
Submitted:
23 December 2023
Posted:
25 December 2023
You are already at the latest version
Abstract
Stalk rot caused by Fusarium fungi is one of the most widespread and devastating diseases of maize, and the introduction of resistant genotypes is one of the most effective strategies for controlling the disease. Breeding genotypes with genetically determined resistance will also allow less use of crop protection products. The aim of the research was to identify molecular markers and associated candidate genes determining maize plant resistance to Fusarium stalk rot. The plant material for the study consisted of 122 maize hybrids, which were sown in plots of 10 m2, in a complete randomized block design, in three repetitions, at two locations (Smolice: 51 42’ 58.904’’ N, 17 13’ 29.13’’ E and Kobierzyce: 50 58’ 19.411’’ N, 16 55’ 47.323’’ E). The analyzed genotypes were simultaneously subjected to next-generation sequencing using the Illumina platform. The results of the observation of the degree of infection and sequencing were used for association mapping, which ultimately resulted in the selection of 10 molecular markers important at both places. Among the identified markers, two SNP markers that are located inside candidate genes play an important role. Marker 4772836 is located inside the serine/threonine-protein kinase bsk3 gene, while marker 4765764 is located inside the histidine kinase 1 gene. Both genes can be associated with plant resistance to Fusarium stalk rot, and these genes can also be used in breeding programs to select resistant varieties.
Keywords:
maize
; next generation sequencing
; SNP markers
; SilicoDArT markers
; stalk rot
1. Introduction
In the last decade, the increase in temperature, expansion of areas and intensification of cultivation, the introduction of agrotechnical simplifications and the appearance of new pest species have resulted in a significant increase in the threat to the amount and quality of maize yields [1]. Crops of this cereal are frequently infected by pathogenic fungi, resulting in serious yield losses [2]. Currently, fungi of the genus Fusarium can be classified as the most dangerous pathogens causing serious losses in maize plantations. The fungus of the genus Fusarium graminearum causes stalk rot in maize [3,4]. This pathogen not only causes yields losses, but also poses a huge threat related to the presence of mycotoxins [5,6,7]. Fungi of the genus Fusarium colonize soil, crop residues of maize and cereals, and are also present on the surface of grains. They reach plants directly from resting places and through wind, water, soil fragments (dust), insects and animals. Maize is exposed to Fusarium fungi from the moment the grain is sown [8]. As a result of infection, maize plants often lodge, leading to a decline in planting density.
Maize plant lodging caused by Fusarium fungus infection is often observed in fields where seed is sown untreated with a fungicide. The development of the pathogen can also be favored by weather factors, such as a cool and rainy spring, heavy rainfall or sowing two deeply on heavy soil [9]. Infection can, of course, be prevented, and several studies have shown that soil fertilization had a positive effect on plant tolerance to pathogenic fungi, such as the use of fertilizers containing zinc or potassium phosphite [10,11,12]. Stalk rot usually occurs when plant tissues are damaged by Ostrinia nubilalis. The feeding of this pest is largely responsible for the development of the disease, especially since Ostrinia nubilalis is also a vector of this fungus and, migrating between plants, transfers fungal spores from diseased plants to healthy ones [13]. There are usually two generations of Ostrinia nubilalis larvae per year: the first generation attacks plants in the middle or end of the vegetative phase, and the second generation attacks plants in the reproductive phase (from early milk stage to maturity) [14]. Strong decomposition of stem tissues, including nodes, leads to whole plants falling over, making harvesting difficult and sometimes impossible.
Fighting fusarium diseases that appear at the end of the growing season is difficult, especially when no treatments have applied beforehand to directly or indirectly limit infection [15]. Currently, the most effective method of combating this pathogen is the cultivation of varieties with genetically determined resistance to Fusarium graminearum. A dynamically changing climate and an increase in demand for maize, driven by natural increase, are catalysts for research into genes and genomic regions important to agronomy [16,17]. Scientists have long used next-generation sequencing (NGS) and association mapping methods to identify markers associated with resistance genes. Genome-wide association studies (GWAS) are a useful tool for identifying candidate genes, especially when combined with QTL mapping to validate loci for quantitative traits. Zila et al. [18,19] performed GWAS tests in maize to detect SNPs associated with increased resistance to fusarium. They identified 10 SNP markers significantly associated with resistance to this pathogen. The introduction of molecular analyzes made it possible to develop a methodology for genetic marker-assisted selection (MAS). The scope of application of this methodology depends, of course, on the progress of knowledge of the genome of the species [20,21]. The completion of the sequencing of the human genome has opened up a wide range of possibilities for understanding the genomic sequences of crop plants [22,23,24,25]. For several years, maize breeding around the world has been supported by useful molecular markers. Many authors state in their publications that breeding supported by molecular marker accelerates yield increases not only in the United States, but also in other countries, offering great potential to increase the productivity and value of maize germplasm [26,27].
Therefore, the aim of this study was to identify new SNP and SilicoDArT markers linked to candidate genes for maize stalk rot resistance.
2. Materials and Methods
2.1. Plant material
The plant material used in the study comes from Smolice Plant Breeding Ltd. IHAR Group (51°42′12″N 17°10′10″E) and Małopolska Plant Breeding Ltd. (50°58′17″N 16°55′50″E). The plant material includes 122 maize hybrids. Some of the lines analyzed were characterized by lines with flint grain of three different origins: F2 (a group related to the F2 line bred at INRA in France from the Lacaune population), EP1 (a group related to the EP1 line, bred in Spain from a population originating in the Pyrenees) and German Flint. The second part of the plant material consisted of dent-type kernels; these were derived from various groups of origin from the United States: Iowa Stiff Stalk Synthetic (BSSS), Iowa Dent (ID) and Lancaster.
2.2. Methods
2.2.1. Field experiment
The experiment was established in plots of 10 m2, in three repetitions, in a randomized complete blocks design, in two localities (Smolice: 51° 42’ 58.904’’ N, 17° 13’ 29.13’’ E and Kobierzyce: 50° 58’ 19.411’’ N, 16° 55’ 47.323’’ E). Health evaluation was carried out on ten plants from each plot by calculating the percentage of stalks infected by Fusarium stalk rot.
2.2.2. Weather conditions
In 2022, the average rainfall in Smolice was 34.45 mm, which was 13.82 mm lower than the multi-year average. The highest rainfall was recorded in July (55 mm) and the lowest in March (15 mm). The average air temperature this year in Smolice was 9.54°C, higher than the long-term average temperature by 0.8°C. The warmest month in 2004 was August (20°C), while the lowest temperature was recorded in December (1.1°C). In 2022, the amount of rainfall and temperature were unfavorable during the initial development of maize plants. Despite the early sowing date, maize remained at the 2–3 leaf phase for a long time, on which purple discoloration was visible due to difficulties in taking up phosphorus from the soil. May had an abundance of rainfall, which had a positive effect on maize development. In Kobierzyce, the average rainfall in 2022 was 51.52 mm, 3.22 mm higher than the average multi-year rainfall. The highest rainfall was recorded in May, with 95.8 mm, and the lowest in March, with 21.6 mm. The average air temperature in Kobierzyce was 11.46°C, which was higher than the multi-year average air temperature by 2.58°C. Among the warmest months in 2022 in Kobierzyce, as in Smolice, was August (22°C), and the coldest was December (1.5°C).
2.2.3. DNA isolation
DNA isolation from 122 maize hybrids was carried out using a ready-made set of reagents from Symbios. The isolated DNA samples were subjected to next-generation sequencing. The concentration and purity of the isolated DNA samples were determined using a DS-11 spectrophotometer from DeNovix. The isolated DNA matrix was adjusted to a uniform concentration of 100 ng μl–1 by dilution with deionized distilled water.
2.2.4. Genotyping
DArTseq technology, based on next-generation sequencing, was used for genotyping. Isolated DNA from the 122 maize hybrids tested at 50 µl at a concentration of 100 ng was transferred on two 96-well Eppendorf plates for analyzes to identify SilicoDArT and SNP polymorphisms. The analyzes were performed at Diversity Arrays Technology, University of Canberra, Australia. The methods used are described in detail on the Diversity Arrays Technology website: (https://www.diversityarrays.com/technology-and-resources/dartseq/).
2.2.5. Association mapping using GWAS analysis
Based on the results obtained from genotyping and phenotyping, association mapping was performed using GWAS analyses. Associations were made for 12 analyzed maize hybrids. Genotypic data were obtained from DArTseq analysis, while phenotypic data are results from observations of the degree of stem infection and lodging of maize. Based on GWAS analysis, SilicoDArT and SNP markers with the highest level of significance, i.e. those that were most strongly associated with plant resistance to Fusarium stalk rot, were selected for further research. For the association analysis, only SilicoDArT and SNP sequences meeting the following criteria were selected: one SilicoDArT and/or SNP within a given sequence (69 nt), minor allele frequency (MAF) > 0.25 and the missing observation fractions <10%.
