Submitted:
18 December 2023
Posted:
19 December 2023
You are already at the latest version
Abstract
The lengthy maturation period is the key cause of the reduced hot pepper market value. An experiment was performed to assess the effects of R/B/W light qualities on reducing the maturation period. The maturation period was shortened by 16 days under 4R:1B:5W compared to the control. The appearance of the first flower is a limiting factor affecting the maturation period. It exhibited a dark purple color and an abnormal petal under 4R:1B:5W compared to the control, which was associated with increased contents of flavonoids, total organic acids, and phenolics and decreased total amino acid and carbohydrate contents. Plant hormone, flavonoid, ABC transporters, key metabolites (such as amino acid, carbohydrate, organic acid) was associated with first flower development. Ethylene response factors (ERFs) mediated by light, especially ERF021, were identified as potential hub regulators during the development of the first flower due to their involvement in all the associated metabolic pathways.
Keywords:
hot peppe
; light quality
; metabolom
; maturation period
; transcriptome
1. Introduction
Hot pepper is one of the most vegetables consumed worldwide, while their lengthy growth period has been considered the primary cause of pepper commercial value loss. The maturity period in hot peppers usually ranges from 60 days to 90 days, and is highly affected by the genotype of the cultivars and the environment[1]. Full fruit coloration and mature seeds are the key phenotypic signs of fruit maturity, and their delayed occurrence has been the principal factor resulting in the reduced market price of hot peppers. Promoting earlier flowering and fruit coloration are critical steps to accelerate maturation and enhance pepper marketability. Therefore, a strategy should be performed to alter the flowering behavior and fruit set in hot pepper. If hot pepper is grown in a controlled environment growth chamber (such as the artificial lighting climate rooms used on an industrial scale) but not in natural conditions, the period of fruit maturation will be shortened. Therefore, exploring optimal growth conditions to accelerate hot pepper maturation is key.
Light is the primary factor affecting the plants' metabolites synthesis and morphogenesis, and these effects result in changes in plant growth and development, including flowering, fruit coloration, and seed maturity[2]. In terms of physiological and biochemical changes, light intensity and quality parameters were shown to affect carbohydrate accumulation in plants cells[3] and to alter the levels of endogenous hormones[4]. Light quality (wavelength) is the most significant factor affecting these changes, and different light qualities were shown to regulate the nutrient assimilation in plants, thereby, in turn, affecting reproductive development. Results from Solanaceae plants grown under different R: B ratios showed that modifying the R: B spectral distribution is crucial for flowering and fruit-setting attributes[5]. A recent study in pepper demonstrated that R- and B-light directly affected plant growth, photosynthetic characteristics and flowering time, with R light promoting pepper fruit coloration through increased pigment accumulation[6,7]. These evidences indicate that comprehensive research on fine tuning the light quantity and quality properties is necessary to meet the light requirements of the plants more accurately and maximize their productivity and quality.
Optimal light qualities regulating biological processes can be achieved through light-responsive photoreceptors-mediated signaling pathways. These photoreceptors, such as phytochromes, cryptochromes, and phototropins, can absorb both red and blue light and transmit the corresponding signals to downstream metabolic pathways, thereby modulating gene and transcription factor expression involved in hormone signaling and circadian rhythms. For example, PHYA and PHYB, the primary phytochromes responsible for perceiving and mediating responses to far-red (FR) and R light signals, can interact with downstream signaling intermediates (such as PIFs and HY5) and then control the expression of target genes, such as FT and CO, thereby promoting flowering[8,9]. Similarly, cryptochromes (CRYs) operating as blue-light photoreceptors, and their flavin adenine dinucleotide (FAD) cofactor, which allows cryptochromes and photolyases to absorb blue light, further stimulated the transcription of Phot1, Phot2, COP1, DFR1, playing key roles in pigment accumulation, inhibition of hypocotyl elongation, fruit coloration and circadian rhythm regulation [10,11]. These experimental findings confirmed the critical contribution of light quality parameters in biological process, however, the precise regulatory mechanisms underlying these processes have not been investigated in depth in hot peppers.
Therefore, it remains challenging to fully understand through which mechanisms light quality shortens the fruit maturity period in hot pepper. Although the integrative analysis of the phenotype, physiology, transcriptome, and metabolome provides an effective way to contribute to the identification of functional genes and elucidate pathways involved in fruit maturity, targeted or untargeted metabolomics studies have not been conducted in hot pepper. In this study, we performed a combined transcriptomics-metabonomic analysis, combined with phenotypic, physiological and quantitative gene expression analyses, to investigate of the mechanisms underlying the accelerating maturity process of hot pepper under different LED light qualities. The results of this study will help shorten the maturity period and provide the basis for speed- breeding-based programs in hot pepper.
2. Results
2.1. Fruit coloring in hot pepper under different LED light qualities
Fruit development of hot pepper was divided into three distinct stages based on fruit coloration characteristics: the initial stage, the coloration stage, and the maturity stage. The fruit color was purple at the initial stage, and as the fruit maturation progressed, it turned to white, yellow, and orange, and then at the fruit coloration stage to orange-red, and eventually at the maturity stage, the fruits were fully turned red (Figure 1A). Discrepant fruit coloration was observed under different LED light qualities (Figure 1B and Table S4). The percentage of red fruits (PRF) per hot pepper plant was significantly increased in the T1 (13.77 %) and T2 (14.48 %) treatment groups, compared to the control (3.85 %). The total percentage of orange, orange-red and red fruits was over 25.0 % in the T2 group. In contrast, the fruit color features in the T3 and T4 groups showed that the fruit coloration process was delayed by the excessive LED light wavelength ratios and was significantly different compared to the T2 group. The PRFs were only 5.93 % and 4.76 % in the T3 and T4 groups, which were nearly equal to that in the control treatment (3.85 %). Further, some parameters involved in fruit ripening such as total sugar, total protein and fruit firmness was determined to confirm the ripening process. As shown in Figure 1CD, along with fruit ripening, total protein and sugar content were increased and T2 group possessed the highest content than other groups and control. Contrary, fruit firmness began to decrease during fruit ripening, and at the maturity stage (red fruit), the lowest firmness was observed in T2 group. Further, some parameters involved in fruit ripening such as total sugar, total protein and fruit firmness was determined to confirm the ripening process. As shown in Figure 1CD, along with fruit ripening, total protein and sugar content were increased and T2 group possessed the highest content than other groups and control. Contrary, fruit firmness began to decrease during fruit ripening, and at the maturity stage (red fruit), the lowest firmness was observed in T2 group. These results suggested that T2 group promotes fruit coloring and maturity.
2.2. Changes in fruit maturity period under LED light qualities
In control, the time to commercial fruit maturity was at least 80 days (88 days), while this time was significantly changed when hot pepper plants were grown under different LED light qualities. This effect was more pronounced in the S1 (the stage from seed to first flower), S4 (the stage from fruit set to fruit coloration), and S5 (the stage from fruit coloration to commercial fruit maturity) stages of hot pepper growth (Figure 2A). In the T2 treatment group the maturity period was 72 days, with the duration of growth stages S1, S4, and S5 being only 29 days, 17 days and 8 days respectively, significantly shorter compared to the control treatment. Similar patterns were also observed in the T1 group (a maturity period of 74 days), despite the duration of the S1 stage not being different compared to the control. Unexpectedly, the T3 and T4 treatments did not effectively shorten the fruit maturity period, which was 86 days and 88 days, respectively. An extension of the S1 and S4 stage duration in these two treatments was the key cause of this result. The S1 stage is the longest compared to other stages across the control and all the treatment groups, indicating that the S1 stage may be a critical contributor to the fruit maturity period.
The application of different LED light qualities had no apparent effects on seed coloration during fruit maturity (which ranged from white to pale yellow). In contrast, the seed number per fruit was affected by LED light qualities. Although yield per hot pepper plant and weight per fruit was significantly higher in the T2 treatment group compared to the control (Figure 2CD), the seed number per fruit was reduced, and no significant differences were observed in the thousand-seed weight (Figure 2AB). In the other treatments, these parameters, including thousand-seed weight, yield per plant, and weight per fruit, were almost equal or even lower compared to the control. Therefore, it was confirmed that LED light qualities could affect seed development.
Interestingly, at the yellow fruit stage, the control plants produced seeds that could not germinate, while a 12.3 % germination percentage was observed in the seeds when the hot pepper fruits reached the orange color stage. In contrast, the seed could germinate in the yellow fruit stage of the T1, T2, and T3 groups, and the percentage of seed germination in the T3 group reached over 34.0 % (Table S2). This suggested that pepper seeds can be harvested before the full fruit maturity for speed-breeding and molecular breeding practices. Furthermore, the germination percentage of seeds red fruits in the T2 treatment group reached 84.1 %, nearly equal to that in the control, at the same fruit maturity stage. On the other hand, deed germination in the other light quality treatments was lower than that in the control. Seeds from purple and white-colored fruits from all treatments could not germinate. Therefore, in combination with the results presented in Figure 2CD, we considered the T2 treatment the best LED light quality regime for fruit coloration and seed maturation and germination.
2.3. Change of metabolites under LED light qualities
As mentioned above, the S1 stage was the key stage affecting the total pepper growth period, and flower bud differentiation is usually completed at the S1 stage. Therefore, the metabolites in the first flowers of hot pepper plants from the Control, T2 (light quality with the best results), and T4 (light quality with the worst results) treatments were quantified. As shown in Figure 3A, the T2 and T4 treatments resulted in dark purple flower phenotypes, while the flower color in control was purple-white. A fully open corolla was observed in the control and T2 treatments but not in T4. Further, the total metabolites in the first flower of hot pepper plants were quantified. The base chromatograms depicting the metabolite peaks are shown in Figure S1. The samples from the treatments were clearly separated according to their metabolite profiles under the different LED light qualities across PC1 (explaining 23.6 % of the total variance), whereas PC2 mostly discerned within treatment variation (explaining 18 % of the total variance). The analysis of the PCA score plot revealed that the T2 and T4 treatments were clearly distinguished from the control (Figure 3B).
Venn diagram analysis revealed that 105 DEMs were shared jointly by the control, T2, and T4 treatments, and 20 and 25 DEMs were unique to the T2 and T4 treatments, respectively (Figure 3C). Among these DEMs, vitexin, a flavonoid compound, maybe a significant metabolite responsible for the differences between treatments, due to its increased Z-core observed in the T2 and T4 treatment when compared to the control (Figure 3D).
