
Submitted:
19 December 2023
Posted:
19 December 2023
You are already at the latest version
Abstract
Silkworm ovary-derived BmN4 cells rely on chromatin-induced spindle assembly to form microtubule-based square mitotic spindles that ensure accurate segregation of holocentric chromosomes during cell division. The chromosome passenger protein Aurora B regulates chromosomal condensation and segregation, spindle assembly checkpoint, and cytokinesis; however, its role in holocentric organisms needs further clarification. This study examined the architecture and dynamics of spindle microtubules during prophase and metaphase in BmN4 cells and those with siRNA-mediated BmAurora B knockdown using immunofluorescence labelling. Anti-α-tubulin and anti-γ-tubulin antibodies revealed faint γ-tubulin signals co-localized with α-tubulin in early prophase during nuclear membrane rupture, which intensified as prophase progressed. At this stage, bright regions of α-tubulin around and on the nuclear membrane surrounding the chromatin suggested the start of microtubule assembling in the microtubule organizing centers (MTOCs). In metaphase, the number and size of γ-tubulin foci increased and were detected on both sides of the chromosomes. This resulted in a distinctive multipolar square spindle with holocentric chromosomes aligned at the metaphase plate. siRNA-mediated BmAurora B knockdown significantly reduced the γ-tubulin foci during prophase, impacting microtubule nucleation and spindle structure in metaphase. Spatiotemporal BmAurora B expression analysis provided new insights into the regulation of this mitotic kinase in silkworm larval gonads during gametogenesis. Our results suggest that BmAurora B is crucial for the formation of multipolar square spindles in holocentric insects, possibly through the activation of γ-tubulin ring complexes in multiple centrosome-like MTOCs.
Keywords:
Bombyx mori
; Chromatin-induced spindle assembly
; siRNA
; cell division
; mitosis
1. Introduction
In addition to the regular changes in chromatin during the cell cycle, there are also remarkable changes in the non-chromatin phase in the cytoplasm and nucleus. Spindles are dynamic structures composed of microtubule (MT) polymers and hundreds of related factors that can be customized according to the cellular environment. A pair of sister chromatids involved in the regulation of interphase replication is precisely distributed to two daughter cells in the later phase of mitosis [1]. In animal cells containing centrosomes, a pair of right-angled centrioles is near the interphase nuclei. In the S phase, one side of each centriole is joined to form a new centriole [2]. Each pair of centrioles is surrounded by a highly ordered mass of dense material, called pericentriolar material, and together they form the microtubule organizing centers (MTOCs).
The γ-tubulin ring near the basal base of each pair of centrioles is the starting point for microtubule assembly [3]. Just before mitosis, short star-shaped radial arrays named asters form around each pair of centrioles. The two centrosomes move apart along the microtubules until they are on opposite sides at the outer edge of the nuclear membrane. When they reach this position, they determine the cell division pole [4]. At this point, the spindles of the two poles are formed. The spindle microtubules that extend from the two poles are called polar microtubules. The microtubules from each pole form half a spindle. Higher plant cells have no centrioles or stars but can still form square shaped spindles and undergo mitosis due to the presence of substances around the centrioles [5]. In cells without centrosomes, such as female meiotic cells, the chromosomes must nucleate and stabilize microtubules to form a bioriented spindle [6].
Acentrosomal microtubule assembly involves the formation of nuclei near chromosomes or on existing MTs, driven by a RanGTP-dependent mechanism [7]. There are two primary assembly mechanisms: one initiated by chromosomes (i.e. chromosome-mediated spindle assembly) and the other via the augmin pathway, an MT-dependent MT nucleation mechanism [8]. The former mechanism occurs via two pathways that depend on the RanGTP complex [9] and the chromosomal passenger complex (CPC) [10]. The Ran GEF RCC1 protein associates with chromatin and generates a RanGTP concentration gradient that decreases from the center of the chromosome towards the periphery [11,12,13]. By strongly binding importin-β, RanGTP promotes the release of spindle assembly factors (SAFs) from importin-α/-β, facilitating MT nucleation, stability and assembly around chromosomes. Kinetochore-related CPC, consisting of the mitotic kinase Aurora B in a complex with INCENP, Survivin and Borealin, promotes MT stability around chromosomes. Aurora B-dependent phosphorylation and inhibition of MT instability factors such as MCAK and OP18 create a favorable microenvironment for MT assembly. Previously, Aurora B CPC was described to play a crucial role in the maturation of oocytes in Drosophila melanogaster and Xenopus laevis [14], in MT stabilization and spindle formation [15].
BmN4 cells derived from Bombyx mori ovarian tissue are highly heteroploidy and polyploid cell lines [16], and the holo-centromere of BmN4 cells has lost the traditional centromeric epigenetic marker CENH3 and the kinetochore protein CENPC [17]. In contrast to the typical monocentric chromosomes, silkworm’s CPC and chromatin-driven spindles are located together on the upper side of the prometaphase chromosomes without forming the typical biaxial spindles. RNAi of the gene encoding Aurora B CPC in BmN4 cells (i.e., BmAurora B) arrests cell cycle progression in prometaphase and disrupts the microtubule network of chromatin-controlled spindles [18]. The mitotic spindle of BmN4 cells was recently analyzed using live-cell imaging and shows a square shape reminiscent of the square spindle of holocentric plant cells [19]. The contribution of BmAurora B to this spindle architecture needs further verification. In the present study, we examined the architecture and dynamics of spindle microtubules during prophase and metaphase in BmN4 cells and those with siRNA-mediated BmAurora B knockdown by immunofluorescence labelling. In addition, we performed qRT-PCR expression analysis of BmAurora B in different larval tissues and throughout the 5th larval instar to gain new insights into its regulation during silkworm growth and development. Our results suggest that BmAurora B is crucial for the formation of multipolar square spindles in silkworm holocentric chromosomes, possibly through the activation of γ-tubulin complexes in multiple centrosome-like MTOCs.
2. Materials and Methods
2.1. Cell Culture and Insects
BmN4 cells from silkworm ovaries were cultured at 27°C in TC-100 insect cell culture medium (BBI Life Sciences, Shanghai), supplemented with 10% fetal calf serum (Gibco, Invitrogen, Shanghai).
The inbred silkworm strain Dazao (P50) was maintained at Nanyang Normal University, reared on fresh white mulberry leaves at 25°C ± 2°C under a 12-hour photo phase. For spatiotemporal analysis of BmAurora B gene expression, fifth instar larvae were dissected daily until molting and samples of the body wall, epidermis, midgut, Malpighian tubules, fat body, silk gland, testis, and ovary were collected, snap-frozen in liquid nitrogen and stored at -80°C until use.