2.2.6. Statistical Analysis and Association Mapping
Data were analyzed using analysis of variance in a model with fixed effects of location and random effects of genotypes and genotype × location interaction. Association mapping, based on SilicoDArT and SNP data and average trait values obtained in each location. All analyses and visualizations of the results were performed in GenStat 23.1 [28]. Significance of association between Fusarium stalk rot and SilicoDArT and SNP markers was assessed using P values corrected for multiple testing using the Benjamini-Hochberg method.
2.2.7. Physical mapping
SilicoDArT and SNP marker sequences selected based on GWAS analysis were subjected to Basic Local Alignment Search Tool (BLAST) analysis, which involves searching databases to find sequences with high homology to the selected silicoDArT and SNP marker sequences. Publicly available web browsers were used for this purpose: CEPH Genotype database http://www.cephb.fr/en/cephdb/ NCBI Map Viewer http://www.ncbi.nlm.nih.gov/projects/mapview/ UCSC Genome Browser http://genome.ucsc.edu/ Ensembl Map View http://ensembl.fugu-sg.org/common/helpview?kw=mapview;ref. The programs used were applied to indicate the chromosomal locations of the searched sequences for, similar to the sequences analyzed, and to determine their physical location. The sequences of all genes located in the designated area on the chromosome were further analyzed.
2.2.8. Functional analysis of gene sequences
Functional analysis was carried out using Blast2GO software. The sequences of all genes located in the chromosome regions determined by BLAST analysis were analyzed. The goal was to obtain information on the biological function of gene sequences located in the designated chromosome region.
2.2.9. Design of primers for identified SilicoDArT and SNP polymorphisms related to yield and its characteristics
Primer 3 Plus was used to design the primers. The program is supported on the website and require no download or installation. Primer 3 Plus offers various options. These range from different ways to specify the sequence for which the primers are to be designed, through our general expectations of the primers (size, melting point – for both primers and products, %GC, complementarity, etc.), to very precise parameter settings for the primers.
3. Results
3.1. Phenotyping
The observed Fusarium stalk rot was characterized by a normal distribution about both localities, which made it possible to carry out association mapping. Analysis of variance indicated that the main effects of genotype and location, as well as genotype × location interaction were significant for Fusarium stalk rot (Table 1). The differences between the locations were large (11.39% in Kobierzyce and 1.20% in Smolice), which was the result of large differences in weather conditions between the two locations (Figure 1).
The Fusarium stalk rot values ranged from 1.65% (for genotype G01.10) to 31.18% (for genotype G03.07) in Kobierzyce and from 0.00% (for 58 genotypes) to 6.36% (G05.03) in Smolice. On average for both localities, the Fusarium stalk rot values ranged from 0.82% (for genotype G01.10) to 17.08% (for genotype G03.07) (Table 2). The variability of the observed values (expressed in standard deviations) also differed between locations: from 0% to 16.65% in Kobierzyce and from 0% to 9.45% in Smolice (Table 2).
Despite very large differences between Fusarium stalk rot values, their correlation was observed between Kobierzyce and Smolice (r=0.36, p<0.001).
3.2. Genotyping
Illumina sequencing identified 60,436 SilicoDArT markers and 32,178 SNP markers (92,614 in total). For association mapping, 32,900 markers (26,234 SilicoDArT and 6,666 SNPs) metting the assumed assumptions (MAF > 0.25 and the number of missing observations <10%) were used. Based on the identified SilicoDArT and SNP molecular markers, a dendrogram of genetic similarity between the 122 hybrids analyzed was prepared. The dendrogram distinguished three groups of similarity. Group I contained 53 hybrids, Group II 50 hybrids, while 19 hybrids were classified in Group III (Figure 2).
The association mapping identified 11,282 markers (8,405 SilicoDArT and 2,877 SNPs) in Smolice (Table 3, Figure 3) and 9,439 markers (7,148 SilicoDArT and 2,291 SNPs) in Kobierzyce (Table 4, Figure 4) significant at 0.05 level. The percentage of variation explained by each marker ranged from 2.4% to 34.4% in Smolice and from 2.4% to 27.0% in Kobierzyce (Table 3 and Table 4). 8,702 markers were statistically significant simultaneously in both localities. 91 of them were highly statistically significant – LOD>7.5 (Table 5). The percentage of variation explained by highly significant markers in both localities ranged from 22.0% (for four SilicoDArT markers: 9682713, 2432042, 2384252 and 4766257) to 30.5% (for SNP marker 4772836|F|0-17:C>A-17:C>A) in Smolice and from 22.0% (for four markers: two SilicoDArTs – 4766257 and 73750965 as well as two SNPs – 4586493|F|0-10:T>C-10:T>C and 25947704|F|0-38:C>T-38:C>T) to 27.0% (for SNP marker 4771464|F|0-65:C>T-65:C>T) in Kobierzyce (Table 5).
Of the 8,702 statistically significant markers in both localities (Smolice and Kobierzyce), ten significantly associated with plant resistance to Fusarium stalk rot at both Kobierzyce and Smolice locations (the most significant with the largest coefficients of determination) were selected for physical mapping (Table 6). Using the BLAST database, the location of the selected markers was determined and the associated candidate genes were provided. The next step was to design primers that would be used to identify the 10 selected markers. The sequences of the primer are shown in Table 7. Among the identified markers, two SNP markers that are located inside candidate genes play an important role. Marker 4772836 is located inside the serine/threonine-protein kinase bsk3 gene, while marker 4765764 is located inside the histidine kinase 1 gene. Both genes may be associated with plant resistance to Fusarium stalk rot.
4. Discussion
Maize is currently the leading crop in the global grain market, next to rice and wheat [29]. Its largest producers are the United States and China, with 2018 production of 392 million tons and 257 million tons, respectively [30]. In terms of yield growth rate in production, maize dominates among cereal plants, with yield potential reaches 12–15 tons of dry matter per 1 ha. The high fertility of maize is the result of many favorable physiological and morphological traits of this species [31]. However, it should be remembered that yield levels are affected by weather conditions during a given growing season and local environmental conditions [32]. Maize has successfully acclimatized to Polish soil and climatic conditions [33,34]. The reason for this phenomenon is the introduction of linear hybrids into maize cultivation, providing access to varieties with an appropriate growing season. Maize, as a crop, is of great importance in the world, both in terms of utility and economy. This is due to the possibility of using virtually all of the plant's biomass as feed, food or industrial raw material [30].
Plants, including maize, are constantly exposed to many stress factors, both abiotic and biotic, that can affect their growth and development [32]. Biotic stress factors include viruses, bacteria, fungi and insect pests. Often the consequence of stress factors is the occurrence of pathogenic diseases, which can interfere with normal plant growth, development and function, and cause significant yield reduction [35]. Maize diseases occurring worldwide include: common smut of maize, maize small leaf spot, maize sheath blight, common rust of maize, maize helminthosporium, and maize stalk rot [36].
Fusarium stalk rot of maize is caused by fungi of the genus Fusarium. Damage caused by European maize stalk rot promotes the development of the pathogenic fungus [37]. Characteristic traits of the disease are wilting and drying of leaf blades, weakening of the stem and often lodging [38]. In addition, Fusarium fungi increase the content of mycotoxins, which in turn disqualify grain for further processing [39,40,41].
In own research the field experiment made it possible to perform and analyze measurements of the degree of fusarium infection of maize stalks. The observed trait was characterized by a normal distribution at both localities, which made it possible to perform association mapping. Analysis of variance indicated that the main effects of genotype and location as well as genotype and location interaction were significant for Fusarium stalk rot. The differences between the locations were large (11.39% in Kobierzyce and 1.20% in Smolice), duo to large differences in weather conditions between the two locations.
The most important mycotoxins include aflatoxins, fumonisins, ochratoxin A, deoxynivalenol, zearalenone and ergot alkaloids, which are mainly produced by the genera Aspergillus, Penicillium, Fusarium and Claviceps [6]. Selection of resistant genotypes based on their phenotype can be unreliable and is time- and labor-consuming. Phenotyping maize for fusarium resistance requires field trials conducted in several environments due to the high interaction of genotype and environment [42]. In addition, proper management of inoculation timing is required due to the diversity of genotypes in terms of flowering [43]. Currently, the most effective and fastest methods to support selection are transcriptomic and genomic association studies (GWAS). They are useful tools for identifying candidate genes, especially when combined with QTL mapping to map and validate loci for quantitative traits [44]. Combining these methods has allowed us to overcome their limitations [45,46].