Correlation analyses (>0.9) revealed a positive relationship between vitexin and flavonoids, organic acids, and phenolics. At the same time, a negative correlation was observed between carbohydrates and the total free amino acids (Table S5). Further analysis confirmed that flavonoids were involved in hot pepper flowering due to their higher content observed in the T2 and T4 treatments than that in the control. Moreover, the T2 treatment had more elevated levels of ODs than the T4 group. Similar results were also observed in the organic acid and phenolics content (Figure 4A). The total phenolics content in the T2 treatment was higher than that in the control and T4 treatment. In contrast, no statistically significant differences were observed between the control and T4, even though the content in T4 was lower than that in the control. In addition, the content of total free amino acids and carbohydrates in the T2 and T4 treatments was decreased compared to the control, with the T4 treatment resulting in lower contents compared to the T2 treatment (Figure 4A). Vitexin was only one of the differential metabolites, as network pathway analysis based on KEGG enrichment (shown in the green box of Figure 4B) revealed that plant hormones located on the network hub positions might stimulate or inhibit the synthesis of metabolites such as flavonoids, phenylpropane, linoleic acid, sucrose, etc. (Figure 4B and Figure S2). Light qualities may induce plant hormones pathway mediated by 27 DEMs from photosynthesis system, including 12 members of Psb family photosynthesis II, 10 members of Psa family from photosynthesis I, 2 members of Pet family from cytochrome b6/f complex, 3 members from photosynthetic electron transport and 3 members from Fe-type ATPase, and 12 DEMs from light-harvesting chlorophyll protein complex (Figure 4B and Figure S4). Therefore, it was speculated that plant hormones mediated by these DEMs from photosynthesis system may play a key role in the flower development of hot pepper.
2.4. Changes in DEGs under different LED light qualities
In order to confirm the deduction, a transcriptome analysis was conducted and showed an obvious separation among control, T2 and T4 groups according to 98 % PCA score plot (PC1 31 % and PC2 67 %) (Figure S4). The rigorous quality estimation and data cleaning resulted in 64.61-67.76 M clean reads and their Q20 and Q30 ranges were 98.42%~98.5% and 95.1%~95.3%, respectively. Moreover, the N percentage was 0.00% in all samples (Table S6).
A total of 3043, 4099 and 5032 DEGs were found in the comparisons of the Control vs T2, Control vs T4, and T2 vs T4 groups, respectively, and 456 DEGs were shared by the three comparisons (Figure 5A and Table S7). GO terms reflected that TOP 10 of these DEGs are annotated into 30 functional subdivisions in three major categories including cellular components (CC), biological processes (BP) and molecular functions (MF). Endomembrane system (GO:0012505), catalytic activity (GO:0140096) and carbohydrate metabolic process (GO:0005975) were significantly changed in the first flowers of hot pepper under different LED light qualities (Figure 5B). KEGG analysis indicated that these DEGs involved in plant hormone signaling transduction was enriched significantly as comparison to other ones (Figure 5C). Similar patterns were also observed in flavonoid synthesis, phenylpropanoid synthesis, starch and sucrose metabolism, endocytosis, MAPK signaling pathway, galactose metabolism, pentose and glucuronate interconversions, alpha-linolenic acid metabolism, animo sugar and nucleotide metabolism. These results revealed that there was difference in development of the first flower between control and treatment groups and a complex biological process was observed during flowering.
2.5. Integrative analysis of DEGs and DEGs and qRT-PCR
DEGs and DEGs related to biosynthesis of plant secondary metabolites, flavonoid biosynthesis, biosynthesis of phenylpropanoids, ABC transporters and biosynthesis of plant hormones were enriched (Figure 6). These DEGs associated with One-to-one correspondence DEGs were listed in Table 1. Especially, only one key gene from ERF family was identified among all ABC transporters. Further, we screened key genes from these DEGs to confirm their expression by qRT-PCR. 9 DEGs were found in Control vs T2 and Control vs T4 groups respectively, and almost all target DEGs was closely related to ethylene-responsive transcription factors (ERFs) (Table S8). This also was confirmed by the result of Figure 4D, which showed that enriched number of DEGs of ERFs were most among all transcriptional factors. qPCR analysis revealed that ERF021 gene possessed the highest transcriptional levels among all detected and represent key genes, and expression of these genes, including ERF021, FAD2, ERF1B, ERF026, TM9SF7, ERF091 ERF012, TM9SF2 and ERF110, in T2 group was higher than that in control and T4 group. Pattern in expression of these represented genes including ACC, SAM and ATR1, was similar with that of the aboved detected genes (Figure 6A). Correspondingly, improved expression of genes involved in flavonoid pathway in T2 group also was observed in T4 group, and F3H expression in the T2 group even reached over 500 (Figure 7B), suggesting a positive relationship between flavonoid and ERFs expression. Furthermore, some DEGs encoding photosynthesis related metabolites were detected and found that improved expression of these genes, especially COP1, FT, CRR1 genes and gene encoding photosystem II polypeptide, was observed in T2 group than that of the control, while that in T4 group is lower when compared with that in the control (Figure 7C). There was significance in expression of genes related amino acid and sugar metabolism among control, T2 and T4 groups, and these genes, such as CAAT7, COP1, AVT16, NEC1, STC, STP8 ect., showed the decreased expression in T2 and T4 group as comparison to the control, but that in T4 group is lower than that in T2 group (Fic. 7D). The results suggested that ERFs expression showed the positively relationship to synthesis of flavonoid and photosynthesis but negatively one to amino acid and carbohydrate under LED light qualities in hot pepper.
3. Discussion
Hot pepper originates from tropical regions with sufficient sunlight, and their fruit maturation varies depending on the duration of the growth stages and environmental conditions (such as the light spectrum) [12]. Uniform and full fruit coloration is an important criterion that reflects fruit maturation and is a key target for breeders because bright and fully colored fruits are one of the key traits attracting consumers' attentio This study investigated the e=ffect of different LED light ratios on hot pepper fruit coloration and found that 4R:1B:5W LED light ratios (T2 group) significantly accelerated the process of fruit coloration. This is potentially associated with the blue light component due to the positive effects of blue light on carotenoid accumulation resulting in color formation[13]. This is due to the fact that the light, after being absorbed by the chloroplasts in the early stages of ripening, activates photoreceptors in fruits, thereby allowing a gradual synthesis of carotenoids in the peel and pulp with the onset of chromoplasts' development. However, excessive red light leads to an overall carotenoid content reduction[14]. In the present study, the decreased PRFs observed in the T4 treatment group with 7R:1B:2W LED light ratios support the viewpoint. In addition, the yield per hot pepper plant and weight per fruit was increased in the T2 treatment group compared to the control, which is critical for the commercial value of hot peppers due to the significance of weight or yield increase in enhancing fruit quality and lengthening the hot pepper shelf life.
Hot pepper consumption has prominent periodic seasonal fluctuation characteristics; therefore, an important challenge in the hot pepper industry is how to shorten the fruit maturity period to enhance its economic value and market competitiveness. Regulation of LED light quality through variable wavelength ratios provides an effective way to achieve these objectives. In the current study, the time of full fruit ripening was shortened by at least 16 days in the T2 treatment compared to the control, a finding that, to our knowledge, is reported for the first time in the literature. This may be related to the heightened expression of CO and FT genes during flowering. Flowering, especially the appearance of the first flower, is the key sign of reproductive growth and is strongly induced by specific wavelengths of the light spectrum[15,16]. Red light induces the differentiation of flower buds and further promotes flowering through the interaction between the red light photoreceptor PHYA and the CO protein, which positively regulate FT expression[17]. Blue light also is an important signal that regulates flowering. The expression levels of genes that are potentially involved in light signaling (PhyB, PIFs, and HY5) and circadian rhythms (LHY and CO) in plants were altered under blue light [18]. However, we did not evaluate the monochromatic light of full R or full B in our experiments. Still, the combination of R/B/W LED light spectra is the most optimal for hot pepper because plant growth and secondary metabolism are increased under a full light spectrum[19], ensuring normal plant growth.
The improved yield and fruit weight in the T2 treatment group confirm the critical role of combining different light qualities in fruit development. In addition, seed germination could be achieved even at earlier fruit maturation stages, when the fruit skin turns yellow color. At the same time, this phenomenon did not occur in control, which indicates that seeds can be harvested earlier, shortening the crop life cycle and enabling faster breeding. Early seed harvest is particularly important for reducing the breeding program length and accelerating the generation of new plant cultivars[20].
The phenomenon of petal color change during the flowering process is uncommon in nature. Anthocyanins, a major flavonoid compound class, play key roles in determining flower coloration[21]. Naturally, the flowers of cultivated pepper are usually white with little pigmentation, which restricts the research on the role of flavonoids in pepper flower coloration. As we used a hot pepper cultivar with purple flowers as test material in the present study, we found that the LED light ratios promoted flavonoid accumulation in the flowers. Certain DEGs or DEMs related to flavonoid synthesis were enriched significantly and the changes in petal color were accompanied by an increased expression of anthocyanin-related genes such as F3H, ANS, CHS, MYB, etc. when hot pepper plants were grown under 4R:1B:5W (T2 treatment). This finding suggests that light qualities can induce the biosynthesis of flavonoids in the flowers of hot pepper plants. The study by [22], which reported that red and blue light increased anthocyanin content and the expression of the relevant genes, supports these findings. However, the key upstream regulator of flavonoid accumulation induced by specific LED light spectra during the flower development of hot pepper remains to be identified, and further studies should be performed in the future.
The LED light ratios evaluated in the T2 treatment upregulated the CO, FT, and FLS gene expression compared to the control. In contrast, the T4 treatment had a contrasting effect resulting in their down-regulation. These results were associated with our findings that the S1 stage was prolonged in the T4 treatment but was shortened in the T2 treatment. This may be the key cause resulting in the altered fruit maturation period. Flower bud differentiation regulated by R/B light has been recently studied[23]. Higher blue light and lower red light intensities promote early and extended flowering, while higher red light and lower blue light intensities delay flowering[15]. Therefore, blue light may be more important for flowering initiation, as it controls the flowering time distinctively via CO and FT [24]. On the other hand, excess red light (monochromatic red light) was the most effective in inhibiting flowering [24]. This was potentially the cause underlying the S1 stage prolongation in the T4 treatment compared to the control. In addition, different wavelengths of supplemental blue light accelerate the flowering of plants early via promoting ethylene production, also suggesting the key role of blue light in early flowering[25].
Notably, the petals in the T4 treatment exhibited an abnormal opening, which was potentially affected by decreased contents of metabolites such as carbohydrates, total amino acids, and phenolics and low expression levels of genes involved in these pathways. Plants use distinct photoreceptors (such as PhyA and PhyB) to sense different wavelengths. PhyA and PhyB signaling has essential roles in the control of primary metabolism in response to light, including the sugar content and other metabolites such as amino acids, polyamines, and metabolites of the tricarboxylic cycle[26]. Our results divulged that abnormal flower development caused by excess red light resulted in the reduction of these metabolites in hot pepper flowers.
