Submitted:
15 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
Phage therapeutics offer a potentially powerful approach to combat multidrug-resistant bacterial infections. However, to be effective, phage therapy must overcome existing and developing phage resistance. While phage cocktails can reduce this risk by targeting multiple receptors in a single therapeutic, bacteria have mechanisms of resistance beyond receptor modification. A rapidly growing body of knowledge describes a broad and varied arsenal of antiphage systems encoded by bacteria to counter phage infection. We sought to understand the types and frequencies of antiphage systems present in a highly diverse panel of Pseudomonas aeruginosa clinical isolates utilized to characterize novel antibacterials. Using web-server tool PADLOC (Prokaryotic Antiviral Defense Locator), putative antiphage systems were identified in these P. aeruginosa clinical isolates based on sequence homology to a validated and curated catalog of known defense systems. Coupling this host bacterium sequence analysis with host range data for 70 phages, we observed a correlation between existing phage resistance and the presence of higher numbers of antiphage systems in bacterial genomes. We were also able to identify antiphage systems that were more prevalent in highly phage-resistant P. aeruginosa strains, suggesting their importance in conferring the resistance.
Keywords:
Pseudomonas aeruginosa
; phage therapy
; phage resistance
; PADLOC
; antiphage systems
; antiphage systems/phage resistance correlation
1. Introduction
The coevolutionary history of bacteria and bacteriophages is ancient. While the exact origin of viruses is uncertain, a prominent hypothesis argues that viruses evolved from ancient cells before the last universal common ancestor of cellular life [1]. That would mean that the evolutionary history of phages and their bacterial hosts is as old or nearly as old as bacteria themselves, and it is often described as an “arms race.” Bacteria frequently alter or hide phage receptors on the cell surface to evade detection by phages; these are often lipopolysaccharides (LPS) (for gram-negative bacteria), capsule, or various surface localized proteins. In response, selection for mutations in receptor-binding proteins (RBP) enable phages to use modified or alternative receptors [2,3,4]. But this is not the full picture. Bacteria and phages have also developed numerous other active measures to counter phage predation and to subsequently counter resultant antiphage defenses. In recent years, dozens of distinct antiphage systems have been identified and characterized [5,6,7,8,9,10,11,12,13,14,15,16,17], adding to long-known mechanisms that counter phage infection such as restriction-modification and CRISPR-Cas systems [18]. These systems can work synergistically, indicating the importance of a more comprehensive understanding of bacterial antiphage strategies and how they work together. Given the abundance and diversity of phages and their hosts, a complete picture is only likely to emerge far in the future. Phage defense genes are frequently grouped into “islands” in bacterial genomes, and sequence analysis has found uncharacterized adjacent genes are often co-localized with these islands, suggesting that many additional antiphage genes are yet to be discovered [19]. Beyond distinct, dedicated antiphage enzymes, bacteria also employ broader strategies including nutrient depletion or production of small molecules with antiphage properties [20,21,22,23,24]. Taken together, the current body of knowledge provides a preliminary picture of what is likely a profoundly vast and varied array of antiphage mechanisms that can work synergistically to inhibit or block phage predation.
Conversely, phages have coevolved to counter bacterial mechanisms that reduce or block infection. In addition to RBP modification to adsorb to the cell surface to facilitate infection [25], phages are also known to alter CRISPR protospacer adjacent motif (PAM) sequences, restriction recognition sites, or other targets of antiphage systems [26,27]. More recently, it has become clear that phages are actively countering antiphage systems, producing enzymes that neutralize bacterial antiphage enzymes in a variety of ways [28]. The discovery of these phage anti-defense (“anti-antiphage”) enzymes is rapidly emerging with improved sequencing and analysis of phage genes. Efforts to identify phage anti-defense systems are made difficult by the large proportion of phage genomes that remains uncharacterized, and of genes encoding products of unknown function. However, recent work to compile known antiphage systems into catalogs is improving efforts at discovery [29]. It is expected that more anti-defense mechanisms will be discovered with the current intensity of phage genomic and genetic studies.
Phages have been proposed for use in both therapeutic and industrial contexts, starting almost immediately after their discovery [30,31,32]. Successful application of phages in human compassionate treatment cases has been widely reported, and there are ongoing and future clinical trials on phage therapy against drug-resistant infections [33,34,35]. While early efforts show promise, the development of effective, durable phage therapeutics will require the reduction of the emergence of phage resistance during treatment. So far, efforts have focused on using multiple phages in a cocktail that target different surface receptors [36,37], because multiple receptors have to be altered to overcome the activity of the cocktail. Also, phage-resistant mutants that arise under the pressure of multiple phages using different receptors tend to be less fit or viable because mutations removing or modifying cell surface molecules often come at a fitness cost for the bacterial cell, and altering two different receptors at once can have a much steeper cost.
However, focusing solely on surface receptors ignores the growing picture that bacteria not only modify surface elements to prevent adsorption and entry, but also employ a wide array of antiphage strategies inside the cell. Antiphage systems are often grouped in “defense islands” and carried on mobile genetic elements, which indicates that these islands can move en bloc from bacterium to bacterium via horizontal gene transfer (HGT) [9,38]. The role of these antiphage systems in phage resistance must be addressed in the development of phage therapeutics, especially durable off-the-shelf phage cocktails. The relationship of antiphage systems to phage resistance must be better understood so that phage therapeutics can be developed in a careful and rational manner, to counter the myriad ways that bacterial pathogens become resistant. The natural presence of anti-antiphage systems in phages or their introduction via engineering could potentially be of great benefit in the development of more effective phage therapeutics.
To apply current knowledge of antiphage systems in the development of phage therapeutics, we utilized a web-based search tool known as PADLOC (Prokaryotic Antiviral Defense Locator) [39] for screening a panel of 100 highly diverse clinical isolates of Pseudomonas aeruginosa. The numbers of detected antiphage systems correlated with susceptibility data for 70 phages: more phage-resistant strains carried significantly more antiphage systems. Some correlation was also found between phage resistance and the prevalence of certain antiphage systems.
2. Results
2.1. Identification of antiphage systems in the genomes of 100 diverse P. aeruginosa clinical isolates
We analyzed genome sequences from 100 highly diverse clinical isolates of P. aeruginosa. This diversity panel was selected based on core-genome multilocus sequence typing (MLST) from a large repository of multidrug-resistant isolates collected from across the U.S. Military Health System [40]. These isolates originated from multiple hospitals in the United States, as well as from Guam and Afghanistan, and include 91 distinct sequence types (STs). Genome sequence files deposited in the database at the National Center for Biotechnology Information (NCBI) were uploaded and analyzed using the PADLOC webserver tool [39].
Multiple antiphage systems were found in the P. aeruginosa strain panel, predicted based on sequence homology to a catalog of known defense systems. A total of 75 distinct antiphage systems were identified in the 100 strains (Figure 1). The number of systems reported does not reflect distinct subtypes [CBASS type I, II, III, other], where there are multiple subtypes for many of the defense systems. Additionally, putative, non-experimentally validated systems identified through “guilt-by-embedding” were identified in the diversity panel with the recent release of an updated catalog, PADLOC-DB v2.0.0 [41]. A total of 44 such phage defense candidate (PDC) systems were identified after PADLOC analysis (Figure 2). While not all these PDC genes may ultimately be confirmed as such, these results indicate areas for future discovery and the potential breadth of undiscovered antiphage systems in this bacterium.
Antiphage systems were not evenly distributed throughout the strains; many were found only in a small number of isolates, perhaps suggesting relatively recent acquisition of these defense systems by P. aeruginosa strains via HGT. The average defense system was present in 13.25 genomes but ranged from 97 strains (PD-T4-6) to 1 strain (multiple systems). All but four of the identified systems were present in fewer than half of the diversity panel strains (Figure 1). Four antiphage systems were identified in 50 or more 100 strains: PD-T4-6, DMS-other, RM Type I, and SoFic. Thirty-seven systems were present in five or fewer strains. However, DMS-other is a catch-all grouping that reflects potential incomplete antiphage systems, new subtypes, or novel systems with known protein domains, thus it should not be viewed as a distinct, broadly conserved system. Since the diversity panel represents 91 different STs that were collected in different geographic locations over multiple years (Table S1), the strains are likely to have acquired distinct sets of antiphage systems via HGT. One example to note is the distribution of PsyrTA, a toxin-antitoxin system that provides phage protection via abortive infection. PsyrTA was present in 11 strains which belong to four STs, 235, 3002, 3043, and 2387.
The predicted antiphage systems provide defense in a variety of manners, including targeting phage nucleic acid invasion through its degradation, termination of infection by several different mechanisms (abortive lytic cycle), modification of phage DNA and RNA via a variety of epigenetic marks or incorporation of unusual bases, protein modification or degradation, production of RNA molecules that function in an RNAi-like manner, depletion of nutrients and cofactors, or physiological changes to the cell, among others that remain to be characterized. Some systems may have multiple mechanisms of action as early investigation into the Zorya defense system suggests [17]. Predicted diverse nucleases and abortive infection systems were the most prevalent among the 100-strain panel (Table 1). The PDC systems represent an additional group of putative antiphage systems for which characterization remains to be done. Some of these systems are reported to share sequence homology with other characterized systems. For example, PDC-S14 shares some overlap with GAO_29, a defense system that appears to act via a restriction-like mechanism [19].
