
Submitted:
21 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
The rapid and accurate detection of parasites is crucial for timely curative intervention in parasitosis and for epidemiological surveillance. To meet the needs of clinical diagnosis, it is imperative to develop a diagnostic tool based on nucleic acid that combines the sensitivity and specificity of established nucleic acid amplifica-tion tests with the speed, cost-effectiveness, and convenience of isothermal amplification methods. A new nu-cleic acid detection method, utilizing the clustered regularly interspaced short palindromic repeats (CRISPR)-associated (Cas) nuclease, holds promise for point-of-care testing. Specifically, the CRISPR-Cas12a system has demonstrated numerous advantages for detecting parasites, with hopeful outcomes for identify-ing malaria, toxoplasmosis, and other parasitic ailments. This review provides an overview of how CRISPR-Cas12a can be utilized for parasite detection, evaluates its advantages and disadvantages, and sug-gests ways to improve the efficiency and sensitivity of CRISPR-Cas12a-based assays.
Keywords:
detection
; CRISPR
; suboptimal crRNA
; light-activated crRNA
; tandem repeats
; POCT
1. Introduction
Parasitosis, caused by parasitic infection, is a prevalent cause of morbidity among humans worldwide. Tropical zones, particularly those that are impoverished, conflicted, or unsanitary, serve as endemic foci for a range of parasitic diseases [1]. The World Health Organization (WHO) has reported that annually 48.4 million cases and 59,724 deaths are attributed to the prevalence of 14 significant parasites, representing a total burden of 8.78 million disability-adjusted life years (DALYs). Of these instances, 48% represent foodborne parasitic diseases, accounting for 76% of the DALYs [2]. Transmission through contaminated food is prevalent in low- and middle-income countries [2]. Approximately 241 million cases of malaria and 627,000 deaths resulting from malaria were reported across the world in 2020. Innumerable deaths were caused by other parasitic infections, most notably neglected tropical diseases (NTDs) [3,4].
Unlike the vast majority of bacterial and viral infections, which have an incubation period as short as hours to days, parasitic diseases tend to have an incubation period of weeks or even months [4]. The incubation period of specific parasitic diseases, like alveolar echinococcosis, can extend up to 10 years [5]. Therefore, early and precise detection of parasitosis is imperative for timely curative interventions and the prevention of pandemics.
A promising new method for nucleic acid detection utilizes the CRISPR-associated (Cas) nuclease, which can overcome the limitations of instrument dependence and laborious operational processes. The present review compares the CRISPR-Cas12a system with alternative molecular methods for detecting parasitic diseases. Emphasis is placed on the enhancement of one-pot recombinase polymerase amplification (RPA) Cas12a and even the improvement of CRISPR-Cas12/Cas13 detection techniques.
2. Application of Nucleic Acids Amplification Tests in Parasite Detection
Currently, the diagnosis of parasitic diseases relies on various methods, including microscopy, epidemiology, pathophysiology, immunodiagnostics, and nucleic acid amplification tests (NAATs). Among these options, the microscopic detection of parasites remains the most reliable [6,7]. In underdeveloped regions with high rates of parasitosis, however, skilled operators are often scarce, making this technique challenging to implement [8]. Furthermore, this approach is unsuitable for conditions linked to parasites at developmental stages that are challenging to detect in blood or stool specimens.
Immunoassay-based diagnostic measures have been in use for decades and are widely utilized for detecting parasites. However, the application of immunoassays for parasitosis diagnosis has been limited by several drawbacks, including the possibility of false negatives and false positives [9,10].
Molecular detection of nucleic acids demonstrates superior sensitivity, specificity, and reproducibility compared to alternative methods (Table 1). As a result, NAATs are the preferred molecular detection tools due to their ability to amplify trace amounts of DNA and RNA, allowing for highly specific detection by complementary nucleotide pairing. Although these techniques have been used to establish dependable parasitosis diagnostic methods for malaria, filariasis, toxoplasmosis, and echinococcosis [11,12,13,14], such methods require long reaction times, complex handling, expensive laboratory equipment, and highly skilled technicians.
Isothermal amplification technology compensates for these shortcomings. In contrast to PCR, isothermal amplification technology, exemplified by loop-mediated isothermal amplification (LAMP) and RPA, significantly reduces the reaction time and dependence on instruments. RPA is an efficient method for on-site detection due to its simple primer design, low temperature requirements, and easy storage [15,16]. However, because the reaction occurs at low temperatures, non-specific amplification by RPA can lead to low specificity. Isothermal amplification technology has been employed in diagnosing various parasitic diseases, including malaria, leishmaniasis, and schistosomiasis [15,16,17].
In addition to the selection of detection methods for NAATs, which has a significant impact on the accuracy and sensitivity of parasitic disease diagnosis, the selection of target genes is also a key point in the establishment of detection methods. Along with 18S ribosomal RNA (rRNA), Internal Transcribed Spacer (ITS), and mitochondrial genes, stable tandem repeats are beginning to come into focus. In most parasites, repetitive sequences make up a substantially greater proportion of the genome when compared to coding sequences, comprising an estimated 20% or even exceeding 30% [18,19]. Numerous tandem repeats have been utilized to detect multiple protozoans and worms, such as Trypanosoma cruzi, Onchocerca volvulus, and Schistosoma mansoni (Table 2).
Point-of-care testing (POCT), a priority for strategies relying on mass drug administration to control many neglected tropical diseases (NTDs), is a medical diagnostic tool that can be used near or at the point-of-care, allowing for on-site testing [43]. Therefore, it is imperative to develop nucleic acid-based diagnostic tools that combine the sensitivity and specificity of established NAAT with the convenience, cost-effectiveness, and speed of isothermal amplification methods. CRISPR-based diagnostics have the potential to fulfill all these requirements (Table 3).
3. CRISPR-Cas12a for POCT
3.1. Discovery of CRISPR
The CRISPR-Cas system was initially identified by Ishino in 1987 [44] and named in 2002 [45]. Extensive research has since focused on identifying and characterizing the proteins and molecules involved in the CRISPR-Cas system [46]. CRISPR-Cas systems are composed of Cas genes organized in operons and a CRISPR array, which consists of unique genome-targeting sequences (called spacers) interspersed with identical repeats [47]. Jennifer Doudna and Emmanuelle Charpentier were the first to illustrate the potential of the CRISPR-Cas9 system as a means of gene editing [47]. Subsequently, Janice Chen and Feng Zhang led the primary studies of CRISPR-Cas12a [48] and CRISPR-Cas13a [49] for the purpose of detection.
3.2. CRISPR-Cas12a Is More Suitable for Rapid On-Site Detection
The Cas12a effector protein, also referred to as the Cpf1 effector protein, is a programmable RNA-guided DNA nuclease that was identified as part of the type-V class Ⅱ CRISPR-Cas system [50]. This protein may have come from a distinct TnpB transposase gene family [51]. Compared to Cas9, the design of Cas12a is simpler and more cost-effective since it only needs one CRISPR RNA (crRNA) and no trans-activating crRNA (tracrRNA).
Cas12a accurately identifies target sequences and double-stranded DNA, creating gaps by recognizing T-rich protospacer adjacent motif (PAM) sequences and catalyzing its own crRNA maturation [52]. It was later discovered that Cas12a exhibits collateral activity and can cleavage single-stranded DNA without the presence of a complementary crRNA sequence [48]. The non-target strand and RuvC domains are highly flexible, with the target strand being particularly flexible when located at the nuclease active site. As a result, the RuvC domain becomes significantly active during R-loop formation, enabling the entry of single-stranded DNA into the enzyme's active center for degradation [53].