KEGG enrichment analysis demonstrated the significance of the ethylene signaling pathway in flowering, and ERF TFs were shown to have a core role in the development of flowers, according to the combined transcriptome-metabolome results. ERFs have been shown to regulate anthocyanin biosynthesis [27], and 175 members of the ERF family have been annotated in the pepper genome[28]. Blue light treatment led to an induction of the ethylene signaling pathway and resulted in the promotion of ethylene biosynthesis by the regulation of transcript levels of ERF TFs [29]. Interactions between these ERFs and MYB TFs enhance the stable bindings between MYBs and bHLHs, which is the primary cause of promoting the accumulation of blue light-induced flavonoids[30]. There is limited research confirming the role of red light in ERF expression regulation, but excess red light certainly caused decreased expression of ERFs genes based on our results (Figure 4A). The current study has demonstrated that the expression changes of ERFs were correlated with the changes in flavonoid content and the expression of genes, such as MYBs and bHLHs, related to anthocyanin synthesis. These findings indicate that the change of anthocyanin content under different light ratios is associated with ERF TF expression, with blue light being the main factor affecting anthocyanin accumulation.
The phenolics and organic acid content also showed a similar trend to that of the flavonoid content in this study. Phenolics and flavonoid biosynthesis are commonly initiated from the phenylalanine metabolic pathway. Genes upstream of the pathway, such as phenylalanine ammonia-lyase (PAL), are responsible for increased flavonoid and phenolics content[31]. It should be noted that a positive relationship between the expression levels of ERF and PAL has been identified[32], and recent studies have also shown that the biosynthesis of phenolics is positively regulated by the several ERF TFs, such as ERF115[33]. A potential conclusion is that the increase in phenolics content is associated with PAL expression mediated by ERFs. Furthermore, ERF TFs expression also significantly increased the total organic acid content[34], which is in agreement with our results. Interestingly, the amino acid and carbohydrate content and the expression of the related genes in the flowers were decreased when hot pepper plants were grown under different LED light qualities. This is potentially a novel finding because most studies show a positive relationship between ERFs and these two metabolite classes (based on the Web of Science literature from 2013 to 2023). A possible deduction is that the ERF TFs are not the key regulators, and excess light could lead to reactive oxygen (ROS) outburst, disrupting amino acid and carbohydrate metabolism. However, such mechanisms need to be studied further. Importantly, the expression of the ERF021 gene was highly induced by the 4R:1B:5W LED light ratio, and it is considered a candidate gene for the follow-up studies. The gene is located in chromosome 01 of pepper and encodes 136 amino acids[28], and its functional characterization needs to be further explored in the future.
This study investigated the effect of different LED light ratios on hot pepper maturation and found that the 4R:1B:5W LED light ratio significantly accelerated the process of fruit coloration and increased the yield and quality of hot peppers, which is key for their commercial value. The time for full fruit ripening was shortened by at least 16 days, and the seeds could be harvested earlier under the 4R:1B:5W LED light ratio treatment compared to the control, indicating that the period of fruit maturation in hot pepper was shortened significantly through light quality regulation. The S1 stage, from the seeds to the appearance of the first flower, is the limiting factor affecting the fruit maturation period. It was also revealed that 4R:1B:5W LED light ratios increase flavonoid, organic acid, and phenolics content and decrease amino acids and carbohydrates content in the first flower of hot pepper plants. Correspondingly, the relative expression of key genes relevant to these metabolites was also changed. ERF TFs function as key regulatory factors, especially ERF021, which plays a key role in flower development due to its very high upregulation and regulation of the associated metabolic pathways under the 4R:1B:5W LED light ratio. In view of the above results, the 4R:1B:5W light ratio was considered to be the optimal light quality for shortening the maturation period and fruit development of hot pepper. This finding will provide key insights for accelerating the pepper breeding process and improving the hot pepper fruit commercial value.
4. Materials and Methods
4.1. Plant materials and treatments
The hot pepper cultivar 'Z1' with a maturity period of 75~90 days, provided by the Horticulture Group, School of Agriculture, Ningxia University, was used for the current study. The seeds were sown in an indoor controlled environment room with artificial LED lighting. The environmental conditions were as follows: air temperature of 26~28 °C, relative humidity of 60~70 %, and light period of 16 h day/8 h dark. The photosynthetic photon flux at about 30 cm from the plant canopy top was set to 240 µmol m-2·s-1. The treatments with different LED light qualities were initiated 1-day post sowing (DPS) and ended at 70 DPS (at the mature fruit stage) in hot pepper using red light LED panels (with a peak wavelength at 700 nm), blue light LED panels (with a peak wavelength at 465 nm) and white LED panels. The lighting equipment used in this study was provided by Chenghui Equipment Agricultural Technology Co., LTD., China. The experiment consisted of four treatments, including 3R:1B:6W (T1 group), 4R:1B:5W (T2 group), 5R:1B:4W (T3 group) and 7R:1B:2W (T4 group) (Table S3). The full spectrum white LED light was assigned as the control. Each group included twenty pepper plants.
4.2. Contributed parameters related to phenotype and physiology during fruit ripening
At 70 DPS, all fruits and seeds were individually collected from each pepper plant, and the fruit color frequency distribution in each treatment was estimated by the formula ①. Yield per pepper plant and weight per pepper fruit in each treatment group were measured using digital electronic scales (JY3003, Guangzhou, China). Germinated seeds were counted, and the percentage of seed germination and thousand seed weight were recorded by the formulas ② and ③, respectively. In addition, the color phenotypes of fruits and seeds were also evaluated and recorded. Soluble sugar content was measured according to the method of [35]. The protein content in pectin solutions (concentration of 0.5%, w/v) was determined according to the description of [36]. The firmness property of chili pepper fruit was examined utilizing a texture analyzer (JC-GY-1, Qingdao Juchuang Environmental Protection Group Co. Ltd, China) following the protocols described in our previous publication[37].
4.3. Untarget metabolite profiling using liquid chromatography-mass spectrometry (LC-MS)
Fully opened first flowers of pepper plants (about 33 DPS) were sampled from the control and each treatment group. The procedures were conducted according to the description of [38] and [39]. For extraction, the collected samples were placed into tubes with methanol (containing 2-Cl-L-phenylalanine) and then centrifuged for 10 min at 12000 rpm at 4 ℃. The supernatant was filtered and transferred into the detection vial of the Thermo Vanquish dual LC-MS high-performance liquid chromatography (HPLC) system (Thermo Fisher Scientific, USA). Chromatography was carried out with a ACQUITY UPLC® HSS T3 chromatographic column (Waters, Milford, MA, USA). Mass spectrometric detection of metabolites was performed on Q Exactive Focus (Thermo Fisher Scientific, USA) with electrospray ionization ion source.
The raw data were processed using the XCMS software package, based on R language, for feature detection, retention time correction, and peak alignment. The metabolites were identified by mass accuracy and MS/MS data which were matched with HMDB (http://www.hmdb.ca), MassBank (http://www.massbank.jp/), LipidMaps (http://www.lipidmaps.org), mzCloud (https://www.mzcloud.org) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) (http://www.genome.jp/kegg/). The robust LOESS (Locally Weighted Scatterplot Smoothing) signal correction (QC-RLSC) was applied for data normalization. After normalization, in quality control (QC) assessment, only ion peaks with relative standard deviations (RSDs) less than 30 %, were kept to ensure proper metabolite identification. The Ropls software was used for variable modeling using principal component analysis (PCA). The metabolic profiles were visualized as a score plot. The corresponding loading plots and S-plots were generated to provide information on the metabolites that influence the clustering of the samples. Orthogonal Partial Least Squares-Discriminant Analysis (OPLS-DA) was performed for the determination of discriminating metabolites using the variable importance on projection (VIP) scores. The P values, the VIP scores obtained by OPLS-DA, and fold change (FC) were applied to discover the contributions by variable for classification. Finally, metabolites with a P value < 0.05 and |FC| values > 2 were considered to be statistically significant differentially expressed metabolites (DEMs).
These DEMs were subjected to pathway analysis using the MetaboAnalyst platform (version 4.0), combining pathway enrichment analysis and pathway topology analysis. The identified metabolites were then mapped to their corresponding KEGG pathways for biological interpretation of higher-level system functions. The metabolites and corresponding pathways were visualized using the KEGG Mapper tool (https://www.kegg.jp/kegg/mapper/).
4.4. Measurement of target metabolites
The fully opened first flower of pepper plants (about 33 DPS) from the control and each treatment group was sampled for the determination of total phenolics, carbohydrates, organic acid, flavonoid, and total amino acid contents.
The total phenolics content was measured using an HPLC system (Thermo Fisher Scientific, USA). In brief, the procedure was as follows: (1) 1.0 g samples were ground in a mortar, and the powder, acetonitrile, and HCl were added to a tube and appropriately mixed. (2) The mixture was incubated at 20 °C for 2 h and then centrifuged for 10 min at 4 °C. The supernatant was extracted and evaporated using a nitrogen evaporator (6 Position N-EVAP, Organomation, USA). (3) The extract was redissolved in 80 % methanol and filtered through a membrane filter for HPLC analysis. (4) A Symmetry™ C18 chromatographic column (5μm, 3.9 mm × 20 mm, Waters, USA) was used, with a flow rate of 0.8 mL min−1. The mobile phase A was aqueous 0.3% phosphoric acid in distilled water, and phase B was absolute methanol. (5) A standard curve with the standard mass concentration on the X-axis and the peak area on the Y-axis was drawn using a standard solution in incremental dilutions. (6) The chromatographic analysis of the samples was carried out, and the phenolics content was calculated after peak integration by the peak area. Carbohydrate content was measured according to the description of [40] but a little modification. Briefly, 1.0 g dried sample was grinded in a mortar, and the powder, 50 μL phenol solution (5%) and 300 μL sulfuric acid were mixed in a tube and incubated for 30 min. Then, the supernate was pipetted and added in a cuvette. The absorbance of the sample was recorded at 490 nm using spectrophotometer instrument (UV-3300, Pumeida, Shanghai, China). Total carbohydrate content was expressed in milligrams of glucose equivalents per 100 milligrams sample.
The carbohydrate content was measured according to the protocol described by [41]. Briefly, a 1.0 g dried sample was ground in a mortar, and the powder, 50 μL phenol solution (5%), and 300 μL sulfuric acid were mixed in a tube and incubated for 30 min. Then, the supernatant was transferred to a cuvette. The sample absorbance was recorded at 490 nm using a spectrophotometer (UV-3300, Pumeida, Shanghai, China). The total carbohydrate content was expressed in milligrams of glucose equivalents per 100 milligrams sample.
