Submitted:
13 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
Background: In the domestic pig, neural and non-neuronal cholinergic systems in the gallbladder and extrahepatic bile ducts need a detailed morphological study to identify the type and localization of their structural components. We aimed to quantitatively analyze biliary cholinergic ganglia and nerves as well as non-neuronal cholinergic cells; Methods: Immunohistochemical detection of choline acetyltransferase expression was performed to define cholinergic nerve and non-neuronal structures in gallbladder and extrahepatic bile ducts; Results: The number and dimensions of cholinergic ganglia, perikaryons and nerves forming ganglionic nerve plexuses in the propria, muscular and subserosal layers was estimated. The immunoexpression of choline acetyltransferase in biliary glandular cells and mast cells was also detected and analyzed. Conclusions: Qualitative and quantitative analysis of choline acetyltransferase positive nerves, ganglionic and non-ganglionic nerve plexuses of acetylcholine supplemented the morphological knowledge of the cholinergic innervation of the extrahepatic biliary pathways and non-neuronal sources: glandular cholinergic cells and cholinergic-type mast cells. Original data of the number and dimensions of intramural nerves, mast cells, ganglia and ganglion perikaryons could be used as reference values in the diagnosis of neuropathological processes in biliary pathways. Clarifying the precise mechanisms of gallbladder smooth muscle contractility will contribute to the improvement of therapeutic strategies to prevent biliary stasis and biliary disorders.
Keywords:
mast cells
; ganglia
; choline acetyltransferase
; tryptase
; bile ducts
; pigs
1. Introduction
Acetylcholine (ACh) is well known neurotransmitter, although it is known to be expressed in non-neuronal cells in animals and man, which are elements of a non-neuronal cholinergic system, performing a number of functions other than neurotransmitter [1,2]. As a neurotransmitter, its action is mediated by nicotinic and muscarinic receptors. Nicotinic ACh receptors (nAChRs) are proteins ensuring chemical neurotransmission in neurons, ganglia, interneurons, and motor plate. Muscarinic ACh receptors (mAChRs) are proteins mediating chemical neurotransmission in neurons and effector organs: heart, smooth muscle cells, and glands.
It is important to note that some non-neuronal cells use the same receptors as cholinergic neurons do to perform their functions and maintain local homeostasis [3].
Evidence for the synthesis of acetylcholine is the establishment of positive anti-ChAT immunoreactivity revealing the presence of the enzyme choline acetyltransferase (ChAT), synthesizing acetylcholine, as well as the content of acetylcholine in most of cells belonging to non-neuronal cholinergic system, especially well studied in humans and laboratory animals [1,2,3,4]. These cells exert their regulatory role by releasing non-neuronal acetylcholine in auto- and paracrine way on different cell types. Such cells are epithelial cells in the respiratory tract, intestines, skin, vagina, placenta, cornea, urothelium, granulosa cells, endothelial and immune cells, adipose tissue. In these cells both precursors for the synthesis of acetylcholine - choline and acetyl-CoA - exist [3].
It is known that neuronal acetylcholine is released by exocytosis, but less data exist on the release mechanisms of non-neuronal acetylcholine. Suitable proteins for mediating non-neuronal acetylcholine release are thought to be the organic cation transporters (OCTs), which are expressed in every cell type [4]. For example, in the human placenta, OCT subtype 3 is involved in the release of nonneuronal acetylcholine (4), whereas in respiratory epithelium OCT subtypes 1 and 2 are present.
In gastrointesinal tract of many species such as guinea pig, dog and human was found that cholinergic and non-cholinergic neurons exist, however neurons in the gallbladder ganglia are cholinergic and immunoreactive for choline acetyltransferase (ChAT) [5,6]. Immunohistochemistry and retrograde tracing have expanded knowledge regarding the presence and distribution of several neuroactive peptides and synthetic enzymes in gall bladder and sphincter of Oddi (SO) neurons. Immunohistochemistry and retrograde tracing help expression of variety of neurotransmitters in intramural neurons, including acetylcholine, tachykinins, enkephalins, nitric oxide synthase (NO), vasoactive intestinal peptide (VIP), somatostatin (SOM), galanin, pituitary adenylate cyclase-activating polypeptide (PACAP), cocaine, and amphetamine-regulated transcript (CART) of gall bladder and sphincter of Oddi (SO) neurons. The distribution of these neuroactive substances in the extrahepatic biliary tract are site- and species-specific [7].
Neuronal plexuses found in human gallbladder are cholinergic [5] that mediate neuronal-induced gallbladder`s smooth muscle contraction by acetylcholine from vagal nerve terminals. Neurons in guinea pig gallbladder are divided into two subpopulations based on the type of transmitters. The larger population, representing over 80% of neurons, was immunoreactive for ChAT, OFQ, CART, SP, NPY, and SOM. The remaining neurons were immunoreactive for ChAT, OFQ, CART, VIP, PACAP, and NOS [6,8,9]. Chemical coding of nerve fibers reveals that in the paravascular plexus CGRP and SP co-localize with PACAP in extrinsic sensory fibers. Subpopulations of nerves in the perivascular plexus are represented by extrinsic sensory nerves, coexpressing PACAP/SP and by sympathetic postganglionic nerves, co-expressing OFQ, TH/D H and NPY [6,10].
In contrast to the gallbladder, in the sphincter of Oddi (SO) of the guinea pig, excitatory and inhibitory neurons constitute two distinct subpopulations, with NOS and ChAT not colocalized [11,12]. Excitatory neurons predominate and coexpress ChAT, tachykinin, and enkephalin (ENK)-endorphin. Many excitatory motor neurons in the SO showed immunoreactivity for OFQ, and some of them were positive for calretinin, SP, ENK, and NOS, but not calbin.
According to the response of SO to cholecystokinin stimulation, animal species can be categorized as: those [13] with SO reacting to cholecystokinin stimulation with increased motility and those with SO reacting with relaxation [13]. Human SO, as well as pig, cat, and dog SO, refer to second category [14]. Pigs, which are less expensive for research works than cats or dogs, appear to be a more suitable species for experimental study of neuronal and humoral control of SO.
The bile production in biliary tract is regulated by the autonomic nervous system [15]. Cholinergic ganglion cells and nerve fibers located in the papilla of Vater and gallbladder are important in coordinating gallbladder empting and bile flow through extrahepatic biliary ducts. The normal function and biliary disorders of gallbladder and extrahepatic bile duct depend on the innervation. Puzikov and Lychkova (16) established that intramural adrenergic neurons synaptically connected to preganglionic parasympathetic fibers are involved in the mechanism of vagal inhibition of sphincter of Oddi motility.
Therefore, when determining the therapy, the regulatory effect of autonomic innervation on bile passage deserves significant attention. Bile acids and autonomic innervation interact to control the response of cholangiocytes to liver injury [18]. LeSage et al. [19] found that cholinergic nerves regulate bile secretion in the biliary tract.In this regard, the present study was undertaken to quantitatively characterize biliary cholinergic neural structures and to provide data on the normal range in ganglion number, number of neurons per ganglion of ganglionic nerve plexuses in gallbladder (GB), extrahepatic bile ducts (EHBD) and major duodenal papilla (PDM) in healthy pigs. These data would serve for a more accurate assessment of biliary neuropathology.