Therefore, the target DNA has the potential to act as an activator, triggering both cis- and trans-cleavage events of the Cas12a nuclease. The FQ reporter in the system is cleaved, releasing fluorescence signal that is then measured to detect pathogen. The CRISPR-Cas12a system has found extensive application in detecting COVID-19, with studies demonstrating a detection limit as low as 5-10 copies utilizing the CRISPR-Cas12a system and isothermal amplification technology [54,55]. The sensitivity of the method was comparable to that of qPCR, and results were obtained via naked-eye observation within 45 minutes. These advantages enable the detection of parasites through the CRISPR-Cas12a system and offers benefits beyond those of other nucleic acid detection methods (Table 2).
3.3. CRISPR-Cas12a Has Been Applied to Parasite Detection
The technique's application to diagnose parasitic diseases, like malaria, has undergone a thorough evaluation. Asymptomatic carriers considerably hinder controlling and eradicating the parasite. Achieving malaria eradication mandates a hyper-sensitive diagnosis of infections with a low parasitic load [4]. Unfortunately, resource-limited areas experience frequent malaria outbreaks, posing a challenge to screening parasite carriers. Lee and colleagues developed a nucleic acid diagnostic method to detect Plasmodium falciparum by combining CRISPR-Cas12a with RT-RPA [56].
The method involves heating human serum, whole blood, or dried blood spots in buffer at 95 °C for 10 minutes, followed by the transfer of the suspended sample to a pre-mixed Cas12a-RPA system. The mixture is then incubated at 40 °C for 30 minutes. The reaction outcomes can be observed using a plate reader or a handheld fluorometer, facilitating on-site detection. This technique significantly lowers the LOD to 0.36 parasites per microliter, which is well under the WHO's rapid diagnostic test threshold of 200 parasites per microliter [57].
Additionally, the CRISPR-Cas12a combination with RPA has been successfully utilized for detecting Toxoplasma gondii [58]. The detection system achieved a sensitivity of 3.3 genome copies per microliter, surpassing real-time fluorescent RPA (33 genome copies per microliter) and other comparable methods [59]. Furthermore, this system was utilized to examine a range of parasites, including Cryptosporidium parvum [60,61], Enterocytozoon hepatopenaei [62], and Heterodera schachtii [63] (Table 4).
It is estimated that 47% of the worldwide population lacks adequate access to medical diagnostic tools, particularly in underdeveloped areas [64]. Cas12a and RPA-based diagnostic technologies are anticipated to resolve this problem since the method satisfies almost all of the POCT requirements [65]. Nevertheless, many concerns still need to be addressed because of the brief time of research and limited large-scale clinical trials. The sensitivity of these assays for POCT, particularly concerning specific sample preparation, ought to be enhanced. Furthermore, the time required to attain results should be decreased.
4. Optimization of the CRISPR-Cas12a One-Pot Detection Assay
Due to the low initial concentration of the target gene in a sample and the kinetic rates that result in an amplification-free LOD in the picomolar range under standard assay conditions [66], Cas12a detection often requires an amplification process prior to application. That means that signal amplification usually involves two processes. Target genes were initially amplified using RPA or LAMP techniques. The resulting amplified products were subsequently transferred to the Cas12a system for cleavage, followed by fluorescence signal generation. To streamline operations and prevent cross-contamination during field tests, the one-pot method is now predominantly utilized. This assay allows the amplification and cleavage of Cas12a to occur simultaneously. However, this leads to the cis-cleavage of Cas12a, which reduces the concentration of the target genes while RPA enhances it. Therefore, it is crucial to optimize amplification in the initial phase of the reaction.
4.1. One-Pot and One Step
4.1.1. Determinants of Cas12a Enzyme Kinetics
Several studies have reported rapid single-turnaround, cis-cleavage reactions at low target concentrations, with typical reaction times of approximately 100 seconds [67]. A Michaelis-Menten model for Cas12a trans-cleavage activity was established and validated by a team from Stanford University. This was achieved through the utilization of varying concentrations of substrates, targets, and crRNAs [66,67]. The authors suggest that the concentration of trans-cleavage product formed over time can be described using the following scaling equation:
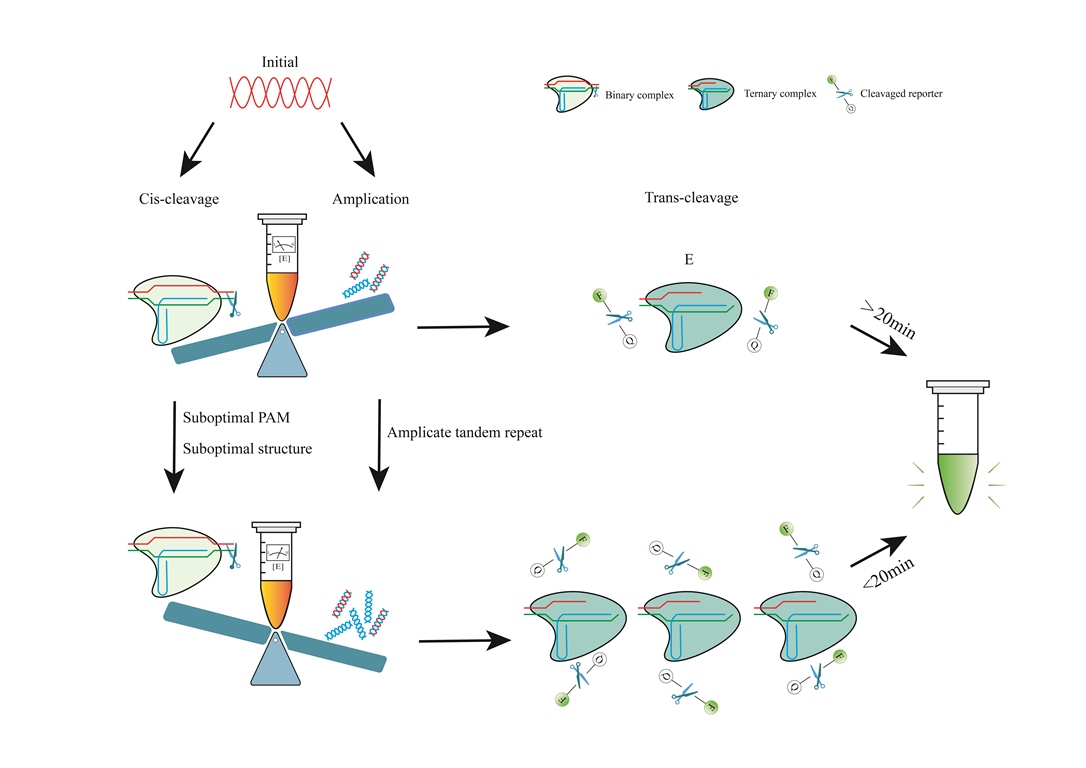
The production efficiency of trans-cleavage product P is influenced by both reaction time and τ. To refer to the target-activated Cas12-crRNA-target DNA complex, use E, and subsequently, [E] represents the concentration of this complex. The characteristic time to complete trans-cleavage is governed by the time scale τ, which is proportional to KM and inversely proportional to kcat and [E] [67]. The rate constant kcat/KM of enzymatic reactions is affected by the Cas type, crRNA, incubation time, pH, and temperature [66]. During the early stages of the reaction, [E] equals the concentration of the target molecule (c), which depends on c0, amplification, and cis-cleavage. Therefore, one could use a suboptimal crRNA to weaken cis-cleavage or employ other methods to ensure that amplification dominates the pre-reaction period, resulting in a rapid increase of [E] (Figure 1).
