
Submitted:
12 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
Predatory mites are a type of biological control agent used in many countries against various vegetable pests, particularly spider mites. Despite the significant presence of predatory mites in the Tibetan plateau, there is limited research on their fitness and prey capacity in the area. Herein, four predatory mite species, including one species Neoseiulus californicus and three species from genus Amblyseius (Amblyseius swirskii, Amblyseius barkeri, and Amblyseius cucumeris) were investigated. Specifically, their fitness and prey capacity on two-spotted mites (Tetranychus urticae) at highland conditions were evaluated. Compared to the other three predatory species, A. swirskii had the highest fecundity (Mean ± SEM=11.60 ± 0.34) and the highest pre-adult survival rate (Mean ± SEM = 83.33 ± 3.33%). Since their juvenile Survival Rate (SR) was extremely low (Mean ± SEM =13.33% ± 5.77%), most A. barkeri nymphs died before emergence. Compared to the other three predatory mites, A. swirskii had the highest predation capacity on T. urticae adult mites at 15 d post-release (Mean ± SEM =14.28 ± 2.24). Collectively, these findings imply notable advantages in employing A. swirskii for controlling two-spotted mites in the Tibetan Plateau. Furthermore, based on the results, A. swirskii was the most effective, and A. barkeri was the least effective in controlling two-spotted mites in the Tibetan Plateau. These findings could inform the development of a feasible biological control method based on suitable predatory mite species to manage T. urticae in the Tibetan Plateau, and would be helpful in delaying the rise or emergence of chemical pesticide resistance in two-spotted mites.
Keywords:
fitness
; predatory mite
; biological control
; spider mite
; Tibetan Plateau
1. Introduction
In all mite families, the Tetranychidae family, which encompasses > 1300 species of spider mites (Acari: Tetranychidae), holds a significant position regarding its impact on agricultural production [1]. Specifically, more than 100 pest spider mite species could harm agricultural and forestry crops [2]. The two-spotted mite (Tetranychus urticae), a generalist herbivore that could feed on > 1150 host plant species (including a multitude of important crops such as corn, soybean, tomato, cucumber, and so on) [1,3] is the most widely distributed and destructive spider mite species worldwide. Furthermore, multiple T. urticae populations could strongly resist major acaricides and insecticides in various regions [4]. Since 1983, T. urticae has been widespread in most regions in China and is currently the dominant spider mite species [5,6]. Specifically, in the Tibetian Autonomous Region of China, T. urticae was reported to harm numerous vegetable crops and was also difficult to control [7]. However, few studies on the biological management of spider mites have been conducted in Tibet, the world’s highest plateau.
Based on the drawbacks of insecticide or pesticide resistance and pollution, biological control is considered one of the most environmentally friendly and economical methods for managing insect pests [8]. Predatory mites are one of the most important natural biological control agents. Specifically, phytoseiid mites are vital biological control agents, with Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) as the most successful predatory mite, having been employed in > 50 countries globally [9,10]. Other biological factors used for controlling the disastrous harm caused by two-spotted mites include predatory mite species such as Neoseiulus californicus and many other mites of the genus Amblyseius [11,12,13].
With an average elevation of >4000 m above sea level, the Tibetan Plateau is the highest of China’s three major landform ladders. It is characterized by low temperatures and hypoxia, the most substantial threats to the lives of arthropods and other animals in the area [14]. Invertebrates often develop various useful strategies to combat adverse highland conditions. For example, to combat hypoxia conditions (2% O2), the bean weevil increases the synthesis of metabolites, including carbohydrates, amino acids, and organic acids [15]. Furthermore, since the cytochrome oxidase activity is higher in Tibetan locusts, they show a higher hypoxia tolerance than lowland locusts [16]. Additionally, as a means of surviving the high-latitude environment, the enemy insect Bombus pyrosoma would upregulate energy metabolism genes [17]. Despite these useful insights, there are no recent explorations on the effects of highland conditions on the fitness and prey capacity of predatory mites.
The predator N. californicus was first reported by McGregor in 1954 in California, United States [18,19]. As a polyphagous predator, N. californicus mainly fed on Tetranychid spider mites (especially T. urticae, and other spider mites such as T. evansi) [20,21,22]. It could also consume various small insects, including other mite species or thrips; hence, McMurtry classified it as a type II specialist predatory mite (selective predators of Tetranychid mites) [10].
On the other hand, predatory mites of the genus Amblyseius, popular for feeding on various prey including a multitude of mites from many families (such as Tetranychidae, Eriophyidae, Pyroglyphidae, and so on) and many other arthropods including enormous pest insects (such as whiteflies, thrips, nematodes, and so on) [10,23], were always considered as generalist predators, hence were listed as subtype III-b in type III lifestyle predators by McMurtry [23]. However, although the species N. californicus and other predators also belong to the genus Amblyseius, the impact of highland conditions on them has received little research attention, and their predatory capacity remains largely unknown.
Herein, four predatory mite species, including species N. californicus and three species from the genus Amblyseius (A. swirskii, A. barkeri, and A. cucumeris) were examined to determine the most suitable predatory mite species for controlling T. urticae in the Tibetan Plateau, the world’s highest plateau. The fitness and predatory capacities of the four species under highland conditions were assessed. The results showed that in the Tibetan Plateau, A. swirskii and A. barkeri were the most effective and least effective predatory mites in preying on T. urticae, respectively.
2. Materials and Methods
2.1. Establishment of the T. urticae and predatory mite populations
The T. urticae population was collected from soybean leaves in the solar greenhouse of the Lhasa Agricultural Science and Technology Park (roughly E91°07′, N29°38′). Spider mite species identification was confirmed by COI sequencing [24]. After introducing the spider mites into the laboratory, the isofemale line of T. urticae mites was established and grown on the leaves of kidney beans (Phaseolus vulgaris L.) in controlled conditions. The T. urticae population was used as the target or objective pest mite to determine the prey capacity of the predatory mites.
Herein, four predatory mite species, including A. swirskii, Amblyseius barkeri (Hughes), Amblyseius cucumeris (Oudemans), and N. californicus were examined. All predatory mite species were purchased from Fujian Yanxuan Biological Control Technology Company (Fuzhou, China) and were categorized based on three characteristics: spermatheca, abdominal shield, and dorsal shield [25].
2.2. Fitness experiment for the four predatory mite species
The fitness of all four predatory species was assessed through a life-table experiment after rearing in highland conditions in Lhasa, Tibet Autonomous Region, China (roughly E91°07′, N29°38′) for more than ten generations (approximately six months). The rearing conditions were as follows: 1. Temperatures of 25.5 ± 1°C; 2. Relative Humidity (RH) of 80 ± 5%; 3. A photoperiod of 16:8 h (L: D); and 4. A mean altitude of 3642 m above sea level. First, thirty new 1-day-old emerged female adults and thirty new 1-day-old emerged male adults were selected for each mite species to oviposit for 4 h in one dish (9 cm). Following that, 30 eggs of each species were randomly selected to conduct the subsequent test, and one egg was placed in one dish with a kidney bean leaf (about 7 cm) as a replicate, with each mite species having 30 replicates. In each dish, five T. urticae eggs and five T. urticae nymphs (mixed age stages) were added every five days as a diet. To calculate the juvenile survival and developmental rate of different predatory mite species, the stages of all predatory mites were monitored and recorded every 12 h. Ten predatory mites were regarded as a replicate for juvenile survival calculation. Each mite species experiment was performed in triplicate.
Once a female predatory mite emerged, one newly emerged male adult was introduced to mate with one female adult. Ten female adults of each mite species were randomly selected each day to record fecundity. Given that the pre-adult Survival Rate (SR) of A. barkeri was extremely low, ten 1-day-old female A. barkeri adults were selected from other leaf dishes. Fecundity and hatchability data were first tested for normality (Kolmogorov–Smirnov test) and then homogeneity of group variances (Levene’s test). Subsequently, one-way ANOVA analysis (SPSS 21.0) was employed to determine the variance of fecundity and the developmental rate among all four predatory mites. Given that the SRs did not follow a normal distribution, they were analyzed by a Kruskal–Wallis test, and multiple comparisons were performed using Dunn’s test with Bonferroni correction (SPSS v21.0). The Log-rank (Mantel-Cox) test was used to compare the survival curves of each predatory mite’s female adults (GraphPad Prism v9.0.2).
2.3. Functional response experiment
The functional response experiments on the ability of the four predatory mite species to prey on T. urticae (different stages) were conducted under 25℃ conditions. First, an equivalent amount of T. urticae at three variant stages (egg, nymph, and adult) was selected at six density gradients (1, 3, 5, 7, 9, and 11), and the same number of T. urticae eggs, nymphs, and adults were all placed in a dish, while one 1-day-old female adult predatory mite was placed in the same dish after 24 h of starvation. After 24 h, the predation activities of the predatory mites on different stages of T. urticae were recorded. The preying of predatory mites on different stages of T. urticae was measured using a predator-prey model, specifically the Holling type II functional response [26]:
Where Na = the number of T. urticae mites preyed on, ɑ = the attack rate (proportion of prey captured by each predator per unit of searching time), Th = the handling time (the time it takes for predatory mites to identify, kill, and consume T. urticae mites), N=the prey density, and T = the time it takes for predators to find the prey (in this test, T is 1 d).
2.4. Predator interference by different densities of predatory mites
The predatory abilities of different mites at various densities were measured. Herein, six predatory mite density gradients were used (1, 3, 5, 7, 9, and 11). Each mite species was prepared per each density, and these combinations were replicated three times. For every replicate, forty T. urticae female adults were first put in a leaf dish. One selected density of 1-day-old female adults from a single predatory species was then placed in the same dish, and the predation situation was recorded after 24 h. Predator interference analysis was performed using Watt’s model of the effect of the densities of attacked and attacking species on the number of the attacked (Watt, 1959):
Where A = the number of T. urticae female adults preyed on, ɑ = the attack rate without competition, X = the density of predators per leaf dish, and b = the intraspecific competition parameter.