There is a diverse function of non-neuronal acetylcholine. The stimulatory effects of acetylcholine on respiratory epithelial cilia movement, mucus secretion, are mediated mainly through mAChRs [20]. Acetylcholine inhibits the apical sodium transport and stimulates the basolateral potassium transport, thereby controlling ion and water transport of lining epithelium and secretory cells of glands. Hirota and McKay [21] reported such effects that are mediated by acetylcholine in gut. Twenty years ago, data emerged regarding the role of the non-neuronal cholinergic system in immune cells, such as T and B cells, dendritic cells, granulocytes, macrophages [22]. Activation of the T-cell receptor enhances acetylcholine synthesis. Acetylcholine can modify immune responses by modulating immune cells activity through auto- and paracrine mechanisms [23]. Therefore, vagus nerve transmits information about the state of the immune system to the brain [24].
Data on ChAT positive mast cells are scarce. There are data on AChE immunoreactivity in human and rodent mast cells. Hagforsen et al. [25] reported acetyl cholinesterase (AChE)+ mast cells in normal skin and in inflammatory skin processes, while Nechushtan et al. [26] observed the AChE expression in mast cells in human colon. Interactions between Ach, histamine and mast cells in inflammatory responses are known. Mast cells are known to express cholinergic receptors. Nicotinic receptors were detected in the HMC-1 mast cell line [27] but not in lung mast cells. Muscarinic receptors were identified in rodent mast cells [28]. According to Jones et al. [29], M1 receptor regulates degranulation of rat mast cells and mast cells in human bronchi [30].
Mast cell involvement in various cholangiopathies was proven in several studies (31-33). Bile is modified by cholangiocytes lining inside the bile ducts which are target cells in cholangiopathies such as PSC and PBC [34,35].
Mast cell participation in gallstone disease in humans is well known. Rau et al. (36) provide important data on the increased number of tryptase-positive mast cells in the mucosa and muscular layer of the gallbladder involved in the generation of symptoms in children with gallstones and biliary dyskinesia. The authors hypothesize an interaction between mast cells and pain in this pathology, which could be explained by the following mechanisms: stretch sensitivity associated with mast cell degranulation in the gastrointestinal tract and through the occurrence of gallbladder spasms stimulated by mast cell biologically active substances. In this regard, some authors demonstrated that histamine and cysteinyl leukotrienes stimulate the excitability and contractility of smooth muscle cells in gallbladder [37]. Similar to mast cells in gallbladder disease, their number increases in the ileum and colon of adults with irritable bowel.
To date, information about studies on mast cells relevant to cholelithiasis in pigs, and studies on the normal localization and numbers of ChAT immunoreactive mast cells in GB and EHBD is not available. We believe that the pig, as a suitable experimental model, could be successfully used in future research in this direction, especially taking into account the evidence of human gall bladder diseases that also occur in pigs, such as cholelithiasis, chronic cholecystitis, mucosal hyperplasia, adenocarcinoma, adenoma, biliary and peribiliary cysts.
Based on the lack of detailed morphological study on the cholinergic neural and non-neuronal cholinergic system in the porcine gallbladder, ductus cysticus, ductus hepaticus communis, ductus choledochus and papilla duodeni major we aimed to quantitatively analyze biliary cholinergic ganglia and nerves as well as non-neuronal cholinergic cells: glandular cells and mast cells in the mentioned organs.
2. Materials and Methods
2.1. Animals
In the present study, tissue samples from the extrahepatic bile ducts and gallbladders of male clinically healthy pigs (Bulgarian White×Landrace cross) aged 6 months (92–100 kg) were used. All procedures were approved by the Bulgarian Food Safety Agency with license № 174. Tissue samples from the common hepatic duct (near the terminal part of ductus cysticus), from the ductus cysticus near collum vesicae biliaris, from beginning of the ductus choledochus (DCHO) near the junction with the ductus cysticus, from the intramural part of ductus choledochus (DCHI), as well as from the bottom (fundus vesicae biliaris), body (corpus vesicae biliaris) and neck (collum vesicae biliaris) of the gallbladder were collected after slaughtering the animals in slaughterhouse, then put in 10% aqueous formalin.
2.2. Histological methods
Tissue paraffin sections with thickness of 5 µm were mounted on glass slides, then deparaffinized and rehydrated. A portion of the serial histological cross-sections were stained with hematoxylin and eosin to rule out presence of pathological findings. Another part of the sections was processed immunohistochemically to determine the expression of choline acetyltransferase and tryptase.
2.3. An immunohistochemical method for visualization of choline acetyltransferase and tryptase reactive structures
Serial sections were put in 0.1 M PBS, then in 1.2% hydrogen peroxide in methanol for 30 min. Antigen retrieval was performed in buffer (pH 9.0) for 20 min. Between steps, sections were washed with EnVisionFlex Wash Buffer, then incubated in a humid chamber overnight at 4 °C with primary antibodies: a polyclonal rabbit choline acetyltransferase (ChAT) (PA 1-4738), Thermo Fischer and a monoclonal mouse antihuman mast cell tryptase (MBS9510697), which is ready to use. After washing with PBS, the sections were incubated with an EnVi-sion detection system (DAKO) for 24 h at 4 °C. Antibody-antigen reaction was visualized with diaminobenzidine. Serial sections were sequentially stained with choline acetyltransferase and tryptase.
PBS instead primary antibody was used as a negative control.
2.4. Statistical analysis
The number of cholinergic nerves was counted on a field of view X400, and the number of ganglia was determined on cross-sections of the examined organs using a light microscope (LEICA DM1000) equipped with a digital camera (LEICA DFC 290) and quantitative analysis software ( LAS V4.10.0 2016). The number of perikaryons per a ganglion was also determined. Endocrine cell density was determined by counting the number of endocrine cells per microscopic field X100 as well as per gland cross-section. Metric data were processed by GraphPad Prism 6 for Windows (GraphPad Software, Inc., USA) by one-way ANOVA followed by Tukey-Kramer post-hoc test. P-values less than 0.05 were considered statistically significant. Data are presented as mean ± SD (Standard deviation).
Terminology is consistent with Nomina Histologica Veterinaria [38].
3. Results
3.1. Choline acetyltransferase positive nerve structures
Cholinergic nerves and ganglia in the GB and EHBD were identified by immunohistochemical expression of the enzyme choline acetyltransferase (ChAT). Choline acetyltransferase positive (ChAT+) structures in GB and EHBD were represented by nerves, perikaryons and outgrowths of ganglion neurons, mast cells, as well as immunopositive glandular cells were found in EHBD (Figure 1).
ChAT+ perikaryons and outgrowths of ganglion neurons with moderate enzyme reactivity were found mainly in the muscle and subserosal layers, less in the propria of the GB and EHBD, but in PDM – one- and two-neuron ganglia were located in the subglandular connective tissue layer (SGCT) of the propria adjacent to the sphincter of Oddi (SO) (Figure 1, Table 1, Table 2, Table 3 and Table 4).