4.1.2. Reduced crRNA Efficiency by PAM
In the CRISPR-Cas system, the effector nuclease must identify the PAM neighboring the target site to initiate target recognition [68]. Studies of the crystal structure of the LbCas12a-crRNA binary complex [69] and the AsCas12a-crRNA-target DNA ternary complex [70,71] disclose the mechanisms involved in Cas12a and crRNA recognition, as well as the operations of crRNA-directed DNA targeting and PAM recognition. These findings suggest that the PAM-binding channel of Cas12a is flexible in conformation, allowing for the identification of both canonical and non-canonical PAMs [72]. LbCas12a and AsCas12a identify TTTV and CTTV/TCTV/TTCV as canonical and non-canonical PAM, respectively [72,73].
In 2022, a Chinese team conducted a one-pot test called sPAMC, which refers to a suboptimal PAM of Cas12a-based test [74]. In comparing collateral activity, crRNAs utilizing suboptimal PAMs demonstrated lower potency and slower kinetics in comparison to those utilizing canonical PAMs. Nevertheless, over 80% of 120 suboptimal PAMs displayed quicker reactions than those of canonical PAMs in the one-pot reactions.
The emergence of the target amplicon occurred 2 minutes after the one-pot reaction utilizing suboptimal PAM, as opposed to 8–10 minutes for canonical PAM. Using a suboptimal PAM with varied concentrations of Cas12a/crRNA ribonucleoprotein produced steady kinetic curves, unlike with traditional PAMs. In one-pot reactions, numerous uncommon PAMs (such as VTTV, TCTV, and TTVV) and some TRTV, TTNT, and YYYN PAMs (excluding TTTV) outperformed canonical PAMs. The SARS-CoV-2 diagnostic method established using suboptimal PAM yields a sensitivity comparable to that of qPCR. The reaction time is only 15 minutes, and the variation among samples is less than 30% [74].
4.1.3. Reduced crRNA Efficiency by Structure
Suboptimal crRNAs can be selected based on their structure while ensuring specificity. If CRISPR-Cas9 cleavage is an energy-driven process, its efficiency relies heavily on nucleotide hybridization and changes in folding free energy [75]. The stability of gRNA-DNA for gRNAs exhibiting different efficiencies significantly varies. When local sliding is examined, an energy model accurately predicts the efficiency of gRNAs. Similar studies for Cas12a are currently unavailable. However, it is important to consider the use of optimal crRNAs when developing a one-pot detection method. Initial support for the feasibility of this approach is demonstrated by our recent experimental findings.
4.2. One-Pot but Two Steps
4.2.1. Light-Activated crRNA to Initiate Cleavage
Controlling chemical reactions through photocontrolled techniques can be achieved in a non-contact manner within seconds. This technology has been extensively used in both CRISPR-Cas9 research and practice [76,77,78], and it has also been progressively refined for CRISPR-Cas12a detection [79]. Initially, the CRISPR-Cas12a system was blocked by a photo-cleaved linker containing crRNA to ensure optimal RPA performance. After amplification, the Cas12a detection system was activated via light to initiate trans-cleavage and produce fluorescence signals [79]. However, this approach requires ongoing optimization of the ratio of the photo-cleaved linker to crRNA, and the lack of pre-binding of crRNA to Cas protein negatively impacts the stability of the Cas12a-crRNA complex.
The same group then developed a novel CRISPR-Cas12a detection assay that uses 6-nitropiperonyloxymethyl-caged thymidine (NPOM-dt) to modify crRNA [80]. This method involves caging crRNA to prevent base pairing between it and the target, rather than binding it to the Cas enzyme. The rapid activation can be attained by photoinduced decaying, which makes this approach simpler, faster, and more stable. It should be noted that optimizing the irradiation time and the number and position of NPOM may need to be reconsidered for different pathogens. In regards to POCT, challenges persist with reagent storage conditions, actual amplification time, and the portability of illumination devices.
4.2.2. Physically Separate the Two Processes
In addition to performing two reactions simultaneously in one tube, it is also possible to physically separate the two reaction systems in one tube to allow for sequential progression [43,81,82,83]. The CRISPR/Cas12a reagents were spun down for cleavage after DNA amplification by leveraging the physical property of the protein-containing liquid's enhanced surface tension [84]. Initially, the RPA reaction takes place at the bottom of the tube, while the CRISPR/Cas12a reaction is located at the lid, separate from the reaction. After amplifying for 20 minutes, briefly spin the CRISPR/Cas12a reagent into the reaction mixture without opening the tube. The reaction will continue, and the RPA amplicon will activate the Cas12a nuclease to trans-cleave the fluorescent ssDNA-FQ molecule, resulting in a fluorescent signal. However, this method can be cumbersome, particularly in large-scale POCT.
In brief, cis-cleavage plays a crucial role as the rate-determining step for overall performance in one-pot reactions [67,74]. During the initial stage, low-concentration targets are diminished due to cis-cleavage, which results in a slow and unstable accumulation of amplicons. Consequently, the growth of the signal decreases or may even disappear altogether (Figure 2). The kcat/KM of the enzyme can be reduced by utilizing a suboptimal PAM or structure, which slows cis-cleavage. This results in a balance between the two signal amplification processes of RPA and trans-cleavage. Through careful engineering of primer design, crRNA design [74,85], reaction system [86,87], reporter selection [88,89], and reaction conditions, isothermal amplification and CRISPR detection can be effectively combined in a one-pot reaction. The light-activated CRISPR RNA and spatially isolated reagents may enhance the efficiency of the one-pot (but two steps) process. The resulting method provides a streamlined and efficient approach to detecting specific nucleic acid sequences with high sensitivity and specificity.
5. Conclusions
Parasites are prevalent in the natural world, particularly in underdeveloped regions, and result in high disability-adjusted life years and substantial economic losses annually. The emergence of CRISPR, and specifically recent examinations of Cas12a, compensates for the limitations of isothermal amplification and presents a fresh approach for POCT. With the collateral-activity of Cas12a, results can be evaluated intuitively via the inclusion of fluorophores. Combined with RPA, samples with even small amounts of pathogens can be quickly and accurately tested at the point of detection.
For POCT, the one-pot method is without a doubt the best option due to its ability to prevent cross-contamination and significantly simplify the procedure. Nonetheless, current one-pot detection techniques come with several limitations, including extended reaction times, low sensitivity, complicated operation, and reliance on sample pretreatment. Additionally, the utilization of RPA has restricted the advancement of CRISPR assays somewhat. As the most commonly used partner of CRISPR one-pot method, RPA kits are only sold by few companies, with high price and unstable supply [90].
By balancing the two processes of amplification and cleavage with a suboptimal PAM or structure, the detection performance of the one-pot method can be improved. With suboptimal conditions, PAM or crRNA's limitation on target genes may be weakened, thereby expanding the pools of target genes. In addition, light-activated crRNA and spatial isolation enable two reactions to proceed in one-pot, one after the other, without opening the lid. Furthermore, incorporating tandem repeats as targets can significantly enhance the detection's amplification efficiency and sensitivity, regardless of sample preparation methods. One-pot RPA-Cas12a is significantly improved with the use of these methods. These ways can enhance not only RPA-Cas12a but also all Cas12a detection methods involving amplification. Furthermore, it is important to assess these concepts not only in Cas12a, but also in other CRISPR systems, including Cas12b and even Cas13.
In the future, for the application of CRISPR-Cas12a to POCT, it is necessary to continually optimize one-pot method detection efficiency and identify a more compatible isothermal amplification technology. Also, the sample preparation for testing is crucial; otherwise, CRISPR detection would remain limited to the laboratory.
Author Contributions
FH: XZ, and XL conceived this manuscript; XL, WT, HZ, and JS searched and collated literature; XL, ZD and RJ drew figures and tables; XL wrote the manuscript; FH and XZ revised the manuscript. All authors read and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the Open Project Program of State Key Laboratory of Hulless Barley and Yak Germplasm Resources and Genetic Improvement (XZNKY-CZ-2022-016-08).
