Submitted:
20 December 2023
Posted:
20 December 2023
You are already at the latest version
Abstract
The impact of drought stress on plant growth in arid regions is a critical concern, necessitating the exploration of strategies to enhance plant drought resistance, particularly during the early stages of drought stress. This study focuses on the ectomycorrhizal fungus Cenococcum geophilum, renowned for its extensive genetic diversity and broad host compatibility, making it a crucial ally for host plants facing external stresses. We utilized approximately 5-month-old Pinus massoniana seedlings inoculated with different ecotypic strains of C. geophilum for 7 days of drought stress. The results showed that the inoculation of most strains of C. geophilum enhanced the drought resistance of P. massoniana seedlings under the early stages of drought stress, by influencing the water content, photosynthesis, accumulation of osmotic adjustment substances, and antioxidant enzyme activities in both shoots and roots of seedlings. These responses were further validated through transcriptome analysis. Notably, the level of drought resistance observed in mycorrhizal seedlings was irrespective of the level of drought tolerance of C. geophilum strains. This study contributes essential data for understanding the drought response mechanisms of mycorrhizal P. massoniana seedlings inoculated by distinct C. geophilum ecotypes, and guidance on selecting candidate species of ectomycorrhizal fungi for mycorrhizal afforestation in drought areas.
Keywords:
Pinus massoniana
; Cenococcum geophilum
; drought stress
; antioxidant activity
; mycorrhiza
1. Introduction
With the changing climate, the global reduction in precipitation has resulted in an increased occurrence of droughts [1]. Drought stress can significantly influence plant root growth by impeding elongation and branching, ultimately diminishing a plant's capacity to absorb water and nutrients in soils. Furthermore, it can lead to reduced photosynthesis and nutrient accumulation, cellular damage, and, in the end, plant wilting and mortality [2,3]. Notably, it has been documented that from 2010 to 2017, drought events were responsible for the demise of approximately 129 million trees in California [4]. Likewise, severe drought has resulted in widespread mortality of Pinus tabulaeformis in central and eastern China [5]. The global impact of drought on plant growth and crop yields is undeniable, and climate-induced and soil-related droughts are projected to intensify further due to ongoing global climate change [6]. When plants experience drought stress, their phenotypic properties and physiological traits can undergo responses, and the degree to which each property responds in the early stage of drought is closely linked to the drought resistance of the plants [7]. Consequently, it is imperative to explore strategies aimed at enhancing the drought resistance of plants under the early stage of drought stress. Pinus massoniana stands as one of the most vital timber resources in southern China, distinguished by its rapid growth, high yield, and multifaceted utility, all while contributing significantly to ecological value [8]. Nonetheless, during the early stages of afforestation, the survival rate of P. massoniana seedlings is notably hindered by factors such as inadequate shade and soil dryness [9]. Therefore, it becomes imperative to enhance the drought resistance of P. massoniana seedlings.
Ectomycorrhiza (ECM) represents a mutualistic symbiosis formed between ectomycorrhizal fungi (ECMF) and non-lignified absorbing roots [10]. ECMF can establish symbiotic relationships with plant families such as Pinaceae, Fagaceae, Salicaceae, and Betulaceae [11,12,13,14]. Under conditions of drought stress, ECM demonstrates the capacity to expand the root absorption zone, thereby enhancing water and nutrient uptake through external hyphae. This expansion results in improved root water retention and notable enhancements in root traits, including increased root biomass, total length, average diameter, and the number of root tips in the host plant. These factors collectively play a crucial role in augmenting drought resistance [15]. Furthermore, Zou, et al. [16] have observed that under water-deficient conditions, ECM hyphae can extend into soil gaps, providing additional water resources to mitigate drought effects. Additionally, ECM symbiosis contributes to increased water and nutrient absorption, facilitates photosynthesis, regulates the synthesis of antioxidant enzymes, and facilitates the accumulation of osmotic substances within plants [17]. Notably, Ahmed, et al. [18] have reported that ECMF promotes the formation of osmotic adjustment substances, such as plant soluble proteins, thereby maintaining cellular water balance and enhancing plant drought tolerance. In a study by Zhao, et al. [19], ECMF inoculation was found to elevate antioxidant enzyme activity in P. sylvestris. These findings collectively underscore the significant role of ECM symbiosis in enhancing plant resilience to drought, a phenomenon that holds paramount importance in the face of increasing environmental challenges. Therefore, the investigation of the influence of the different ECMF species on P. massoniana drought resistance, and the underlying molecular mechanisms governing is indispensable to the early stages of afforestation of P. massoniana.
Cenococcum geophlium Fr., a commonly encountered ectomycorrhizal fungus in natural ecosystems, typically exists in soil in mycorrhizal and sclerotial forms, without the formation of spores, and is characterized by a profusion of external hyphae [20]. This fungal species exhibits a broad spectrum of host compatibility and displays robust adaptability and stress resistance, making it a valuable ally for plant growth across diverse and challenging environments [21]. Notably, C. geophlium demonstrates exceptional drought tolerance and is often employed as a prominent candidate in water-scarce forest ecosystems. Research by Coleman, et al. [22] has illuminated its role in preserving cellular integrity under conditions of water deficit while mitigating the surge in reactive oxygen species (ROS) typically observed during drought stress. Furthermore, recent findings by Li, et al. [23] underscore the ability of different ecotypes of C. geophlium to induce the expression of drought-related genes when subjected to drought conditions, thereby enhancing drought resistance. It is important to note that different ecotypes of C. geophlium exhibit distinct responses to drought, presenting an intriguing avenue for further exploration in this context.
In this study, we undertook the inoculation of P. massoniana seedlings with eight ecotypes of C. geophlium. By meticulously simulating the early stage of drought stress conditions for 7 days, we aimed to investigate the diverse impacts of these ecotypes of C. geophlium on the growth and physiological responses of P. massoniana seedlings when subjected to the early stage of drought stress. Subsequently, we selected mycorrhizal seedlings that performed better (drought-tolerant) and worse (drought-sensitive) under the early stage of drought stress for in-depth RNA sequencing. The comparative assessment of differentially expressed genes in drought-tolerant and drought-sensitive mycorrhizal seedlings under the early stage of drought stress provided valuable insights into the physiological and molecular mechanisms underlying the capacity of various ecotypes of C. geophlium to enhance the drought tolerance of P. massoniana. This research endeavor aims to furnish a robust theoretical foundation for future afforestation efforts involving mycorrhizal seedlings, ultimately contributing to more resilient ecosystems in the face of drought challenges.
2. Materials and Methods
2.1. The mycelial growth of different ecotypes of C. geophilum under drought treatment
Eight ecotypic strains of C. geophilum (Jacg16, Jacg21, Jacg37, Jacg81, Jacg121, Jacg189, Jacg243, Chcg57) were used in this study. The strains in Japan were isolated by Laboratory of Forest Symbiosis at the Graduate School of Agricultural and Life Sciences, the University of Tokyo, while the strain in China was provided by the International Joint Laboratory of Forest Symbiosis at Fujian Agriculture and Forestry University. The source information of each strain is shown in Table S1. The mycelia of C. geophilum strains were pre-cultured in a modified Melin-Norkrans (MMN) agar medium [24] at 25 ℃ in the dark for 30 days. Subsequently, an agar plug of C. geophilum with a 7 mm diameter was transferred to the center of a 90 mm petri dish, which was lined with cellophane membrane at the bottom. Each dish contained 10 mL of MMN liquid medium with 0% or 10% polyethylene glycol (PEG-6000, simulated drought) and then cultured in dark at 25 ℃. Three replicates of each strain were conducted for each treatment. After culturing for 30 days, the mycelial growth area of each strain was measured by X-Plan 380dⅢ, Ushikata (Kantum Ushikata Co., LTD., Yokohama, Japan).
2.2. Preparation of mycorrhizal seedlings of P. massoniana
Pinus massoniana seeds were obtained from the Wuyi National Forest Farm in Fujian Province. These seeds were sterilized in 1% sodium hypochlorite (NaCIO, v/v) for 10 min, followed by rinsing with sterile deionized water five times. The sterilized seeds were then sown in sterilized vermiculite and cultured in a controlled environment for 6 weeks (25 ℃, 16 h light cycle). Simultaneously, the eight C. geophilum strains were pre-cultured in MMN agar medium at 25 ℃ in the dark for 45 days.
The substrate used for planting the mycorrhizal seedlings of P. massoniana inoculated by C. geophilum consisted of a mixture of forest soil and red jade soil (1:2, v/v). The forest soil was collected from the Soil and Water Moisturizing Garden of Fujian Agriculture and Forestry University in Fujian Province, China. The mixture was then sterilized by autoclaving at 121 ℃ for 3 h. Six-week-old seedlings of P. massoniana with consistent growth were transplanted into a rectangle rhizobox (23.5×8.5×1.6 cm) and then C. geophlium plugs were inoculated onto the lateral roots, while the roots of non-inoculated seedlings were covered with the agar plugs with the same size containing MMN medium. These seedlings were placed in a growth chamber with a light cycle of 16 h at 25 ℃. Throughout the cultivation period, the seedlings were weekly applied by 0.1% Hoagland nutrient solution. After 45 days of cultivation, the seedlings of P. massoniana with mycorrhizal root rate exceeding 90% were selected for further drought stress experiments.
2.3. Experimental design for drought stress of P. massoniana seedlings
Each of aforementioned seedlings was transplanted into a 50 mL centrifuge tube filled with 40.0 g of a soil mixture (forest soil:red jade soil=1:2, v/v) and then cultivated in a greenhouse under controlled conditions with an average relative humidity of 70% and the illumination duration was set at 16 h at 25 ℃. During the cultivation period, the moisture level in soil was consistently maintained at a ranged of 85%-90% of its field capacity. After 2 months of growth, the seedlings were treated by different drought stress. During the treatment period, each tube with seedling was weighted, and watered daily at 9:00 am to keep the water content in soil at the required level. For the drought treatment, the field capacity in soil was reduced at 30%-35%, while for the non-drought treatments, the field capacity in soil was maintained at 85%-90%. In each treatment, each seedling inoculated by different ecotypic strains of C. geophilum had 15 replicates. Following 7 days of different drought treatments, the seedlings were collected for the further analysis.
2.4. Determination of photosynthetic index
After the 7 days of drought treatments, the gas exchange parameters of P. massoniana needles were measured using a photosynthesis instrument with a red-blue light source (LI-6400XT, Lincoln, NE, USA). The parameters measured included the net photosynthetic rate (Pn), transpiration rate (Tr), intercellular CO2 concentration (Ci), and stomatal conductance (Gs).