Immunohistochemical examination showed that ChAT immunoreactive intramural nerves and ganglia formed ganglionic nerve plexuses (Figure 1). The most ganglia of muscle layer were found in the gallbladder`s neck and body - 67% each, less in the bottom - 63%. These ganglia contained up to 8 perikaryons in the bottom, up to 10 in the body, and up to 12 in the neck of the gallbladder. Single ganglion cells were found in lamina propria of the bladder. Their number was the most in the propria of the body - 29%, followed by that in the neck - 22%, and the smallest number (12%) - in the bottom. The submucosal plexus is formed mainly by ganglion cells in the propria and by the ganglion outgrowths projected from muscular layer. The nerve fibers making up the submucosal plexus reached the basement membrane of the epithelium, but did not penetrate between the epithelial cells. In the fundus wall of the porcine GB, it was found that 25% of the ganglia were located in the subserosal layer near the muscle layer, 63% in the muscle layer, and 12% in the propria near the muscle layer. In the bladder`s body, 4% of the ganglia were found in the subserosal layer near the muscle layer, 67% in the muscle layer, 29% in the propria near the muscle layer.
The largest ganglia were located in collum vesicae biliaris, followed by the corpus and fundus vesicae biliaris (Table 1). In the wall of the ductus cysticus, the size of the ganglia was similar to the size of those located at fundus vesicae biliaris. The size of ganlia decreased in direction from GB to DHC, followed by intramural part of DCH, DC, extramural part of DCHT and the smallest ganglia were detected in PDM (Table 1).
The number of ganglia per cross section decreased from GB (more ganglia in body compared to neck and bottom) to PDM (Table 1). GB was followed by DC, DHC, DCHO, DCHT, PDM and DCHI (Table 1).
The diameter of the perikaryons in the ganglia decreased in direction from GB to PDM (Table 1). The largest perikaryons were identified in the ganglia of GB (with the largest diameter in the body, followed by the neck and bottom), followed by those in DC, DHC, DCHO, DCHT, DCHI and PDM.
The number of perikaryons per a ganglion was greatest in the GB (most perikaryons in the neck, followed by the body and bottom), followed by DHC, DCH, DC and PDM (Table 1).
Immunohistochemical expression of ChAT allowed for the first time to establish the localization of ChAT+ nerves in the muscular and subserosal ganglion nerve plexus, in the propria between the glands and near the superficial biliary epithelium as well as in the muscular layer and subserosa of porcine GB and EHBD (Figure 1).
The number and diameter of ChAT+ nerves were highest in the serosa and adventitia of the GB and EHBD, followed by the muscle layer and lowest in the propria (Table 2, Table 3). In the subglandular layer of DCHI and PDM, ChAT+ nerves were fewer but larger in diameter compared to nerves in the subepithelial layer.
3.2. Choline acetyltransferase positive glandular cells (ChAT+Cs)
Choline acetyltransferase positive cells (ChAT+Cs) with moderate to strong reactivity were found in the EHBD glands, but were absent in the GB. Their density was greatest in the glands of DCHI, followed by PDM, DHC, DCHO and DC (Table 4) (Figure 1).
The highest density of ChAT+Cs in the superficial glandular alveoli was observed in DCHO, followed by that in DHC, PDM, DCHI and DC. More ChAT+Cs were found in superficial glandular alveoli compared to deep glandular alveoli in DCHI and PDM, whereas deep glands in DHC, DCHO and DC were immunonegative (Table 4).
3.3. Immunohistochemical localization of ChAT+ mast cells in GB, EHBD and PDM
To establish the presence of choline acetyltransferase positive mast cells (MCsChAT+), immunohistochemical identification of tryptase positive mast cells was performed on serial sections from the wall of GB, DC, DHC, extra- and intramural part of DCH, as well as from the wall of PDM. Both types of mast cells: MCsChAT+ and MCstr+, showed similar morphology (immunopositive granules in the cytoplasm) and similar localization in the propria, muscle and subserosal layers of the examined organs (Figure1).
MCsChAT+ and MCstr+ were found adjacent to capillaries, arterioles, and venules in the propria and basement membrane of the biliary epithelium of the mucosa of the GB, EHBD and majorduodenal papilla (Figure 1).
In the muscle layer of these organs, MCsChAT+ and MCstr+ were observed near blood vessels, superficial and deep glands, and between bundles of smooth muscle cells, but some mast cells were found adjacent to smooth muscle cells.
In adventitia or serosa of the organs, MCsChAT+ and MCstr+ were localized most often in the adventitia of the muscular-type arteries and adjacent to the nerves of the GB, DC, DHC and DCH.
4. Discussion
Regarding gallbladder innervation, it is known that there is both internal and external innervation, which primarily participates in regulation of contractility of the smooth muscles and secretory activity of the organ. The internal innervation of the gallbladder is similar to the enteric nervous system (ENS) in the digestive tract and consists of neurons located in lamina propria, muscle layer and subserosa that form ganglion plexuses [13]. The number and precise localization of ganglion plexuses in the gallbladder wall depends on the animal species [7,39]. Our findings support the statement of Meedeniya et al. [39] that porcine gallbladder ganglion plexus localisation resembles those of large animals and the statement of Mawe [7] that this type of plexus differs from those in rodents. The current study provided original information on the number and dimensions of intramural ChAT+ nerves, ganglia, and perikaryons of ganglion neurons, showing that ganglia in porcine GB, EHBD and PDM are cholinergic. In addition, a similar localization of intramural nerves and ganglia was found in the porcine GB and EHBD to that in man, studied by Alexander [40]. The intramural nerves of the gallbladder, common hepatic and common bile ducts formed nerve plexuses in subserosa, muscular and submucosal layer, containing different numbers and dimensions of ganglia. In the human GB, subserosal ganglia are more numerous in the neck than in the body or fundus of the organ. In the pig, we found that the ChAT+ serosal ganglionated plexuses were the most in the bottom - 25% of the total number of ganglia in the three layers of the bladder wall, followed by the neck - 11% and least in the body - 4%. The muscular nerve plexus of the human GB contains numerous small ganglia, consisting of one to five or six cells, throughout the corpus and fundus of the gallbladder. In the pig, the most musclular layer ganglia were in the neck and body - 67% each, less in the bottom - 63%. Single ganglion cells were found in the lamina propria of the bladder in both man and pig. Similar to the human, the submucosal plexus was found to arise mainly from these cells and from the outgrowths of the cells in the muscular layer of GB. The fibers making up the submucosal plexus reached the epithelium but did not penetrate between the epithelial cells.
The present study identified ChAT immunoreactive ganglion perikaryons. The largest ganglia were located in the neck of the porcine GB, followed by its body and bottom. In the wall of the ductus cysticus, the size of the ganglia was similar to the size of those located at the bottom of the bladder.