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
CRISPR: Clustered Regularly Interspaced Short Palindromic Repeats; crRNA, CRISPR RNA; DALY, disability adjusted life year; NAAT, nucleic acid amplification test; NTD, neglected tropical disease; PAM, protospacer adjacent motif; POCT, Point-of-care testing.
References
- WHO. Working to overcome the global impact of neglected tropical diseases: first WHO report on neglected tropical diseases. Available online: (accessed on 11 February).
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fevre, E.M.; Sripa, B.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS Med 2015, 12, e1001920. [Google Scholar] [CrossRef] [PubMed]
- WHO. World malaria report 2021. Available online: (accessed on 6 December).
- Theel, E.S.; Pritt, B.S. Parasites. Microbiol Spectr 2016, 4. [Google Scholar] [CrossRef]
- Meinel, T.R.; Gottstein, B.; Geib, V.; Keel, M.J.; Biral, R.; Mohaupt, M.; Brügger, J. Vertebral alveolar echinococcosis—a case report, systematic analysis, and review of the literature. The Lancet Infectious Diseases 2018, 18, e87–e98. [Google Scholar] [CrossRef]
- Daly, R.; Chiodini, P.L. Laboratory investigations and diagnosis of tropical diseases in travelers. Infectious disease clinics of North America 2012, 26, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.S.; Fung, K.S.; Chau, S.; Poon, R.W.; Wong, S.C.; Yuen, K.Y. Molecular diagnosis in clinical parasitology: when and why? Exp Biol Med (Maywood) 2014, 239, 1443–1460. [Google Scholar] [CrossRef] [PubMed]
- Kettelhut, M.M.; Chiodini, P.L.; Edwards, H.; Moody, A. External quality assessment schemes raise standards: evidence from the UKNEQAS parasitology subschemes. Journal of clinical pathology 2003, 56, 927–932. [Google Scholar] [CrossRef]
- Rosenblatt, J.E. Laboratory diagnosis of infections due to blood and tissue parasites. Clin Infect Dis 2009, 49, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Kalogeropoulos, D.; Sakkas, H.; Mohammed, B.; Vartholomatos, G.; Malamos, K.; Sreekantam, S.; Kanavaros, P.; Kalogeropoulos, C. Ocular toxoplasmosis: a review of the current diagnostic and therapeutic approaches. International ophthalmology 2022, 42, 295–321. [Google Scholar] [CrossRef]
- Qian, W.; Yan, W.; Lv, C.; Bai, R.; Wang, T.; Wei, Z.; Zhang, M. Molecular Detection and Genotyping of Toxoplasma gondii and Neospora caninum in Slaughtered Goats in Central China. Foodborne pathogens and disease 2020, 17, 348–356. [Google Scholar] [CrossRef]
- Waitumbi, J.N.; Gerlach, J.; Afonina, I.; Anyona, S.B.; Koros, J.N.; Siangla, J.; Ankoudinova, I.; Singhal, M.; Watts, K.; Polhemus, M.E.; et al. Malaria prevalence defined by microscopy, antigen detection, DNA amplification and total nucleic acid amplification in a malaria-endemic region during the peak malaria transmission season. Trop Med Int Health 2011, 16, 786–793. [Google Scholar] [CrossRef]
- Alhassan, A.; Li, Z.; Poole, C.B.; Carlow, C.K. Expanding the MDx toolbox for filarial diagnosis and surveillance. Trends Parasitol 2015, 31, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Momcilovic, S.; Cantacessi, C.; Arsic-Arsenijevic, V.; Otranto, D.; Tasic-Otasevic, S. Rapid diagnosis of parasitic diseases: current scenario and future needs. Clin Microbiol Infect 2019, 25, 290–309. [Google Scholar] [CrossRef] [PubMed]
- Lalremruata, A.; Nguyen, T.T.; McCall, M.B.B.; Mombo-Ngoma, G.; Agnandji, S.T.; Adegnika, A.A.; Lell, B.; Ramharter, M.; Hoffman, S.L.; Kremsner, P.G.; et al. Recombinase Polymerase Amplification and Lateral Flow Assay for Ultrasensitive Detection of Low-Density Plasmodium falciparum Infection from Controlled Human Malaria Infection Studies and Naturally Acquired Infections. J Clin Microbiol 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.A.; Faisal, K.; Chowdhury, R.; Nath, R.; Ghosh, P.; Ghosh, D.; Hossain, F.; Abd El Wahed, A.; Mondal, D. Evaluation of molecular assays to detect Leishmania donovani in Phlebotomus argentipes fed on post-kala-azar dermal leishmaniasis patients. Parasit Vectors 2021, 14, 465. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Rong, R.; Zhang, H.Q.; Shi, C.J.; Zhu, X.Q.; Xia, C.M. Sensitive and rapid detection of Schistosoma japonicum DNA by loop-mediated isothermal amplification (LAMP). Int J Parasitol 2010, 40, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A.; von Sternberg, R. Why repetitive DNA is essential to genome function. Biol Rev Camb Philos Soc 2005, 80, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhang, W.; Zhang, L.; Zhang, Z.; Li, J.; Lu, G.; Zhu, Y.; Wang, Y.; Huang, Y.; Liu, J.; et al. The genome of the hydatid tapeworm Echinococcus granulosus. Nat Genet 2013, 45, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Prediger, E.; Huecas, M.E.; Nogueira, N.; Lizardi, P.M. Minichromosomal repetitive DNA in Trypanosoma cruzi: its use in a high-sensitivity parasite detection assay. Proc Natl Acad Sci U S A 1984, 81, 3356–3360. [Google Scholar] [CrossRef] [PubMed]
- Homan, W.L.; Vercammen, M.; De Braekeleer, J.; Verschueren, H. Identification of a 200- to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. Int J Parasitol 2000, 30, 69–75. [Google Scholar] [CrossRef]
- Demas, A.; Oberstaller, J.; DeBarry, J.; Lucchi, N.W.; Srinivasamoorthy, G.; Sumari, D.; Kabanywanyi, A.M.; Villegas, L.; Escalante, A.A.; Kachur, S.P.; et al. Applied genomics: data mining reveals species-specific malaria diagnostic targets more sensitive than 18S rRNA. J Clin Microbiol 2011, 49, 2411–2418. [Google Scholar] [CrossRef]
- Singh, R.; Singh, D.P.; Gupta, R.; Savargaonkar, D.; Singh, O.P.; Nanda, N.; Bhatt, R.M.; Valecha, N. Comparison of three PCR-based assays for the non-invasive diagnosis of malaria: detection of Plasmodium parasites in blood and saliva. European journal of clinical microbiology & infectious diseases : official publication of the European Society of Clinical Microbiology 2014, 33, 1631–1639. [Google Scholar] [CrossRef]
- Azam, M.; Upmanyu, K.; Gupta, R.; Sruthy, K.S.; Matlani, M.; Savargaonkar, D.; Singh, R. Development of Two-Tube Loop-Mediated Isothermal Amplification Assay for Differential Diagnosis of Plasmodium falciparum and Plasmodium vivax and Its Comparison with Loopamp™ Malaria. Diagnostics 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Rosenzvit, M.C.; Canova, S.G.; Kamenetzky, L.; Ledesma, B.A.; Guarnera, E.A. Echinococcus granulosus: Cloning and characterization of a tandemly repeated DNA element. Exp. Parasitol. 1997, 87, 65–68. [Google Scholar] [CrossRef]
- Abbasi, I.; Hamburger, J.; Raoul, F.; Craig, P.S.; Campos-Ponce, M.; Branzburg, A.; Hafez, S.K.A. Copro-Diagnosis of Echinococcus Granulosus Infection in Dogs by Amplification of a Newly Identified Repeated DNA Sequence. The American Journal of Tropical Medicine and Hygiene 2003, 69, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.; Vallejo, V.; Mossie, K.G.; Ortiz, D.; Agabian, N.; Flisser, A. Isolation and characterization of species-specific DNA probes from Taenia solium and Taenia saginata and their use in an egg detection assay. J Clin Microbiol 1995, 33, 1283–1288. [Google Scholar] [CrossRef]
