
Submitted:
13 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
A rather large portion of the population is sleep restricted in modern society, either by necessity or by choice. Epidemiological studies have observed that lack of sleep and poor sleep quality are associated with several pathological states, ranging from neuropsychiatric to metabolic diseases. To date, only a few studies have investigated the molecular consequences of sleep loss at the level of peripheral organs involved in maintaining metabolic homeostasis such as the white adipose tissue (WAT). From a molecular point of view, two key transcriptional regulators of WAT function are sirtuin 1 (SIRT1) and peroxisome proliferator-activated receptors gamma (PPARγ). Both genes interact with specific targets involved in multiple metabolic processes, including adipocyte differentiation, browning and lipid metabolism. In this study, we assessed the effects of chronic sleep restriction (CSR) on the interplay between SIRT1 and PPARγ in mediating WAT metabolism in young adult mice. We found that CSR up-regulated expression of SIRT1 and some of its transcriptional targets involved in lipid metabolism and mitochondrial functions of adipocytes. Results suggest that CSR induces SIRT1-mediated metabolic remodeling of WAT to support an increased energy demand due to sleep loss.
Keywords:
sleep deprivation
; adipose tissue
; mouse
; qPCR
1. Introduction
White adipose tissue (WAT) is one of the most important organs involved in the regulation of metabolic homeostasis. WAT influences both the endocrine homeostasis and the immune system function by secreting several adipokines, such as leptin [1], adiponectin [2], TNFα [3], and resistin [4] among many others [5]. In addition, WAT adipocytes show significant phenotypic plasticity as demonstrated by their involvement in the browning phenomenon (i.e. a switch from white to beige-like adipocytes) [6]. WAT transcriptional signaling is finely orchestrated by networks of transcription factors that regulate the expression of several target genes. In this regard, Sirtuin 1 (SIRT1) is a NAD+-dependent deacetylase involved in regulating key metabolic pathways in adipocytes [7,8]. SIRT1-related molecular network includes several fatty acid sensors such as peroxisome proliferator-activated receptors (PPARs) [9], and transcription factor coregulators such as peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α), and PR domain containing 16 (PRDM16) [7]. It has recently been reported that the interplay between PPARγ and SIRT1 seems to be critical for WAT browning [10]. This phenotypic change is likely to be protective against obesity-associated metabolic disorders [11]. The importance of SIRT1 signaling in WAT is supported by several in vitro and in vivo studies [10,12]. In fact, deregulation of SIRT1-mediated pathway is closely associated with a range of metabolic alterations as observed in a SIRT1 adipose-specific knockdown mouse model which results in fat accumulation, alterations of glucose and lipid metabolism as well as impaired insulin sensitivity, and development of obesity [12]. There is also evidence that SIRT1 plays an anti-inflammatory effect in WAT by inhibiting the expression of inflammatory response genes [13,14].
One of the environmental factors that can modulate metabolism and has been linked to increased risks of developing obesity is sleep deprivation. Many studies described an inverse or U-shaped relationship between sleep duration and weight gain [15,16,17] with sleep deprivation being associated with increased hunger and appetite, by affecting the leptin and ghrelin signaling pathways [18], increased intake for caloric food [19,20] and higher Body Mass Index (BMI) [16]. In addition to increasing food intake, sleep restriction is responsible for causing endocrine and metabolic alterations, including decreased glucose tolerance, decreased insulin sensitivity [21,22], and increased evening concentrations of cortisol [23,24]. In addition, epidemiologic data correlate short sleep habits with the onset of metabolic-related disorders such as obesity and type-2-diabetes Mellitus, T2DM [25,26]. It has also been suggested that sleep restriction may be an important regulator of energy metabolism in peripheral tissues by increasing insulin resistance in human adipocytes [27]. However, to date, very little is known about the effect of sleep loss on WAT metabolic homeostasis, especially at the molecular level. Seen the progressively decrease in sleep time and the concomitant marked increase in obesity witnesses in the last decades in modern societies, investigating the causal link and the physiological mechanisms involved in sleep loss and WAT metabolism could lead to the identification of new strategies to ameliorate health.
Here, we investigated whether chronic sleep-restriction (CSR), a condition of prolonged short sleep, affects the transcriptional regulation of SIRT1 signaling in mice WAT. Gene-expression analysis was used to identify key Sirt1-regulated metabolic pathways in WAT. The data demonstrated that CSR increased the expression levels of SIRT1 and some of its transcriptional targets (i.e. PPARs, PGC-1A, and sterol regulatory element-binding transcription factor 1/2 - SREBF) involved in lipid metabolism and mitochondrial functions of adipocytes. These data provide new insights into how SIRT1 signaling regulates adipogenic pathways in sleep-restricted mice.
2. Materials and Methods
2.1. Animals
14-week-old male C57BL/6J mice were used for the experiment. Mice were housed in groups of two in environmentally controlled cages for the duration of the experiment (12 h light/dark cycle, light on at 8:00 P.M., the temperature of 24 ± 1°C; food and water available ad libitum and replaced daily at 9:00 A.M.). The bodyweight of mice was measured before and after experimental conditions. All the experiments were performed according to the local Institutional Animal care and Use Committee and the European Communities Council Directives (2010/63/EU) and complied with the 3Rs-Australian Code for the care and use of animals for scientific purposes (Replacement of animals by alternatives wherever possible, Reduction in number of animals used, and Refinement of experimental conditions and procedures to minimize the harm to animals) [28].
2.2. Experimental conditions
The experimental design is summarized in Figure 1. Mice were weight-balanced and divided into two experimental groups: 1) Chronic sleep-restricted (CSR) group (n=8), where mice were sleep restricted for 14 days (24/24h) by an automated sleep deprivation chamber (Pinnacle Technology inc.). The effectiveness of this automated sleep deprivation method has been proved in previous experiments using EEG recording in rodents [29,30]. The procedure consists of a rotating bar placed at a short distance above the cage floor, lightly nudging the animal from sleep and encouraging low levels of activity until the animal maintains wakefulness on its own. The rotating bar was set counterclockwise, at a velocity of 2rpm (one turn each 30s). 2) Control (CTR) group (n=8), where mice were placed in the same sleep deprivation chambers and allowed to sleep undisturbed, except for 3 hours/day (during the dark period, when mice are usually awake) during which the bar rotation was activated to expose the mice of this group to the experience of the bar movement and the stress associated with it. Animal behavior was daily assessed by direct visual observation. After 14 days, all mice were sacrificed between 9:00 and 11:00 A.M. to maintain the time of tissue collection within the same 2-h time of day window for all experimental groups.