Data on the predatory capacity, handling time, attack coefficient, and max predatory amount were tested for normality (Kolmogorov–Smirnov test) and homogeneity of group variances (Levene’s test) if they followed a normal distribution and One-way ANOVA analysis (SPSS 21.0) was performed to determine the variance among all four predatory mites. On the other hand, the data were subjected to a Kruskal–Wallis test, and multiple comparisons were performed using Dunn’s test with Bonferroni correction if they followed a non-normal distribution (SPSS 21.0).
3. Results
3.1. Compared to other mites in the highland area N. californicus had the fastest developmental rate
One-way ANOVA was used in this part of the results as all the mites’ developmental time data followed a normal distribution. When the development times of the four predatory mites from eggs to emergence were compared, A. barkeri (Mean ± SEM =14.50 ± 0.61) and A. cucumeris (Mean ± SEM = 13.54 ± 0.15) had a significantly longer juvenile period in the Tibet area. On the other hand, N. californicus had the fastest developmental rate (Mean ± SEM = 9.50 ± 0.16) (P < 0.05, One-way ANOVA, SPSS v21.0), implying that it had an applicability advantage over the remaining three predatory mite species grown in Tibet (Figure 1).
In terms of the specific developmental stages, A. swirskii and N. californicus had similar developmental rates for the egg period and larva stage, showing that they could spend non-significantly different periods to complete both growth stages. On the other hand, A. barkeri and A. cucumeris took significantly longer periods to complete the two growth stages (P < 0.05, One-way ANOVA, SPSS v21.0) (Figure S1A and S1B). Additionally, compared to the other three mite species, N. californicus had the fastest developmental rate for the protonymph stage. On the other hand, although the growth time of N. californicus during the deutonymph stage was significantly shorter than that of A. barkeri and A. cucumeris, it was comparable to that of A. swirskii (Figure S1C and S1D).
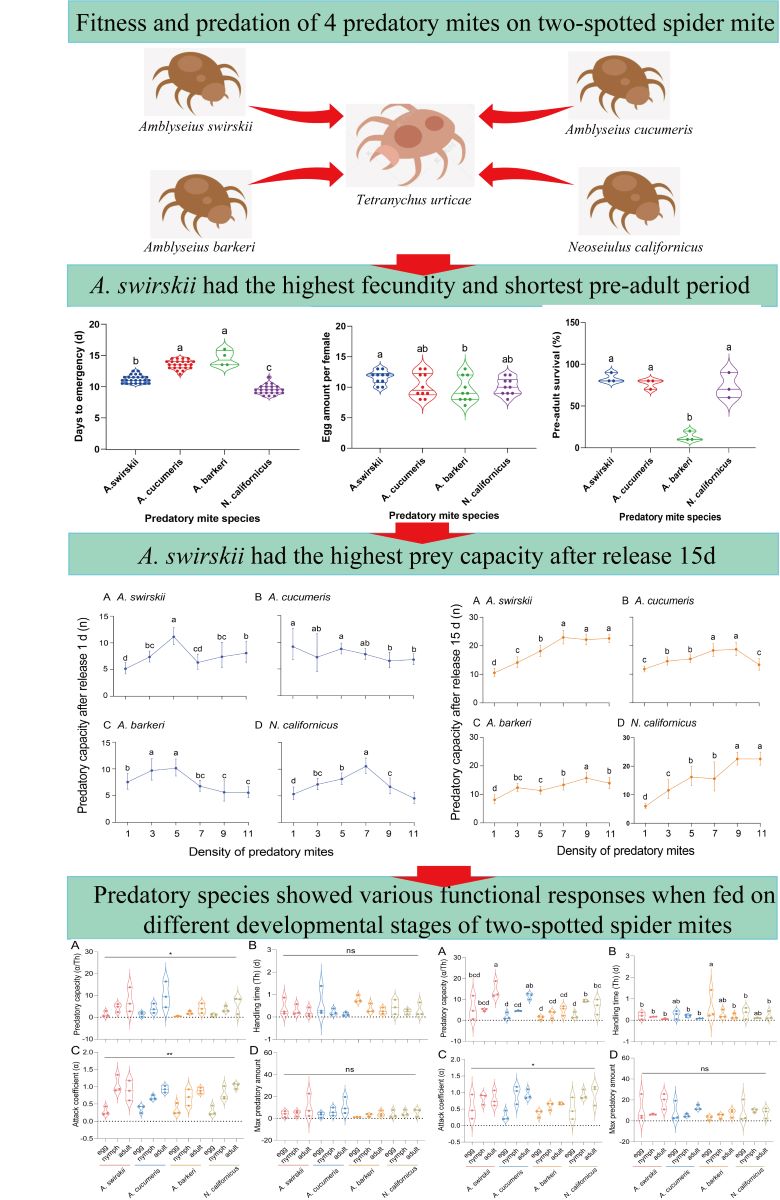
3.2. A. barkeri had relatively less fecundity than A. swirskii
Egg counts of all predatory mites were determined per female animal. Consistent with the developmental time data, all fecundity data followed a normal distribution. According to the results, A. swirskii (Mean ± SEM =11.60 ± 0.65) had a significantly higher fecundity compared to the other four predatory species, followed by A. barberi (Mean ± SEM = 9.60 ± 0.34) (P < 0.05, One-way ANOVA). Compared to those of A. swirskii or A. barberi, the fecundities of A. cucumeris (Mean ± SEM = 10.30 ± 0.633) and N. californicus (Mean ± SEM =10.10 ± 0.43) were non-significantly different (Figure 2).
3.3. Most A. barkeri mites died before emergence in Tibet
During rearing in the Tibet plateau, A. swirskii had the highest pre-adult SR (Mean ± SEM =83.33% ± 5.77%), while A. barberi had the lowest SR (Mean ± SEM =13.33% ± 5.77%), significantly lower than that of the other three predatory species (P < 0.05, Kruskal–Wallis test and Dunn’s test). At the same time, the pre-adult SRs of A. cucumeris and N. californicus were not significantly different compared to that of A. swirskii (Figure 3).
3.4. All four predatory mite species shared similar longevities in high altitudes
Each species’ longevity from mite emergence to death was measured to further clarify the influence of highland conditions on the life span of the four predatory species. According to the results, there were no significant differences among all four predatory species (P = 0.13, Log-rank (Mantel-Cox) test), but A. barberi had the greatest longevity (Median survival = 38 d). (Figure 4).
3.5. The predatory capacity of predatory mites varied at different densities under different release times
The prey capacities of the four predatory mite species with different predator densities were measured at 1 d and 15 d post-release. At 1 d post-release, most species exhibited the highest predatory capacity on T. urticae mites at density 5. Specifically, A. swirskii had the highest predatory capacity at density 5 (Mean ± SD = 11.20 ± 1.30), and other two species; A. barkeri (Mean ± SD=10.20 ± 1.30) and N. californicus (Mean ± SD = 8.20 ± 0.84,) also had a high predatory capacity at the same density. On the other hand, A. cucumeris had the highest predatory capacity on spider mites at density 1 (Mean ± SD = 9.40 ± 2.07) (Figure 5).
At 15 d post-release, the highest predatory capacity was observed at density 9 for all mites except A. swirskii, which had the highest capacity at density 7 (Mean ± SD = 23.00 ± 1.87). Compared to A. cucumeris and A. barkeri, N. californicus had a higher predatory capacity at density 9 (Mean ± SD = 22.60 ± 1.82) (Figure 6). At the same time, data in Figure 5 and Figure 6 successfully fitted into the Holling II disc equation and the Attack coefficient (α), Handling time (Th (d)), and Maximum predation capacity (1/ Th) results are all summarized in Table 1 (1 d post-release) and Table 2 (15 d post-release). Notably, the predatory mites’ prey capacity was always significantly higher at 15 d post-release than at 1d post-release (Figure 7).
3.6. Four predatory species showed various functional responses at different developmental stages of two-spotted spider mites, whether at 1 d or 15 d after release
The functional response experimental results were subjected to the Holling type II functional response analysis to obtain the functional response models of each predatory mite species’ prey capacities on different stages of T. urticae. All experiments were performed in triplicate (Table 1, Table 2, Table 3, Table 4, Table 5 and Table 6). At 1 d post-release, data on the predatory capacity of all four predatory mite species did not follow a normal distribution. Consequently, nonparametric analysis was employed, showing that the predatory capacities of all four predatory mite species were significantly different (P < 0.05, Kruskal–Wallis tests). However, multiple comparisons through Dunn’s tests with Bonferroni correction showed no significant difference between any two groups, with A. cucumeris (Mean ± SEM = 10.14 ± 3.41) having the highest predatory capacity on adult T. urticae mites, and A. swirskii (Mean ± SEM = 7.58 ± 3.25) also having a high predatory capacity on adult T. urticae mites (Figure 8A). Additionally, the four predatory mite species’ data on handling time and attack coefficient did not follow a normal distribution. Although no statistically significant difference was detected among the four species in handling time (P = 0.293, Kruskal–Wallis tests) (Figure 8B), there was a statistically significant difference between them in the attack coefficient data (P < 0.01, Kruskal–Wallis tests). On the other hand, Dunn’s tests with Bonferroni correction for multiple comparisons revealed no statistically significant difference in the attack coefficient data (Figure 8C). The Max predatory amount data of all four predatory mite species followed a normal distribution, but with no significant difference [F (11, 24) = 1.272, P = 0.298, one-way ANOVA] (Figure 8D).
At 15 d post-release, the predatory mites’ data on predatory capacity, handling time, and max predatory amount all followed a normal distribution. A statistically significant difference was found in all mites’ predatory capacity [F (11, 24) = 7.473, P < 0.001, one-way ANOVA]. Specifically, A. swirskii (Mean ± SEM = 14.28 ± 2.24) had the highest predatory capacity on adult T. urticae mites, with A. cucumeris (Mean ± SEM =11.62 ± 1.48) also having a relatively high predatory capacity on adult T. urticae mites. On the other hand, A. barkeri had the lowest predatory capacity on various stages of T. urticae mites (Figure 9A). No statistically significant difference was found in handling time [F (11, 24) = 1.245, P = 0.312, one-way ANOVA] (Figure 9B). The attack coefficient data did not follow a normal distribution, and a significant difference was found in the attack coefficients among the four predatory mite species (P < 0.05, Kruskal–Wallis tests). On the other hand, Dunn’s tests with Bonferroni correction for multiple comparisons found no statistically significant difference in the attack coefficient data (Figure 9C). There was no significant difference detected in mite species’ max predatory amount data [F (11, 24) = 1.385, P = 0.242, one-way ANOVA] (Figure 9D).