- González, L.M.; Montero, E.; Harrison, L.J.; Parkhouse, R.M.; Garate, T. Differential diagnosis of Taenia saginata and Taenia solium infection by PCR. J Clin Microbiol 2000, 38, 737–744. [Google Scholar] [CrossRef]
- Hamburger, J.; Turetski, T.; Kapeller, I.; Deresiewicz, R. Highly repeated short DNA sequences in the genome of Schistosoma mansoni recognized by a species-specific probe. Mol Biochem Parasitol 1991, 44, 73–80. [Google Scholar] [CrossRef]
- Mwangi, I.N.; Agola, E.L.; Mugambi, R.M.; Shiraho, E.A.; Mkoji, G.M. Development and Evaluation of a Loop-Mediated Isothermal Amplification Assay for Diagnosis of Schistosoma mansoni Infection in Faecal Samples. Journal of parasitology research 2018, 2018, 1267826. [Google Scholar] [CrossRef]
- Hamburger, J.; Xu, Y.X.; Ramzy, R.M.; Jourdane, J.; Ruppel, A. Development and laboratory evaluation of a polymerase chain reaction for monitoring Schistosoma mansoni infestation of water. Am J Trop Med Hyg 1998, 59, 468–473. [Google Scholar] [CrossRef]
- Hamburger, J.; He, N.; Abbasi, I.; Ramzy, R.M.; Jourdane, J.; Ruppel, A. Polymerase chain reaction assay based on a highly repeated sequence of Schistosoma haematobium: a potential tool for monitoring schistosome-infested water. Am J Trop Med Hyg 2001, 65, 907–911. [Google Scholar] [CrossRef]
- Hertel, J.; Hamburger, J.; Haberl, B.; Haas, W. Detection of bird schistosomes in lakes by PCR and filter-hybridization. Exp Parasitol 2002, 101, 57–63. [Google Scholar] [CrossRef]
- Lodh, N.; Caro, R.; Sofer, S.; Scott, A.; Krolewiecki, A.; Shiff, C. Diagnosis of Strongyloides stercoralis: Detection of parasite-derived DNA in urine. Acta tropica 2016, 163, 9–13. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, L.A.; DeSimone, S.M.; Williams, S.A. Cloning and comparison of repeated DNA sequences from the human filarial parasite Brugia malayi and the animal parasite Brugia pahangi. Proc Natl Acad Sci U S A 1986, 83, 797–801. [Google Scholar] [CrossRef]
- Albers, A.; Sartono, E.; Wahyuni, S.; Yazdanbakhsh, M.; Maizels, R.M.; Klarmann-Schulz, U.; Pfarr, K.; Hoerauf, A. Real-time PCR detection of the HhaI tandem DNA repeat in pre- and post-patent Brugia malayi Infections: a study in Indonesian transmigrants. Parasit Vectors 2014, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; McCarthy, J.; Bierwert, L.; Lizotte-Waniewski, M.; Chanteau, S.; Nutman, T.B.; Ottesen, E.A.; Williams, S.A. A polymerase chain reaction assay for detection of the parasite Wuchereria bancrofti in human blood samples. Am J Trop Med Hyg 1996, 54, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.U.; Atkinson, L.J.; Ramzy, R.M.; Helmy, H.; Farid, H.A.; Bockarie, M.J.; Susapu, M.; Laney, S.J.; Williams, S.A.; Weil, G.J. A real-time PCR-based assay for detection of Wuchereria bancrofti DNA in blood and mosquitoes. Am J Trop Med Hyg 2006, 74, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Saul, A.; Yeganeh, F.; Howard, R.J. Cloning and characterization of a novel multicopy, repetitive sequence of Plasmodium falciparum, REP51. Immunology and cell biology 1992, 70 ( Pt 5), 357-359. [CrossRef]
- Hotterbeekx, A.; Raimon, S.; Abd-Elfarag, G.; Carter, J.Y.; Sebit, W.; Suliman, A.; Siewe Fodjo, J.N.; De Witte, P.; Logora, M.Y.; Colebunders, R.; et al. Onchocerca volvulus is not detected in the cerebrospinal fluid of persons with onchocerciasis-associated epilepsy. Int J Infect Dis 2020, 91, 119–123. [Google Scholar] [CrossRef]
- Macfarlane, C.L.; Quek, S.; Pionnier, N.; Turner, J.D.; Wanji, S.; Wagstaff, S.C.; Taylor, M.J. The insufficiency of circulating miRNA and DNA as diagnostic tools or as biomarkers of treatment efficacy for Onchocerca volvulus. Scientific reports 2020, 10, 6672. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.M.; Fessler, M.K.; Bloomfield, R.A.; Sandke, W.D.; Malekshahi, C.R.; Keroack, C.D.; Duignan, P.J.; Torquato, S.D.; Williams, S.A. A novel quantitative real-time PCR diagnostic assay for fecal and nasal swab detection of an otariid lungworm, Parafilaroides decorus. International journal for parasitology. Parasites and wildlife 2020, 12, 85–92. [Google Scholar] [CrossRef]
- van Dongen, J.E.; Berendsen, J.T.W.; Steenbergen, R.D.M.; Wolthuis, R.M.F.; Eijkel, J.C.T.; Segerink, L.I. Point-of-care CRISPR/Cas nucleic acid detection: Recent advances, challenges and opportunities. Biosens Bioelectron 2020, 166, 112445. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. Journal of bacteriology 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Embden, J.D.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol Microbiol 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Mohanraju, P.; Makarova, K.S.; Zetsche, B.; Zhang, F.; Koonin, E.V.; van der Oost, J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science 2016, 353, aad5147. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Ma, E.B.; Harrington, L.B.; Da Costa, M.; Tian, X.R.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR-Cas systems. Nature reviews. Microbiology 2017, 15, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Mol Cell 2015, 60, 385–397. [Google Scholar] [CrossRef]
- Swarts, D.C.; Jinek, M. Mechanistic Insights into the cis- and trans-Acting DNase Activities of Cas12a. Mol Cell 2019, 73, 589–600 e584. [Google Scholar] [CrossRef]
- Strohkendl, I.; Saifuddin, F.A.; Rybarski, J.R.; Finkelstein, I.J.; Russell, R. Kinetic Basis for DNA Target Specificity of CRISPR-Cas12a. Mol Cell 2018, 71, 816–824 e813. [Google Scholar] [CrossRef]
- Wang, R.; Qian, C.; Pang, Y.; Li, M.; Yang, Y.; Ma, H.; Zhao, M.; Qian, F.; Yu, H.; Liu, Z.; et al. opvCRISPR: One-pot visual RT-LAMP-CRISPR platform for SARS-cov-2 detection. Biosens Bioelectron 2021, 172, 112766. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhong, M.; Liu, Y.; Ma, P.; Dang, L.; Meng, Q.; Wan, W.; Ma, X.; Liu, J.; Yang, G.; et al. Rapid and sensitive detection of COVID-19 using CRISPR/Cas12a-based detection with naked eye readout, CRISPR/Cas12a-NER. Sci Bull (Beijing) 2020, 65, 1436–1439. [Google Scholar] [CrossRef]
- Lee, R.A.; Puig, H.; Nguyen, P.Q.; Angenent-Mari, N.M.; Donghia, N.M.; McGee, J.P.; Dvorin, J.D.; Klapperich, C.M.; Pollock, N.R.; Collins, J.J. Ultrasensitive CRISPR-based diagnostic for field-applicable detection of Plasmodium species in symptomatic and asymptomatic malaria. Proc Natl Acad Sci U S A 2020, 117, 25722–25731. [Google Scholar] [CrossRef]
- WHO. Malaria rapid diagnostic test performance: summary results of WHO product testing of malaria RDTs: Round 1-8 (2008–2018). Available online: (accessed on 23 April).