2.3. WAT Collection
All mice were deeply anesthetized with isoflurane 3%. Mice were sacrificed by cervical dislocation, immediately immersed in a freezing 2-methylbutane solution for a few seconds and dissected for WAT collection. To get access to visceral organs, the central skin near the genital organs was elevated and a small incision was performed. Then, the scissor was put horizontally into the opened hole, allowing to cut the abdominal muscle along linea alba (for ~4 cm, depending on the mouse size) from the genitals to the rib cage. Consistently, WAT was collected in the same specific area for all mice. The fat used in this study was perigonadal fat (PGF), localized around the reproductive organs. PGF was collected by separating it from surrounding structures (epididymis, the testis, and the ductus deferens). All samples were immediately frozen in dry ice and subsequently stored in a -80 °C freezer for later analysis.
2.4. Molecular Analyses
Total RNA was extracted from 50 mg of WAT using 0.5 mL QIAzol® Lysis Reagent according to the manufacturer’s instructions (QIAzol® Lysis Reagent, QIAGEN® Sample & Assay Technologies). RNA concentration and purity were assessed spectrophotometrically at an absorbance of 260/280 nm, and the integrity was confirmed by electrophoresis through 1% agarose gels stained with ethidium bromide. The complementary DNA (cDNA) was synthesized from 2 µg of total RNA in 20 µL of total volume reaction using 4 µl of 5X All-In-One RT MasterMix according to the manufacturer’s instructions (abm®, Cat. No. G592). The optimized 5X RT MasterMix contains abm’s proprietary OneScript® Hot Reverse Transcriptase, RNaseOFF Ribonuclease Inhibitor, temperature-sensitive DNase, dNTPs, and a finely balanced ratio of Oligo (dT)s and Random Primers. The mixture was incubated at 37°C for 15 minutes, followed by 45°C for 60 minutes. The cDNA synthesis reaction was stopped by heating at 85°C for 2 minutes. SYBR green-based real-time PCR (q-PCR) was used to evaluate expression profiles of PPARG, PPARA, SIRT-1, LEPT, PGC-1A, PRDM16, C-Myc, SREBF, CEBP, UCP-1 target genes. The well-known housekeeping gene, Glucose-6-phosphate dehydrogenase (G6PDH), was used as a reference gene for the qPCR analysis since its expression was unaffected by experimental conditions [31]. All the primer sequences are reported in Table 1. The expression of individual gene targets was analyzed using the ABI 7300 Real-Time PCR System (Applied Biosystems Inc.). All reactions were prepared in 20 µl of total volume using BlasTaq™ 2X qPCR MasterMix according to the manufacturer’s instructions. Briefly, the reaction included 10 μL BlasTaq™ 2X qPCR MM (Abm®), 10 μmol L−1 each of forward and reverse primers, 1 μL cDNA template, and sterile distilled water (Abm®). Thermocycling for all reactions was for 3 min 95 °C, followed by 40 cycles of 15 s at 95 °C and 60 s at 60 °C. Fluorescence was monitored at the end of every cycle. Melting curve analysis has been performed to confirm the presence of a single amplicon of interest. Results were calculated using the relative 2 -ΔΔCt method [39] and means of mRNA levels were expressed with respect to control mice ± standard deviation (SD).
2.5. STRING network visualization
Cluster analysis of genes selected in the present study was performed using the K-means clustering method in STRING (Search Tool for the Retrieval of Interacting Genes/Proteins) [40,41] including only interactions with a confidence score higher than 0.7. The STRING v11.5 web resource [42] is a database which gives an association score for interacting proteins, calculated using various parameters such as neighborhood score, fusion, co-occurrence, homology, co-expression, experimental, database and text mining score [43]. Recently, STRING v11.5 has been also successfully applied to identify gene interaction networks [41,44]. To simplify the dense network and to obtain a better visual representation and comprehension of the interactions, the resulting groups were separated manually.
2.6. Statistical Analysis
q-PCR results were expressed as normalized fold change corrected for G6PDH and with respect to the control group (2 -ΔΔCt method). Data were first examined for their fit to a normal distribution and homogeneity of variance using Kolmogorov–Smirnov and Levene median tests. Student’s t-test was used to determine statistical differences between CSR and Control group. All statistical analyses were performed with GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA).
3. Results
The expression levels of SIRT1 were significantly increased in CSR mice with respect to control animals (Figure 2A, p<0.001). Similarly, the signaling pathway downstream of activated SIRT1 (i.e., PPARα/γ, PGC-1α, LEPT, and SREBF) was significantly up-regulated by sleep restriction (Figure 2A, p<0.05). On the contrary, CEBP-β and C-myc were down-regulated in sleep restricted mice (Figure 2A, p<0.05). Both PRDM16 and UCP-1 did not show any significant difference in mRNA levels between experimental groups (Figure 2A). The K-means clustering analysis in STRING identified four main clusters (Figure 2B). All genes within and between each cluster are strongly interconnected, reflecting a high degree of functional association, and suggesting an interplay among the numerous pathways related to the gene network. In this regard, we identified genes that 1) are involved in the transcriptional regulation of lipid homeostasis (green group), 2) are defined as DNA-binding transcription factors during the adipogenesis process (yellow group), 3-4) act as regulators for adipose tissue development (LEPT) and fat cell differentiation (UCP-1). Our results also suggest that induction of SIRT1-dependent molecular pathways prevents weight gain. Indeed, body weights of CSR mice remained almost unchanged after 14 days of treatment, while the body weights of control mice were found significantly increased at the end of the experiment (Figure 2C, p<0,001).
