
Submitted:
16 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
The consumption of foods with fiber and phenolic compounds can be part of treating obesity through interaction with the gastrointestinal tract. Treatment with common bean leaves (BL) for obesity was evaluated in mice on a high-fat, high-fructose diet (HFFD). The proximal and phytochemical characterization of BL was determined. Obesity was induced with a HFFD for 14 weeks. Immediately after, the animals were supplemented with BL (10%) or standard diet (SD) as a strategy to seem a healthy diet (12 additional weeks). Changes in body composi-tion, lipid profile, and intestinal integrity were analyzed. BL contained 27.2% dietary fiber, total phenolics (475.04 mg/100 g), and saponins (2.2 mg/100 g). The VAT consumption of the BL group decreased by 52% compared to the HFFD group. Additionally, triglyceride concentrations were 23% lower in the BL consumption group compared to the HFFD group. The improvement in lipid profile was attributed to the (1.77-fold) higher fecal lipid excretion in the BL consumption group compared to the HFFD group. Likewise, pancreatic lipase decreased by 29%. Furthermore, BL supplementation reduced the level of IL-6 (4.4-fold) and FITC-dextran in serum by 50% compared with that in the HFFD group. Metabolic endotoxemia was inhibited after BL supplementation (-33%) compared to the HFFD group. BL consumption as a treatment in obese mice reduces adipose tissue accumulation and improves the lipid profile. Furthermore, we report for the first time that BL consumption improves intestinal integrity.
Keywords:
bean leaves
; obesity treatment
; intestinal integrity
1. Introduction
Obesity is a pathology characterized by excess adipose tissue and metabolic alterations such as dyslipidemia, hyperglycemia, and insulin resistance [1]. One of the main causes of obesity is excessive consumption of fats and sugars [2,3]. It is also known that excessive consumption of fructose and saturated fats modifies the synthesis of metabolites in the colon by the intestinal microbiota [4].
Furthermore, lipid intake stimulates the synthesis of pro-inflammatory metabolites such as indoxyl sulfuric, arachidonic, and stearic acids in the colon which promote intestinal permeability by inhibiting the action of tight junction proteins. This inhibition promotes intestinal permeability and allows the passage of toxin-producing bacteria, such as lipopolysaccharides (LPS), from the intestinal lumen into the systemic circulation, reaching organs such as the liver and brain and promoting inflammation characterized in part by the increase of proinflammatory cytokines such as IL-6 and IL-2 [4,5].
In previous studies in murine animals, it has been reported that the type of macronutrients that make up the diet affects intestinal adaptation. Increasing intake favors cell proliferation, the length of the villi, and the depth of the crypts; while nutrient deficiency generates the opposite[6,7]. Specifically, it has been reported that the consumption of saturated fats in mice increases the length of the villi, which in turn favors the surface area for nutrient absorption. [6].
Previously, a relationship between consuming that include fats and sugars has been associated with diets and developing alterations in intestinal integrity has been suggested [8]. On the other hand, there are diets that have been related to their health benefits. The WHO recommends the daily intake of 400 g of vegetables, including legumes as part of a healthy diet, since they are the main source of fiber and phytochemical compounds in the diet; in addition, it provides vitamins and minerals [9].
Beans are one of the most produced plants in Mexico. From the harvest of the seeds that are part of the basic diet of the population, the leaves that are mostly obtained are discarded [10]. However, some rural populations consume this part of the plant that is classified as “quelites” or edible plants [11]. These quelites contain 24.20% crude protein and 24.10% total dietary fiber and a minimum contribution of lipids [12]. Likewise, bean leaves are a source of iron (1.65 mg/g) and phytochemical compounds such as total phenols (2 459 mg/100g), soluble phenols (474 mg/100g) and flavonoids (949 mg/100g) [12,13].
The nutraceutical properties of its leaves have been scarcely investigated. Until now, it has been reported that the consumption of common bean leaves (Phaseolus vulgaris L.) prevents the development of obesity in rats fed with HFFD, since it reduces weight gain (12%), the abdominal fat accumulation (10%) and early insulin resistance (6%). Interestingly, the improvement of metabolic alterations is related to the higher concentration of SCFAs (54%) in the cecal content p < 0.05 [12]. In another study, supplementation with bean leaves (Phaseolus vulgaris L.) of the Eugenia variety for 13 weeks prevented hepatic lipotoxicity in rats fed HFFD [14].
With the above, it is suggested a strategy to decreased obesity is by improving intestinal integrity. Bean leaves have been recommended for the prevention of obesity. However, its effect as a treatment and relationship with the improvement of intestinal integrity is unknown. Therefore, the main objective of this study was to determine the effect of bean leaves ingestion as a treatment on metabolic complications and intestinal integrity in C57BL/6 mice with obesity induced by a high-fat, high-fructose diet.
2. Materials and Methods
2.1. Bean leaves
The Phaseolus vulgaris L. variety Eugenia plant was harvested in June 2019 by sowing at the Amazcala experimental campus, Autonomous University of Queretaro (Faculty of Natural Sciences, UAQ). The bean plants were grown in the open field and kept free of pesticides. After harvesting, green leaves (6–15 cm long and 3–11 cm wide) were selected, washed with distilled water, and dried in a convection oven at 45 °C up to constant weight. The dried bean leaves (BLs) were ground, sieved to 0.2 mm (Thomas-Wiley mill model 4) and stored at -80°C.
2.2. Bromathological and phytochemical characterization of the common bean leaves
2.2.1. Bromatological characterization of the common bean leaves
To determine the humidity of the BL powder, 2 g of the sample was weighed in porcelain crucibles, previously placed at constant weight in a drying oven at 65 °C. The BL sample was dried to constant weight, then removed from the oven, placed in a desiccator for 15 minutes, weighed again, and the difference was calculated. Ash content was determined by calcination (AOAC 942.05). The protein was analyzed by the total nitrogen macro-Kjeldahl method (AOAC 2001). Lipids were determined by the ethereal extraction method (AOAC 920.39). Total dietary fiber was obtained by an enzymatic method using a kit (Total Dietary Fiber Assay Kit, Sigma-Aldrich) with the modifications reported by Heredia-Ortega et al. (2014) [15]. All techniques were performed in triplicate on dry matter.
2.2.2. Extraction of phenolic compounds
The sample (5 g of dried BL flours) was extracted with 10 mL of 70:30 methanol: water and sonicated three times for 30 s, with a resting of 15 s between cycles. The sample was centrifuged at 10,000 g for 10 min at 4 °C, and the supernatant was recovered and concentrated by using a rotary vacuum evaporator to evaporate the methanol [16]. The extract was dissolved in 1 mL of sterile distilled water and lyophilized at −60 °C [17]. For the determination of anthocyanins, the extracted sample (0.5 g) was dissolved in 4 mL of acidified ethanol (85 mL of 95% ethanol + 15 mL of 1.0 N HCl), mixed for 2 min and the pH was adjusted to 1.0 with HCl. Subsequently, the sample was shaken for 30 min, centrifuged at 10,000 g for 20 min, and stored at −80 °C until analysis.
2.2.3. Determination of total phenols
The quantification of total phenols was performed by the Folin-Ciocalteu assay [18]. In a 96-well plate, 12.5 µL of the acetone extract plus 50 µL of water were placed; 32 µL of 1 N Folin-Ciocalteu reagent was added and the oxidation reaction was neutralized with 156 µL of 7% sodium carbonate. The sample was incubated in the dark for 2 h at room temperature and the absorbance was measured at 760 nm in a plate reader (Varioskan Flash Multimode Reader, Thermo-Scientific, MA, USA). The total phenol content was calculated from a calibration curve using gallic acid as a standard. The results are expressed in µg gallic acid equivalents/g dry sample (µg GAE/g dry sample).
2.2.4. Determination of total flavonoids
The quantification was carried out as described by Oomah et al. (2005) [19]. For this 50 µL of the acetone extract was placed in a 96-well plate and 180 µL of methanol was added, followed by 20 µL of a 1% solution of 2-aminoethyldiphenylborate in methanol. Absorbance was measured at 404 nm in a plate reader. The total flavonoid content was calculated from a calibration curve using rutin as a standard and the results are expressed as µg of rutin equivalents/g of dry sample (µg RE/g of dry sample).
2.2.5. Quantification of condensed tannins
It was carried out by the vanillin test described by Deshpande and Cheryan (1987), and modified to a microplate. 50 µL of the acetone extract was placed in a 96-well plate and 200 µL of a freshly prepared 0.5% vanillin mixture (1% vanillin in methanol and 8% HCl in methanol in a 1:1 ratio). In addition, a blank was prepared by adding 50 µL of the extract and 200 µL of 4% HCl. The absorbance was measured in a plate reader at 492 nm. To calculate the concentrations of condensed tannins, a calibration curve was obtained with (+)-catechin (0-0.2 mg/mL). The results are expressed as mg (+)-catechin equivalents/g dry sample (mg ECAT/g dry sample) [20].
2.2.6. Quantification of total saponins
BL powder (1 g) was weighed and 10 mL of 80% methanol was added. The methanol was removed by rotary evaporation under vacuum conditions. 1 mL of distilled water and 200 µL of acetone were added to the extract, and it was centrifuged at 2,500 g for 15 min to obtain the methanol extract. Subsequently, 0.1 mL of the methanol extract was placed and incubated with 1 mL of 72% sulfuric acid and 0.8% vanillin in ethanol. The mixture was incubated in a water bath at 60 °C for 20 min and subsequently cooled with an ice bath. Absorbance were measured at 544 nm in a plate reader. A saponin standard (0.012-0.36 mg/mL) was used to determine the amount of saponins in the extract [21].
2.2.7. Identification and quantification of phenolic and flavonoids in bean leaves by UPLC-ESI-MS/MS
From the extracts previously obtained, the samples were resuspended in 200 μL of methanol and filtered (0.45 μm PTFE). Subsequently, the samples were prepared at a concentration of 10mg of extract/mL, 2µL were injected into a C18 reverse-phase column (Acquity UPLCr BEH C18 1.7 μm, 2.1 mm X 100 mm), at a flow rate of 0.25mL. /min. Spectrometric conditions included a capillary voltage of 2.15kV, cone of 30V, source output 60V, source temperature of 150 ºC, desolvation temperature of 400 ºC, a cone gas flow of 150L/h, collision gas flow of 0.15mL/h, nebulization gas flow of 7.00 bar. The collision energy in MS mode of 2.00 and in MS/MS mode of 20.00. The mixture of water: 7.5 mM formic acid was used for phase (A) and acetonitrile as phase (B) at a flow rate of 0.25 mL/min and sample volumes of 2 μL. As elution gradient (0 min, 97% A, 3% B; 1.23 min, 91% A, 9% B; 3.82 min, 84% A, 16% B; 11.40 min, 50% A, 50% B; 13.24 min, 97% A, 3% B; 15 min, 98% A, 2% B)..The results were expressed in μg/g of extract [16].