Organic acids were determined using HPLC (Thermo Fisher Scientific, USA) according to the protocol described by(Lee, Yi, Choi, & Lee, 2020). Briefly, 2.0 g samples were ground and placed into a tube with a water–methanol mixture solution (75:25, v/v). The mixture was centrifuged at 25 °C for 30 min using a centrifuge (5424R, Eppendorf, USA). The upper phase was filtered, and the filtrates were collected. An HPLC system equipped with a UV absorbance detector set at 214 nm was used. Chromatographic separation was performed on a Shodex RSpak KC-118 model ion-exchange organic acid column (300 mm × 8 mm i.d.). The mobile phase was 0.1% (w/v) of phosphoric acid in distilled water (HPLC grade) with a flow rate of 0.8 mL/min. The concentrations of organic acids were estimated from peak areas of known concentrations of standards automatically computed by the data processor.
The flavonoid content measurements were performed according to the method of[42]. The collected, frozen samples were ground to a powder and extracted with an extracting solution (3 M 1HCl: 3H2O:16methanol, volume ratio) overnight at 4 °C in the dark. After centrifugation for 10 min, the absorbance of the supernatant was measured at 530 and 657 nm. The relative flavonoid content in pepper leaves was calculated on the basis of optical density (OD) per g fresh weight by the formula ④
The total amino acid content was measured according to the method of[43]. Approximately 1.0 g dried sample powder was placed into digestion bottle with 20 mL HCl, and digested for 24 h at 110 °C. Then, the mixture was filtered and diluted to 100 mL using ultrapure water. 2 mL of the solution was added into a tube and evaporated using a water bath, and then the crystallization was dissolved with 0.02 mol L−1 HCl and filtered. This mixture was measured for the amino acids content using an amino acids analyzer (S433, SYKAM, Germany).
4.5. RNA extraction and analysis of trancriptom
0.1 g flowers were ground in liquid nitrogen, and the total RNA was extracted using the Trizol reagent.0. The concentration of the RNA was determined, and the isolated RNA was purified further using the RNeasy Mini Kit (DP432, QIAGEN, China). High-quality RNA was sequenced by the Illumina platform, and raw reads were generated. Quality control was performed on the raw reads, and clean reads were obtained, which were then mapped to Capsicum reference genomes (https://www.ncbi.nlm.nih.gov/datasets/genome/?taxon=4072). Gene expression in the transcriptome was measured by RNA-seq by Expectation Maximization (RSEM). Genes with a q value <0.05 and |FC| > 2 were considered to be statistically significant differentially expressed genes (DEGs). PCA analysis was performed on the DEGs from the treatments and the control. Gene Ontology (GO) and KEGG pathway analysis was also conducted to assess the function and the associated metabolic and signaling pathways of the DEGs.
4.6. Quantitative real-time PCR analysis (qPCR)
The total RNA was converted into first-strand cDNA using a PrimeScript Kit (Takara, Dalian, China). The resulting cDNA was used as a template for quantitative PCR amplification using SYBR Green as the fluorescent reporter (BlazeTaq™ SYBR® Green qPCR mix 2.0, GeneCopoeia, USA). Using sequence information from GenBank, primers were designed, and their sequences are listed in Table S1. qRT-PCR was conducted using a fluorescent quantitative PCR instrument (qTower3G, Jena, Germany). Ubi3 gene was selected as the reference gene. The 2-△△CT method was used to determine the relative gene expression, calculated as the mean from three biological replicates.
4.7. Statistical Analysis
All the parameters assessed in this study were measured in triplicates. The data were analyzed using one-way analysis of variance (ANOVA)in SPSS 19.0 software (SPSS Inc., USA). Statistical significance was inferred at p < 0.05. The analyzed data were presented as means ± standard deviation (SD) of three biological replicates.
5. Conclusions
This study investigated the effect of different LED light ratios on hot pepper maturation and found that the 4R:1B:5W LED light ratio significantly accelerated the process of fruit coloration and increased the yield and quality of hot peppers, which is key for their commercial value. The time for full fruit ripening was shortened by at least 16 days, and the seeds could be harvested earlier under the 4R:1B:5W LED light ratio treatment compared to the control, indicating that the period of fruit maturation in hot pepper was shortened significantly through light quality regulation. The S1 stage, from the seeds to the appearance of the first flower, is the limiting factor affecting the fruit maturation period. It was also revealed that 4R:1B:5W LED light ratios increase flavonoid, organic acid, and phenolics content and decrease amino acids and carbohydrates content in the first flower of hot pepper plants. Correspondingly, the relative expression of key genes relevant to these metabolites was also changed. ERF TFs function as key regulatory factors, especially ERF021, which plays a key role in flower development due to its very high upregulation and regulation of the associated metabolic pathways under the 4R:1B:5W LED light ratio. In view of the above results, the 4R:1B:5W light ratio was considered to be the optimal light quality for shortening the maturation period and fruit development of hot pepper. This finding will provide key insights for accelerating the pepper breeding process and improving the hot pepper fruit commercial value.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1 The treatment of light quality in the experiment; Table S2 Seed germination in fruit with different color under LED light qualities; Table S3 Primers used in this study; Table S4 Percentage of fruit color under different LED light qualities; Table S5 The correlation index between vitexin and metabolites; Table S6 Estimation of RNA-sequencing data; Table S7 Statistics of expression difference analysis results. In addition, 3 supplementary figure also were included in this study and listed as a pptx file (to see “Supplementary file-Figures”): Figure S1 The base chromatograms depicting the metabolite peaks; Figure S2 Accumulation of metabolic component based on KEGG analysis; Figure S3 Principal component analysis under different light ratios.
Author Contributions
Conceptualization, Shuo Yang. and Xiao-Juan Ma; methodology, Shuo Yang; formal analysis, Xuan Zheng and Yi Hou; investigation, Shuo Yang; resources, Guo-Xin Cheng and Xiao-Min Wang; writing—original draft preparation, Shuo Yang; writing—review and editing, Guo-Xin Cheng.; supervision, Guo-Xin Cheng; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Key Research and Development Project of Ningxia (2021YFD1600302), National Key Research and Development Project of China (2021BBF02024), Nature and Science Foundation of Ningxia (2022AAC05023), and Key Research and Development Plan (Major Projects) of Ningxia (NXNYYZ202001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank MOgoEdit company for providing language help.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Liu, K.; He, R.; He, X.; Tan, J.; Chen, Y.; Li, Y.; Liu, R.; Huang, Y.; Liu, H. Speed Breeding Scheme of Hot Pepper through Light Environment Modification. Sustainability-basel 2022, 14, 12225. [Google Scholar] [CrossRef]
- van der Meer, M.; Lee, H.; de Visser, P. H. B.; Heuvelink, E.; Marcelis, L. F. M. Consequences of interplant trait variation for canopy light absorption and photosynthesis. Front. Plant Sci. 2023, 14, 1012718. [Google Scholar] [CrossRef] [PubMed]
- Larsen, D. H.; Li, H.; van de Peppel, A. C.; Nicole, C. C. S.; Marcelis, L. F. M.; Woltering, E. J. High light intensity at End-Of-Production improves the nutritional value of basil but does not affect postharvest chilling tolerance. Food Chem. 2022, 369, 130913. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Lafuente, M. T. , LED Blue Light-induced changes in phenolics and ethylene in citrus fruit: Implication in elicited resistance against Penicillium digitatum infection. Food Chem. 2017, 218, 575–583. [Google Scholar] [CrossRef]
- Rashidi, A.; Tehranifar, A.; Samiei, L. Modifying spectral distributions during the seedling stage influences the flowering and branching of Petunia × hybrida. Sci. Hortic-Amsterdam 2023, 309, 111664. [Google Scholar] [CrossRef]
- Nie, W.-F.; Li, Y.; Chen, Y.; Zhou, Y.; Yu, T.; Zhou, Y.; Yang, Y. Spectral light quality regulates the morphogenesis, architecture, and flowering in pepper (Capsicum annuum L.). J. Photoch. Photobio. B. 2023, 241, 112673–112673. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Qin, J.; Xie, F.; Zhou, K.; Xi, W. Red light-transmittance bagging promotes carotenoid accumulation through xanthophylls esterification during the ripening of blood orange fruit. Food Chem. 2023, 404, 134578. [Google Scholar] [CrossRef]
- Su, L.; Hou, P.; Song, M.; Zheng, X.; Guo, L.; Xiao, Y.; Yan, L.; Li, W.; Yang, J. Synergistic and Antagonistic Action of Phytochrome (Phy) A and PhyB during Seedling De-Etiolation in Arabidopsis thaliana. Food Chem. 2023, 404, 134578. [Google Scholar] [CrossRef]