Our study identified 60,436 SilicoDArT markers and 32,178 SNP markers (92,614 in total). For association mapping, 32,900 markers (26,234 DArTs and 6,666 SNPs) were selected, meeting assumptions about the distribution of values (MAF > 0.25) and the number of missing observations (<10%). The association mapping identified 11,282 markers (8,405 silicoDArT and 2,877 SNPs) in the Smolice experiment and 9,439 markers (7,148 silicoDArT and 2,291 SNPs) in the Kobierzyce experiment. Of the 8,702 markers for physical mapping, ten were selected that were statistically significant in both localities (Smolice and Kobierzyce).
In recent years, many authors have attempted to identify molecular markers associated with functionally important traits in maize. Using next-generation sequencing, Sobiech et al. [47] identified markers associated with Fusarium resistance in maize plants. Bocianowski et al. [48] used NGS technology and association mapping to identify markers associated with the heterosis effect in maize. A Similar study was conducted by Tomkowiak et al. [49], who identified six SNP markers (1818, 14506, 2317, 3233, 11657 and 12812) located within genes, on chromosomes 8, 9, 7, 3, 5 and 1, respectively, associated with maize yield. Other authors [50] identified four genes: the isoform × 2 gene of sucrose synthase 4, the isoform × 1 gene of phosphoinositide phosphatase, the putative isoform × 1 gene of the SET domain-containing protein family, and the grx_c8–glutaredoxin subgroup iii gene, which significantly regulate the level of vigor and germination of maize seeds.
In the study presented here, of the 10 markers selected, two of them are located inside genes. SNP marker 4772836 located on chromosome 2, is located inside the serine/threonine-protein kinase bsk3 gene, while SNP marker 4765764, located on chromosome 5, is inside the histidine kinase 1 gene. Histidine kinase are typically trans-membrane proteins of the transferase class that play a role in signal transduction across the cellular membrane. The vast majority of histidine kinases (HKs) are homodimers that exhibit autokinase, phosphotransfer and phosphatase activity [51]. Unlike other classes of protein kinases, HKs are usually parts of a two-component signal transduction mechanisms, in which the HK transfers a phosphate group from ATP to a histidine residue inside the kinase and then to an aspartate residue in the receiver domain of the response regulator protein (or sometimes on the kinase itself). Histidine kinases have been implicated in cytokinin and ethylene signaling, in regulation responses to environmental stress [52]. These, it can be speculated that they also involved in the immune response associated with fusarium infection of plants.
The second gene analyzed, a probable serine/threonine kinase, acts as a positive regulator of brassinosteroid (BR) signaling. Brassinosteroids, another class of growth-promoting regulator, regulate many aspects of plant growth and development, including cell expansion and elongation (in combination with auxin) and vascular differentiation. The necessity of brassinosteroids in plant growth and development has been demonstrated by the identification of many BR-deficient dwarf mutants and subsequent BRs treating experiments [53]. Brassinosteroid-signaling kinases (BSKs) are critical in BRs signal transduction. BSK3 is the only BSK member involved in BR-mediated plant root growth [54].
To confirm the involvement of the two candidate genes in the immune response to infection of maize plants by Fusarium fungi, their expression will be analyzed. If the involvement of these genes in the immune reaction is confirmed, SNP markers associated with them can be used to select resistant varieties.
5. Conclusions
In maize cultivation, it is crucial to reduce the severity of diseases that cause a drastic decrease in yields. Excessive use of chemical protection negatively affects the natural environment, so the best method of crop protection is to use varieties with genetically determined resistance in breeding. Understanding the genetic basis and molecular mechanisms controlling Fusarium resistance is a key requirement for developing maize varieties with enhanced resistance. Because resistance to Fusarium fungi is a quantitative trait based on a distributed architecture of multiple genes with small effectors, the best approach for future molecular breeding is MAS. Using next-generation sequencing (NGS), two significant markers were identified in candidate genes associated with Fusarium resistance in maize plants. SNP marker 4772836, located on chromosome 2, is inside the serine/threonine-protein kinase bsk3 gene, while SNP marker 4765764, located on chromosome 5, is inside the histidine kinase 1 gene. If the involvement of these genes in the immune response is confirmed, SNP markers associated with them can be used to select resistant varieties.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The author declares no conflicts of interest.
References
- Zijlstra, C.; Lund, I.; Justesen, A.; Nicolaisen, M.; Bianciotto, V.; Posta, K.; Balestrini, R.; Przetakiewicz, A.; Czembor, E.; van de Zande, J. Combining novel monitoring tools and precision application technologies for integrated high-tech crop protection in the future (a discussion document). Pest Manag. Sci. 2011, 67, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Galiano-Carneiro, A.; Miedaner, T. Genetics of resistance and pathogenicity in the maize/Setosphaeria turcica pathosystem and implications for breeding. Front. Plant Sci. 2017, 8, 1490. [Google Scholar] [CrossRef]
- Vigier, B.; Reid, L.M.; Dwyer, L.M.; Stewart, D.W.; Sinha, R.C.; Arnason, J.T.; Butler, G. Maize resistance to gibberella ear rot: symptoms, deoxynivalenol, and yield. Can. J. Plant Pathol. 2001, 23, 99–105. [Google Scholar] [CrossRef]
- Sun, Y.; Ruan, X.; Wang, Q.; Zhou, Y.; Wang, F.; Ma, L.; Wang, Z.; Gao, X. Integrated gene co-expression analysis and metabolites profiling highlight the important role of ZmHIR3 in maize resistance to Gibberella stalk rot. Front. Plant Sci. 2021, 12, 664–733. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ceron, D.; Lowe, R.G.T.; McKenna, J.A.; Brain, L.M.; Dawson, C.S.; Clark, B.; Berkowitz, O.P.; Whelan, J.; Bleackley, M.R.; Anderson, M.A. Extracellular vesicles from Fusarium graminearum contain protein effectors expressed during infection of corn. J. Fungi 2021, 7, 977. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, R.A.; Jebur, A.; Kang, W.; El-Demerdash, F. An overview on the major mycotoxins in food products: characteristics, toxicity, and analysis. J. Future Foods 2022, 2, 91–102. [Google Scholar] [CrossRef]
- Hoppe, K.; Chełkowski, J.; Błaszczyk, L.; Bocianowski, J. Observation of changes in Fusarium mycotoxin profiles in maize grain over the last decade in Poland. Food Control 2024, 158, 110248. [Google Scholar] [CrossRef]
- Deepthi, B.V.; Deepa, N.; Vanitha, P.R.; Sreenivasa, M.Y. Stress responses on the growth and mycotoxin biosynthesis of Fusarium proliferatum associated with stored poultry feeds. Appl. Food Res. 2022, 2, 100091. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Europ. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Alsamir, M.; Al-Samir, E.; Kareem, T.A.; Abass, M.; Trethowan, R. The application of zinc fertilizer reduces Fusarium infection and development in wheat. Aust. J. Crop Sci. 2020, 14, 1088–1094. [Google Scholar] [CrossRef]
- Nowembabaz, A.; Taulya, G.; Tinzaara, W.; Karamura, E. Effect of integrated potassium nutrition on Fusarium wilt tolerance in apple bananas. Afr. J. Plant Sci. 2021, 15, 257–265. [Google Scholar] [CrossRef]
- Gilardi, G.; Vasileiadou, A.; Garibaldi, A.; Gullino, M.L. The effects of biological control agents, potassium phosphite and calcium oxide on race 1 of Fusarium oxysporum f. sp. lactucae of lettuce in closed soilless cultivation systems. J. Phytopathol. 2022, 170, 626–634. [Google Scholar] [CrossRef]
- Szóke, C.; Zsubori, Z.; Pók, I.; Rácz, F.; Illés, O.; Szegedi, I. Significance of the European corn borer (Ostrinia nubilalis Hóbn.) in maize production. Acta Agron. Hungarica 2002, 50, 447–461. [Google Scholar] [CrossRef]
- Blandino, M.; Reyneri, A.; Vanara, F.; Pascale, M.; Haidukowski, M.; Campagna, C. Management of fumonisin contamination in maize kernels through the timing of insecticide application against European corn borer Ostrinia nubilalis Hübner. Food Addit. Contam. A 2009, 26, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Unterseer, S.; Bauer, E.; Haberer, G.; Seidel, M.; Knaak, C.; Ouzunova, M.; Meitinger, T.; Strom, T.M.; Fries, R.; Pausch, H.; Bertani, C.; Davassi, A.; Mayer, K.F.; Schön, C.C. A powerful tool for genome analysis in maize: development and evaluation of the high density 600 k SNP genotyping array. BMC Genomics 2014, 15, 823. [Google Scholar] [CrossRef] [PubMed]