2.2. Immunofluorescence
BmN4 cells on 12-well glass coverslips were first fixed using 4% paraformaldehyde for 10 min at room temperature, then permeabilized with 0.25% Triton X-100 for 5 min, and subsequently blocked using 3% BSA for 30 min. α-Tubulin staining used α-tubulin mouse monoclonal antibody (Wuhan Sanying Biotechnology Co., Ltd.) at 1:300 dilution, followed by Alexa Fluor 488-conjugated goat anti-mouse secondary antibody (Jackson ImmunoResearch) at 1:1000 dilution. Meanwhile, γ-tubulin was stained with γ-tubulin polyclonal antibody (Wuhan Sanying Biotechnology Co., Ltd.) at 1:400 dilution, followed by Alexa FluorR 594-conjugated goat anti-rabbit secondary antibody (Jackson ImmunoResearch) at 1:1000 dilution. For H3Thr3ph staining, we used a custom rabbit polyclonal anti-H3Thr3ph antibody at 1:500 dilution, obtained by immunizing rabbits with a chemically-synthesized peptide AR(pT)KQTARKC of H3Thr3ph protein (GL Biochem, Shangai, China), followed by Alexa FluorR 594-conjugated goat anti-rabbit secondary antibody (Jackson ImmunoResearch) at 1:1000 dilution. In all cases, cells were incubated overnight at 4°C with the primary antibody and for 2 hours at 37°C with the secondary antibody. Each step was followed by a thorough triple rinse with PBS 1X (137 mM NaCl; 10 mM Na2HPO4; 1.8 mM KH2PO4; 2.7 mM KCl; pH 7.4). Finally, nuclei were stained with 4’,6-diamino-2-phenylindole (DAPI; 0.01 mg/mL in 90% glycerol). Fluorescence images were acquired using a ZEISS inverted fluorescence microscope Axio Observer 7.
2.3. Quantitative RT-PCR (qRT-PCR)
Total RNA was extracted from silkworm samples using TRIZOL (TaKaRa, DaLian, China) and subjected to DNase I treatment to remove potential DNA contamination. Total RNA concentration was measured using the NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA). Complementary DNA (cDNA) was then synthesized from 2 μg of total RNA using the SuperScript™ III First-Strand Synthesis System (Invitrogen, Shanghai, China).
SYBR green real-time PCR master mix (Applied Biosystems, Foster City, CA) was used for 20-μl qRT-PCR reactions. Each reaction contained 10 μl of the 2x concentrated SYBR green real-time PCR master mix, 5 pmol of each primer (Table 1), and 0.2 μg of cDNA template. The PCR was performed in triplicate on an ABI 7500 Real-Time PCR System (Applied Biosystems). The reactions followed a program consisting of incubation at 50°C for 2 min, then at 95°C for 15 min, followed by 40 cycles of denaturation at 95°C for 15 s, annealing at 60°C for 30 s, and extension and read-out at 72°C for 30 s. The relative expression levels were calculated using the 2−ΔΔCt method [20]. The silkworm cytoplasmic actin A3 gene (Bmactin3, GenBank accession No. U49854) was used as an internal reference control.
2.4. Synthesis of BmAurora B-specific siRNA fragments
Four siRNA fragments, sense (-s) and antisense (-as) strands, targeting the open reading frame of BmAurora B (GenBank Acc. No. NM_001287844) were designed using the free online tool siDESIGN Center (https://horizondiscovery.com/en/ordering-and-calculation-tools/sidesign-center). The BmAurora B-specific siRNA fragments (siBmAurora B) and a (FAM)-labelled Negative Control siRNA (NC-FAM), used for easy monitoring of siRNA uptake by fluorescence microscopy, are shown in Table 1 and were purchased from Oligobio (Beijing, China).
2.5. Cell transfection
The BmN4 cells were inoculated into 12-well cell culture plates 24 h before transfection. The cells were transfected with FAM-labelled NC siRNA and BmAurora B-specific siRNA fragments using the transfection reagent LIPO8000 (Beyotime Biotech Company, Shanghai, China), according to the manufacturer’s instructions. A concentration of 50nM siRNA per well plate was used for transfection. The transfected cells were incubated for 48 h to promote BmAurora B turnover. Subsequently, transfection efficiency was assessed by immunofluorescence and qRT-PCR analysis as described above.
2.6. Statistics
One-way analysis of variance (ANOVA) in conjunction with Tukey’s post-test at P < .05 as a significance level was performed to determine differences between siRNA treatments during qRT-PCR analysis. Three biological replicates were used for each experiment, and similar results were obtained. The standard error of the means was used to compare replicates.
3. Results
3.1. Spindle microtubule assembly in BmN4 cells
In early prophase, immunofluorescence analysis revealed dispersed α-tubulin signals outside the nucleus with an undefined assembly pattern, accompanied by low levels of the chromosome condensation marker, histone H3 Thr3 phosphorylated (H3Thr3ph) (Figure 1A). As prophase progressed, α-tubulin formed a ring-shaped signal with uneven brightness around the nuclear membrane. Simultaneously, scattered H3Thr3ph foci emerged at specific chromatin sites. Notably, the α-tubulin signal was most intense at two particular sites, indicating the onset of spindle microtubule assembly (Figure 1B). In metaphase, α-tubulin had formed bright regions around the nuclear membrane, and H3Thr3ph had distributed across the chromosome (Figure 1C, D). These results corroborate spindle microtubule assembly spanning prophase to metaphase in BmN4 cells.
3.2. Co-localisation of α-tubulin and γ-tubulin displays square spindles in metaphase BmN4 cells.
To further investigate spindle assembly in BmN4 cells, we conducted α-tubulin and γ-tubulin co-localization analysis. In the early prophase, faint γ-tubulin signals co-localized with α-tubulin were observed during nuclear membrane rupture (Figure 2A). As prophase advanced, γ-tubulin signals increased in both number and intensity. α-Tubulin appeared as multiple bright regions around the nuclear membrane, indicating multiple microtubules organizing centers (MTOCs). Most MTOCs were observed on the nuclear membrane and surrounding the chromatin, potentially assembling α-tubulin on chromatin (Figure 2B).
In metaphase, the γ-tubulin foci increased in size and number. They scattered on both sides of the chromosomes, forming a multipolar square microtubule spindle with chromosomes aligned to the metaphase plate (Figure 2C). Closer inspection revealed that only part of the α-tubulin accumulated at γ-tubulin sites. As this stage progressed, the square structure became more evident, with chromosomes and spindle microtubules interacting more dynamically. Interestingly, only 3 to 4 γ-tubulin foci were detected on both sides of the chromosome of late metaphase cells, which were significantly larger than those previously scattered in early metaphase cells, suggesting possible MTOC fusions (Figure 2D).