2.3. Experimental design
2.3.1. Experimental animals
For the obesity-induced model, 72 8-week-old male C57BL/6 mice (25 to 30 g) were purchased from the Bioterio Animal of the Institute of Neurobiology (INB-UNAM Campus Juriquilla). Animals were housed by placing 4 mice per box in standard boxes (dimensions: 19 x 29 x 12 cm), with solid continuous floors and walls, removable lids, and sterile shaving litter (1.5 cm high). The animals were kept at temperature control (22 ± 3 °C) and relative humidity (60 ± 5%) conditions under a regulated 12-h light/dark cycle. Food and water were administered ad libitum. The cleaning of the cages, the bottles, and the feeders was carried out every third day. After one week of acclimatization, mice were randomly assigned into 8 different groups. The experiments on animals were performed by the Animal Care and Use protocol (NOM-062-ZOO-1999), approved by the Ethics Committee of the Autonomous University of Queretaro (Project identification code CBQ22/114, approved September 21, 2022). C57BL/6 male mice were randomly assigned into 6 groups (n = 12); the experimental groups included: 1) and 2) standard diet (SD), 3) and 4) high fat and fructose diet (HFFD), 5) HFFD diet + Bean leaves (BL consumption), 6) HFFD diet followed by SD standard diet (change to healthy diet) to simulate a change to the recommended diet.
2.3.2. Design of diets for experimental animals
The SD diet (Rodent Lab Chow 5001) is formulated for adequate nutrition during the life cycle of rodents and includes the nutritional and energy requirements (3.36 Kcal/g, 28.5% proteins, 13.5% fats, and 58% carbohydrates). The BL diet was prepared by adding the BL powder to the HFFD group (10 % w:w), thus simulating the recommended daily intake of vegetables per day. At the beginning of study, the animals had one week of adaptation to the powdered food base.
2.3.3. Energy and nutritional content of diets
The energy and nutritional composition of the diets for three diets the SD, HFFD and BL consumption groups are shown in Table 1. The diets corresponding to the HFFD and BL consumption groups were designed to be isocaloric. To keep protein intake similar between both diets (isoprotein diets), calcium caseinate was added. Additionally, to promote the absorption of dietary lipids, 0.15% sodium cholate was added.
2.3.4. Experimental design
The mice in groups 1 and 2 were fed an SD diet, while groups 3 – 6 were fed an HFFD diet. Changes in weight gain, food intake, and visceral circumferences were monitored weekly. After 14 weeks, the body weight of the HFFD-fed animals increased by≥ 10% compared to that of the SD group and it was set as the obesity-induced period, according to the indicator of obesity reported in the literature [22,23]. Therefore, the animals in groups 1 (SD, n = 12) and 3 (HFFD, n = 12) were sacrificed; their large intestine was removed and washed to confirm alterations in intestinal integrity. Afterward, the animals in the rest of the groups (2, 4-6) were supplemented according to the assigned treatments shown in Table 2 for an additional 12 weeks.
2.4. Changes in body composition from the consumption of bean leaves and the change to a healthy diet.
2.4.1. Body composition
To evaluate the distribution of the body fat, the thoracic and abdominal circumferences were measured, and the abdominal circumference/thoracic circumference ratio was determined in the week previous to the euthanasia before the 14-week obesity-induced period and at the end of the experiment (26 weeks).
2.4.2. Quantification of adipose tissue
To determine the distribution of body fat, the animals were subjected to magnetic resonance imaging (Service Unit/Laboratory of the National Magnetic Resonance Imaging Laboratory, LANIREN INB-UNAM, Campus Juriquilla, Qro.) in the last week before euthanasia, using isoflurane 2% as an inhalation anesthetic. The images were analyzed with the ITK-SNAP program version 3.8.0 (University of Pennsylvania, Pennsylvania, USA) and the results were expressed as the volume in mm3.
2.5. Role of bean leaves consumption and the change to a healthy diet in lipid metabolism.
2.5.1. Determination of the content of total cholesterol and triglycerides in serum.
From the serum of the different groups obtained from the euthanasia, the concentrations of cholesterol and triglycerides were analyzed with the corresponding enzymatic-colorimetric kits (Spinreact, Spain). The results were expressed as mg/dL.
2.5.2. Total lipids and triglycerides in feces
Feces were collected over 24 h in the last week of the treatment phase and stored at −80°C. 200 mg of feces was homogenized with 2 mL of 0.9% saline and shaken for 1 min, then 2 mL of chloroform-methanol (2:1 v/v) was added and the samples were shaken for 1 min. Subsequently, the samples were centrifuged at 10,000 g for 10 min, and the lower phase was recovered and evaporated under vacuum for 24 h. The difference in initial and final weight was determined and the results were expressed as mg of total lipids per g of feces [24]. Additionally, triglycerides in feces lipid extract were determined using an enzymatic–colorimetric kit (Spin react, Spain). Results were expressed as mg of triglycerides per g of total lipids in feces.
2.5.3. Pancreatic lipase activity in vitro
To demonstrate the inhibition of pancreatic lipase activity of bean leaves, a sample of bean leaves powder was mixed in a water-soluble extract and determined following the method proposed by Pérez Ramírez et al. (2017) [25]. The results were expressed as a percentage of inhibition, and tetrahydrolipstatin was used as a positive control.
2.6. Euthanasia
The euthanasia procedure in both phases (at 14 and 26 weeks) was performed by two methods, according to the subsequent analyses. In this regard, half of the animals in each group were sacrificed by administering CO2 by inhalation with a flow rate of 20% of the chamber volume per minute, for 60 s, and blood was subsequently obtained by cardiac puncture. Afterward, the animals were dissected, the gastrointestinal tract was removed and washed with saline solution (0.9%). The other half of the animals (n = 6) were sacrificed by intraperitoneal administration of pentobarbital (120 mg/kg) dissolved in saline solution and subsequent cardiac puncture to draw blood. Blood samples were collected in yellow vacutainer tubes. Serum was obtained by centrifugation at 3000 g for 20 min and stored in aliquots at −20 °C until analysis. The cleansed intestines were placed in a 10% formaldehyde solution for paraffin embedding and the tissues were fixed on slides until histology morphometry analysis.
2.7. Impact of bean leaves consumption and healthy diet change on intestinal integrity and metabolic endotoxemia.
2.7.1. Intestinal permeability by the FITC-dextran assay
The intestinal permeability was assessed by the in vivo FITC-dextran permeability assay [26] a week before the both euthanasia of all animals. After a 12-h fast, the animals were administered a dose of FITC-dextran (Sigma-Aldrich, USA) (600 mg/kg body weight) dissolved in sterile phosphate-buffered saline (Sigma-Aldrich) by oral cannulation. 2.5 h after FITC-dextran administration, approximately 200 µL of blood was drawn via the femoral vein and centrifuged for 15 min at 3,500 g at 4 °C; the plasma was placed in new tubes and stored at -80 °C for later analysis. The plasma FITC-dextran concentrations were assessed in a plate reader with an excitation wavelength of 490 nm and an emission wavelength of 520 nm [27]. The results are expressed in µg/mL of FITC-dextran in plasma.
2.7.2. Histopathological and morphometric analysis of jejunum and proximal colon
Sections of the jejunum and the proximal colon tissues were fixed in Carnoy’s solution (ethanol: chloroform: acetic acid 6:3:1) and stained with hematoxylin and eosin to determine the extent of damage due to HFFD consumption. For this, 3 fields per section were observed in 3 sections of each mouse sample under a microscope (Velab, VE-BC3PLUS, MEX) and images were captured. The evaluation of histological damage parameters was performed by using a mouse histological guide to determine the extent of damage and the level of inflammation in crypts and villi of the jejunum and colon tissues [28].
2.7.3. Determination of LPS and IL-6 levels in serum
To evaluate the effect of BL consumption on metabolic endotoxemia, serum concentrations of the LPS (Abbexa LDT, Cambridge, GBR) and the proinflammatory interleukin 6 (IL-6) (Express Biotech International, Wedgewood, USA) were measured by the ELISA techniques using the corresponding kits and following the manufacturer’s instructions.
2.8. Statistical analysis
The results of bean leaves composition, changes in body composition, cholesterol and triglycerides, food and water consumption, intestinal permeability test, and the serum levels of LPS and IL-6 are expressed as the means ± standard error (SEM). The Chi-square test was employed to analyze the histopathological changes of the colon and jejunum portions. Changes in body composition, lipid profile, and intestinal integrity parameters were associated with the different diets using Pearson correlation. Analysis of variance (ANOVA) was used to determine significant differences among groups followed by post hoc Tukey’s test (p ≤ 0.05). The statistical program GraphPad Prism 8 (1995-2022 GraphPad Software, LLC.) was used for the graphs and the SPSS Statistical Software version 18.0 (IBM) was used for the analyses.
The results are presented as the mean and standard deviation (n=2). The content is expressed as the amount in µg/g of extract.
3. Results
3.1. Quantification of dietary fiber and phytochemical compounds.
Bean leaves composition included 26.91% of protein, 27.2% of dietary fiber, and a minimal amount of lipids. On the other hand, the phytochemical characterization showed a total phenolic content of 475.04 ± 25.47 mg GAE/100 g, flavonoids of 243.53 ± 98.40 mg EC/100 g, and tannins of 1.38 ± 0.17 mg EC/100 g. While, the total saponins content was 217.08 ± 35.69 mg/ 100 g.With the identification of phenolic acids and flavonoids, it was found that the pre-dominant compounds in bean leaves are coumaric acid (1283 ug/g), caftaric acid (207 ug/g), 4-hydroxybenzoic acid (106 ug /g), quinic acid (97 ug/g) and ferulic acid derivative (95 ug/g). While the predominant flavonoids are Quercetin 3-O-ß-glucuronide (6,892 ug/g), rutin (971 ug/g) and Quercetin 3-O-glucoside (73 ug/g) (Table 3).
3.2. Effect of bean leaves on energy intake
During the 14-wk obesity induction phase, the daily food intake (g/day) and water consumption were not affected by the type of diet; however, the energy density of the HFFD increased significantly the energy consumption of the HFFD 14-wk group by 41 % compared to that of the SD 14-wk group. However, no difference in the energy intake was observed between the HFFD 14-wk and the HFFD 26-wk groups. Unexpectedly, the energy intake of the SD 26-wk group was 18 % lower than the energy intake of the SD 14-wk group. On the other hand, the HFFD 26-wk and HFFD + BL supplementation groups had an increase in energy intake by 89.6 and 63.3%, respectively, compared to the SD 26-wk group. As expected, the change to healthy diet (SD) decreased the energy intake by 32% when compared to the HFFD 26-wk group (Table 4).