4. Discussion
Herein, the fitness and prey capacity of four predatory mite species (A. swirskii, A. barkeri, A. cucumeris, and N. californicus) were assessed using the life-table and functional response experiments. According to the results, A. swirskii had the highest fecundity and pre-adult SR, N. californicus took the least time in transitioning from the egg stage to emergence, and A. barkeri could not survive in highland conditions, as most of its nymphs died before emergence. Furthermore, based on the functional response results, A. swirskii had the highest prey capacity on T. urticae mites compared to the other three predatory species on many parameters. Our findings collectively implied that among the four tested groups, A. swirskii could be the best predatory mite species to biologically control two-spotted mites in the Tibetan Plateau.
The two-spotted mite (T. urticae) is one of the world’s most disastrous agricultural arthropod pests, damaging a wide range of host plants, including many vital crops such as vegetables, cotton, maize, and so on [27,28]. In China, chemical acaricides and insecticides were the primary methods for controlling T. urticae. However, over the past few decades, T. urticae rapidly developed a global resistance to these chemical agents, especially in China [29,30,31,32]. In Tibet, the world’s largest highland, chemical pesticides were widely applied to combat pests, and the resulting significant residues became an issue of concern [33]. Interestingly, no treatments other than chemical control were used in Tibet to effectively prevent the harm caused by two-spotted mites. Despite being important biological control agents against many pest species, including spider mites, predatory mites were rarely used in Tibet, and pertinent research was also extremely limited even in our studies; hence, the fitness and predatory capacity of many predatory mites on T. urticae in this region remained unclear [33,34,35]. Adding to the literature, this study comprehensively explored the fitness and predatory capacity of the four widely used predatory mite species in the Tibetan area.
Multiple studies have reported that high-attitude conditions could significantly influence arthropods’ development, morphological characteristics, and body size, among many other aspects [36,37,38,39,40]. For example, a significant link was found between insects’ body size and altitude in a study on wing-reduced stonefly in New Zealand [40]. Furthermore, insects’ fitness was found to decrease with increasing elevation, as demonstrated by the reduced fecundity of the willow leaf beetle (Chrysomela aeneicollis) under high altitude conditions [41]. Herein, we confirmed that a high attitude significantly adversely affected A. barkeri, resulting in an extremely low SR before emergence (13.33% ± 5.77%), whereas the other three predator species had a better SR as they all had a relatively better fitness, especially the A. swirskii female adult mites, which had the highest fecundity and pre-adult SR (Figure 2 and Figure 3).
It has been reported that A. swirskii is one of the most successful biocontrol agents globally, as it can prey on many severe pests and is easy to rear [9]. Furthermore, another study in China indicated that A. swirskii had an advantage in the predation of whiteflies over other two predatory mite species; Amblyseius orientalis and Neoseiulus californicus [42]. These results are consistent with our findings, as A. swirskii showed a significantly higher predatory capacity on T. urticae than N. californicus at 15 d post-release (Figure 9A), and its fitness in Tibet was comparable to that of similar strains reared under normal conditions [43]. As for N. californicus, our results indicated that although most fitness parameters including developmental time, pre-adult SR, and longevity in Tibet were comparable to those of mites reared in low altitude conditions [44,45,46], its fecundity in Tibet (Mean ± SEM = 10.10 ± 0.43) was significantly lower compared to that under relatively lower elevation laboratory conditions (Mean ± SEM =38.31±2.52) [44]. This finding was unsurprising as, just like other arthropods, mites’ fitness could be influenced by many abiotic and biotic factors [44,47,48].
Phytoseiid mites, including N. californicus are widely distributed biological agents for controlling spider mites. They have been used for decades and have attracted substantial attention given their satisfactory capacity to control the harm caused by T. urticae [44,49,50,51]. Notably, the prey capacity of Phytoseiid mites on spider mites varies under various conditions, including different temperature ranges [47,51], biotic factors such as host plants and prey quality and prey densities [23]. Here, we elucidated the influence of high altitudes on the prey capacities of four predatory mite species and deduced that compared to the other three predators, A. swirskii has the highest predation capacity on adult T. urticae mites at 15 d post-release (Mean ± SEM = 14.28 ± 2.24), indicating an applicability advantage over the other three species in controlling two-spotted mites in the Tibetan Plateau.
Overall, four predatory species, including one species N. californicus, and three species from the genus Amblyseius (A. swirskii, A. barkeri, and A. cucumeris) were investigated herein to compare their fitness and prey capacities on T. urticae in the Tibetan Plateau. We found that A. swirskii had a relatively higher fitness and better predatory capacity than the other predatory mites for controlling T. urticae in a highland area. Furthermore, we discovered that the density of nine predatory mites per plant was the most effective in controlling spider mites and that A. barkeri was an unsuitable biological control agent in highland conditions. Our findings could inform the development of feasible biological control methods for managing T. urticae in the Tibetan Plateau using suitable predatory mite species. They could also be useful in delaying the emergence or increase of resistance to chemical pesticides in two-spotted mites.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, D.X. and K.Y.; methodology, D.X., K.Y. and H.-H.Z.; software, W.Z. and K.Y.; validation, K.Y.; formal analysis, L.X. and K.Y.; data curation, K.Y.; writing-original draft preparation, D.X.; writing-review and editing, K.Y. and Y.-C.W.; visualization, K.Y.; supervision, D.X. and K.Y.; funding acquisition, D.X. and K.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Qingdao Agricultural University High-level Talent Fund (663-1121025), Lhasa Comprehensive Experimental Station of National Technical System of Bulk Vegetable Industry of China, CARS-23-G39, The Central Guide Local Projects of China (XZ202301YD0042C), The Central Guide Local Projects of China (XZ202001YD0014C), ZUNSHI BS [2023] 5
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article and Supplementary Materials.
Acknowledgments
The authors would like to thank all the editors and reviewers for the modifications and suggestions regarding the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider Mites Web: A comprehensive database for the Tetranychidae. Trends Acarol. 2011, 557–560. [Google Scholar]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Dermauw, W.; Grbic, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2013, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Ding, X.L.; Zhang, J.P.; Hong, X.Y. Tetranychus urticae (green form) on Gossypium hirsutum in China: two records confirmed by aedeagus morphology and RFLP analysis. Syst. Appl. Acarol. 2013, 18, 239–244. [Google Scholar]
- Lu, W.; Wang, M.; Xu, Z.; Shen, G.; Wei, P.; Li, M.; Reid, W.; He, L. Adaptation of acaricide stress facilitates Tetranychus urticae expanding against Tetranychus cinnabarinus in China. Ecol Evol. 2017, 7, 1233–1249. [Google Scholar] [CrossRef]
- Dai, W.A.; Yang, J.; Luo, B.; Chen, H.Q.; Zhou, J.; Luo, Y.; Wang, Y.X. Regularity of outbreak and prevention of carmine spider mite in Tibet. Tibet Journal of Agricultural Sciences 2011, 33, 27–29. (in Chinese). [Google Scholar]
- Cock, M.J.W.; van Lenteren, J.C.; Brodeur, J.; Barratt, B.I.P.; Bigler, F.; Bolckmans, K.; Consoli, F.I.; Haas, F.; Mason, P.G.; Parra, J.R.P. Do new access and benefit sharing procedures under the convention on biological diversity threaten the future of biological control? Biocontrol 2010, 55, 199–218. [Google Scholar] [CrossRef]
- Calvo, F.J.; Knapp, M.; van Houten, Y.M.; Hoogerbrugge, H.; Belda, J.E. Amblyseius swirskii: what made this predatory mite such a successful biocontrol agent? Exp. Appl. Acarol. 2015, 65, 419–433. [Google Scholar] [CrossRef]
- McMurtry, J.A.; Croft, B.A. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol. 1997, 42, 291–321. [Google Scholar] [CrossRef]
- Badii, M.H.; Hernández-Ortiz, E.; Flores, A.E.; Landeros, J. Prey stage preference and functional response of Euseius hibisci to Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae). Exp. Appl. Acarol. 2004, 34, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, J.S.; Schausberger, P.; Croft, B.A. Prey-stage preference in generalist and specialist phytoseiid mites (Acari: Phytoseiidae) when offered Tetranychus urticae (Acari: Tetranychidae) eggs and larvae. Environ. Entomol. 2001, 30, 1103–1111. [Google Scholar] [CrossRef]