Immunohistochemical expression of ChAT allowed for the first time to establish the localization of ChAT+ nerves in the muscular and subserosal ganglion nerve plexus, in the interglandular propria and near the superficial biliary epithelium, in the muscular layer and subserosa of porcine GB and EHBD, which are most likely efferent nerve fibers because ChAT is a well-known marker of motor efferent nerve fibers [41]. Therefore, ChAT negative nerve fibers are afferent with a sensory role similar to those in the esophagus and intestine. A number of authors have found that primary afferent fibers serve as receptors for muscle stretch, mechanical and chemical mucosal stimuli, as well as mucosal distension [41].
Baumgarten & Lange [43] and Cai & Gabella, [42] revealed the absence of intramuscular cholinergic and adrenergic nerve fibers in the GB of guinea pig and cat, which were located mainly in the lower part of the common bile duct. In contrast to the ementioned authors, the present study found a well-defined network of intramuscular cholinergic nerves in the wall of the GB and EHBD in the domestic pig. Even the density of intramuscular cholinergic nerves in the GB musclular layer was greater than that in DHC and DCH but less than in DC.
According to Mawe [44], the ganglionic plexus of the gallbladder contains preganglionic vagus fibers that terminate in gallbladder neurons as well as sympathetic postganglionic fibers. Hormone cholecystokinin (CCK) acts in the gallbladder ganglia to activate gallbladder emptying after a meal. This communication reveals the existence of hormonal modulation of ganglion neuron function in regulation of gallbladder motility. Cholinergic nerves in lamina propria and muscular layer of the GB and EHBD in pigs were observed near the epithelial cells, which according to Gonda et al. [45], indicates the role of nerves in the regulation of secretory activity and muscle contractility of the gallbladder.
The mechanisms of neural regulation of the biliary epithelium in the intrahepatic bile ducts can be explained by results of Zanchi et al., [46]. According to these authors these mechanisms include direct nerve-cholangiocytic interaction, deposition of neurotransmitter in the peribular, extracellular environment (as described in alimentary canal) [47], and interaction of nerves with hepatic stellate cells (HSC) [48].
The present micromorphometric study on the neural structures of the porcine GB and EHBD provided original data on the number and dimensions of intramural nerves, ganglia and ganglion perikaryons that could be used as reference values in the diagnosis of neuropathological processes [15] in these organs, where the amount of neural structures is known to be affected.
The expression of acetylcholine in non-neuronal cells in animals and human are regarded as elements of a non-neuronal cholinergic system, performing number functions other than neurotransmitter [1,2].
The immunohistochemical reactions carried out in this study to establish the expression of ChAT allowed for the first time the determination of non-neuronal cholinergic system elements in the GB and EHBD of the domestic pig, represented by neuroendocrine cells and ChAT+ mast cells.
ChAT+ endocrine cells were found to be of the closed type in porcine EHBD in contrast to intestinal ChAT+ endocrine cells [49]. These data are similar to the data for ChAT immunoreactivity in rat and guinea pig duodenal, jejunum, and ileal glands, but lack data for ChAT expression in gastric or colonic mucosa. We also did not detect ChAT+ neuroendocrine cells in the mucosa of the porcine GB.
Given ChAT+ endocrine cell distribution in the intramural biliary glands of the EHBD, we can hypothesize that, similar to the intestine, cholinergic endocrine cells may initiate both secretory and muscle activity in organs examined [50,51].
It is important to clarify the precise mechanisms of gallbladder smooth muscle contractility to prevent biliary stasis and disorders.
The current study for the first time provided data on the number and size of choline acetyltransferase immunoreactive mast cells (ChAT+M) in porcine GB and EHBD. Acetylcholine appears to mediate interaction between mast cells and acetylcholine neurons, which are well represented in the intestine of mice and humans [26] similarly to porcine GB and EHBD as demonstrated in the present study. Neural regulation of mast cell degranulation is species specific. In humans, acetylcholine - inhibits [30], whereas in rats and rabbits, acetylcholine stimulates mast cell degranulation [28]. The sensitization status of the host may also affect the mast cell response to acetylcholine due to acetylcholine-induced histamine release [28]. On the other hand, antigenic stimulation that leads to mast cell degranulation causes an increase in ACh release from enteric neurons [26]. Interactions between mast cells and neurons may potentially contribute to the pathogenesis of many diseases in gastrointestinal tract [26].
Pathophysiological effects of cholinergic stimulation on mast cells are largely due to the mast cell inflammatory mediator histamine, involved in vasodilation, increased vascular permeability, gastric secretion, and contraction of gastrointestinal smooth muscle cells. Mast cell histamine exerts its effect through the histamine receptors on smooth muscles of the gallbladder. For example, activation of H1 receptors induces contraction by depolarizing smooth muscle cells, and H2 receptors induce relaxation by hyperpolarizing smooth muscle cells with the involvement of KATP channels [37]. It can be assumed that increased degranulation of mast cells are related to acute cholecystitis, through the release of endogenous histamine, which exerts an excitatory effect on the smooth muscle layer of the gallbladder to induce its contractility as a protective effect.
One of the first studies regarding MCs and PBC was done by Nakamura et al. [53], in which the number of mast cells was found to be increased around the portal tract in patients suffering from PBC. The authors concluded that increased nerve stimulation induces mast cell migration and activation leading to fibrosis.
The expression of choline acetyltransferase found by us for the first time in the mast cells of porcine GB, and EHBD and PDM suggests, on the one hand, the possibility of its synthesis and release from these cells, and on the other hand, its involvement in modulating the interaction between mast cell and intramural neurons in the studied organs. Similar to the gut, in the GB, and EHBD and PDM, interactions between mast cells and neurons could also contribute to the pathogenesis of many diseases. Data of the present study are a solid basis for future studies aimed at elucidating the role of acetylcholine produced by mast cells in the physiological and pathological processes occurring in the gallbladder, extrahepatic bile ducts, and the large major duodenal papilla.
5. Conclusions
This immunohistochemical study allowed for qualitative and quantitative analysis of ChAT+ nerves, ganglionic and non-ganglionic nerve plexuses, contributing to the morphological knowledge of the cholinergic innervation of the GB, EHBD and PDM. At the same time, data were provided on the non-neuronal sources of acetylcholine: glandular cholinergic cells and cholinergic-type mast cells. The original data on the number and dimensions of intramural nerves, ganglia and ganglion perikaryons could be used as reference values in the diagnosis of neuropathological processes in GB and EHBD, which are known to affect the amount of neural structures. Clarifying the precise mechanisms of gallbladder smooth muscle contractility will contribute to the improvement of therapeutic strategies to prevent biliary stasis and biliary disorders.
Author Contributions
I am the only author.
Funding
This work was supported by Bulgarian Ministry of Education and Science (MES) in the frames of Bulgarian National Recovery and Resilience Plan, Component "Innovative Bulgaria", the Project No. BG-RRP-2.004-0006-C02 “Development of research and innovation at Trakia University in service of health and sustainable well-being”.