2.5. Determination of morphological and physiological indicators
The collected seedlings were cleaned with double distilled water, and the excess water was removed with absorbent paper. Subsequently, three seedlings for each treatment were randomly selected to measure the fresh weight of shoots and roots. The freshly weighed seedlings were then dried at 80 ℃ until constant weight, and their dry weight was recorded using a digital scale. The water content in both shoot and root of seedlings was determined by gravimetric method. The remaining fresh seedlings were immediately treated with liquid nitrogen and stored in a refrigerator at -80 ℃ for the determination of osmotic adjustment substances, malondialdehyde (MDA) and antioxidant enzymes. To determinate each index, 0.2 g of fresh shoot or root tissue from each sample were quickly ground into a powder after freezing with liquid nitrogen. The determination of antioxidant enzymes and MDA followed the methods described by Zhang [25]: Catalase (CAT) activity was assessed using the ultraviolet absorption method, peroxidase (POD) activity was determined by employing guaiacol and hydrogen peroxide as substrates in the reaction and measuring the absorbance at 470 nm, superoxide dismutase (SOD) activity was measured using the nitroblue tetrazolium (NBT) reduction method, and the content of MDA in the seedlings was quantified utilizing the thiobarbituric acid (TBA) method. In addition, the proline (Pro) content was determined at 520 nm, following the procedure outlined by Bates, et al. [26], while the soluble protein (SP) content was assessed using the Coomassie brilliant blue method as described by Bradford [27].
2.6. RNA-seq and qRT-PCR analysis
After evaluating the drought tolerance of different ecotypic strains of C. geophilum and mycorrhizal seedlings of P. massoniana inoculated with these strains of C. geophilum, the Jacg121-inoculated (drought-tolerant) and Chcg57-inoculated (drought-sensitive) seedlings were selected for transcriptome analysis. The preparation method and drought stress treatment for these mycorrhizal seedlings remained consistent with the procedures previously described. After 7 days of different drought stress treatment, the shoots and roots of each seedling were frozen in liquid nitrogen and stored at -80 ℃. The frozen shoots and roots were sent to the Biomarker Technologies (Beijing, China) for total mRNA extraction, cDNA library construction, and sequencing. Briefly, the frozen samples were ground into powder by mortar and pestle, and immediately transferred to the lysis extraction buffer. Total mRNA from both the shoots and roots of seedlings was extracted using the RNAsimple total RNA extraction kit provided by Tiangen Biotech (Beijing, China). The cDNA library was constructed, and the sequencing was performed by an Illumina NovaSeq 6000 platform in accordance with standard protocols. The clean reads of sequencing were assembled using Trinity software to obtain an Unigene library [28]. Differentially expressed genes analysis was performed by DESeq2 software [29]. The novel genes identified in the enrichment analysis were ruled out in further analysis. Additional annotation and functional analysis of identified DEGs were conducted using Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) databases [30,31], as well as PlantGSEA analysis [32].
To confirm the gene expression levels obtained from the RNA-seq, a real-time quantification polymerase chain reaction (RT-qPCR) analysis was performed. An aliquot of the RNA sample prepared for RNA sequencing were also used for RT-qPCR analysis. The total RNA reverse transcription and RT-qPCR reactions were performed using Novozyme HiScript II Q RT SuperMix for qPCR (+gDNA wiper) Kit and ChamQ Uniweisal SYBR qPCR Master Mix Kit (Vazyme Biotech, Nanjing, China). Gene-specific primers were designed based on the unigene sequence by the tool Premier 5.0 (http://www.premierbiosoft.com/; accessed on 2 June 2022). The Aquaporin protein gene (AQP) was amplified as an internal reference gene to detect the effectiveness of template preparation. The expression of each gene was confirmed in at least three rounds of independent RT-qPCR reactions.
2.7. Statistical analysis
A principal component analysis (PCA) for comprehensively assessing the drought tolerance of plant was employed to evaluate the drought tolerance of mycorrhizal seedlings of P. massoniana inoculated by different ecotypic strains of C. geophilum, as reported by Zou, et al. [33]. This evaluation was based on parameters including water content, photosynthetic indexes, and various physiological indexes.
The formula for calculating the membership function of comprehensive index of mycorrhizal seedlings inoculated by different ecotypic strains of C. geophilum is as below:
Index weight:
Comprehensive evaluation of drought tolerant level:
Where CIj was the jth comprehensive index, CImin is the minimum value of the jth comprehensive index, CImax was the maximum value of the jth comprehensive index, and Pj was the contribution index of CIj.
SPSS (Statistical Product and Service Solutions 21.0) was used for principal component analysis, Pearson correlation analysis, Mann-Whitney U test and Student’s t-test. Pictures was drawn using Origin software.
3. Results
3.1. Assessing the drought tolerance of various C. geophilum ecotypic strains and their impact on the growth of P. massoniana seedlings through inoculation
Under non-drought condition, eight C. geophilum ecotypic strains exhibited variations in their mycelial growth (Figure 1). Jacg37 showed the largest mycelial growth, followed by Jacg189, Chcg57, and Jacg16. Conversely, Jacg21 exhibited the smallest mycelial growth. Comparatively, when cultured to the drought treatment with 10% DEG-6000, the mycelial growth of Jacg16, Jacg21, Jacg37, Jacg81, Jacg121, and Jacg189 was not significantly affected. However, the mycelial growth of Jacg243 and Chcg57 was significantly enhanced and inhibited, respectively.
The growth of P. massoniana seedlings non-inoculated and inoculated with eight C. geophilum strains was showed in Figure 2. In comparison to non-inoculated seedlings, those inoculated with each C. geophilum ecotypic strain exhibited increase in fresh and dry weight for both shoots and roots. Notably, compared with non-inoculated seedlings, the shoots and roots fresh weights of seedlings inoculated with Jacg121, Jacg37 and Chcg57 were remarkably increased (shoots, 74.00%, 38.00% and 30.67%; roots, 281.58%, 119.21% and 99.21%), the shoots fresh weights of seedlings inoculated with Jacg189 and Jacg21 showed a significant enhance (34.67% and 33.33%), and the roots fresh weights of seedlings inoculated with Jacg81 and Jacg243 remarkably increased by 87.10% and 86.58%, respectively. For dry weights, the shoots and roots of seedlings inoculated with Jacg121 and Chcg57 showed a remarkable increase (shoots, 33.45% and 19.89%; roots, 148.60% and 58.16%), the root dry weights of seedlings inoculated with Jacg37 remarkably increased by 46.93%, when compared to the non-inoculated seedlings.
3.2. Effects of inoculation with different ecotypic strains of C. geophilum on water content and photosynthetic parameters of P. massoniana seedlings during the early stage of drought stress
Compared to non-inoculated seedlings, the water content in shoots of seedlings inoculated with all different C. geophilum stains exhibited significant increases by 12.68% to 25.07%, respectively; the roots water content of seedlings inoculated with different ecotypes of C. geophilum increased significantly (21.17% to 33.46%) after 7 days of drought stress treatment except for Chcg57 (Figure 3a,b). Notably, the shoot and root water content for seedlings inoculated with Jacg121 showed the highest increase of 25.07% and 33.46%, respectively.
Inoculation with most strains of C. geophilum, except for Jacg243 with the strongest drought-tolerant mycelia also had a positive impact on the photosynthetic performance of the seedlings of P. massoniana when compared to non-inoculated seedlings after 7 days of drought stress treatment, (Table 1), especially for Jacg121-inoculated and Jacg37-inoculated seedlings. In comparison to non-inoculated seedlings, Pn, Gs, Ci and Tr of Jacg121-inoculated and Jacg37-inoculated seedlings remarkably increased by 230.37%, 496.68%, 87.47%, 142.22% and 196.57%, 366.28%, 73.51%, 85.76%, respectively.
3.3. Effects of inoculation with different ecotypic strains of C. geophilum on physiological indexes of P. massoniana seedlings during the early stage of drought stress
Compared to non-inoculated seedlings, the Pro contents in both the shoots and roots of seedlings inoculated by all strains of C. geophilum increased after 7 days of drought stress treatment (Figure 4a), especially the seedlings inoculated with Jacg121 with a 360.25% and 361.29% increase in Pro content in shoot and root. The shoots and roots of seedlings inoculated with Jacg121 and Jacg189 remarkably increased in SP content (shoots, 11.19% and 9.81%; roots, 16.71% and 10.79%) (Figure 4b). The SP content in the roots of seedlings inoculated with Jacg21 and Jacg16 increased 10.58% and 8.63%, respectively.
For MDA, the inoculation of Jacg21, Jacg81, Jacg189, Jacg243, and Chcg57 did not affect the contents in shoots of seedlings after 7 days of drought stress treatment, while inoculation of Jacg16, Jacg37, Jacg121 significantly decreased the MDA content in shoots and roots of seedlings compared to the non-inoculated seedlings (Figure 4c).
The responses of antioxidant enzyme activities (CAT, POD and SOD) in seedlings of P. massoniana to drought stress treatment of 7 days were showed in Figure 4d–f. In contrast to the non-inoculated seedlings, the inoculation of Jacg81 and Jacg121 resulted in significant decrease of CAT activity of seedlings in both the shoots (61.38% and 52.46%) and roots (53.44% and 47.44%) (Figure 4d). The CAT activity in the shoots of seedlings inoculated with Jacg37 showed a remarkably decrease with 45.79%, and the roots of seedlings inoculated with Jacg21 was significantly reduced by 39.33%. For POD activity, the inoculation of most strains of C. geophilum did not affect POD activity in shoots, while inoculation of Jacg121 and Jacg243 significantly increased POD activity (Figure 4e). In the roots, the inoculation of with Jacg121, Jacg243, Jacg37 and Jacg189 led to a significant increase in POD activity. The inoculation of most strains of C. geophlium also had a positive impact in SOD activity in seedlings. The SOD activity in shoots and roots was significantly increased by inoculation of Jacg121, Jacg81 and Jacg37, and inoculation with Jacg189 significantly increased SOD activity in shoots, when compared to the non-inoculated seedlings (Figure 4f).