- Liburd, O.E.; White, J.C.; Rhodes, E.M.; Browdy, A.A. The residual and direct effects of reduced-risk and conventional miticides on two-spotted spider mites, Tetranychus urticae (Acari: Tetranychidae), and predatory mites (Acari: Phytoseiidae). Fla. Entomol. 2007, 90, 249–257. [Google Scholar] [CrossRef]
- Zheng, D.; Zhao, DS. Characteristics of natural environment of the Tibetan Plateau. Science and Technology Review 2017, 35, 13–22. (in Chinese). [Google Scholar]
- Cui, S.F.; Wang, L.; Qiu, J.P.; Liu, Z.C.; Geng, X.Q. Comparative metabolomics analysis of Callosobruchus chinensis larvae under hypoxia, hypoxia /hypercapnia and normoxia. Pest Manag. Sci. 2017, 73, 1267-1276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Chen, B.; Zhao, D.J.; Kang, L. Functional modulation of mitochondrial cytochrome c oxidase underlies adaptation to high-altitude hypoxia in a Tibetan migratory locust. Proc. Royal Soc. B. 2013, 280, 20122758. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, H.; Luo, Q.; Yang, Y.; Zhang, G.; Zhou, Z.; Naeem, M.; An, J. De novo transcriptomic and metabolomic analyses reveal the ecological adaptation of high-altitude Bombus pyrosoma. Insects 2020, 11, 631. [Google Scholar] [CrossRef]
- Barber, A.; Campbell, C.A.M.; Crane, H.; Lilley, R.; Tregidga, E. Biocontrol of two-spotted spider mite Tetranychus urticae on dwarf hops by the phytoseiid mites Phytoseiulus persimilis and Neoseiulus californicus. Biocontrol Sci. Technol. 2003, 13, 275–284. [Google Scholar] [CrossRef]
- Fraulo, A.B.; McSorley, R.; Liburd, O.E. Effects of the biological control agent Neoseiulus californicus (Acari: Phytoseiidae) on arthropod community structures in north Florida strawberry fields. Fla. Entomol. 2008, 91, 336–345. [Google Scholar] [CrossRef]
- Elmoghazy, M.M.E.; El-Seidy, E.M.A.; Romeith, A.H.M. Integrated control of the two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) on faba bean Vicia faba (L.) in an open field at Behaira Governorate, Egypt. Int. J. Environ. Sci. Eng. 2011, 2, 93–100. [Google Scholar]
- Escudero, L.A.; Ferragut, F. Life-history of Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) on four spider mite species as prey, with special reference to Tetranychus evansi (Acari: Tetranychidae). Biol. Control 2005, 32, 378–384. [Google Scholar] [CrossRef]
- Toldi, M.; Ferla, N.J.; Dameda, C.; Majelo, F. Biology of Neoseiulus californicus feeding on two-spotted spider mite. Biotemas. 26, 105–111. [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourassou, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef]
- Ovalle, T.M.; Vásquez-Ordóñez, A.A.; Jimenez, J.; Parsa, S.; Cuellar, W.J.; L opez-Lavalle, L.A.B. A simple PCR-based method for the rapid and accurate identification of spider mites (Tetranychidae) on cassava. Sci. Rep. 2020, 10, 19496. [Google Scholar] [CrossRef]
- Sun, B.; Zhang, Y.K.; Xue, X.F.; Li, Y.X.; Hong, X.Y. Effects of Wolbachia infection in Tetranychus urticae (Acari: Tetranychidae) on predation by Neoseiulus cucumeris (Acari: Phytoseiidae). Syst. Appl. Acarol. 2015, 20, 591–602. [Google Scholar]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Ilias, A.; J. ; Tsagkarakou, V.A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. A bioassay for evaluation of the resistance of Tetranychus urticae (Acari: Tetranychidae) to selected acaricides. Syst. Appl. Acarol. 2015, 20, 579–590. [Google Scholar]
- Kwon, D.H.; Song, D.Y.; Kang, S.; Ahn, J.J.; Lee, J.H.; Choi, B.R.; Lee, S.W.; Kim, J.H.; Lee, S.H. Residual contact vial bioassay for the on-site detection of acaricide resistance in the two-spotted spider mite. J. Asia-Pacific Entomol. 2010, 13, 333–337. [Google Scholar] [CrossRef]
- Xu, D.D.; He, Y.Y.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. Status of pesticide resistance and associated mutations in the two-spotted spider mite, Tetranychus urticae, in China. Pestic. Biochem. Physiol. 2018, 150, 89–96. [Google Scholar] [CrossRef]
- Vassiliou, V.A.; Kitsis, P. Acaricide resistance in Tetranychus urticae (Acari: Tetranychidae) populations from Cyprus. J. Eco. Entomol. 2013, 106, 1848–1854. [Google Scholar] [CrossRef]
- Wang, X.G.; Xiang, X.; Yu, H.L.; Liu, S.H.; Yin, Y.; Cui, P.; Wu, Y.Q.; Yang, J.; Jiang, C.X.; Yang, Q.F. Monitoring and biochemical characterization of beta-cypermethrin resistance in Spodoptera exigua (Lepidoptera: Noctuidae) in Sichuan Province, China. Pestic. Biochem. Physiol. 2018, 146, 71–79. [Google Scholar] [CrossRef]
- Fan, R.Y.; Wang, B.H.; Zai, Q.; Wang, W.F.; Zhao, Y.Q.; Wei, Q.; et al. Composition and occurrence of wheat pests and green control technology in Tibet. Tibet Journal of Agricultural Science 2019, 41, 133–137. (in Chinese). [Google Scholar]
- 34. Deqingzhuoga, Xiang, D.; Nima, Y.Z.; Wang, Z.; Chen, H.Q. Study on the characteristics and green prevention and control technology of vegetable pests in plateau. Tibet Journal of Agricultural Science 2019, 4, 81–84. (in Chinese).
- Xiang, D.; Huang, H.J.; Wang, Z. ; Deqingzhuoga. Preliminary report on control effect of several predator mites on leaf mites of fruit trees. Tibet Journal of Agricultural Science 2019, 2, 39–42. (in Chinese). [Google Scholar]
- Atkinson, D.; Sibly, R.M. Why are organisms usually bigger in colder environments? Making sense of a life history puzzle. Trends. Ecol. Evol. 1997, 12, 235–239. [Google Scholar] [CrossRef]
- Dahlhoff, E.P.; Dahlhoff, V.C.; Grainger, C.A.; Zavala, N.A.; Otepola-Bello, D.; Sargent, B.A.; Roberts, K.T.; Heidl, S.J.; Smiley, J.T.; Rank, N.E. Getting chased up the mountain: High elevation may limit performance and fitness characters in a montane insect. Funct. Ecol. 2019, 33, 809–818. [Google Scholar] [CrossRef]
- Gaston, K.J.; Chown, S.L.; Evans, K.L. Ecogeographical rules: Elements of a synthesis. J. Biogeogr. 2008, 35, 483–500. [Google Scholar] [CrossRef]
- Greenslade, A.F.C.; Chapman, J.W.; Reynolds, D.R. High-altitude migration of Psylloidea (Hemiptera) over England. Entomol. Gaz. 2021, 72, 189–198. [Google Scholar] [CrossRef]
- McCulloch, G.A.; Waters, J.M. Does wing reduction influence the relationship between altitude and insect body size? A case study using New Zealand's diverse stonefly fauna. Ecol. Evol. 2018, 8, 953–960. [Google Scholar] [CrossRef]
- Dahlhoff, E.P.; Dahlhoff, V.C.; Grainger, C.A.; Zavala, N.A.; Otepola-Bello, D.; Sargent, B.A.; Roberts, K.T.; Heidl, S.J.; Smiley, J.T.; Rank, N.E. Getting chased up the mountain: High elevation may limit performance and fitness characters in a montane insect. Funct. Ecol. 2019, 33, 809–818. [Google Scholar] [CrossRef]
- Guo, Y.; Lv, J.; Jiang, X.; Wang, B.; Gao, Y.; Wang, E.; Xu, X. Intraguild predation between Amblyseius swirskii and two native Chinese predatory mite species and their development on intraguild prey. Sci Rep. 2016, 6, 22992. [Google Scholar] [CrossRef]
- Midthassel, A.; Leather, S.R.; Wright, D.J.; Baxter, I.H. Compatibility of Amblyseius swirskii with Beauveria bassiana: two potentially complimentary biocontrol agents. BioControl 2016, 61, 437–447. [Google Scholar] [CrossRef]
- Khanamani, M.; Fathipour, Y.; Talebi, A.A.; Mehrabadi, M. Linking pollen quality and performance of Neoseiulus californicus (Acari: Phytoseiidae) in two-spotted spider mite management programmes. Pest Manag. Sci. 2017, 73, 452–461. [Google Scholar] [CrossRef]
- Maroufpoor, M.; Ghoosta, Y.; Pourmirza, A.A. Life table parameters of Neoseiulus californicus (Acari: Phytoseiidae), on the European red mite, Panonychus ulmi (Acari: Tetranychidae) in laboratory condition. Persian J. Acarol. 2013, 2, 265–276. [Google Scholar]
- Rahmani, H.; Fathipour, Y.; Kamali, K. Life history and population growth parameters of Neoseiulus californicus (Acari: Phytoseiidae) fed on Thrips tabaci (Thysanoptera: Thripidae) in laboratory conditions. Syst. Appl. Acarol. 2009, 14, 91–100. [Google Scholar] [CrossRef]
- Ghazy, N.A.; Suzuki, T.; Amano, H. Development and reproduction of Neoseiulus californicus (Acari: Phytoseiidae) and Tetranychus urticae (Acari: Tetranychidae) under simulated natural temperature. Environ. Entomol. 2018, 47, 1005–1012. [Google Scholar] [CrossRef]
- Marafeli, P.P.; Reis, P.R.; Silveira, E.C.; Souza-Pimentel, G.C.; de Toledo, M.A. Life history of Neoseiulus californicus (McGregor, 1954) (Acari: Phytoseiidae) fed with castor bean (Ricinus communis L.) pollen in laboratory conditions. Braz, J, Biol. 2014, 74, 691–697. [Google Scholar] [CrossRef]
- Greco, N.M.; Sánchez, N.E.; Liljesthröm, G.G. Neoseiulus Californicus (Acari: Phytoseiidae) as a potential control agent of Tetranychus urticae (Acari: Tetranychidae): effect of pest/predator ratio on pest abundance on strawberry. Exp. Appl. Acarol. 2005, 37, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Olaniyi, O.G.; Rhodes, E.M.; Chase, C.A.; Liburd, O.E. The effect of summer cover crops and strawberry cultivars on the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae) and the predatory mite, Neoseiulus californicus (Acari: Phytoseidae) in organic strawberry production systems in Florida. J. Econ. Entomol. 2021, 114, 2135–2146. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Nguyen, D.T.; Audenaert, J.; Verhoeven, R.; Gobin, B.; Tirry, L.; Clercq, P.D. Prey consumption by phytoseiid spider mite predators as affected by diurnal temperature variations. BioControl 2015, 60, 595–603. [Google Scholar] [CrossRef]
Figure 1.
Developmental time of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
Figure 1.
Developmental time of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.

Figure 2.
Fecundity of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
Figure 2.
