International journal of biological macromolecules 2019, 140, 653-660. [CrossRef]
- Turner, C.J.; Badu-Nkansah, K.; Crowley, D.; van der Flier, A.; Hynes, R.O. alpha5 and alphav integrins cooperate to regulate vascular smooth muscle and neural crest functions in vivo. Development 2015, 142, 797-808. [CrossRef]
- Topol, E.J.; Byzova, T.V.; Plow, E.F. Platelet GPIIb-IIIa blockers. Lancet 1999, 353, 227-231. [CrossRef]
- Hartman, G.D.; Egbertson, M.S.; Halczenko, W.; Laswell, W.L.; Duggan, M.E.; Smith, R.L.; Naylor, A.M.; Manno, P.D.; Lynch, R.J.; Zhang, G.; et al. Non-peptide fibrinogen receptor antagonists. 1. Discovery and design of exosite inhibitors. Journal of medicinal chemistry 1992, 35, 4640-4642. [CrossRef]
- Almeida, G.O.; de Oliveira, I.S.; Arantes, E.C.; Sampaio, S.V. Snake venom disintegrins update: insights about new findings. The journal of venomous animals and toxins including tropical diseases 2023, 29, e20230039. [CrossRef]
- Lang, S.H.; Manning, N.; Armstrong, N.; Misso, K.; Allen, A.; Di Nisio, M.; Kleijnen, J. Treatment with tirofiban for acute coronary syndrome (ACS): a systematic review and network analysis. Current medical research and opinion 2012, 28, 351-370. [CrossRef]
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Junior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Frontiers in pharmacology 2020, 11, 1132. [CrossRef]
- Scarborough, R.M.; Rose, J.W.; Hsu, M.A.; Phillips, D.R.; Fried, V.A.; Campbell, A.M.; Nannizzi, L.; Charo, I.F. Barbourin. A GPIIb-IIIa-specific integrin antagonist from the venom of Sistrurus m. barbouri. The Journal of biological chemistry 1991, 266, 9359-9362.
- Roth, G.A.; Huffman, M.D.; Moran, A.E.; Feigin, V.; Mensah, G.A.; Naghavi, M.; Murray, C.J. Global and regional patterns in cardiovascular mortality from 1990 to 2013. Circulation 2015, 132, 1667-1678. [CrossRef]
- Podrez, E.A.; Byzova, T.V.; Febbraio, M.; Salomon, R.G.; Ma, Y.; Valiyaveettil, M.; Poliakov, E.; Sun, M.; Finton, P.J.; Curtis, B.R.; et al. Platelet CD36 links hyperlipidemia, oxidant stress and a prothrombotic phenotype. Nature medicine 2007, 13, 1086-1095. [CrossRef]
- Venturini, W.; Olate-Briones, A.; Valenzuela, C.; Mendez, D.; Fuentes, E.; Cayo, A.; Mancilla, D.; Segovia, R.; Brown, N.E.; Moore-Carrasco, R. Platelet Activation Is Triggered by Factors Secreted by Senescent Endothelial HMEC-1 Cells In Vitro. International journal of molecular sciences 2020, 21. [CrossRef]
- Huilcaman, R.; Venturini, W.; Fuenzalida, L.; Cayo, A.; Segovia, R.; Valenzuela, C.; Brown, N.; Moore-Carrasco, R. Platelets, a Key Cell in Inflammation and Atherosclerosis Progression. Cells 2022, 11. [CrossRef]
- Bledzka, K.; Smyth, S.S.; Plow, E.F. Integrin alphaIIbbeta3: from discovery to efficacious therapeutic target. Circulation research 2013, 112, 1189-1200. [CrossRef]
- Rivas-Mercado, E.A.; Garza-Ocañas, L. Disintegrins obtained from snake venom and their pharmacological potential. Medicina Universitaria 2017, 19, 32-37. [CrossRef]
- Canas, C.A.; Castano-Valencia, S.; Castro-Herrera, F.; Canas, F.; Tobon, G.J. Biomedical applications of snake venom: from basic science to autoimmunity and rheumatology. Journal of translational autoimmunity 2021, 4, 100076. [CrossRef]
- Cook, J.J.; Huang, T.F.; Rucinski, B.; Strzyzewski, M.; Tuma, R.F.; Williams, J.A.; Niewiarowski, S. Inhibition of platelet hemostatic plug formation by trigramin, a novel RGD-peptide. The American journal of physiology 1989, 256, H1038-1043. [CrossRef]
- Monteiro, D.A.; Kalinin, A.L.; Selistre-de-Araujo, H.S.; Vasconcelos, E.S.; Rantin, F.T. Alternagin-C (ALT-C), a disintegrin-like protein from Rhinocerophis alternatus snake venom promotes positive inotropism and chronotropism in fish heart. Toxicon : official journal of the International Society on Toxinology 2016, 110, 1-11. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