4. Discussion
In this study, we investigated the effects of CSR on SIRT1-dependent molecular pathways in WAT of mice models. The expression levels of SIRT1 were significantly increased in CSR mice with respect to control animals. Similarly, the signaling pathway downstream of activated SIRT1 (i.e., PPARα/γ, PGC-1α, LEPT, and SREBF) was significantly up-regulated by sleep restriction. On the contrary, CEBP-β and C-myc were down-regulated in sleep restricted mice. Both PRDM16 and UCP-1 did not show any significant difference in mRNA levels between experimental groups. Our findings are consistent with the role of SIRT1 activation in adipose tissue signaling as previously suggested by different studies and well described by Boutant and Cantó [7]. Indeed, up-regulation of SIRT1 was found to decrease the mRNA levels of key WAT transcription factors and up-regulate those of brown adipose tissue (BAT) genes. Importantly, genes whose expression is induced by SIRT1-activation include those involved in mitochondrial biogenesis such as PGC-1α, and fatty acid-oxidation such as PPARα [9,45]. Majeed et al. [10] reported that PGC-1α expression levels were promoted by SIRT1 as demonstrated using SIRT1-depleted adipocyte models which in turn showed reduced mitochondrial mass and respiratory capacity. On the other hand, activation of PPARα is pivotal in regulating lipid metabolism and promoting fatty acid oxidation [45]. SIRT1:PPARα interaction was protective in cardiac hypertrophy [46] and deregulation of its-related signaling pathway was associated with reduced fatty acid-oxidation in the liver [47]. Interestingly, the CSR-induced up-regulation of PPARα paired that of SREBF. As previously demonstrated, SREBF1 overexpression in 3T3-L1 adipocytes promoted fatty acid metabolism and resulted in the production of lipids that work as PPARγ ligands [48,49]. Therefore, these findings could explain, at least in part, the increase in PPARγ expression observed in our study. There is indeed evidence that, in WAT, SIRT1 usually acts as PPARγ repressor rather than inducer [50]. This effect is most likely due to a SIRT1-dependent deacetylation of PPARγ that subsequently causes the recruitment of PRDM16, a transcriptional co-regulator of the BAT genetic program. Importantly, SIRT1-dependent deacetylation of PPARγ is responsible for UCP-1 induction in WAT [51]. However, we found no change in UCP-1 nor PDRM16 expression, which could indicate unaffected acetylation levels of PPARγ in WAT. Other studies found increased UCP mRNA and protein levels in BAT and muscles of chronic sleep restricted rats suggesting increased energy expenditure associated with prolonged sleep loss [52,53].
Our study also indicates that sleep deprivation significantly increased the expression of the adipokine leptin in the CSR group with respect to control mice. From a molecular point of view, it is possible that a SIRT1-mediated increase in CEBP-α transcriptional activity is responsible for the induction of the leptin gene in response to CSR. Stimulation of leptin transcription was previously found to be SIRT1-dependent in preadipocytes supplemented with the NAD+ booster nicotinamide mononucleotide (NMN) [10]. In addition, leptin expression is highly correlated with PPARγ expression, which was also increased in CSR mice [54]. However, in the literature there are conflicting results regarding leptin circulation in sleep curtailment, with its level found to be both reduced [55,56,57] and elevated [58,59,60,61]. Discrepancies may be related to the duration and quality of sleep disturbances applied.
Overall, our data suggest the presence of a functional interaction among SIRT1, PPARα, SREBF and PGC-1α that results in reduced fat accumulation and enhanced fat consumption probably due to a stimulation of the adipose thermogenic capacity. Together with leptin, all these genes act as metabolic regulators whose expression patterns are sensitive to CSR in a SIRT1-dependent manner. The CSR-induced downregulation of genes associated with the adipogenic pathway (i.e., CEBP-β, and C-Myc) is also consistent with the reduction in fat accumulation. Earlier evidence has shown that adipogenesis as well as adipocyte differentiation is driven by two subsequent waves of transcription factor activation during which CEBP-β plays a pivotal function [62,63]. CEBP-β is a well-known early mediator of adipocyte differentiation and is involved in the activation of several targets including PPARγ, which in turn mediate adipogenic differentiation [64]. Interestingly, CEBP-β shows polylysine acetylation which may be a deacetylation target of SIRT1 [65]. Consequently, SIRT1-dependent deacetylation alters CEBP-β function as a transcription factor in adipocytes. In line with this, SIRT1 was shown to inhibit C-Myc signaling as demonstrated using SIRT1-silenced preadipocytes [66]. SIRT1-mediated deacetylation of C-Myc resulted in reduced preadipocyte hyperplasia, lipid accumulation and inflammation [66]. In addition, Tóth et al. [67] suggested that C-Myc could participate in the regulation of adipocyte-thermogenic function by modulating UCP1 gene expression. Overall, the CSR-induced overexpression of SIRT1 matches repression of both CEBP-β and C-Myc.
Our results also suggest that induction of SIRT1-dependent molecular pathways prevents weight gain. Indeed, body weights of CSR mice remained almost unchanged after 14 days of treatment, while the body weights of control mice were found significantly increased at the end of the experiment. We did not measure food consumption nor metabolic expenditure is in line with what observed by other groups. Chronic sleep restriction in rodents is known to cause a cluster of syndromes first described by Rechtschaffen and Bergmann [68] and that include hyperphagia, weight loss, elevated energy expenditure, increased plasma catecholamines, hypothyroidism, reduction in core temperature, deterioration in physical appearance [69] levels of anabolic hormones [70] and declines in integrity of the immune system [71]. Reduced weight gain occurs consistently despite unchanged or increased levels of food consumption [52,72,73]. Our findings suggest that, in addition to increased energy expenditure associated with sleep deprivation, overexpression of SIRT-1 signaling could be involved in the phenomenon of loss of body weight, mostly by inducing a decrease in fat accumulation.
Finally, the K-means clustering analysis in STRING identified four main clusters. All genes within and between each cluster are strongly interconnected, reflecting a high degree of functional association, and suggesting an interplay among the numerous pathways related to the gene network. In this regard, we identified genes that 1) are involved in the transcriptional regulation of lipid homeostasis (green group), 2) are defined as DNA-binding transcription factors during the adipogenesis process (yellow group), 3) act as regulators for adipose tissue development (LEP) and fat cell differentiation (UCP-1). As a whole, the obtained gene network highlights the crucial role of SIRT1-dependent molecular pathways in mediating the metabolic remodeling of adipose tissue induced by CSR.
5. Conclusions
Altogether, our results indicate that CSR induces SIRT1-mediated metabolic remodeling of WAT. Overexpression of SIRT1 decreases lipid accumulation probably by inducing fat oxidation and increasing adaptive thermogenesis. However, further studies are needed to characterize the temporal dynamics of thermogenic gene expression and the long-term effects of WAT adaptation to CSR.