Figure 1.
Antiphage systems identified in 100-strain diversity panel of P. aeruginosa and the number of strains encoding each system.
Figure 1.
Antiphage systems identified in 100-strain diversity panel of P. aeruginosa and the number of strains encoding each system.
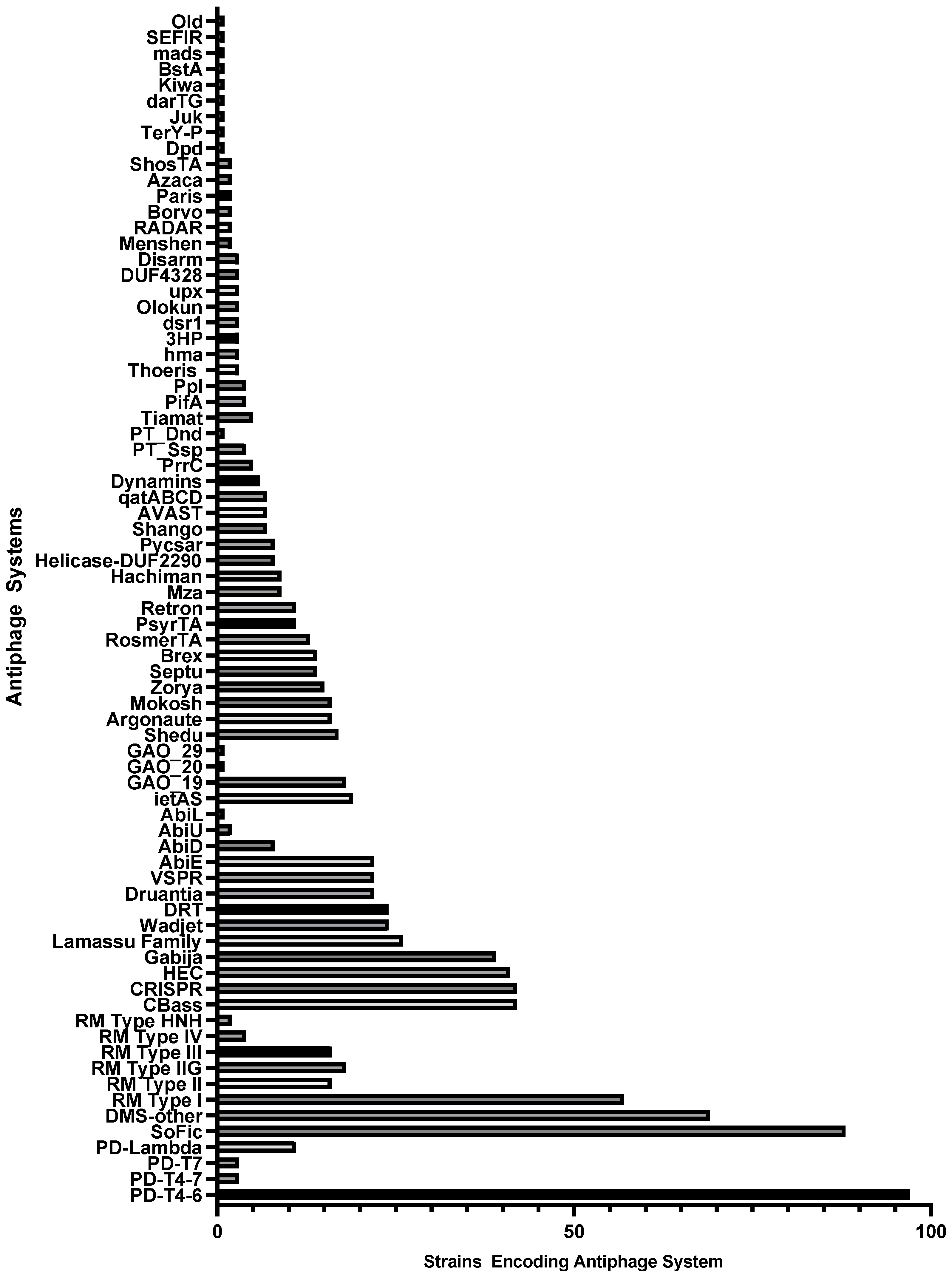
Figure 2.
Phage defense candidates (PDCs) identified by a “guilt-by-embedding” approach and the number of strains from the diversity panel encoding each system.
Figure 2.
Phage defense candidates (PDCs) identified by a “guilt-by-embedding” approach and the number of strains from the diversity panel encoding each system.


Table 1.
Experimentally validated or predicted mechanism of antiphage activity for identified systems in the diversity panel.
Table 1.
Experimentally validated or predicted mechanism of antiphage activity for identified systems in the diversity panel.
| Mechanism of Antiphage Action | System | Number of Strains Encoding | References |
|---|---|---|---|
| Nuclease Activity – Nucleic Acid Degradation | R-M systems | 67 | [18] |
| CRISPR-cas systems* | 42 | [42] | |
| Gabija | 39 | [17] | |
| Wadjet | 24 | [17,43] | |
| Druantia | 22 | [14,17] | |
| Shedu | 17 | [44] | |
| Mokosh | 16 | [7] | |
| Zorya* | 15 | [17] | |
| Septu | 14 | [14,17] | |
| mza | 9 | [19] | |
| qatABCD | 7 | [14,19] | |
| Ppl | 4 | [45] | |
| Olokun | 4 | [7] | |
| DISARM | 3 | [46] | |
| Upx | 3 | [19] | |
| Menshen | 2 | [7,47] | |
| Azaca | 2 | [7] | |
| Kiwa | 1 | [17,48] | |
| Old | 1 | [8] | |
| Abortive Infection or Cell Dormancy | PD Systems (T4-6, T4-7, T7, λ) | 100 | [10] |
| Abi systems (E, D, U, L) | 30 | [49,50,51,52] | |
| GAO_19 | 18 | [19] | |
| CBASS | 42 | [53] | |
| Lamassu | 26 | [7] | |
| ietAS | 19 | [19,26] | |
| RosmerTA | 13 | [7,54] | |
| PsyrTA | 11 | [7,55] | |
| Hachiman | 9 | [17,26,56] | |
| Helicase-DUF2290 | 8 | [8] | |
| Pycsar | 8 | [57] | |
| AVAST | 7 | [19,58] | |
| PrrC | 5 | [59,60] | |
| PifA | 4 | [61] | |
| Paris | 2 | [8] | |
| ShosTA | 2 | [7] | |
| darTG | 1 | [55,62] | |
| BstA | 1 | [63] | |
| Nucleic Acids Modification/Reverse Transcriptases/Expression Modification | DRT | 24 | [19] |
| Argonaute | 16 | [64,65,66] | |
| BREX | 14 | [67] | |
| Retrons | 11 | [68,69] | |
| PT_Ssp and PT_Dnd | 5 | [70] | |
| RADAR | 2 | [19] | |
| Dpd | 1 | [71] | |
| MADS | 1 | [72] | |
| Protein Modification | SoFic | 88 | [7] |
| Borvo | 2 | [7] | |
| TerY-P | 1 | [19] | |
| Other | Dynamins (Lysis Delayance) | 6 | [73] |
| Thoeris (NAD+ depletion) | 3 | [74] | |
| Dsr (NAD+ depletion) | 3 | [21] | |
| SEFIR (NAD+ depletion) | 1 | [7] | |
| Unknown | DMS other | 69 | [39] |
| HEC (Hma embedded candidates) | 41 | [39] | |
| Shango | 7 | [7] | |
| Tiamat | 5 | [7] | |
| Hma | 3 | [56] | |
| 3HP | 3 | [8] | |
| DUF4238 | 3 | [8] | |
| Juk | 1 | [75] |
Antiphage systems can be grouped by general mechanism into several types. For instance, 19 different major validated nucleic acid degradation or uncharacterized nuclease domain-containing systems were identified, not including subtypes. Eighteen types and multiple subtypes of abortive infection systems were identified. Other antiphage strategies included nucleic acid modification, formation of non-coding RNAs, or epigenetic modification of invading phage nucleic acids, modification of proteins or altered protein activity, nutrient and cofactor depletion, or retardation of lysis. Many of these systems have only been recently identified, so more thorough characterization is still required. Some systems may display multiple mechanisms of action or have context-dependent mechanisms of action. The breadth of antiphage mechanisms demonstrates the numerous, overlapping, and complimentary approaches that bacteria utilize to counter phage infection.