3.3. Treatment with bean leaves and its effect on body composition
3.3.1. Body weight and the increase in the abdominal circumference.
During the obesity induction phase, the body weight of the animals in the HFFD 14-wk group increased significantly by 23.4% compared to that of the SD 14-wk group, confirming the development of obesity; additionally, the abdominal circumference increased significantly in HFFD 14-wk group (26 %) compared to that of the SD 14-wk group. In the treatment phase, the abdominal circumference and the body weight of the animals in the HFFD 26-wk group were 25.5 and 6.6% higher, respectively, compared to those of the SD 26-wk group. Meanwhile, the consumption of bean leaves did not exert any difference in both indicators in the animals of the HFFD 26-wk group. Although no significant decrease in body weight was observed, the abdominal circumference was significantly reduced by 18.7% by the change to a healthy diet compared to that of HFFD 26-wk group (Table 5).
3.3.2. Determination of volume adipose tissue.
As expected, the visceral adipose tissue (VAT) and subcutaneous adipose tissue (SCAT) of the animals in the SD 26-wk group remained 56% and 48% lower, respectively, compared to those values of the animals fed HFFD 26-wk (Figure 1A). Interestingly, the consumption of bean leaves reduced the values of VAT by 52% and SCAT by 57%, compared to VAT and SCAT values of the animals fed HFFD 26-wk. Unexpectedly, although the final body weight and the abdominal circumference significantly decreased in the animals with the change to a healthy diet, no effect on adipose tissue was observed (Figure 1 B–C).
3.4. Impact of the consumption of different diets on the lipid profile.
3.4.1. Serum cholesterol and triglycerides concentration.
The serum triglyceride level in the animals of the HFFD 14-wk group was 1.75 times higher than in the animals of the SD 14-wk group (p < 0.05). Similarly, the serum cholesterol level (123 mg/dL) in the animals fed with HFFD 14-wk was higher (p < 0.05) compared to that in animals fed with SD 14-wk of (93 mg/dL). Similarly, at 26 weeks serum cholesterol and triglyceride concentrations were significantly higher in the HFFD 26-wk group (120 mg/dL and 141 mg/dL, respectively) compared to those of the SD 26-wk group (98 mg /dL and, 99 mg/ dL). On the other hand, triglyceride levels decreased significantly by 19 % in the animals fed HFFD and supplemented with BL, as well as in the animals fed with a healthy diet after induction of obesity by 21 %, compared to the values of the animals fed with HFFD 26-wk (p < 0.05) (Table 5).
3.4.2. Lipid excretion
Figure 2A shows the total lipids in feces of the different experimental groups. According to the diet composition and the energy intake, the lipid content of both groups fed the standard diet was similar. However, although the HFFD group consumed more dietary fat, its lipid content was slightly lower to those of the SD and healthy diet groups. Interestingly, bean leaves consumption increased total lipid content by 1.74-fold compared to that of the HFFD 26-wk group (p < 0.05). Regarding triglycerides in feces, a similar trend was observed, being significantly higher in animals fed with BL (107 mg/g) compared to animals fed with HFFD (41 mg/g) (Figure 2 B).
3.4.3. Inhibition of pancreatic lipase in vitro.
To determine whether the decrease in the accumulation of adipose tissue and the concentration of triglycerides was attributed to a lower absorption of dietary fat, the in vitro activity of pancreatic lipase was determined. It was shown that the BL methanolic extract inhibits pancreatic lipase activity by 29% (Figure 2 C).
3.5. Changes in morphometry and histopathology in the intestine and colon
3.5.1. Relative weight and length of the small intestine and colon.
On the other hand, the small intestine length decreased by 19% in the HFFD 14-wk group as compared to that of the SD 14-wk group. Additionally, the length of the colon was reduced by 27% in the HFFD 14-wk group compared to that of the SD 14-wk, probably as an inflammation indicator. The lengths of both intestines after the consumption of bean leaves and the change to a healthy diet was not different with HFFD 26-wk or SD 26-wk. Unexpectedly, a 23.3% decrease in the length of the colon was observed when switching to a healthy diet compared to the length of the colon of animals in the SD 26-wk group (Table 6).
Similarly, the consumption of HFFD 26-wk decreased the relative weight of the small intestine and colon tissues by 18 and 33%, respectively, compared to those of the group fed SD 26-wk. As expected, the change to a healthy diet recuperated the relative weight of the colon by 23%, compared to that of animals fed with HFFD 26-wk; however, no significant difference was found between BL consumption group and HFFD 26-wk group (Table 7).
3.5.2. Histological change in intestine and colon.
Figure 3 shows the representative micrographs and histological characteristics of the jejunum and colon portions of the animals of the different experimental groups. In the HFFD 26-wk group, 50% of the colon samples showed lymphoid hyperplasia and 25% developed villous atrophy, and in the jejunum, 50% of the analyzed structures showed infiltration of lymphoid tissue and the 25% had lymphoid hyperplasia (p < 0.05). Furthermore, the consumption of bean leaves improved the altered histological structures of the jejunum by 75% and the colon by 80%. Unexpectedly, 80% of the structures of the jejunum and 60% of the structures of the colon showed villous atrophy in the histological structures of the animals in the change to a healthy diet group. Likewise, infiltration of lymphoid tissue was observed in 20% of the samples analyzed from the jejunum and colon (p < 0.05). The small intestine and the colon tissues of the animals fed with SD 26-wk presented normal structures.
3.6. Intestinal permeability and metabolic endotoxemia.
3.6.1. Intestinal permeability.
One of the main characteristics of the altered intestinal integrity is the increase in the permeability of the small intestine. In this regard, the serum concentration of FITC-dextran in the HFFD group increased 4-fold compared with the serum concentration of FITC-dextran in the SD group (p < 0.05). The intestinal permeability of the animals that consumed BL or change to a healthy diet decreased by 50 and 55%, respectively, compared with that of animals fed HFFD for 26-wk (Figure 4).
3.6.2. Metabolic endotoxemia and inflammation
In this investigation, the serum LPS concentration in the 14-wk HFFD group was 324% higher (p < 0.05) compared with that in the 14-wk SD group. On the other hand, animals supplemented with bean leaves had a decrease in serum LPS concentration (33%), as well as animals with a change in feeding style (30%, p < 0.05) (Figure 5a). Furthermore, the serum IL-6 concentration in the 14-wk HFFD group was 2.3 times higher compared with the serum IL-6 concentration in the 14-wk SD group (p < 0.05). Furthermore, animals that consumed BL and THE change to a healthy diet had a significantly lower 4.5 and 3.19-fold serum IL-6 concentration, respectively, compared to the 26-wk HFFD group (Figure 5b).
3.6.3. Intake of dietary fiber and total phenols.
The HFFD 26-wk group had a lower intake of total dietary fiber and insoluble fiber of 4.5 and 3.56 times, respectively, compared to the intake of the SD 26-wk group. However, supplementation with BL increased the consumption of both total dietary fiber and insoluble fiber by 3.30 and 3.16 times, respectively, compared to those of the group that only consumed HFFD 26-wk. Also, the supplementation with 10% of bean leaves increases the dietary total phenolic content by 52% compared to the intake of total phenolic content of the HFFD 26-wk (Table 8).
4. Discussion
Bean leaves are a food source of bioactive compounds. Thus, in this study, we confirmed that bean leaves contain dietary fiber, mainly insoluble fiber, and are a source of phenols, flavonoids, and tannins. Furthermore, for the first time, we report the saponin content in bean leaves. Bioactive compounds are mainly metabolized by colonic microbiota, which synthesizes metabolites with potentially beneficial effects related to the maintenance of intestinal integrity [29]. The concentration of flavonoids in BL was similar to that reported in a previous study in the pinto villa variety [13]. Furthermore, our results are similar to those of Becerril-Campos (2022) who demonstrated that in rats with HFFD-induced obesity, 10% bean leaves prevented the development of obesity and increased the total SCFA concentration in the cecal contents [12].
Furthermore, the presence of phenolic acids was identified, among which coumaric acid stands out. Previously, administration of coumaric acid (10 mg/kg) in mice fed a high-fat diet was shown to decrease the amount of adipose tissue, adipocyte size, and leptin concentrations [30]. The proposed mechanism of action of coumaric acid is from the inhibition of the expression of genes in white adipose tissue; it also promotes the hepatic oxidation of fatty acids [30]. Meanwhile, among the flavonoids identified in BL, quercetin and kaempferol stand out, which have been shown to reduce the metabolic alterations of obesity. Administration of quercetin (0.05%) in a high-fat diet was able to decrease the transcription of proinflammatory cytokines in mice [31]. Meanwhile, kaempferol has been used in various studies in murine with high-fat diets, demonstrating that at doses between 10 to 200 mg/kg body weight, it can reduce body weight, serum concentrations of glucose, cholesterol and triglycerides, and the accumulation of adipose tissue [32,33,34]. When comparing the nutritional composition of bean leaves with the most consumed quelites such as quintonil (17.5 to 20.3% of proteins and 2.6% of crude fiber) quelite cenizo (4% of protein in fresh weight and flavonoids 3935 mg/g of extract) and purslane (total phenols 9.12 mg/g and total flavonoids 1.44 mg/g in dry weight) [35], bean leaves prove to be a competitive alternative with these quelites to be included in the diet.
In this study, the presence of tannins was also reported. Tannins are hydrolyzed into gallic acid, pyrogallol and phloroglucinol, which are precursors to acetate and butyrate. Also, urolithins are one of the main metabolites derived from tannins; More importantly, both urolithin A and urolithin B have been shown to be related to the prevention of obesity in rats fed a high-fat diet by reducing lipid accumulation in the liver and increasing lipid and fecal excretion [36,37]. With the above, the intake of bean leaves that contain fiber, phenolic compounds, saponins and tannins could be part of a healthy diet.
In relation to the model used to evaluate the effect of BL consumption as a treatment for obesity, animal models with obesity induced with diets rich in fat and fructose have been successfully reported to evaluate various intervention strategies. In this sense, the induction of obesity was demonstrated with the ingestion of a diet with fructose (20%) and lard (40%) for 14 weeks, characterized by a 10% overweight with respect to the body weight of the animals fed with SD. Likewise, animals fed HFFD presented greater abdominal circumference, greater accumulation of adipose tissue and increased serum levels of glucose, cholesterol and triglycerides when compared to the SD group. In addition to the above, average food consumption (g/day) was positively correlated with energy consumption (r = 0.749). These changes were attributed to excess fat that provides greater energy density which promotes body weight gain (r = 0.290), VAT (r = 0.457) and SCAT (r = 0.473). Meanwhile, it has been shown that the addition of fructose to the diet can exacerbate the metabolic alterations of obesity [38]. In previous studies, it has been confirmed that high-fat diets favor the development of obesity in a short time, while persistent intake of fructose favors the development of non-alcoholic fatty liver [39,40,41]. Therefore, the combination of lard and fructose in a high-calorie diet aggravates metabolic complications.