Fecundity of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
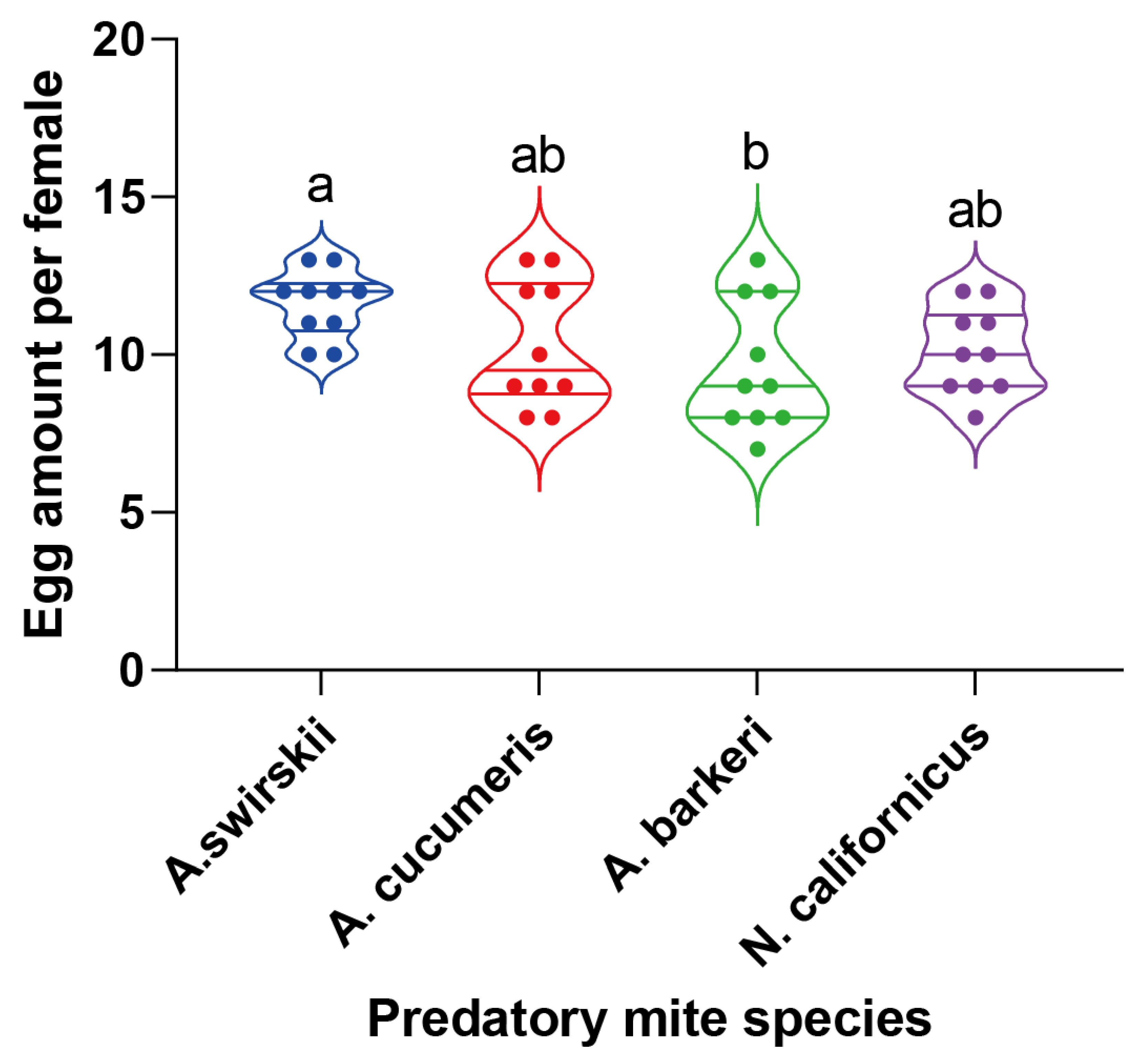
Figure 3.
Pre-adult survival rate of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following by Kruskal–Wallis tests and Dunn’s tests with Bonferroni correction for multiple comparisons.
Figure 3.
Pre-adult survival rate of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following by Kruskal–Wallis tests and Dunn’s tests with Bonferroni correction for multiple comparisons.
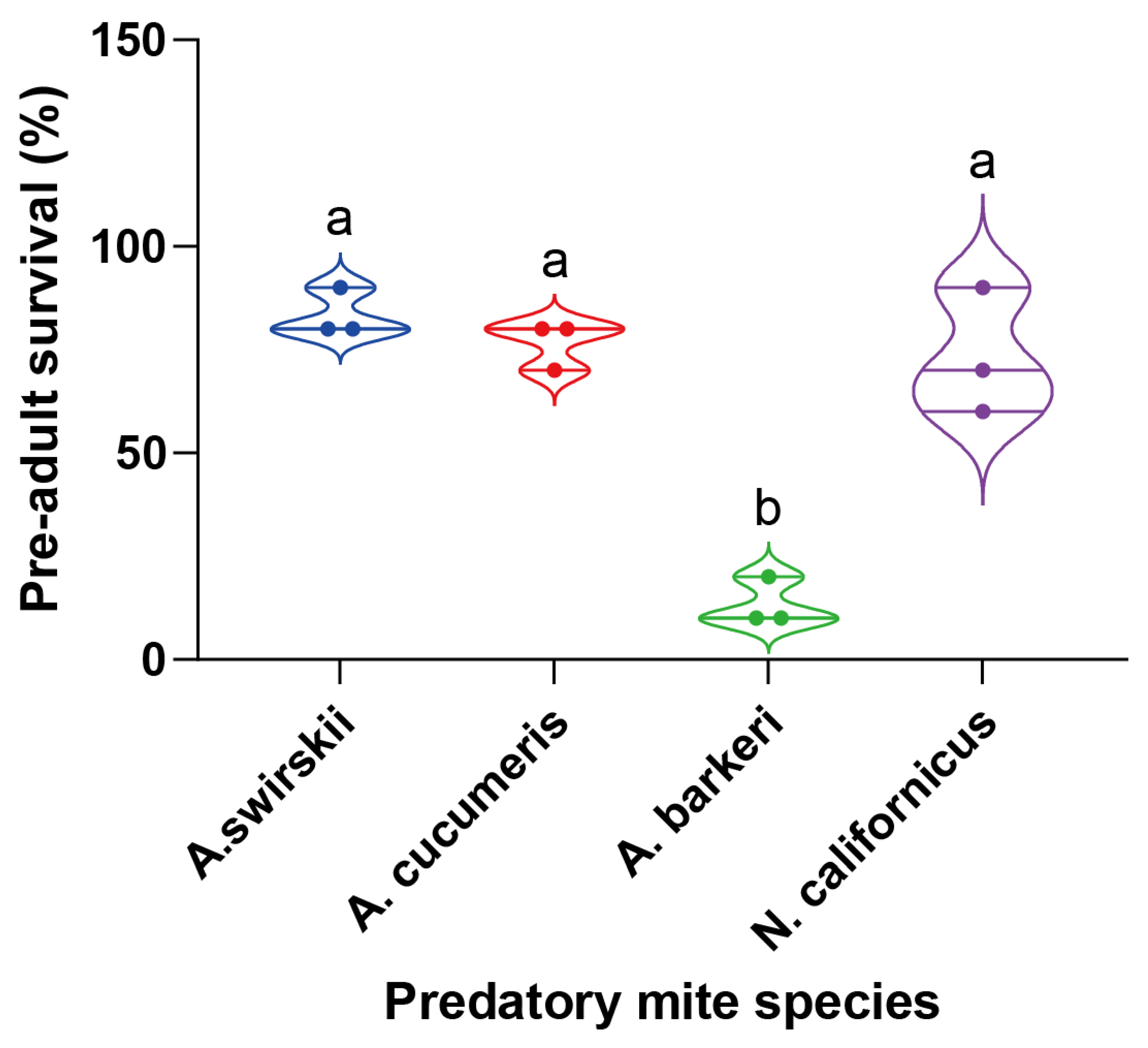
Figure 4.
Longevity of four predatory mite species in Tibetan Plateau. Survival curves for individual hosts were compared using the Kaplan–Meier method and log-rank test.
Figure 4.
Longevity of four predatory mite species in Tibetan Plateau. Survival curves for individual hosts were compared using the Kaplan–Meier method and log-rank test.
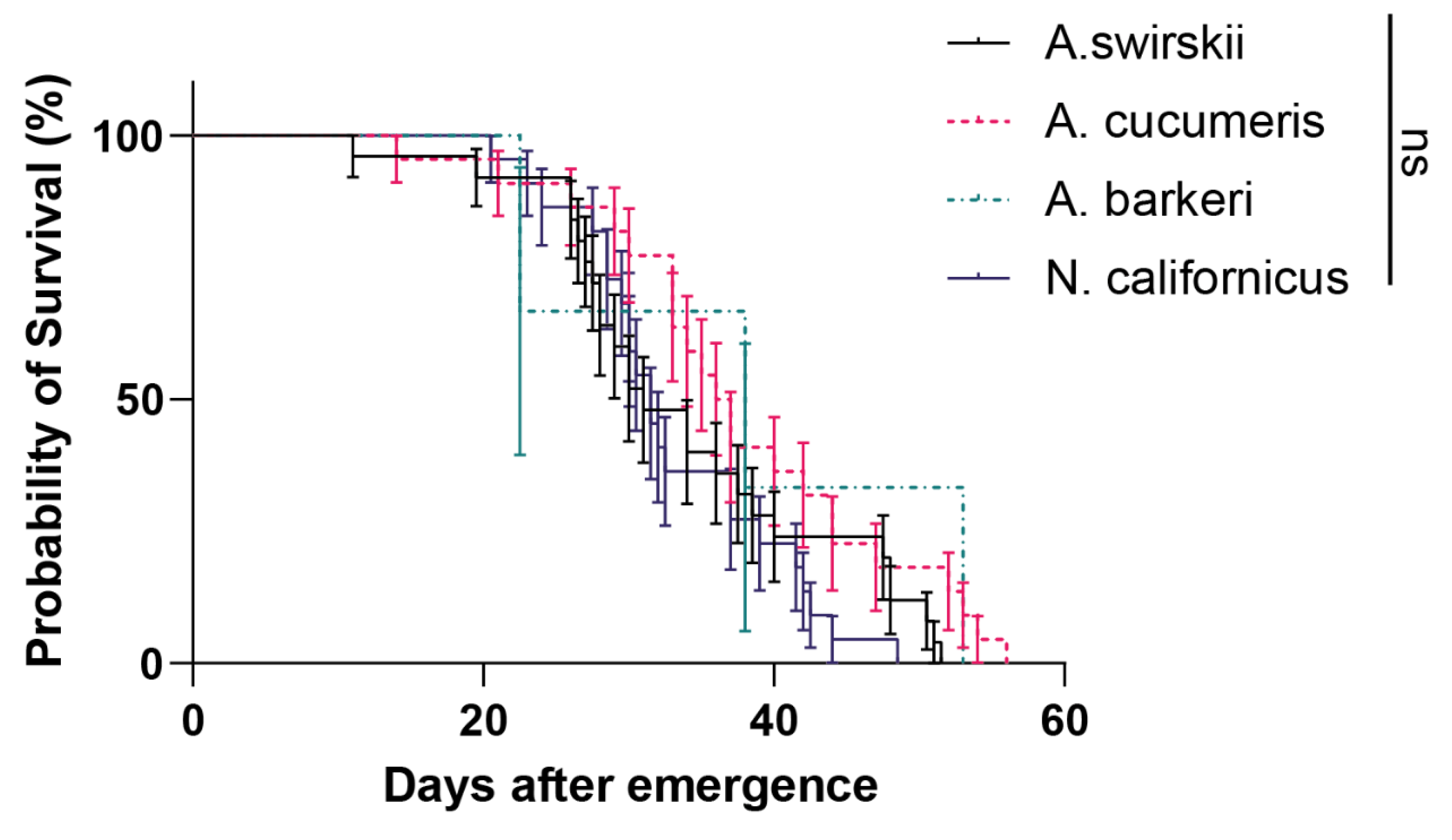
Figure 5.