- Shah, T.R.; Prasad, K.; Kumar, P. Maize – A potential source of human nutrition and health. Cogent Food Agriculture 2016, 2, 1. [Google Scholar] [CrossRef]
- Zila, C.T.; Ogut, F.; Romay, M.C.; Gardner, C.A.; Buckler, E.S. Genome wide association study of Fusarium ear rot disease in the U.S.A. maize inbred line collection. BMC Plant Biol. 2014, 14, 372. [Google Scholar] [CrossRef] [PubMed]
- Zila, C.T.; Samayoa, L.F.; Santiago, R.; Butrón, A.; Holland, J.B. A genome-wide association study reveals genes associated with fusarium ear rot resistance in a maize core diversity panel. G3-Genes Genom. Genetic 2013, 3, 2095–2104. [Google Scholar] [CrossRef]
- Eathington, S.R.; Crosbie, T.M.; Edwards, M.D.; Reiter, R.S.; Bull, J.K. Molecular markers in a commercial breeding program. Crop Sci. 2007, 47, S-154–S-163. [Google Scholar] [CrossRef]
- Xu, C.; Ren, Y.; Jian, Y.; Guo, Z.; Zhang, Y.; Xie, C.; Fu, J.; Wang, H.; Wang, G.; Xu, Y.; Li, P.; Zou, C. Development of a maize 55 K SNP array with improved genome coverage for molecular breeding. Mol. Breed. 2017, 37, 20. [Google Scholar] [CrossRef] [PubMed]
- Cerrudo, D.; Cao, S.; Yuan, Y.; Martinez, C.; Suarez, E.A.; Babu, R.; Zhang, X.; Trachsel, S. Genomic selection outperforms marker assisted selection for grain yield and physiological traits in a maize doubled haploid population across water treatments. Front. Plant Sci. 2018, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; Dreisigacker, S.; Singh, R.; Zhang, X.; Gowda, M.; Roorkiwal, M.; Rutkoski, J.; Varshney, R.K. Genomic selection in plant breeding: Methods, models, and perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Heslot, N.; Jannink, J.-L.; Sorrells, M.E. Perspectives for genomic selection applications and research in plants. Crop Sci. 2015, 55, 1–12. [Google Scholar] [CrossRef]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic selection for drought tolerance using genome-wide SNPs in maize. Front. Plant Sci. 2017, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Dhliwayo, T.; Pixley, K.; Menkir, A.; Warburton, M. Combining ability, genetic distances, and heterosis among elite CIMMYT and IITA tropical maize inbred lines. Crop Sci. 2009, 49, 1201–1210. [Google Scholar] [CrossRef]
- Chander, S.; Guo, Y.Q.; Yang, X.H.; Zhang, Z.; Lu, X.Q.; Yan, J.B.; Song, T.M.; Rocheford, T.R.; Li, J.S. Using molecular markers to identify two major loci controlling carotenoid contents in maize grain. Theor. Appl. Genet. 2008, 116, 223–233. [Google Scholar] [CrossRef] [PubMed]
- VSN International Genstat for Windows, 23rd ed.; VSN International: Hemel Hempstead, UK, 2023.
- Chung, U.; Gbegbelegbe, S.; Shiferaw, B.; Robertson, R.; Yun, J.I.; Tesfaye, K.; Hoogenboom, G.; Sonder, K. Modeling the effect of a heat wave on maize production in the USA and its implications on food security in the developing world. Weather Clim. Extremes 2014, 5–6, 67–77. [Google Scholar] [CrossRef]
- Byerlee, D. The globalization of hybrid maize, 1921–70. J. Global History 2020, 15, 101–122. [Google Scholar] [CrossRef]
- Cassidy, E.S.; West, P.C.; Gerber, J.S.; Foley, J.A. Redefining agricultural yields: From tonnes to people nourished per hectare. Environ. Res. Lett. 2013, 8, 034015. [Google Scholar] [CrossRef]
- Burdon, J.J.; Zhan, J. Climate change and disease in plant communities. PLoS Biol. 2020, 18, e3000949. [Google Scholar] [CrossRef] [PubMed]
- Bocianowski, J.; Szulc, P.; Nowosad, K. Soil tillage methods by years interaction for dry matter of plant yield of maize (Zea mays L.) using additive main effects and multiplicative interaction model. J. Integr. Agric. 2018, 17, 2836–2839. [Google Scholar] [CrossRef]
- Bocianowski, J.; Nowosad, K.; Szulc, P. Soil tillage methods by years interaction for harvest index of maize (Zea mays L.) using additive main effects and multiplicative interaction model. Acta Agric. Scand. Sect. B 2019, 69, 75–81. [Google Scholar] [CrossRef]
- Picot, A.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Lannou, C.; Richard-Forget, F. Factors of the Fusarium verticillioides-maize environment modulating fumonisin production. Crit. Rev. Microbiol. 2010, 36, 221–231. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Wenda-Piesik, A.; Lemańczyk, G.; Twarużek, M.; Błajet-Kosicka, A.; Kazek, M.; Grajewski, J. Fusarium head blight incidence and detection of Fusarium toxins in wheat in relation to agronomic factors. Eur. J. Plant Pathol. 2017, 149, 515–531. [Google Scholar] [CrossRef]
- Ogliari, J.B.; Guimarães, M.A.; Geraldi, I.O.; Camargo, L.E.A. New resistance genes in the Zea mays – Exserohilum turcicum pathosystem. Genet. Mol. Biol. 2005, 28, 435–439. [Google Scholar] [CrossRef]
- Tagele, S.B.; Kim, S.W.; Lee, H.G.; Lee, Y.S. Potential of novel sequence type of Burkholderia cenocepacia for biological control of root rot of maize (Zea mays L.) caused by Fusarium temperatum. Int. J. Mol. Sci. 2019, 20, 1–18. [Google Scholar] [CrossRef]
- Bocianowski, J.; Szulc, P.; Waśkiewicz, A.; Nowosad, K.; Kobus-Cisowska, J. Ergosterol and Fusarium mycotoxins content in two maize cultivars under different forms of nitrogen fertilizers. J. Phytopathol. 2019, 167, 516–526. [Google Scholar] [CrossRef]
- Bocianowski, J.; Szulc, P.; Waśkiewicz, A.; Cyplik, A. The Effect of Agrotechnical Factors on Fusarium Mycotoxins Level in Maize. Agriculture 2020, 10, 528. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, S.; Li, W.X.; Liu, J.; Guo, W.; Xu, M.; Xu, Y. QTL mapping and genomic selection for Fusarium ear rot resistance using two F2:3 populations in maize. Euphytica 2022, 218, 131. [Google Scholar] [CrossRef]
- Maschietto, V.; Cinzia, C.; Pirona, R.; Pea, G.; Strozzi, F.; Marocco Adriano-Rossini, L.; Lanubile, A. QTL Mapping and candidate genes for resistance to Fusarium Ear Rot and fumonisin contamination in Maize. BMC Plant Biol. 2017, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: a review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef]
- Brachi, B.; Faure, N.; Horton, M.; Flahauw, E.; Vazquez, A.; Nordborg, M.; Bergelson, J.; Cuguen, J.; Roux, F. Linkage and association mapping of Arabidopsis thaliana flowering time in nature. PLoS Genet. 2010, 6, e1000940. [Google Scholar] [CrossRef] [PubMed]
- Bocianowski, J.; Nowosad, K.; Wróbel, B.; Szulc, P. Identification of Associations between SSR Markers and Quantitative Traits of Maize (Zea mays L.). Agronomy 2021, 11, 182. [Google Scholar] [CrossRef]
- Sobiech, A.; Tomkowiak, A.; Bocianowski, J.; Szymańska, G.; Nowak, B.; Lenort, M. Identification and analysis of candidate genes associated with maize fusarium cob resistance using next-generation sequencing technology. Int. J. Mol. Sci. 2023, 24, 16712. [Google Scholar] [CrossRef] [PubMed]
- Bocianowski, J.; Tomkowiak, A.; Bocianowska, M.; Sobiech, A. The use of DArTseq technology to identify markers related to the heterosis effects in selected traits in maize. Curr. Issues Mol. Biol. 2023, 45, 2644–2660. [Google Scholar] [CrossRef] [PubMed]
- Tomkowiak, A.; Nowak, B.; Sobiech, A.; Bocianowski, J.; Wolko, Ł.; Spychała, J. The use of DArTseq technology to identify new SNP and SilicoDArT markers related to the yield-related traits components in maize. Genes 2022, 13, 848. [Google Scholar] [CrossRef] [PubMed]
- Nowak, B.; Tomkowiak, A.; Bocianowski, J.; Sobiech, A.; Bobrowska, R.; Kowalczewski, P.; Bocianowska, M. The Use of DArTseq Technology to Identify Markers Linked to Genes Responsible for Seed Germination and Seed Vigor in Maize. Int. J. Mol. Sci. 2022, 23, 14865. [Google Scholar] [CrossRef] [PubMed]
- Muñiz, L.M.; Royo, J.; Gómez, E.; Baudot, G.; Paul, W.; Hueros, G. Atypical response regulators expressed in the maize endosperm transfer cells link canonical two component systems and seed biology. BMC Plant Biol. 2010, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Dong, H.; Mu, J.; Ren, B.; Zheng, B.; Ji, Z.; Yang, W.-C.; Liang, Y.; Zuo, J. Arabidopsis histidine kinase CKI1 acts upstream of HISTIDINE PHOSPHOTRANSFER PROTEINS to regulate female gametophyte development and vegetative growth. Plant Cell 2010, 22, 1232–1248. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shen, J.; Liang, J. Genome-wide identification, expression profile, and alternative splicing analysis of the brassinosteroid-signaling kinase (BSK) family genes in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 1138. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Willige, B.C.; Jaillais, Y.; Geng, S.; Park, M.Y.; Gray, W.M.; Chory, J. BRASSINOSTEROIDSIGNALING KINASE 3, a plasma membrane-associated scaffold protein involved in early brassinosteroid signaling. PLoS Genet. 2019, 15, e1007904. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Density charts showing the distribution of the Fusarium stalk rot.
