
Submitted:
18 December 2023
Posted:
20 December 2023
You are already at the latest version
Abstract
We investigated the prevalence and genetic diversity of Wolbachia pipientis strains in Aedes albopictus populations in Greece. Using a combination of PCR and Sanger sequencing techniques, we genotyped Wolbachia strains in 105 mosquitoes collected across eight different prefectures in 2021. We found a high prevalence of Wolbachia in both male (90%) and female (97%) mosquitoes. Among the infected samples, 84% had double infections with both wAlbA and wAlbB strains, while 16% had infections with only wAlbB. Our comparison of the Multi-Locus Sequence Typing (MLST) profile, employing gatB-coxA-hcpA-ftsZ-fbpA genotyping, revealed a single MLST profile for each wAlbA and wAlbB strain in Greek populations. The same MLST profiles were also reported in populations from China, Russia, and Argentina, suggesting low levels of global diversity in wAlbA and wAlbB strains. Furthermore, our results indicated a significant association between temperature and the prevalence of single infections (P = 6.498E-07), with higher temperatures correlating with an increased likelihood of single infections. Although male-bias showed a tendency towards single infections, the effect was marginally non-significant (P = 0.053). These results were confirmed using a bootstrap-with-replacement analysis approach. Overall, our findings offer novel insights into the distribution and species diversity of Wolbachia strains in Greek Ae. albopictus populations, emphasizing the importance of understanding the short-term plastic and adaptive responses of these organisms to environmental stressors and rapid climate change.
Keywords:
Asian tiger mosquito
; Insect-borne disease
; Microbial ecology
; Microbiome dynamics
; pathogen vector
; MLST typing
; Wolbachia supergroups
Introduction
The Asian tiger mosquito, Aedes albopictus, is a highly invasive mosquito endemic to Southeast Asia, and its adaptation to urban environments has led to its global spread [1]. The first detection of this mosquito in Greece occurred in Corfu and Thesprotia in 2003/4, and since then, it has been found in nearly every prefecture [2,3]. Due to its ability to transmit many important human pathogens, such as Zika, Chinkungunya (CHIKV), Dengue (DENV- 1, 2, 3, and 4), and West Nile (WNV) viruses, the Asian tiger mosquito has attracted substantial research interest [4,5]. With traditional use of chemical insecticides raising serious concern about toxic effects on organisms and ecosystems, as well as, about the increasing mosquito resistance, alternative environment-friendly methods for the control of mosquito populations have arisen [6]. The Sterile Insect Technique (SIT), a genetic control method that has been used against agricultural pests since decades, is reconsidered as a promising approach against disease transmitting mosquito species, including Ae. albopictus, exhibiting considerable progress towards this goal in recent years [7]. Furthermore, symbiont-based approaches are gaining more interest as promising tools for vector control [8].
Wolbachia pipientis is a Gram-negative, intracellular, endosymbiotic bacterium found in several arthropod species and filarial nematodes. It is classified into 17 supergroups A-R (except G) based on genomic diversity [9]. Supergroups A and B are the most prevalent found in arthropod species [10]. Initial reports proposed that 20% of arthropod species are infected by Wolbachia [11,12], but a later meta-analysis suggested that the number is closer to 60% [13]. The empirical evidence regarding the intraspecific prevalence of Wolbachia infection seems to follow an “all or nothing” pattern, with either very low (<10%) or very high (>90) infection rates in each species [13]. The spread of Wolbachia in arthropod species is closely linked to the manipulations and phenotypes that the bacterium induces in its hosts. The most frequently observed reproductive phenotype is cytoplasmic incompatibility (CI). The maternally inherited Wolbachia strain induces a modification during spermatogenesis in males, which can be reversed if a compatible strain from the mated female is present in the embryo. If the strains are not compatible, the eggs will not hatch. It is important to note that a non-infected male is compatible with any infected female. This phenotype explains the ability of Wolbachia to establish infections in natural populations with a high prevalence, close to 100% [10,14,15]. In mosquitoes, another important Wolbachia-associated observed phenotype is pathogen interference (PI). Certain Wolbachia strains can reduce the replication and transmission of specific viruses and other pathogens. This phenotype has been observed in natural infections and has been most successfully demonstrated through transfections [10,16,17]. [10,16,17] In recent years, Wolbachia has emerged as a promising tool for reducing mosquito-transmitted diseases, through the introduction of appropriately infected mosquitoes that can exert CI or PI effects in natural populations [8]. It is widely accepted that careful consideration and thorough research on the effects of a particular Wolbachia strain in a specific mosquito species must precede the release of mosquitoes in nature to prevent unintended consequences. Furthermore, extensive screening of natural populations for Wolbachia prevalence and the strain composition is an equally important step before any Wolbachia-based control effort can be considered [8].
In the natural populations of Ae. albopictus tested, mainly in Asia and south America, Wolbachia infection is established with a high prevalence (>95%) [18,19,20,21]. Two commonly found Wolbachia strains in Ae. albopictus are wAlbA and wAlbB, belonging to supergroups A and B, respectively [22]. These strains often co-infect the same individual, although single infections with wAlbB, more commonly observed in males, have also been reported [23]. Enviromental factors have been shown to affect the density of the Wolbachia strains in A. albopictus, with higher temperature and lower nutrition resulting to lower Wolbachia density [24]. Genetic diversity of the wAlbA and wAlbB strains has been studied by the MLST (MultiLocus Sequence Typing) system [25], a typing system based on the polymorphism of five conserved genes (gatB, coxA, hcpA, ftsZ and fbpA), in natural populations [25]. Both strains were found stable with wAlbA presenting a unique profile and wAlbB two very similar ones [18,25,26,27]. In addition to the naturally occurring Wolbachia strains in Ae. albopictus, wAlbA and wAlbB, several Wolbachia transfections have been explored in the species, including: (a) stable transfection with the wMel strain from Drosophila melanogaster, which showed positive results in blocking DENV and CHIKV transmission [28,29], (b) the wAu strain from Drosophila simulans, which displayed resistance to Zika and DENV with a moderate fitness cost to the mosquitoes [30], and (c) the wPip strain from Culex pipiens, which led to a reduction in field populations when triple-infected (wAlbA, wAlbB and wPip) (line HC) or single infected (wPip)(line ARwP) male mosquitoes were released in large field trials [31,32,33,34].
To date, the prevalence and genetic diversity of Wolbachia strains in Ae. albopictus populations in Europe remain poorly understood. To our knowledge only one study reports a prevalence of 79% in field population of the Iberian Peninsula, with no information provided about the type of stains (wAlbA or wAlbB) or genotypic diversity [35]. Apart from a field trial of the IIT method with the ARwP line in Italy [34], no Wolbachia-based management practices are currently underway in Europe. Notably, most cases of viruses transmitted by Ae. albopictus are not autochthonous [36,37]. However, sporadic autochthonous cases of DENV-1 in Italy, Croatia, and southern France [38,39,40,41], as well as CHIKV outbreaks in Italy and France [40,41,42,43,44], and WNV cases in Italy, Germany, the Netherlands, France [45,46,47,48,49,50,51], have been reported. The frequency of Ae. albopictus-transmitted diseases is expected to increase in Europe in the coming years, driven by climate change [52,53,54], and thus increased efforts should be made to study and control the mosquito populations, including the use of Wolbachia-based methods.