- Hajdu, A.; Adam, E.; Sheerin, D. J.; Dobos, O.; Bernula, P.; Hiltbrunner, A.; Kozma-Bognar, L.; Nagy, F. High-level expression and phosphorylation of phytochrome B modulates flowering time in Arabidopsis. Plant J. 2015, 83, 794–805. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Y. C.; Sang, Y.; Li, Q. H.; Yang, H. Q. A role for Arabidopsis cryptochromes and COP1 in the regulation of stomatal opening. Pro. Natl Aca. Sci. USA 2005, 102, 12270–12275. [Google Scholar] [CrossRef]
- Holub, D.; Kubar, T.; Mast, T.; Elstner, M.; Gillet, N. , What accounts for the different functions in photolyases and cryptochromes: a computational study of proton transfers to FAD. Phys. Chem. Chem. Phy. 2019, 21(22), 11956–11966. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-Y.; Koh, S. C. , Fruit Development and Quality of Hot Pepper (Capsicum annuum L.) under Various Temperature Regimes. Hort. Sci. Technol. 2019, 37, 313–321. [Google Scholar] [CrossRef]
- Xiang, N.; Qi, X.; Hu, J.; Wang, S.; Guo, X. l-Tryptophan synergistically increased carotenoid accumulation with blue light in maize (Zea mays L.) sprouts. Food Chem. Mol. Sci. 2023, 6, 100161. [Google Scholar] [CrossRef] [PubMed]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. Inter. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, X.; Liu, X.; Xue, J.; Ren, X.; Zhai, Y.; Zhang, X. The red/blue light ratios from light-emitting diodes affect growth and flower quality of Hippeastrum hybridum 'Red Lion'. Front. Plant Sci. 2022, 13, 1048770. [Google Scholar] [CrossRef] [PubMed]
- Walker, C. H.; Bennett, T. A distributive '50% rule' determines floral initiation rates in the Brassicaceae. Nat. Plants 2019, 5, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Jaudal, M.; Wen, J.; Mysore, K. S.; Putterill, J. Medicago PHYA promotes flowering, primary stem elongation and expression of flowering time genes in long days. Bmc Plant Biol. 2020, 20, 329. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, Y.; Li, X.; Chen, Q.; Zhang, Y.; Luo, Y.; Liu, Z.; Wang, Y.; Lin, Y.; Zhang, Y.; Wang, X.; Tang, H. Transcriptome Profile Analysis of Strawberry Leaves Reveals Flowering Regulation under Blue Light Treatment. Int. J. Genomics 2021, 2021, 5572076. [Google Scholar] [CrossRef] [PubMed]
- Danziger, N.; Bernstein, N. Light matters: Effect of light spectra on cannabinoid profile and plant development of medical cannabis (Cannabis sativa L.). Ind. Crop. Prod. 2021, 164, 113351. [Google Scholar] [CrossRef]
- Gonzalez-Barrios, P.; Bhatta, M.; Halley, M.; Sandro, P.; Gutierrez, L. Speed breeding and early panicle harvest accelerates oat (Avena sativa L.) breeding cycles. Crop Sci. 2021, 61, 320–330. [Google Scholar] [CrossRef]
- Vidana Gamage, G. C.; Lim, Y. Y.; Choo, W. S. Anthocyanins From Clitoria ternatea Flower: Biosynthesis, Extraction, Stability, Antioxidant Activity, and Applications. Front. Plant Sci. 2021, 12, 792303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, L.; Li, Y.; Chen, Q.; Yuntian, Y.; Zhang, Y.; ya, L.; Sun, B.; Wang, X.-r.; Tang, H. Effect of Red and Blue Light on Anthocyanin Accumulation and Differential Gene Expression in Strawberry (Fragaria × ananassa). Molecules 2018, 23, 820. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, M.; Dang, S.; Zhou, J.; Zhang, Y. Comparative transcriptomic analysis of transcription factors and hormones during flower bud differentiation in 'Red Globe' grape under red‒blue light. Sci. Rep. 2023, 13, 8932–8932. [Google Scholar] [CrossRef] [PubMed]
- Prisca, M.; Maarten, V.; Jan, V. D.; Bart, N.; Wouter, S.; Timo, H.; Barbara, D. C.; Bram, V. d. P. Blue and far-red light control flowering time of woodland strawberry (Fragaria vesca) distinctively via CONSTANS (CO) and FLOWERING LOCUS T1 (FT1) in the background of sunlight mimicking radiation. Environ. Exp. Bot. 2022, 198, 104866. [Google Scholar] [CrossRef]
- He, R.; Wei, J.; Zhang, J.; Tan, X.; Li, Y.; Gao, M.; Liu, H. Supplemental Blue Light Frequencies Improve Ripening and Nutritional Qualities of Tomato Fruits. Front. Plant Sci. 2022, 13, 888976. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Tohge, T.; Lalor, P.; Dockery, P.; Devaney, N.; Esteves-Ferreira, A. A.; Fernie, A. R.; Sulpice, R. Phytochrome A and B Regulate Primary Metabolism in Arabidopsis Leaves in Response to Light. Front. Plant Sci. 2017, 8, 1394. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; He, Y.; Li, S.; Shi, S.; Li, L.; Liu, Y.; Chen, H. Genome-wide characterization and expression analysis of AP2/ERF genes in eggplant (Solanum melongena L.). Plant Physiol. Biochem. 2021, 167, 492–503. [Google Scholar] [CrossRef]
- Jin, J.-H.; Wang, M.; Zhang, H.-X.; Khan, A.; Wei, A.-M.; Luo, D.-X.; Gong, Z.-H. Genome-wide identification of the AP2/ERF transcription factor family in pepper (Capsicum annuum L.). Genome 2018, 61, 663–674. [Google Scholar] [CrossRef]
- Gong, D.; Cao, S.; Sheng, T.; Shao, J.; Song, C.; Wo, F.; Chen, W.; Yang, Z. Effect of blue light on ethylene biosynthesis, signalling and fruit ripening in postharvest peaches. Sci. Hortic-Amsterdam 2015, 197, 657–664. [Google Scholar] [CrossRef]
- Ni, J.; Bai, S.; Zhao, Y.; Qian, M.; Tao, R.; Yin, L.; Gao, L.; Teng, Y. Ethylene response factors Pp4ERF24 and Pp12ERF96 regulate blue light-induced anthocyanin biosynthesis in ‘Red Zaosu’ pear fruits by interacting with MYB114. Plant Mol. Biol. 2019, 99, 67–78. [Google Scholar] [CrossRef]
- Medda, S.; Dessena, L.; Mulas, M. Monitoring of the PAL Enzymatic Activity and Polyphenolic Compounds in Leaves and Fruits of Two Myrtle Cultivars during Maturation. Agriculture-Basel 2020, 10, 39. [Google Scholar] [CrossRef]
- Min, T.; Liu, E.; Xie, J.; Yi, Y.; Wang, L.; Ai, Y.; Wang, H. Effects of Vacuum Packaging on Enzymatic Browning and Ethylene Response Factor (ERF) Gene Expression of Fresh-cut Lotus Root. Hortscience 2019, 54, 331–336. [Google Scholar] [CrossRef]
- Sun, M.; Shi, M.; Wang, Y.; Huang, Q.; Yuan, T.; Wang, Q.; Wang, C.; Zhou, W.; Kai, G. The biosynthesis of phenolic acids is positively regulated by the JA-responsive transcription factor ERF115 in Salvia miltiorrhiza. J. Exp. Bot. 2019, 70, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Mitsuyama, K.; Komiyama, Y.; Yagi, M.; Andoh, A.; Sata, M. Preventive effects of enzyme-treated rice fiber in a restraint stress-induced irritable bowel syndrome model. Int. J. Mol. Med. 2010, 25, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Hu, D.; Yu, J.; Liu, H. Effect of green, yellow and purple radiation on biomass, photosynthesis, morphology and soluble sugar content of leafy lettuce via spectral wavebands "knock out". Sci. Hortic-Amsterdam 2018, 236, 10–17. [Google Scholar] [CrossRef]
- Li, M.; Wen, X.; Peng, Y.; Wang, Y.; Wang, K.; Ni, Y. Functional properties of protein isolates from bell pepper (<i>Capsicum annuum</i> L. var. <i>annuum</i>) seeds. Lwt-Food Sci. Technol. 2018, 97, 802–810. [Google Scholar]
- Mi, S.; Zhang, X.; Wang, Y.; Zheng, M.; Zhao, J.; Gong, H.; Wang, X. Effect of different genotypes on the fruit volatile profiles, flavonoid composition and antioxidant activities of chilli peppers. Food Chem. 2022, 374, 131751. [Google Scholar] [CrossRef] [PubMed]
- Zelena, E.; Dunn, W. B.; Broadhurst, D.; Francis-McIntyre, S.; Carroll, K. M.; Begley, P.; O'Hagan, S.; Knowles, J. D.; Halsall, A.; Wilson, I. D.; Kellt, D. B.; Consortium, H. Development of a Robust and Repeatable UPLC-MS Method for the Long-Term Metabolomic Study of Human Serum. Analytical Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Want, E. J.; Masson, P.; Michopoulos, F.; Wilson, I. D.; Theodoridis, G.; Plumb, R. S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J. K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S. I.; Lee, Y. C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Analytical Biochemistry 2005, 339(1), 69–72. [Google Scholar] [CrossRef]
- Lee, J. G.; Yi, G.; Choi, J. H.; Lee, E. J. Analyses of targeted/untargeted metabolites and reactive oxygen species of pepper fruits provide insights into seed browning induced by chilling. Food Chem. 2020, 332, 127406. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.-Y.; Cheng, G.-X.; Zhang, Z.; Sun, J.-T.; Ali, M.; Jia, Q.-L.; Luo, D.-X.; Gong, Z.-H.; Li, D.-W. CaMYC, A Novel Transcription Factor, Regulates Anthocyanin Biosynthesis in Color-leaved Pepper (Capsicum annuum L.). J. Plant Growth Regul. 2019, 38, 574–585. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, S.; He, X.; Li, Y.; Zhang, Y.; Chen, W. Response of total phenols, flavonoids, minerals, and amino acids of four edible fern species to four shading treatments. Peerj 2020, 8, 8354. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The phenotypes of fruit in hot pepper under different LED light qualities (A) the process of fruit maturity in hot pepper; (B) fruit coloration in hot pepper under different LED light qualities; (C) Total sugar content; (D) Total protein content; (E) Fruit firmness. T1, 3R:1B:6W; T2, 4R:1B:5W; T3, 5R:1B:4W; T4, 7R:1B:2W; PRF, percentage of red fruit. Three biological replicates were conducted in the experiment.
Figure 1.
The phenotypes of fruit in hot pepper under different LED light qualities (A) the process of fruit maturity in hot pepper; (B) fruit coloration in hot pepper under different LED light qualities; (C) Total sugar content; (D) Total protein content; (E) Fruit firmness. T1, 3R:1B:6W; T2, 4R:1B:5W; T3, 5R:1B:4W; T4, 7R:1B:2W; PRF, percentage of red fruit. Three biological replicates were conducted in the experiment.