2.2. Phage susceptibility of the diverse P. aeruginosa strains
We determined the susceptibility of the 100 strains of P. aeruginosa to 70 phages that belong to 14 genera, Pbunavirus, Nankokuvirus, Pakpunavirus, Phikzvirus, Yuavirus, Septimatrevirus, Epaquintavirus, Phikmvvirus, Pifdecavirus, Bruynoghevirus, Kochitakasuvirus, Litunavirus, Warsawvirus, and Hollowayvirus (Table S2). Two P. aeruginosa strains, MRSN 3587 and MRSN 8141, replaced previous members of the diversity panel after host range data was collected for some phage-host pairs. Consequently, for those two strains phage-host interaction data is available for only 44 phages. Plaque formation was used to determine the overall phage susceptibility percentage for each strain. Strains from the panel were susceptible to phages over a range of 0%-90.0% (Figure 3a). The number of antiphage systems encoded by each strain was plotted against the phage susceptibility percentage to address whether presence of more antiphage systems is correlated with higher phage resistance. It was observed that as strains encoded more antiphage systems, their resistance to phages increased (Figure 3a). The relationship between phage resistance and the presence of antiphage systems was statistically significant (p < 0.0001). The trend was weakly correlated with an R squared value of 0.2620, probably due to the role of other elements that mediate phage host tropism which includes surface receptors and RBPs, presence or absence of prophages, and other physiological traits of the host strains. The diversity panel represents 91 STs of P. aeruginosa. The structure of LPS, type IV pilus and other common phage receptors [77] has not yet been determined for the 100 strains used in this study. However, one can expect that the bacterial panel posseses diverse phage receptors, since analysis of P. aeruginosa isolates collected in the United States possessed extensive diversity in O-antigen serotypes [78]. Also, the receptors for some of the phages used in this sudy are yet to be identified. These host-phage interaction factors were not considered in this analysis, which was focused solely on the presence of antiphage systems and phage susceptibility.
There were clear outlier strains which might make for interesting followup analysis looking at the presence of prophages, unusual type of LPS or other surface structures, or other elements. Of interest are strains like MRSN 317, which possesses only six antiphage systems and yet is lysed by only 15% of the phages tested (Tables S3 and S4). Other strains that exhibit very low susceptibility to the phages but have relatively few identified antiphage systems include MRSN 6220, MRSN 6678, and MRSN 13488. These are among the 10 most phage-resistant strains in the panel (<3% phages show productive infection) and yet only possess 12, 13, or 9 antiphage systems, respectively. Outliers that were more phage-susceptible even with many predicted antiphage systems included strain MRSN 11976 and MRSN 1899. These strains possess 26 and 18 predicted antiphage systems, respectively; MRSN 11976 and MRSN 1899 are lysed by 63% and 68% of the phages tested, respectively (Tables S3 and S4). The average number of antiphage systems possessed by the most resistant strains was 19.2 (Table 2). That means that these highly phage-susceptible strains had the number of defense systems average for the most resistant strains or even higher.
Figure 3.
(a) Susceptibility of P. aeruginosa strains to a panel of phages plotted against the number of antiphage systems identified in each strain via PADLOC analysis. (b) Average efficiency of plating for a panel of phages plotted agianst the number of antiphage systems.
Figure 3.
(a) Susceptibility of P. aeruginosa strains to a panel of phages plotted against the number of antiphage systems identified in each strain via PADLOC analysis. (b) Average efficiency of plating for a panel of phages plotted agianst the number of antiphage systems.
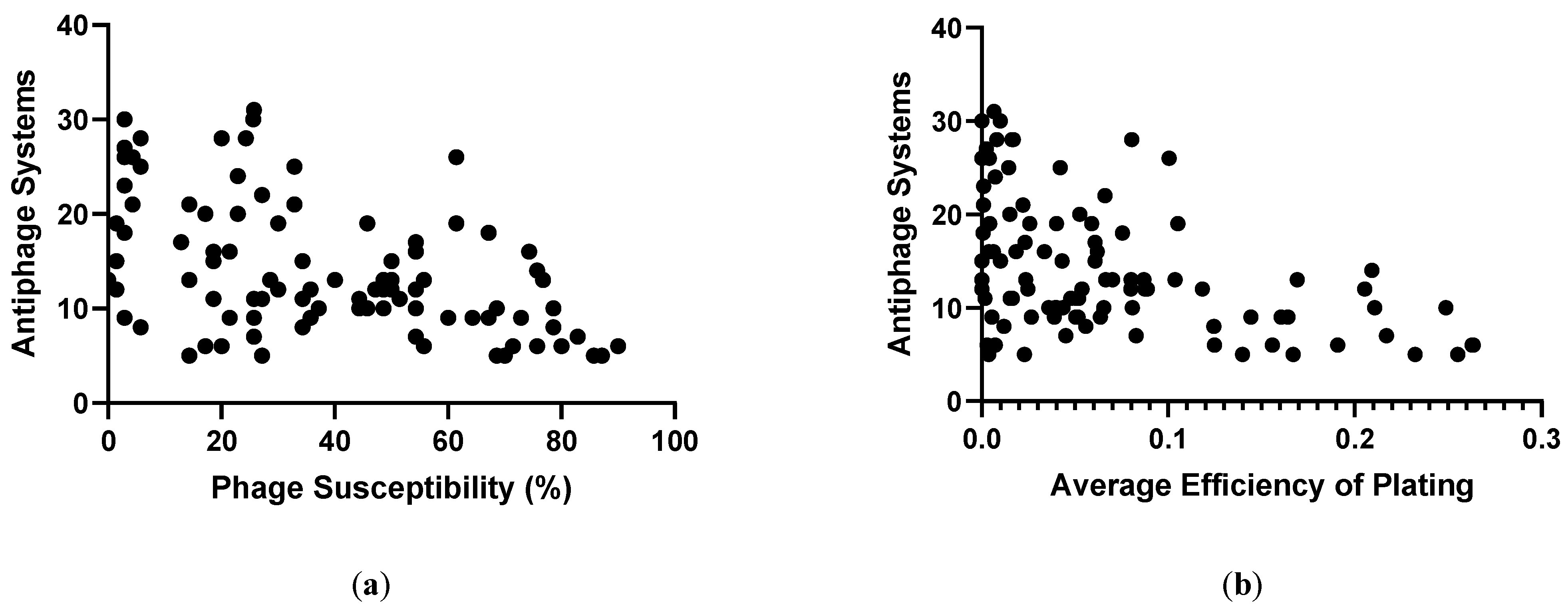
Antiphage systems may not fully prevent productive lysis, but instead reduce the efficiency of plating (EOP), so the impact of antiphage systems on efficiency of plating was assessed (Figure 3b). Phage titers were normalized to the highest value for that phage on the 100-strain panel. For any strain, all EOP values of the 70 phages were then averaged. This yielded an average EOP ranging from 0 (MRSN 6678) to 0.26 (MRSN 11281) (Table S5). The results showed similar trends to those observed for the presence or absence of plaque formation. The presence of higher numbers of antiphage systems was correlated with lower average EOP of phages on that strain (Figure 3b). This trend was stastically significant (p < 0.0001) but with a weak correlation (R squared = 0.2058). The relationship of antiphage systems with average EOP and phage susceptibility together was then analyzed. A general trend was observed that as strains had more antiphage systems, both phage susceptibility and EOP were lower (Figure S1). Along with the correlation of antiphage systems prevalence with , there were outliers which may represent interesting followup with investigation into receptor structure, presence or absence of prophages, or assessment of the activity of predicted antiphage systems. For example, MRSN 435288 was predicted to encode only five antiphage sytems, yet the 70 phages plated on this strain have a very low average EOP of 0.0037 (Table S5).
We hypothesized that certain antiphage systems could be particuarly important in driving phage resistance. Then it was determined if some antiphage systems are more prevalent in phage-resistant strains, and if some are relatively rare. The median point in phage susceptibility was approximately 34%. The 100 strains were divided into four separate groups: most resistant (lysed by <3%); intermediate resistant (lysed by 3-35%); intermediate susceptible (lysed by 35-75.5%); and most susceptible (lysed by >75.5%) (Figure S2). The most resistant and intermediate resistant strains possessed an average of 19.2 and 16.8 antiphage systems per strain, repsectively. Conversely, intermediate susceptible and most susceptible strains had 11.9 and 7.6 antiphage systems per strain (Table 2).
Table 2.
Average number of antiphage systems present per strain in different phage susceptibility groups.
Table 2.
Average number of antiphage systems present per strain in different phage susceptibility groups.