3.3. siRNA-mediated BmAurora B silencing in BmN4 cells
BmN4 cells were transfected with fluorescein amidite (FAM)-labelled negative control siRNA (NC-FAM) and four distinct siRNA fragments targeting various regions of BmAurora B mRNA. Forty-eight hours post-transfection, numerous dot-shaped green fluorescent signals from NC-FAM were evident in the cytoplasmic region outside the nuclear membrane (Figure 3A), indicating successful siRNA fragment introduction.
qRT–PCR results demonstrated a significant decrease in BmAurora B mRNA levels with each siRNA fragment. Notably, siAurora B-1 and siAurora B-4 fragments exhibited the most effective knockdown effects, achieving an approximately 85% reduction (Figure 3B). Consequently, these two fragments were selected for further functional analysis.
3.4. BmAurora B-depleted cells do not form square spindles in metaphase
We investigated spindle formation in siAurora B-1- and siAurora B-4-transfected BmN4 cells during prophase and metaphase by colocalization of α-tubulin and γ-tubulin. In prophase, depletion of BmAurora B significantly reduced γ-tubulin complexes, which are critical for spindle microtubule assembly near chromatin. Furthermore, α-tubulins were scattered in siRNA-mediated BmAurora B-depleted cells with obvious difficulties in microtubule nucleation compared to NC-FAM-transfected cells (Figure 4, left panels).
In metaphase, siAurora B-transfected cells showed only two γ-tubulin foci, indicative of spindle poles, in contrast to control cells with multiple γ-tubulin foci distributed on both sides of the chromosome forming a multipolar, square microtubule spindle with chromosomes aligned to the metaphase plate. In BmAurora B-depleted cells, microtubules faced challenges in nucleation, resulting in poorly aligned chromosomes to the metaphase plate. As a consequence, the resultant spindle structure took on a quasi-form, closely resembling the canonical oval spindle typically observed in monocentric organisms (see Figure 4, right panels).
3.5. Spatiotemporal analysis of BmAurora B expression in silkworm larval tissues
To gain new insights into the role of BmAurora B in insect growth and development, we analyzed its expression pattern in different tissues of silkworm larvae at the fifth (last) instar stage by quantitative RT-PCR. The results showed that BmAurora B was highly expressed in the testis, followed by the body wall, ovary and fat body, while the expression was very low in the silk gland, midgut, Malpighian tubules and epidermis of the larvae on the third day of the fifth instar (5L3d) (Figure 5A). Further analysis of BmAurora B expression in the testis and ovary during the entire fifth instar stage showed fluctuating accumulation in the testis (Figure 5B) and progressive accumulation in the ovary (Figure 5C), with both reaching highest levels at the onset of ecdysis.
4. Discussion
In the present study, we have characterized the multipolar square spindle of mitotic BmN4 cells using immunofluorescence experiments. The multipole formed by γ-tubulin ring complexes eventually fused into a bipolar or near-bipolar state, which is crucial for the successful division of later sister chromatids into two poles. Previously, Mon et al. (2014) [18] observed that the spindle in BmN4 cells of silkworm was positioned on the upper side of the prometaphase chromosome, and α-tubulin antibody immunofluorescence displayed typical biorientation at metaphase. These findings align with the positioning results of α-tubulin obtained in our study. However, our α-tubulin and γ-tubulin colocalization analysis revealed that chromatin-mediated spindle assembly in metaphase BmN4 cells was multipoint, with multiple γ-tubulin foci still present.
The formation of multipolar square spindles in mitotic BmN4 cells was impaired after siRNA-mediated knockdown of the BmAurora B gene. In a Drosophila S2 cell line, silencing of the Aurora B gene resulted in multipolar spindle formation in some cells in early mitosis and reduced microtubule density in the central region of the spindle in late mitosis, preventing cytokinesis and polyploid formation [21]. This shows that Aurora B gene is involved in spindle microtubule assembly. The different phenotypes in BmN4 and S2 cell lines could be associated with the latter having monocentric chromosomes, whereas the chromosome of BmN4 cells is a diffuse centromeric chromosome without CENH3 or CENPC. In Caenorhabditis elegans, a nematode with the same diffuse centromeric chromosome, Aurora B, even when inactivated after mitosis, disrupts cytokinesis and causes defects in apical structure, suggesting that it is involved in the regulation of epithelial polarization after cytokinesis [22]. Aurora B can also promote sister chromatid adhesion release in late-stage hypothyroid oocytes by phosphorylating the adhesion protein subunit Rec8 [23]. Recent studies have shown that the kinase activity of Aurora B is crucial for acentrosomal-mediated bipolar spindle assembly in nematode cells [24]. In our study, siRNA-treated cells showed a decrease in the number of γ-tubulin foci and high α-tubulin dispersion, indicating obvious difficulties in microtubule nucleation compared to untreated control cells. These results suggest that BmAurora B-mediated phospho-regulation is crucial for the organization of multipolar square spindles in holocentric BmN4 cells, not described previously, probably through direct or indirect phosphorylation of downstream effectors involved in de novo assembly of γ-tubulin ring complexes and spindle formation.
In BmN4 cells, mitotic microtubule generation depends on the formation of the γ-tubulin ring complex. This protein complex, composed of γ-tubulin complex proteins (GCP family proteins), forms connections between its components, creating a structure in which a ring of γ-tubulin molecules mimics the plus end of a microtubule, serving as a crucial starting point for microtubule assembly [25]. Mitotic kinases are involved in the activation of several GCP proteins that regulate various processes at the γ-tubulin ring complexes, such as biogenesis and duplication of centrioles [26,27].
Some spindle proteins, particularly the motor kinesin proteins, are also regulated by mitotic kinases. In Caenorhabditis elegans, phosphorylation of Kin-5 by Aurora B kinase is crucial for its activation [28]. Interestingly, Kin-5 and Kin-14 have already been shown to be important orchestrators that play a crucial role in the organization of the square spindle assembly in BmN4 cells [29]. In addition, Aurora B can phosphorylate and activate other kinases during mitosis, such as the CPC INCENP [30] and Polo-like kinase 1 (PlK1), which in turn phosphorylates mitotic centromere-associated kinesin (MCAK) to stimulate its microtubule depolymerase activity [31]. In both cases, the cascade of phosphorylation processes is crucial for accurate chromosome segregation. In another example, the microtubule-destabilizing Oncoprotein 18/Stathmin (Op18) protein is phosphorylated by Aurora B during spindle assembly [32]. Surprisingly, an increase in Aurora B activity is also detrimental to cells, and in yeast, it causes defects in chromosome segregation and spindle assembly checkpoint activation [33]. This interconnected network of molecular interactions clearly demonstrates the complexity and precision underlying the regulation of microtubule assembly, in which phosphorylation processes involving the mitotic kinase BmAurora B are of fundamental importance.
Aurora B is constitutively expressed in mitotically active cells and up-regulated in highly proliferative tissues. A previous study [34] revealed high expression of the BmAurora B gene in the testis and ovary of silkworms during the final larval instar. This coincided with the cell proliferation phase of both female and male gametogenesis in silkworms. Building on this research, in our study, we investigated the kinetics of BmAurora B expression during the last larval stage in more detail. In particular, we found different BmAurora B mRNA accumulation patterns in the testis and ovary, indicating differences in mitotic progression between these tissues.