Author Contributions
Conceptualization, F.A.P., M.R. and M.B.; methodology, M.B., F.A.P. and P.C..; software, F.A.P., P.C. and M.R.; validation, F.A.P.; formal analysis, M.R. and P.C..; investigation, M.R. and P.C..; resources, M.B., d.V.L. and F.P.A..; data curation, F.A.P and P.C..; writing—original draft preparation, M.R..; writing—review and editing, F.A.P. and M.B..; visualization, F.A.P.; supervision, M.B., d.V.L. and F.A.P..; project administration, F.A.P and M.B..; funding acquisition, F.A.P. and M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All the experiments were performed according to the local Institutional Animal care and Use Committee and the European Communities Council Directives (2010/63/EU).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S. Leptin Levels in Human and Rodent: Measurement of Plasma Leptin and Ob RNA in Obese and Weight-Reduced Subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A Novel Serum Protein Similar to C1q, Produced Exclusively in Adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Arner, P.; Atkinson, R.L.; Spiegelman, B.M. Differential Regulation of the P80 Tumor Necrosis Factor Receptor in Human Obesity and Insulin Resistance. Diabetes 1997, 46, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The Hormone Resistin Links Obesity to Diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef]
- Rodríguez, A.; Ezquerro, S.; Méndez-Giménez, L.; Becerril, S.; Frühbeck, G. Revisiting the Adipocyte: A Model for Integration of Cytokine Signaling in the Regulation of Energy Metabolism. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E691–714. [Google Scholar] [CrossRef] [PubMed]
- Kuryłowicz, A.; Puzianowska-Kuźnicka, M. Induction of Adipose Tissue Browning as a Strategy to Combat Obesity. Int. J. Mol. Sci. 2020, 21, 6241. [Google Scholar] [CrossRef] [PubMed]
- Boutant, M.; Cantó, C. SIRT1 Metabolic Actions: Integrating Recent Advances from Mouse Models. Mol. Metab. 2014, 3, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Kajimura, S.; Yang, W.; Chin, S.; Rohas, L.M.; Uldry, M.; Tavernier, G.; Langin, D.; Spiegelman, B.M. Transcriptional Control of Brown Fat Determination by PRDM16. Cell Metab. 2007, 6, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zhang, Y.; Breyer, M.D. The Role of PPARs in the Transcriptional Control of Cellular Processes. Drug News Perspect. 2002, 15, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Majeed, Y.; Halabi, N.; Madani, A.Y.; Engelke, R.; Bhagwat, A.M.; Abdesselem, H.; Agha, M.V.; Vakayil, M.; Courjaret, R.; Goswami, N.; et al. SIRT1 Promotes Lipid Metabolism and Mitochondrial Biogenesis in Adipocytes and Coordinates Adipogenesis by Targeting Key Enzymatic Pathways. Sci. Rep. 2021, 11, 8177. [Google Scholar] [CrossRef]
- Smorlesi, A.; Frontini, A.; Giordano, A.; Cinti, S. The Adipose Organ: White-Brown Adipocyte Plasticity and Metabolic Inflammation: Adipocyte Plasticity and Adipose Organ. Obes. Rev. 2012, 13 Suppl 2, 83–96. [Google Scholar] [CrossRef]
- Li, F.; Li, H.; Jin, X.; Zhang, Y.; Kang, X.; Zhang, Z.; Xu, M.; Qian, Z.; Ma, Z.; Gao, X.; et al. Adipose-Specific Knockdown of Sirt1 Results in Obesity and Insulin Resistance by Promoting Exosomes Release. Cell Cycle 2019, 18, 2067–2082. [Google Scholar] [CrossRef]
- Chalkiadaki, A.; Guarente, L. High-Fat Diet Triggers Inflammation-Induced Cleavage of SIRT1 in Adipose Tissue to Promote Metabolic Dysfunction. Cell Metab. 2012, 16, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Milne, J.C.; Imamura, T.; Schenk, S.; Sonoda, N.; Babendure, J.L.; Lu, J.-C.; Smith, J.J.; Jirousek, M.R.; Olefsky, J.M. SIRT1 Exerts Anti-Inflammatory Effects and Improves Insulin Sensitivity in Adipocytes. Mol. Cell. Biol. 2009, 29, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Cappuccio, F.P.; Taggart, F.M.; Kandala, N.-B.; Currie, A.; Peile, E.; Stranges, S.; Miller, M.A. Meta-Analysis of Short Sleep Duration and Obesity in Children and Adults. Sleep 2008, 31, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kruisbrink, M.; Wallace, J.; Ji, C.; Cappuccio, F.P. Sleep Duration and Incidence of Obesity in Infants, Children, and Adolescents: A Systematic Review and Meta-Analysis of Prospective Studies. Sleep 2018, 41. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhai, L.; Zhang, D. Sleep Duration and Obesity among Adults: A Meta-Analysis of Prospective Studies. Sleep Med. 2014, 15, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, K.; Leproult, R.; L’hermite-Balériaux, M.; Copinschi, G.; Penev, P.D.; Van Cauter, E. Leptin Levels Are Dependent on Sleep Duration: Relationships with Sympathovagal Balance, Carbohydrate Regulation, Cortisol, and Thyrotropin. J. Clin. Endocrinol. Metab. 2004, 89, 5762–5771. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, A.M.; Dinges, D.F.; Goel, N. Sex and Race Differences in Caloric Intake during Sleep Restriction in Healthy Adults. Am. J. Clin. Nutr. 2014, 100, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Shi, C.; Park, C.G.; Zhao, X.; Reutrakul, S. Effects of Sleep Restriction on Metabolism-Related Parameters in Healthy Adults: A Comprehensive Review and Meta-Analysis of Randomized Controlled Trials. Sleep Med. Rev. 2019, 45, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.N.; Neylan, T.C.; Grunfeld, C.; Mulligan, K.; Schambelan, M.; Schwarz, J.-M. Subchronic Sleep Restriction Causes Tissue-Specific Insulin Resistance. J. Clin. Endocrinol. Metab. 2015, 100, 1664–1671. [Google Scholar] [CrossRef] [PubMed]