In this study, our aim was to elucidate the unknown prevalence of Wolbachia in Ae. albopictus mosquitoes in Greece. As mentioned, Ae. albopictus was first reported in Greece relatively recently (around 2003), and in the subsequent years, its populations have expanded throughout the country [3]. We also aimed to determine the detected strains, wAlbA or wAlbB, and the occurrence of double and single infections. Additionally, we sought to shed light on the genetic diversity of wAlbA or wAlbB strains in northeastern Europe by comparing our MLST genotypes with those in other studies. Furthermore, we recorded the sex of the mosquitoes and the temperature environment of the studied populations to provide empirical evidence regarding the association between these factors and the propensity for double or single infections. Our experimental design involved a field sampling campaign across eight different prefectures in Greece between July and September 2021, during which we collected a total of 114 adult Ae. albopictus mosquitoes. The sex of each mosquito was recorded, and we employed PCR and Sanger sequencing techniques to detect and genotype the Wolbachia strains. To assess the impact of environmental temperature, we collected temperature recordings four times a day for 30 days prior to the sampling date. We then analyzed this data in relation to the observed patterns of occurrence of double/single infections. Overall, our findings provide valuable insights into the distribution and genetic diversity of Wolbachia in Greek Ae. albopictus populations, Furthermore, they highlight the importance of understanding the organisms’ short-term plastic and adaptive responses to environmental stressors and rapid climate change. We anticipate these results to contribute to the development of effective Wolbachia-based approaches for controlling mosquito-borne diseases in Ae. albopictus in Greece but also in other regions.
Materials and methods
Mosquito sampling
Adult mosquitoes were collected between July and September of 2021, in the frame of standard entomological surveillance of mosquito control projects run in four regions of Greece (Central Macedonia, Western Macedonia, Western Greece and Thessaly). For the entomological surveillance, CDC-style traps were used which were supplied with CO2 and UV light to attract mosquitoes with a constant CO2 -release rate of 0.5 l/min. The traps are used for wide-area mosquito control projects all over the country since 2010 and were designed, fabricated, and operated by Ecodevelopment S.A. (https://ecodev.gr/en/ecodev-english/). Random individuals were selected from the hundreds of mosquitoes captured in the traps. These individuals were morphologically identified to species level by scientific personnel of the entomological laboratory of Ecodevelopment using dichotomous identification keys [55,56,57]. Overall, samples from 8 Greek prefectures were obtained: 10 from Achaea, 15 from Aetolia-Acarnania, 9 from Imathia, 8 from Thessaloniki, 27 from Karditsa, 19 from Kozani, 13 from Pieria and 4 from Serres (Table 1). A total of 105 adult Ae. albopictus were analyzed, comprising 74 females (70.5%), 31 males (29.5%) (Supplementary Table 1).
DNA extraction and Wolbachia detection
DNA was extracted from single Ae. albopictus mosquitoes using a modified DNA extraction CTAB protocol [58]. To detect Wolbachia infection, PCR was performed using the wspecF/wspecR primer pair (5΄-YATACCTATTCGAAGGGATAG-3΄/5΄-AGCTTCGAGTGAAACCAATTC-3΄) which amplifies a 438 bp fragment of the 16S rRNA gene [11]. The PCR cycling protocol was as follows: 95 oC for 5 min, followed by 30 cycles of 95 oC for 1 min, 55 oC for 1 min, and 72 oC for 1 min, followed by 72 oC for 5 min [11,59]. The DNA from a Wolbachia-infected Drosophila suzukii laboratory strain was used as a positive control in each reaction. For DNA quality control, the 12SC primer pair (12SCFR: 5΄-GAGAGTGACGGGCGATATGT-3΄/12SCRR: 5΄-AAACCAGGATTAGATACC-CTATTAT-3΄), which amplifies a 377 bp fragment of the mosquito 12S rRNA gene was used. The PCR cycling protocol for this control reaction was: 95 oC for 5 min, followed by 30 cycles of 95 oC for 1 min, 53 oC for 1 min and 72 oC for 1 min, followed by 72oC for 5 min [60,61,62]. Both PCR reactions were carried out in a final volume of 15 μl, containing 1X reaction buffer, 2.5 mM MgCl2, 1 Unit of Taq DNA polymerase (BIOTAQ, Bioline), 0.2 mM dNTPs (Jena Bioscience), 0.5 mM of each primer and 1μl of the template DNA. The PCR products were analyzed by electrophoresis on a 1.5% agarose gel using the 100bp DNA ladder PLUS BLUE (GeneOn). Samples were characterized as negative for Wolbachia only when they repeatedly failed to give an amplification signal for wspec (at least two amplification tests) although producing a strong positive signal for 12S rRNA.
Strain characterization and MLST genotyping
To characterize the Wolbachia subgroup identity in the samples that were positive of Wolbachia presence, two primer pairs were used: wAlbA_328F/wsp691R (5΄-CCAGCAGATACTATTGCG-3΄/5΄-AAAAATTAAACGCTACTC-CA-3΄) for detection of the wAlbA strain, and wAlbB_183F/wsp691R (5΄-AAGGAACCGAAGTTCATG-3΄/5΄-AAAAATTAAACGCTACTCCA-3΄) for detection of the wAlbB strain. The primer pairs produce amplicons of 341 bp and 463 bp, respectively. A multiplex PCR was conducted with the following cycling profile: 95oC for 5 min, followed by 35 cycles of 95oC for 1 min, 55oC for 1 min and 72oC for 1 min, followed by 72oC for 3 min [22]. The PCR reaction was performed in a final volume of 15μl containing 1X reaction buffer, 2.5 mM MgCl2, 1 Unit of Taq DNA polymerase (BIOTAQ, Bioline), 0.2 mM dNTPs (Jena Bioscience), 0.5 mM of each primer and 1μl of the template DNA. The results were analyzed using a 1.5% agarose gel and insects were classified as “double-infected” if they exhibited both wAlbA (341bp) and wAlbB (463bp) specific bands, or as “single-infected” if they showed only a single band.
MLST genotyping was conducted on 11 selected samples: four (two females, two males) double-infected from THE, three (two females, one male) double-infected and one (male) single-infected from KAR, and one (male) double-infected and two (one female, one male) single-infected from AET. For each individual, five loci of the MLST system, as developed for Wolbachia by Baldo et al. (2006) were PCR-amplified and Sanger-sequenced. For four out of the five loci (gatB, hcpA, ftsZ, fbpA) wAlbA and wAlbB strain-specific primers were used to amplify the respective alleles (Baldo et al 2006). For locus coxA the wAlbA-specific primers were used to produce the A allele, while universal primers in wAlbB single-infected individuals were used to amplify the B allele, since the wAlbB-specific primers failed to produce the expected amplicon. The PCR cycling profile was: 94 oC for 2 min, followed by 37 cycles of 94 oC for 30 sec, 55-62 oC for 45 sec and 72 oC for 1.5 min, followed by 72 oC for 10 min (https://pubmlst.org/organisms/Wolbachia-spp). The PCR reactions were performed in a final volume of 20μl, consisting of 1X reaction buffer, 1.5mM MgCl2, 1 Unit of Taq DNA polymerase (BIOTAQ, Bioline), 0.2mM dNTPs (Jena Bioscience), 1mM of each primer and 1μl of the extracted DNA (Gerth et al., 2013). The success of each amplification was evaluated using 1.5% agarose gels. The PCR products were cleaned using the ExoSAP-IT™ (New England Biolabs) protocol and Sanger-sequenced in both directions by GENEWIZ (Takeley, UK). The chromatograms were inspected using the Geneious Prime software version 2023.0.4 (www.geneious.com). The allele number for each locus was determined according to the Wolbachia MLST database (https://pubmlst.org/organisms/Wolbachia-spp). All sequences have been submitted to GenBank (Accession Numbers: OQ985406-OQ985492).
Temperature data collection and statistical analysis
For each sampling location, we calculated the mean temperature from the 30 days prior to the date of sampling. This approach captures the temperature exposure of the mosquito populations over a longer period than the temperature of the day of sampling. Using the latitude and longitude of each trap, we recovered for each day the temperature readings at four time points: 6 o’clock in the morning and in the evening, midday, and midnight. We thus averaged 120 temperature readings for each trap for each sampling event. We thus also accounted for the variability in daily temperature, Temperature data were collected from the OpenWeather platform (www.openweather.org) via its API using the Python package ‘requests’. OpenWeather is a widely used weather data provider used by a variety of industries, including agriculture, transportation, and energy.