| Phage Susceptibility Group | Average Antiphage Systems/Strain |
|---|---|
| Most resistant 10% | 19.2 |
| Intermediate resistant | 16.8 |
| Intermediate susceptible | 11.9 |
| Most susceptible 10% | 7.6 |
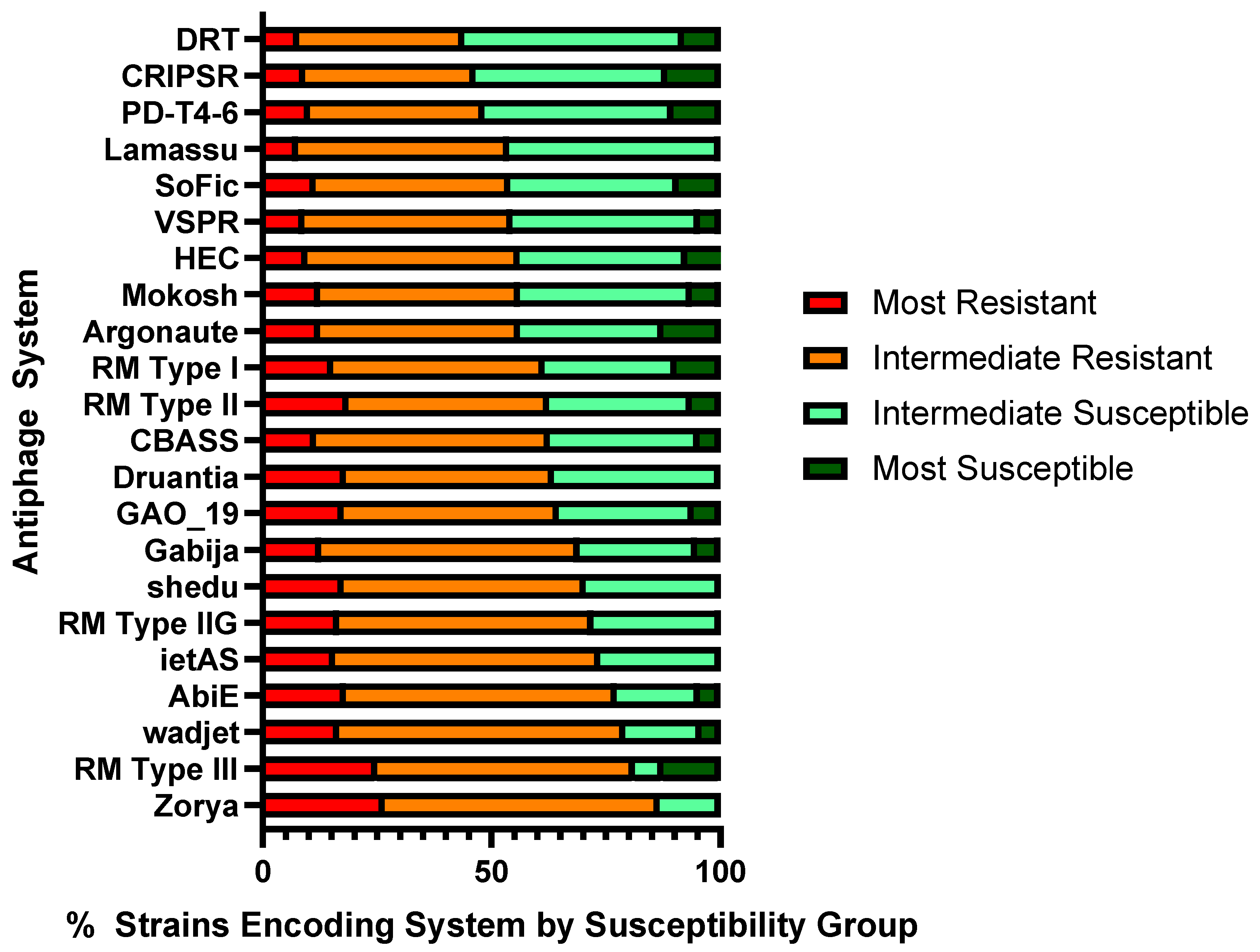
Altogether, 22 validated antiphage systems and three PDC systems were present in 15 or more of the P. aeruginosa strains. An antiphage system was considered prevalent in a phage-resistant strain if it represented 52.5% or greater of the total strains in the panel encoding that particular system. Of the 22 validated systems, 19 were prevalent in the phage-resistant strains (Figure 4). Prevalence ranged from 86.7% phage-resistance in the strains encoding Zorya (n = 15) to 53.8% resistance in the strains encoding Lamassu Family systems (n = 26). One of the three remaining systems, PD-T4-6, was intermediate with no prevalence in resistant or susceptible strains (Table S6). Two other systems, CRISPR-Cas with all Cas types and DRT of all types, were prevalent in susceptible strains, with 53.5% of the 43 strains encoding CRSIPR-Cas systems and 56% of the 25 strains encoding DRT in the susceptible groups.
Other defense systems were also prevalent in resistant strains but were less represented within the 100-strain panel. These included RosmerTA (69.2% in resistant strains), PsyrTA (81.8% in resistant strains), and retrons (54.6% in resistant strains) (Table S6). Some antiphage systems showed no prevalence in resistant or susceptible strains, including BREX and Septu, which showed a 50/50 split (both systems are predicted to be encoded by 14 strains). A few rare systems showed relative prevalence in susceptible strains, including 10 strains encoding PD-Lambda systems (60% of them were phage-susceptible), seven strains encoding AVAST (71% of them were susceptible), and nine strains encoding mza (56% of them were susceptible) (see Table S6).
The three PDC systems found in 15 or more strains were prevalent in resistant strains. PDC-S02 was predicted in 39 strains, of which 59.0% were resistant. PDC-S06 was predicted in 38 strains, of which 74.4% were resistant. Lastly, PDC-M30 was predicted in 32 strains, of which 68.8% were resistant. Other more rarely occurring PDC systems were found at a higher frequency in resistant or susceptible groups. For example, PDC-S09 and PDC-S11 showed strong prevalence in resistant strains, with 82% and 90% of 11 and 10 strains encoding them, respectively. One system, PDC-S08, showed an even 50/50 split, and PDC-S14 showed spread in susceptible strains (57% of 14 strains; Table S6).
3. Discussion
This study provides a snapshot of distribution of the currently identifiable antiphage systems in a highly diverse panel of 100 P. aeruginosa strains, and of how the presence of these antiphage systems relates to susceptibility to a collection of 70 phages that includes 14 genera, comprising myo-, sipho- and podophages. These diverse panels of bacterial strains and phages provide both a significant sample of the defense systems present in P. aeruginosa and an indication of how effective they are against the broader diversity of Pseudomonas phages. Understanding the representation and frequency of different antiphage systems in P. aeruginosa clinical isolates can provide critical information for the development of rationally designed phage therapeutic cocktails. This knowledge can empower the choice of therapeutic phages that counter these defense systems, either by evasion or by production of dedicated anti-defense enzymes [28,29,79]. Coupling this new information with the current standards of phage selection, including safe genomic properties, broad host range, robust lytic and anti-biofilm activity, using different receptors, and confirmation of synergy with other phages and antibiotics can enable the rational design of more effective and durable therapeutic phage cocktails [31]. As phage engineering approaches also continue to improve [80], incorporation of anti-antiphage genes that counter identified antiphage systems into candidate therapeutic phages could improve their efficacy and long-term durability as therapeutics.
An initial avenue of possible utilization of these results is the identification of candidate phages encoding anti-defense genes. Some of these genes have been identified and characterized [29,81]. In this work, we identified some phages that broadly lyse strains encoding certain antiphage systems (i.e. pycsar) for which known anti-defense genes have been identified. For example, phage KEN5 lyses 7/8 strains encoding a pycsar effector, while KEN3 is able to lyse 6/8 of these strains. These strains are genetically diverse and belong to seven different STs, indicating that these phages may be broadly active against more diverse P. aeruginosa strains. While the information on LPS types and RBPs is currently unavailable, it seems clear that if the putative pycsar genes are active, these two phages are somehow unaffected by the system that is prevalent in phage-resistant strains (62.5% of strains with pycsar are resistant) (Tables S6 and S7). This dataset could also provide initial information for identifying novel anti-defense enzymes. Shango has been discovered and characterized in Escherichia coli and P. aeruginosa [5,7], but no phage-encoded anti-Shango enzymes have been identified. A Pbunavirus phage EPa11 can lyse 5/7 strains encoding putative Shango systems (Table S8). If the predicted Shango systems are active in these strains, it suggests that this phage is either somehow unaffected by or can counter this defense system. While these data alone are insufficient to establish that novel anti-defense enzymes are present, these phages are certainly candidates for the discovery of new anti-defense systems. Further discovery of these systems can be leveraged to suppress the emergence of phage resistance in the design of improved phage therapeutics.
In addition, our analysis could provide information on types of phages that are not well targeted by certain defense systems, a phenomenon that has been observed for multiple antiphage systems [19]. With respect to our pycsar example, KEN3 is a podovirus in the genus Bruynoghevirus, while KEN5 is a myovirus in the genus Pakpunavirus (Table S2). Broader analysis of phage-host interaction data could potentially define preferences for antiphage systems related to either taxonomic or structural factors. These are important avenues of future investigation and require further in silico analysis and laboratory work.
The diversity in the types of antiphage systems that were identified in the 100-strain P. aeruginosa diversity panel is remarkable. Most of the strains encoded both an abortive infection strategy and a nucleic acid degradation mechanism. Substantial numbers of strains encoded more unique systems with a variety of mechanisms, including nucleic acid modification and protein modification or degradation. Countering different steps of phage infection with multiple and different antiphage systems likely contributes to phage resistance in a synergistic manner. Such synergy has been observed for many antiphage systems, including those that seem mutually exclusive per comparative genomic analysis [82]. The coupling of nucleic acid-degrading, phage gene expression control, and cell-suicide mechanisms may represent a complementary array of antiphage mechanisms that provide flexibility in speed of response and cost to the bacterial cell [83]. In our analysis, CRISPR-Cas systems were not prevalent in phage-resistant strains. This paradoxical outcome is in agreement with recent data of Lood et al. [81], who observed that strains of P. aeruginosa encoding CRISPR-Cas systems were more susceptible to a panel of 14 phages than strains lacking CRISPR-Cas. More work is clearly needed to understand synergies and antagonisms among antiphage systems.