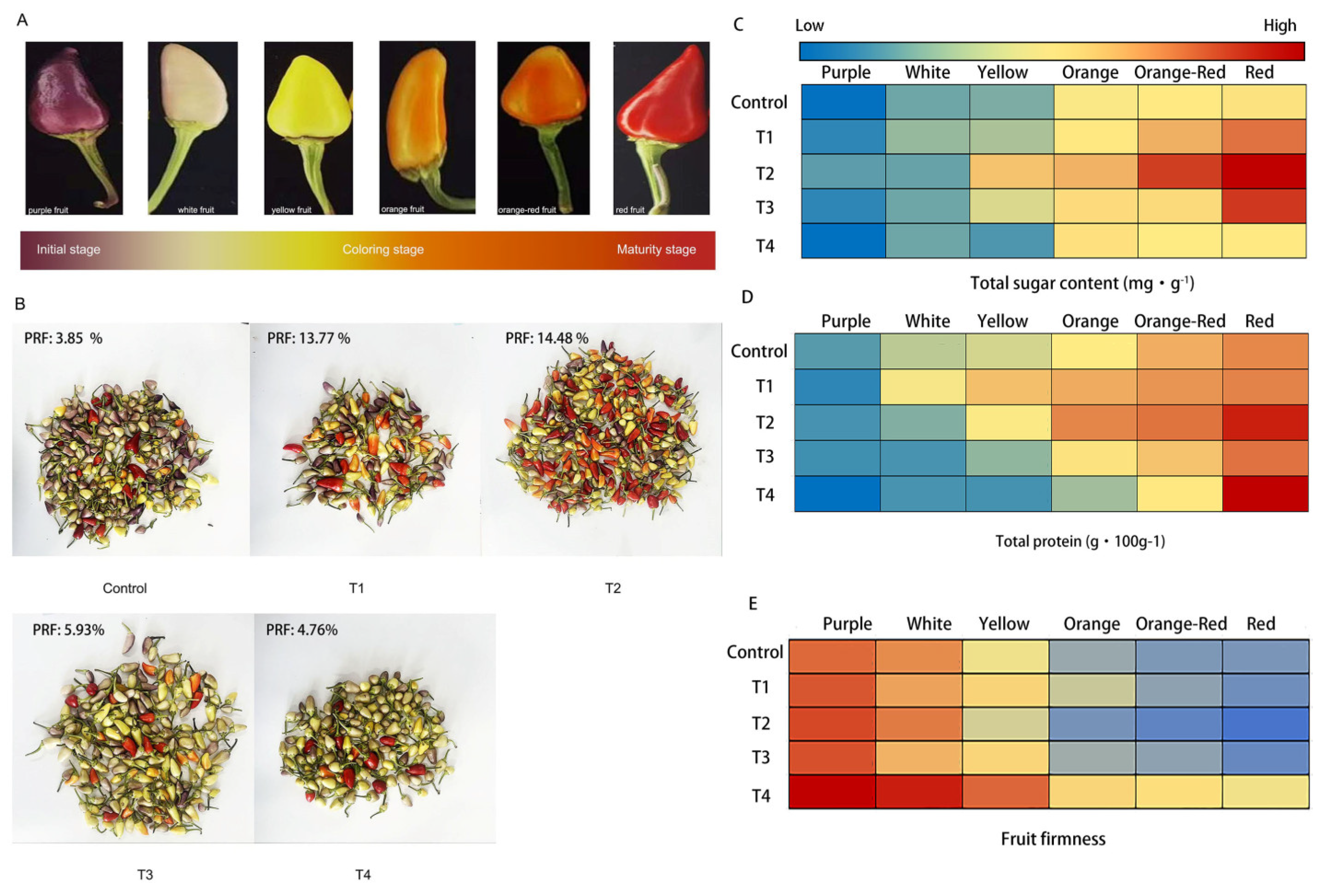
Figure 2.
Period of fruit maturity and seed development in hot pepper under different LED light qualities (A) Period of fruit maturity and seed development from different colored hot peppers; (B) thousand-seed weight; (C) Yield per pepper individual; (D) Weight per fruit; The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.
Figure 2.
Period of fruit maturity and seed development in hot pepper under different LED light qualities (A) Period of fruit maturity and seed development from different colored hot peppers; (B) thousand-seed weight; (C) Yield per pepper individual; (D) Weight per fruit; The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.
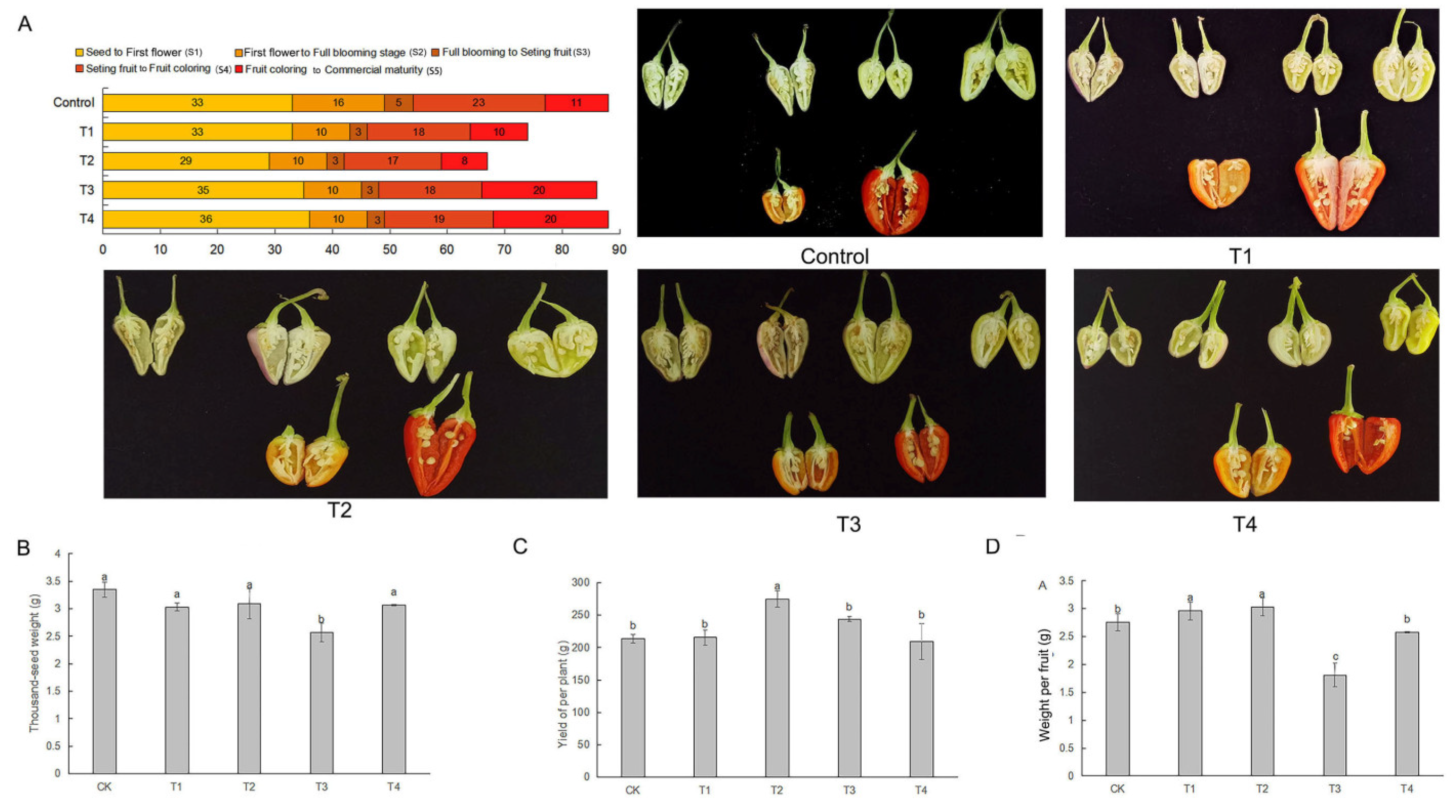
Figure 3.
The development of first flowers and changes of their untarget different expression metabolites (DEMs) in hot pepper based on metabolome analysis under different LED light qualities (A) Phenotype of first flowers; (B) PCA principal component analysis; (C) Venn diagram of untarget DEMs; (D) Z-score analysis of DEMs; In the figure D, the vertical coordinate is names of the metabolite, and the color of the points represents different groups. The horizontal coordinate is the relative content of the metabolite in the group obtained by Z-score conversion. The more to the right, the more the metabolite in the group. Three biological replicates were conducted in the experiment.
Figure 3.
The development of first flowers and changes of their untarget different expression metabolites (DEMs) in hot pepper based on metabolome analysis under different LED light qualities (A) Phenotype of first flowers; (B) PCA principal component analysis; (C) Venn diagram of untarget DEMs; (D) Z-score analysis of DEMs; In the figure D, the vertical coordinate is names of the metabolite, and the color of the points represents different groups. The horizontal coordinate is the relative content of the metabolite in the group obtained by Z-score conversion. The more to the right, the more the metabolite in the group. Three biological replicates were conducted in the experiment.
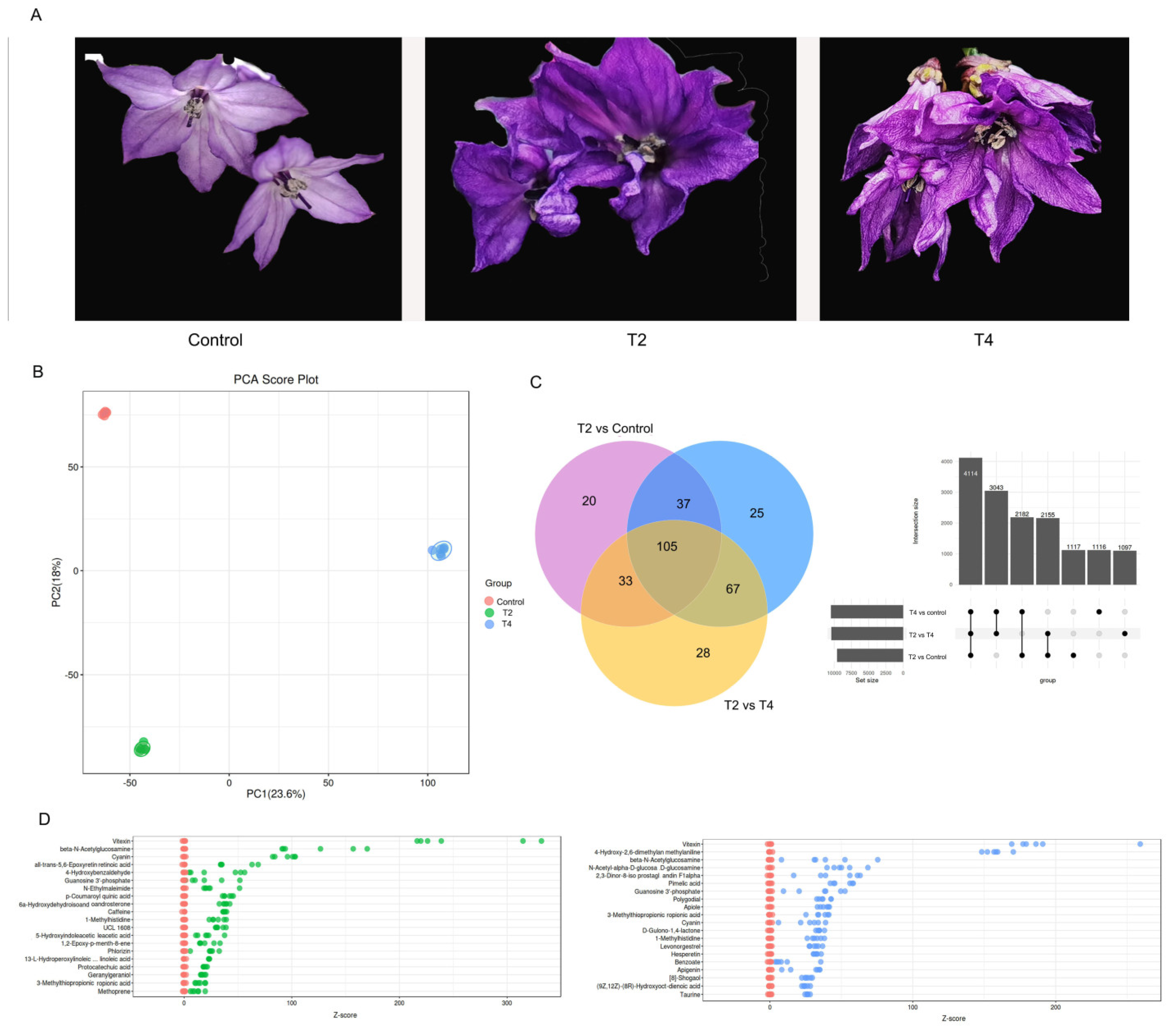
Figure 4.