- Nedeltcheva, A.V.; Kessler, L.; Imperial, J.; Penev, P.D. Exposure to Recurrent Sleep Restriction in the Setting of High Caloric Intake and Physical Inactivity Results in Increased Insulin Resistance and Reduced Glucose Tolerance. J. Clin. Endocrinol. Metab. 2009, 94, 3242–3250. [Google Scholar] [CrossRef] [PubMed]
- Omisade, A.; Buxton, O.M.; Rusak, B. Impact of Acute Sleep Restriction on Cortisol and Leptin Levels in Young Women. Physiol. Behav. 2010, 99, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Guyon, A.; Morselli, L.L.; Balbo, M.L.; Tasali, E.; Leproult, R.; L’Hermite-Balériaux, M.; Van Cauter, E.; Spiegel, K. Effects of Insufficient Sleep on Pituitary-Adrenocortical Response to CRH Stimulation in Healthy Men. Sleep 2017, 40. [Google Scholar] [CrossRef] [PubMed]
- Itani, O.; Jike, M.; Watanabe, N.; Kaneita, Y. Short Sleep Duration and Health Outcomes: A Systematic Review, Meta-Analysis, and Meta-Regression. Sleep Med. 2017, 32, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Semba, E.; Yamakawa, T.; Terauchi, Y. Associations of Impaired Glucose Tolerance and Sleep Disorders with Mortality among the US General Population. BMJ Open Diabetes Res. Care 2021, 9, e002047. [Google Scholar] [CrossRef]
- Broussard, J.L.; Ehrmann, D.A.; Van Cauter, E.; Tasali, E.; Brady, M.J. Impaired Insulin Signaling in Human Adipocytes after Experimental Sleep Restriction: A Randomized, Crossover Study: A Randomized, Crossover Study. Ann. Intern. Med. 2012, 157, 549–557. [Google Scholar] [CrossRef]
- Schuppli, C.A.; Fraser, D. The Interpretation and Application of the Three Rs by Animal Ethics Committee Members. Altern. Lab. Anim. 2005, 33, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Fenzl, T.; Romanowski, C.P.N.; Flachskamm, C.; Honsberg, K.; Boll, E.; Hoehne, A.; Kimura, M. Fully Automated Sleep Deprivation in Mice as a Tool in Sleep Research. J. Neurosci. Methods 2007, 166, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, B.M.; Kushida, C.A.; Everson, C.A.; Gilliland, M.A.; Obermeyer, W.; Rechtschaffen, A. Sleep Deprivation in the Rat: II. Methodology. Sleep 1989, 12, 5–12. [Google Scholar] [CrossRef]
- Barbu, A.; Hedlund, G.P.; Lind, J.; Carlsson, C. Pref-1 and Adipokine Expression in Adipose Tissues of GK and Zucker Rats. Mol. Cell. Endocrinol. 2009, 299, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mandard, S.; Zandbergen, F.; Tan, N.S.; Escher, P.; Patsouris, D.; Koenig, W.; Kleemann, R.; Bakker, A.; Veenman, F.; Wahli, W.; et al. The Direct Peroxisome Proliferator-Activated Receptor Target Fasting-Induced Adipose Factor (FIAF/PGAR/ANGPTL4) Is Present in Blood Plasma as a Truncated Protein That Is Increased by Fenofibrate Treatment. J. Biol. Chem. 2004, 279, 34411–34420. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Qu, D.; Zhang, Q.; Jiang, J.; Zhou, H.; Jiang, R.; Li, Y.; Zhang, Y.; Yan, H. Corrigendum: Phytosterol Esters Attenuate Hepatic Steatosis in Rats with Non-Alcoholic Fatty Liver Disease Rats Fed a High-Fat Diet. Sci. Rep. 2017, 7, 46884. [Google Scholar] [CrossRef] [PubMed]
- Nøhr, M.; Bobba, N.; Richelsen, B.; Lund, S.; Pedersen, S. Inflammation Downregulates UCP1 Expression in Brown Adipocytes Potentially via SIRT1 and DBC1 Interaction. Int. J. Mol. Sci. 2017, 18, 1006. [Google Scholar] [CrossRef] [PubMed]
- Volat, F.E.; Pointud, J.-C.; Pastel, E.; Morio, B.; Sion, B.; Hamard, G.; Guichardant, M.; Colas, R.; Lefrançois-Martinez, A.-M.; Martinez, A. Depressed Levels of Prostaglandin F2α in Mice Lacking Akr1b7 Increase Basal Adiposity and Predispose to Diet-Induced Obesity. Diabetes 2012, 61, 2796–2806. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Kido, Y.; Asahara, S.-I.; Kaisho, T.; Tanaka, T.; Hashimoto, N.; Shigeyama, Y.; Takeda, A.; Inoue, T.; Shibutani, Y.; et al. Ablation of C/EBPbeta Alleviates ER Stress and Pancreatic Beta Cell Failure through the GRP78 Chaperone in Mice. J. Clin. Invest. 2010, 120, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Ciribilli, Y.; Borlak, J. Correction: Oncogenomics of c-Myc Transgenic Mice Reveal Novel Regulators of Extracellular Signaling, Angiogenesis and Invasion with Clinical Significance for Human Lung Adenocarcinoma. Oncotarget 2018, 9, 37269–37269. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jia, Y.; Yang, G.; Zhang, X.; Boddu, P.C.; Petersen, B.; Narsingam, S.; Zhu, Y.-J.; Thimmapaya, B.; Kanwar, Y.S.; et al. PPARα-Deficient Ob/Ob Obese Mice Become More Obese and Manifest Severe Hepatic Steatosis Due to Decreased Fatty Acid Oxidation. Am. J. Pathol. 2015, 185, 1396–1408. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Poswar, F. de O.; Farias, L.C.; Fraga, C.A. de C.; Bambirra, W., Jr; Brito-Júnior, M.; Sousa-Neto, M.D.; Santos, S.H.S.; de Paula, A.M.B.; D’Angelo, M.F.S.V.; Guimarães, A.L.S. Bioinformatics, Interaction Network Analysis, and Neural Networks to Characterize Gene Expression of Radicular Cyst and Periapical Granuloma. J. Endod. 2015, 41, 877–883. [CrossRef]
- Kumar, R.; Samal, S.K.; Routray, S.; Dash, R.; Dixit, A. Publisher Correction: Identification of Oral Cancer Related Candidate Genes by Integrating Protein-Protein Interactions, Gene Ontology, Pathway Analysis and Immunohistochemistry. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- STRING: Functional Protein Association Networks. Available online: http://string.embl.de (accessed on 21 November 2023).