To explore, per sampling date, the correlation between the mean temperature, the ratio of single to double Wolbachia infections, and the sex (female, male) of the infected mosquitoes, we employed the non-parametric Kruskal-Wallis test [63] given the limited number of independent observations (n=11). To further examine the effects of different groups, we performed a post-hoc Dunn test [64]. Additionally, to validate our findings and assess their robustness, we utilized a bootstrap-with-replacement approach. We generated 10,000 bootstrap resamples by randomly selecting observations from the original dataset, allowing for duplicates (i.e., with replacement) in each resampled dataset. Kruskal-Wallis and Dunn tests were then performed in each bootstrap resample to assess the effects of temperature and sex ratio. By employing this approach, we were able to estimate the confidence intervals of the observed correlations and strengthen the reliability of our findings.
Results
Wolbachia prevalence
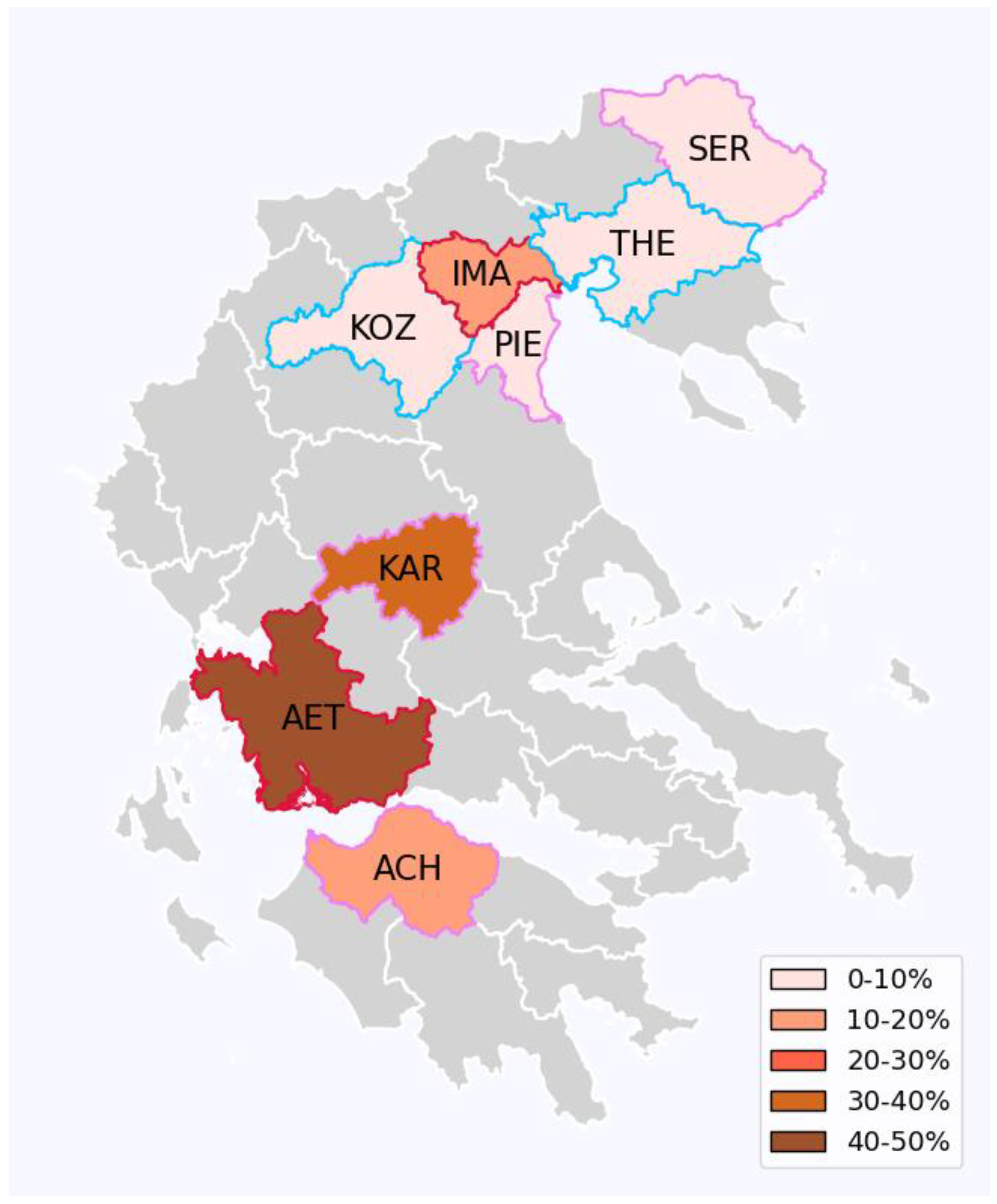
Out of the 105 Ae. albopictus individuals examined, 100 (95%) were found to carry a Wolbachia infection. The prevalence of infection in males was 90% (28/31) and in females it was 97% (72/74). The infection rates in each district prefecture were as follows: 90% in ACH (9/10), 87% in AET (13/15), 93% in KAR (25/27) and 100% in IMA (9/9), THE (8/8), KOZ (19/19), PIE (13/13) and SER (4/4) (Table 2; Supplementary Table 1).
Wolbachia strain characterization
The 100 Wolbachia-infected mosquitoes were screened for wAlbA and wAlbB. The PCR essay detected a double infection (both wAlbA and wAlbB) in 84%, while a single infection with the wAlbB strain was found in 16% of the infected samples. No mosquitoes infected solely with the wAlbA strain were found (Supplementary Table 1). Most female mosquitoes were double infected (68/74 female mosquitoes or 92%), whereas double infection was present in 52% of males (16/31 male mosquitoes). When inspecting the results by prefecture, single infected individuals were only found in four regions: ACH (11%), AET (46%), KAR (32%) and IMA (11%) (Table 2; Supplementary Table 1).
Figure 1.
The distribution of Wolbachia proportion of single (wAlbB) to double infection in the natural populations of Ae. albopictus at each of the studied Greek prefectures (colored areas). Prefecture abbreviations are as in Table 1 and Table 2. Prefecture colors depict the different single infection proportion levels as shown in the legend. Prefecture boundaries’ color indicates average temperature as cyan (23.067 – 25.341°C), pink (25.341 – 27.615°C), or red (27.615 – 29.888°C). These ranges represent equidistant bins across the observed minimum-maximum temperature range. When samples from multiple time points were taken from a prefecture, the overall temperature average was used for this visualization. Additional information is provided in Supplementary Table 1.
Figure 1.
The distribution of Wolbachia proportion of single (wAlbB) to double infection in the natural populations of Ae. albopictus at each of the studied Greek prefectures (colored areas). Prefecture abbreviations are as in Table 1 and Table 2. Prefecture colors depict the different single infection proportion levels as shown in the legend. Prefecture boundaries’ color indicates average temperature as cyan (23.067 – 25.341°C), pink (25.341 – 27.615°C), or red (27.615 – 29.888°C). These ranges represent equidistant bins across the observed minimum-maximum temperature range. When samples from multiple time points were taken from a prefecture, the overall temperature average was used for this visualization. Additional information is provided in Supplementary Table 1.
MLST genotyping
To gain a deeper understanding of the diversity of Wolbachia strains present in Ae. albopictus populations in Greece, 11 samples were genotyped by MLST. We genotyped single and double infected samples from both sexes from three different prefectures, attempting to capture possible genetic variation and different MLST profiles (if any). The MLST profile obtained for the wAlbA strain was identical across all samples. According to the numbering of the Wolbachia MLST database the identified alleles were 3, 229, 2, 10, and 3 for the loci gatB, coxA, hcpA, ftsZ, and fbpA, respectively, presenting four matches with ST code 2. Similarly, the wAlbB strain exhibited identical profiles across all samples (alleles 242,229, 166, 210, 27) matching ST code 464. It should be noted that for double-infected individuals the allele for the coxA of the wAlbB strain has been assumed to be the same with the one in the wAlbB single-infected individuals, given the monomorphic outcome observed for all loci in all samples.