3.4. Comprehensive evaluation of drought tolerance in P. massoniana mycorrhizal seedlings inoculated by various ecotypes of C. geophilum
The combined responses of total 18 indexes including water contents (2), photosynthetic parameters (4), and physiological factors (12) in P. massoniana seedlings inoculated and non-inoculated by 8 ecotypic strains of C. geophilum after 7 days of drought stress treatment were analyzed by PCA reported by Zou, et al. [33]. The result showed that the contribution rate of the first three comprehensive indexes (CI1, CI2, CI3) was 68.426%, 12.359% and 7.722%, respectively, with a total of 88.507% (Table S3). Therefore, this comprehensive evaluation of the combined responses of multiple indexes was employed to assess the drought tolerance of P. massoniana seedlings inoculated by each ecotypic strain of C. geophilum. The comprehensive evaluation of drought tolerant level (D value) of the seedlings inoculated by each strain of C. geophilum was higher than that of non-inoculated seedlings (CK) (Table 2), indicating that the inoculation of all strains of C. geophilum could enhance the drought tolerance of P. massoniana seedlings. Of these C. geophilum inoculated seedlings, the Jacg121 inoculated seedlings showed the drought tolerance, while those inoculated by Chcg57 showed the drought sensitive.
3.5. Transcriptional responses in C. geophilum inoculated P. massoniana seedlings with various drought tolerance under early stage of drought stress
To evaluate the transcriptional responses of P. massoniana seedlings with various drought tolerance inoculated with C. geophilum after 7 days of drought stress treatment, we selected the Jacg121 (drought-tolerant) and Chcg57 (drought-sensitive) inoculated P. massoniana seedlings for RNA sequencing. Compared to the seedlings of non-drought stress, 4113 DEGs (up-regulated: 1935; down-regulated: 2178) in shoots and 7014 DEGs (up-regulated: 2398; down-regulated: 4616) in roots of Jacg121 inoculated seedlings were identified, respectively (Figure 5a,c). Similarly, the Chcg57 inoculated seedlings exhibited 3174 DEGs (up-regulated: 1163; down-regulated: 2011) in shoots and 3001 DEGs (up-regulated: 1615; down-regulated: 1386) in roots compared to non-drought stressed seedlings (Figure 5b,d). Of these DEGs, 1946 were shared in the shoots, while 1343 were common in the roots of Jacg121 and Chcg57 inoculated seedlings, respectively (Figure 5e,f).
To pinpoint the potential functions of the identified DEGs in Jacg121 and Chcg57 inoculated seedlings under the early stage of drought stress, the GO term enrichment analysis was performed (Figure 6 and Figure 7). In terms of biological process, the top five terms of DEGs enrichment in the shoots and roots of both Jacg121 and Chcg57 inoculated seedlings in response to drought were mainly associated with “metabolic process”, “single-organism process”, “cellular process”, “biological regulation” and “response to stimulus”. As for cellular component, DEGs responsive to drought were primarily enriched in “membrane”, “cell”, and “organelle”. In terms of molecular function, the enrichment was mainly observed in “binding” and “catalytic activity”. Notably, the DEGs of shoots and roots of seedlings inoculated with Jacg121 and Chcg57 were also significantly enriched in the “antioxidant activity” of molecular function, indicating that the DEGs induced by mycorrhizal seedlings in the early stage of drought stress were mostly related to metabolism and reduction-oxidation reaction.
The KEGG pathway enrichment analysis was further performed to identify the active metabolic pathways involved in drought stress responses (Figure 8). Under the early stage of drought stress, pathways such as flavonoid biosynthesis (ko00941), plant hormone signal transduction (ko04075), photosynthesis (ko00195), mitogen-activated protein kinase (MAPK) signaling pathway-plant (ko04016), phenylpropanoid biosynthesis (ko00940) and starch and sucrose metabolism (ko00500) were significantly enriched in the shoots of both Jacg121 and Chcg57 inoculated seedlings. The response of these two kinds of mycorrhizal seedlings shoots to drought stress was mainly related to the synthesis of secondary metabolites (phenylpropanoid) and energy metabolism (photosynthesis, starch and sucrose metabolism). In the roots of drought-tolerant mycorrhizal seedlings, the top two pathways were phenylalanine metabolism (ko00360) and selenocompound metabolism (ko00450), followed by cysteine and methionine metabolism (ko00270) and isoquinoline alkaloid biosynthesis (ko00950). The DEGs of drought-sensitive mycorrhizal seedlings were mainly enriched in pathways such as pentose and glucuronate interconversions (ko00040), flavonoid biosynthesis (ko00941), starch and sucrose metabolism (ko00500) and DNA replication (ko03030). These results showed that the response in roots of drought-tolerant mycorrhizal seedlings to drought stress was related to the improvement of plant resistance and the removal of reactive oxygen species (ROS), while the synthesis of secondary metabolites and energy metabolism played a more important role in the roots of drought-sensitive mycorrhizal seedlings.
3.6. Analysis of key genes for drought tolerance of P. massoniana seedlings inoculated by C. geophilum under the early stage of drought stress
Combined with the KEGG pathway, we selected the significantly up/down-regulated DEGs of the Jacg121 and Chcg57 inoculated seedlings in phenylpropanoid biosynthesis (ko00940), flavonoid biosynthesis (ko00941), glutathione metabolism (ko00480) and phenylalanine metabolism (ko00360) pathways to draw a heatmap to screen out the key genes of P. massoniana seedlings in response to the early stage of drought stress (Figure 9). A total of 11 common DEGs were selected in the phenylpropanoid biosynthesis pathway. Among them, 7 DEGs were significantly down-regulated, and TRINITY_DN11509_c1_g1, TRINITY_DN20940_c0_g2 and TRINITY_DN1881_c1_g1 were significantly up-regulated in the shoots and roots of the drought-tolerant and drought-sensitive mycorrhizal seedlings under the early stage of drought stress. Seven DEGs were selected in the flavonoid biosynthesis pathway. Except for TRINITY_DN18667_c0_g1 and TRINITY_DN1881_c1_g1, the expression levels of the remaining 5 DEGs were significantly down-regulated in the shoots and roots of drought-sensitive mycorrhizal seedlings, and significantly up-regulated in shoots and significantly down-regulated in roots of drought-tolerant mycorrhizal seedlings. Five DEGs were selected in the glutathione metabolism pathway. Except for TRINITY_DN_1340_c0_g2, the other 4 DEGs were significantly down-regulated in the shoots and roots of the two kinds of mycorrhizal seedlings under the early stage of drought stress. Six DEGs were selected in the phenylalanine metabolism pathway. Under the early stage of drought stress, TRINITY_DN1073_c0_g1, TRINITY_DN1104_c0_g1 and TRINITY_DN246562_c0_g2 were significantly down-regulated in the shoots and roots of the drought-tolerant and drought-sensitive mycorrhizal seedlings. The expression of TRINITY_DN14578_c0_g1 and TRINITY_DN7317_c0_g1 in shoots of the two kinds of mycorrhizal seedlings was significantly up-regulated.
3.7. RT-qPCR verification of DEGs
In order to verify the reliability of the sequencing results, we selected two genes in the phenylpropanoid biosynthesis pathway (TRINITY_DN3930_c0_g1, TRINITY_DN20940_c0_g2), three genes in the flavonoid biosynthesis metabolic pathway (TRINITY_DN40689_c0_g2, TRINITY_DN12846_c0_g3, TRINITY_DN18667_c0_g1), and one gene in the glutathione metabolism pathway (TRINITY_DN6096_c0_g1) for RT-qPCR validation. As shown in Figure 10, the RNA sequencing results were consistent with the expression profiles of the TRINITY_DN20940_c0_g2, TRINITY_DN40689_c0_g2, TRINITY_DN12846_c0_g3, TRINITY_DN18667_c0_g1 and TRINITY_DN6096_c0_g1, genes.
4. Discussion
The current literature on enhancing the drought resistance of P. massoniana through the use of ECMF has predominantly concentrated on various ECMF species [34,35]. However, there has been limited attention dedicated to investigating the impact of different ecotypes within each ECMF species. Cenococcum geophilum, characterized by abundant genetic diversity, holds ecological significance linked to its genetic variations [21]. Numerous studies have highlighted the varying resistance levels of different ecotypic strains of C. geophilum to stresses such as drought, high temperature, heavy metals, and salt [23,36,37,38]. Nevertheless, existing studies primarily focus on the strain's inherent stress resistance mechanism, with limited exploration of the symbiotic relationship between strains and plants in facing diverse stresses. In this study, we delved into the enhancement and mechanisms of drought resistance in P. massoniana through the inoculation of eight ecotypes of C. geophilum, each with distinct drought tolerances during the early stages of drought stress. Notably, in mycelial culture experiments, Jacg243 exhibited the strongest drought resistance, Chcg57 displayed the weakest, and the remaining six ecotypic strains showed no evident growth response to drought stress. Intriguingly, our findings suggest that the inoculation of all eight ecotypes effectively enhanced the drought resistance of P. massoniana seedlings, regardless of the drought tolerance exhibited by C. geophilum mycelia.
During the early stage of drought stress, the inoculation of most ecotypes of C. geophilum increased both shoot and root water contents, photosynthetic levels, and the contents of Pro and SP in P. massoniana seedlings. These responses align with previous studies emphasizing the importance of maintaining high water content, enhancing photosynthetic activity, and accumulating osmotic regulators for plant drought resistance [39]. Photosynthetic indexes not only affect the water potential of stomatal guard cells, induce water absorption or water loss, regulate leaves stomatal opening and closing, but also carry out photosynthesis through gas exchange between internal and external environment to ensure energy supply [40]. As the main osmotic regulators of plants, Pro and SP are hydrophilic organic solvents, which can stabilize protein structure protect macromolecular substances and are positively correlated with drought tolerance of plants [41]. Wang, et al. [42] showed that inoculation with ECMF increased the water content, photosynthetic rate and accumulation of osmotic adjustment substances in P. tabulaeformis under drought stress. Rasouli, et al. [43] also reported that inoculation with mycorrhizal fungi can help Satureja hortensis resist drought stress by increasing its water content, photosynthetic levels, and accumulation of osmotic regulators. KEGG pathway enrichment analysis highlighted the significance of pathways such as phenylpropanoid biosynthesis, photosynthesis, and starch and sucrose metabolism in both drought-tolerant and drought-sensitive mycorrhizal seedlings. As an important phenolic compound, the synthesis and accumulation of phenylpropanoids can enhance the thickness of the needle cell wall and reduce the loss of water transpiration to resist the negative effects caused by drought stress [44]. Feng, et al. [45] have shown that genes such as DHNs, LEA, Annexin D2, and NAC in the phenylpropanoid biosynthesis pathway play an important role in protecting plant cell membrane permeability. These results further demonstrated that inoculation with C. geophilum could help P. massoniana resist drought stress by regulating energy supply and reducing water loss.