- Korbel, J.O.; Jensen, L.J.; von Mering, C.; Bork, P. Analysis of Genomic Context: Prediction of Functional Associations from Conserved Bidirectionally Transcribed Gene Pairs. Nat. Biotechnol. 2004, 22, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. bioRxiv 2018. [CrossRef] [PubMed]
- Monsalve, F.A.; Pyarasani, R.D.; Delgado-Lopez, F.; Moore-Carrasco, R. Peroxisome Proliferator-Activated Receptor Targets for the Treatment of Metabolic Diseases. Mediators Inflamm. 2013, 2013, 549627. [Google Scholar] [CrossRef] [PubMed]
- Planavila, A.; Iglesias, R.; Giralt, M.; Villarroya, F. Sirt1 Acts in Association with PPARα to Protect the Heart from Hypertrophy, Metabolic Dysregulation, and Inflammation. Cardiovasc. Res. 2011, 90, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Purushotham, A.; Schug, T.T.; Xu, Q.; Surapureddi, S.; Guo, X.; Li, X. Hepatocyte-Specific Deletion of SIRT1 Alters Fatty Acid Metabolism and Results in Hepatic Steatosis and Inflammation. Cell Metab. 2009, 9, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 Promotes Adipocyte Differentiation and Gene Expression Linked to Fatty Acid Metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Wright, H.M.; Wright, M.; Spiegelman, B.M. ADD1/SREBP1 Activates PPARgamma through the Production of Endogenous Ligand. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 4333–4337. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado De Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 Promotes Fat Mobilization in White Adipocytes by Repressing PPAR-Gamma. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown Remodeling of White Adipose Tissue by SirT1-Dependent Deacetylation of Pparγ. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef]
- Koban, M.; Swinson, K.L. Chronic REM-Sleep Deprivation of Rats Elevates Metabolic Rate and Increases UCP1 Gene Expression in Brown Adipose Tissue. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E68–74. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Tononi, G. Uncoupling Proteins and Sleep Deprivation. Arch. Ital. Biol. 2004, 142, 541–549. [Google Scholar] [PubMed]
- Yamauchi, T.; Waki, H.; Kamon, J.; Murakami, K.; Motojima, K.; Komeda, K.; Miki, H.; Kubota, N.; Terauchi, Y.; Tsuchida, A.; et al. Inhibition of RXR and PPARγ Ameliorates Diet-Induced Obesity and Type 2 Diabetes. J. Clin. Invest. 2001, 108, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Lin, L.; Austin, D.; Young, T.; Mignot, E. Short Sleep Duration Is Associated with Reduced Leptin, Elevated Ghrelin, and Increased Body Mass Index. PLoS Med. 2004, 1, e62. [Google Scholar] [CrossRef] [PubMed]
- Chaput, J.-P.; Després, J.-P.; Bouchard, C.; Tremblay, A. Short Sleep Duration Is Associated with Reduced Leptin Levels and Increased Adiposity: Results from the Québec Family Study. Obesity (Silver Spring) 2007, 15, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Grant, A.S.; Thomson, C.A.; Tinker, L.; Hale, L.; Brennan, K.M.; Woods, N.F.; Chen, Z. Short Sleep Duration Is Associated with Decreased Serum Leptin, Increased Energy Intake and Decreased Diet Quality in Postmenopausal Women. Obesity (Silver Spring) 2014, 22, E55–61. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.C.; Dorrian, J.; Liu, P.Y.; Van Dongen, H.P.A.; Wittert, G.A.; Harmer, L.J.; Banks, S. Impact of Five Nights of Sleep Restriction on Glucose Metabolism, Leptin and Testosterone in Young Adult Men. PLoS One 2012, 7, e41218. [Google Scholar] [CrossRef] [PubMed]
- Simpson, N.S.; Banks, S.; Dinges, D.F. Sleep Restriction Is Associated with Increased Morning Plasma Leptin Concentrations, Especially in Women. Biol. Res. Nurs. 2010, 12, 47–53. [Google Scholar] [CrossRef]
- Hayes, A.L.; Xu, F.; Babineau, D.; Patel, S.R. Sleep Duration and Circulating Adipokine Levels. Sleep 2011, 34, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Pejovic, S.; Vgontzas, A.N.; Basta, M.; Tsaoussoglou, M.; Zoumakis, E.; Vgontzas, A.; Bixler, E.O.; Chrousos, G.P. Leptin and Hunger Levels in Young Healthy Adults after One Night of Sleep Loss: Leptin after One Night of Sleep Loss. J. Sleep Res. 2010, 19, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr; Liu, X.S.; et al. PPARgamma and C/EBP Factors Orchestrate Adipocyte Biology via Adjacent Binding on a Genome-Wide Scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, X.; Tang, Q.-Q. Transcriptional Regulation of Adipocyte Differentiation: A Central Role for CCAAT/Enhancer-Binding Protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Siersbæk, R.; Nielsen, R.; Mandrup, S. Transcriptional Networks and Chromatin Remodeling Controlling Adipogenesis. Trends Endocrinol. Metab. 2012, 23, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Ceseña, T.I.; Cardinaux, J.-R.; Kwok, R.; Schwartz, J. CCAAT/Enhancer-Binding Protein (C/EBP) Beta Is Acetylated at Multiple Lysines: Acetylation of C/EBPbeta at Lysine 39 Modulates Its Ability to Activate Transcription. J. Biol. Chem. 2007, 282, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Abdesselem, H.; Madani, A.; Hani, A.; Al-Noubi, M.; Goswami, N.; Ben Hamidane, H.; Billing, A.M.; Pasquier, J.; Bonkowski, M.S.; Halabi, N.; et al. SIRT1 Limits Adipocyte Hyperplasia through C-Myc Inhibition. J. Biol. Chem. 2016, 291, 2119–2135. [Google Scholar] [CrossRef] [PubMed]
- B Tóth, B.; Barta, Z.; Barta, Á.B.; Fésüs, L. Regulatory Modules of Human Thermogenic Adipocytes: Functional Genomics of Large Cohort and Meta-Analysis Derived Marker-Genes. BMC Genomics 2021, 22, 886. [Google Scholar] [CrossRef] [PubMed]
- Rechtschaffen, A.; Gilliland, M.A.; Bergmann, B.M.; Winter, J.B. Physiological Correlates of Prolonged Sleep Deprivation in Rats. Science 1983, 221, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Rechtschaffen, A.; Bergmann, B.M. Sleep Deprivation in the Rat: An Update of the 1989 Paper. Sleep 2002, 25, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Everson, C.A.; Crowley, W.R. Reductions in Circulating Anabolic Hormones Induced by Sustained Sleep Deprivation in Rats. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E1060–70. [Google Scholar] [CrossRef] [PubMed]
- Everson, C.A.; Toth, L.A. Systemic Bacterial Invasion Induced by Sleep Deprivation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R905–16. [Google Scholar] [CrossRef] [PubMed]
- Barf, R.P.; Van Dijk, G.; Scheurink, A.J.W.; Hoffmann, K.; Novati, A.; Hulshof, H.J.; Fuchs, E.; Meerlo, P. Metabolic Consequences of Chronic Sleep Restriction in Rats: Changes in Body Weight Regulation and Energy Expenditure. Physiol. Behav. 2012, 107, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Baud, M.O.; Magistretti, P.J.; Petit, J.-M. Sustained Sleep Fragmentation Affects Brain Temperature, Food Intake and Glucose Tolerance in Mice: Sleep Fragmentation Induces Metabolic Impairments. J. Sleep Res. 2013, 22, 3–12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental Design. Young adult mice were divided into two groups: Chronic sleep restriction (CSR) and Control group (CTR). Mice were exposed to either CSR or 3hr bar movement for 14 days, then tissue was collected for molecular analysis.