Temperature and sex ratio effect on single Wolbachia infection
The Kruskal-Wallis test revealed a highly significant effect of mean temperature and sex bias on the ratio of single to double Wolbachia infection in the studied mosquito populations (P = 1.09E-06; statistic = 27.46). Further analysis using a post-hoc Dunn test distinguished between a highly significant effect of mean temperature (P = 6.50E-07; Figure 2A) and a marginally non-significant effect of sex ratio (P = 0.053). Although the latter may be influenced by the limited number of observations (n = 11), the observed trend appears to be strong (Figure 2B), and the removal of a single outlier value resulted in a significant outcome (P = 0.040). To this end, the bootstrap-with-replacement approach confirmed these observations over 10,000 bootstrap resamples. The mean of P-values for the effect of temperature was P = 2.27E-06, with a high 95% confidence limit of P = 1.69E-05. The mean of P-values for the effect of sex ratio was P = 0.097, with 52.45% of the resamples at P < 0.05 (Figure 3). Overall, these findings suggest a positive relationship between higher environmental temperatures and single Wolbachia infections in Ae. albopictus mosquitoes, with a negative influence from female sex bias (Figure 2).
Discussion
High Wolbachia infection rate in Ae. albopictus population in Greece. The high infection rate (95%) in the natural population of Ae. albopictus in Greece supports previous studies from other regions and confirms that Wolbachia infection has been fixed in Ae. albopictus worldwide [13,21,65]. However, we observed slight differences in the infection rates among different prefectures. All the examined prefectures of Northern Greece exhibited an infection rate of 100% unlike the prefectures in central and southern Greece (88% mean rate). Variance in infection rates due to different geographical regions has been previously reported [18]. Understanding the infection dynamics of Wolbachia in Ae. albopictus holds the promise of establishing biologically based management practices, as the density of the endosymbiont Wolbachia plays an important role in crossing sterility (e.g. Werren et al. 1995 [12]).
Low MLST profile diversity for Wolbachia strains in Greek populations and worldwide | The MLST profiles for the wAlbA and wAlbB strains were monomorphic in the tested Greek populations, which is consistent with the low diversity observed in the MLST genes in the Ae. albopictus Wolbachia strains worldwide [18,25,26,27]. Specifically, the sole identified profile for the wAlbB strain (242, 229, 166, 210, and 27 as per the numbering of the MLST database) was also the most common profile identified in almost all populations studied from China, Russia, and Argentina [18,25,26,27]. There is only one other profile reported for the wAlbB strain in China, which differs only in the gatB allele [18]. The sole identified wAlbA profile (3, 229, 2, 10, and 3) shares identical alleles at four out of the five loci with samples from China and Thailand [18,66]. It only differs in the coxA locus, where we detected the same allele (229) as in the wAlbB strain. Since it has been proposed that the 229 allele has moved from supergroup-A to supergroup-B through recombination [26], it may be that the supergroup-A-specific primers mis-amplified the 229 allele of the wAlbB strain as it is expected to be more abundant in the double infected individuals [67]. Notably, as mentioned in Badieritakis et al. (2018) [3], the Greek populations of Ae. albopictus have been found to have some genetic affinity with those of Thailand [68]. As such, our results seem to align with those earlier findings. The Greek populations of Ae. albopictus have also been established relatively recently. For example, the first reports in Serres, Aetolia-Acarnania, and Thessaloniki were made between 2007 and 2009 [3]. This recent occurrence of populations may have a role in the observed low MLST diversity due to founder effects and limited time for divergence.
Differential strain distribution of Wolbachia by sex | By examining the double and single infection patterns in relation to the sex of the individual mosquitoes, we noted that almost all female mosquitoes were double infected (92%), whereas only about half of males carried both strains (52%). We observed that in males, the single infection with the wAlbB strain is more common in marginally non-significant level (Figure 2B), which agrees with the scientific literature [19,20,21,23,69]. There are different hypotheses for the detection of both strains in females and only the wAlbB strain in males. It is believed that male mosquitoes lose their wAlbA infection as they get older as an evolutionary selection of nuclear countermeasures to Wolbachia manipulation [23]. Female Ae. albopictus carry a fixed infection with both strains, with rare instances of single infected females [19,20,21,23,69]. The mechanism of CI suggests that females need both strains, to reverse the sperm modification each Wolbachia strain induces in males to maximize mating compatibility [10,70]. However, males with a single or no infection remain compatible with double infected females [10]. Male embryos carry both infections since their mothers are almost always double infected, but they tend to lose the wAlbA infection as they age. The same pattern is not observed in the wAlbB strain. The first detected Wolbachia infected Ae. albopictus (before 1970) carried only the wAlbA strain, indicating that the infection with the wAlbB strain is more recent [65].
Differential strain distribution of Wolbachia by environmental temperature | The prefecture-specific infection rates highlighted that non-infected individuals were only found in the three southernmost prefectures examined (ACH, AET, KAR). Furthermore, single-infected individuals were also mainly found in these three prefectures (with one single-infected individual found in Imathia as well) (Table 1). This observation aligns with previous results from China showing that Wolbachia prevalence in Ae albopictus natural populations is affected by geographical location and climate type [18]. It also agrees with hypotheses that higher temperatures lead to decreased Wolbachia density in laboratory populations, affecting both infections but particularly wAlbA [24,71]. Our statistical analysis strongly supported this hypothesis, as the effect of the mean temperature from 30 days prior to the sampling date had a highly significant impact on the ratio of single infections in the mosquito populations (P = 6.50E-07; Figure 2A). The mechanistic underpinnings of this observation are not fully understood. Some insight could be provided by studies concluding that maternal transmission under high temperatures and larval density is more often unsuccessful for the wAlbA than for the wAlbB strain [67,71]. Even so, when scoring single- to double-infected individuals it should not be disregarded that the density of the wAlbA strain is smaller than that of wAlbB in double-infected individuals [18,67] and, thus, it is possible that what appears like absence of the wAlbA infection could in fact reflect a reduction beyond PCR detection limits [15,71]. Nevertheless, the above findings raise important points regarding the molecular ecology of the Wolbachia infections, which may also be of applied interest.
Conclusions: Altogether: this study provides important insights into the distribution and diversity of Wolbachia strains in Ae. albopictus populations in Greece and the broader northeastern European region. We report the first MLST genotype information of Wolbachia strains in Ae. albopictus in Europe and identify a high infection rate in the natural population in Greece, confirming that Wolbachia infection has become fixed in populations worldwide. We found a positive correlation between higher environmental temperatures and single Wolbachia infections in Ae. albopictus mosquitoes, demonstrating that ecological and evolutionary factors play a role in Wolbachia dynamics. Overall, this study contributes to our understanding of Wolbachia dynamics in natural populations of Ae. albopictus. We anticipate that future research with even denser sampling will evaluate our findings, and it will have profound implications for the potential use of Wolbachia-based approaches in controlling mosquito-borne diseases in the area.
Acknowledgments
We thank Prof. Alexander Triantafyllids and Dr. Nikoleta Karaiskou from the Department of Biology of the Aristotle University of Thessaloniki for hosting the molecular work conducted in this work.