Plants have evolved complex defense mechanism to resist external stress, which can effectively remove the accumulation of ROS and reduce the damage caused by membrane lipid peroxidation to plant [46]. Antioxidant enzymes, as an important protective enzyme in plants, play an important role in plant stress resistance, by eliminating excessive ROS, reducing cell membrane damage, and protecting cell membranes integrity [47]. Our results showed that the activities of POD and SOD in P. massoniana seedlings, inoculated with C. geophilum were generally higher compared to non-inoculation during the early stage of drought stress, while the inoculation seedlings exhibited lower MDA content, indicating that C. geophilum inoculation can help P. massoniana to resist drought stress by reducing cell damage and scavenging ROS. Alvarez, et al. [48] have reported that ECMF inoculation can influence the activity of ROS scavenging enzyme, thereby regulating host resistance to stress. [19], Zhao, et al. [49] also found that ECMF inoculation could promote SOD activity and reduce MDA content in P. sylvestris, which was consistent with our research results. Pan, et al. [50] also suggested that plants exhibiting lower MDA content under stress had stronger drought tolerance. The results of transcriptome showed that the DEGs in P. massoniana seedlings inoculated with C. geophilum during early stage of drought stress, were significantly enriched in pathways such as flavonoid biosynthesis, plant hormone signal transduction, and MAPK signaling pathway-plant. Flavonoids are mainly derived from the phenylpropanoid pathway, which can enhance the antioxidant activity of plants and help plants resist various biotic and abiotic stresses [51]. Moreover, Hodaei, et al. [52] have shown that drought stress can lead to a significant up-regulation of flavonoid metabolism-related genes in Chrysanthemum morifolium, and a significant increase in flavonoid compounds, thereby improving the antioxidant capacity of C. morifolium. The MAPK signaling pathway is an important tool for plants to respond to abiotic and biotic stress, with ROS accumulation serving as an activator of this pathway [53]. Liu, et al. [54] found that GhMAPKKK49 in Gossypium hirsutum was induced by abscisic acid (ABA) and ROS and involved in ROS and ABA-mediated responses to various abiotic stresses. Zhu, et al. [55] reported that up-regulation of StMAPK11 in Solanum tuberosum under drought conditions can promote the activity of antioxidant enzymes, thus improving drought resistance. In our study, the CAT activity of P. massoniana seedlings inoculated with most ecotypic strains of C. geophilum was significantly lower than that of non-inoculation, possibly due to the competitive relationship between CAT and POD on the substrate H2O2 [56]. However, our results showed that the POD activity in P. massoniana seedlings inoculated with C. geophilum was higher than that of non-inoculation, indicating that C. geophilum mainly decomposed H2O2 by increasing POD activity of P. massoniana.
In this study, we found that the drought resistance of mycorrhizal seedlings did not correlate with the inherent drought resistance of the C. geophilum strain itself. Jacg121, despite showing no growth response to drought in mycelium culture, exhibited the strongest drought resistance when forming symbionts with P. massoniana. Conversely, the mycorrhizal seedlings of P. massoniana inoculated with Jacg243 of the strongest drought resistance in mycelium culture, demonstrated comparatively weak drought resistance. Under the same water condition, the biomass of P. massoniana seedlings inoculated with Jacg121 was significantly higher than that of Jacg243, especially the fresh and dry weight of roots. This implies that the establishment of a symbiotic relationship with Jacg121 significantly increased the root absorption area, enhancing the seedlings' moisture absorption capacity. Rasouli, et al. [43] have already reported that ECMF can expand the root absorption area by establishing a symbiotic relationship, improve the water status and nutrient metabolism of host plants, and avoid or slow down the drought stress to plants. This emphasizes the importance of promoting root growth to help plants overcome drought stress [57,58]. Based on our findings, it is suggested to cultivate drought-resistant ECM seedlings using strains that not only exhibit drought resistance but also possess the ability to significantly enhance plant growth in mycorrhizal afforestation in drought areas.
5. Conclusions
This study revealed that the inoculation of various ecotypes of C. geophilum had a positive impact on both shoot and root water contents, photosynthesis, osmotic adjustment substance accumulations, and antioxidant enzyme activities in P. massoniana seedlings, offering varying degrees of support against early-stage drought stress. However, the level of drought resistance of mycorrhizal seedlings of P. massoniana did not correlate with the inherent drought resistance of the C. geophilum strain itself. The strains of C. geophilum that exhibit greater efficacy in enhancing the growth, particularly the development of roots in P. massoniana seedlings, demonstrate superior capabilities in augmenting the drought resistance of these seedlings during early-stage drought stress, indicating that by applying ECMF to reforest vegetation in drought areas, the species that exhibit both drought resistance and the ability to significantly enhance plant growth are available.
Supplementary Materials
Figure S1: The number of DEGs in drought-tolerant and drought-sensitive mycorrhizal seedlings under drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments; Table S1: Geographic information of different ecotypes of Cenococcum geophilum; Table S2: Correlation coefficient matrix of single index of different ecotypes mycorrhizal seedlings after 7 days of drought stress (field capacity 30%-35%); Table S3: The index coefficient and proportion of comprehensive coefficient CI (comprehensive index); Table S4: List of transcriptome data quality; Table S5: Unigene length distribution statistics; Table S6: Sequences of primers used for RT-qPCR analysis.
Author Contributions
Conceptualization, C.L. and S.Z.; Methodology, X.Z.; Software, X.Z.; Validation, X.Z., C.L. and S.Z.; Formal analysis, X.Z.; Investigation, X.Z., J.Z., J.H., M.L., N.M. and Q.G.; Resources, X.Z., M.L. and C.L.; Data curation, X.Z., C.L. and S.Z.; Writing—original draft, X.Z.; Writing—Review & Editing, N.M.; Q.G.; C.L. and S.Z., Visualization, X.Z.; Supervision, C.L. and S.Z.; Project administration, C.L. and S.Z.; Funding acquisition, C.L. and S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by [The Foundation for Forest Science Peak Project of College of Forestry, Fujian Agriculture and Forestry University] grants number [71201800701], [Natural Science Foundation of Jiangsu Province] grants number [BK20210166], [“Double Innovation Doctor” Fund for High-Level Entrepreneurial and Innovative Talents Introduction of Jiangsu Province of China] grants number [JSSCBS20211313(032102903)] and [Open Fund Project of Jiangsu Key Laboratory for the Research and Utilization of Plant Resources] grants number [JSPKLB202204].
Data Availability Statement
The data presented in this study are available within the article and supplementary materials. Data for sequence reads are available in a publicly available repository [NCBI], reference number [PRJNA1048730].
Acknowledgments
We are also very thankful to Yuyu Shi and Chao Yuan for their contributions to the laboratory basic samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Praba, M.L.; Cairns, J.E.; Babu, R.C.; Lafitte, H.R. Identification of physiological traits underlying cultivar differences in drought tolerance in rice and wheat. J. Agron. Crop Sci. 2010, 195, 30–46. [Google Scholar] [CrossRef]
- Avery, W.; Easterling, D.R.; Kunkel, K.E.; Lewis, K.L.M.; Crimmins, A. Impacts, risks, and adaptation in the united states: fourth national climate assessment, volume II. U.S. Glob. Change Res. Program 2018, 572–603. [Google Scholar]
- Wang, H.; Zhang, Z.; Kong, X.; Lui, S.; Shen, Z. Preliminary deduction of potential distribution and alternative hosts of invasive pest, Dendroctonus valens (Coleoptera: Scolytidae). Sci. Silvae Sin. 2007, 43, 71–76. [Google Scholar]
- Choat, B. Predicting thresholds of drought-induced mortality in woody plant species. Tree Physiol. 2013, 33, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.N.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. Comparative effects of salinity and water stress on photosynthesis, water relations and growth of Jatropha curcas plants. J. Arid Environ. 2010, 74, 1130–1137. [Google Scholar] [CrossRef]
- Xiao, F.; Zhao, Y.; Wang, X.; Liu, Q.; Ran, J. Transcriptome analysis of needle and root of Pinus massoniana in response to continuous drought stress. Plants 2021, 10, 769. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Tang, S.; Chen, H. Advances of drought resistance in Pinus massoniana. Guangxi For. Sci. 2017, 46, 7. [Google Scholar]
- Liu, Y.; Li, X.; Kou, Y. Ectomycorrhizal Fungi: participation in nutrient turnover and community assembly pattern in forest ecosystems. Forests 2020, 11, 453. [Google Scholar] [CrossRef]
- Vaario, L.; Xing, S.; Xie, Z.; Lun, Z.; Sun, Y. In situ and in vitro colonization of Cathaya argyrophylla (Pinaceae) by ectomycorrhizal fungi. Mycorrhiza 2006, 16, 137–142. [Google Scholar] [CrossRef]