Institutional Review Board Statement
All procedures were in accordance with Ordinance 20 of 01.11.2012 on the minimum requirements for the protection and humane treatment of experimental animals and the requirements for facilities for use, breeding and/or supply, Directive 2010/63/EU of the European Union Parliament and the Council - September 22, 2010 for the protection of animals used for scientific purposes and approved by the Ethics Committee of the Bulgarian Food Safety Agency with license № 174.
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
I would like to acknowledge the technical support of associate professor Dimitrinka Atanasova from Department of Anatomy, Medical Faculty, Trakia University, Stara Zagora, Bulgaria.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kurzen, H.; Wessler, I.; Kirkpatrick, C.J.; Kawashima, K.; Grando, S.A. The non-neuronal cholinergic system of human skin. Horm Metab Res 2007, 39, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Lips, K.S.; Luhrmann, A.; Tscherni, T.; Stoeger, T.; Alessandrini, F.; Grau, V.; et al. Down-regulation of the non-neuronal acetylcholine synthesis and release machinery in acute allergic airway inflammation of rat and mouse. Life Sci 2007, 80, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: the non-neuronal cholinergic system in humans. Br J Pharmacol 2008, 154, 1558–1571. [Google Scholar] [CrossRef]
- Wessler, I.; Panter, H.; Bittinger, F.; Kriegsmann, J.; Kirkpatrick, C.J.; Kawashima, K.; et al. Subcellular location of choline acetyltransferase (ChAT) and acetylcholine (ACh) in human placenta. Naunyn Schmiedebergs Arch Pharmacol (Suppl) 2001, 363, R23. [Google Scholar]
- Talmage, E.K.; Pouliot, W.A.; Schemann, M.; Mawe, G.M. Structure and chemical coding of human, canine and opossum gallbladder ganglia. Cell Tissue Res 1996, 284, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.M.; Ellis, L.M. Chemical coding of intrinsic and extrinsic nerves in the guinea pig gallbladder: distributions of PACAP and orphanin FQ. Anat Rec 2001, 262, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.M. Neurobiology of the gallbladder and sphincter of Oddi. In Neurogastroenterology: from the basics to the clinics.H.J. Krammer. M.V. Singer, (eds). New York: Kluwer Academic Publishers and Falk Foundation. 2000; pp. 288–302.
- Talmage, E.K.; Pouliot, W.A.; Cornbrooks, E.B.; Mawe, G.M. Transmitter diversity in ganglion cells of the guinea pig gallbladder: an immunohistochemical study. J Comp Neurol. 1992, 317, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Talmage, E.K.; Mawe, G.M. NADPH-diaphorase and VIP are colocalized in neurons of gallbladder ganglia. J Auton Nerv Syst 1993, 4, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.; Gershon, MD. Structure, afferent innervation, and transmitter content of ganglia of the guinea pig gallbladder: Relationship to the enteric nervous system. J Comp Neurol 1989, 283, 374–390. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.G.; Talmage, E.K.; Mawe, G.M. Immunohistochemical identification of neurons in ganglia of the guinea pig sphincter of Oddi. J Comp Neurol. 1995, 352, 106–116. [Google Scholar] [CrossRef]
- O’Donnell AM, Ellis LM, Riedl MS, Elde RP, Mawe, GM. Distribution and chemical coding of orphanin FQ/nociceptin-immunoreactive neurons in the myenteric plexus of guinea pig intestines and sphincter of Oddi. J Comp Neurol 2001, 430, 1–11. [CrossRef]
- Dahlstrand, C.; Theodorsson, E.; Dahlstfm .;, Ahlman, H. VIP antisera inhibit the relaxatory motor responses of the feline sphincter of Oddi and gall-bladder induced by VIP or vagal nerve stimulation. Acta Physiol Scand 1989, 137, 375–378. [CrossRef] [PubMed]
- Boyden, E.A. The sphincter of Oddi in man and certain representative mammals. Surgery 1937, 1, 25–37. [Google Scholar] [CrossRef]
- Ritter, U. Autonomous nervous system and bile ducts. Leber Magen Darm 1979, 9, 81–4. [Google Scholar] [PubMed]
- Puzikov, A.M; Lychkova, A.E. Nervous regulation of biliary tract motility. Eksp Klin Gastroenterol 2016, (7), 62–5. [Google Scholar]
- Mizuno, K.; Ueno, Y. Autonomic nervous system and the liver. Hepatol Res. 2017, 47, 160–16. [Google Scholar] [CrossRef] [PubMed]
- Fava, G.; Marzioni, M.; Francis, H.; Glaser, S.; Demorrrow, S.; Ueno, Y.; et al. Novel interaction of bile acid and neural signaling in the regulation of cholangiocyte function. Hepatol Res 2007, 37 (Supp. 1), S420–429. [Google Scholar] [CrossRef] [PubMed]
- LeSage, G.; Glaser S, Alpini G. Regulatory mechanisms of ductal bile secretion. Dig Liver Dis 2000, 32, 563–566. [CrossRef] [PubMed]
- Acevedo, M. Effect of acetylcholine on ion transport in sheep tracheal epithelium. Pflugers Arch. 1994, 427, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Hirota, C.L.; McKay, D.M. Cholinergic regulation of epithelial ion transport in the mammalian intestine. Br J Pharmacol. 2006, 149, 463–479. [Google Scholar] [CrossRef]
- Kawashima, K.; Yoshikawa, K.; Fujii, Y.X.; Moriwaki, Y.; Misawa, H. Expression and function of genes encoding cholinergic components in murine immune cells. Life Sci 2007, 80, 2314–2319. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.J.; Ja¨nig, W.; Levine, J.D. Vagal branches involved in inhibition of bradykinin-induced synovial plasma extravasation by intrathecal nicotine and noxious stimulation in the rat. J Physiol 1997, 498, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Hagforsen, E.; Einarsson, A.; Aronsson, F.; Nordlind, K.; Michaelsson, G. The distribution of choline acetyltransferase- and acetylcholinester-ase-like immunoreactivity in the palmar skin of patients with palmoplantar pustulosis. Br J Dermatol 2000, 142, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Nechushtan H, Soreq H, Kuperstein V, Tshori S, Razin E. Murine and human mast cell express acetylcholinesterase. FEBS Lett 1996, 379, 1–6. [CrossRef] [PubMed]
- Sudheer, P.S.; Hall, J.E;, Donev, R., Rea; G., Rowbottom, A.; Williams, P.E. Nicotinic acetylcholine receptors on basophils and mast cells. Anaesthesia 2006, 61, 1170–1174. [CrossRef] [PubMed]
- Masini, E.; Fantozzi, R.; Blandina, P.; Brunelleschi, S.; Mannaioni, P.F. Muscarinic cholinergic receptor binding in rat mast cells. Agents Actions 1983, 13, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.V.P.; Choi, O.H.; Beaven, M.A. Carbachol induces secretion in a mast cell line (RBL-2H3) transfected with the ml muscarinic receptor gene. FEBS Lett. 1991, 289, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Reinheimer, T.; Mohlig, T.; Zimmermann, S.; Hohle, K.D.; Wessler, I. Muscarinic control of histamine release from airways. Inhibitory M1-receptors in human bronchi but absence in rat trachea. Am J Respir Crit Care Med 2000, 162, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.V.; Vlaykova, T.I. Mast cells in human bile duct obstruction. J Mol Histol 2004, 35, 791–801. [Google Scholar] [CrossRef]
- Lindor, K.D.; Gershwin, M.E.; Poupon, R.; Kaplan, M.; Bergasa, N.V.; Heathcote, E.J. Primary biliary cirrhosis. Hepatology 2009, 50, 291–308. [Google Scholar] [CrossRef]
- Grizzi, F.; Caro, G.D.; Laghi, L.; Hermonat, P; Mazzola, P.; Nguyen, D.D. et al. Mast cells and the liver aging process. Immun Ageing 2013, 10, 9. [CrossRef]
- Glaser, S.S.; Gaudio, E.; Miller, T.; Alvaro, D.; Alpini, G. Cholangiocyte proliferation and liver fibrosis. Expert Rev Mol Med 2009, 11, e7. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–62. [Google Scholar] [CrossRef]
- Rau, B.; Friesen, C.A.; Daniel, J.F.; Qadeer, A.; You-Li, D.; Robert, C.C.; et al. Gallbladder wall inflammatory cells in pediatric patients with biliary dyskinesia and cholelithiasis: a pilot study. J Pediatr Surg 2006, 41, 1545–1548. [Google Scholar] [CrossRef]
- Jennings, L.J.; Salido, G.; Pozo, M.; Davison, J;, Sharkey, K.; Lea, R. et al. The source and action of histamine in the isolated guinea-pig gallbladder. Inflamm Res. 1995, 44, 447–453. [CrossRef] [PubMed]
- Seeger, J.; Stoffel, M.; Simoence, P. Nomina Histologica Veterinaria, submitted by The International Committee on Veterinary Histological Nomenclature (ICVHM) to the World Association of Veterinary Anatomists. ( 2017, submitted.