References
- Swan, T.; Russell, T.L.; Staunton, K.M.; Field, M.A.; Ritchie, S.A.; Burkot, T.R. A Literature Review of Dispersal Pathways of Aedes Albopictus across Different Spatial Scales: Implications for Vector Surveillance. Parasit. Vectors 2022, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Samanidou-Voyadjoglou, A.; Patsoula, E.; Spanakos, G.; Vakalis, N.C. Confirmation of Aedes Albopictus (Skuse) (Diptera: Culicidae) in Greece. Eur. Mosq. Bull. 2005, 19, 10–11. [Google Scholar]
- Badieritakis, Ε.; Papachristos, D.; Latinopoulos, D.; Stefopoulou, A.; Kolimenakis, A.; Bithas, K.; Patsoula, Ε.; Beleri, S.; Maselou, D.; Balatsos, G.; et al. Aedes Albopictus (Skuse, 1895) (Diptera: Culicidae) in Greece: 13 Years of Living with the Asian Tiger Mosquito. Parasitol. Res. 2018, 117, 453–460. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical Review of the Vector Status of Aedes Albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes Albopictus, an Arbovirus Vector: From the Darkness to the Light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Egid, B.R.; Coulibaly, M.; Dadzie, S.K.; Kamgang, B.; McCall, P.J.; Sedda, L.; Toe, K.H.; Wilson, A.L. Review of the Ecology and Behaviour of Aedes Aegypti and Aedes Albopictus in Western Africa and Implications for Vector Control. Curr. Res. Parasitol. Vector-Borne Dis. 2022, 2, 100074. [Google Scholar] [CrossRef]
- Bouyer, J.; Yamada, H.; Pereira, R.; Bourtzis, K.; Vreysen, M.J.B. Phased Conditional Approach for Mosquito Management Using Sterile Insect Technique. Trends Parasitol. 2020, 36, 325–336. [Google Scholar] [CrossRef]
- Bourtzis, K.; Dobson, S.L.; Xi, Z.; Rasgon, J.L.; Calvitti, M.; Moreira, L.A.; Bossin, H.C.; Moretti, R.; Baton, L.A.; Hughes, G.L.; et al. Harnessing Mosquito–Wolbachia Symbiosis for Vector and Disease Control. Acta Trop. 2014, 132, S150–S163. [Google Scholar] [CrossRef] [PubMed]
- Inácio da Silva, L.M.; Dezordi, F.Z.; Paiva, M.H.S.; Wallau, G.L. Systematic Review of Wolbachia Symbiont Detection in Mosquitoes: An Entangled Topic about Methodological Power and True Symbiosis. Pathogens 2021, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Werren, J.H.; Windsor, D.M. Wolbachia Infection Frequencies in Insects: Evidence of a Global Equilibrium? Proc. R. Soc. London. Ser. B Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Windsor, D.; Guo, L. Distribution of Wolbachia among Neotropical Arthropods. Proc. R. Soc. B Biol. Sci. 1995, 262, 197–204. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How Many Species Are Infected with Wolbachia? €“ a Statistical Analysis of Current Data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Sinkins, S.P. Wolbachia and Cytoplasmic Incompatibility in Mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Sinkins, S. P.; Braig, H. R.; O’Neill, S.L. Wolbachia Superinfections and the Expression of Cytoplasmic Incompatibility. Proc. R. Soc. London. Ser. B Biol. Sci. 1995, 261, 325–330. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Walker, T.; O’ Neill, S.L. Wolbachia and the Biological Control of Mosquito-borne Disease. EMBO Rep. 2011, 12, 508–518. [Google Scholar] [CrossRef]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Exploiting Intimate Relationships: Controlling Mosquito-Transmitted Disease with Wolbachia. Trends Parasitol. 2016, 32, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xi, Z.; Liu, X.; Wang, J.; Guo, Y.; Ren, D.; Wu, H.; Wang, X.; Chen, B.; Liu, Q. Identification and Molecular Characterization of Wolbachia Strains in Natural Populations of Aedes Albopictus in China. Parasit. Vectors 2020, 13, 28. [Google Scholar] [CrossRef]
- Afizah, An.; Roziah, A.; Nazni, W.; Lee, H. Detection of Wolbachia from Field Collected Aedes Albopictus Skuse in Malaysia. Indian J. Med. Res. 2015, 142, 205. [Google Scholar] [CrossRef]
- Albuquerque, A.L. de; Magalhães, T.; Ayres, C.F.J. High Prevalence and Lack of Diversity of Wolbachia Pipientis in Aedes Albopictus Populations from Northeast Brazil. Mem. Inst. Oswaldo Cruz 2011, 106, 773–776. [Google Scholar] [CrossRef]
- Baimai, V.; Kitrayapong, P.; O’Neill, S.L. Field Prevalence of Wolbachia in the Mosquito Vector Aedes Albopictus. Am. J. Trop. Med. Hyg. 2002, 66, 108–111. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neill, S. Phylogeny and PCR–Based Classification of Wolbachia Strains Using Wsp Gene Sequences. Proc. R. Soc. London. Ser. B Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef]
- Tortosa, P.; Charlat, S.; Labbé, P.; Dehecq, J.-S.; Barré, H.; Weill, M. Wolbachia Age-Sex-Specific Density in Aedes Albopictus: A Host Evolutionary Response to Cytoplasmic Incompatibility? PLoS One 2010, 5, e9700. [Google Scholar] [CrossRef]
- Wiwatanaratanabutr, I.; Kittayapong, P. Effects of Crowding and Temperature on Wolbachia Infection Density among Life Cycle Stages of Aedes Albopictus. J. Invertebr. Pathol. 2009, 102, 220–224. [Google Scholar] [CrossRef]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus Sequence Typing System for the Endosymbiont Wolbachia Pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.; Bogacheva, A.; Rakova, V.; Ganushkina, L.; Ilinsky, Y. Wolbachia Symbionts in Mosquitoes: Intra- and Intersupergroup Recombinations, Horizontal Transmission and Evolution. Mol. Phylogenet. Evol. 2019, 134, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Chuchuy, A.; Rodriguero, M.S.; Ferrari, W.; Ciota, A.T.; Kramer, L.D.; Micieli, M. V. Biological Characterization of Aedes Albopictus (Diptera: Culicidae) in Argentina: Implications for Arbovirus Transmission. Sci. Rep. 2018, 8, 5041. [Google Scholar] [CrossRef]
- Blagrove, M.S.C.; Arias-Goeta, C.; Failloux, A.-B.; Sinkins, S.P. Wolbachia Strain w Mel Induces Cytoplasmic Incompatibility and Blocks Dengue Transmission in Aedes Albopictus. Proc. Natl. Acad. Sci. 2012, 109, 255–260. [Google Scholar] [CrossRef]
- Blagrove, M.S.C.; Arias-Goeta, C.; Di Genua, C.; Failloux, A.-B.; Sinkins, S.P. A Wolbachia WMel Transinfection in Aedes Albopictus Is Not Detrimental to Host Fitness and Inhibits Chikungunya Virus. PLoS Negl. Trop. Dis. 2013, 7, e2152. [Google Scholar] [CrossRef]