- Mrnka, L.; Kuchár, M.; Cieslarová, Z.; Matějka, P.; Száková, J.; Tlustoš, P.; Vosátka, M. Effects of endo- and ectomycorrhizal fungi on physiological parameters and heavy metals accumulation of two species from the family salicaceae. Water Air Soil Pollut. 2012, 223, 399–410. [Google Scholar] [CrossRef]
- Wu, B.; Gao, C.; Chen, L.; Buscot, F.; Goldmann, K.; Purahong, W.; Ji, N.; Wang, Y.; Lü, P.; Li, X. Host phylogeny is a major determinant of Fagaceae-associated ectomycorrhizal fungal community assembly at a regional scale. Front. Microbiol. 2018, 9, 2409. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, C.; Chen, L.; Ji, N.; Wu, B.; Li, X.; Lü, P.; Zheng, Y.; Guo, L. Host plant phylogeny and geographic distance strongly structure Betulaceae-associated ectomycorrhizal fungal communities in Chinese secondary forest ecosystems. FEMS Microbiol. Ecol. 2019, 95, fiz037. [Google Scholar] [CrossRef]
- Plassard, C.; Louche, J.; Ali, M.A.; Duchemin, M.; Legname, E.; Cloutier-Hurteau, B. Diversity in phosphorus mobilisation and uptake in ectomycorrhizal fungi. Ann. For. Sci. 2011, 68, 33–43. [Google Scholar] [CrossRef]
- Zou, Y.; Srivastava, A.K.; Ni, Q.D.; Wu, Q. Disruption of mycorrhizal extraradical mycelium and changes in leaf water status and soil aggregate stability in rootbox-grown trifoliate orange. Front. Microbiol. 2015, 6, 203. [Google Scholar] [CrossRef]
- Tong, L.; Zhang, H.; Yao, X. Prospects of exploitation and utilization of ecto-mycorrhiza. Anhui Agric. Sci. Bull. 2008, 14, 4. [Google Scholar]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhriss, M.; Abdullah, F.B. Exogenous proline effects on photosynthetic performance and antioxidant defense system of young olive tree. J. Agric. Food Chem. 2010, 58, 4216–4222. [Google Scholar] [CrossRef]
- Zhao, M.; Hao, W.; Ning, X.; Hao, L.; Yan, H.; Mu, Y.; Bai, S. Screening of Excellent Ectomycorrhizal Fungi-tree for Drought Resistant with Pinus sylvestris var. mongolica. Bull. Bot. Res. 2020, 40, 133–140. [Google Scholar]
- Park, J.Y. Effects of field inoculation with mycorrhizae of Cenococcum graniforme on Basswood growth. Bim. Res. Notes 1970, 26, 27–28. [Google Scholar]
- Guo, Z.; Wang, Y.; Wu, B.; Xu, Y.; Yao, H.; Li, Z.; Yu, Q.; Li, X.; Guo, L. Population genetic diversity and structure of ectomycorrhizal fungus Cenococcum geophilum. Mycosystema 2021, 40, 920–935. [Google Scholar]
- Coleman, M.D.; Bledsoe, C.S.; Lopushinsky, W. Pure culture response of ectomycorrhizal fungi to imposed water stress. Can. J. Bot. 1989, 67, 29–39. [Google Scholar] [CrossRef]
- Li, M.; Yuan, C.; Zhang, X.; Pang, W.; Zhang, P.; Xie, R.; Lian, C.; Zhang, T. The transcriptional responses of ectomycorrhizal fungus, Cenococcum geophilum, to drought stress. J. Fungi 2022, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Marx, D.H. The influence of ectotrophic mycorrhizal fungi on the resistance of pine roots to pathogenic infections. II. Production, identification, and biological activity of antibiotics produced by Leucopaxillus cerealis var. piceina. Phytopathology 1969, 59, 153–163. [Google Scholar]
- Zhang, Z. Plant physiology experiment instruction, 2nd ed.; Higher Education Press: Beijing, China, 1990. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; Chen, Z.; Mauceli, E.; Hacohen, N.; Gnirke, A.; Rhind, N.; Palma, F.d.; Birren, B.W.; Nusbaum, C.; Lindblad-Toh, K.; Friedman, N.; Regev, A. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Minoru, K.; Susumu, G.; Shuichi, K.; Yasushi, O.; Masahiro, H. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; Harris, M.A.; Hill, D.P.; Issel-Tarver, L.; Kasarskis, A.; Lewis, S.; Matese, J.C.; Richardson, E.; Ringwald, M.; Rubin, G.M.; Sherlock, G. Gene Ontology: tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Yi, X.; Du, Z.; Su, Z. PlantGSEA: a gene set enrichment analysis toolkit for plant community. Nucleic Acids Res. 2013, 41, W98–W103. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Hu, W.; Li, Y.; He, J.; Zhu, H.; Zhou, Z. Screening of drought resistance indices and evaluation of drought resistance in cotton (Gossypium hirsutum L.). J. Integr. Agric. 2020, 19, 495–508. [Google Scholar] [CrossRef]
- Peng, S.; Wang, X.; Li, J.; Xia, D.; Ge, Z.; Xue, J. Effects of ectomycorrhizal fungi inoculation on growth and photosynthetic characteristics of Broussonetia papyrifera seedlings under drought stress. Chin. J. Ecol. 2021, 40, 2719–2726. [Google Scholar]
- Wang, Y.; Tu, G. Effects of drought and rewatering on physiological properties of five species of Pinus massoniana seedlings. J. Cent. South Univ. For. Technol. 2021, 41, 12. [Google Scholar]
- Yan, T.; Zhang, P.; Pang, W.; Zhang, X.; Lian, C.; Zhang, T. Effects of high temperature-triggered transcriptomics on the physiological adaptability of Cenococcum geophilum, an ectomycorrhizal fungus. Microorganisms 2022, 10, 2039. [Google Scholar] [CrossRef]
- Shi, Y.; Yan, T.; Yuan, C.; Li, C.; Rensing, C.; Chen, Y.; Xie, R.; Zhang, T.; Lian, C. Comparative physiological and transcriptome analysis provide insights into the response of Cenococcum geophilum, an ectomycorrhizal fungus to cadmium stress. J. Fungi 2022, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C.; Tsuruta, M.; Matsushita, N.; Goto, S.; Shen, Z.; Tsugama, D.; Zhang, S.; Lian, C. Physiological and transcriptional responses of the ectomycorrhizal fungus Cenococcum geophilum to salt stress. Mycorrhiza 2022, 32, 327–340. [Google Scholar] [CrossRef]
- Khoyerdi, F.F.; Shamshiri, M.H.; Estaji, A. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. Sci. Hortic. 2016, 198, 44–51. [Google Scholar] [CrossRef]
- Zhao, L.; Li, Y. Effects of shading and drought on light-induced stomatal dynamics in Betula platyphylla seedling. Chin. J. Appl. Ecol. 2022, 33, 2331–2338. [Google Scholar]
- Firdos, K.; Aisha, A.N.; Muhammad, S.; Fahad, A.-Q.; Muhammad, A. Trehalose: a key organic osmolyte effectively involved in plant abiotic stress tolerance. J. Plant Growth Regul. 2019, 38, 608–618. [Google Scholar]
- Wang, J.; Zhang, H.; Gao, J.; Zhang, Y.; Tang, M. Effects of ectomycorrhizal fungi (Suillus variegatus) on the growth, hydraulic function, and non-structural carbohydrates of Pinus tabulaeformis under drought stress. BMC Plant Biol. 2021, 21, 171. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, F.; Amini, T.; Skrovankova, S.; Asadi, M.; Hassanpouraghdam, M.B.; Ercisli, S.; Buckova, M.; Mrazkova, M.; Mlcek, J. Influence of drought stress and mycorrhizal (Funneliformis mosseae) symbiosis on growth parameters, chlorophyll fluorescence, antioxidant activity, and essential oil composition of summer savory (Satureja hortensis L.) plants. Front. Plant Sci. 2023, 14, 1151467. [Google Scholar] [CrossRef] [PubMed]
- Mckiernan, A.B.; Hovenden, M.J.; Brodribb, T.J.; Potts, B.M.; Davies, N.W.; O’Reilly-Wapstra, J.M. Effect of limited water availability on foliar plant secondary metabolites of two Eucalyptus species. Environ. Exp. Bot. 2014, 105, 55–64. [Google Scholar] [CrossRef]
- Feng, X.; Yang, Z.; Wang, X. Tissue-specific transcriptome analysis of drought stress and rehydration in Trachycarpus fortunei at seedling. PeerJ 2021, 9, e10933. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Breusegem, F.V. ROS signaling: the new wave? Trends Plant Sci. 2011, 6, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Huygens, D.; Fernandez, C.; Yessy, G.; Olivares, E.; Saavedra, I.; Alberdi, M.; Valenzuela, E. Effect of ectomycorrhizal colonization and drought on reactive oxygen species metabolism of Nothofagus dombeyi roots. Tree Physiol. 2009, 29, 1047–1057. [Google Scholar] [CrossRef]
- Zhao, M.; Hao, W.; Ning, X.; Hao, L.; Yan, H.; Mu, Y.; Bai, S. Screening of excellent ectomycorrhizal fungi-tree for drought resistant with Pinus sylvestris var. mongolica. Bull. Bot. Res. 2020, 40, 133–140. [Google Scholar]
- Pan, X.; Li, J.; Wang, J.; He, Q.; Su, Y.; Ma, J.; Du, K. The impact of drought stress on physiological indicators of four shrub species on the Qinghai-Tibet Plateau. For. Res. 2013, 26, 352–358. [Google Scholar]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Wang, L. The phenylpropanoid pathway and plant defence - A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali Wani, O.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive oxygen species in plants: from source to sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhu, M.; Hao, L.; Chen, X.; Gao, Y.; Guo, X.; Li, H. GhMAPKKK49, a novel cotton (Gossypium hirsutum L.) MAPKKK gene, is involved in diverse stress responses. Acta Physiol. Plant. 2016, 13, 28. [Google Scholar]
- Zhu, X.; Zhang, N.; Liu, X.; Li, S.; Yang, J.; Hong, X.; Wang, F.; Si, H. Mitogen-activated protein kinase 11 (MAPK11) maintains growth and photosynthesis of potato plant under drought condition. Plant Cell Rep. 2021, 40, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Liu, L.; Li, Q.; Wang, J.; Du, B. A cascade enzyme system integrating peroxidase mimic with catalase for linear range expansion of H2O2 assay: a mechanism and application study. Small 2023, 19, e2300444. [Google Scholar] [CrossRef] [PubMed]
- Bengough, A.G.; Mckenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: a review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
Figure 1.
The mycelial growth of eight ecotypes of Cenococcum geophilum under drought and non-drought treatments. Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between 0% PEG and 10% PEG treatments is tested by Student’s t-test (** P<0.01).
Figure 1.
The mycelial growth of eight ecotypes of Cenococcum geophilum under drought and non-drought treatments. Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between 0% PEG and 10% PEG treatments is tested by Student’s t-test (** P<0.01).