- Meedeniya, A.C.; Schloithe, A.C.; Toouli, J.; Saccon, G.T. Characterization of the intrinsic and extrinsic innervation of the gall bladder epithelium in the Australian brush-tailed possum (Trichosurus vulpecula). Neurogastroenterol Motil. 2003, 15, 383–392. [Google Scholar] [CrossRef]
- Alexander, W.F. The innervation of the biliary system. J Comp Neurol 1940, 72, 357–370. [Google Scholar] [CrossRef]
- Powley, T.L.; Baronowsky, E.A.; Gilbert, J.M.; Hudson, C.N.; Martin, F.N.; Mason, J.K. , et al. Vagal afferent innervation of the lower esophageal sphincter. Auton Neurosci 2013, 177, 129–142. [Google Scholar] [CrossRef]
- Cai, W.; Gabella, G. Innervation of the gallbladder and biliary pathways in the guinea pig. J Anat 1983, 136, 97–109. [Google Scholar]
- Baumgarten, H.G.; Lange, W. Extrinsic adrenergic innervation of the extrahepatic biliary system in guinea-pigs, cats and rhesus monkeys. Z Zellforsch Mik Ana 1969, 100, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.M. Nerves and Hormones Interact to Control Gallbladder Function. News Physiol Sci 1998, 13, 84–90. [Google Scholar] [CrossRef]
- Gonda, T. , Akiyoshi, H., Ichihara, K. Scanning electron microscopic observations of nerves in the guinea-pig gallbladder after an acetylcholinesterase histochemistry. J Smooth Muscle Res, 1995, 31, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Zanchi, A.; Reidy, J.; Feldma, H.J.; Qualter, J.; Gouw, A.S.; Osbeck, J.; et al. Innervation of the proximal human biliary tree. Virchows Arch, 2020, 477, 385–392. [Google Scholar] [CrossRef]
- Gershon, MD. Nerves, reflexes, and the enteric nervous system: pathogenesis of the irritable bowel syndrome. J. Clin. Gastroenterol, 2005, 39, S184–S193. [Google Scholar] [CrossRef] [PubMed]
- Bioulac-Sage, P.; Lafon, M.E.; Saric, J.; Balabaud, C. Nerves and perisinusoidal cells in human liver. J Hepatol 1990, 10, 105–112. [Google Scholar] [CrossRef] [PubMed]
- McMurray, G; Chris, S.; Johnston, C.F.; Halton, D.W. Choline acetyltransferase (ChAT) immunoreactivity in a sub-population of mammalian intestinal endocrine cells. Comp. Biochem Physiol C Pharmacol Toxicol Endocrinol 1993, 106, 509–515. [CrossRef] [PubMed]
- Pearse, A,G,E. Histochemistry, 3rd edn, Edinburgh, London: Churchill Livingstone 1968, pp. 70–76.
- Furness, J.B.; Bornstein, J.C.; Smith, T.K.; Murphy, R.; Pompolo, S. Correlated functional and structural analysis of enteric neural circuits. Arch Histol Cytol 1989, 52S, 161. [Google Scholar] [CrossRef]
- Nakamura, A.; Yamazaki, K.; Suzuki, K.; Sato, S. Increased portal tract infiltration of mast cells and eosinophils in primary biliary cirrhosis. Am J Gastroenterol 1997, 92, 2245–2249. [Google Scholar]
Figure 1.
A) Moderate reactivity of ChAT+ perikaryons (arrowhead) of ganglia in the muscle layer, moderate to strong reactivity of ChAT+ nerves (large arrows) in the muscle layer, and moderate to strong reactivity of ChAT+ mast cells (small arrows) in the propria (PR) and muscular layer (TM) of ductus cysticus. G – glands with single ChAT+ cells. Bar = 50 µm. B) Moderate reactivity of a ChAT+ neuron (arrowhead) and a ChAT+ nerve (arrows) located in the subserosal layer (S) of ductus cysticus. Bar = 50 µm. C) Moderate to strong reactivity of ChAT+ glandular cells (arrowheads) as well as moderate to strong reactivity of ChAT+ mast cells (small arrows) in lamina propria of the intramural part of the ductus choledochus. D) moderate to strong reactivity of MCstr+ (arrow) in lamina propria of intramural part of ductus choledochus. GL – glands. Bar = 50 µm.
Figure 1.
A) Moderate reactivity of ChAT+ perikaryons (arrowhead) of ganglia in the muscle layer, moderate to strong reactivity of ChAT+ nerves (large arrows) in the muscle layer, and moderate to strong reactivity of ChAT+ mast cells (small arrows) in the propria (PR) and muscular layer (TM) of ductus cysticus. G – glands with single ChAT+ cells. Bar = 50 µm. B) Moderate reactivity of a ChAT+ neuron (arrowhead) and a ChAT+ nerve (arrows) located in the subserosal layer (S) of ductus cysticus. Bar = 50 µm. C) Moderate to strong reactivity of ChAT+ glandular cells (arrowheads) as well as moderate to strong reactivity of ChAT+ mast cells (small arrows) in lamina propria of the intramural part of the ductus choledochus. D) moderate to strong reactivity of MCstr+ (arrow) in lamina propria of intramural part of ductus choledochus. GL – glands. Bar = 50 µm.