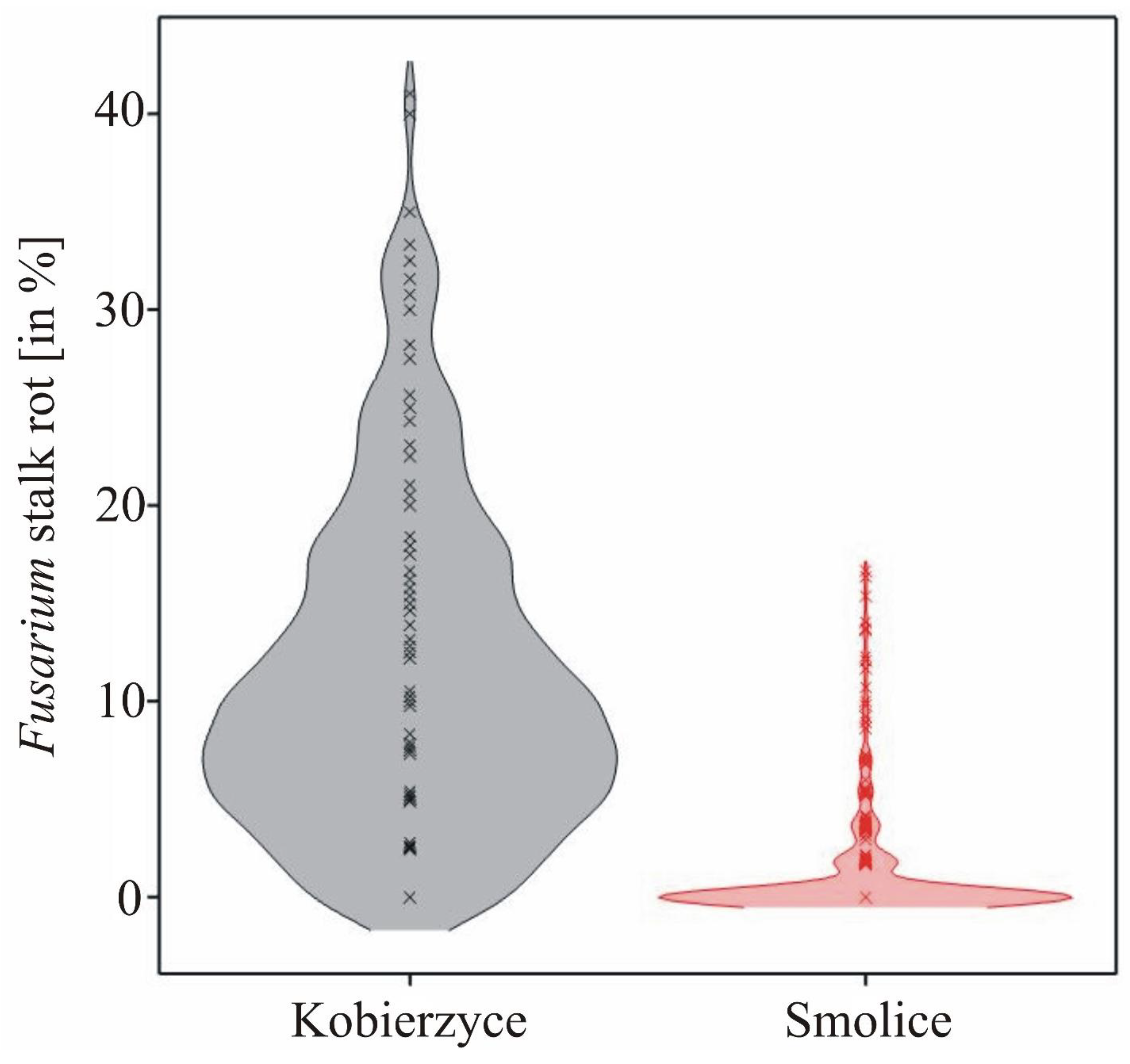
Figure 2.
Dendrogram showing the genetic similarity between the 122 analyzed hybrids constructed based on the observation of 32,900 markers (26,234 SilicoDArTs and 6,666 SNPs).
Figure 2.
Dendrogram showing the genetic similarity between the 122 analyzed hybrids constructed based on the observation of 32,900 markers (26,234 SilicoDArTs and 6,666 SNPs).
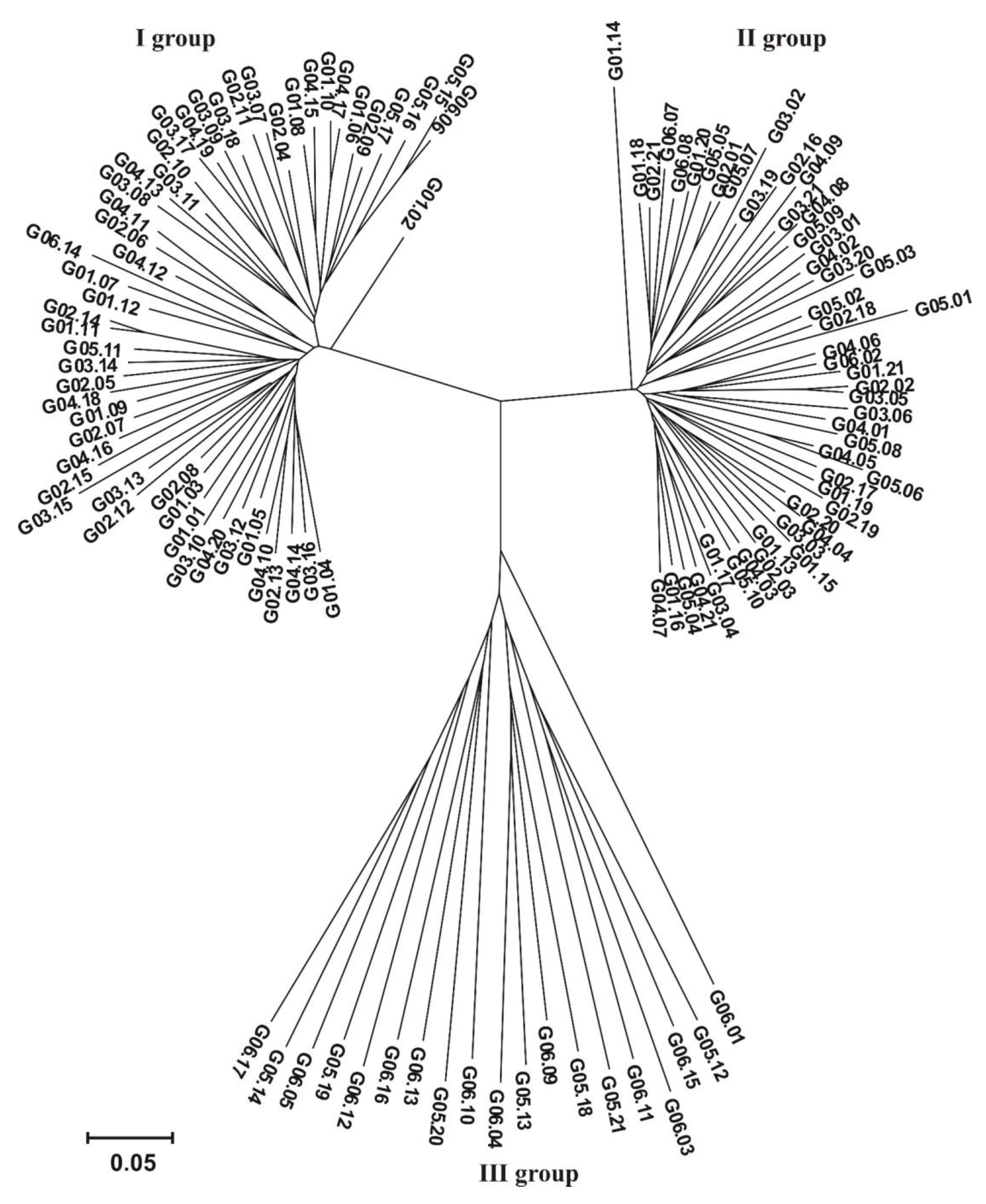
Figure 3.