- Lei, R.; Li, L.; Wu, P.; Fei, X.; Zhang, Y.; Wang, J.; Zhang, D.; Zhang, Q.; Yang, N.; Wang, X. RPA/CRISPR/Cas12a-Based On-Site and Rapid Nucleic Acid Detection of Toxoplasma gondii in the Environment. ACS Synth Biol 2022, 11, 1772–1781. [Google Scholar] [CrossRef]
- Galvani, A.T.; Christ, A.P.G.; Padula, J.A.; Barbosa, M.R.F.; de Araújo, R.S.; Sato, M.I.Z.; Razzolini, M.T.P. Real-time PCR detection of Toxoplasma gondii in surface water samples in São Paulo, Brazil. Parasitol Res 2019, 118, 631–640. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, K.; Wang, Y.; Li, D.; Cui, Z.; Huang, J.; Zhang, S.; Li, X.; Zhang, L. CRISPR/Cas12a-based on-site diagnostics of Cryptosporidium parvum IId-subtype-family from human and cattle fecal samples. Parasit Vectors 2021, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, F.; Hall, T.; Vesey, G.; Goldys, E.M. CRISPR/Cas12a-powered immunosensor suitable for ultra-sensitive whole Cryptosporidium oocyst detection from water samples using a plate reader. Water research 2021, 203, 117553. [Google Scholar] [CrossRef]
- Kanitchinda, S.; Srisala, J.; Suebsing, R.; Prachumwat, A.; Chaijarasphong, T. CRISPR-Cas fluorescent cleavage assay coupled with recombinase polymerase amplification for sensitive and specific detection of Enterocytozoon hepatopenaei. Biotechnology reports (Amsterdam, Netherlands) 2020, 27, e00485. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Peng, D.; Jiang, C.; Zhao, W.; Li, G.; Huang, W.; Kong, L.; Gao, H.; Zheng, J.; Peng, H. Rapid and Visual Detection of Heterodera schachtii Using Recombinase Polymerase Amplification Combined with Cas12a-Mediated Technology. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef]
- Fleming, K.A.; Horton, S.; Wilson, M.L.; Atun, R.; DeStigter, K.; Flanigan, J.; Sayed, S.; Adam, P.; Aguilar, B.; Andronikou, S.; et al. The Lancet Commission on diagnostics: transforming access to diagnostics. Lancet 2021, 398, 1997–2050. [Google Scholar] [CrossRef]
- Kosack, C.S.; Page, A.L.; Klatser, P.R. A guide to aid the selection of diagnostic tests. Bull World Health Organ 2017, 95, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Huyke, D.A.; Ramachandran, A.; Bashkirov, V.I.; Kotseroglou, E.K.; Kotseroglou, T.; Santiago, J.G. Enzyme Kinetics and Detector Sensitivity Determine Limits of Detection of Amplification-Free CRISPR-Cas12 and CRISPR-Cas13 Diagnostics. Anal Chem 2022, 94, 9826–9834. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Santiago, J.G. CRISPR Enzyme Kinetics for Molecular Diagnostics. Anal Chem 2021, 93, 7456–7464. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.E.; Dupuis, M.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Ren, K.; Qiu, X.; Zheng, J.; Guo, M.; Guan, X.; Liu, H.; Li, N.; Zhang, B.; Yang, D.; et al. The crystal structure of Cpf1 in complex with CRISPR RNA. Nature 2016, 532, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Yang, H.; Rajashankar, K.R.; Huang, Z.; Patel, D.J. Type V CRISPR-Cas Cpf1 endonuclease employs a unique mechanism for crRNA-mediated target DNA recognition. Cell research 2016, 26, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Nishimasu, H.; Zetsche, B.; Hirano, H.; Slaymaker, I.M.; Li, Y.; Fedorova, I.; Nakane, T.; Makarova, K.S.; Koonin, E.V.; et al. Crystal Structure of Cpf1 in Complex with Guide RNA and Target DNA. Cell 2016, 165, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Zetsche, B.; Ishitani, R.; Zhang, F.; Nishimasu, H.; Nureki, O. Structural Basis for the Canonical and Non-canonical PAM Recognition by CRISPR-Cpf1. Mol Cell 2017, 67, 633–645 e633. [Google Scholar] [CrossRef]
- Kim, H.K.; Song, M.; Lee, J.; Menon, A.V.; Jung, S.; Kang, Y.M.; Choi, J.W.; Woo, E.; Koh, H.C.; Nam, J.W.; et al. In vivo high-throughput profiling of CRISPR-Cpf1 activity. Nat Methods 2017, 14, 153–159. [Google Scholar] [CrossRef]
- Lu, S.; Tong, X.; Han, Y.; Zhang, K.; Zhang, Y.; Chen, Q.; Duan, J.; Lei, X.; Huang, M.; Qiu, Y.; et al. Fast and sensitive detection of SARS-CoV-2 RNA using suboptimal protospacer adjacent motifs for Cas12a. Nat Biomed Eng 2022, 6, 286–297. [Google Scholar] [CrossRef]
- Corsi, G.I.; Qu, K.; Alkan, F.; Pan, X.; Luo, Y.; Gorodkin, J. CRISPR/Cas9 gRNA activity depends on free energy changes and on the target PAM context. Nat Commun 2022, 13, 3006. [Google Scholar] [CrossRef]
- Zhuo, C.; Zhang, J.; Lee, J.-H.; Jiao, J.; Cheng, D.; Liu, L.; Kim, H.-W.; Tao, Y.; Li, M. Spatiotemporal control of CRISPR/Cas9 gene editing. Signal Transduction and Targeted Therapy 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Brown, W.; Bardhan, A.; Delaney, M.; Ilk, A.S.; Rauen, R.R.; Kahn, S.I.; Tsang, M.; Deiters, A. Spatiotemporal Control of CRISPR/Cas9 Function in Cells and Zebrafish using Light-Activated Guide RNA. Angewandte Chemie International Edition 2020, 59, 8998–9003. [Google Scholar] [CrossRef]