Aurora B kinases, displaying remarkable sequence similarity throughout evolution, possess a highly conserved catalytic domain. This domain exhibits serine/threonine kinase activity, acquired through auto-phosphorylation of a conserved residue at Thr232 upon binding of a co-factor during mitosis [35]. This mitotic kinase plays a central role in orchestrating mitotic progression and exerts its influence by phosphorylating multiple substrates. Mass spectrometry analyses of HeLa cells arrested in the G1 and mitotic phases of the cell cycle have unveiled a staggering array of over 1000 distinct mitotic-regulated phosphoproteins, shedding light on the extensive regulatory network governed by Aurora B in these cells [36]. While similarities and differences in the function of Aurora B between monocentric and holocentric organisms have been described [37], unknown facets remain to be explored. In this context, future studies on the phosphoproteins in BmN4 cells could help to identify new BmAurora B targets to unravel the intricacies of the functions of this kinase during mitosis in holocentric species, providing valuable insights into the broader landscape of cell division regulation.
In summary, we have shown that BmAurora B is crucial for the formation of multipolar square spindles in holocentric BmN4 cells, possibly through the activation of γ-tubulin complexes, and we found that its high expression in the gonads of the last instar larvae follows distinct patterns. Given Aurora B's identification as a marker for cancer cells and its association with multipolar spindles in some cancer cells, further exploring BmAurora B's functions in BmN4 mitosis may offer promising avenues for advances in cancer diagnostics and therapeutics.
Author Contributions
Conceived and designed the experiments, Z.B., L.D. and K.Y.; performed the experiments, Z.B., L.X. and S.M.; analyzed the data, Z.B., L.X. and K.Y.; wrote the paper, Z.B. and C.A-P.; project administration, Z.B. C.A-P., and K.Y.; funding acquisition, Z.B. and L.D. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 31870917, Key Research Project Plan for Higher Education Institutions in Henan Province, grant number 22A180007, Science and Technology Research Projects of Henan Provincial Department of Science and Technology, grant number 232102310396 and Special Funded Projects of Nanyang Normal University, grant number ZX2016001.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
We are grateful for the assistance of all staff members and students in the Henan Key Laboratory of Insect Biology in Funiu Mountain, Henan International Joint Laboratory of Insect Biology, College of Life Science and Agricultural Engineering, Nanyang Normal University, Henan, China.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Helmke, K.J.; Heald, R.; Wilbur, J.D. Interplay between spindle architecture and function. Intern. Rev. Cell Mol. Biol. 2013, 306, 83–125. [Google Scholar] [CrossRef]
- Bornens, M. Centrosome organization and functions. Curr. Opin. Struc. Biol. 2021, 66, 199–206. [Google Scholar] [CrossRef]
- Rosselló, C.A.; Lindström, L.; Eklund, G.; Corvaisier, M.; Kristensson, M.A. γ-Tubulin-γ-tubulin interactions as the basis for the formation of a meshwork. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Akhmanova, A.; Kapitein, L.C. Mechanisms of microtubule organization in differentiated animal cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 541–558. [Google Scholar] [CrossRef]
- Zhang, H.; Dawe, R.K. Mechanisms of plant spindle formation. Chromosome Res. 2011, 19, 335–344. [Google Scholar] [CrossRef]
- Mogessie, B.; Scheffler, K.; Schuh, M. Assembly and positioning of the oocyte meiotic spindle. Annu Rev. Cell Dev. Biol. 2018, 34, 381–403. [Google Scholar] [CrossRef]
- Meunier, S.; Vernos, I. Microtubule assembly during mitosis - from distinct origins to distinct functions? J. Cell Sci. 2012, 125, 2805–2814. [Google Scholar] [CrossRef]
- Meunier, S.; Vernos, I. Acentrosomal microtubule assembly in mitosis: the where, when, and how. Trends Cell Biol. 2016, 26, 80–87. [Google Scholar] [CrossRef]
- Clarke, P.R.; Zhang, C. Spatial and temporal coordination of mitosis by Ran GTPase. Nature Rev. Mol. Cell Biol. 2008, 9, 464–477. [Google Scholar] [CrossRef]
- Carmena, M.; Wheelock, M.; Funabiki, H.; Earnshaw, W.C. The chromosomal passenger complex (CPC): from easy rider to the godfather of mitosis. Nature Rev. Mol. Cell Biol. 2012, 13, 789–803. [Google Scholar] [CrossRef]
- Carazo-Salas, R.E.; Guarguaglini, G.; Gruss, O.J.; Segref, A.; Karsenti, E.; Mattaj, I.W. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature 1999, 400, 178–181. [Google Scholar] [CrossRef]
- Kaláb, P.; Pralle, A.; Isacoff, E.Y.; Heald, R.; Weis, K. Analysis of a RanGTP-regulated gradient in mitotic somatic cells. Nature 2006, 440, 697–701. [Google Scholar] [CrossRef]
- Nachury, M.V.; Maresca, T.J.; Salmon, W.C.; Waterman-Storer, C.M.; Heald, R.; Weis, K. Importin beta is a mitotic target of the small GTPase Ran in spindle assembly. Cell 2001, 104, 95–106. [Google Scholar] [CrossRef]
- Sampath, S.C.; Ohi, R.; Leismann, O.; Salic, A.; Pozniakovski, A.; Funabiki, H. The chromosomal passenger complex is required for chromatin-induced microtubule stabilization and spindle assembly. Cell 2004, 118, 187–202. [Google Scholar] [CrossRef]
- Tseng, B.S.; Tan, L.; Kapoor, T.M.; Funabiki, H. Dual detection of chromosomes and microtubules by the chromosomal passenger complex drives spindle assembly. Dev. Cell 2010, 18, 903–912. [Google Scholar] [CrossRef]