The levels of target metabolites and changes of metabolic pathway under in first flowers of hot pepper under different LED light qualities (A) The contents in flavonoid, carbohydrate, total phenolics, organic acid and total amino acid; (B) Analysis on metabolic pathway based on metabolome sequencing; The most complex metabolic pathway is enclosed using green box. The red points represent metabolic pathways, and the others means various kinds of metabolic product. The size of the pathway point indicates that the more metabolites are associated with it, the larger the point; T2, 4R:1B:5W; T4, 7R:1B:2W; The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.
Figure 4.
The levels of target metabolites and changes of metabolic pathway under in first flowers of hot pepper under different LED light qualities (A) The contents in flavonoid, carbohydrate, total phenolics, organic acid and total amino acid; (B) Analysis on metabolic pathway based on metabolome sequencing; The most complex metabolic pathway is enclosed using green box. The red points represent metabolic pathways, and the others means various kinds of metabolic product. The size of the pathway point indicates that the more metabolites are associated with it, the larger the point; T2, 4R:1B:5W; T4, 7R:1B:2W; The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.
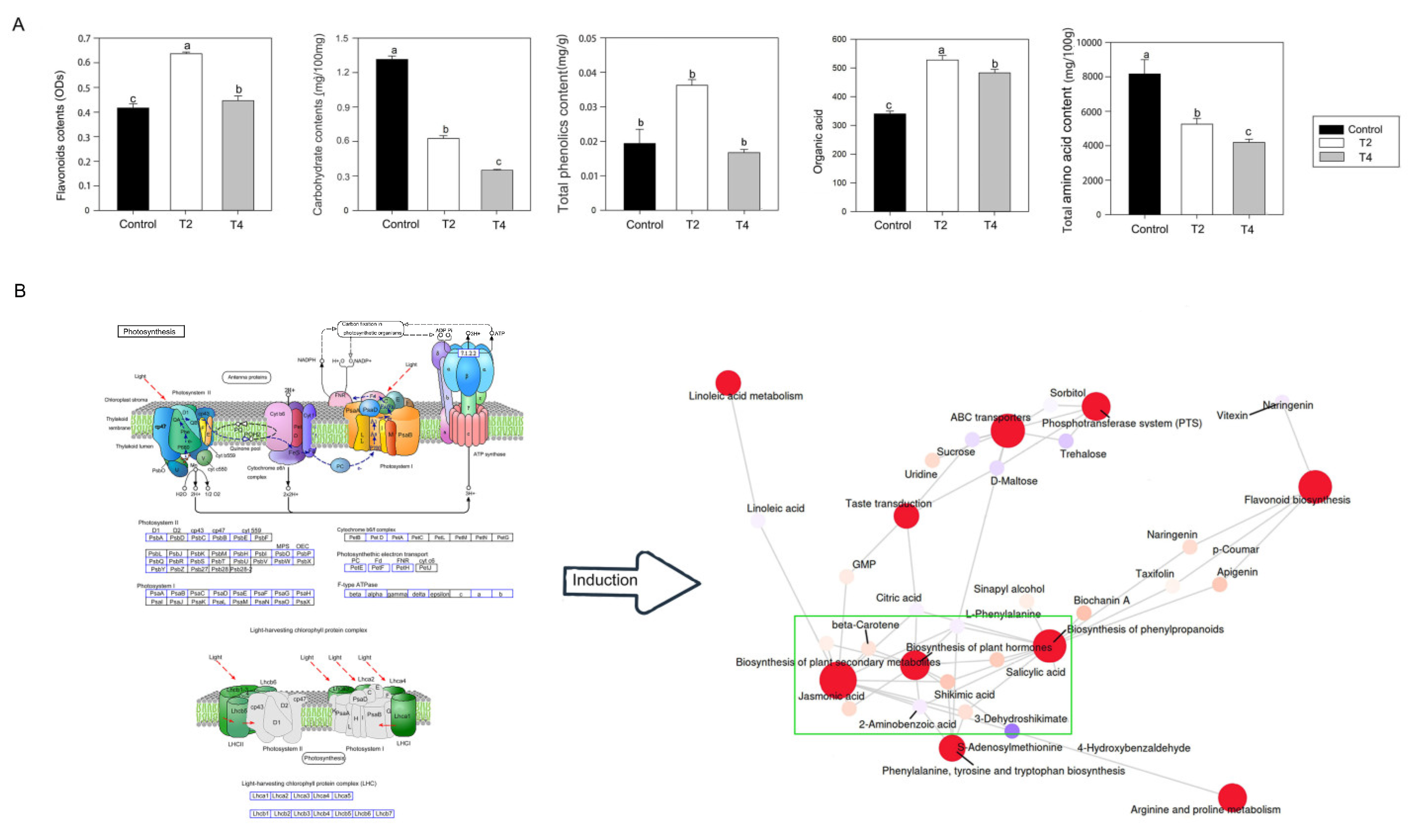
Figure 5.
Screening and analysis of different expression genes (DEGs) based on RNA-sequencing in the first flower of hot pepper under different LED light qualities (A) DEGs enrichment; (B) GO annotation; (C) KEGG analysis; (D) Number of TFs among all DEGs; The DEGs were further analysis based on RNA clean data, and RNA sequencing was conducted using the Illumina platform; Three biological replicates were conducted in the experiment.
Figure 5.
Screening and analysis of different expression genes (DEGs) based on RNA-sequencing in the first flower of hot pepper under different LED light qualities (A) DEGs enrichment; (B) GO annotation; (C) KEGG analysis; (D) Number of TFs among all DEGs; The DEGs were further analysis based on RNA clean data, and RNA sequencing was conducted using the Illumina platform; Three biological replicates were conducted in the experiment.
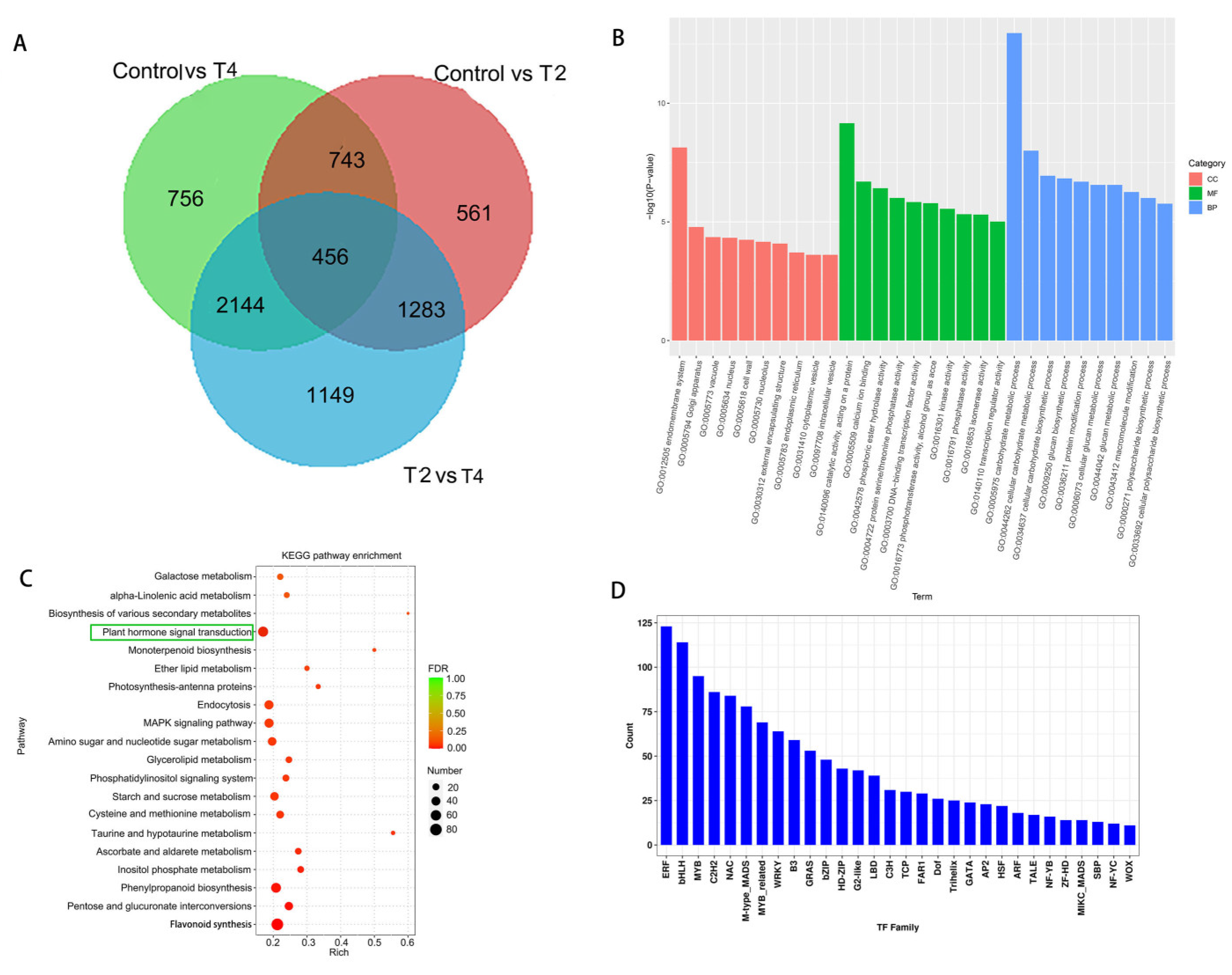
Figure 6.
Integrative analysis of DEGs and DEGs.