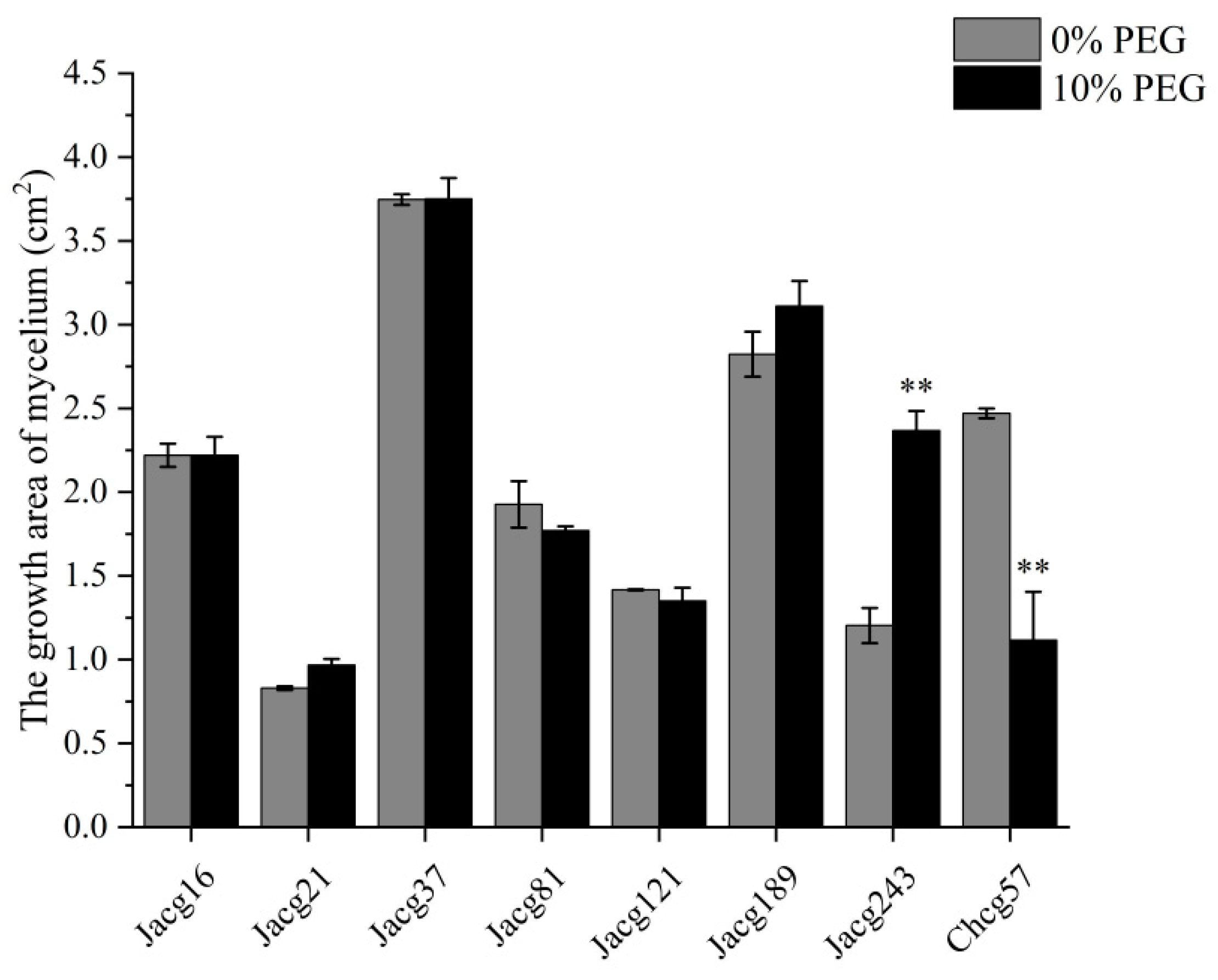
Figure 2.
Effects of different Cenococcum geophilum ecotypes inoculation on (a) fresh weights and (b) dry weights of Pinus massoniana seedlings. Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between different Cenococcum geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01; *** P<0.001). CK, non-inoculation.
Figure 2.
Effects of different Cenococcum geophilum ecotypes inoculation on (a) fresh weights and (b) dry weights of Pinus massoniana seedlings. Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between different Cenococcum geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01; *** P<0.001). CK, non-inoculation.
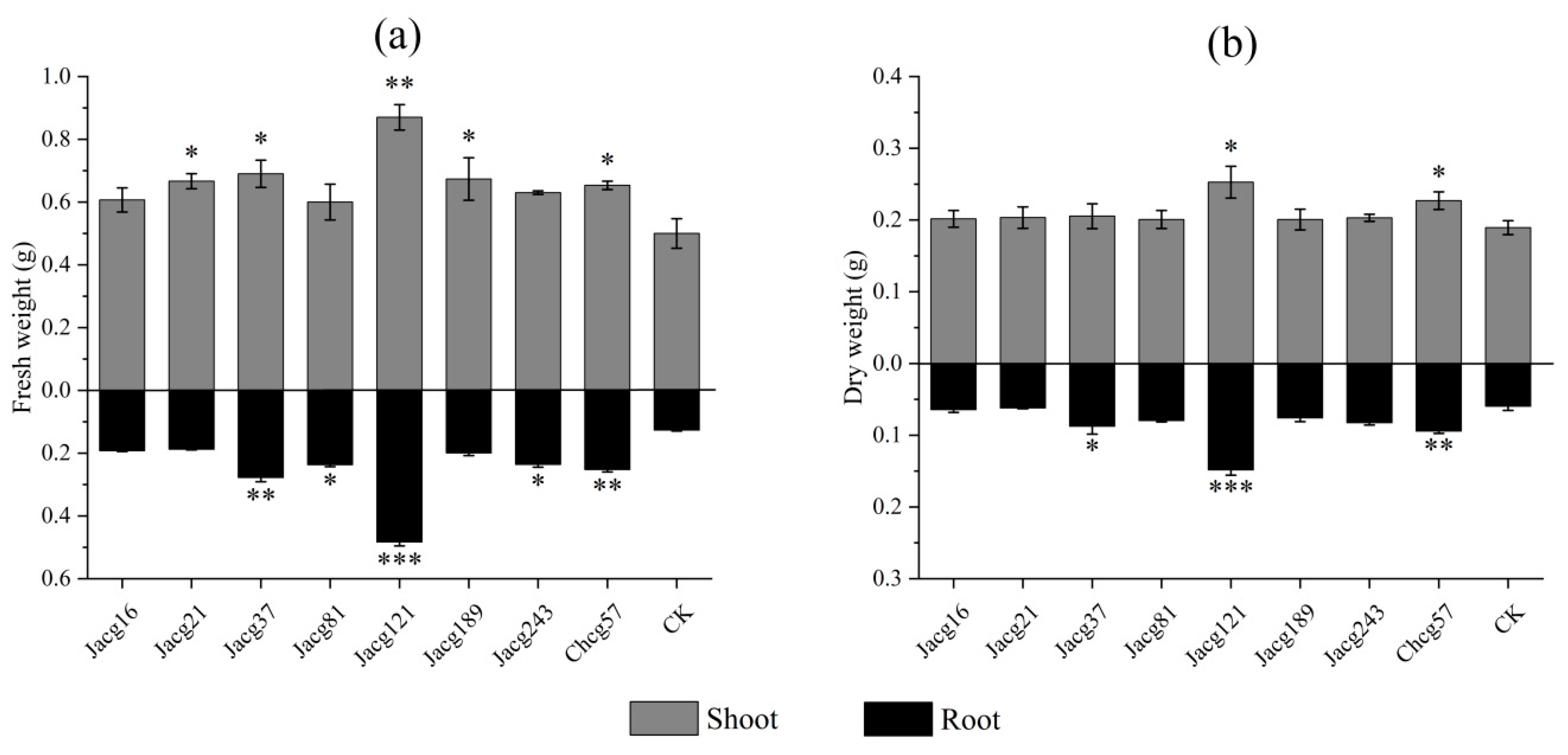
Figure 3.
Effects of different Cenococcum geophilum ecotypes inoculation on the (a) shoot and (b) root water contents of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%). Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between different C. geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01). CK, non-inoculated.
Figure 3.
Effects of different Cenococcum geophilum ecotypes inoculation on the (a) shoot and (b) root water contents of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%). Data and bars are shown as mean and ± SE of the replicates, respectively (n=3). The statistically significant difference between different C. geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01). CK, non-inoculated.
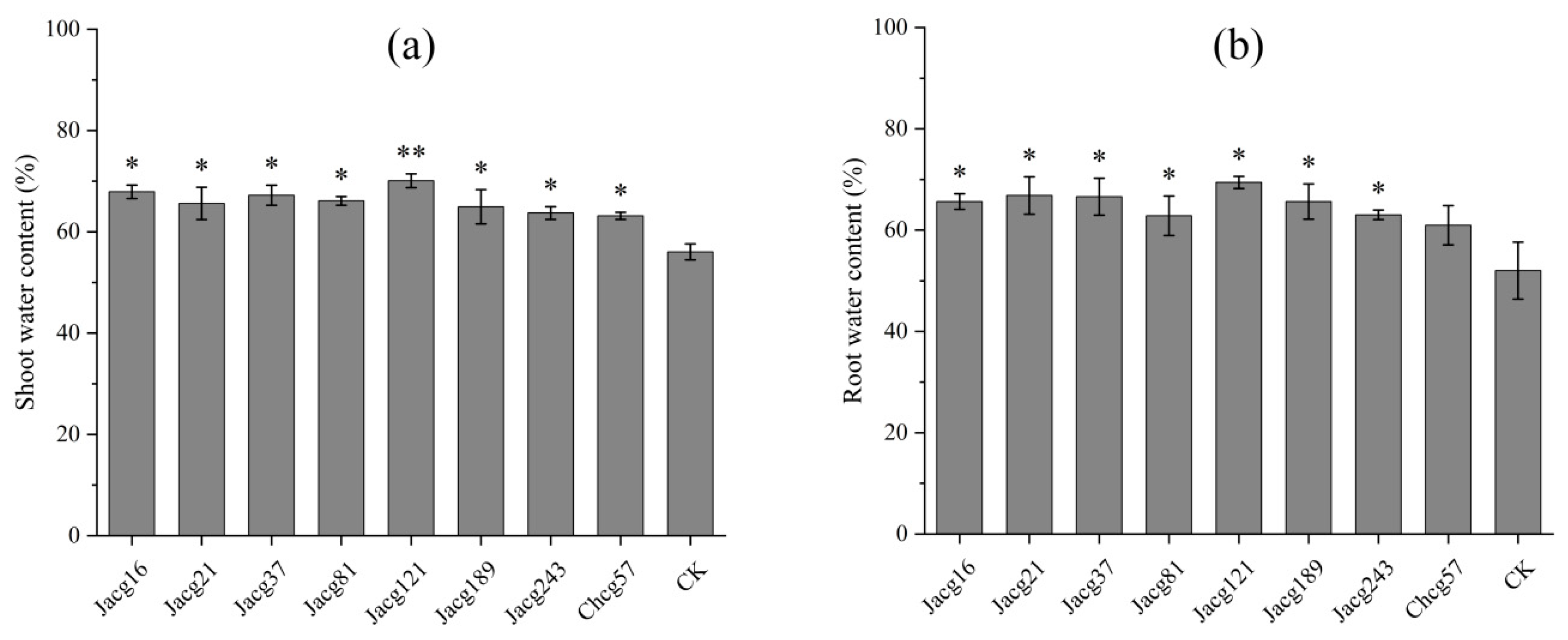
Figure 4.