Figure 1.
Experimental Design. Young adult mice were divided into two groups: Chronic sleep restriction (CSR) and Control group (CTR). Mice were exposed to either CSR or 3hr bar movement for 14 days, then tissue was collected for molecular analysis.
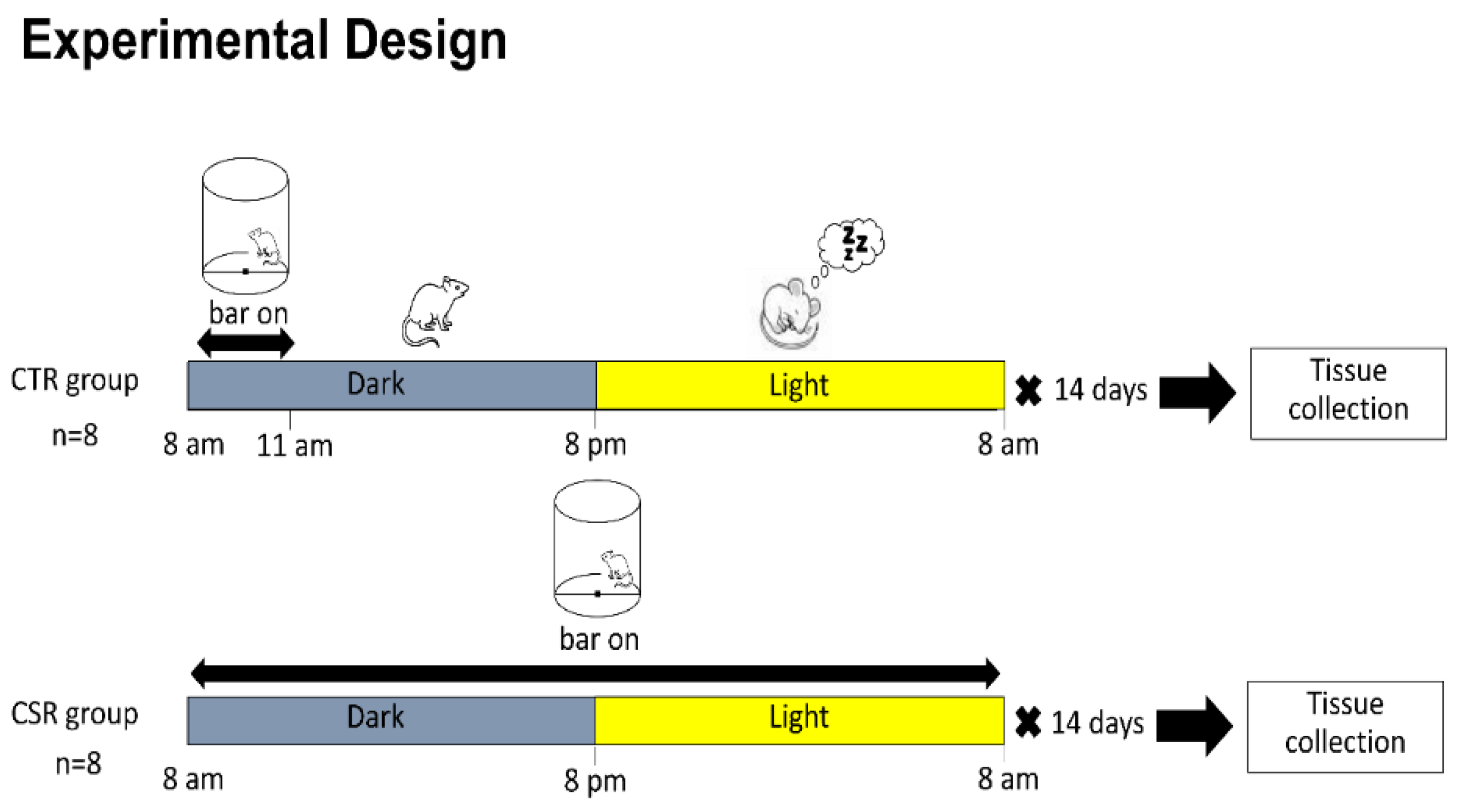
Figure 2.
Effects of chronic sleep restriction (CSR) on mRNA-expression of white adipose tissue (WAT) related genes (A) and body weight (C). Sirtuin-1 (SIRT1); Peroxisome proliferator activated receptor gamma (PPARγ); Peroxisome proliferator activated receptor alpha (PPARα); Sterol regulatory element-binding transcription factor 1/2 (SREBF); Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1); Leptin (LEPT); CCAAT/enhancer-binding protein beta (CEBP-β); Myc (C-Myc); PR domain containing 16 (PRDM16); Uncoupling Protein 1 (UCP1). Gene expression is shown as fold changes relative to the control group. Data are mean ± 95% C.I. (n = 8). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. Clustered gene association network using STRING v11.5 (B). The solid and the dotted lines indicate connection within the same and different cluster, respectively. +/- indicate CSR-induced upregulation/downregulation of selected genes. = indicates no changes in mRNA levels of UCP-1 and PRDM16 genes in CSR mice.
Figure 2.