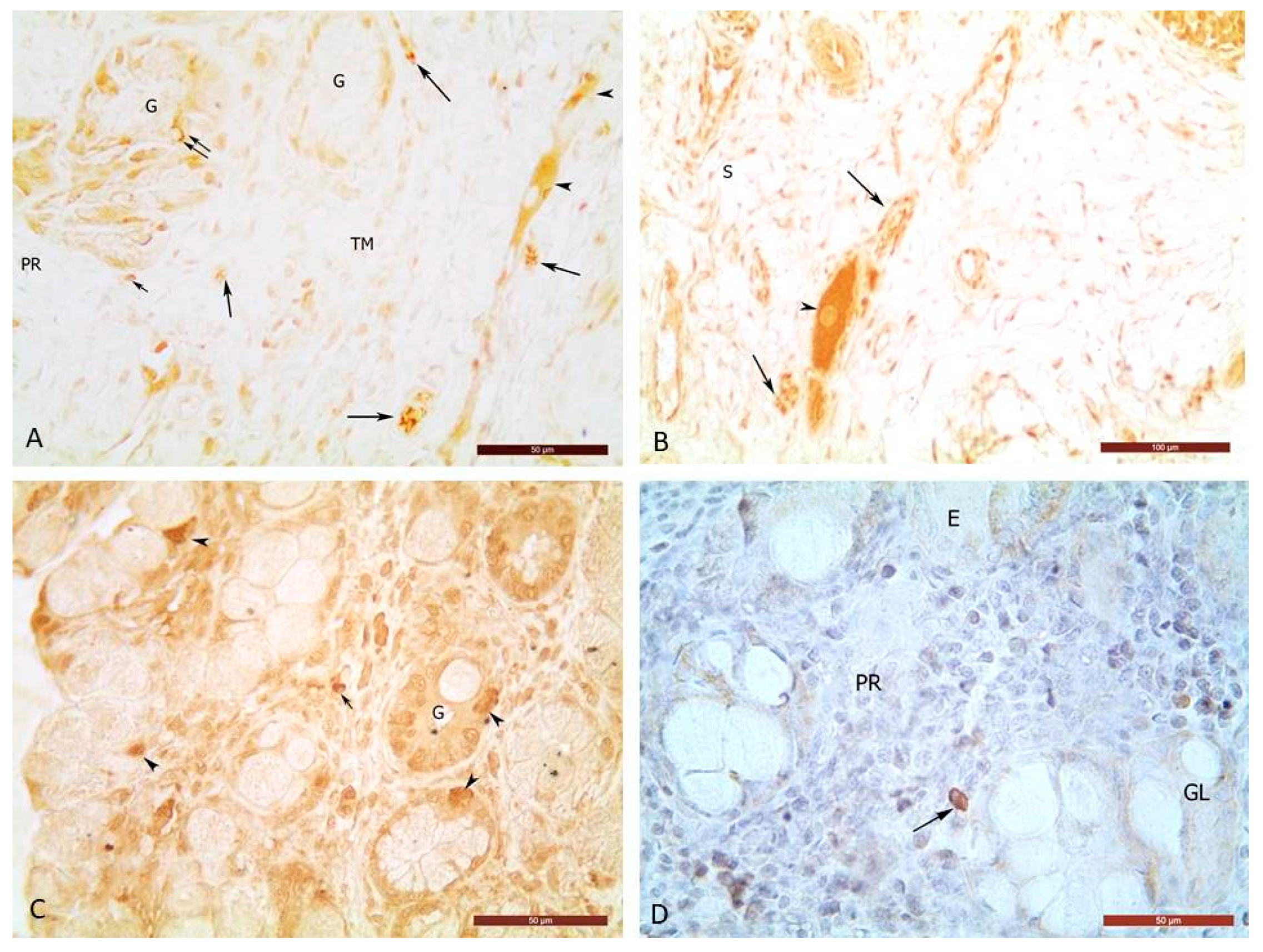

Table 1.
Density (number of ganglia per cross section) and area (µm2) of ganglia in different parts of the gallbladder and in the wall of the ductus cysticus, ductus hepaticus communis (DHC), ductus choledochus (DCH) and papilla duodeni major (PDM) in 6-month-old pigs..
Table 1.
Density (number of ganglia per cross section) and area (µm2) of ganglia in different parts of the gallbladder and in the wall of the ductus cysticus, ductus hepaticus communis (DHC), ductus choledochus (DCH) and papilla duodeni major (PDM) in 6-month-old pigs..
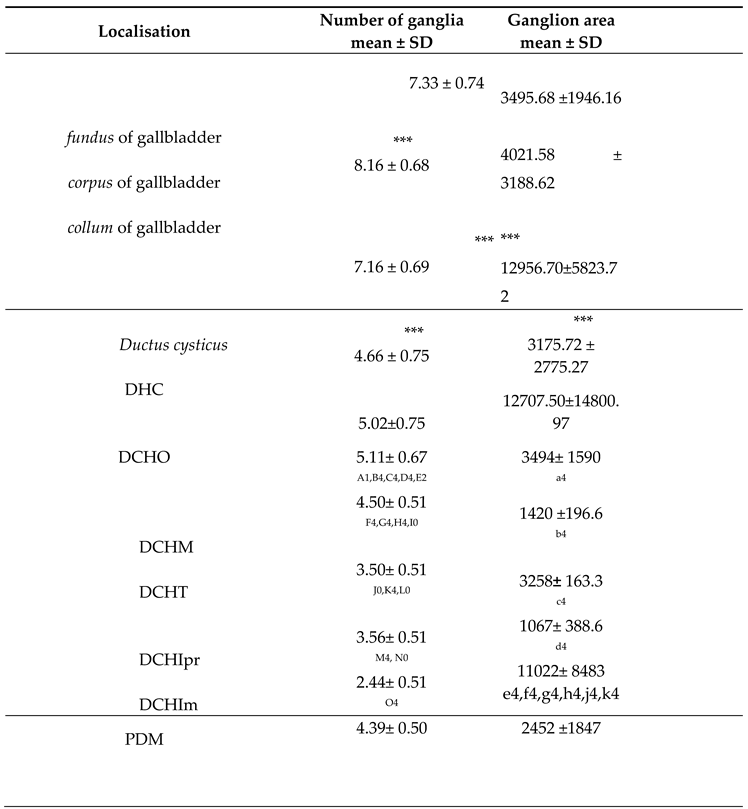
* * p <0,05, ** p <0,01, *** p <0,001, - statistically significant difference compared to the upper row. DCHO - initial segment of DCH; DCHM - middle segment of DCH; DCHT - terminal segment of DCH; DCHIpr - propria in the intramural segment of DCH; DCHMm - tunica muscularis of the intramural segment of DCH 1,2,3,4 - P ˂ 0.05 / 0.01 / 0.001 / 0.0001 A1 - statistically significant difference compared to DCHM; B4 - statistically significant difference compared to DCHT; C4 - statistically significant difference compared to DCHIpr; D4 - statistically significant difference compared to DCHIm; E2 - statistically significant difference compared to PDM; F4 - statistically significant difference compared to DCHT; G4 - statistically significant difference compared to DCHIpr; H4 - statistically significant difference compared to DCHIm; I0 - lack of statistically significant difference compared to PDM; J0 - lack of statistically significant difference compared to DCHIpr; K4 - statistically significant difference compared to DCHIm; L0 - no statistically significant difference compared to PDM; M4 - statistically significant difference compared to DCHIm; N0 - no statistically significant difference compared to PDM; O4 - statistically significant difference to PDM a4 - statistically significant difference compared to DCHIm; b4 - statistically significant difference compared to DCHIm; c4 - statistically significant difference compared to DCHIm; d4 - statistically significant difference compared to DCHIm; e4 - statistically significant difference compared to DCHIm; f4 - statistically significant difference compared to DCHO; g4 - statistically significant difference compared to DCHM; h4 - statistically significant difference compared to DCHT; j4 - statistically significant difference compared to DCHIpr.