Regarding the effect of BL consumption on body composition and lipid profile, body weight did not decrease. However, the accumulation of VAT and SCAT, as well as serum triglycerides, was inhibited. Meanwhile, a change to a healthy diet reduced abdominal circumference and decreased serum triglyceride concentrations.
Increased abdominal circumference has been considered an independent predictor of morbidity, associated with greater accumulation of VAT [42]. This increase in adipose tissue can promote chronic low-grade inflammation, insulin resistance, and cardiovascular risk [43]. Interestingly, the reduction in adipose tissue due to BL consumption can be attributed to the contribution of phenolic compounds (r = - 0.451).
It was previously reported that the consumption of bean leaves in rats on an obesogenic diet prevents weight gain and the amount of adipose tissue without decreasing energy consumption [12,14].
In this study, body weight was negatively correlated with total phenol consumption (r = - 0.306) and subcutaneous adipose tissue (SCAT, r = - 0.452). It was shown that despite not decreasing body weight, BL consumption decreases adipose tissue accumulation and serum triglyceride concentrations in animals fed HFFD. It is relevant for future research to evaluate changes in other body compartments such as muscle mass, water content and bone tissue to understand why there is no difference in body weight.
The increase in triglycerides and their accumulation is considered a risk factor for the development of diabetes and cardiovascular diseases [44]. In this study, BL intake and switching to a healthy diet decreased serum triglyceride concentrations that were previously increased with HFFD. Previous studies evaluated the preventive effect of consuming cooked bean seeds of different varieties (60% of the diet) and bean pod extract (300 mg/kg body weight) in obesogenic models, which decreased serum triglycerides, which was combined with a protective effect on the development of diabetes and cardiovascular diseases [45,46]. In this study we co-tested the treatment effect of BL consumption on hypertriglyceridemia.
The above could be due in part to the increase in the excretion of lipids and triglycerides in feces partially due to the inhibition of pancreatic lipase since the in vitro assay showed an enzymatic inhibition of 29%. The above is of great relevance since pancreatic lipase is released in the small intestine to break down fats along with bile salts so that they can be absorbed [47]. Previous studies in murine models have shown that in vitro inhibition of pancreatic lipase in proportions similar to those in this study is closely related to the reduction of metabolic alterations of obesity [47,48].
As part of the effect of BL treatment on intestinal integrity, changes in the length and weight of the small intestine and colon were evaluated. Previously, it was reported that diets high in fat (greater than 30%) and sugar promote alterations in intestinal integrity[49,50,51]. In this study, the small intestine and colon of animals fed HFFD had a lower relative weight, which was not recovered with BL consumption or the change to a healthy diet. Previously, in mice fed a high-fat diet, it has been reported that decreased intestinal length and weight are associated with intestinal inflammation that could exacerbate intestinal permeability [26].
In addition to the above, histopathological changes in regions of the small intestine and colon related to alterations in intestinal integrity were analyzed. In the jejunum of the HFFD-fed group, increased lymphocytic infiltration and hyperplasia will be observed in parallel with a decrease in colonic length and an increase in circulating IL-6. Meanwhile, treatment with 10% BL restored the villi and no lymphocyte infiltration was observed, which could be related to less inflammation [52]. Interestingly, 75% of the jejunum portions and 80% of the colon portions of animals consuming 10% BL showed no histopathological changes, suggesting a beneficial effect against intestinal inflammation.
It was previously described that villous atrophy is associated with lower nutrient absorption, as in the case of this study, in the healthy diet change group. The alterations in colon and jejunum histologies induced by the consumption of HFFD for 14 weeks were restored with the consumption of 10% BL, protecting against potential intestinal inflammation.
HFFD intake promotes intestinal permeability and metabolic endotoxemia that decreases both with BL consumption and with the change to a healthy diet. In this study, the increase in serum FITC-dextran concentration associated with intestinal permeability was demonstrated. While, the change to a healthy diet and the consumption of BL reduced the levels of LPS and IL-6 in sleep. Regarding the development of metabolic endotoxemia, HFFD consumption was found to increase serum concentrations of LPS and IL-6 (r = 0.98 and r = 0.99, respectively). In contrast, switching to a healthy diet was negatively correlated with serum LPS and IL-6 concentrations (r = -0.885 and r = -0.996, respectively). Furthermore, BL consumption decreased serum LPS concentrations and was inversely correlated with serum IL-6 concentration (r = -0.806). It should be noted that in this study the serum concentration of IL-6 decreased 4.4 times with the consumption of bean leaves, which is related to less systemic inflammation [41].
Metabolic endotoxemia increases due to alterations in intestinal integrity that allow bacterial translocation and this has been linked to an increase in low-grade systemic inflammation [53]. Supplementation with microbiota-accessible carbohydrates (15%) for 15 weeks has been reported to improve intestinal eubiosis by decreasing metabolic endotoxemia caused by a high-fat, low-fiber diet in mice [26]. It has also been reported that the inclusion in the diet of foods rich in fiber such as cereals, vegetables and various legumes has been related to the improvement of the intestinal microbiota and the decrease in systemic inflammation with lower concentrations of LPS and IL-6 in circulation [39,54,55]. With the above, the consumption of bean leaves not only reduces the complications of obesity but also reduces intestinal permeability and metabolic endotoxemia.
Finally, the gastrointestinal tract, in addition to digesting and absorbing nutrients, also protects against pathogenic infections and exerts an immune function. An unhealthy diet high in fat and sugar decreases microbial diversity, causing leaky gut and chronic inflammation; On the other hand, a nutritionally balanced diet is essential to maintain a healthy intestinal microbiome, intestinal barrier integrity, immune tolerance and normal intestinal physiology [56].
Until now we demonstrated the role of consuming bean leaves as a treatment for obesity and the decrease in permeability and metabolic endotoxemia. However, in future research the analysis of the intestinal microbiota, its metabolites and the association with the reduction of obesity and inflammation from consumption.
5. Conclusions
Bean leaves, known as “quelites”, are low-cost and easily accessible edible plants that provide fiber and phenolic compounds that can be part of a correct diet. In addition, they can be used to treat obesity and maintain intestinal integrity. The intake of 10% bean leaves helps prevent and treat the accumulation of adipose tissue and the increase in serum triglycerides induced by the consumption of HFFD, by inhibiting the absorption of dietary lipids and protecting from the development of comorbidities. Furthermore, for the first time we report that bean leaf consumption improves intestinal integrity by reducing permeability and metabolic endotoxemia and reducing inflammation. It is necessary to investigate the changes in the intestinal microbiota and microbial metabolites derived from the consumption of bean leaves. As well as, elucidate the mechanism of action regarding the improvement of intestinal integrity, such as the regulation of the secretion of mucins and tight junction proteins. As well as associating microbial metabolites derived from fiber and phenolic compounds with their potential beneficial effects.
Author Contributions
All the authors contribute to the conceptualization, methodology, and writing—review and editing; P.V.O.A. performed formal analysis, visualization, and writing—original draft; P.V.O.A., L.L.V.R., M.R.G. and S.M.A.-S. were in charge of the investigation, project administration, and funding acquisition; N.E. R.G., A.M.O.R., R.R.C., M.R.G., and S.M.A.-S. supervised and validated all the procedures. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by the Fondo de Proyectos Especiales de Rectoría-UAQ (FOPER: 2022-FQU02686) and Fondo para el desarrollo del conocimiento FONDEC 2021 (FNN-2021-05). PVOA was financed by Consejo Nacional de Ciencia y Tecnología (CONAHCYT: CVU 927759).
Institutional Review Board Statement
All procedures performed in animals followed bioethical standards (NOM-062-Z00-1999 and ARRIVE guidelines) and were previously approved by the Ethics Committee of School OF Chemistry, Autonomous University of Queretaro (CBQ22/114, 21 September 2022).
Data Availability Statement
The data will be available by contacting the corresponding author.
Acknowledgments
We are grateful to Candelario Mondragón, Juan Ortiz, Martín García Servín, Alejandra Castilla León, María A. Carbajo Mata, María Eugenia Ramos, Azucena R. Aguilar, Sofía Yolanda Díaz, Adriana González Gallardo, Aurora Jáuregui, Tércia Reis and Alexandro Escobar for their technical support. As well as María de Jesús Guerrero Carrillo and Paola Jazmin Vargas for their support in the histopathological diagnosis.
Conflicts of Interest
The authors declare that there are no conflictS of interest regarding the publication of this paper. The funders had no role in any step of this research.
References
- Kim, S.J.; Kim, S-E.; Kim, A-R.; Kang, S.; Park, M-Y.; Sung, M-K. Dietary fat intake and age modulate the composition of the gut microbiota and colonic inflammation in C57BL/6J mice. BMC Microbiol. 2019; 19, 1–11. [CrossRef]
- Bouwman, M.S.; Nieuwenhuizen, A.G.; Swarts, J.M.; Piga, R.; van Schothorst, E.M.; Keijer, J. Metabolic effects of the dietary monosaccharides fructose, fructose–glucose, or glucose in mice fed a starch-containing moderate high–fat diet. Physiol Rep. 2020; 8, 1–12. [CrossRef]
- Pérez-Campos Mayoral, L.; Mayoral Andrade, G.; Pérez-Campos Mayoral, E.; Hernandez Huerta, T.; Pia Canseco, S.; Rodal Canales, F.J.; Cabrera-Fuentes, H.A.; Martinez Cruz, M.; Pérez Santiago, A.D.; Alpuche, J.J.; Zentero, E.; Martinez Ruiz, H.; Martinez Cruz, R.; Hernandez Jeronimo, J.; Perez-Campos, E. Obesity subtypes, related biomarkers & heterogeneity. Indian J Med Res. 2020;151, 11–21. [CrossRef]
- Agustí, A.; García-Pardo, M.P.; López-Almela, I.; Campillo, I.; Maes, M.; Romani-Perez, M.; Sanz, Y. Interplay between the gut-brain axis, obesity and cognitive function. Front Neurosci. 2018; 12, 1–17. [CrossRef]
- Horne, R.G.; Yu, Y.; Zhang, R.; Abdalqadir, N.; Rossi, L.; Surette, M.; Sherman, P.M.; Adeli, K. High fat-high fructose diet-induced changes in the gut microbiota associated with dyslipidemia in Syrian hamsters. 2020, 12, 1–20. [CrossRef]
- Soares, A.; Beraldi, E.J.; Ferreira, P.E.; Bazotte, R.B.; Buttow, N.C. Intestinal and neuronal myenteric adaptations in the small intestine induced by a high-fat diet in mice. BMC Gastroenterol. 2015; 15, 1-9. [CrossRef]
- Dailey, M.J. Nutrient-induced intestinal adaption and its effect in obesity. Physiol Behav. 2014;0, 74–78. [CrossRef]
- Álvarez, J.; Fernández Real, J.M.; Guarner, F.; Guelmonde, M.; Rodriguez, J.M.; Saens de Pipaon, M.; Sanz, Y. Gut microbes and health. Gastroenterol Hepatol. 2021; 44, 519–535. [CrossRef]
- Organización Mundial de la Salud (OMS). (2018). Dieta saludable. Página web. Recuperado, Junio 2022. Available online: https://www.who.int/es/news-room/fact-sheets/detail/healthy-diet.