In the PADLOC analysis used here (based on sequence homology), many antiphage systems that are multi-gene (located in operons) were readily identified. However, there are also single-gene antiphage systems, including SoFic, some PD systems (e.g., PD-T4-6), and others that are more subject to false calls. We have not independently assessed whether these systems are expressed in a given strain and if so, whether they play any role in phage defense. The number of strains, systems, and possibility for redundancy makes such analysis very time- and labor-consuming. This can be even more important to assess if some systems may have roles in the bacterial cell beyond phage defense. For example, wadjet systems have been reported to provide protection from exogenous DNA, including not only phage DNA but also plasmids, transposons, or other mobile genetic elements [43]. Some wadjet systems were shown to be involved in phage defense, but some may only be activated in response to certain signals or phage components. Understanding the context of defense system activation is important. Even growth conditions in the laboratory could impact their expression. For example, in one P. aeruginosa strain, CRISPR-Cas type I-F was under control of a two-component system involved in regulating alginate biosynthesis. Phages hijacked a repressor of this two-component system to silence expression of the CRISPR-Cas genes [84]. While we have an initial picture of antiphage machinery in the genomes of diverse P. aeruginosa strains, whether, how, and when these systems respond to phage infection is yet to be elucidated. As antiphage systems continue to be identified, characterized, and their role in phage-host interactions is established, this picture will continue to become clearer.
To find if there was a correlation between the number of antiphage systems and phage resistance, we first relied on the presence or absence of plaque formation. This provides data if a strain is resistant under laboratory conditions but may lead to an underappreciation of “soft” resistance. Where antiphage systems have been characterized, they frequently reduce efficiency of plating (EOP) without eliminating lytic activity. For example, the PD-T4-6 system identified by Vassallo and colleagues conferred an approximately 4-log reduction in EOP for phage T4, but plaques were still formed [10]. Consequently, we calculated the average EOP for the panel of 70 phages on each of the P. aeruginosa diversity panel strains to provide an approximation of how well phages plate on each strain. EOP was correlated with the number of antiphage systems predicted by the PADLOC analysis and some protection was revealed in the form of reduced EOP. However, numerous factors can influence the EOP, including the presence of primary or secondary phage receptors, characteristics of the strains, or even the presence or absence of certain plasmids [85,86]. To empirically assess whether the presence of a system affects EOP, it would be necessary to conduct comparative EOP analysis with antiphage systems knocked out in isogenic strains. However, given the diversity and number of antiphage systems, strains, and phages, vast resources exceeding our current capacity would be needed to carefully assess synergy in analyzed strains beyond the marker that was selected.
While we found a statistically significant correlation between the number of antiphage systems and phage resistance in P. aeruginosa strains, this trend may not be applicable to all bacterial species. For instance, a recent analysis of E. coli phage-host interactions found no relation between antiphage systems and phage susceptibility [87]. Phages of P. aeruginosa and other gram-negative bacteria often adsorb to LPS, a molecule with complex structure and high diversity, particularly in the O-antigen [77]. There are twenty characterized O-antigen serotypes within P. aeruginosa [88], while E. coli has a much greater diversity, with approximately 180 O-antigen serotypes [89]. The restrictions that higher receptor diversity impose on host range may reduce the need to maintain a diverse arsenal of antiphage systems. For a species with less diversity in common phage receptors, more antiphage systems may be necessary to provide adequate protection against infection to be successful in a particular niche. Despite the lower number of antiphage systems in E. coli, Gaborieau and colleagues were able to identify a weak but statistically significant correlation between antiphage systems and reduced viral infectivity [87]. In species with a greater diversity in phage receptors, antiphage systems may play a more secondary role by reducing the infectious efficacy of the phages that adsorb to the bacterial cell. Consequently, an analysis of antiphage systems in phage-host dynamics should consider all of the elements that drive phage host tropism in a particular bacterium.
4. Materials and Methods
4.1. Bacterial strains used in this study
The 100 P. aeruginosa clinical isolates used in this study were provided by the Multidrug-resistant organism Repository and Surveillance Network (MRSN) which developed this panel to maximize genetic diversity [40].
4.2. Phages used in this work
Phages used in this study represent a diverse group of 50 unique lytic Pseudomonas phages isolated from sewage and environmental waters over almost 10 years at Walter Reed Army Institute of Research (WRAIR), as well as 20 unique phages isolated by WRAIR’s overseas laboratories, in Kenya at the U.S. Army Medical Research Directorate – Africa (KEN phages) and in Thailand at the Armed Forces Research Institute of Medical Sciences (AFRIMS, AFR phages).
4.3. Handling of bacterial cultures and phages
All strains were cultured overnight in Heart Infusion Broth (HIB, BD, Franklin Lakes, NJ, USA) at 37˚C with shaking prior to use for host range testing. All phages were purified from environmental samples following our standard approach [90]. Phages were stored in propagation medium (HIB + 0.1% glucose, 2mM MgCl2, 0.5mM CaCl2) at 4˚C until the use in phage host range assays.
4.4. Analysis of bacterial genomes for identification of antiphage systems
We utilized the PADLOC webserver (https://padloc.otago.ac.nz/padloc/), employing their full catalog of defense systems when collecting search results [39]. This means that antiphage systems not fully characterized or experimentally validated, or reported in pre-prints were included. We wanted to obtain the broadest representation possible of antiphage systems present in the P. aeruginosa diversity panel and thus did not exclude these predicted, putative systems. Genome sequences of the 100 P. aeruginosa panel strains available in the National Center for Biotechnology Information (NCBI) database [40] were used as queries for the webserver search tool, and results were collected following a run with CRISPRdetect analysis. Results were analyzed such that total numbers of strains encoding each system were determined.
4.5. Phage susceptibility testing
Phage susceptibility was assessed using a micro-spot plaque assay [91], where phage dilution series are plated on all 100 strains of the panel and plaque formation monitored. ”Lysis from without” or non-replicative lysis was considered as a negative result (phage resistance).
4.6. Determination of phage susceptibility groups
To provide a breakdown of strain susceptibility to the phages, a division at the median phage susceptibility was made, where 50 strains based on phage susceptibility calculations were considered more resistant and the other 50 strains more susceptible. Within the halves, the 10 most resistant and 10 most susceptible strains were placed in groups. The number of antiphage systems present in each strain within each group was averaged to yield the average number of antiphage systems present within that group.
4.7. Correlation of antiphage systems with phage susceptibility
To address the core question of how antiphage system genome relates to phage susceptibility, a plot comparing these datasets was generated and a XY correlation analysis was performed to assess statistical significance using available statistical analysis tools from GraphPad Prism (version 9.5.1).
4.8. Correlation of antiphage systems with average efficiency of plating
To account for the possibility of partial antiphage activity that does not fully prevent productive lysis, we determined the average EOP for each strain. To do so, each phage’s titer was normalized to the highest titer observed on any of the 100 strains. For phage-strain interactions with no plaque formation or non-productive lysis, these were considered an efficiency of 0. The EOPs for all 70 phages on each strain were averaged together. The calculated average EOP values were plated against the number of antiphage systems and a XY correlation analysis was run to assess statistical significance (GraphPad Prism 9.5.1).
4.9. Assessment of prevalence of antiphage systems in phage susceptibility groups
The distribution of strains encoding each antiphage system in a phage susceptibility group was determined by reviewing the PADLOC results. To determine if a particular system was prevalent in resistant or susceptible strains, the distribution of antiphage systems was assessed across the susceptibility groups (most resistant, intermediate resistant, intermediate susceptible, and most susceptible). If >52.5% of strains encoding a system belonged to resistant or susceptible groups, that antiphage system was considered prevalent in either resistant or susceptible strains.
5. Conclusions
Phages have coevolved with their bacterial hosts for billions of years, and in a very long evolutionary process (that likely preceded the divergence of the bacteria from the rest of life). The host cell has developed various means to avoid being infected with these viruses, as the viruses in turn develop means to overcome or circumvent these defenses. Complex and multi-layered phage-bacteria interactions continue to be unraveled in the current renaissance of phage research. Bacterial resistance to phages represents a major and persistent challenge in the development of efficacious phage therapeutics, as has been experienced in the antibiotic saga. Efforts to subvert or prevent phage resistance in therapeutic applications have largely focused on designing multi-phage cocktails that target multiple receptors with complementary and overlapping component host ranges. These efforts are important and should continue toward developing effective and durable phage therapeutics, but in this endeavor, attention must be paid to addressing bacterial defense strategies. While much remains to be discovered, early analysis is already showing that certain antiphage systems have clear specificities for the types of phages they target [19]. With a better understanding of the types of antiphage mechanisms present within a species of interest, such as P. aeruginosa, rational decisions can be made to select therapeutic phages that are less subject to, or that can evade or counter prevalent antiphage systems. To our knowledge this is the first work relating the in-silico analysis of antiphage system genome content using PADLOC with a broad analysis of susceptibility of a highly diverse panel of host strains to a diverse collection of phages. Our results indicate that the multiple and varied antiphage systems present in the genomes of diverse P. aeruginosa clinical isolates have distinct patterns of association with phage resistance and susceptibility, and thus are likely play an important role in mediating phage resistance. Understanding these interactions is critical for the future development of phage-based antibacterial drugs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Multivariate analysis of 100 P. aeruginosa strains examining relationship between number of predicted antiphage systems with average efficiency of plating and overall phage susceptibility; Figure S2: Box and whiskers plot of 100 strains of P. aeruginosa showing median phage susceptibility and the 10 most extreme strains Table S1: Phages used in this work; Table S2: P. aeruginosa strains used in this work; Table S3: Antiphage systems and phage susceptibility of the 100 diverse P. aeruginosa strains; Table S4: Host ranges of 70 phages against 100 diverse strains of P. aeruginosa. Table S5: Phage efficiency of plating; Table S6: Prevalence of antiphage systems in P. aeruginosa strains of different phage susceptibility groups; Table S7: Host ranges of phages KEN3 and KEN5 and their activity against P. aeruginosa strains encoding predicted pycsar system; and Table S8: Host range of phage EPa11 and its activity against P. aeruginosa strains encoding predicted Shango system.