- Mancini, M.V.; Herd, C.S.; Ant, T.H.; Murdochy, S.M.; Sinkins, S.P. Wolbachia Strain WAu Efficiently Blocks Arbovirus Transmission in Aedes Albopictus. PLoS Negl. Trop. Dis. 2020, 14, e0007926. [Google Scholar] [CrossRef] [PubMed]
- MORETTI, R.; CALVITTI, M. Male Mating Performance and Cytoplasmic Incompatibility in a w Pip Wolbachia Trans-infected Line of Aedes Albopictus ( Stegomyia Albopicta ). Med. Vet. Entomol. 2013, 27, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zheng, X.; Xi, Z.; Bourtzis, K.; Gilles, J.R.L. Combining the Sterile Insect Technique with the Incompatible Insect Technique: I-Impact of Wolbachia Infection on the Fitness of Triple- and Double-Infected Strains of Aedes Albopictus. PLoS One 2015, 10, e0121126. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and Sterile Insect Techniques Combined Eliminate Mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Caputo, B.; Moretti, R.; Virgillito, C.; Manica, M.; Lampazzi, E.; Lombardi, G.; Serini, P.; Pichler, V.; Beebe, N.W.; della Torre, A.; et al. A Bacterium against the Tiger: Further Evidence of the Potential of Noninundative Releases of Males with Manipulated Wolbachia Infection in Reducing Fertility of Aedes Albopictus Field Populations in Italy. Pest Manag. Sci. 2023, 79, 3167–3176. [Google Scholar] [CrossRef]
- Lucati, F.; Delacour, S.; Palmer, J.R.B.; Caner, J.; Oltra, A.; Paredes-Esquivel, C.; Mariani, S.; Escartin, S.; Roiz, D.; Collantes, F.; et al. Multiple Invasions, Wolbachia and Human-Aided Transport Drive the Genetic Variability of Aedes Albopictus in the Iberian Peninsula. Sci. Rep. 2022, 12, 20682. [Google Scholar] [CrossRef] [PubMed]
- Amraoui, F.; Failloux, A.-B. Chikungunya: An Unexpected Emergence in Europe. Curr. Opin. Virol. 2016, 21, 146–150. [Google Scholar] [CrossRef]
- Maria, A.T.; Maquart, M.; Makinson, A.; Flusin, O.; Segondy, M.; Leparc-Goffart, I.; Le Moing, V.; Foulongne, V. Zika Virus Infections in Three Travellers Returning from South America and the Caribbean Respectively, to Montpellier, France, December 2015 to January 2016. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Gjenero-Margan, I.; Aleraj, B.; Krajcar, D.; Lesnikar, V.; Klobučar, A.; Pem-Novosel, I.; Kurečić-Filipović, S.; Komparak, S.; Martić, R.; Đuričić, S.; et al. Autochthonous Dengue Fever in Croatia, August–September 2010. Eurosurveillance 2011, 16. [Google Scholar] [CrossRef]
- Barzon, L.; Gobbi, F.; Capelli, G.; Montarsi, F.; Martini, S.; Riccetti, S.; Sinigaglia, A.; Pacenti, M.; Pavan, G.; Rassu, M.; et al. Autochthonous Dengue Outbreak in Italy 2020: Clinical, Virological and Entomological Findings. J. Travel Med. 2021, 28. [Google Scholar] [CrossRef]
- Tomasello, D.; Schlagenhauf, P. Chikungunya and Dengue Autochthonous Cases in Europe, 2007–2012. Travel Med. Infect. Dis. 2013, 11, 274–284. [Google Scholar] [CrossRef]
- Gould, E.A.; Gallian, P.; de Lamballerie, X.; Charrel, R.N. First Cases of Autochthonous Dengue Fever and Chikungunya Fever in France: From Bad Dream to Reality! Clin. Microbiol. Infect. 2010, 16, 1702–1704. [Google Scholar] [CrossRef] [PubMed]
- Roiz, D.; Boussès, P.; Simard, F.; Paupy, C.; Fontenille, D. Autochthonous Chikungunya Transmission and Extreme Climate Events in Southern France. PLoS Negl. Trop. Dis. 2015, 9, e0003854. [Google Scholar] [CrossRef]
- Calba, C.; Guerbois-Galla, M.; Franke, F.; Jeannin, C.; Auzet-Caillaud, M.; Grard, G.; Pigaglio, L.; Decoppet, A.; Weicherding, J.; Savaill, M.-C.; et al. Preliminary Report of an Autochthonous Chikungunya Outbreak in France, July to September 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Venturi, G.; Di Luca, M.; Fortuna, C.; Remoli, M.E.; Riccardo, F.; Severini, F.; Toma, L.; Del Manso, M.; Benedetti, E.; Caporali, M.G.; et al. Detection of a Chikungunya Outbreak in Central Italy, August to September 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Bagnarelli, P.; Marinelli, K.; Trotta, D.; Monachetti, A.; Tavio, M.; Del Gobbo, R.; Capobianchi, M.R.; Menzo, S.; Nicoletti, L.; Magurano, F.; et al. Human Case of Autochthonous West Nile Virus Lineage 2 Infection in Italy, September 2011. Eurosurveillance 2011, 16. [Google Scholar] [CrossRef]
- Pietsch, C.; Michalski, D.; Münch, J.; Petros, S.; Bergs, S.; Trawinski, H.; Lübbert, C.; Liebert, U.G. Autochthonous West Nile Virus Infection Outbreak in Humans, Leipzig, Germany, August to September 2020. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Bachmann, F.; Choi, M.; Kurvits, L.; Schmidt, M.L.; Bergfeld, L.; Meier, I.; Zuchowski, M.; Werber, D.; Hofmann, J.; et al. Autochthonous West Nile Virus Infection in Germany: Increasing Numbers and a Rare Encephalitis Case in a Kidney Transplant Recipient. Transbound. Emerg. Dis. 2022, 69, 221–226. [Google Scholar] [CrossRef]
- Vlaskamp, D.R.; Thijsen, S.F.; Reimerink, J.; Hilkens, P.; Bouvy, W.H.; Bantjes, S.E.; Vlaminckx, B.J.; Zaaijer, H.; van den Kerkhof, H.H.; Raven, S.F.; et al. First Autochthonous Human West Nile Virus Infections in the Netherlands, July to August 2020. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef]
- Ramalli, L.; Grard, G.; Beck, C.; Gallian, P.; L’Ambert, G.; Desvaux, S.; Jourdan, M.; Ortmans, C.; Paty, M.-C.; Franke, F. West Nile Virus Infections in France, July to November 2018. Eur. J. Public Health 2019, 29. [Google Scholar] [CrossRef]
- Laidoudi, Y.; Durand, G.; Watier-Grillot, S.; Dessimoulie, A.-S.; Labarde, C.; Normand, T.; Andréo, V.; Guérin, P.; Grard, G.; Davoust, B. Evidence of Antibodies against the West Nile Virus and the Usutu Virus in Dogs and Horses from the Southeast of France. Transbound. Emerg. Dis. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- Young, J.J.; Haussig, J.M.; Aberle, S.W.; Pervanidou, D.; Riccardo, F.; Sekulić, N.; Bakonyi, T.; Gossner, C.M. Epidemiology of Human West Nile Virus Infections in the European Union and European Union Enlargement Countries, 2010 to 2018. Eurosurveillance 2021, 26. [Google Scholar] [CrossRef]