- SAGARPA. Planeación agrícola nacional 2017-2030. Frijol mexicano. Secretaría de Agricultura, Ganadería, Desarrollo rural, Pesca y Alimentación. 2017. México. Recuperado junio 2022. Available online: https://docplayer.es/83182578-Planeacion-agricola-nacional-frijol.html.
- Mateos-Maces, L.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Villagómez-González, B.B. Edible leafy plants from Mexico as sources of antioxidant compounds, and their nutritional, nutraceutical and antimicrobial potential: A review. Antioxidants, 2020, 9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Becerril-Campos, A.A.; Ocampo-Anguiano, P. V.; Mondragón-Jacobo, C.; Escobar-García, K.; Camacho-Barrón, M.; Anaya-Loyola, M.A.; Feregrino-Pérez, AA; García-Gasca, G.; Ahumada-Solórzano, S.M. Phaseolus vulgaris L. leaves increase short-chain fatty acid (SCFA) production, ameliorating early metabolic alterations. Plant Foods Hum Nutr. 2022; 77, 421–426. [CrossRef]
- Martinez-Zavala, M.; Mora-Avilés, M.A.; Anaya-Loyola, M.A.; Guzmán-Maldonado, O.; Aguilera-Barreyro, A.; Blanco-Labra, A.; García-Gasca, T. Common bean leaves as a source of dietary iron. Plant Foods Hum Nutr. 2016: 71, 1–6. [CrossRef]
- Becerril-Campos, A.A.; Ramos-Gómez, M.; de los Ríos-Arellano, E.A.; Ocampo-Anguiano, P.V.; Gónzalez-Gallardo, A.; Macotela, Y.; García-Gasca, T.; Ahumada-Solórzano, S.M. Bean leaves ameliorate lipotoxicity in fatty liver disease. Nutrients. 2023; 15, 1–23. [CrossRef]
- Heredia-Ortega, J.; Aguilera, T.; Resis, M.; Bernal, K. Fibra dietética total: métodos de determinación en pastas de oleaginosas. Facultad de Ciencias Naturales UAQ. 2020, 91–102. [Google Scholar]
- Herrera-Rocha, K.M.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Larrosa-Pérez, M.; Moreno-Jiménez, M.R. Phenolic acids and flavonoids in acetonic extract from quince (Cydonia oblonga Mill.): Nutraceuticals with antioxidant and anti-inflammatory potential. Molecules. 2022; 27, 1–14. [CrossRef]
- Baek, S.; Choi, N.H.; Lee, K.P.; Jhun, H.; Kim, J. Smallanthus sonchifolius leaf attenuates neuroinflammation. J Exerc Nutr Biochem 2018, 22, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.; Rosi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic. 1965; 16, 58–144. [CrossRef]
- Campos-Vega, R.; Reynoso-Camacho, R.; Pedraza-Aboytes, G.; Acosta Gallegos, J.A.; Guzmán-Maldonado, S.H.; Paredes López, O.; Oomah, B.D.; Loarca-Piña, G. Chemical composition and in vitro polysaccharide fermentation of different beans (Phaseolus vulgaris L.). J Food Sci. 2009; 74, 59–65. [CrossRef]
- Feregrino-Pérez, A.A.; Berumen, L.C.; García-Alcocer, G.; Guevara-Gonzalez, R.G.; Ramos-Gomez, M.; Reynoso-Camacho, R.; Acosta-Gallegos, J.A.; Loarca-Piña, G. Composition and chemopreventive effect of polysaccharides from common beans (Phaseolus vulgaris L.) on azoxymethane-induced colon cancer. J Agric Food Chem. 2008; 56, 8737–8744. 8744. [CrossRef]
- Dini, I.; Tenore, G.C.; Dini, A. Saponins in ipomoea batatas tubers: Isolation, characterization, quantification and antioxidant properties. Food Chem. 2009; 113, 411–419. [CrossRef]
- Buettner, R.; Schölmerich, J.; Bollheimer, LC. High-fat diets: modeling the metabolic disorders of human obesity in rodents. Obesity 2007, 15, 798–808. [CrossRef]
- Martínez-Orozco, H.; Reyes-Castro, L.A.; Lomas-Soria, C.; Sandoval-Salazar, C.; Ramírez-Emiliano, J.; Díaz-Cintra, S.; Solís-Ortiz, S. High-fat and combined high-fat–high-fructose diets impair episodic-like memory and decrease glutamate and glutamine in the hippocampus of adult mice. Nutr Neurosci 2022, 25, 2479-2489. [CrossRef]
- Kraus, D.; Yang, Q.; Kahn, B. Lipid Extraction from Mouse Feces. BIO-PROTOCOL. 2015; 5, 1–5. 5. [CrossRef]
- Pérez-Ramírez, I.F.; González-Dávalos, M.L.; Mora, O.; Gallegos-Corona, M.A; Reynoso-Camacho, R. Effect of Ocimum sanctum and Crataegus pubescens aqueous extracts on obesity, inflammation, and glucose metabolism. J Funct Foods. 2017; 35, 24–31. [CrossRef]
- Shi, H.; Wang, Q.; Zheng, M.; Hao, S.; Lum, J.S.; Chen, X.; Huang, X.F.; Yu, Y.; Zheng, K. Supplement of microbiota-accessible carbohydrates prevents neuroinflammation and cognitive decline by improving the gut microbiota-brain axis in diet-induced obese mice. J Neuroinflammation. 2020; 17, 1–21. [CrossRef]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain–gut axis alterations. J Physiol. 2018; 596, 4923–4944. [CrossRef]
- Quesenberry KE, Carpenter JW. Rats and mice. In Book Ferrets, rabbits and todents: Clinical medicine and surgery. 4th ed.; Elsevier; United States, 2012, pp. 345–365.
- Martinez, K.B.; Mackert, J.D.; McIntosh, M.K. Chapter 18: Polyphenols and Intestinal Health. Editor: Ronald Ross Watson, Academic Press; United States, 2017, pp. 191–210. [CrossRef]
- Yoon, D.S.; Cho, S.Y.; Yoon, H.J.; Kim, S.R.; Jung, U.J. Protective effects of p-coumaric acid against high-fat diet-induced metabolic dysregulation in mice. Biomed Pharmacother. 2021;142, 1–8. [CrossRef]
- Jihyeon, Y.; Chu-Sook, K.; Thai Hien, T.; Min-Seon, K.; Tsuyoshi, G.; Teruo, K.-, Myug-Sook, C.; Taesun, P.; Mi-Kyung, S.; Jong Won, y.; Suck-Young, C.; Jee Hye, L.; Yeonsoo, J.; Hye-Seon, C.; Sung Hoon, B.; Hun Taeg, C.; Rina, Y. Quercetin protects obesity-induced hypothalamic inflammation by reducing microglia-mediated inflammatory responses via HO-1 induction. Nutrients. 2017; 9, 1–14. Nutrients. [CrossRef]
- Alkhalidy, H.; Moore, W.; Wang, A.; Luo, J.; McMillan, P.; Wang, Y.; Zhen, W.; Hulver, M.T.; Liu, D. Kaempferol ameliorates hyperglycemia through suppressing hepatic gluconeogenesis and enhancing hepatic insulin sensitivity in diet-induced obese mice. J Nutr Biochem. 2018; 58, 1–25. [CrossRef]
- Wang, T.; Wu, Q.; Zhao, T. Preventive effects of kaempferol on high-fat diet-induced obesity complications in C57BL/6 Mice. Biomed Res Int. 2020; 2020, 1–20. [CrossRef]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, W.; Liu, Z.; Ito, K.; Zhang, B. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J Nutr Biochem. 2022; 99, 1–11. [CrossRef]
- Santiago-Saenz, Y.O.; Hernández-Fuentes, A.D.; López-Palestina, C.U.; Garrido-Cauich, J.H.; Alatorre-Cruz, L.M.; Monroy-Torres, R. Nutritional importance and biological activity of compounds from quelites consumed in Mexico. Rev. Chil. Nutr. 2019; 46, 593–605. [CrossRef]
- Sallam, I.E.; Abdelwareth, A.; Attia, H.; Aziz, R.K.; Homsi, M.N.; von Bergen, M.; Farag, M.A. Effect of gut microbiota biotransformation on dietary tannins and human health implications. Microorganisms. 2021; 9, 1-34. [CrossRef]
- Toney, A.M.; Albusharif, M.; Works, D.; Polenz, L.; Schlange, S.; Chaidez, V.; Ramer-Tait, A. E.; Chung, S. Differential effects of whole red raspberry polyphenols and their gut metabolite urolithin a on neuroinflammation in BV-2 microglia. International Journal of Environmental Research and Public Health. 2021; 18, 1–11. [CrossRef]
- Do, M.H.; Lee, E.; Oh, M.J.; Kim, Y.; Park, H.Y. High-glucose or-fructose diet cause changes of the gut microbiota and metabolic disorders in mice without body weight change. Nutrients. 2018; 10, 1–14. [CrossRef]
- Lutsiv, T.; Weir, T.L.; McGinley, J.N.; Neil, E.S.; Wei, Y.; Thompson, H.J. Compositional changes of the high-fat diet-induced gut microbiota upon consumption of common pulses. Nutrients. 2021; 13, 1–20. [CrossRef]
- Moreno-Fernández, S.; Garcés-Rimón, M.; Vera, G.; Astier, J.; Landrier, J.F.; Miguel, M. High fat/high glucose diet induces metabolic syndrome in an experimental rat model. Nutrients. 2018; 10, 1–15. [CrossRef]
- Tan, R.; Dong, H.; Chen, Z.; Jin, M.; Yin, J.; Li, H.; Shi, D.; Shao, Y.; Wang, H.; Chen, T.; Yang, D.; Li, J. Intestinal microbioa mediate high-fructose and high-fat diets to induce chronic intestinal inflammation. Front Cell Infect Microbiol. 2021; 11, 1–14. [CrossRef]
- Shamah-Levy, T.; Vielma-Orozco, E.; Heredia-Hernández, O. Encuesta Nacional de Salud y Nutrición 2018-19 Resultados Nacionales.; 2020. Accessed 4 December 2022. Available online: https://ensanut.insp.mx/encuestas/ensanut2018/doctos/informes/ensanut_2018_informe_final.pdf.