- Jain, P.K.; Ramanan, V.; Schepers, A.G.; Dalvie, N.S.; Panda, A.; Fleming, H.E.; Bhatia, S.N. Development of Light-Activated CRISPR Using Guide RNAs with Photocleavable Protectors. Angewandte Chemie (International ed. in English) 2016, 55, 12440–12444. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Qiu, Z.; Bi, Z.; Tian, T.; Jiang, Y.; Zhou, X. Photocontrolled crRNA activation enables robust CRISPR-Cas12a diagnostics. Proc Natl Acad Sci U S A 2022, 119, e2202034119. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Liu, R.; Qiu, Z.; Cao, F.; Tian, T.; Lu, Y.; Jiang, Y.; Zhou, X. Light-Start CRISPR-Cas12a Reaction with Caged crRNA Enables Rapid and Sensitive Nucleic Acid Detection. Angewandte Chemie International Edition 2023, 62. [Google Scholar] [CrossRef]
- Pang, B.; Xu, J.; Liu, Y.; Peng, H.; Feng, W.; Cao, Y.; Wu, J.; Xiao, H.; Pabbaraju, K.; Tipples, G.; et al. Isothermal Amplification and Ambient Visualization in a Single Tube for the Detection of SARS-CoV-2 Using Loop-Mediated Amplification and CRISPR Technology. Anal Chem 2020, 92, 16204–16212. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, Y.; Chen, Y.; Yang, Z.; Wu, H.; Zhou, Z.; Li, J.; Ping, J.; He, L.; Shen, H.; et al. Contamination-free visual detection of SARS-CoV-2 with CRISPR/Cas12a: A promising method in the point-of-care detection. Biosens Bioelectron 2020, 169, 112642. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Liu, Y.; Yang, M.; Zheng, J.; Liu, C.; Ye, W.; Song, S.; Bai, T.; Song, C.; Wang, M.; et al. The engineered CRISPR-Mb2Cas12a variant enables sensitive and fast nucleic acid-based pathogens diagnostics in the field. Plant Biotechnology Journal 2023, 21, 1465–1478. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, B.; Yang, L.; Zhao, C.; Wang, Y.; Tang, Y.; Yang, G.; Wang, P.; Gao, S. CRISPR/Cas12a combined with recombinase polymerase amplification for rapid and sensitive detection of <italic>Vibrio vulnificus</italic> in one tube. Acta Biochimica et Biophysica Sinica 2023, 55, 322–326. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Smith, B.M.; Jain, P.K. Enhancement of trans-cleavage activity of Cas12a with engineered crRNA enables amplified nucleic acid detection. Nat Commun 2020, 11, 4906. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhou, X. CRISPR-Based Biosensing Strategies: Technical Development and Application Prospects. Annual Review of Analytical Chemistry 2023, 16, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, W.; Ma, S.; Li, Z.; Yao, Y.; Fei, T. A chemical-enhanced system for CRISPR-Based nucleic acid detection. Biosens Bioelectron 2021, 192, 113493. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, M.; Merlo, R.; Bagheri, N.; Moscone, D.; Valenti, A.; Saha, A.; Arantes, P.R.; Ippodrino, R.; Ricci, F.; Treglia, I.; et al. Enhancement of CRISPR/Cas12a trans-cleavage activity using hairpin DNA reporters. Nucleic Acids Res 2022, 50, 8377–8391. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Hu, R.; Xia, J.; Xu, Z.; Chen, D.; Xi, J.; Liu, B.F.; Zhu, J.; Li, Y.; Yang, Y.; et al. G-triplex: A new type of CRISPR-Cas12a reporter enabling highly sensitive nucleic acid detection. Biosens Bioelectron 2021, 187, 113292. [Google Scholar] [CrossRef]
- Lobato, I.M.; O'Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Analyt Chem 2018, 98, 19–35. [Google Scholar] [CrossRef]
Figure 1.
Effect of amplification and cleavage on detection efficiency. E is the target-activated Cas12-crRNA-target DNA complex. During the initial stage of the one-pot procedure, cis-cleavage is immediate, and the enzyme concentration is much greater than the target concentration. As amplification becomes more dominant, such as when amplification efficiency is increased and cis-cleavage speed is reduced, the amount of [E] is substantially larger and can be quickly increased, resulting in a significant improvement in reaction efficiency. On the other hand, if the target is quickly depleted in the initial stage, the emission of the fluorescent signal will decrease.
Figure 1.
Effect of amplification and cleavage on detection efficiency. E is the target-activated Cas12-crRNA-target DNA complex. During the initial stage of the one-pot procedure, cis-cleavage is immediate, and the enzyme concentration is much greater than the target concentration. As amplification becomes more dominant, such as when amplification efficiency is increased and cis-cleavage speed is reduced, the amount of [E] is substantially larger and can be quickly increased, resulting in a significant improvement in reaction efficiency. On the other hand, if the target is quickly depleted in the initial stage, the emission of the fluorescent signal will decrease.
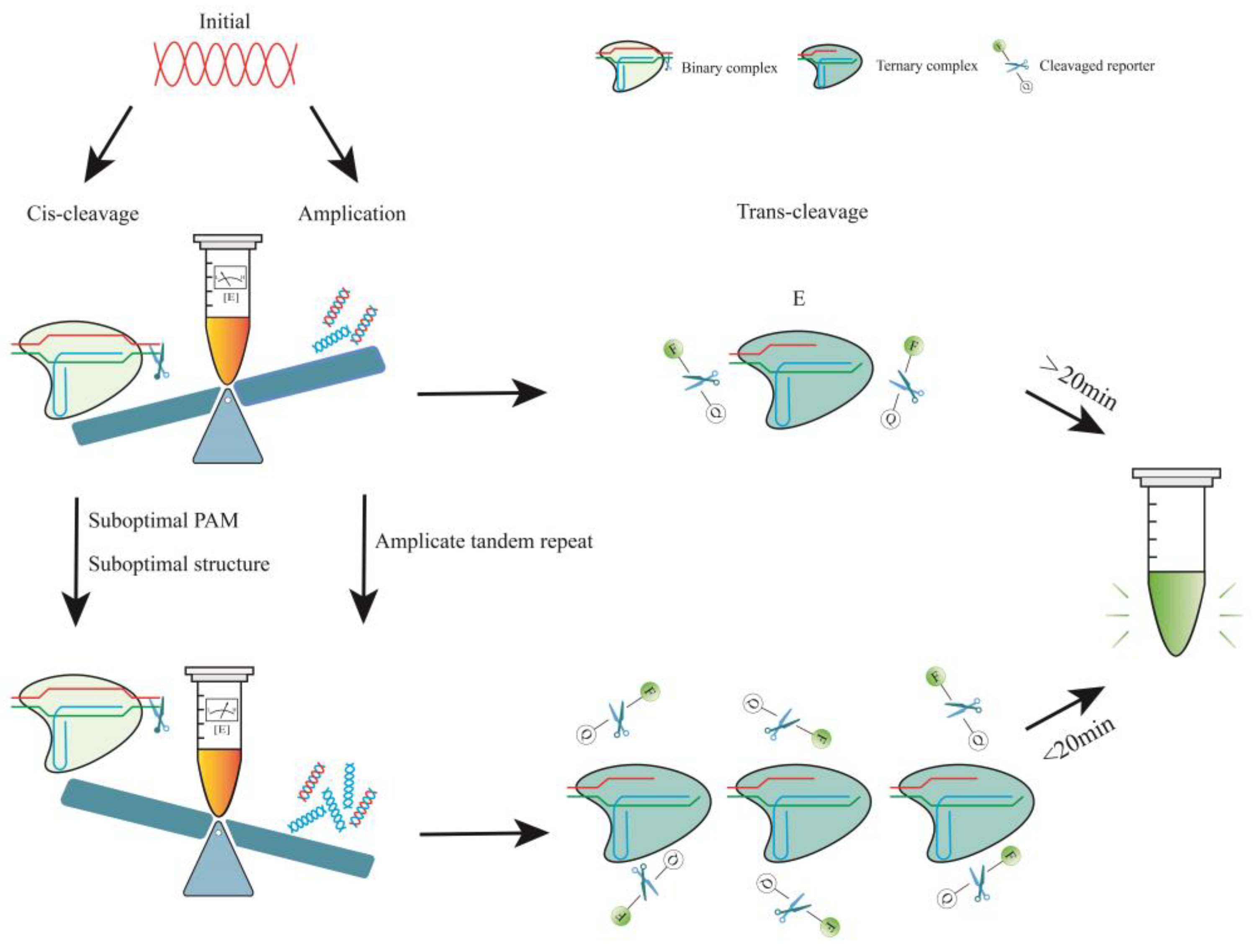
Figure 2.
A schematic diagram of the relationship between amplification and cleavage in the one-pot method. a. Comparison of amplification and cleavage efficiency over time when using suboptimal protospacer or suboptimal crRNA to attenuate the early cleavage efficiency of Cas12a in the one-pot method. b. Comparison of amplification and cleavage efficiency over time using optimal crRNA in the one-pot method. c. Variation in substrate concentrations over time when using suboptimal and optimal crRNA in the one-pot method.