- Lillepold, K.; Rocklöv, J.; Liu-Helmersson, J.; Sewe, M.; Semenza, J.C. More Arboviral Disease Outbreaks in Continental Europe Due to the Warming Climate? J. Travel Med. 2019, 26. [Google Scholar] [CrossRef] [PubMed]
- Semenza, J.C.; Suk, J.E. Vector-Borne Diseases and Climate Change: A European Perspective. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Whitehorn, J.; Yacoub, S. Global Warming and Arboviral Infections. Clin. Med. (Northfield. Il). 2019, 19, 149–152. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.B.; Dahl, C.; Kaiser, A. Mosquitoes. Identification, Biology and Control, 3rd ed.; Springer: Switzerland, 2020; ISBN 9783030116224. [Google Scholar]
- Darsie, R.F.; Samanidou-Voyadjoglou, A. Keys for the Identification of the Mosquitoes of Greece. J. Am. Mosq. Control Assoc. 1997, 13, 247–254. [Google Scholar]
- Samanidou-Voyadjoglou, A.; Harbach, R.E. Keys to the Adult Female Mosquitoes (Culicidae) of Greece. Eur. Mosq. Bull. 2001, 10, 13–20. [Google Scholar]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Springer Berlin Heidelberg: Berlin, Heidelberg, 1991; pp. 283–293. [Google Scholar]
- Sakamoto, J.M.; Feinstein, J.; Rasgon, J.L. Wolbachia Infections in the Cimicidae: Museum Specimens as an Untapped Resource for Endosymbiont Surveys. Appl. Environ. Microbiol. 2006, 72, 3161–3167. [Google Scholar] [CrossRef] [PubMed]
- Hanner, R.; Fugate, M. Branchiopod Phylogenetic Reconstruction from 12s RDNA Sequence Data. J. Crustac. Biol. 1997, 17, 174. [Google Scholar] [CrossRef]
- Kounatidis, I.; Papadopoulos, N.; Bourtzis, K.; Mavragani-Tsipidou, P. Genetic and Cytogenetic Analysis of the Fruit Fly Rhagoletis Cerasi (Diptera: Tephritidae). Genome 2008, 51, 479–491. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Santos-Garcia, D.; Dionyssopoulou, E.; Moreira, M.; Papapanagiotou, A.; Scarvelakis, M.; Doudoumis, V.; Ramos, S.; Aguiar, A.F.; Borges, P.A. V.; et al. Detection and Characterization of Wolbachia Infections in Natural Populations of Aphids: Is the Hidden Diversity Fully Unraveled? PLoS One 2011, 6, e28695. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple Comparisons Using Rank Sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Armbruster, P.; Damsky, W.E.; Giordano, R.; Birungi, J.; Munstermann, L.E.; Conn, J.E. Infection of New- and Old-World Aedes albopictus (Diptera: Culicidae) by the intracellular parasite Wolbachia: implications for host mitochondrial DNA evolution. J. Med. Entomol. 2003, 40, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Werren, J.H. Revisiting Wolbachia Supergroup Typing Based on WSP: Spurious Lineages and Discordance with MLST. Curr. Microbiol. 2007, 55, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Dutton, T.J.; Sinkins, S.P. Strain-Specific Quantification of Wolbachia Density in Aedes Albopictus and Effects of Larval Rearing Conditions. Insect Mol. Biol. 2004, 13, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Guglielmino, C.R.; Scolari, F.; Vega-Rúa, A.; Failloux, A.B.; Somboon, P.; Lisa, A.; Savini, G.; Bonizzoni, M.; Gomulski, L.M.; et al. Genetic Evidence for a Worldwide Chaotic Dispersion Pattern of the Arbovirus Vector, Aedes Albopictus. PLoS Negl. Trop. Dis. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chung, J.; Robinson, K.L.; Schmidt, T.L.; Ross, P.A.; Liang, J.; Hoffmann, A.A. Sex-Specific Distribution and Classification of Wolbachia Infections and Mitochondrial DNA Haplogroups in Aedes Albopictus from the Indo-Pacific. PLoS Negl. Trop. Dis. 2022, 16, e0010139. [Google Scholar] [CrossRef] [PubMed]
- ENGELSTÄDTER, J.; HAMMERSTEIN, P.; HURST, G.D.D. The Evolution of Endosymbiont Density in Doubly Infected Host Species. J. Evol. Biol. 2007, 20, 685–695. [Google Scholar] [CrossRef] [PubMed]
- WIWATANARATANABUTR, S.; KITTAYAPONG, P. Effects of Temephos and Temperature on Wolbachia Load and Life History Traits of Aedes Albopictus. Med. Vet. Entomol. 2006, 20, 300–307. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Boxplots showing the distribution of the proportion of mosquitoes infected with single Wolbachia strains across (A) three temperature bins and (B) six female:male sex ratio bins in the studied populations of mosquitoes. The three bins in each cases represent equal splits across the observed min-max range (N = 1, 3, and 8 in temperature bins from lower to higher oC and N = 7, 3, and 2 in sex ratio bins from lower to higher female bias). The lower and upper hinges of the boxes represent the first and third quartiles, while the line inside the box represents the median. Whiskers extend to the lowest and highest values within 1.5 times the interquartile range, and points beyond this range are shown as individual data points. The plot (A) shows that the proportion of single infected mosquitoes increases as temperature increases and the plot (B) shows that the same proportion decreases towards more female-biased sex ratios.
Figure 2.
Boxplots showing the distribution of the proportion of mosquitoes infected with single Wolbachia strains across (A) three temperature bins and (B) six female:male sex ratio bins in the studied populations of mosquitoes. The three bins in each cases represent equal splits across the observed min-max range (N = 1, 3, and 8 in temperature bins from lower to higher oC and N = 7, 3, and 2 in sex ratio bins from lower to higher female bias). The lower and upper hinges of the boxes represent the first and third quartiles, while the line inside the box represents the median. Whiskers extend to the lowest and highest values within 1.5 times the interquartile range, and points beyond this range are shown as individual data points. The plot (A) shows that the proportion of single infected mosquitoes increases as temperature increases and the plot (B) shows that the same proportion decreases towards more female-biased sex ratios.
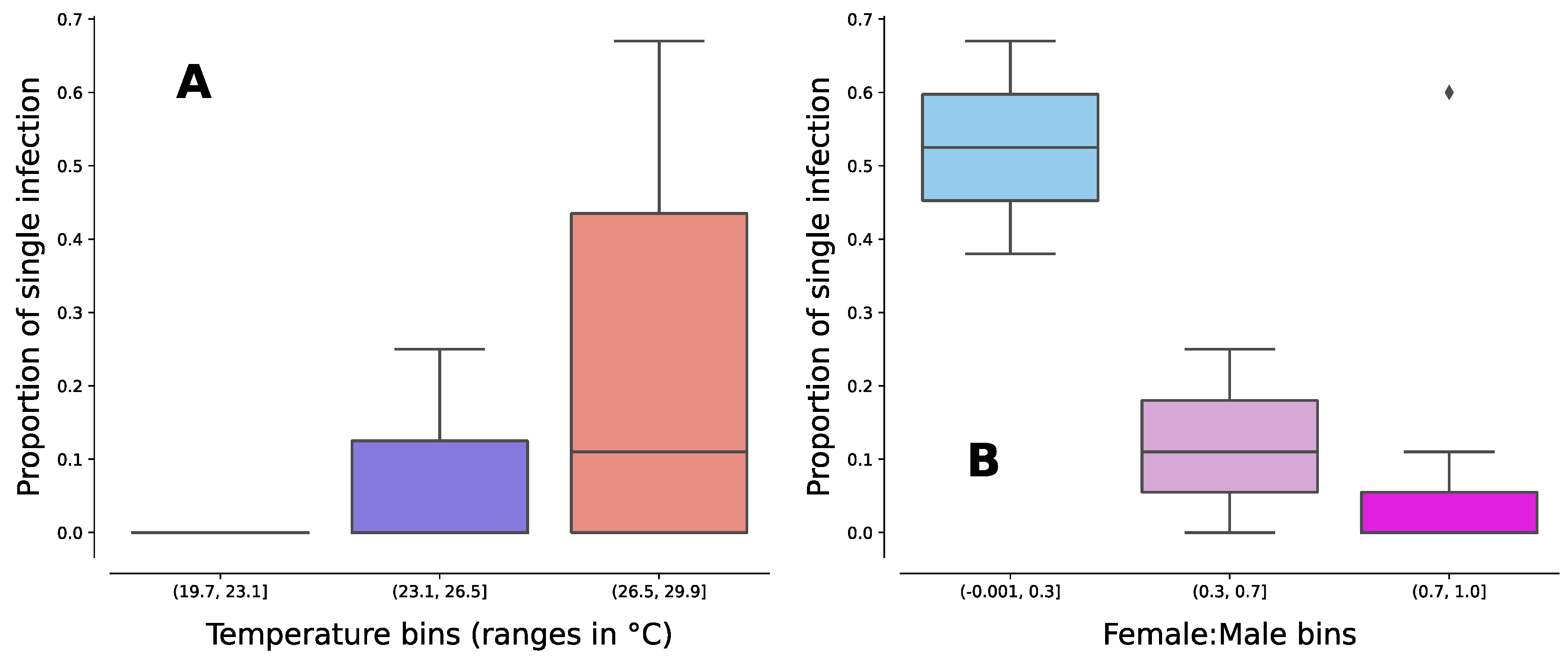
Figure 3.