- Bala, C.; Craciun, A.E.; Hancu, N. Updating the concept of metabolically healthy obesity. Acta Endocrinol (Copenh). 2016; 12, 197–205. [CrossRef]
- Zou, Y.; Sheng, G.; Yu, M.; Xie, G. The association between triglycerides and ectopic fat obesity: An inverted U-shaped curve. PLoS One 2020, 15, 1-13. [CrossRef]
- Obonyo, H.R.; Selvi, S.V. Evaluation of hydrated extract of Phaseolus vulgaris L. (bean plant) on hypoglycemia and hypolipidemic in streptozotocin-induced diabetic albino wistar rats. Int J Res Pharm Sci. 2019; 10, 3704–3710. [CrossRef]
- Thompson, H.J.; McGinley, J.N.; Neil, E.S.; Brick, M.A. Beneficial effects of common bean on adiposity and lipid metabolism. Nutrients. 2017; 9, 1–12. [CrossRef]
- Li, N.; Liu, X.; Zhang, J.; Yan-Zhi, L.; Lu, L.; Mi, J.; You-Long, C.; Ya-Mei, Y.; Lin-Wu, R. Preventive effects of anthocyanins from lycium ruthenicum murray in high-fat diet-induced obese mice are related to the regulation of intestinal microbiota and inhibition of pancreatic lipase activity. Molecules. 2022; 27, 1–18. [CrossRef]
- Escobar-Ortiz, A.; Hernández-Saavedra, D.; Lizardi-Mendoza, J.; Pérez-Ramírez, I.F.; Mora-Izaguirre, O.; Ramos-Gómez, M.; Reynoso-Camacho, R. Consumption of cricket (Acheta domesticus) flour decreases insulin resistance and fat accumulation in rats fed with high-fat and -fructose diet. J Food Biochem. 2022; 46, 1–15. [CrossRef]
- Horne, R.G.; Yu, Y.; Zhang, R.; Abdalqadir, N.; Rossi, L.; Surette, M.; Sherma, P.M.; Adeli, K. High fat-high fructose diet-induced changes in the gut microbiota associated with dyslipidemia in Syrian hamsters. Nutrients. 2020; 12, 1–20. [CrossRef]
- Jang, C.; Hui, S.; Lu, W.; Cowan, A.J.; Morscher, R. J.; Lee, G.; Liu, W.; Tesz, G.J.; Birnbaun, M.J.; Rabinowitz, J.D. The small intestine converts dietary fructose into glucose and organic acids. Cell Metab. 2018; 27, 351–361. [CrossRef]
- Lecomte, V.; Kaakoush, N.O.; Maloney, C.A.; Raipuria, M.; Huinao, K.D.; Mitchell, H.M.; Morris, M.J. Changes in gut microbiota in rats fed a high fat diet correlate with obesity-associated metabolic parameters. PLoS One. 2015; 10, 1–22. [CrossRef]
- Costes, L.M.M.; Lindenbergh-Kortleve, D.J.; van Berkel, L.A.; Veenbergen, S.; Raatgeep, H.C.; Simons-Oosterhuis, Y.; van Haaften, D.H.; Karrich, J.J.; Escher, J.C.; Groeneweg, M.; Clausen, B.E.; Cupedo, T.; Samson, J.N. IL-10 signaling prevents gluten-dependent intraepithelial CD4+ cytotoxic T lymphocyte infiltration and epithelial damage in the small intestine. Mucosal Immunol. 2019; 12, 479–490. [CrossRef]
- Ye, X.; Liu, Y.; Hu, J.; Gao, Y.; Ma, Y.; Wen, D. Chlorogenic acid-induced gut microbiota improves metabolic endotoxemia. Front Endocrinol (Lausanne). 2021; 12, 1–22. [CrossRef]
- McGinley, J.N.; Fitzgerald, V.K.; Neil, E.S.; Omerigic, H.M.; Heuberger, A.L.; Weir, T.L.; McGee, R.; Vandemark, G.; Thompson, H.J. Pulse crop effects on gut microbial populations, intestinal function, and adiposity in a mouse model of diet-induced obesity. Nutrients. 2020; 12, 1–17. [CrossRef]
- Wei, B.; Zhong, Q.W.; Ke, S.Z.; Zhou, T.S.; Wu, Q.L.; Wang, S.J.; Chen, J.W.; Zhang, H.W.; Jin, W.H.; Wang, H. Sargassum fusiforme polysaccharides prevent high-fat diet-induced early fasting hypoglycemia and regulate the gut microbiota composition. Mar Drugs. 2020; 18, 1–13. [CrossRef]
- Di, Tommaso, N.; Gasbarrini, A.; Ponziani, F.R. Intestinal barrier in human health and disease. Int J Environ Res Public Health. 2021; 18, 1-23. [CrossRef]
Figure 1.
Effect of bean leaves consumption and change to healthy diet on adipose tissue in mice with induced-obesity. a) Total adipose tissue (images from magnetic resonance analysis); visceral adipose tissue (VAT, red) and subcutaneous adipose tissues (SCAT, blue), b) visceral adipose tissue (VAT) volume, and c) subcutaneous adipose tissue (SCAT) volume. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Figure 1.
Effect of bean leaves consumption and change to healthy diet on adipose tissue in mice with induced-obesity. a) Total adipose tissue (images from magnetic resonance analysis); visceral adipose tissue (VAT, red) and subcutaneous adipose tissues (SCAT, blue), b) visceral adipose tissue (VAT) volume, and c) subcutaneous adipose tissue (SCAT) volume. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
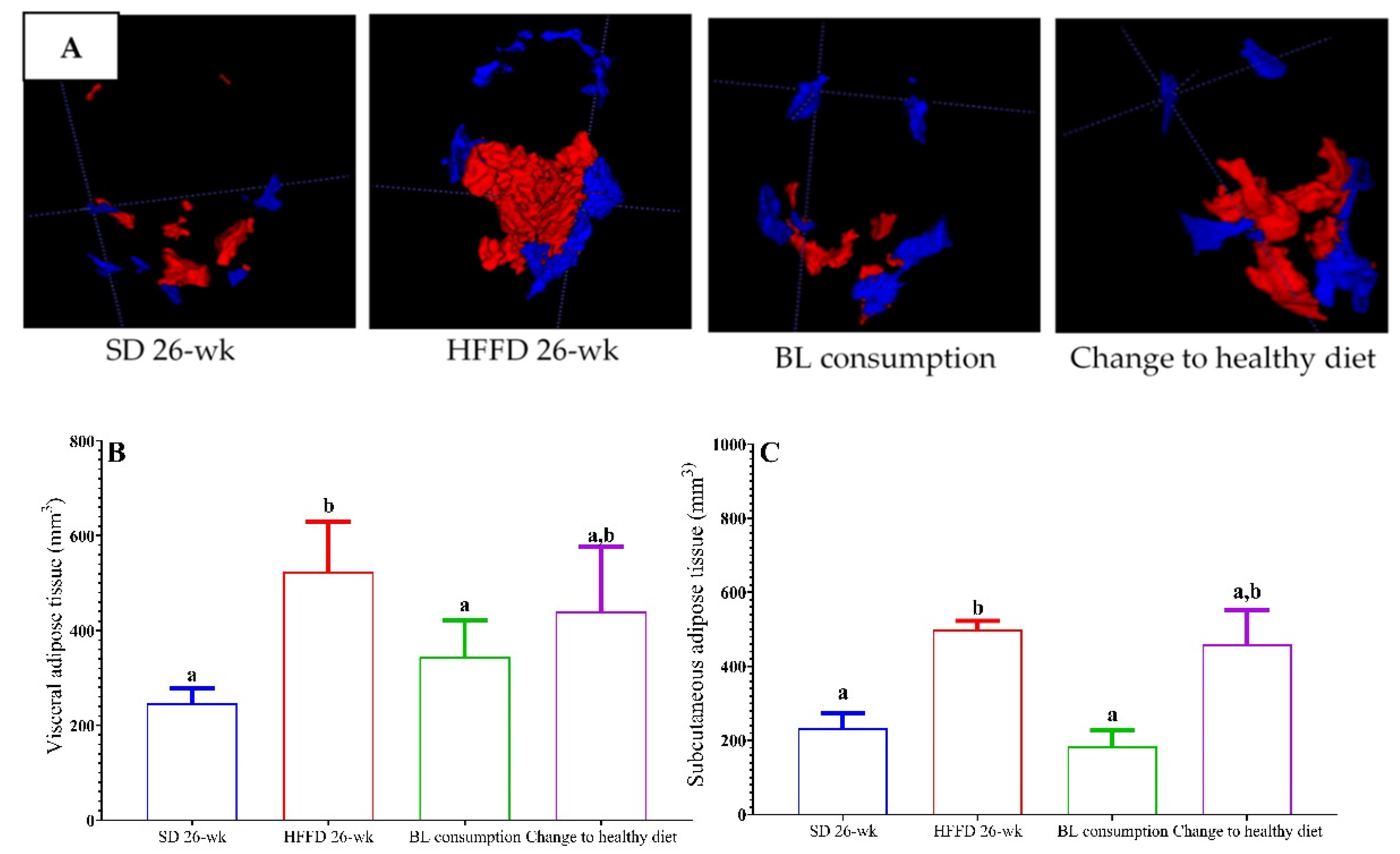
Figure 2.
Effect of bean leaves consumption and change to healthy diet on a) total lipids in feces, b) triglycerides in feces and c) inhibition of pancreatic lipase. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet. Orlistat = positive control of enzyme inhibition. BL extract = bean leaves methanolic extract.
Figure 2.
Effect of bean leaves consumption and change to healthy diet on a) total lipids in feces, b) triglycerides in feces and c) inhibition of pancreatic lipase. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet. Orlistat = positive control of enzyme inhibition. BL extract = bean leaves methanolic extract.
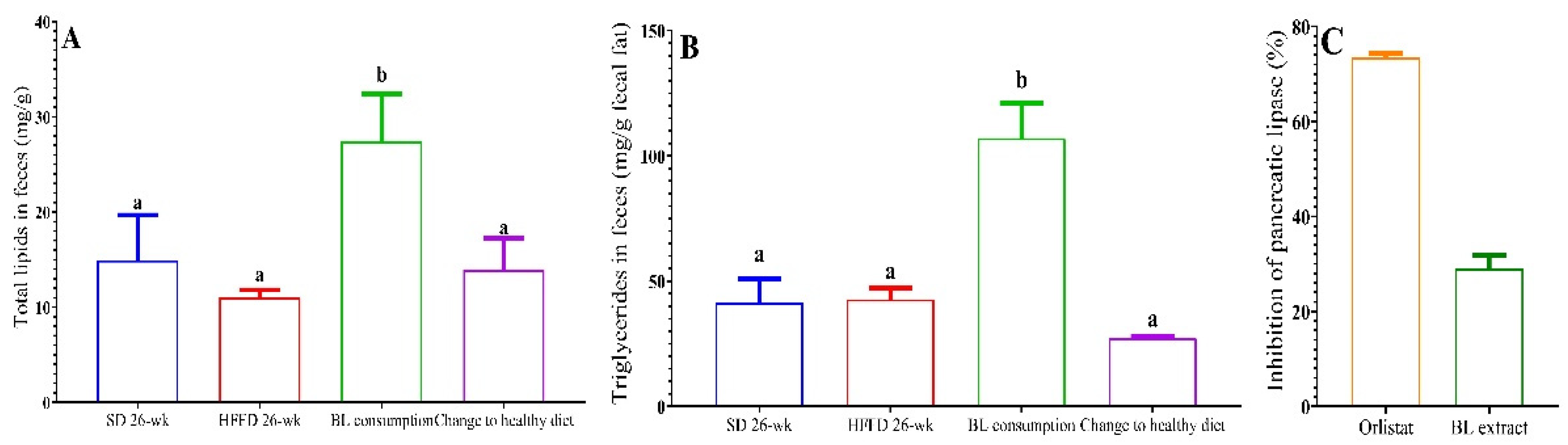
Figure 3.