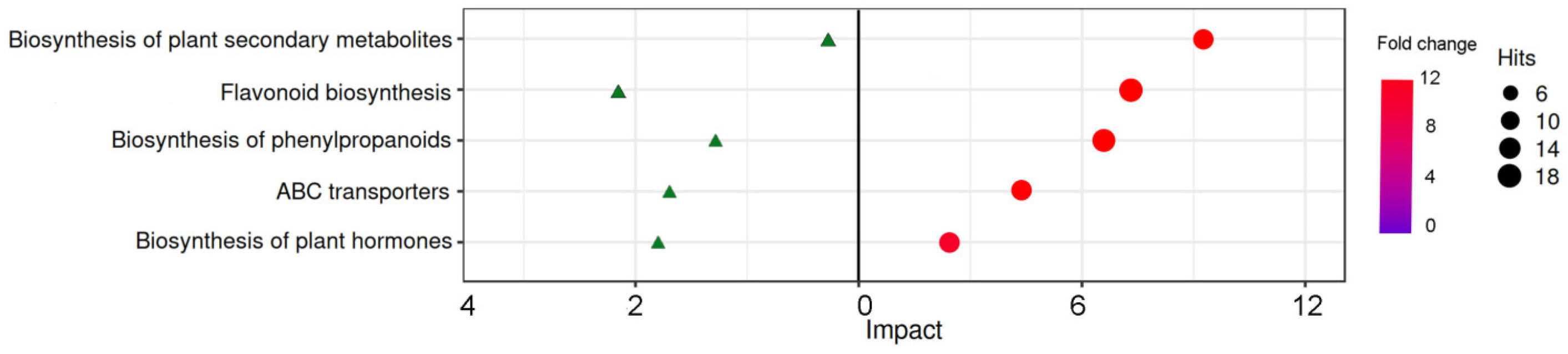
Figure 7.
Expression profiles of DEGs in the first flower of hot pepper under different LED light qualities DEGs expression of (A) Ethylene pathway, (B) Flavonoid pathway, (C) Photosynthesis and photoreceptor, (D) amino acid metabolic pathway and (E) Carbohydrate. The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.
Figure 7.
Expression profiles of DEGs in the first flower of hot pepper under different LED light qualities DEGs expression of (A) Ethylene pathway, (B) Flavonoid pathway, (C) Photosynthesis and photoreceptor, (D) amino acid metabolic pathway and (E) Carbohydrate. The error bars represent SD for three biological replicates, and the lowercases showed the significant level at P < 0.05.


Table 1.
Key genes related metabolites and their description.
| Metabolites | Gene ID | Fold change | Description |
|---|---|---|---|
| Plant hormone | CA09g03230 | 6.43 | pathogenesis-related leaf protein 4-like [Capsicum annuum] |
| CA03g34530 | -5.06 | auxin-induced protein AUX22-like [Capsicum annuum] | |
| CA06g10200 | 2.78 | small auxin-up protein 58 | |
| CA01g31150 | 6.97 | pathogenesis-related protein 1A-like [Capsicum annuum] | |
| CA07g13330 | 4.50 | jasmonic acid-amido synthetase JAR1-like isoform X1 | |
| CA12g03510 | -2.5 | serine/threonine-protein kinase SAPK2-like | |
| CA01g18900 | 2.2 | auxin-responsive protein IAA8-like | |
| CA10g20850 | -3.2 | transcription factor TGA2-like | |
| CA11g01500 | 2.9 | transcription factor HBP-1b(c38)-like | |
| CA03g21920 | 2.2 | uncharacterized protein LOC107864989 | |
| Flavonoid | CA04g12020 | 3.40 | flavonoid 3'-monooxygenase-like |
| CA04g13910 | 6.16 | cytochrome P450 71A1-like | |
| CA01g04460 | 2.22 | flavonoid 3'-monooxygenase-like | |
| CA02g21520 | -2.33 | flavanone 3 beta-hydroxylase | |
| Amino acid | CA03g02860 | -4.09 | probable indole-3-pyruvate monooxygenase YUCCA3 |
| CA03g25930 | -7.10 | cytochrome P450 71A6-like | |
| CA01g33440 | 2.82 | hydroxymethylglutaryl-CoA synthase-lik | |
| CA12g20610 | -2.17 | adenosylhomocysteinase | |
| CA10g05940 | -3.54 | 1-aminocyclopropane-1-carboxylate synthase 4-like | |
| CA08g13390 | 3.43 | uncharacterized protein LOC104242730 | |
| CA04g06460 | -2.17 | 1-aminocyclopropane-1-carboxylate synthase 3-like | |
| CA09g16560 | -2.63 | probable polyamine oxidase 5 | |
| CA03g25960 | -6.88 | LOW QUALITY PROTEIN: (+)-menthofuran synthase-like | |
| CA12g15620 | -3.23 | 1-aminocyclopropane-1-carboxylate oxidase 1 | |
| CA08g07360 | -3.93 | bifunctional L-3-cyanoalanine synthase/cysteine synthase 2, mitochondrial isoform X1 | |
| CA07g11190 | -2.81 | 1-aminocyclopropane-1-carboxylate oxidase 3-like | |
| CA06g11020 | -3.07 | S-adenosylmethionine decarboxylase proenzyme 4-like | |
| CA05g16500 | 2.74 | isoleucine N-monooxygenase 1-like | |
| CA03g02860 | -4.09825977 | probable indole-3-pyruvate monooxygenase YUCCA3 | |
| CA03g25930 | -7.10 | cytochrome P450 71A6-like | |
| CA01g33440 | 2.82 | hydroxymethylglutaryl-CoA synthase-like | |
| CA12g20610 | -2.17 | adenosylhomocysteinase | |
| CA10g05940 | -3.54 | 1-aminocyclopropane-1-carboxylate synthase 4-like | |
| CA08g13390 | 3.43 | uncharacterized protein LOC104242730 | |
| CA04g06460 | -2.17 | 1-aminocyclopropane-1-carboxylate synthase 3-like | |
| CA09g16560 | -2.63 | probable polyamine oxidase 5 | |
| CA03g25960 | -6.88 | LOW QUALITY PROTEIN: (+)-menthofuran synthase-like | |
| CA12g15620 | -3.23 | 1-aminocyclopropane-1-carboxylate oxidase 1 | |
| CA08g07360 | -3.93 | bifunctional L-3-cyanoalanine synthase/cysteine synthase 2, mitochondrial isoform X1 | |
| CA07g11190 | -2.81 | 1-aminocyclopropane-1-carboxylate oxidase 3-like | |
| CA06g11020 | -3.07 | S-adenosylmethionine decarboxylase proenzyme 4-like | |
| CA05g16500 | 2.74 | isoleucine N-monooxygenase 1-like [Capsicum annuum] | |
| Carbohydrate | CA04g15380 | 3.369706641 | 7-deoxyloganetin glucosyltransferase-like |
| CA12g05960 | -2.559152027 | beta-D-glucosyl crocetin beta-1,6-glucosyltransferase-like ] | |
| CA12g19750 | 2.681845577 | limonoid UDP-glucosyltransferase-like isoform X2 | |
| CA12g09020 | 4.314617163 | antifungal protein | |
| CA03g28260 | 2.656431408 | UDP-glycosyltransferase 90A1-like | |
| CA01g22510 | 2.63238818 | anthocyanidin 3-O-glucosyltransferase 2-like | |
| CA02g28410 | 2.57066522 | aldose 1-epimerase-like | |
| CA12g19660 | 2.784774378 | UDP-glycosyltransferase 74E2-like | |
| CA03g00610 | -3.33150609 | germacrene C synthase-like isoform X2 | |
| CA07g09480 | 5.117071724 | antifungal protein | |
| CA12g02820 | 6.208983562 | LOW QUALITY PROTEIN: rhamnogalacturonate lyase-like | |
| CA02g08210 | -2.364017925 | alpha-1,4-glucan-protein synthase [UDP-forming] 1-like | |
| CA03g30170 | 7.594115814 | chitin-binding lectin 1-like | |
| CA12g19670 | 2.84487617 | UDP-glycosyltransferase 74E2-like | |
| CA06g16330 | -3.952708828 | inorganic pyrophosphatase 1-like | |
| CA03g35740 | -2.14010396 | beta-fructofuranosidase, insoluble isoenzyme CWINV1-like | |
| CA06g18490 | 8.087637601 | D: UDP-glucose 6-dehydrogenase 1-like | |
| CA02g17640 | 6.038102413 | alpha-farnesene synthase-like, partial | |
| CA02g21050 | 3.962130051 | acidic 27 kDa endochitinase | |
| CA05g00420 | 5.203134101 | glucan endo-1,3-beta-glucosidase, acidic, partial | |
| CA03g10530 | 2.406883633 | protein NDH-DEPENDENT CYCLIC ELECTRON FLOW 5 | |
| CA07g13240 | 2.470378336 | endochitinase 4 | |
| CA03g24000 | 6.93069508 | UDP-arabinopyranose mutase 3-like isoform X2 | |
| CA07g06280 | -2.608737749 | 7-deoxyloganetin glucosyltransferase-like | |
| CA09g17310 | -2.578056349 | pyruvate dehydrogenase E1 component subunit alpha-3, chloroplastic-like | |
| CA02g21020 | 4.228337947 | basic endochitinase-like | |
| CA01g04790 | -2.085738523 | acid beta-fructofuranosidase | |
| CA05g05560 | -3.708332213 | beta-xylosidase/alpha-L-arabinofuranosidase 2-like | |
| CA01g20700 | -2.639117089 | polygalacturonase-like | |
| CA12g19530 | -3.894926195 | polygalacturonase-like | |
| CA02g24560 | -2.091080347 | probable hexokinase-like 2 protein | |
| CA12g00950 | -2.622367834 | glucan endo-1,3-beta-glucosidase 8-like | |
| CA12g19520 | -5.39008693 | polygalacturonase-like | |
| CA11g08190 | -2.832023018 | polygalacturonase At1g48100-like | |
| CA04g18770 | 6.215998482 | alpha-amylase-like | |
| Organic acid | CA08g15160 | 3.313126727 | 12-oxophytodienoate reductase-like protein isoform X1 |
| CA12g22620 | 5.911464401 | delta(12)-acyl-lipid-desaturase-like | |
| CA07g16860 | 2.109237399 | gamma-glutamyl hydrolase 1-like isoform X1 | |
| CA04g15770 | 3.769308934 | LOW QUALITY PROTEIN: alcohol dehydrogenase-like 1 | |
| CA01g17610 | 2.733481441 | stearoyl-[acyl-carrier-protein] 9-desaturase 6, chloroplastic-like | |
| CA01g02250 | 3.694905784 | phosphoglycerate mutase-like protein AT74 | |
| CA01g00840 | 3.817028485 | 3-oxoacyl-[acyl-carrier-protein] synthase I, chloroplastic-like | |
| CA12g22610 | 3.857220972 | delta(12)-acyl-lipid-desaturase-like | |
| CA10g12320 | 2.630639785 | 12-oxophytodienoate reductase 1-like | |
| CA12g22630 | 4.647795771 | delta(12)-acyl-lipid-desaturase-like | |
| CA09g03700 | 2.316060915 | L-ascorbate peroxidase 2, cytosolic | |
| CA08g04180 | 3.043702607 | delta(12)-fatty-acid desaturase FAD2-like | |
| CA06g17700 | 5.393154636 | phosphoethanolamine N-methyltransferase 1-like | |
| ABC transporter | CA02g05780 | 3.175 | ethylene-responsive transcription factor ERF113-like |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