Manhattan plot for Fusarium stalk rot in Smolice.
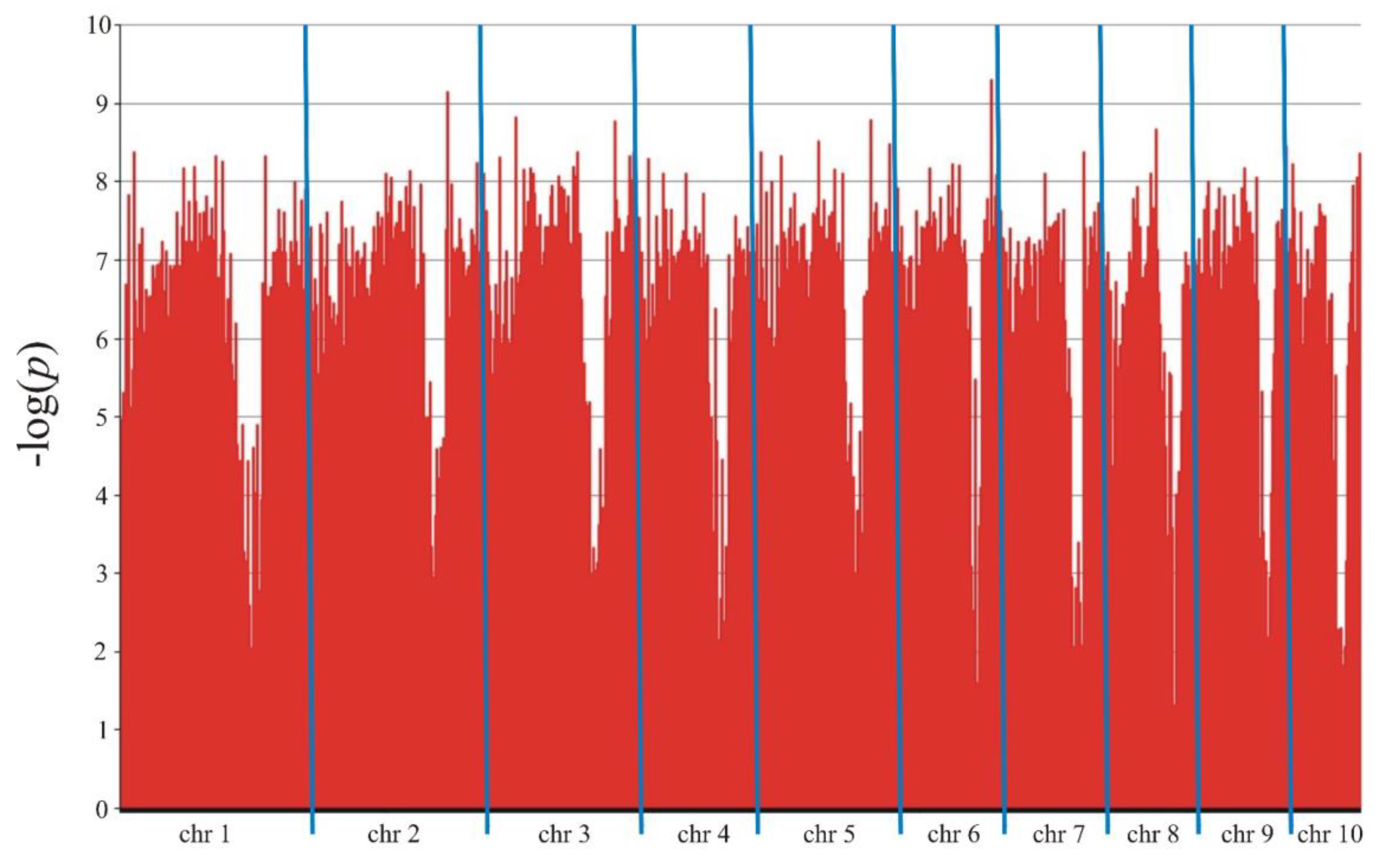
Figure 4.
Manhattan plot for Fusarium stalk rot in Kobierzyce.
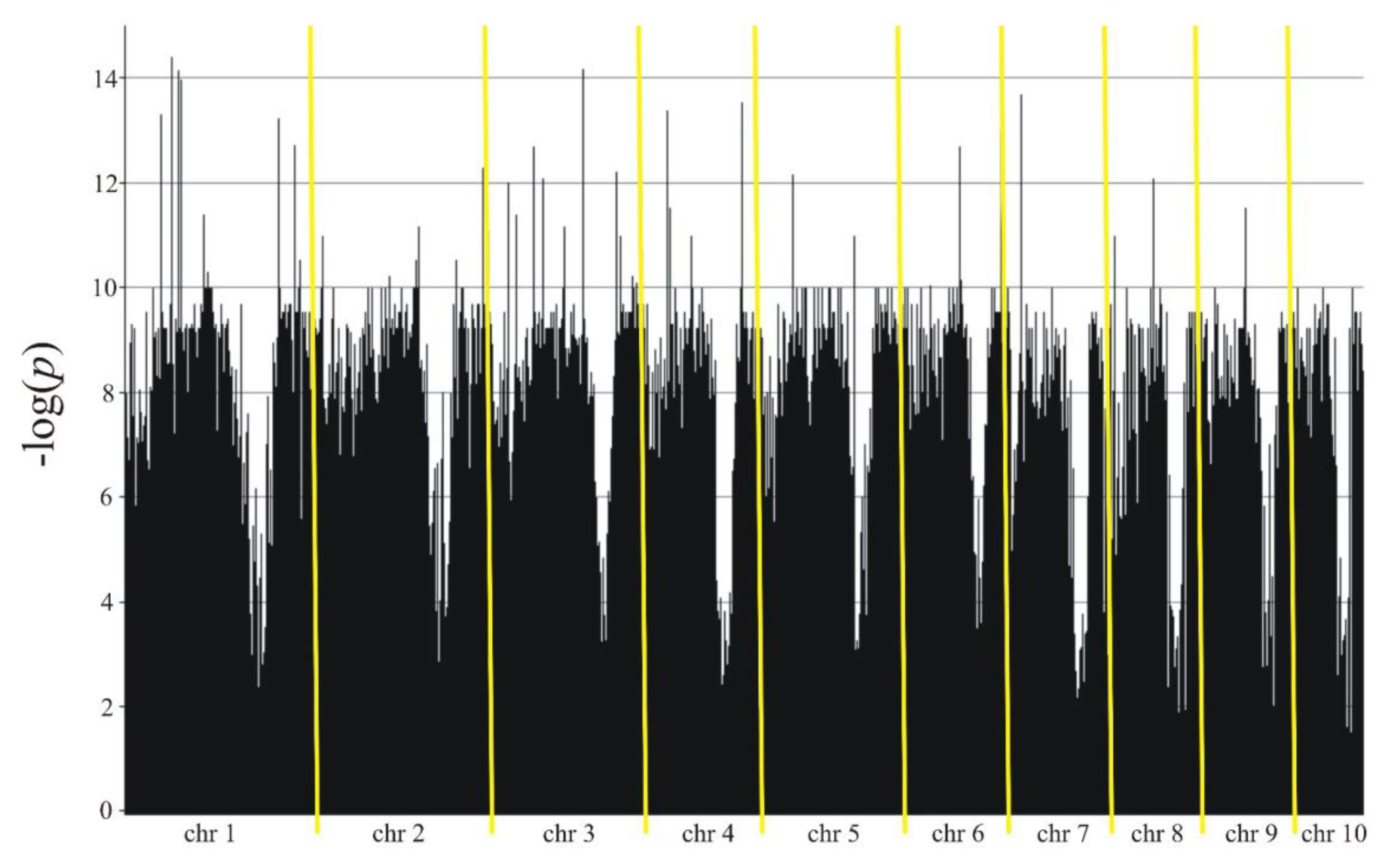

Table 1.
Results of two-way analysis of variance for Fusarium stalk rot.
| Source of variation | The number of degrees of freedom | Sum of square | Mean square | F statistics |
|---|---|---|---|---|
| Genotype | 121 | 8468.68 | 69.99 | 2.45 *** |
| Location | 1 | 18988.97 | 18988.97 | 664.68 *** |
| Genotype × Location | 121 | 5765.31 | 47.65 | 1.67 *** |
| Residual | 488 | 13941.44 | 28.57 | |
| Total | 731 | 47164.40 |
*** p<0.001.
Table 2.
Mean values and standard deviation (s.d.) of Fusarium stalk rot for particular genotypes in two locations as well as for the average of the locations.
Table 2.