Effect of bean leaves consumption on the histopathology of jejunum and colon (n = 3-5). Micrographs of colon and jejunum tissue fragment were stained with H&E and analyzed with the 10X objective. BL consumption = HFFD + 10% bean leaves, HFFD = high fat and fructose diet, Change to a healthy diet = consumption of SD in animals with obesity, SD = standard diet. The black arrows indicate the altered structures identified by histopathological analysis.
Figure 3.
Effect of bean leaves consumption on the histopathology of jejunum and colon (n = 3-5). Micrographs of colon and jejunum tissue fragment were stained with H&E and analyzed with the 10X objective. BL consumption = HFFD + 10% bean leaves, HFFD = high fat and fructose diet, Change to a healthy diet = consumption of SD in animals with obesity, SD = standard diet. The black arrows indicate the altered structures identified by histopathological analysis.
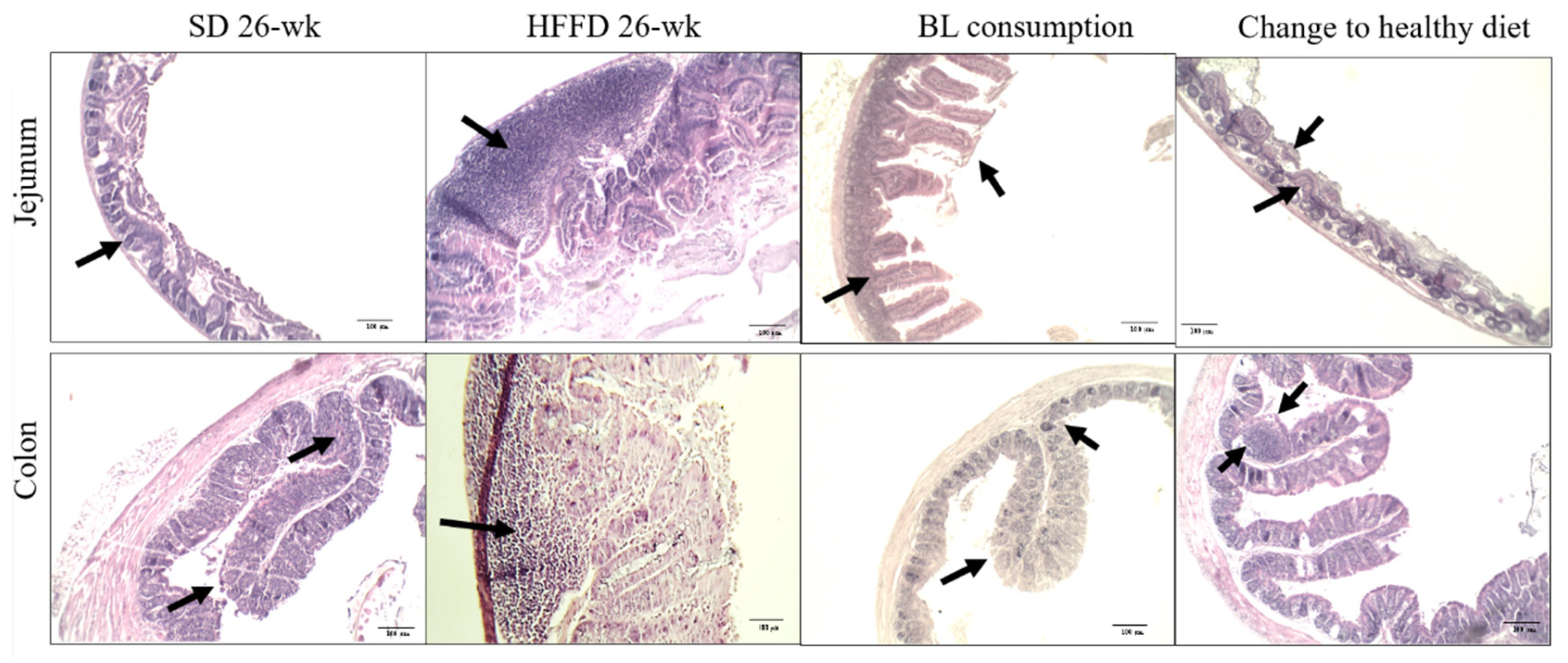
Figure 4.
Effect of bean leaves consumption and change to a healthy diet on intestinal permeability. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Figure 4.
Effect of bean leaves consumption and change to a healthy diet on intestinal permeability. Data are presented as mean values ± SEM (n ≤ 3). Different letters indicate significant differences (p < 0.05). BL consumption = HFFD + bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
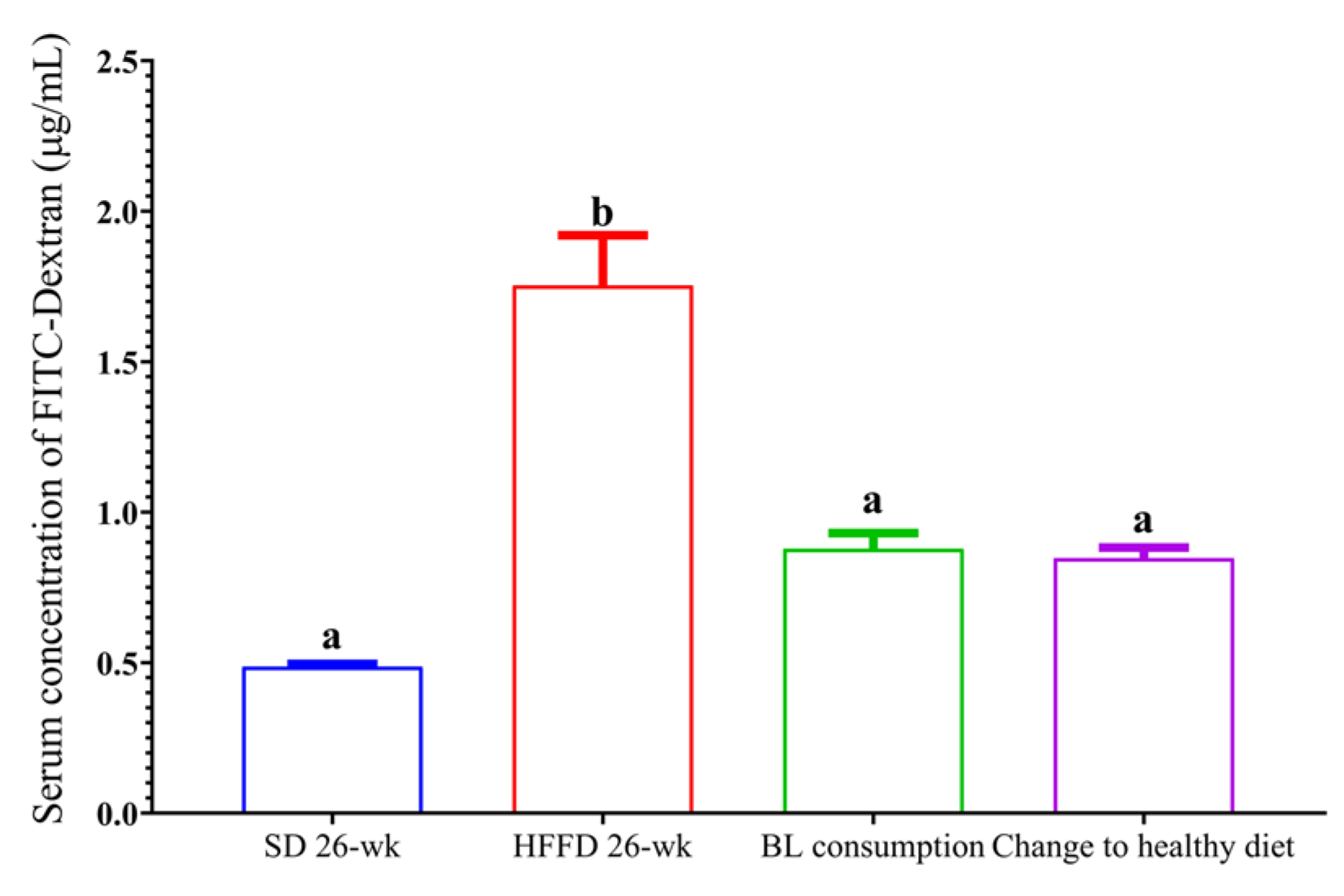
Figure 5.
Effect of bean leaves and on metabolic endotoxemia and inflammation. a) LPS concentration and b) IL-6 concentration. Data are presented as mean values ± SEM (n ≤ 3). Different letters (treatment period) indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Figure 5.