Author Contributions
Conceptualization, K.A.B. and A.A.F.; methodology, K.A.B., C.D.U, N.M., and A.A.F.; validation, K.A.B., A.A.F., N.M., and M.P.N.; formal analysis, K.A.B.; investigation, K.A.B. and A.A.F.; data curation, K.A.B.; writing—original draft preparation, K.A.B.; writing—review and editing, A.A.F., K.A.B., N.M., and M.P.N; supervision, A.A.F. and M.P.N.; project administration, A.A.F. and M.P.N.; funding acquisition, A.A.F. and M.P.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Congressionally Directed Medical Research Programs, Peer Reviewed Medical Research Program, grant number PR182667 and Military Infectious Diseases Research Program, grant number MI230019.
Acknowledgments
Material has been reviewed by the Walter Reed Army Institute of Research. There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the authors, and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. The Multidrug Resistant Organism Repository and Surveillance Network (MRSN) at the Walter Reed Army Institute of Research provided the P. aeruginosa diversity panel used for this work. Some of the phages used in this study were isolated by the U.S. Army Medical Research Directorate – Africa in Nairobi, Kenya (Dr. Lillian Musila and team) and the Armed Forces Research Institute of Medical Sciences in Bangkok, Thailand (Dr. Paphavee Lertsethtakarn and team). We thank Yunxiu He for her technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nasir, A.; Romero-Severson, E.; Claverie, J.M. Investigating the concept and origin of viruses. Trends Microbiol 2020, 28, 959–967. [Google Scholar] [CrossRef]
- Kortright, K.E.; Done, R.E.; Chan, B.K.; Souza, V.; Turner, P.E. Selection for phage resistance reduces virulence of Shigella flexneri. Appl Environ Microbiol 2022, 88, e0151421. [Google Scholar] [CrossRef]
- Tang, M.; Huang, Z.; Zhang, X.; Kong, J.; Zhou, B.; Han, Y.; Zhang, Y.; Chen, L.; Zhou, T. Phage resistance formation and fitness costs of hypervirulent Klebsiella pneumoniae mediated by K2 capsule-specific phage and the corresponding mechanisms. Front Microbiol 2023, 14, 1156292. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Ahn, J. Evolutionary dynamics between phages and bacteria as a possible approach for designing effective phage therapies against antibiotic-resistant bacteria. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Laderman, E.; Huiting, E.; Zhang, C.; Davidson, A.; Bondy-Denomy, J. Core defense hotspots within Pseudomonas aeruginosa are a consistent and rich source of anti-phage defense systems. Nucleic Acids Res 2023, 51, 4995–5005. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, H.; Tong, Y. Unveil the Secret of the bacteria and phage arms race. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Millman, A.; Melamed, S.; Leavitt, A.; Doron, S.; Bernheim, A.; Hor, J.; Garb, J.; Bechon, N.; Brandis, A.; Lopatina, A.; et al. An expanded arsenal of immune systems that protect bacteria from phages. Cell Host Microbe 2022, 30, 1556–1569. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; Depardieu, F.; Miele, S.; Dowding, J.; Laval, A.L.; Lieberman, E.; Garry, D.; Rocha, E.P.C.; Bernheim, A.; Bikard, D. Phages and their satellites encode hotspots of antiviral systems. Cell Host Microbe 2022, 30, 740–753. [Google Scholar] [CrossRef]
- Georjon, H.; Bernheim, A. The highly diverse antiphage defence systems of bacteria. Nat Rev Microbiol 2023, 21, 686–700. [Google Scholar] [CrossRef]
- Vassallo, C.N.; Doering, C.R.; Littlehale, M.L.; Teodoro, G.I.C.; Laub, M.T. A functional selection reveals previously undetected anti-phage defence systems in the E. coli pangenome. Nat Microbiol 2022, 7, 1568–1579. [Google Scholar] [CrossRef]
- Macdonald, E.; Wright, R.; Connolly, J.P.R.; Strahl, H.; Brockhurst, M.; van Houte, S.; Blower, T.R.; Palmer, T.; Mariano, G. The novel anti-phage system Shield co-opts an RmuC domain to mediate phage defense across Pseudomonas species. PLoS Genet 2023, 19, e1010784. [Google Scholar] [CrossRef] [PubMed]
- Akritidou, K.; Thurtle-Schmidt, B.H. OLD family nuclease function across diverse anti-phage defense systems. Front Microbiol 2023, 14, 1268820. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, L.; Song, S.; Kirigo, J.; Battisti, M.E.; Huelgas-Méndez, D.; García-Contreras, R.; Petersen, M.E.; Tomás, M.; Wood, T.K. Toxin/antitoxin systems induce persistence and work in concert with restriction/modification systems to inhibit phage. bioRxiv 2023, 2023.2002.2025.529695. [Google Scholar] [CrossRef]
- Wang, S.; Sun, E.; Liu, Y.; Yin, B.; Zhang, X.; Li, M.; Huang, Q.; Tan, C.; Qian, P.; Rao, V.B.; et al. Landscape of new nuclease-containing antiphage systems in Escherichia coli and the counterdefense roles of bacteriophage T4 genome modifications. J Virol 2023, 97, e0059923. [Google Scholar] [CrossRef]
- Wein, T.; Johnson, A.G.; Millman, A.; Lange, K.; Yirmiya, E.; Hadary, R.; Garb, J.; Steinruecke, F.; Hill, A.B.; Kranzusch, P.J.; et al. CARD-like domains mediate anti-phage defense in bacterial gasdermin systems. bioRxiv 2023, 2023.2005.2028.542683. [Google Scholar] [CrossRef]
- Bravo, J.P.K.; Aparicio-Maldonado, C.; Nobrega, F.L.; Brouns, S.J.J.; Taylor, D.W. Structural basis for broad anti-phage immunity by DISARM. Nat Commun 2022, 13, 2987. [Google Scholar] [CrossRef]
- Doron, S.; Melamed, S.; Ofir, G.; Leavitt, A.; Lopatina, A.; Keren, M.; Amitai, G.; Sorek, R. Systematic discovery of antiphage defense systems in the microbial pangenome. Science 2018, 359. [Google Scholar] [CrossRef]
- Dupuis, M.E.; Villion, M.; Magadan, A.H.; Moineau, S. CRISPR-Cas and restriction-modification systems are compatible and increase phage resistance. Nat Commun 2013, 4, 2087. [Google Scholar] [CrossRef]
- Gao, L.; Altae-Tran, H.; Bohning, F.; Makarova, K.S.; Segel, M.; Schmid-Burgk, J.L.; Koob, J.; Wolf, Y.I.; Koonin, E.V.; Zhang, F. Diverse enzymatic activities mediate antiviral immunity in prokaryotes. Science 2020, 369, 1077–1084. [Google Scholar] [CrossRef]
- Tal, N.; Millman, A.; Stokar-Avihail, A.; Fedorenko, T.; Leavitt, A.; Melamed, S.; Yirmiya, E.; Avraham, C.; Brandis, A.; Mehlman, T.; et al. Bacteria deplete deoxynucleotides to defend against bacteriophage infection. Nat Microbiol 2022, 7, 1200–1209. [Google Scholar] [CrossRef]
- Garb, J.; Lopatina, A.; Bernheim, A.; Zaremba, M.; Siksnys, V.; Melamed, S.; Leavitt, A.; Millman, A.; Amitai, G.; Sorek, R. Multiple phage resistance systems inhibit infection via SIR2-dependent NAD(+) depletion. Nat Microbiol 2022, 7, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Kronheim, S.; Daniel-Ivad, M.; Duan, Z.; Hwang, S.; Wong, A.I.; Mantel, I.; Nodwell, J.R.; Maxwell, K.L. A chemical defence against phage infection. Nature 2018, 564, 283–286. [Google Scholar] [CrossRef]
- Bernheim, A.; Millman, A.; Ofir, G.; Meitav, G.; Avraham, C.; Shomar, H.; Rosenberg, M.M.; Tal, N.; Melamed, S.; Amitai, G.; et al. Prokaryotic viperins produce diverse antiviral molecules. Nature 2021, 589, 120–124. [Google Scholar] [CrossRef]
- Hardy, A.; Kever, L.; Frunzke, J. Antiphage small molecules produced by bacteria - beyond protein-mediated defenses. Trends Microbiol 2023, 31, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Habusha, M.; Tzipilevich, E.; Fiyaksel, O.; Ben-Yehuda, S. A mutant bacteriophage evolved to infect resistant bacteria gained a broader host range. Mol Microbiol 2019, 111, 1463–1475. [Google Scholar] [CrossRef]
- Stokar-Avihail, A.; Fedorenko, T.; Hor, J.; Garb, J.; Leavitt, A.; Millman, A.; Shulman, G.; Wojtania, N.; Melamed, S.; Amitai, G.; et al. Discovery of phage determinants that confer sensitivity to bacterial immune systems. Cell 2023, 186, 1863–1876. [Google Scholar] [CrossRef]