Effects of different Cenococcum geophilum ecotypes inoculation on the (a) proline content, (b) soluble protein content, (c) malondialdehyde (MDA) content, (d) catalase (CAT) activities, (e) peroxidase (POD) activities and (f) superoxide dismutase (SOD) activities of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%). Data and bars are shown as mean and ±SE of the replicates, respectively (n=3). The statistically significant difference between different C. geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01; *** P<0.001). CK, non-inoculation.
Figure 4.
Effects of different Cenococcum geophilum ecotypes inoculation on the (a) proline content, (b) soluble protein content, (c) malondialdehyde (MDA) content, (d) catalase (CAT) activities, (e) peroxidase (POD) activities and (f) superoxide dismutase (SOD) activities of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%). Data and bars are shown as mean and ±SE of the replicates, respectively (n=3). The statistically significant difference between different C. geophilum ecotypes inoculation and non-inoculation treatments is tested by Mann-Whitney U test (* P<0.05; ** P<0.01; *** P<0.001). CK, non-inoculation.
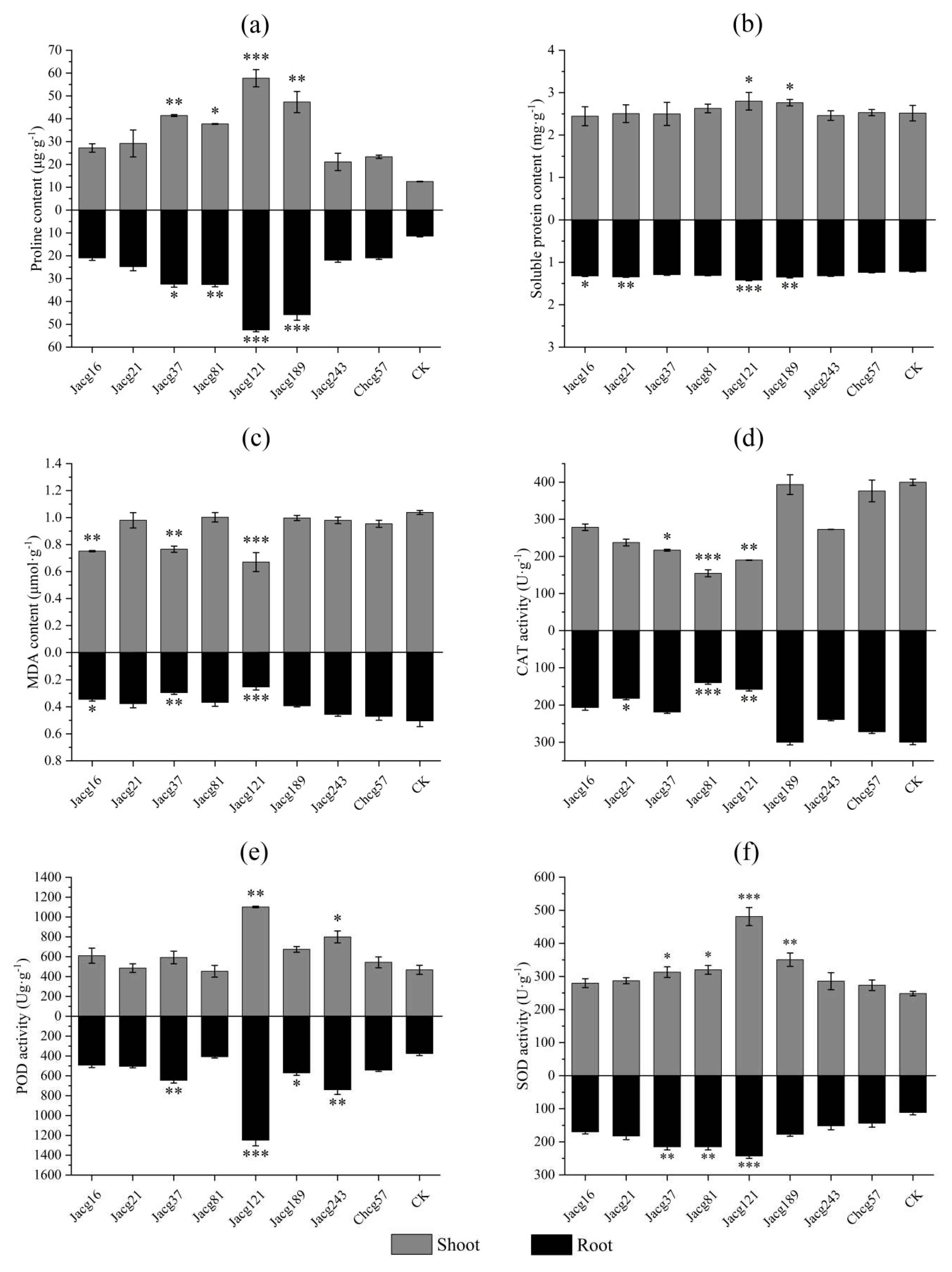
Figure 5.
Volcano (a-d) and Wenn (e, f) maps of differentially expressed genes (DEGs) analysis of drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. Red spot indicates significant up-regulated genes after drought treatment; blue spot indicates significantly down-regulated genes; and gray spot indicates no-change genes. |log2Foldchange| ≥ 2 are used as the screening criteria of DEGs.
Figure 5.
Volcano (a-d) and Wenn (e, f) maps of differentially expressed genes (DEGs) analysis of drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. Red spot indicates significant up-regulated genes after drought treatment; blue spot indicates significantly down-regulated genes; and gray spot indicates no-change genes. |log2Foldchange| ≥ 2 are used as the screening criteria of DEGs.
Figure 6.
Gene ontology (GO) enrichment analysis of the differentially expressed genes (DEGs) in shoots of drought-tolerant (Jacg121) (a) and drought-sensitive (Chcg57) (b) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The abscissa is the GO classification, and the ordinate is the number of genes.
Figure 6.
Gene ontology (GO) enrichment analysis of the differentially expressed genes (DEGs) in shoots of drought-tolerant (Jacg121) (a) and drought-sensitive (Chcg57) (b) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The abscissa is the GO classification, and the ordinate is the number of genes.
Figure 7.
Gene ontology (GO) enrichment analysis of the differentially expressed genes (DEGs) in roots of drought-tolerant (Jacg121) (a) and drought-sensitive (Chcg57) (b) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The abscissa is the GO classification, and the ordinate is the number of genes.
Figure 7.
Gene ontology (GO) enrichment analysis of the differentially expressed genes (DEGs) in roots of drought-tolerant (Jacg121) (a) and drought-sensitive (Chcg57) (b) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The abscissa is the GO classification, and the ordinate is the number of genes.
Figure 8.
The Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of differentially expressed genes (DEGs) in shoots and roots of drought-tolerant (Jacg121) (a, c) and drought-sensitive (Chcg57) (b, d) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The size of the symbol represents the numbers of DEGs involved in the corresponding pathway.
Figure 8.
The Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of differentially expressed genes (DEGs) in shoots and roots of drought-tolerant (Jacg121) (a, c) and drought-sensitive (Chcg57) (b, d) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The size of the symbol represents the numbers of DEGs involved in the corresponding pathway.
Figure 9.
The heatmaps of part up-regulated or down-regulated differentially expressed genes (DEGs) in different pathways of drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The Y- and X-axes represent the DEGs and different samples, respectively. The different colors of the heatmaps, ranging from blue over white to red, represent scaled expression levels of genes with [log2 (FPKM + 1)] across different samples. DS, drought stress; WW, well-watered.
Figure 9.
The heatmaps of part up-regulated or down-regulated differentially expressed genes (DEGs) in different pathways of drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The Y- and X-axes represent the DEGs and different samples, respectively. The different colors of the heatmaps, ranging from blue over white to red, represent scaled expression levels of genes with [log2 (FPKM + 1)] across different samples. DS, drought stress; WW, well-watered.
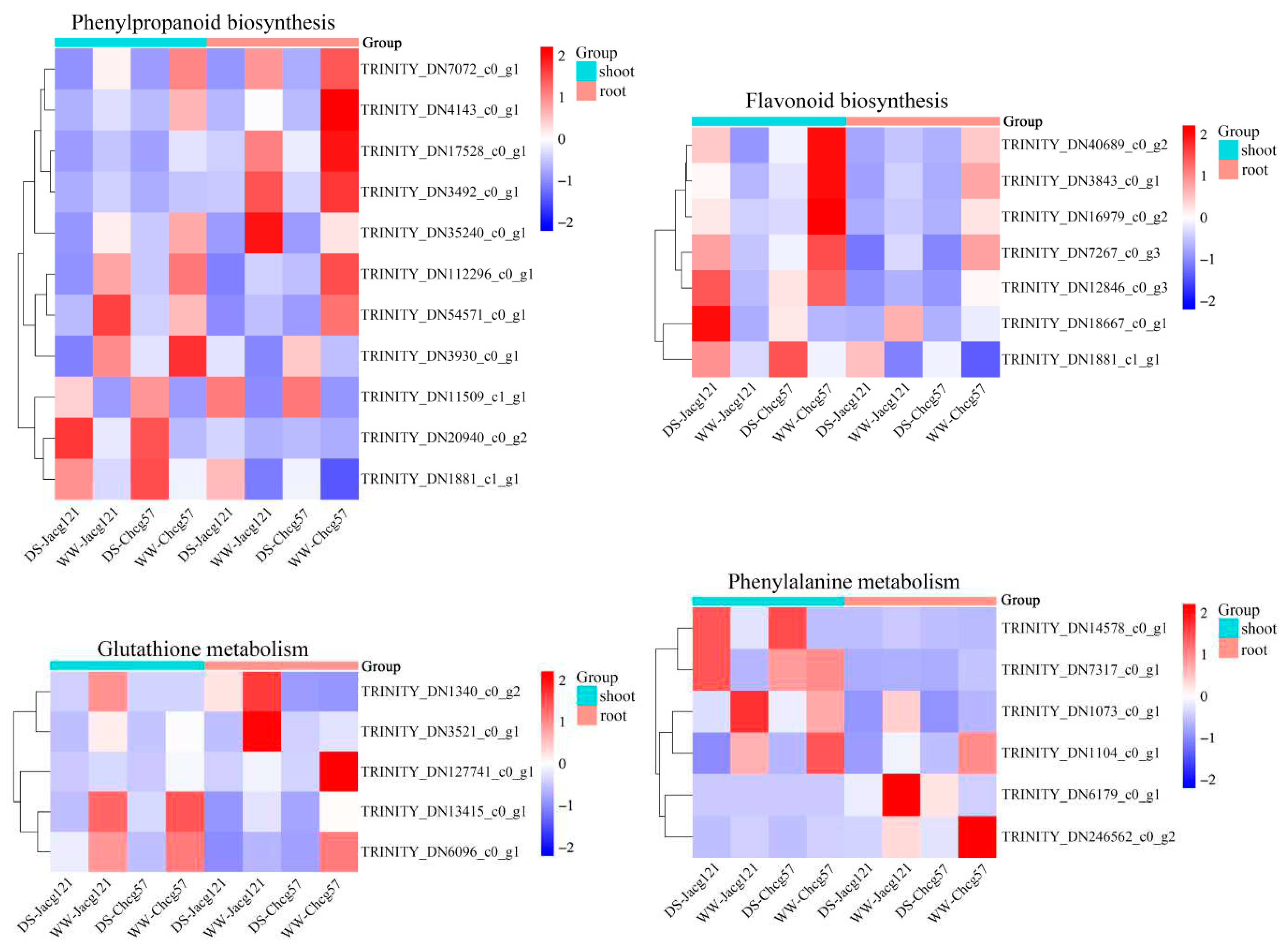
Figure 10.