- Pu, X.; Hong, X.; Chen, M.; Lu, C. Studies on mitosis and chromosomes of BmN cells. Science of Sericulture 2003, 29, 136–141, (In Chinese with English abstract). [Google Scholar]
- Drinnenberg, I.A.; deYoung, D.; Henikoff, S.; Malik, H.S. Recurrent loss of CenH3 is associated with independent transitions to holocentricity in insects. eLife 2014, 3, e03676. [Google Scholar] [CrossRef]
- Mon, H.; Lee, J.M.; Mita, K.; Goldsmith, M.R.; Kusakabe, T. Chromatin-induced spindle assembly plays an important role in metaphase congression of silkworm holocentric chromosomes. Insect Biochem. Mol. Biol. 2014, 45, 40–50. [Google Scholar] [CrossRef]
- Vanpoperinghe, L.; Carlier-Grynkorn, F.; Cornilleau, G.; Kusakabe, T.; Drinnenberg, I.A.; Tran, P.T. Live-cell imaging reveals square shape spindles and long mitosis duration in the silkworm holocentric cells. MicroPub. Biol. 1791. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Giet, R.; Glover, D.M. Drosophila aurora B kinase is required for histone H3 phosphorylation and condensin recruitment during chromosome condensation and to organize the central spindle during cytokinesis. J. Cell Biol. 2001, 152, 669–682. [Google Scholar] [CrossRef]
- Bai, X.; Melesse, M.; Sorensen Turpin, C.G.; Sloan, D.E.; Chen, C.Y.; Wang, W.C.; Lee, P.Y.; Simmons, J.R.; Nebenfuehr, B.; Mitchell, D.; Klebanow, L.R.; Mattson, N.; Betzig, E.; Chen, B.C.; Cheerambathur, D.; Bembenek, J.N. Aurora B functions at the apical surface after specialized cytokinesis during morphogenesis in C. elegans. Dev. (Cambridge, England) 2020, 147. [Google Scholar] [CrossRef]
- Ferrandiz, N.; Barroso, C.; Telecan, O.; Shao, N.; Kim, H.M.; Testori, S.; Faull, P.; Cutillas, P.; Snijders, A.P.; Colaiácovo, M.P.; Martinez-Perez, E. Spatiotemporal regulation of Aurora B recruitment ensures release of cohesion during C. elegans oocyte meiosis. Nat. Commun. 2018, 9, 834. [Google Scholar] [CrossRef]
- Divekar, N.S.; Davis-Roca, A.C.; Zhang, L.; Dernburg, A.F.; Wignall, S.M. A degron-based strategy reveals new insights into Aurora B function in C. elegans. PLoS Genet. 2021, 17, e1009567. [Google Scholar] [CrossRef]
- Oakley, B.R.; Paolillo, V.; Zheng, Y. γ-Tubulin complexes in microtubule nucleation and beyond. Mol. Biol. Cell 2015, 26, 2957–2962. [Google Scholar] [CrossRef]
- Bahtz, R.; Seidler, J.; Arnold, M.; Haselmann-Weiss, U.; Antony, C.; Lehmann, W.D.; Hoffmann, I. GCP6 is a substrate of Plk4 and required for centriole duplication. J. Cell Sci. 2012, 125 Pt 2, 486–496. [Google Scholar] [CrossRef]
- Farache, D.; Emorine, L.; Haren, L.; Merdes, A. Assembly and regulation of γ-tubulin complexes. Open Biol. 2018, 8. [Google Scholar] [CrossRef]
- Bishop, J.D.; Han, Z.; Schumacher, J.M. The Caenorhabditis elegans Aurora B kinase AIR-2 phosphorylates and is required for the localization of a BimC kinesin to meiotic and mitotic spindles. Mol. Biol. Cell. 2005, 16, 742–756. [Google Scholar] [CrossRef]
- Kassamaly, I.T.; Cornilleau, G.; Drinnenberg, I.A.; Tran, P.T. Kinesin-5 and kinesin-14 are partially antagonistic in spindle assembly in the holocentric silkworm B. mori cells. MicroPubl. Biol. 0006. [Google Scholar] [CrossRef]
- Honda, R.; Körner, R.; Nigg, E.A. Exploring the functional interactions between Aurora B, INCENP, and survivin in mitosis. Mol. Biol. Cell 2003, 14, 3325e3341. [Google Scholar] [CrossRef]
- Shao, H.; Huang, Y.; Zhang, L.; Yuan, K.; Chu, Y.; Dou, Z.; Jin, C.; Garcia-Barrio, M.; Liu, X.; Yao, X. Spatiotemporal dynamics of Aurora B-PLK1-MCAK signaling axis orchestrates kinetochore bi-orientation and faithful chromosome segregation. Sci. Rep. 2015, 5, 12204. [Google Scholar] [CrossRef]
- Gadea, B.B.; Ruderman, J.V. Aurora B is required for mitotic chromatin-induced phosphorylation of Op18/Stathmin. Proc. Natl. Acad. Sci. USA. 2006, 103, 4493–4498. [Google Scholar] [CrossRef]
- Muñoz-Barrera, M.; Monje-Casas, F. Increased Aurora B activity causes continuous disruption of kinetochore–microtubule attachments and spindle instability. Proc. Natl. Acad. Sci. USA. 2014, 111, E3996–E4005. [Google Scholar] [CrossRef]
- Gang, X.; Qian, W.; Zhang, T.; Yang, X.; Xia, Q.; Cheng, D. Aurora B kinase is required for cell cycle progression in silkworm. Gene 2017, 599, 60–67. [Google Scholar] [CrossRef]
- Willems, E.; Dedobbeleer, M.; Digregorio, M.; Lombard, A.; Lumapat, P.N.; Rogister, B. The functional diversity of Aurora kinases: a comprehensive review. Cell Div. 2018, 13, 7. [Google Scholar] [CrossRef]
- Dephoure, N.; Zhou, C.; Villén, J.; Beausoleil, S.A.; Bakalarski, C.E.; Elledge, S.J.; Gygi, S.P. A quantitative atlas of mitotic phosphorylation. Proc. Natl. Acad. Sci. USA. 2008, 105, 10762–10767. [Google Scholar] [CrossRef]
- Collette, K.S.; Petty, E.L.; Golenberg, N.; Bembenek, J.N.; Csankovszki, G. Different roles for Aurora B in condensin targeting during mitosis and meiosis. J. Cell Sci. 2011, 124 Pt 21, 3684–3694. [Google Scholar] [CrossRef]
Figure 1.
Chromatin-induced spindle assembly in the early prophase of holocentric BmN4 cells. Immunofluorescence staining of α-tubulin (green) and histone H3 Thr3 phosphorylated (H3Thr3ph, red) in prophase (A and B) and metaphase (C and D) of mitosis. Panels with the chromatin staining with DAPI in blue were merged (left panels) to show the overlap of the two fluorescence patterns. Scale bar, 5 μm.
Figure 1.
Chromatin-induced spindle assembly in the early prophase of holocentric BmN4 cells. Immunofluorescence staining of α-tubulin (green) and histone H3 Thr3 phosphorylated (H3Thr3ph, red) in prophase (A and B) and metaphase (C and D) of mitosis. Panels with the chromatin staining with DAPI in blue were merged (left panels) to show the overlap of the two fluorescence patterns. Scale bar, 5 μm.