- Watson, B.N.J.; Easingwood, R.A.; Tong, B.; Wolf, M.; Salmond, G.P.C.; Staals, R.H.J.; Bostina, M.; Fineran, P.C. Different genetic and morphological outcomes for phages targeted by single or multiple CRISPR-Cas spacers. Philos Trans R Soc Lond B Biol Sci 2019, 374, 20180090. [Google Scholar] [CrossRef]
- Gao, Z.; Feng, Y. Bacteriophage strategies for overcoming host antiviral immunity. Front Microbiol 2023, 14, 1211793. [Google Scholar] [CrossRef]
- Yan, Y.; Zheng, J.; Zhang, X.; Yin, Y. dbAPIS: a database of anti-prokaryotic immune system genes. Nucleic Acids Res 2023. [Google Scholar] [CrossRef]
- Dublanchet, A.; Bourne, S. The epic of phage therapy. Can J Infect Dis Med Microbiol 2007, 18, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Nikolich, M.P.; Filippov, A.A. Bacteriophage therapy: developments and directions. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Polaska, M.; Sokolowska, B. Bacteriophages-a new hope or a huge problem in the food industry. AIMS Microbiol 2019, 5, 324–346. [Google Scholar] [CrossRef]
- McCallin, S.; Sacher, J.C.; Zheng, J.; Chan, B.K. Current state of compassionate phage therapy. Viruses 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.M.; Sagona, A.P. Armed phages are heading for clinical trials. Nat Microbiol 2023, 8, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, N.M.; Devequi Gomes Nunes, D.; Shiach, J.; Valeria Saraiva Hodel, K.; Dantas Viana Barbosa, J.; Alencar Pereira Rodrigues, L.; Coler, B.S.; Botelho Pereira Soares, M.; Badaro, R. Current clinical landscape and global potential of bacteriophage therapy. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage cocktail development for bacteriophage therapy: toward improving spectrum of activity breadth and depth. Pharmaceuticals (Basel) 2021, 14, 1019. [Google Scholar] [CrossRef]
- Li, C.; Shi, T.; Sun, Y.; Zhang, Y. A novel method to create efficient phage cocktails via use of phage-resistant bacteria. Appl Environ Microbiol 2022, 88, e0232321. [Google Scholar] [CrossRef]
- Hochhauser, D.; Millman, A.; Sorek, R. The defense island repertoire of the Escherichia coli pan-genome. PLoS Genet 2023, 19, e1010694. [Google Scholar] [CrossRef]
- Payne, L.J.; Meaden, S.; Mestre, M.R.; Palmer, C.; Toro, N.; Fineran, P.C.; Jackson, S.A. PADLOC: a web server for the identification of antiviral defence systems in microbial genomes. Nucleic Acids Res 2022, 50, W541–W550. [Google Scholar] [CrossRef]
- Lebreton, F.; Snesrud, E.; Hall, L.; Mills, E.; Galac, M.; Stam, J.; Ong, A.; Maybank, R.; Kwak, Y.I.; Johnson, S.; et al. A panel of diverse Pseudomonas aeruginosa clinical isolates for research and development. JAC Antimicrob Resist 2021, 3, dlab179. [Google Scholar] [CrossRef]
- Payne, L. PADLOC-DB v2.0.0. 2023, Available online: https://github.com/padlocbio/padloc-db/releases.
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants. Nat Rev Microbiol 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Deep, A.; Gu, Y.; Gao, Y.Q.; Ego, K.M.; Herzik, M.A., Jr.; Zhou, H.; Corbett, K.D. The SMC-family Wadjet complex protects bacteria from plasmid transformation by recognition and cleavage of closed-circular DNA. Mol Cell 2022, 82, 4145–4159. [Google Scholar] [CrossRef] [PubMed]
- Loeff, L.; Walter, A.; Rosalen, G.T.; Jinek, M. DNA end sensing and cleavage by the Shedu anti-phage defense system. bioRxiv 2023, 2023.2008.2010.552762. [Google Scholar] [CrossRef]
- Wang, S.; Sun, E.; Liu, Y.; Yin, B.; Zhang, X.; Li, M.; Huang, Q.; Tan, C.; Qian, P.; Rao, V.B.; et al. The complex roles of genomic DNA modifications of bacteriophage T4 in resistance to nuclease-based defense systems of E. coli. bioRxiv 2022, 2022.2006.2016.496414. [Google Scholar] [CrossRef]
- Ofir, G.; Melamed, S.; Sberro, H.; Mukamel, Z.; Silverman, S.; Yaakov, G.; Doron, S.; Sorek, R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat Microbiol 2018, 3, 90–98. [Google Scholar] [CrossRef]
- Mariano, G.; Blower, T.R. Conserved domains can be found across distinct phage defence systems. Mol Microbiol 2023, 120, 45–53. [Google Scholar] [CrossRef]
- Todeschini, T.C.; Wu, Y.; Naji, A.; Mondi, R.; Nobrega, F.L. Kiwa rescues RecBCD for anti-phage activity. bioRxiv 2023, 2023.2002.2026.530102. [Google Scholar] [CrossRef]
- Dy, R.L.; Przybilski, R.; Semeijn, K.; Salmond, G.P.; Fineran, P.C. A widespread bacteriophage abortive infection system functions through a Type IV toxin-antitoxin mechanism. Nucleic Acids Res 2014, 42, 4590–4605. [Google Scholar] [CrossRef]
- Garvey, P.; Fitzgerald, G.F.; Hill, C. Cloning and DNA sequence analysis of two abortive infection phage resistance determinants from the lactococcal plasmid pNP40. Appl Environ Microbiol 1995, 61, 4321–4328. [Google Scholar] [CrossRef]
- Dai, G.; Su, P.; Allison, G.E.; Geller, B.L.; Zhu, P.; Kim, W.S.; Dunn, N.W. Molecular characterization of a new abortive infection system (AbiU) from Lactococcus lactis LL51-1. Appl Environ Microbiol 2001, 67, 5225–5232. [Google Scholar] [CrossRef]
- Deng, Y.M.; Liu, C.Q.; Dunn, N.W. Genetic organization and functional analysis of a novel phage abortive infection system, AbiL, from Lactococcus lactis. J Biotechnol 1999, 67, 135–149. [Google Scholar] [CrossRef]
- Millman, A.; Melamed, S.; Amitai, G.; Sorek, R. Diversity and classification of cyclic-oligonucleotide-based anti-phage signalling systems. Nat Microbiol 2020, 5, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.; Arrowsmith, T.J.; Went, S.C.; Blower, T.R. Toxin-antitoxin systems as mediators of phage defence and the implications for abortive infection. Curr Opin Microbiol 2023, 73, 102293. [Google Scholar] [CrossRef] [PubMed]
- Sberro, H.; Leavitt, A.; Kiro, R.; Koh, E.; Peleg, Y.; Qimron, U.; Sorek, R. Discovery of functional toxin/antitoxin systems in bacteria by shotgun cloning. Mol Cell 2013, 50, 136–148. [Google Scholar] [CrossRef]
- Payne, L.J.; Todeschini, T.C.; Wu, Y.; Perry, B.J.; Ronson, C.W.; Fineran, P.C.; Nobrega, F.L.; Jackson, S.A. Identification and classification of antiviral defence systems in bacteria and archaea with PADLOC reveals new system types. Nucleic Acids Res 2021, 49, 10868–10878. [Google Scholar] [CrossRef]
- Tal, N.; Morehouse, B.R.; Millman, A.; Stokar-Avihail, A.; Avraham, C.; Fedorenko, T.; Yirmiya, E.; Herbst, E.; Brandis, A.; Mehlman, T.; et al. Cyclic CMP and cyclic UMP mediate bacterial immunity against phages. Cell 2021, 184, 5728–5739. [Google Scholar] [CrossRef]
- Boyle, T.A.; Hatoum-Aslan, A. Recurring and emerging themes in prokaryotic innate immunity. Curr Opin Microbiol 2023, 73, 102324. [Google Scholar] [CrossRef]
- Jabbar, M.A.; Snyder, L. Genetic and physiological studies of an Escherichia coli locus that restricts polynucleotide kinase- and RNA ligase-deficient mutants of bacteriophage T4. J Virol 1984, 51, 522–529. [Google Scholar] [CrossRef]
- Kaufmann, G. Anticodon nucleases. Trends Biochem Sci 2000, 25, 70–74. [Google Scholar] [CrossRef]
- Cram, D.; Ray, A.; Skurray, R. Molecular analysis of F plasmid pif region specifying abortive infection of T7 phage. Mol Gen Genet 1984, 197, 137–142. [Google Scholar] [CrossRef]
- LeRoux, M.; Srikant, S.; Teodoro, G.I.C.; Zhang, T.; Littlehale, M.L.; Doron, S.; Badiee, M.; Leung, A.K.L.; Sorek, R.; Laub, M.T. The DarTG toxin-antitoxin system provides phage defence by ADP-ribosylating viral DNA. Nat Microbiol 2022, 7, 1028–1040. [Google Scholar] [CrossRef]