Predatory capacity (preyed spider mites per predator) after release for 1 d of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
Figure 5.
Predatory capacity (preyed spider mites per predator) after release for 1 d of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
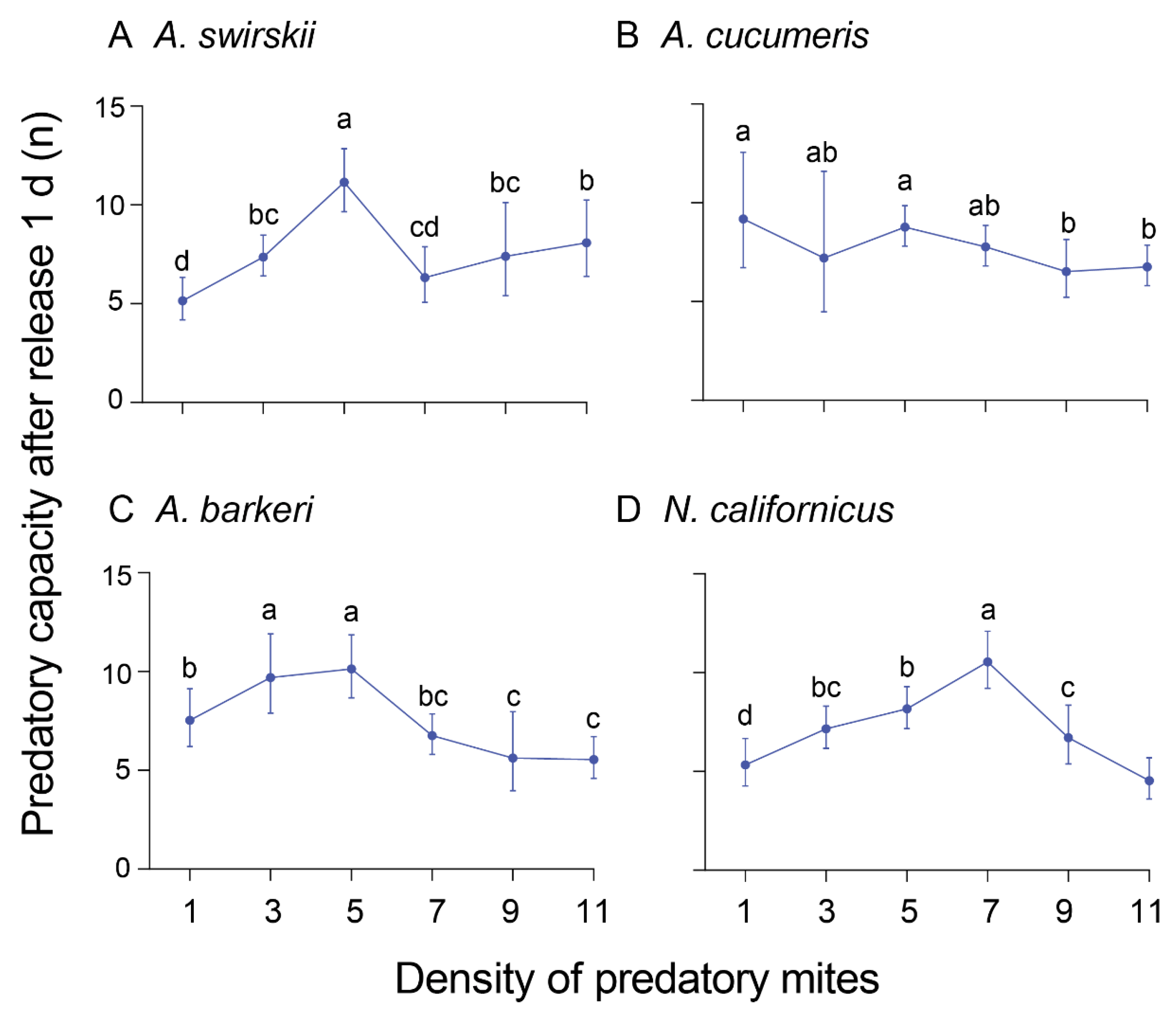
Figure 6.
Predatory capacity (preyed spider mites per predator) after release for 15 d of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
Figure 6.
Predatory capacity (preyed spider mites per predator) after release for 15 d of four predatory mite species in Tibetan Plateau. The same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA.
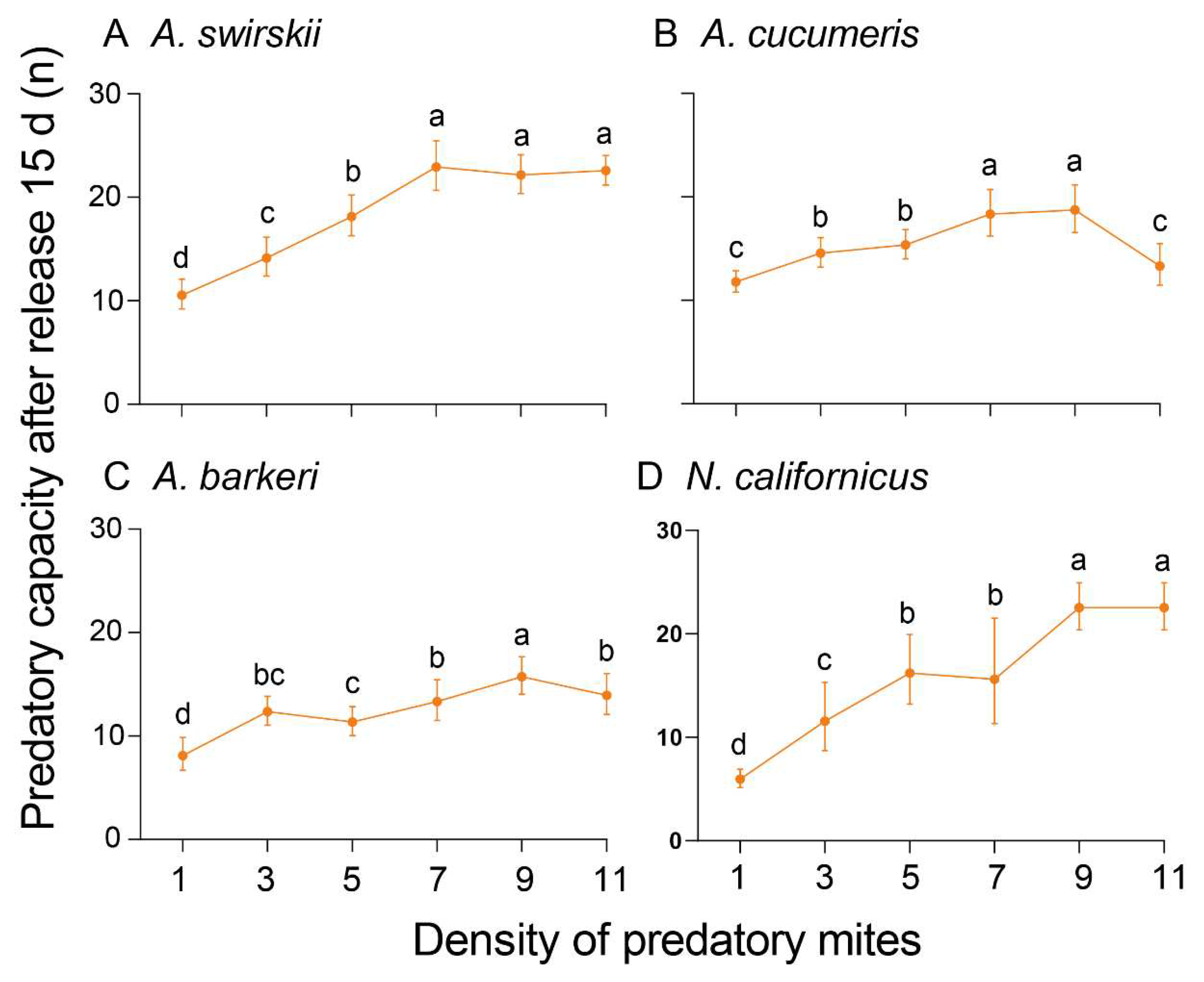
Figure 7.
Predatory capacity (preyed spider mites per predator) between release for 1 d and 15 d of four predatory mite species in Tibetan Plateau. Asterisks above points indicate statistically significant differences (Mann–Whitney U-test, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant).
Figure 7.
Predatory capacity (preyed spider mites per predator) between release for 1 d and 15 d of four predatory mite species in Tibetan Plateau. Asterisks above points indicate statistically significant differences (Mann–Whitney U-test, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant).