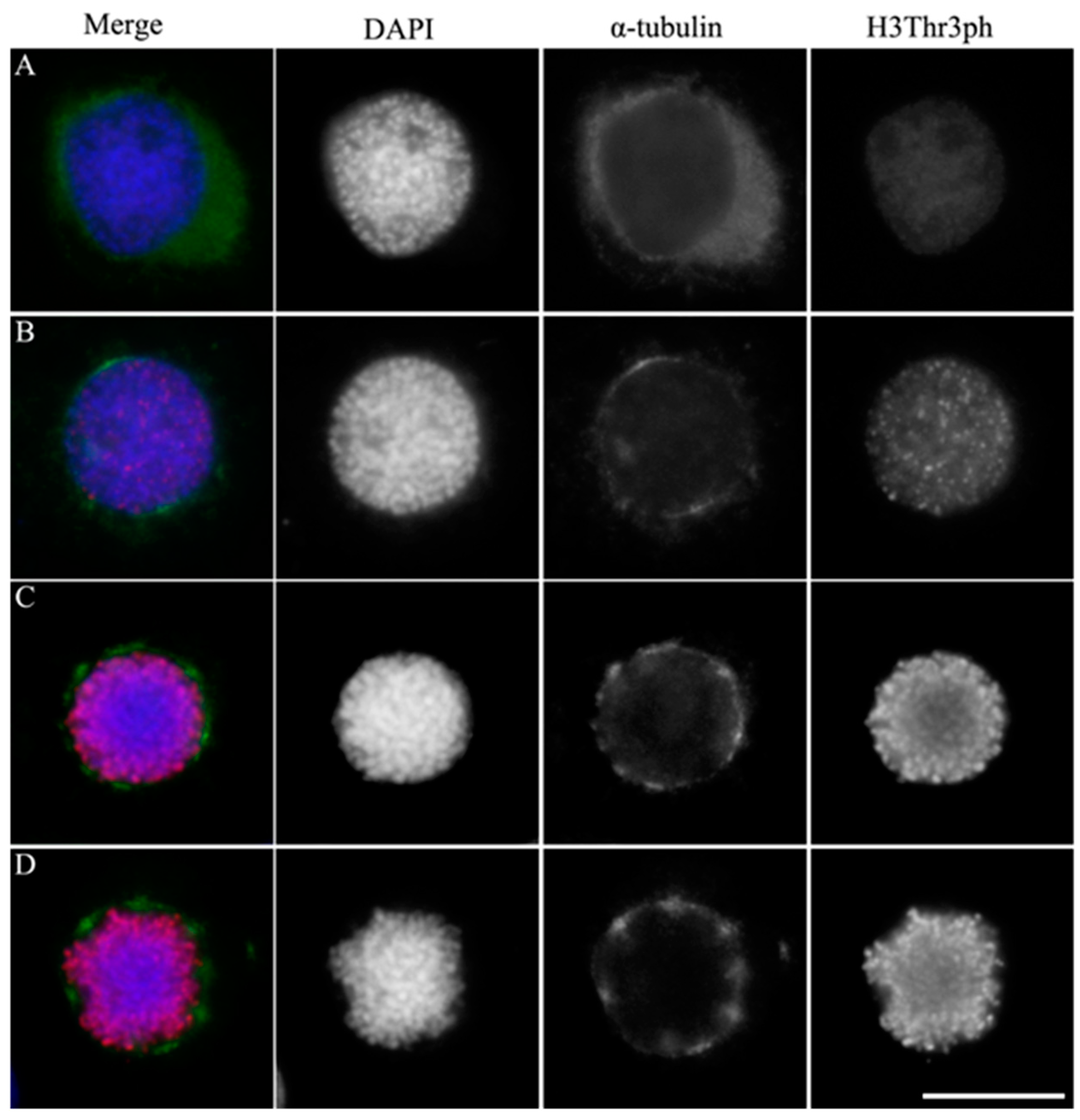
Figure 2.
Mitotic spindle assembly dynamics in holocentric BmN4 cells. Immunofluorescence staining of α-tubulin (green) and γ-tubulin (red) in prophase (A and B) and metaphase (C and D). Panels with the chromatin staining with DAPI in blue were merged (left panels) to show the overlap of the two fluorescence patterns. Scale bar, 5 μm.
Figure 2.
Mitotic spindle assembly dynamics in holocentric BmN4 cells. Immunofluorescence staining of α-tubulin (green) and γ-tubulin (red) in prophase (A and B) and metaphase (C and D). Panels with the chromatin staining with DAPI in blue were merged (left panels) to show the overlap of the two fluorescence patterns. Scale bar, 5 μm.
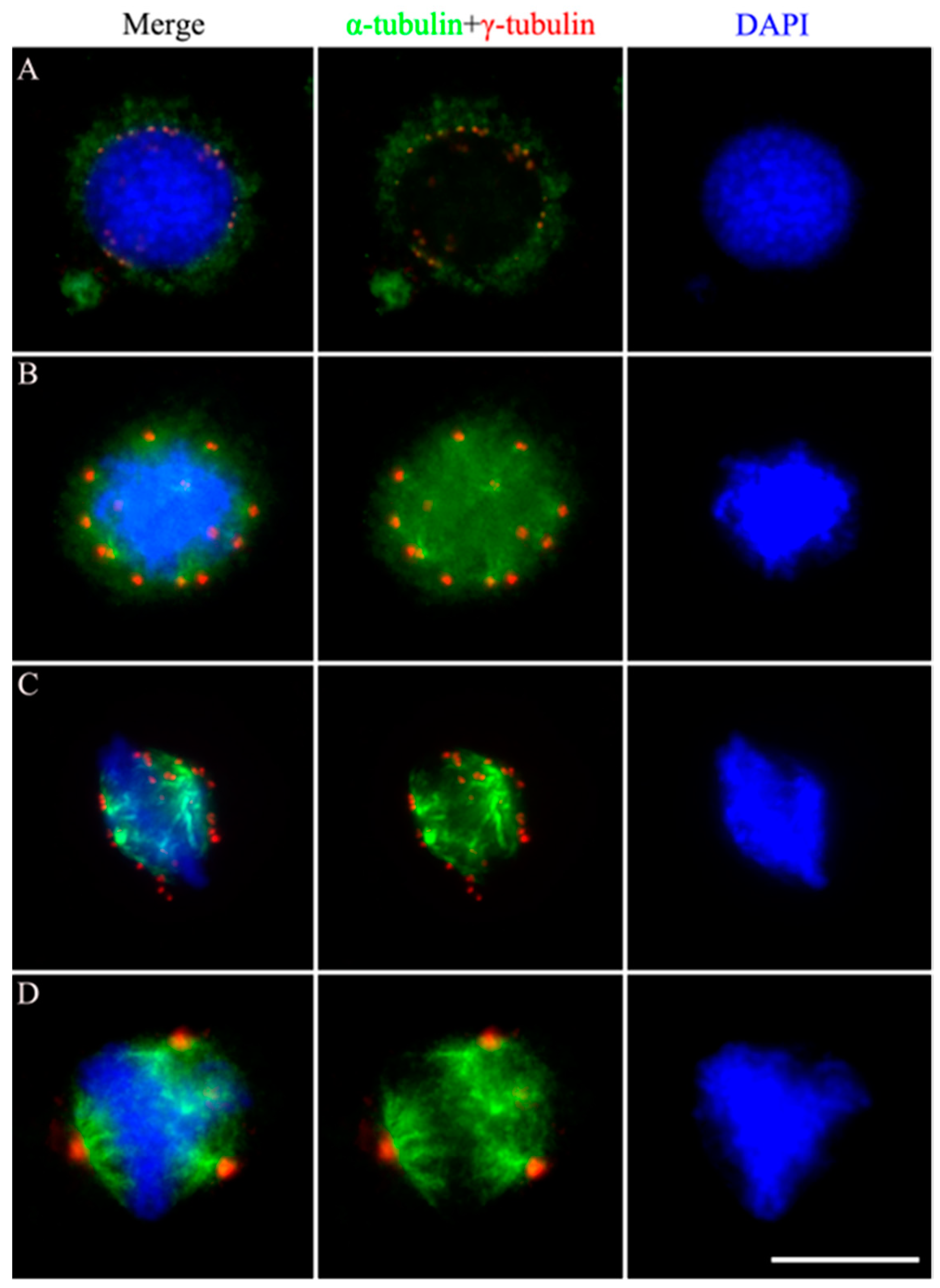
Figure 3.