Table 2.
Diameter of ChAT+ nerves in the layers of extrahepatic bile ducts (EHBD) and gallbladder (GB).
Table 2.
Diameter of ChAT+ nerves in the layers of extrahepatic bile ducts (EHBD) and gallbladder (GB).
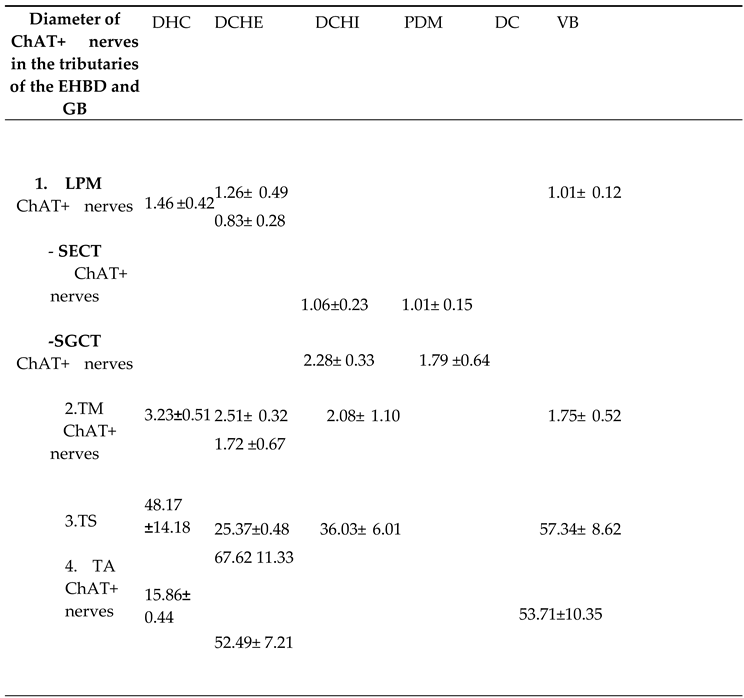
DHC – ductus hepsticus communis, DCHE – extramural part of ductus choledochus, DCHI – intramural part of ductus choledochus, PDM – papilla duodeni major, DC – ductus cisticus, VB – vesica biliaris LPM – lamina propria mucosae, SECT– subepithelial connective tissue layer, SGCT– subglandular connective tissue layer, TM – tunica muscularis, TS – tunica serosa, TA – tunica adventitia.
Table 3.
Density (number of nerves/field of view x400 in LPM, SECT, CGCT, TM, but in TS and TA – number of nerves /1cm) of ChAT+ nerves in EHBD and GB.
Table 3.
Density (number of nerves/field of view x400 in LPM, SECT, CGCT, TM, but in TS and TA – number of nerves /1cm) of ChAT+ nerves in EHBD and GB.
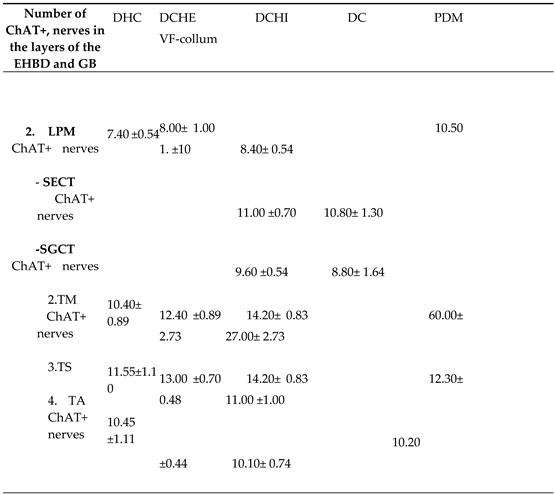
DHC – ductus hepsticus communis, DCHE – extramural part of ductus choledochus, DCHI – intramural part of ductus choledochus, PDM – papilla duodeni major, DC – ductus cisticus, VF– vesica felleaLPM – lamina propria mucosae, SECT– subepithelial connective tissue layer, SGCT– subglandular connective tissue layer, TM – tunica muscularis, TS – tunica serosa, TA – tunica adventitia.
Table 4.
Number of choline acetyltransferase positive cells (ChAT+ C) per microscopic field (ChAT+ C/F) or per gland cross-section (ChAT+ C/GL) in the gallbladder (VF), ductus cysticus (DC), ductus hepaticus communis (DHC), initial segment of the ductus choledochus (DCHO) and intraduodenal part of the ductus choledochus (DCHI), presented as mean ± SD (standard deviation). ChAT+Cot – open type of endocrine cells.
Table 4.
Number of choline acetyltransferase positive cells (ChAT+ C) per microscopic field (ChAT+ C/F) or per gland cross-section (ChAT+ C/GL) in the gallbladder (VF), ductus cysticus (DC), ductus hepaticus communis (DHC), initial segment of the ductus choledochus (DCHO) and intraduodenal part of the ductus choledochus (DCHI), presented as mean ± SD (standard deviation). ChAT+Cot – open type of endocrine cells.
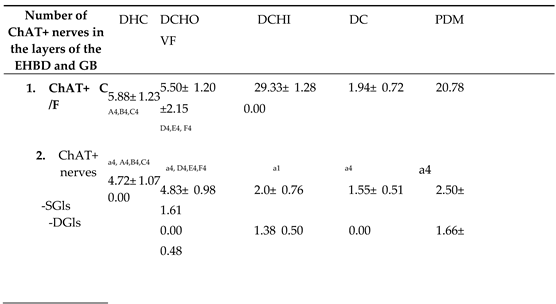
A – Statistically significant difference between DHC and DCHI; B – Statistically significant difference between DHC and DC; C – Statistically significant difference between DHC and PDM; D – Statistically significant difference between DCHO and DCHI; E – Statistically significant difference between DCHO and DC; F – Statistically significant difference between DCHO and PDM; a – Statistically significant difference between superficial glands (SGls) and deep glands (DGls); 1, 2, 3, 4 – p ˂0.05, p ˂0.01, p ˂0.001, p ˂0.0001, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