- Owen, S.V.; Wenner, N.; Dulberger, C.L.; Rodwell, E.V.; Bowers-Barnard, A.; Quinones-Olvera, N.; Rigden, D.J.; Rubin, E.J.; Garner, E.C.; Baym, M.; et al. Prophages encode phage-defense systems with cognate self-immunity. Cell Host Microbe 2021, 29, 1620–1633. [Google Scholar] [CrossRef]
- Burroughs, A.M.; Ando, Y.; Aravind, L. New perspectives on the diversification of the RNA interference system: insights from comparative genomics and small RNA sequencing. Wiley Interdiscip Rev RNA 2014, 5, 141–181. [Google Scholar] [CrossRef]
- Chakravarti, A.; Patel, D.J. Atypical bacterial Argonautes regulate antiphage defense. Cell Res 2023, 33, 655–656. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; van der Oost, J.; Koonin, E.V. Prokaryotic homologs of Argonaute proteins are predicted to function as key components of a novel system of defense against mobile genetic elements. Biol Direct 2009, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, T.; Sberro, H.; Weinstock, E.; Cohen, O.; Doron, S.; Charpak-Amikam, Y.; Afik, S.; Ofir, G.; Sorek, R. BREX is a novel phage resistance system widespread in microbial genomes. EMBO J 2015, 34, 169–183. [Google Scholar] [CrossRef]
- Millman, A.; Bernheim, A.; Stokar-Avihail, A.; Fedorenko, T.; Voichek, M.; Leavitt, A.; Oppenheimer-Shaanan, Y.; Sorek, R. Bacterial retrons function in anti-phage defense. Cell 2020, 183, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Mestre, M.R.; Gonzalez-Delgado, A.; Gutierrez-Rus, L.I.; Martinez-Abarca, F.; Toro, N. Systematic prediction of genes functionally associated with bacterial retrons and classification of the encoded tripartite systems. Nucleic Acids Res 2020, 48, 12632–12647. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Wu, G.; Wei, Y.; Liu, L.; Zhang, Y.; Su, R.; Jiang, X.; Li, M.; Gao, H.; Tian, X.; et al. SspABCD-SspE is a phosphorothioation-sensing bacterial defence system with broad anti-phage activities. Nat Microbiol 2020, 5, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Thiaville, J.J.; Kellner, S.M.; Yuan, Y.; Hutinet, G.; Thiaville, P.C.; Jumpathong, W.; Mohapatra, S.; Brochier-Armanet, C.; Letarov, A.V.; Hillebrand, R.; et al. Novel genomic island modifies DNA with 7-deazaguanine derivatives. Proc Natl Acad Sci U S A 2016, 113, E1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Maestri, A.; Pursey, E.; Chong, C.; Pons, B.J.; Gandon, S.; Custodio, R.; Chisnall, M.; Grasso, A.; Paterson, S.; Baker, K.; et al. Bacterial defences interact synergistically by disrupting phage cooperation. bioRxiv 2023, 2023.2003.2030.534895. [Google Scholar] [CrossRef]
- Guo, L.; Sattler, L.; Shafqat, S.; Graumann, P.L.; Bramkamp, M. A Bacterial dynamin-like protein confers a novel phage resistance strategy on the population level in Bacillus subtilis. mBio 2021, 13, e0375321. [Google Scholar] [CrossRef] [PubMed]
- Ka, D.; Oh, H.; Park, E.; Kim, J.H.; Bae, E. Structural and functional evidence of bacterial antiphage protection by Thoeris defense system via NAD(+) degradation. Nat Commun 2020, 11, 2816. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guan, J.; Hareendranath, S.; Crawford, E.; Agard, D.A.; Makarova, K.S.; Koonin, E.V.; Bondy-Denomy, J. A family of novel immune systems targets early infection of nucleus-forming jumbo phages. bioRxiv 2022, 2022.2009.2017.508391. [Google Scholar] [CrossRef]
- Meeske, A.J.; Nakandakari-Higa, S.; Marraffini, L.A. Cas13-induced cellular dormancy prevents the rise of CRISPR-resistant bacteriophage. Nature 2019, 570, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol Lett 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed]
- Nasrin, S.; Hegerle, N.; Sen, S.; Nkeze, J.; Sen, S.; Permala-Booth, J.; Choi, M.; Sinclair, J.; Tapia, M.D.; Johnson, J.K.; et al. Distribution of serotypes and antibiotic resistance of invasive Pseudomonas aeruginosa in a multi-country collection. BMC Microbiol 2022, 22, 13. [Google Scholar] [CrossRef]
- Hynes, A.P.; Rousseau, G.M.; Agudelo, D.; Goulet, A.; Amigues, B.; Loehr, J.; Romero, D.A.; Fremaux, C.; Horvath, P.; Doyon, Y.; et al. Widespread anti-CRISPR proteins in virulent bacteriophages inhibit a range of Cas9 proteins. Nat Commun 2018, 9, 2919. [Google Scholar] [CrossRef]
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic engineering of bacteriophages against infectious diseases. Front Microbiol 2019, 10, 954. [Google Scholar] [CrossRef]
- Hobbs, S.J.; Wein, T.; Lu, A.; Morehouse, B.R.; Schnabel, J.; Leavitt, A.; Yirmiya, E.; Sorek, R.; Kranzusch, P.J. Phage anti-CBASS and anti-Pycsar nucleases subvert bacterial immunity. Nature 2022, 605, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Garushyants, S.K.; Hurk, A.v.d.; Aparicio-Maldonado, C.; Kushwaha, S.K.; King, C.M.; Ou, Y.; Todeschini, T.C.; Clokie, M.R.J.; Millard, A.M.; et al. Synergistic anti-phage activity of bacterial defence systems. bioRxiv 2023, 2022.2008.2021.504612. [Google Scholar] [CrossRef]
- Tesson, F.; Bernheim, A. Synergy and regulation of antiphage systems: toward the existence of a bacterial immune system? Curr Opin Microbiol 2023, 71, 102238. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.L.; Castro, B.; Govindarajan, S.; Solvik, T.; Escalante, V.; Bondy-Denomy, J. Bacterial alginate regulators and phage homologs repress CRISPR-Cas immunity. Nat Microbiol 2020, 5, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Ngiam, L.; Weynberg, K.D.; Guo, J. The presence of plasmids in bacterial hosts alters phage isolation and infectivity. ISME Commun 2022, 2, 75. [Google Scholar] [CrossRef]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol Biol 2009, 501, 141–149. [Google Scholar] [CrossRef]
- Gaborieau, B.; Vaysset, H.; Tesson, F.; Charachon, I.; Dib, N.; Bernier, J.; Dequidt, T.; Georjon, H.; Clermont, O.; Hersen, P.; et al. Predicting phage-bacteria interactions at the strain level from genomes. bioRxiv 2023, 2023.2011.2022.567924. [Google Scholar] [CrossRef]
- Lam, J.S.; Taylor, V.L.; Islam, S.T.; Hao, Y.; Kocincova, D. Genetic and functional diversity of Pseudomonas aeruginosa lipopolysaccharide. Front Microbiol 2011, 2, 118. [Google Scholar] [CrossRef]
- DebRoy, C.; Fratamico, P.M.; Yan, X.; Baranzoni, G.; Liu, Y.; Needleman, D.S.; Tebbs, R.; O’Connell, C.D.; Allred, A.; Swimley, M.; et al. Comparison of O-antigen gene clusters of all O-serogroups of Escherichia coli and proposal for adopting a new nomenclature for O-typing. PLoS One 2016, 11, e0147434. [Google Scholar] [CrossRef]
- Mencke, J.L.; He, Y.; Filippov, A.A.; Nikolich, M.P.; Belew, A.T.; Fouts, D.E.; McGann, P.T.; Swierczewski, B.E.; Getnet, D.; Ellison, D.W.; et al. Identification and characterization of vB_PreP_EPr2, a lytic bacteriophage of pan-drug resistant Providencia rettgeri. Viruses 2022, 14, 708. [Google Scholar] [CrossRef]
- Sergueev, K.V.; Filippov, A.A.; Farlow, J.; Su, W.; Kvachadze, L.; Balarjishvili, N.; Kutateladze, M.; Nikolich, M.P. Correlation of host range expansion of therapeutic bacteriophage Sb-1 with allele state at a hypervariable repeat locus. Appl Environ Microbiol 2019, 85, e01209–19. [Google Scholar] [CrossRef]
Figure 4.
Antiphage systems present in 15 or more strains, represented by percent distribution in phage susceptibility groups. If greater than 52.5% of the strains encoding a system are categorized as resistant, the system is considered prevalent among phage-resistant strains.
Figure 4.
Antiphage systems present in 15 or more strains, represented by percent distribution in phage susceptibility groups. If greater than 52.5% of the strains encoding a system are categorized as resistant, the system is considered prevalent among phage-resistant strains.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