Confirmation of transient transfection efficiency. (A) Green fluorescence signals in BmN4 cells transfected with fluorescein amidite (FAM)-labelled negative control siRNA (NC-FAM) for 48 h. Cells were stained with DAPI in blue. Scale bar, 5 μm. (B) Real-time quantitative polymerase chain reaction (qRT-PCR) of BmAurora-B after the transfection of specific siRNA (siAurora B-1, siAurora B-2, siAurora B-3, siAurora B-4) and NC-FAM (NC). The silkworm actinA3 gene was used as an internal control. Bars represent means ± SD. Different letters indicate significant differences (P < .05).
Figure 3.
Confirmation of transient transfection efficiency. (A) Green fluorescence signals in BmN4 cells transfected with fluorescein amidite (FAM)-labelled negative control siRNA (NC-FAM) for 48 h. Cells were stained with DAPI in blue. Scale bar, 5 μm. (B) Real-time quantitative polymerase chain reaction (qRT-PCR) of BmAurora-B after the transfection of specific siRNA (siAurora B-1, siAurora B-2, siAurora B-3, siAurora B-4) and NC-FAM (NC). The silkworm actinA3 gene was used as an internal control. Bars represent means ± SD. Different letters indicate significant differences (P < .05).

Figure 4.
Effects of siRNA-mediated BmAurora silencing B on the mitotic spindle assembly dynamics in holocentric BmN4 cells. Immunofluorescence staining of alpha-tubulin (green) and gamma-tubulin (red) signals in siRNA-transfected BmN4 cells in prometaphase and metaphase. Chromatin was stained with DAPI in blue. Scale bar, 5 μm.
Figure 4.
Effects of siRNA-mediated BmAurora silencing B on the mitotic spindle assembly dynamics in holocentric BmN4 cells. Immunofluorescence staining of alpha-tubulin (green) and gamma-tubulin (red) signals in siRNA-transfected BmN4 cells in prometaphase and metaphase. Chromatin was stained with DAPI in blue. Scale bar, 5 μm.
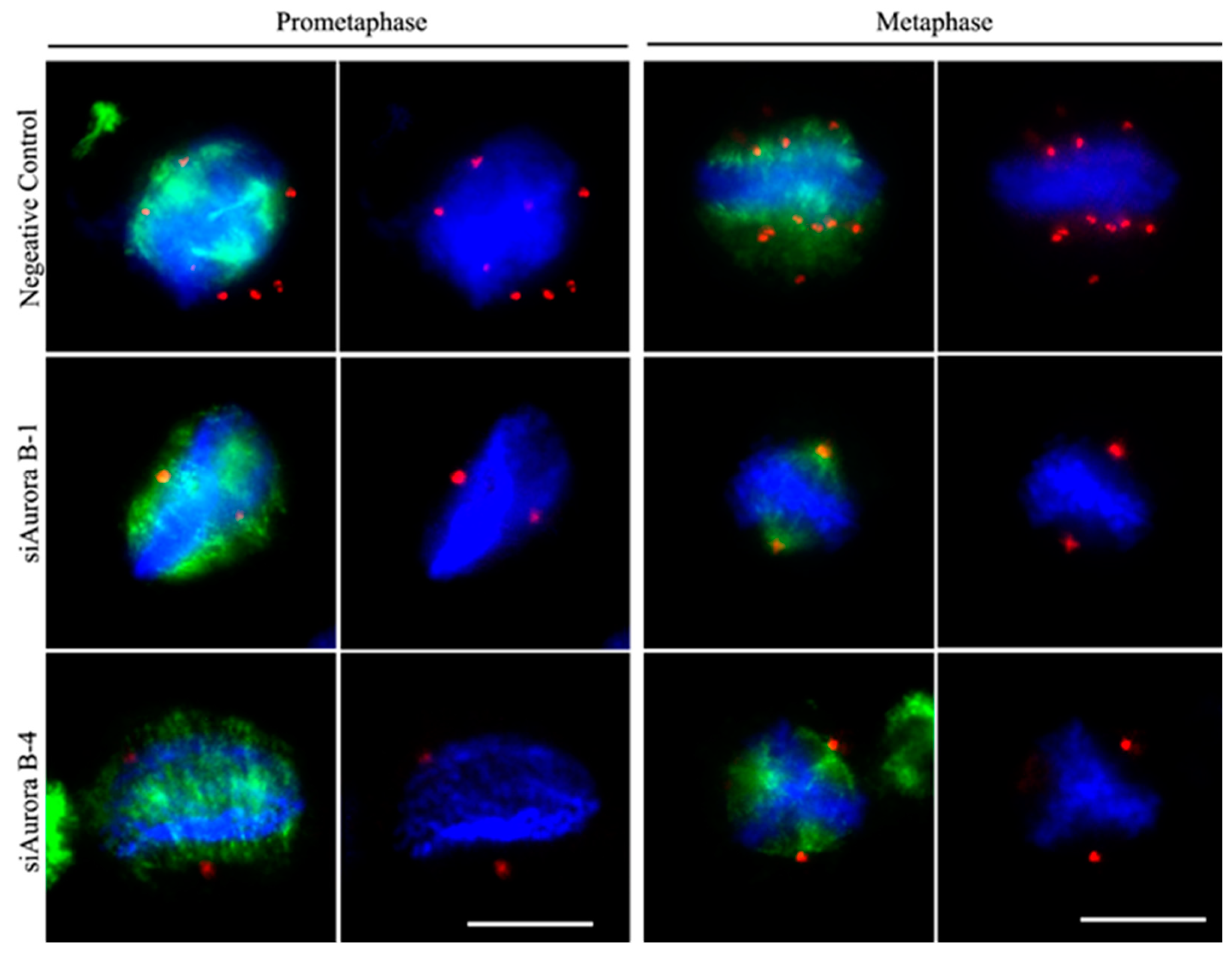
Figure 5.