The relative expression levels of six candidate genes in drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum were analyzed by RT-qPCR after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The relative expression changes of candidate genes in different treatments were calculated by 2- △△Ct method and expressed as mean and ± SE (n=3). The statistically significant difference between drought and well-watered treatments is tested by Student’ s t-test (* P<0.05; ** P<0.01). WW, well-watered; DS, drought stress.
Figure 10.
The relative expression levels of six candidate genes in drought-tolerant (Jacg121) and drought-sensitive (Chcg57) seedlings of Pinus massoniana inoculated by Cenococcum geophilum were analyzed by RT-qPCR after 7 days of drought (field capacity 30%-35%) and well-watered (field capacity 85%-90%) treatments. The relative expression changes of candidate genes in different treatments were calculated by 2- △△Ct method and expressed as mean and ± SE (n=3). The statistically significant difference between drought and well-watered treatments is tested by Student’ s t-test (* P<0.05; ** P<0.01). WW, well-watered; DS, drought stress.
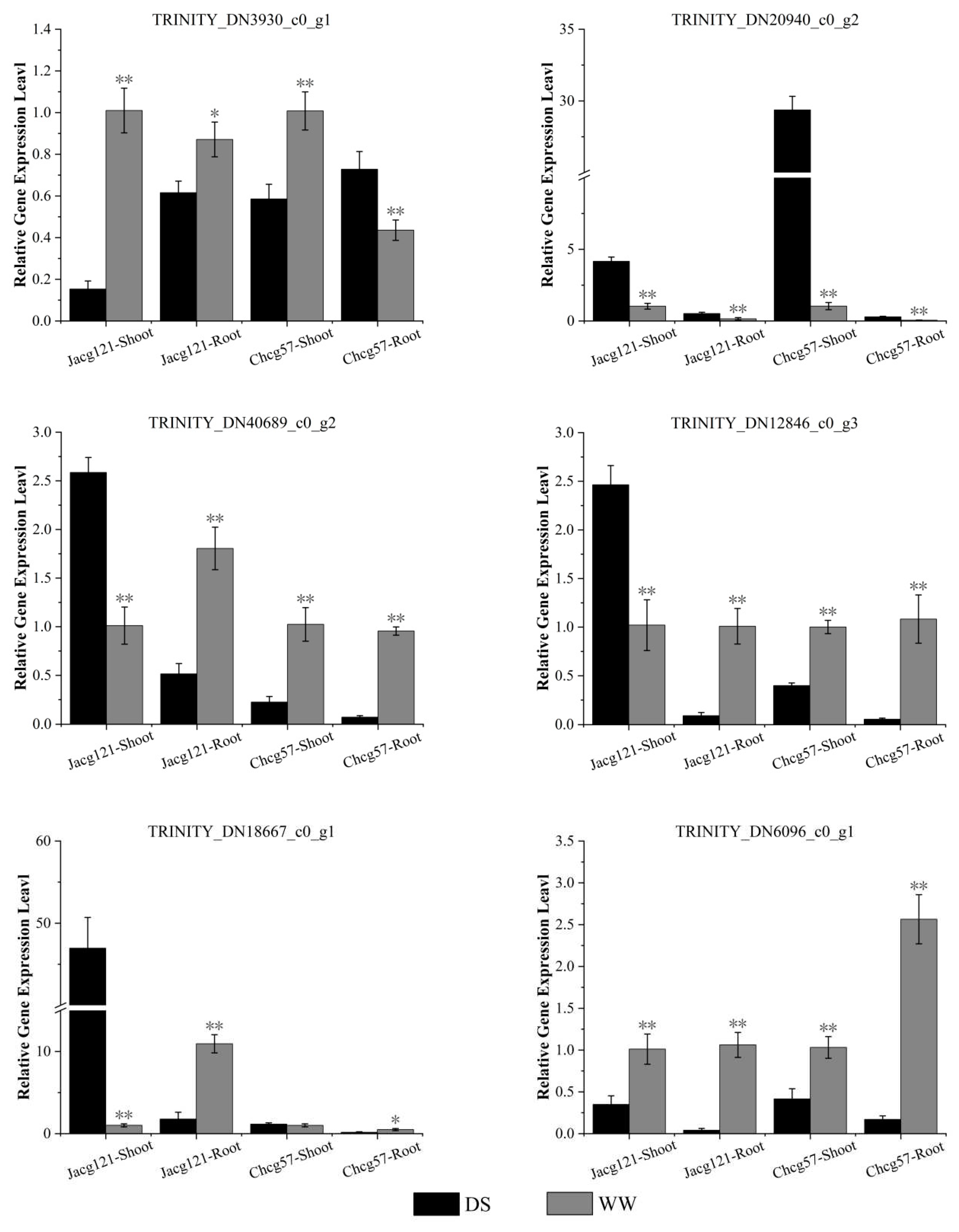

Table 1.
Effects of inoculation with different Cenococcum geophilum ecotypes on photosynthetic parameters of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%).
Table 1.
Effects of inoculation with different Cenococcum geophilum ecotypes on photosynthetic parameters of Pinus massoniana seedlings after 7 days of drought stress treatment (field capacity 30%-35%).
| ID | Pn (μmol·m-2·s-1) |
Gs (mol·m-2·s-1) |
Ci (μmol·m-2·s-1) |
Tr (mmol·m-2·s-1) |
|---|---|---|---|---|
| Jacg16 | 1.57±0.06 | 0.023±0.001 | 322.17±4.37*** | 0.95±0.01 |
| Jacg21 | 2.45±0.32* | 0.019±0.004 | 214.01±3.12 | 1.00±0.02 |
| Jacg37 | 3.22±0.03** | 0.036±0.001*** | 280.97±1.73* | 1.72±0.01*** |
| Jacg81 | 3.53±0.08*** | 0.026±0.007* | 207.72±8.04 | 1.16±0.03* |
| Jacg121 | 3.58±0.12*** | 0.046±0.003*** | 303.57±4.77** | 2.25±0.01*** |
| Jacg189 | 1.93±0.02 | 0.035±0.003** | 327.84±0.94*** | 1.59±0.01** |
| Jacg243 | 1.47±0.04 | 0.020±0.001 | 261.70±3.00 | 0.98±0.01 |
| Chcg57 | 2.30±0.08 | 0.024±0.007 | 280.65±1.61* | 1.20±0.03* |
| CK | 1.08±0.01 | 0.008±0.001 | 161.93±3.55 | 0.93±0.01 |
Note: Data expressed as mean ± SE (n=3). The statistically significant difference between different C. geophilum ecotypes inoculation and non-inoculation treatments was tested by Mann-Whitney U test (* P<0.05; ** P<0.01; *** P<0.001). Pn, net photosynthetic rate; Gs, stomatal conductance; Ci, intercellular CO2 concentration; Tr, transpiration rate; CK, non-inoculation.
Table 2.
The value of comprehensive index (CI), index weight (Wj), µ(Xj), D value (comprehensive evaluation of drought tolerance) and comprehensive evaluation in Pinus massoniana seedlings inoculated by different Cenococcum geophilum ecotypes after 7 days of drought stress (field capacity 30%-35%).
Table 2.
The value of comprehensive index (CI), index weight (Wj), µ(Xj), D value (comprehensive evaluation of drought tolerance) and comprehensive evaluation in Pinus massoniana seedlings inoculated by different Cenococcum geophilum ecotypes after 7 days of drought stress (field capacity 30%-35%).
| Code | CI1 | CI2 | CI3 | μ(X1) | μ(X2) | μ(X3) | D | Comprehensive evaluation* |
|---|---|---|---|---|---|---|---|---|
| Jacg16 | -0.083 | -0.382 | -1.990 | 0.437 | 0.354 | 0.000 | 0.386 | 6 |
| Jacg21 | -0.118 | -1.057 | -0.437 | 0.427 | 0.154 | 0.450 | 0.391 | 5 |
| Jacg37 | 0.555 | -0.492 | -0.107 | 0.610 | 0.321 | 0.545 | 0.564 | 3 |
| Jacg81 | 0.188 | -1.574 | 1.465 | 0.510 | 0.000 | 1.000 | 0.483 | 4 |
| Jacg121 | 1.991 | 0.511 | 0.417 | 1.000 | 0.619 | 0.697 | 0.919 | 1 |
| Jacg189 | 0.319 | 1.792 | 0.337 | 0.546 | 1.000 | 0.673 | 0.621 | 2 |
| Jacg243 | -0.454 | 0.230 | -0.627 | 0.336 | 0.536 | 0.394 | 0.369 | 7 |
| Chcg57 | -0.704 | 0.580 | -0.060 | 0.268 | 0.640 | 0.559 | 0.347 | 8 |
| CK | -1.693 | 0.393 | 1.002 | 0.000 | 0.584 | 0.866 | 0.160 | 9 |
| Index weight (wj) | 0.770 | 0.140 | 0.09 |
Note: CK, non-inoculation; *, the lower level indicates the higher drought tolerance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