Effect of bean leaves and on metabolic endotoxemia and inflammation. a) LPS concentration and b) IL-6 concentration. Data are presented as mean values ± SEM (n ≤ 3). Different letters (treatment period) indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
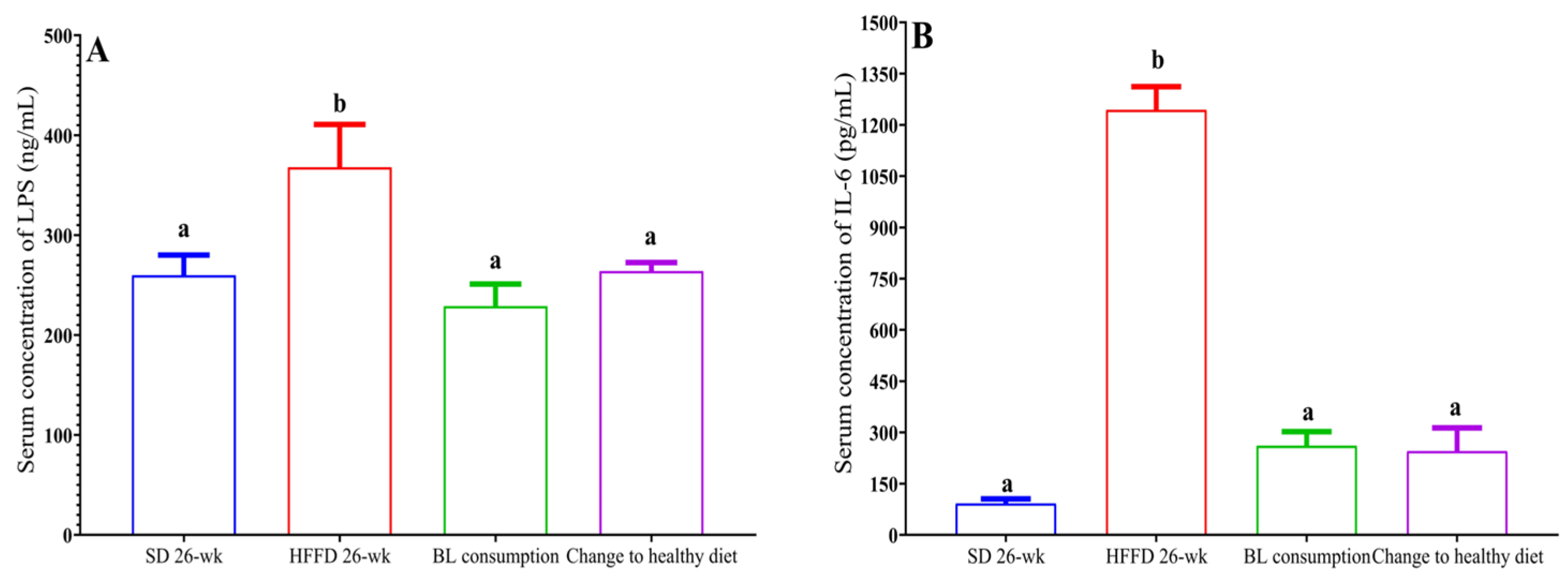

Table 1.
Experimental groups and nutritional content of standard diet (SD) and high fat-high fructose diet (HFFD).
Table 1.
Experimental groups and nutritional content of standard diet (SD) and high fat-high fructose diet (HFFD).
| Energy (kcal/g) |
Proteins (%) | Carbohydrates (%) |
Lipids (%) |
Crude fiber (g /100 g) |
Total dietary fiber (g/100 g) |
Fructose (%) | Lard (%) | Total phenolic compounds (mg/g) | |
|---|---|---|---|---|---|---|---|---|---|
| SD | 3.40 | 28.54 | 58.15 | 13.43 | 5.10 | 36.92 | 0.00 | 0.00 | 2.53 |
| HFFD | 4.70 | 14.28 | 40.90 | 44.81 | 3.07 | 17.31 | 19.98 | 40.17 | 1.55 |
| BL consumption | 4.70 | 13.79 | 41.20 | 45.02 | 3.16 | 21.00 | 20.26 | 40.73 | 3.26 |
Table 2.
Groups and diets of the experimental treatment phase after obesity induction.
| Group | Diet |
|---|---|
| SD 26-wk | Standard diet by 26 weeks |
| HFFD 26-wk | HFFD by 26 weeks |
| BL consumption | HFFD + 10% BL by 12 weeks |
| Change healthy diet | Standard diet by 12 week |
Abbreviations: BL = bean leaves consumption, HFFD = high fat-high fructose diet, SD = standard diet.
Table 3.
Quantification of phenolics and flavonoids in bean leaves by UPLC-ESI-MS/MS.
| Compounds | µg/g |
|---|---|
| Phenolic acids | |
| Quinic acid | 97.44 ± 7.08 |
| Caffeic acid (derivative) | 57.14 ± 3.95 |
| Cinnamic acid (derivative) | 72.41 ± 13.00 |
| Protocatechoic acid | 8.31 ± 2.17 |
| Caftaric acid | 207.21 ± 22.20 |
| 4-hydroxybenzoic acid | 106.59 ± 2.18 |
| Coumaric acid (derivative) | 1283.81 ± 110.87 |
| Ferulic acid (derivative) | 95.53 ± 11.82 |
| 2-hydroxybenzoic acid | 96.76 ± 10.79 |
| Benzoic acid | 49.55 ± 0.54 |
| Flavonoids | |
| Rutin | 971.18 ± 73.19 |
| Quercetin 3-O-ß-glucuronide | 6892.20 ± 99.42 |
| Quercetin 3-O-glucoside | 73.05 ± 7.53 |
| Kaempferol 3-O-glucoside | 11.24 ± 0.10 |
| Naringin | 1.01 ± 0.36 |
| Naringenin | 2.29 ± 0.61 |
Table 4.
Influence of the addition of bean leaves and the change to a healthy diet on food and water consumption in mice with induced obesity.
Table 4.
Influence of the addition of bean leaves and the change to a healthy diet on food and water consumption in mice with induced obesity.
| Obesity induction phase | Experimental treatment phase | |||||
|---|---|---|---|---|---|---|
| SD 14 wk | HFFD 14 wk | SD 26 wk | HFFD 26 wk | BL consumption | Change to healthy diet | |
| Food intake (g/day) |
4.59 ± 0.50 | 4.88 ± 0.30 | 3.79 ± 0.40a | 5.13 ± 0.80a | 4.54 ± 0.60a | 5.09 ± 0.50a |
| Energy intake (Kcal/day) |
15.75 ± 1.50 | 22.94 ± 1.30* | 12.89 ± 1.50a | 23.48 ± 3.80b | 20.80 ± 2.70b | 17.11 ± 1.50a |
| Water intake (mL/day) |
4.62 ± 0.50 | 4.92 ± 0.40 | 5.05 ± 0.40a | 5.61 ± 0.50a | 4.90 ± 0.70a | 6.10 ± 0.80a |
Data are presented as mean values ± SEM (n ≤ 6), p < 0.05. * SD vs HFFD (induced obesity period). Different letters in the same row (treatment period) indicate significant differences. BL consumption = HFFD + 10% bean leaves, Change of healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Table 5.
Effect of bean leaves consumption and change to a healthy diet on body parameters and lipid profile of mice with induced-obesity.
Table 5.
Effect of bean leaves consumption and change to a healthy diet on body parameters and lipid profile of mice with induced-obesity.
| Obesity induction phase | Experimental treatment phase | |||||
|---|---|---|---|---|---|---|
| SD 14-wk | HFFD 14-wk | SD 26-wk | HFFD 26-wk | BL consumption | Change to healthy diet | |
| Body weight (g) | 25.63 ± 0.57 | 33.47 ± 0.72 * | 27.96 ± 0.65a | 30.50 ± 0.52b | 28.09 ± 0.84ab | 29.47 ± 0.42a |
| Abdominal circumference (cm) | 7.50 ± 0.46 | 10.40 ± 0.43* | 7.79 ± 0.29a | 8.33 ± 0.28a | 8.50 ± 0.39a | 6.83 ± 0.17b |
| Thoracic circumference (cm) | 7.13 ± 0.43 | 9.2 ± 0.73 | 7.36 ± 0.26ª | 8.17 ± 0.25ª | 8.50 ± 0.55ª | 7.33 ± 0.33ª |
| Serum cholesterol (mg/dL) | 92.60 ± 8.60 | 122.80 ± 11.90* | 98.30 ± 0.90a | 119.70 ± 3.40b | 114.40 ± 3.50b | 104.90 ± 4.20b |
| Serum triglycerides (mg/dL) | 77.70 ± 7.60 | 137.30 ± 17.50* | 99.50 ± 3.90a | 141.10 ± 4.20b | 110.70 ± 7.30a | 106.80 ± 7.70a |
Data are presented as mean values ± SEM (n ≤ 6). * SD vs HFFD (induced obesity period). Different letters in the same row (treatment period) indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Table 6.
Effect of bean leaves consumption and change to a healthy diet on small intestine and colon length as an indicator of intestinal inflammation.
Table 6.
Effect of bean leaves consumption and change to a healthy diet on small intestine and colon length as an indicator of intestinal inflammation.
| Obesity induction phase | Experimental treatment phase | ||||||
|---|---|---|---|---|---|---|---|
| Length (cm) | SD 14-wk | HFFD 14-wk | SD 26-wk | HFFD 26-wk | BL consumption | Change to healthy diet | |
| Small intestine | 44.00 ± 2.91 | 36.63 ± 1.58* | 39.85 ± 1.72a | 34.31 ± 0.97a | 37.95 ± 1.04a | 37.24 ± 1.67a | |
| Total colon | 8.86 ± 0.50 | 6.98 ± 0.50 | 7.28 ± 0.45a | 6.58 ± 0.52a | 7.41±0.48a | 5.43 ± 0.53b | |
Data are presented as mean values ± SEM (n ≤ 6). * SD vs HFFD (induced obesity period, p < 0.05). Different letters (treatment period) indicate significant differences (p < 0.05).BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Table 7.
Effect of bean leaves consumption and change to a healthy diet on the relative organ weight after treatment.
Table 7.
Effect of bean leaves consumption and change to a healthy diet on the relative organ weight after treatment.
| Percentage (%) | SD 26-wk | HFFD 26-wk | BL consumption | Change to healthy diet |
|---|---|---|---|---|
| Colon | 1.82 ± 0.06a | 1.23 ± 0.03b | 1.24 ± 0.02b | 1.31 ± 0.09b |
| Small intestine | 7.06 ± 0.28a | 5.77 ± 0.27b | 6.25 ± 0.14ab | 6.09 ± 0.14b |
| Liver | 4.81 ± 0.14a | 5.93 ± 0.28ab | 5.58 ± 0.59a | 4.68 ± 0.21ac |
Data are presented as mean values ± SEM (n ≤ 6). * SD vs HFFD (induced obesity period, p < 0.05). Different letters (treatment period) indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change TO A healthy diet = SD after obesity-induced phase, HFFD = high fat and fructose diet, SD = standard diet.
Table 8.
The consumption of bean leaves and the change to a healthy diet increases the fiber intake through the diet.
Table 8.
The consumption of bean leaves and the change to a healthy diet increases the fiber intake through the diet.
| Intake (g/day) | SD 26-wk | HFFD 26-wk | BL consumption | Change to healthy diet |
|---|---|---|---|---|
| Soluble fiber | 0.21 ± 0.02a | 0.08 ± 0.01b | 0.24 ± 0.03a | 0.33 ± 0.04a |
| Insoluble fiber | 0.81 ± 0.08a | 0.23 ± 0.02b | 0.70 ± 0.09a | 1.08 ± 0.13a |
| Total dietary fiber | 1.20 ± 0.10a | 0.27 ± 0.02b | 0.94 ± 0.02c | 1.40 ± 0.16d |
| Total phenolic compounds | 9.59 ± 1.02a | 7.95 ± 1.24a | 14.8 ± 2.72b | 12.87 ± 1.27b |
Data are presented as mean values ± SEM (n ≤ 6). * SD vs HFFD (induced obesity period, p < 0.05). Different letters (treatment period) indicate significant differences (p < 0.05). BL consumption = HFFD + 10% bean leaves, Change to a healthy diet = SD after obesity induction phase, HFFD = high fat and fructose diet, SD = standard diet.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