Spatiotemporal BmAurora B expression analysis in 5th instar larvae of Bombyx mori by qRT-PCR. (A) Quantification of BmAurora B mRNA in multiple tissues of silkworm larvae on day 3 of the fifth instar (5L3d). (B) Quantification of BmAurora B mRNA in the testis from day 1 of the 5th instar larvae (5L1d) to the moulting stage (5LM). (C) Quantification of BmAurora B mRNA in the ovary from day 1 of the 5th instar larvae (5L1d) to the molting stage (5LM). Values represent the relative mRNA expression with respect to the internal control. The data represent the mean ± SD of the triplicates.
Figure 5.
Spatiotemporal BmAurora B expression analysis in 5th instar larvae of Bombyx mori by qRT-PCR. (A) Quantification of BmAurora B mRNA in multiple tissues of silkworm larvae on day 3 of the fifth instar (5L3d). (B) Quantification of BmAurora B mRNA in the testis from day 1 of the 5th instar larvae (5L1d) to the moulting stage (5LM). (C) Quantification of BmAurora B mRNA in the ovary from day 1 of the 5th instar larvae (5L1d) to the molting stage (5LM). Values represent the relative mRNA expression with respect to the internal control. The data represent the mean ± SD of the triplicates.
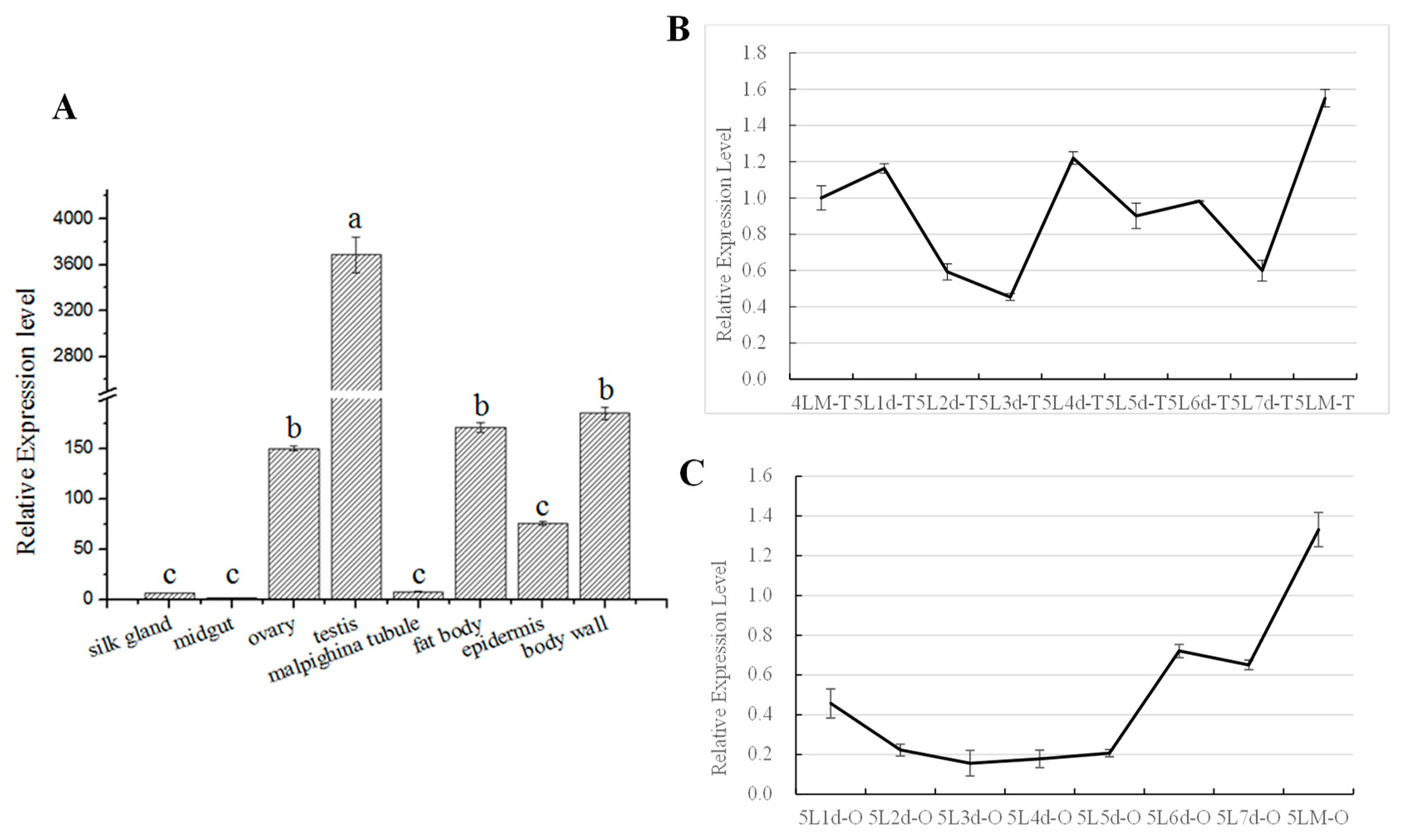

Table 1.
Primers used in this study.
| Primer | Primer sequence (5’-3’) | Purpose |
|---|---|---|
| BmAurora B-RT-F | GGCCAAGGCAAATTCGGACATGTT | qRT-PCR |
| BmAurora B-RT-R | CGTCCTTGGGGTGAATTTGTGAGATGTT | |
| BmA3-RT-F | ATGTGCGACGAAGAAGTTGC | |
| BmA3-RT-R | GTCTCCTACGTACGAGTCCT | |
| siBmAurora B-1-s | CCAGAAAGUAAAGCAGCAATT | siRNA synthesis |
| siBmAurora B-1-as | UUGCUGCUUUACUUUCUGGTT | |
| siBmAurora B-2-s | GGGAAAGCCUCCAUUUGAATT | |
| siBmAurora B-2-as | UUCAAAUGGAGGCUUUCCCTT | |
| siBmAurora B-3-s | CCUGAUGGAGCCAAGGAUUTT | |
| siBmAurora B-3-as | AAUCCUUGGCUCCAUCAGGTT | |
| siBmAurora B-4-s | GCCAAGGAUUUGAUCUCAATT | |
| siBmAurora B-4-as | UUGAGAUCAAAUCCUUGGCTT | |
| NC-FAM-s | UUCUCCGAACGUGUCACGUTT | |
| NC-FAM-as | ACGUGACACGUUCGGAGAATT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