Mean values and standard deviation (s.d.) of Fusarium stalk rot for particular genotypes in two locations as well as for the average of the locations.
| Location | Kobierzyce | Smolice | Average | Location | Kobierzyce | Smolice | Average | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. | Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. |
| G01.01 | 5.83 | 2.89 | 0.00 | 0.00 | 2.92 | 3.68 | G03.20 | 5.83 | 8.04 | 0.61 | 1.05 | 3.22 | 5.87 |
| G01.02 | 6.84 | 3.92 | 0.00 | 0.00 | 3.42 | 4.49 | G03.21 | 25.98 | 7.39 | 5.45 | 9.45 | 15.72 | 13.56 |
| G01.03 | 6.80 | 1.57 | 1.82 | 3.15 | 4.31 | 3.52 | G04.01 | 12.68 | 2.24 | 0.00 | 0.00 | 6.34 | 7.09 |
| G01.04 | 9.11 | 10.12 | 0.00 | 0.00 | 4.55 | 8.11 | G04.02 | 11.67 | 1.44 | 5.85 | 1.92 | 8.76 | 3.53 |
| G01.05 | 7.50 | 2.50 | 0.57 | 0.99 | 4.04 | 4.16 | G04.03 | 8.33 | 8.04 | 0.62 | 1.07 | 4.48 | 6.65 |
| G01.06 | 5.83 | 1.44 | 0.00 | 0.00 | 2.92 | 3.32 | G04.04 | 4.19 | 2.87 | 1.16 | 1.00 | 2.67 | 2.54 |
| G01.07 | 15.11 | 8.97 | 1.25 | 1.09 | 8.18 | 9.50 | G04.05 | 1.67 | 2.89 | 0.61 | 1.05 | 1.14 | 2.03 |
| G01.08 | 4.17 | 1.45 | 0.00 | 0.00 | 2.09 | 2.46 | G04.06 | 11.86 | 6.43 | 2.44 | 2.13 | 7.15 | 6.71 |
| G01.09 | 10.22 | 6.75 | 0.00 | 0.00 | 5.11 | 7.04 | G04.07 | 3.33 | 3.82 | 0.00 | 0.00 | 1.67 | 3.03 |
| G01.10 | 1.65 | 1.43 | 0.00 | 0.00 | 0.82 | 1.28 | G04.08 | 14.46 | 7.48 | 5.18 | 7.46 | 9.82 | 8.39 |
| G01.11 | 14.53 | 7.84 | 0.58 | 1.01 | 7.56 | 9.13 | G04.09 | 5.00 | 4.33 | 0.00 | 0.00 | 2.50 | 3.87 |
| G01.12 | 8.55 | 4.18 | 0.00 | 0.00 | 4.28 | 5.38 | G04.10 | 12.52 | 11.43 | 3.95 | 4.36 | 8. 23 | 9.05 |
| G01.13 | 7.61 | 8.56 | 0.00 | 0.00 | 3.81 | 6.84 | G04.11 | 7.65 | 8.92 | 0.58 | 1.01 | 4.12 | 6.87 |
| G01.14 | 4.27 | 3.92 | 0.00 | 0.00 | 2.14 | 3.41 | G04.12 | 5.09 | 5.00 | 0.00 | 0.00 | 2.54 | 4.22 |
| G01.15 | 23.72 | 2.80 | 0.00 | 0.00 | 11.86 | 13.11 | G04.13 | 3.33 | 2.89 | 0.00 | 0.00 | 1.67 | 2.58 |
| G01.16 | 11.97 | 7.83 | 0.00 | 0.00 | 5.98 | 8.22 | G04.14 | 16.05 | 7.59 | 0.57 | 0.99 | 8.31 | 9.76 |
| G01.17 | 15.83 | 3.82 | 3.33 | 5.77 | 9.58 | 8.13 | G04.15 | 13.33 | 16.65 | 0.00 | 0.00 | 6.67 | 12.81 |
| G01.18 | 8.38 | 6.23 | 0.72 | 1.25 | 4.55 | 5.81 | G04.16 | 23.61 | 9.22 | 1.85 | 3.21 | 12.73 | 13.42 |
| G01.19 | 16.89 | 7.03 | 0.00 | 0.00 | 8.44 | 10.26 | G04.17 | 10.83 | 8.04 | 0.00 | 0.00 | 5.42 | 7.81 |
| G01.20 | 16.09 | 12.78 | 0.00 | 0.00 | 8.05 | 11.96 | G04.18 | 19.38 | 8.27 | 3.57 | 6.18 | 11.48 | 10.85 |
| G01.21 | 16.77 | 9.40 | 0.00 | 0.00 | 8.39 | 10.94 | G04.19 | 10.00 | 2.50 | 0.71 | 1.23 | 5.36 | 5.39 |
| G02.01 | 1.67 | 1.44 | 0.00 | 0.00 | 0.83 | 1.29 | G04.20 | 21.03 | 3.07 | 1.23 | 2.14 | 11.13 | 11.10 |
| G02.02 | 10.00 | 2.50 | 3.61 | 3.64 | 6.81 | 4.48 | G04.21 | 9.40 | 10.36 | 1.36 | 2.36 | 5.38 | 8.04 |
| G02.03 | 2.50 | 0.00 | 0.00 | 0.00 | 1.25 | 1.37 | G05.01 | 5.83 | 1.44 | 1.96 | 2.10 | 3.90 | 2.66 |
| G02.04 | 15.06 | 8.94 | 0.00 | 0.00 | 7.53 | 10.00 | G05.02 | 11.84 | 5.49 | 0.00 | 0.00 | 5.92 | 7.36 |
| G02.05 | 13.42 | 7.50 | 0.00 | 0.00 | 6.71 | 8.75 | G05.03 | 15.11 | 4.24 | 6.36 | 6.06 | 10.74 | 6.70 |
| G02.06 | 15.83 | 1.44 | 0.58 | 1.01 | 8.21 | 8.43 | G05.04 | 9.07 | 3.69 | 0.00 | 0.00 | 4.53 | 5.49 |
| G02.07 | 11.10 | 6.84 | 0.00 | 0.00 | 5.55 | 7.46 | G05.05 | 15.17 | 5.26 | 4.85 | 4.32 | 10.01 | 7.10 |
| G02.08 | 21.90 | 5.60 | 5.64 | 5.84 | 13.77 | 10.27 | G05.06 | 4.23 | 3.90 | 1.26 | 2.18 | 2.74 | 3.26 |
| G02.09 | 4.25 | 3.85 | 0.00 | 0.00 | 2.13 | 3.37 | G05.07 | 9.65 | 8.46 | 2.30 | 3.98 | 5.98 | 7.15 |
| G02.10 | 10.96 | 7.33 | 4.68 | 8.11 | 7.82 | 7.72 | G05.08 | 6.67 | 2.89 | 1.75 | 3.04 | 4.21 | 3.78 |
| G02.11 | 7.91 | 7.31 | 0.62 | 1.07 | 4.26 | 6.15 | G05.09 | 15.83 | 10.10 | 3.07 | 2.76 | 9.45 | 9.63 |
| G02.12 | 16.71 | 13.70 | 1.89 | 1.96 | 9.30 | 11.94 | G05.10 | 5.88 | 3.81 | 0.62 | 1.07 | 3.25 | 3.81 |
| G02.13 | 8.33 | 2.89 | 0.00 | 0.00 | 4.17 | 4.92 | G05.11 | 16.67 | 11.82 | 2.90 | 3.50 | 9.78 | 10.84 |
| G02.14 | 17.27 | 6.23 | 1.19 | 2.06 | 9.23 | 9.74 | G05.12 | 13.14 | 6.84 | 0.57 | 0.99 | 6.86 | 8.16 |
| G02.15 | 4.30 | 5.33 | 0.00 | 0.00 | 2.15 | 4.11 | G05.13 | 21.67 | 7.22 | 0.67 | 1.16 | 11.17 | 12.40 |
| G02.16 | 8.51 | 3.05 | 0.00 | 0.00 | 4.26 | 5.05 | G05.14 | 14.17 | 9.47 | 0.00 | 0.00 | 7.08 | 9.80 |
| G02.17 | 6.67 | 3.82 | 0.00 | 0.00 | 3.33 | 4.38 | G05.15 | 21.94 | 10.55 | 1.19 | 2.06 | 11.57 | 13.25 |
| G02.18 | 11.84 | 2.75 | 0.58 | 1.01 | 6.21 | 6.44 | G05.16 | 3.42 | 5.92 | 0.00 | 0.00 | 1.71 | 4.19 |
| G02.19 | 5.04 | 0.08 | 0.00 | 0.00 | 2.52 | 2.76 | G05.17 | 11.56 | 5.38 | 0.00 | 0.00 | 5.78 | 7.19 |
| G02.20 | 17.68 | 12.84 | 1.72 | 2.99 | 9.70 | 12.08 | G05.18 | 6.10 | 4.08 | 0.00 | 0.00 | 3.05 | 4.22 |
| G02.21 | 3.42 | 2.97 | 0.00 | 0.00 | 1.71 | 2.65 | G05.19 | 12.65 | 5.23 | 4.97 | 2.72 | 8.81 | 5.62 |
| G03.01 | 5.00 | 4.33 | 3.02 | 2.20 | 4.01 | 3.26 | G05.20 | 17.50 | 9.01 | 2.21 | 1.92 | 9.85 | 10.21 |
| G03.02 | 10.94 | 8.37 | 0.63 | 1.09 | 5.79 | 7.77 | G05.21 | 10.92 | 1.37 | 2.35 | 2.67 | 6.64 | 5.07 |
| G03.03 | 11.82 | 10.24 | 0.00 | 0.00 | 5.91 | 9.15 | G06.01 | 9.34 | 10.38 | 1.82 | 3.15 | 5.58 | 8.00 |