Figure 2.
A schematic diagram of the relationship between amplification and cleavage in the one-pot method. a. Comparison of amplification and cleavage efficiency over time when using suboptimal protospacer or suboptimal crRNA to attenuate the early cleavage efficiency of Cas12a in the one-pot method. b. Comparison of amplification and cleavage efficiency over time using optimal crRNA in the one-pot method. c. Variation in substrate concentrations over time when using suboptimal and optimal crRNA in the one-pot method.
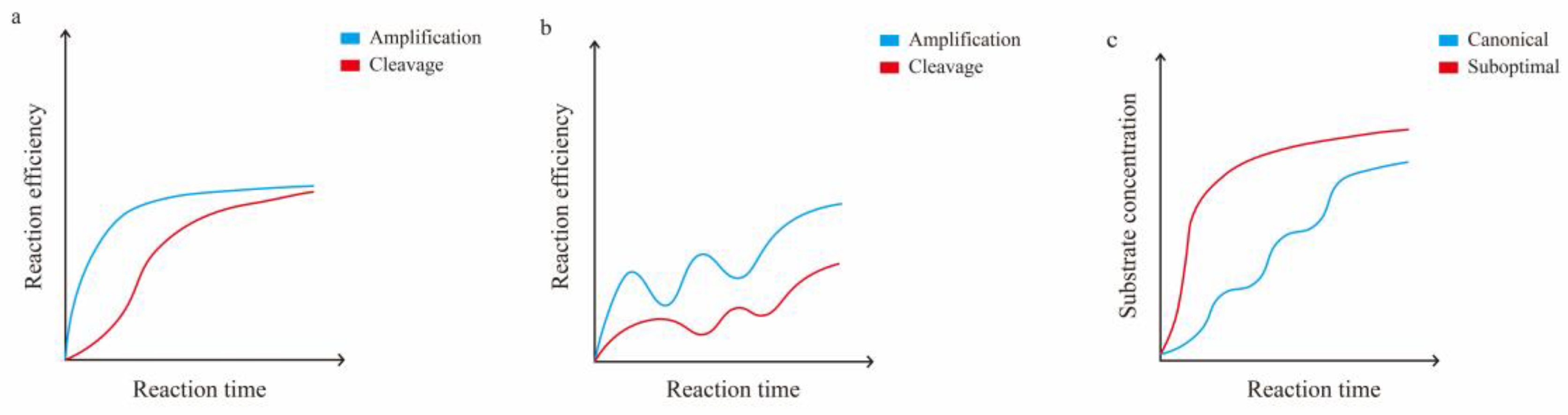

Table 1.
Main strengths and weaknesses of different approaches for parasite detection.
| Discipline | Strength | Weakness |
| Morphology | ·Accuracy (gold standard) ·Can detect multiple species at the same time |
·Lower sensitivity ·Difficulty distinguishing parasite-like egg ·High demand for professional skills |
| Immunology | ·Strong specificity ·Strong sensitivity |
·High cost and time consuming ·False positives for cross-reactivity ·False negatives in immunocompromised patients ·Inability to differentiate between ongoing and past infections |
| Molecular biology | ·Strong specificity ·Strong sensitivity ·Strong repeatability |
·High cost ·Limitations related to sample preparation and equipment ·Logistics systems requiring fresh sample analysis (e.g. cryogenic) |
Table 2.
List of partial parasitic repeat sequences.
| Parasite | Repeat Sequence Name | Length(bp) | Quantity | GenBank Accession | Refs |
|---|---|---|---|---|---|
| Protozoa | |||||
| Trypanosoma cruzi | TCNRE | 195 | 12% of the total genome | K01772 | [20] |
| Toxoplasma gondii | / | 529 | 200-300 copies per genome | AF146527 | [21] |
| Plasmodium falciparum | Pfr364 | 716 | 41 copies per genome | / | [22,23] |
| Plasmodium vivax | Pvr47 | 333 | 14 copies per genome | / | [22,23,24] |
| Cestodes | |||||
| Echinococcus granulosus | EgG1 Hae III repeat | 269 | 6900 copies per haploid genome (1% of E. granulosus genomic DNA) |
DQ157697 | [25,26] |
| Taenia solium | Tsol-9 | 158 | None Related Description | U45987 | [27] |
| Taenia saginata | HDP1 | 1272 | 0.4% of the T. saginata DNA | AJ133764 | [28] |
| Trematodes | |||||
| Schistosoma mansoni | Sml-7(DraI) | 121 | 12% of the total genome | M61098 | [29,30,31] |
| Schistosoma haematobium | DraI | 121 | over 15% of the S. haematobium genome | DQ157698 | [32] |
| Trichobilharzia ocellata | ToSau3A | 396 | 10,000 copies per haploid genome (1.5% of the T. ocellata genome) |
AF442689 | [33] |
| Nematodes | |||||
| Strongyloides stercoralis | / | 765 | None Related Description | AY028262 | [34] |
| Brugia malayi | HhaI repeat | 320 | several thousand copies per haploid genome (about 12% of the genome) |
M12691 | [35,36] |
| Wuchereria bancrofti | SspI | 195 | 300 copies per haploid genome | L20344 | [37] |
| LDR | 1674 | None Related Description | AY297458 | [38] | |
| Onchocerca volvulus | O-150 | 149 | 4500 copies per haploid genome | J04659 | [39,40,41] |
| Parafilaroides decorus | Pd65 | 689 | None Related Description | MT053285 | [42] |
Specific information on tandem repeats that have been used for parasite detection, including GenBank accession numbers and references, is provided.
Table 3.
Comparison of CRISPR-Cas12a and commonly used detection technologies in molecular biology.
| Technology | Device dependency | Specificity | Reaction time (min) |
Number of primers | Quantification | Cost | Results View Method | POCT potential |
| PCR | Moderate | Strong | 60-180 | 2 | No | High | Gel electrophoresis | Moderate |
| q-PCR (qRT-PCR) |
High | Strong | >60 | 2 | Yes | Extremely high | Fluorescent and computer system | LOW |
| d-PCR | High | Strong | >60 | 2 | Yes | Extremely high | Fluorescent and computer system | LOW |
| LAMP | Low | Strong | <60 | 4-6 | No | Low | ·Gel electrophoresis ·Color ·Turbidity |
High |
| RPA | Low | Moderate | 20-60 | 2 | No | Low | ·Gel electrophoresis ·Fluorescent ·Lateral flow |
High |
| Cas12a | Low | Strong | 20-60 | 2 | No | Low | ·fluorescent ·Lateral flow |
High |
Table 4.
Application of the CRISPR/CAS12 system to parasite detection.
| Species | Method | Time (min) | LOD | Specificity | Refs |
|---|---|---|---|---|---|
| Plasmodium falciparum | Cas12a-RPA | 30 (+10) a | 0.36 parasites/μL | 100% | [56] |
| Plasmodium vivax | Cas12a-RPA | 30 (+10) a | 1.2 parasites/μL | 100% | |
| Toxoplasma gondii | Cas12a-RPA | 35 (+20) a | 99~115 copies/μL b | 100% | [58] |
| Cryptosporidium parvum | Cas12a-RPA (two steps) | 30 + 60 (+20) a | 10 oocysts | 100% | [60] |
| Cas12a-RPA | 90 | 1 oocyst | 100% | [61] | |
| Enterocytozoon hepatopenaei | Cas12a-RPA | 60 | 50 copies/μL b | 100% | [62] |
| Heterodera schachtii | Cas12a-RPA | 60 | 10-4 single cysts | 100% | [63] |
a, The time in parentheses is the time required for sample preparation or DNA extraction. b, Copy numbers indicate the copies of target genes rather than the genomes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