Histograms illustrating the distribution of negative log10-transformed P-values from the simulation analysis (10,000 bootstrap resamples) for the effects of (A) temperature and (B) female:male ratio. Vertical lines indicate the observed significance (in blue), while the 95% confidence intervals of the significance from the simulations are represented in purple. Bars colored in pink correspond to simulations falling below the 5% significance threshold [-log10(0.05) = 1.3].
Figure 3.
Histograms illustrating the distribution of negative log10-transformed P-values from the simulation analysis (10,000 bootstrap resamples) for the effects of (A) temperature and (B) female:male ratio. Vertical lines indicate the observed significance (in blue), while the 95% confidence intervals of the significance from the simulations are represented in purple. Bars colored in pink correspond to simulations falling below the 5% significance threshold [-log10(0.05) = 1.3].
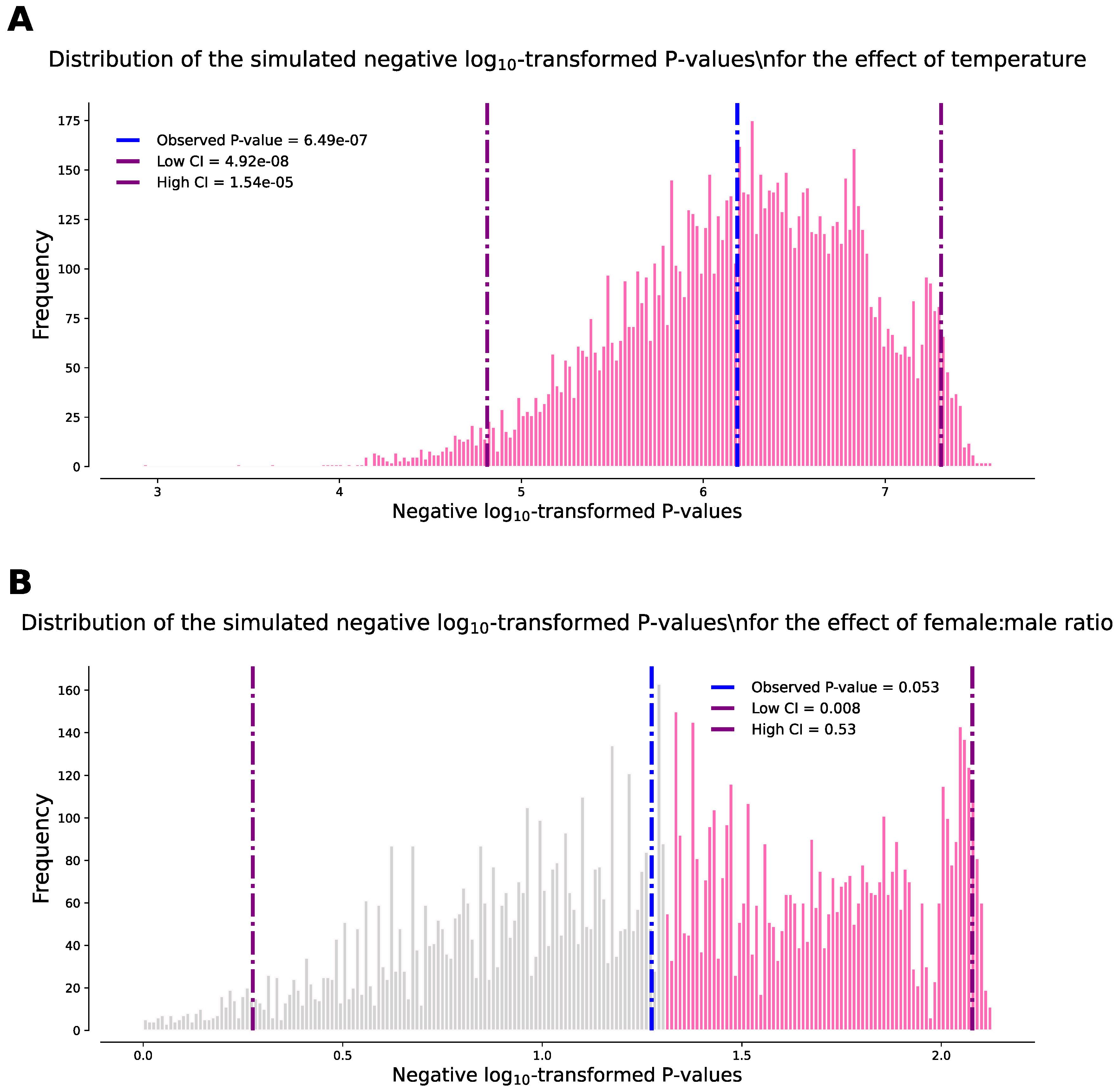

Table 1.
Information about the studied Ae. albopictus sites, the coordinates of the traps, the sampling dates, and the studied sample sizes. More detailed information is provided in the Supplementary Table 1.
Table 1.
Information about the studied Ae. albopictus sites, the coordinates of the traps, the sampling dates, and the studied sample sizes. More detailed information is provided in the Supplementary Table 1.
| Prefecture (abbreviation). | Trap coordinates | Sampling dates | Collected mosquitoes |
|---|---|---|---|
| Achaea (ACH) | 38.314695/ 21.814237 | 30/08 | 10 |
| Aetolia-Acarnania (AET) | 38.61610755/ 21.3825769;38.39032018/ 21.85072874 | 12/07; 11/08 | 15 |
| Imathia (IMA) | 40.536217/ 22.20227 | 26/08 | 9 |
| Karditsa (KAR) | 39.397834/ 22.070087;39.37078032/ 21.93258031 | 27/7; 7/9; 21/9 | 27 |
| Kozani (KOZ) | 40.312092/ 21.822304 | 16/8; 27/9 | 19 |
| Pieria (PIE) | 40.237544/ 22.582148 | 30/8 | 13 |
| Serres (SER) | 41.205796/ 23.074921 | 13/7 | 4 |
| Thessaloniki (THE) | 40.64858/ 22.954067 | 16/9 | 8 |
Table 2.
Results of the sex, the Wolbachia infection and the type of infection (single or double) in the studied mosquitoes. More detailed information, such as the date of the samplings and the average temperature, is provided in the Supplementary Table 1.
Table 2.
Results of the sex, the Wolbachia infection and the type of infection (single or double) in the studied mosquitoes. More detailed information, such as the date of the samplings and the average temperature, is provided in the Supplementary Table 1.
| Prefecture (abbreviation) | No of samples/Sex | Wolbachia prevalence | Double infection (wAlbA+wAlbB) | Single infection (wAlbB) |
|---|---|---|---|---|
| Achaea (ACH) | 5♂,5♀ | 90% (9/10) | 89% (4♂,4♀) | 11% (1♂) |
| Aetolia-Acarnania (AET) | 9♂,6♀ | 87% (13/15) | 54% (5♂,2♀) | 46% (3♂,3♀) |
| Imathia (IMA) | 9♀ | 100% (9/9) | 89% (8♀) | 11% (1♀) |
| Karditsa (KAR) | 14♂,13♀ | 93% (25/27) | 68% (4♂,13♀) | 32% (8♂) |
| Kozani (KOZ) | 19♀ | 100% (19/19) | 100% (19♀) | 0% |
| Pieria (PIE) | 13♀ | 100% (13/13) | 100% (13♀) | 0% |
| Serres (SER) | 4♀ | 100% (4/4) | 100% (4♀) | 0% |
| Thessaloniki (THE) | 3♂,5♀ | 100% (8/8) | 100% (3♂,5♀) | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