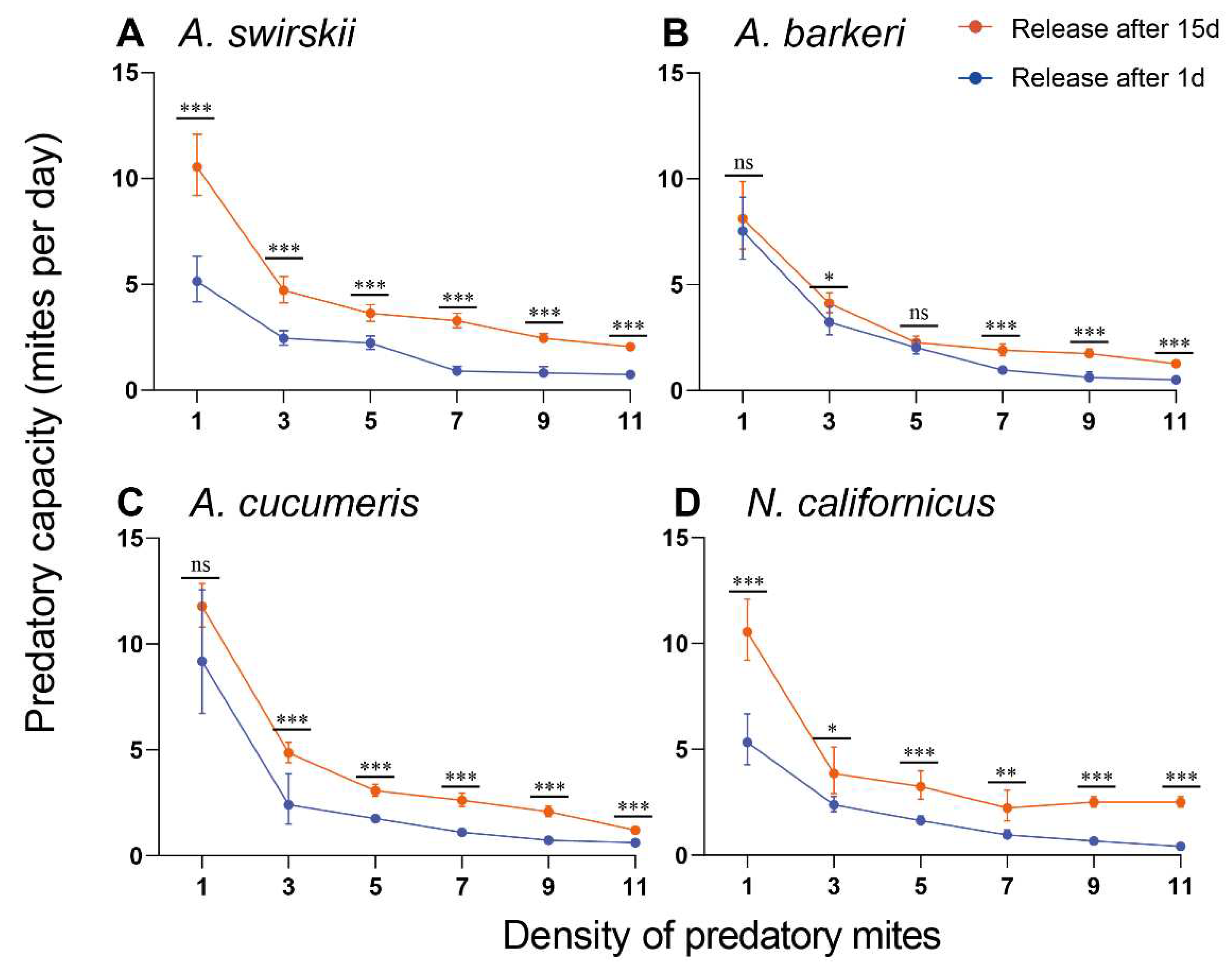
Figure 8.
Predatory capacity (A), handling time (B), attack coefficient (C) and max predatory amount (D) after release for 1 d of four predatory mite species in Tibetan Plateau. The max predatory amount data are analyzed by one-way ANOVA, SPSS 21.0, while the data of predatory capacity, handling time and attack coefficient are analyzed by Kruskal–Wallis tests, SPSS 21.0 (*P < 0.05, **P < 0.01, ns, not significant).
Figure 8.
Predatory capacity (A), handling time (B), attack coefficient (C) and max predatory amount (D) after release for 1 d of four predatory mite species in Tibetan Plateau. The max predatory amount data are analyzed by one-way ANOVA, SPSS 21.0, while the data of predatory capacity, handling time and attack coefficient are analyzed by Kruskal–Wallis tests, SPSS 21.0 (*P < 0.05, **P < 0.01, ns, not significant).
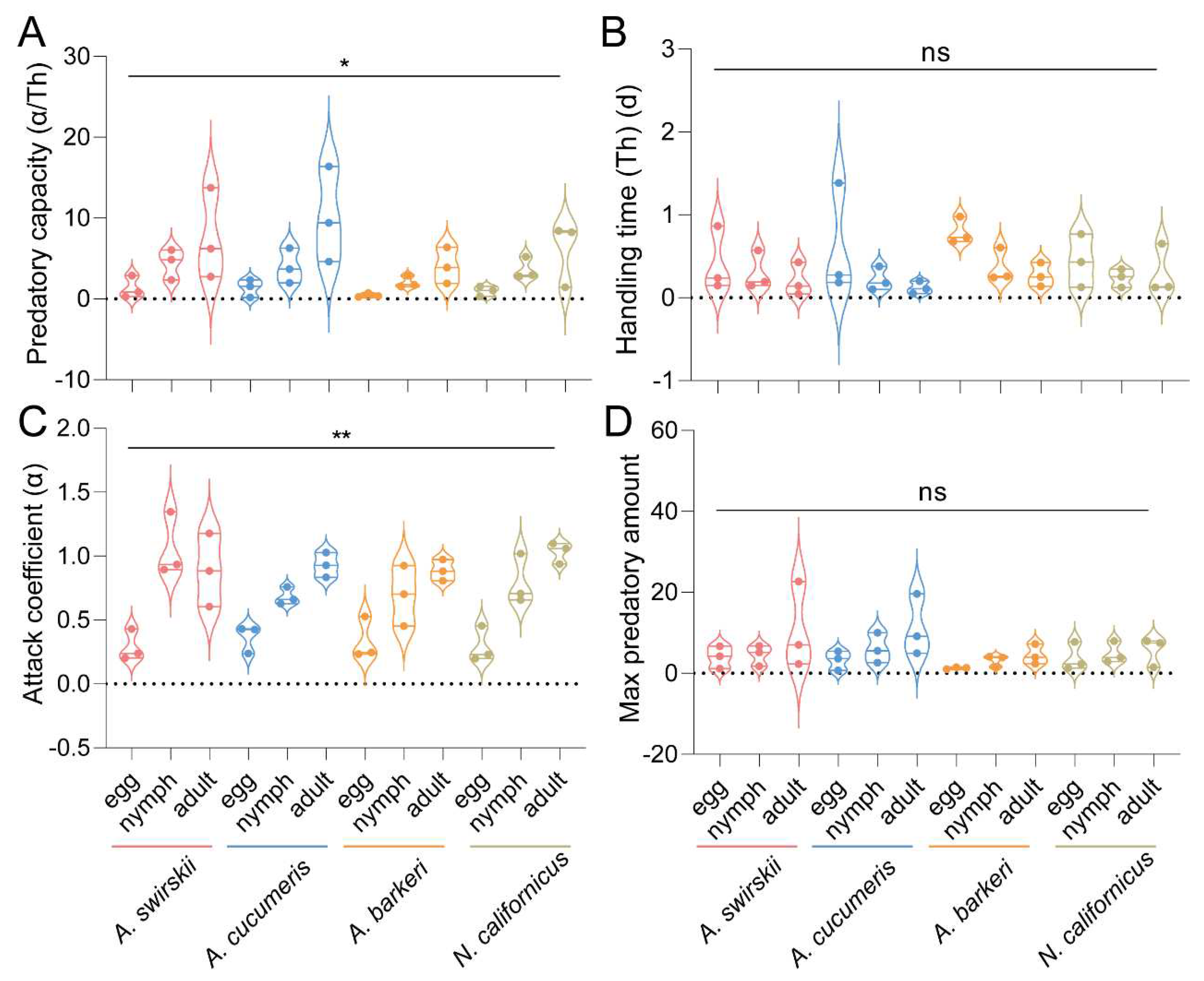
Figure 9.
Predatory capacity (A), handling time (B), attack coefficient (C) and max predatory amount (D) after release for 15 d of four predatory mite species in Tibetan Plateau. Results are shown as box and whiskers; the same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA, except for attack coefficient, which is analyzed by Kruskal–Wallis tests, SPSS 21.0 (*P < 0.05).
Figure 9.
Predatory capacity (A), handling time (B), attack coefficient (C) and max predatory amount (D) after release for 15 d of four predatory mite species in Tibetan Plateau. Results are shown as box and whiskers; the same letters above columns represent homogeneous groups in post-hoc tests (P > 0.05) following an ANOVA, except for attack coefficient, which is analyzed by Kruskal–Wallis tests, SPSS 21.0 (*P < 0.05).