Effects of chronic sleep restriction (CSR) on mRNA-expression of white adipose tissue (WAT) related genes (A) and body weight (C). Sirtuin-1 (SIRT1); Peroxisome proliferator activated receptor gamma (PPARγ); Peroxisome proliferator activated receptor alpha (PPARα); Sterol regulatory element-binding transcription factor 1/2 (SREBF); Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1); Leptin (LEPT); CCAAT/enhancer-binding protein beta (CEBP-β); Myc (C-Myc); PR domain containing 16 (PRDM16); Uncoupling Protein 1 (UCP1). Gene expression is shown as fold changes relative to the control group. Data are mean ± 95% C.I. (n = 8). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. Clustered gene association network using STRING v11.5 (B). The solid and the dotted lines indicate connection within the same and different cluster, respectively. +/- indicate CSR-induced upregulation/downregulation of selected genes. = indicates no changes in mRNA levels of UCP-1 and PRDM16 genes in CSR mice.
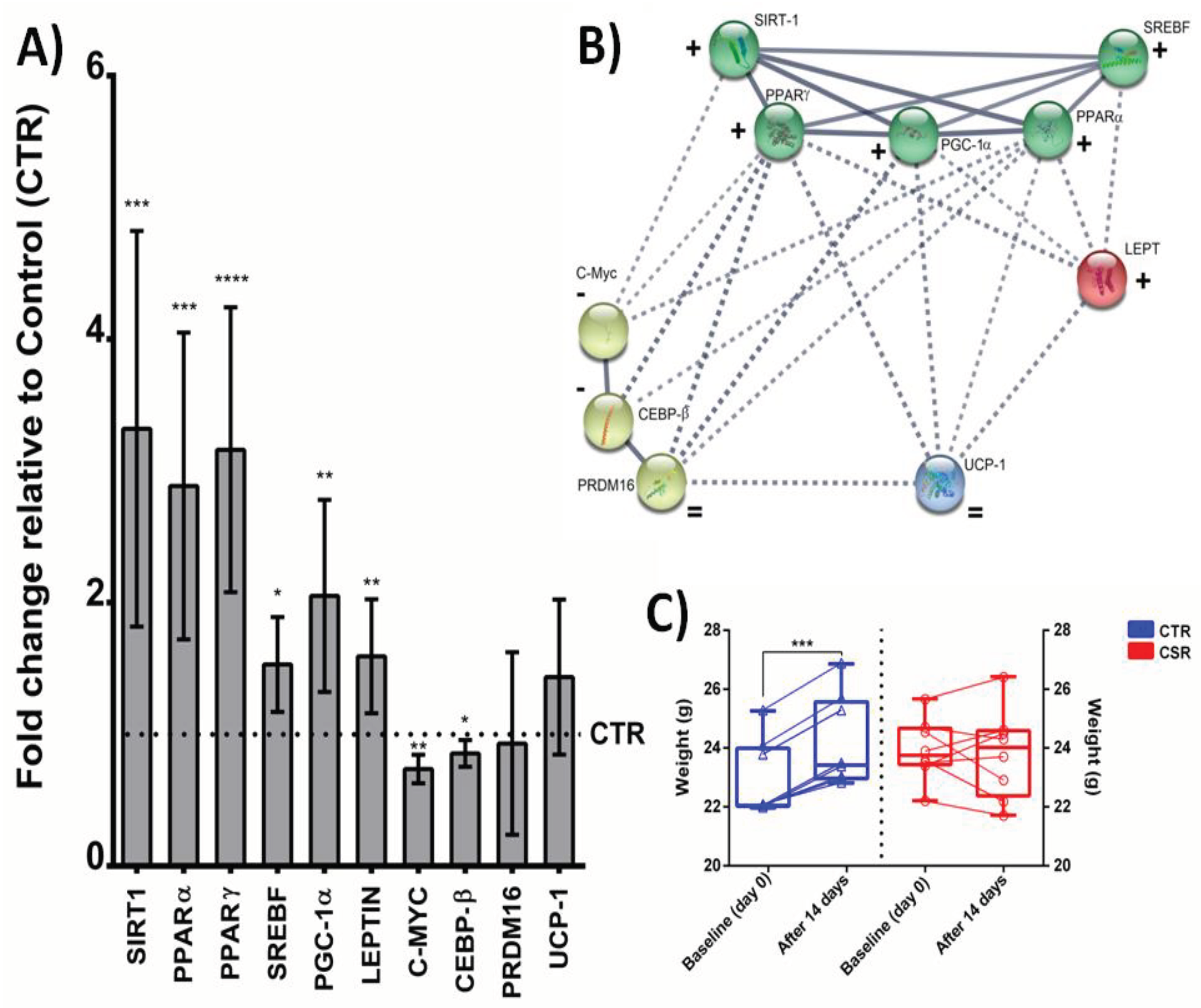

Table 1.
primer sequences (5’ – 3’) used for qPCR analysis.
| Gene | Primer sequence (5’- 3’) | GenBank | Reference |
| PPARγ | ATGGAGCCTAAGTTTGAGTTTGCT GGATGTCCTCGATGGGCTTCA |
XM_006505737.5 | [32] |
| PPARα | CACCCTCTCTCCAGCTTCCA GCCTTGTCCCCACATATTCG |
XM_011245516.4 | [33] |
| SIRT-1 | CGATGACAGAACGTCACACG TCGAGGATCGGTGCCAATCA | NM_019812.3 | [34] |
| LEPT | GACATTTCACACAGGCAGTCG GCAAGCTGGTGAGGATCTGT |
NM_008493.3 | [31] |
| SREBF | GAACAGACACTGGCCGAGAT GAGGCCAGAGAAGCAGAAGAG |
NM_011480.4 | [35] |
| CEBP-β | ACCGGGTTTCGGGACTTGA GTTGCGTAGTCCCGTGTCCA |
X62600.1 | [36] |
| C-Myc | GCCACGTCTCCACACATCAG TGGTGCATTTTCGGTTGTTG |
XM_021216459.1 | [37] |
| PRDM16 | CACGGTGAAGCCATTCATATGCG AGGTTGGAGAACTGCGTGTAGG |
XM_006539171.5 | [34] |
| UCP-1 | GCCATCTGCATGGGATCAAACC TCGTCCCTTTCCAAAGTGTTGAC |
NM_009463.3 | [34] |
| PGC-1α | CTCCAGCCTGACGGCACCC GCAGGGACGTCTTTGTGGCT |
NM_001330751.2 | [38] |
| G6PDH | ATTGACCACTACCTGGGCAA GAGATACACTTCAACACTTTGACCT |
XM_021153829.2 | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