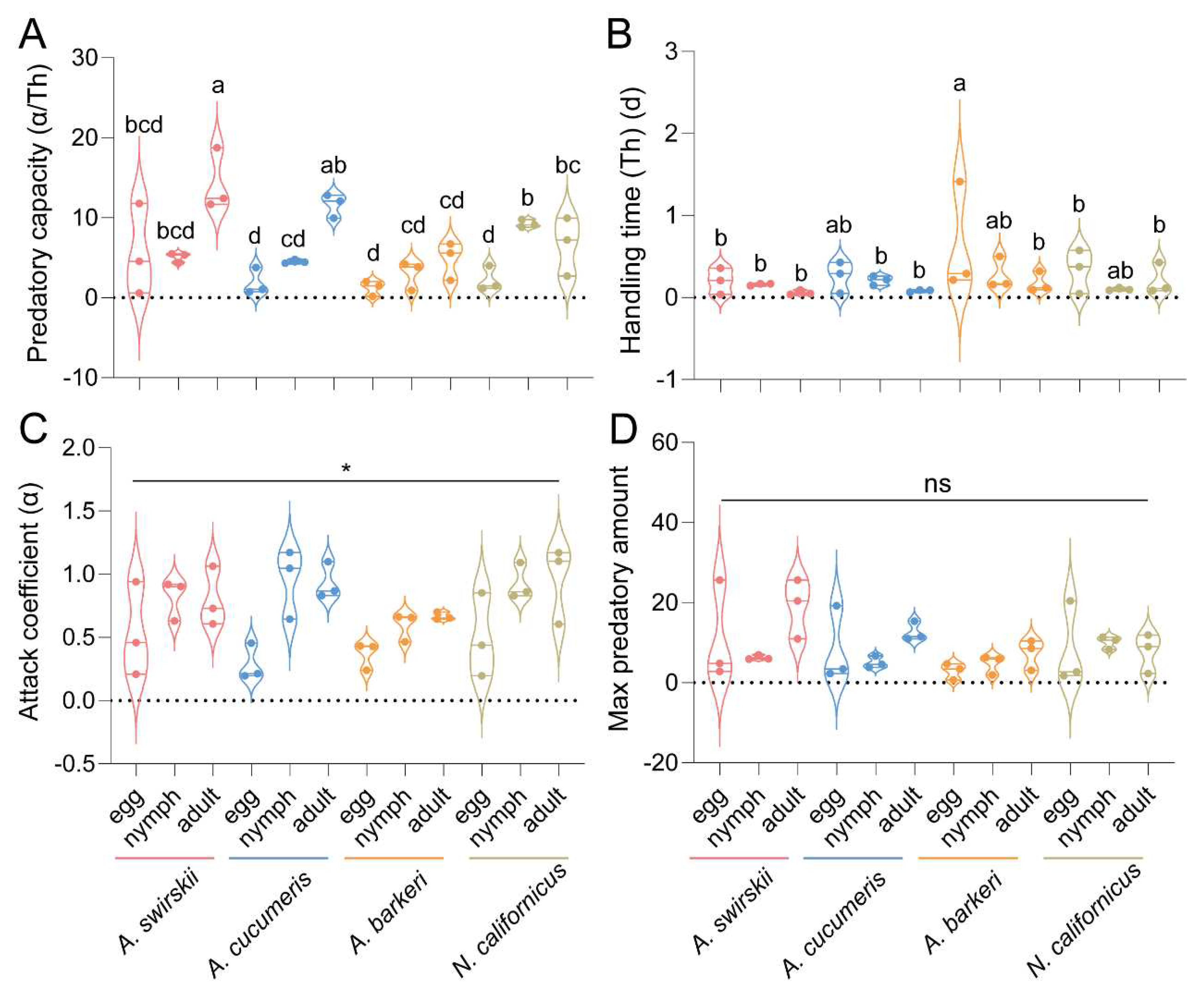

Table 1.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 1st repeat.
Table 1.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 1st repeat.
| Predatory mite species |
Tetranychus urticae (green) stage |
Functional response model (Na=) |
R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.238N0/(1+0.206 N0) | 0.992 | 28.547 |
| nymph | 1.346 N0/(1+0.751 N0) | 0.751 | 16.336 | |
| egg | 1.178 N0/(1+0.506 N0) | 0.767 | 17.043 | |
| Amblyseius cucumeris | adult | 0.238 N0/(1+0.329 N0) | 0.783 | 17.794 |
| nymph | 0.759 N0/(1+0.289 N0) | 0.925 | 24.793 | |
| egg | 0.928 N0/(1+0.186 N0) | 0.922 | 24.672 | |
| Amblyseius barkeri | adult | 0.246 N0/(1+0.241 N0) | 0.988 | 28.335 |
| nymph | 0.925 N0/(1+0.561 N0) | 0.873 | 22.091 | |
| egg | 0.808 N0/(1+0.343 N0) | 0.879 | 22.382 | |
| Neoseiulus californicus | adult | 0.246 N0/(1+0.241 N0) | 0.985 | 28.145 |
| nymph | 0.925 N0/(1+0.561 N0) | 0.965 | 27.032 | |
| egg | 0.808 N0/(1+0.343 N0) | 0.826 | 19.776 |
Table 2.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 2nd repeat.
Table 2.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 2nd repeat.
| Predatory mite species |
Tetranychus urticae (green) stage |
Functional response model (Na=) |
R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.204N/(1+0.048N) | 0.977 | 27.677 |
| nymph | 0.894N/(1+0.132N) | 0.893 | 23.111 | |
| egg | 0.605N/(1+0.027N) | 0.965 | 27.033 | |
| Amblyseius cucumeris | adult | 0.430N/(1+0.079N) | 0.963 | 26.894 |
| nymph | 0.629N/(1+0.063N) | 0.963 | 26.887 | |
| egg | 0.835N/(1+0.043N) | 0.986 | 28.169 | |
| Amblyseius barkeri | adult | 0.235N/(1+0.160N) | 0.978 | 27.745 |
| nymph | 0.453N/(1+0.117N) | 0.949 | 26.092 | |
| egg | 0.973N/(1+0.244N) | 0.875 | 22.189 | |
| Neoseiulus californicus | adult | 0.199N/(1+0.026N) | 0.973 | 27.444 |
| nymph | 1.020N/(1+0.352N) | 0.760 | 16.744 | |
| egg | 1.098N/(1+0.146N) | 0.912 | 24.111 |
Table 3.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 3rd repeat.
Table 3.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 1d), the 3rd repeat.
| Predatory mite species |
Tetranychus urticae (green) stage |
Functional response model (Na=) |
R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.431N/(1+0.064N) | 0.961 | 26.774 |
| nymph | 0.935N/(1+0.180N) | 0.941 | 25.690 | |
| egg | 0.885N/(1+0.126N) | 0.946 | 25.925 | |
| Amblyseius cucumeris | adult | 0.426N/(1+0.117N) | 0.916 | 24.335 |
| nymph | 0.662N/(1+0.118N) | 0.907 | 23.869 | |
| egg | 1.028N/(1+0.112N) | 0.920 | 24.533 | |
| Amblyseius barkeri | adult | 0.529N/(1+0.385N) | 0.886 | 22.573 |
| nymph | 0.703N/(1+0.172N) | 0.914 | 24.212 | |
| egg | 0.882N/(1+0.122N) | 0.940 | 25.616 | |
| Neoseiulus californicus | adult | 0.445N/(1+0.196N) | 0.919 | 24.501 |
| nymph | 0.657N/(1+0.083N) | 0.934 | 25.321 | |
| egg | 1.059N/(1+0.133N) | 0.917 | 24.369 |
Table 4.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 1st repeat.
Table 4.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 1st repeat.
| Predatory mite species | Tetranychus urticae (green) | Functional response equation | R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.209N/(1+0.075N) | 0.982 | 27.993 |
| nymph | 0.631N/(1+0.091N) | 0.939 | 25.573 | |
| egg | 1.064N/(1+0.097N) | 0.929 | 25.036 | |
| Amblyseius cucumeris | adult | 0.214N/(1+0.063N) | 0.995 | 28.684 |
| nymph | 0.645N/(1+0.096N) | 0.915 | 24.256 | |
| egg | 0.867N/(1+0.075N) | 0.979 | 27.810 | |
| Amblyseius barkeri | adult | 0.240N/(1+0.340N) | 0.784 | 17.827 |
| nymph | 0.466N/(1+0.234N) | 0.899 | 23.455 | |
| adult | 0.701N/(1+0.225N) | 0.914 | 24.222 | |
| Neoseiulus californicus | adult | 0.853N/(1+0.491N) | 0.844 | 20.699 |
| nymph | 1.093N/(1+0.131N) | 0.921 | 24.613 | |
| adult | 1.170N/(1+0.503N) | 0.820 | 19.477 |
Table 5.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 2nd repeat.
Table 5.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 2nd repeat.
| Predatory mite species | Tetranychus urticae (green) | Functional response equation | R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.459N/(1+0.018N) | 0.913 | 24.517 |
| nymph | 0.919N/(1+0.154N) | 0.934 | 25.315 | |
| egg | 0.731N/(1+0.029N) | 0.930 | 25.067 | |
| Amblyseius cucumeris | adult | 0.196N/(1+0.010N) | 0.968 | 27.169 |
| nymph | 1.171N/(1+0.308N) | 0.821 | 19.563 | |
| egg | 1.099N/(1+0.100N) | 0.977 | 27.654 | |
| Amblyseius barkeri | adult | 0.428N/(1+0.092N) | 0.956 | 26.504 |
| nymph | 0.663N/(1+0.105N) | 0.930 | 25.101 | |
| adult | 0.644N/(1+0.062N) | 0.939 | 25.545 | |
| Neoseiulus californicus | adult | 0.197N/(1+0.010N) | 0.963 | 26.912 |
| nymph | 0.861N/(1+0.076N) | 0.988 | 28.329 | |
| adult | 1.104N/(1+0.123N) | 0.898 | 23.367 |
Table 6.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 3rd repeat.
Table 6.
Parameter estimates of four predatory species’ functional response when feeding on different stages of T. urticae (green) by Holling Ⅱ functional response model (after release for 15d), the 3rd repeat.
| Predatory mite species | Tetranychus urticae (green) | Functional response equation | R2 | Chi-sqaure value |
|---|---|---|---|---|
| Amblyseius swirskii | adult | 0.940N/(1+0.195N) | 0.918 | 24.449 |
| nymph | 0.903N/(1+0.153N) | 0.917 | 24.385 | |
| egg | 0.608N/(1+0.030N) | 0.947 | 26.031 | |
| Amblyseius cucumeris | adult | 0.455N/(1+0.195N) | 0.910 | 24.001 |
| nymph | 1.046N/(1+0.228N) | 0.852 | 21.062 | |
| egg | 0.833N/(1+0.054N) | 0.950 | 26.173 | |
| Amblyseius barkeri | adult | 0.432N/(1+0.126N) | 0.927 | 24.914 |
| nymph | 0.657N/(1+0.112N) | 0.959 | 26.676 | |
| adult | 0.653N/(1+0.076N) | 0.907 | 23.858 | |
| Neoseiulus californicus | adult | 0.438N/(1+0.164N) | 0.913 | 24.186 |
| nymph | 0.830N/(1+0.078N) | 0.903 | 23.628 | |
| adult | 0.605N/(1+0.051N) | 0.864 | 21.650 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