| G03.04 | 1.67 | 2.89 | 1.17 | 2.03 | 1.42 | 2.25 | G06.02 | 6.84 | 5.93 | 0.00 | 0.00 | 3.42 | 5.30 |
| G03.05 | 19.57 | 10.48 | 6.28 | 8.07 | 12.93 | 11.09 | G06.03 | 9.43 | 5.65 | 1.73 | 1.73 | 5.58 | 5.63 |
| G03.06 | 19.62 | 13.23 | 0.00 | 0.00 | 9.81 | 13.62 | G06.04 | 20.21 | 13.80 | 0.00 | 0.00 | 10.11 | 14.10 |
| G03.07 | 31.18 | 16.18 | 2.98 | 5.16 | 17.08 | 18.81 | G06.05 | 7.54 | 6.59 | 0.00 | 0.00 | 3.77 | 5.87 |
| G03.08 | 24.32 | 7.44 | 0.00 | 0.00 | 12.16 | 14.13 | G06.06 | 17.50 | 5.00 | 1.13 | 0.98 | 9.32 | 9.53 |
| G03.09 | 14.92 | 8.73 | 0.00 | 0.00 | 7.46 | 9.86 | G06.07 | 9.38 | 6.49 | 0.57 | 0.99 | 4.98 | 6.36 |
| G03.10 | 4.23 | 3.90 | 0.57 | 0.99 | 2.40 | 3.24 | G06.08 | 10.00 | 2.50 | 1.15 | 1.00 | 5.58 | 5.14 |
| G03.11 | 13.33 | 8.04 | 0.00 | 0.00 | 6.67 | 8.90 | G06.09 | 9.19 | 7.09 | 0.00 | 0.00 | 4.59 | 6.74 |
| G03.12 | 5.92 | 5.31 | 1.74 | 1.76 | 3.83 | 4.21 | G06.10 | 14.98 | 2.36 | 1.80 | 1.85 | 8.39 | 7.47 |
| G03.13 | 15.61 | 14.03 | 4.55 | 7.88 | 10.08 | 11.84 | G06.11 | 14.17 | 11.55 | 0.00 | 0.00 | 7.08 | 10.66 |
| G03.14 | 15.46 | 4.85 | 0.00 | 0.00 | 7.73 | 9.00 | G06.12 | 25.15 | 7.28 | 0.60 | 1.03 | 12.87 | 14.23 |
| G03.15 | 15.00 | 5.00 | 5.32 | 6.03 | 10.16 | 7.26 | G06.13 | 14.49 | 7.79 | 0.00 | 0.00 | 7.25 | 9.34 |
| G03.16 | 5.88 | 1.41 | 0.00 | 0.00 | 2.94 | 3.34 | G06.14 | 11.84 | 8.17 | 0.00 | 0.00 | 5.92 | 8.29 |
| G03.17 | 13.55 | 1.30 | 6.16 | 9.14 | 9.86 | 7.11 | G06.15 | 10.92 | 6.28 | 0.00 | 0.00 | 5.46 | 7.18 |
| G03.18 | 5.04 | 2.50 | 0.00 | 0.00 | 2.52 | 3.18 | G06.16 | 5.77 | 1.34 | 0.00 | 0.00 | 2.89 | 3.27 |
| G03.19 | 15.28 | 9.29 | 3.64 | 3.57 | 9.46 | 8.96 | G06.17 | 5.83 | 2.89 | 0.00 | 0.00 | 2.92 | 3.68 |
| Location | 11.39 | 8.33 | 1.20 | 2.79 | |||||||||
| LSD0.05 – Genotype: 6.06; Location: 0.78; Genotype x Location: 8.58 | |||||||||||||
LSD – Least Significant Difference.
Table 3.
SilicoDArT and SNP molecular markers significantly associated with maize plant resistance to Fusarium stalk rot in Smolice (significant associations selected at p<0.001 with correction for Benjamini-Hochberg multiple testing).
Table 3.
SilicoDArT and SNP molecular markers significantly associated with maize plant resistance to Fusarium stalk rot in Smolice (significant associations selected at p<0.001 with correction for Benjamini-Hochberg multiple testing).
| Type of markers | All | SilicoDArT | SNP | |
|---|---|---|---|---|
| The number of markers | 11282 | 8405 | 2877 | |
| Negative | Numbers | 5800 | 4336 | 1464 |
| Effects | -17.06 – -2.14 | -13.88 – -2.14 | -17.06 – -2.14 | |
| Percentage variance accounted for | 2.4 – 31.4 | 2.4 – 31.4 | 2.4 – 30.5 | |
| LOD | 1.30 – 12.70 | 1.30 – 16.70 | 1.30 – 11.10 | |
| Positive | Numbers | 5482 | 4069 | 1413 |
| Effects | 2.14 – 8.55 | 2.14 – 8.55 | 2.17 – 7.37 | |
| Percentage variance accounted for | 2.4 – 34.4 | 2.4 – 33.7 | 2.4 – 34.4 | |
| LOD | 1.30 – 14.40 | 1.30 – 14.40 | 1.30 – 13.53 | |
Table 4.
SilicoDArT and SNP molecular markers significantly associated with maize plant resistance to Fusarium stalk rot in Kobierzyce (significant associations selected at p<0.001 with correction for Benjamini-Hochberg multiple testing).
Table 4.
SilicoDArT and SNP molecular markers significantly associated with maize plant resistance to Fusarium stalk rot in Kobierzyce (significant associations selected at p<0.001 with correction for Benjamini-Hochberg multiple testing).
| Type of markers | All | SilicoDArT | SNP | |
|---|---|---|---|---|
| The number of markers | 9439 | 7148 | 2291 | |
| Negative | Numbers | 4523 | 3437 | 1086 |
| Effects | -6.49 – -0.78 | -6.79 – -0.777 | -4.38 – -0.784 | |
| Percentage variance accounted for | 2.4 – 27.0 | 2.4 – 25.7 | 2.4 – 27.0 | |
| LOD | 1.30 – 9.30 | 1.30 – 8.82 | 1.30 – 9.30 | |
| Positive | Numbers | 4916 | 3711 | 1205 |
| Effects | 0.779 – 2.214 | 0.779 – 2.214 | 0.788 – 2.173 | |
| Percentage variance accounted for | 2.4 – 25.3 | 2.4 – 25.3 | 2.4 – 23.9 | |
| LOD | 1.30 – 8.68 | 1.30 – 8.68 | 1.31 – 8.17 | |
Table 5.
Ninety-one markers highly statistically significant (LOD>7.5) simultaneously in both localities: Kobierzyce and Smolice.
Table 5.
Ninety-one markers highly statistically significant (LOD>7.5) simultaneously in both localities: Kobierzyce and Smolice.
| Chromosome | Marker type | CloneID | Kobierzyce | Smolice | ||||
|---|---|---|---|---|---|---|---|---|
| Estimate | Percen1 | LOD | Estimate | Percen | LOD | |||
| 1 | SilicoDArT | 29628241 | 5.902 | 23.4 | 8.02 | 2.167 | 24.3 | 8.33 |
| 1 | SilicoDArT | 82348823 | 6.031 | 24.4 | 8.34 | 2.167 | 24.2 | 8.27 |
| 1 | SilicoDArT | 24026805 | 6.219 | 26.0 | 8.92 | 2.156 | 24.0 | 8.20 |
| 1 | SNP | 4772298|F|0-68:A>G-68:A>G | -6.032 | 24.5 | 8.40 | -2.127 | 23.4 | 8.00 |
| 1 | SilicoDArT | 2488934 | -5.894 | 22.1 | 7.54 | -2.166 | 22.9 | 7.84 |
| 1 | SNP | 2439850|F|0-12:A>C-12:A>C | -6.061 | 24.9 | 8.52 | -2.092 | 22.7 | 7.76 |
| 1 | SilicoDArT | 4774875 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 1 | SilicoDArT | 9626410 | 6.623 | 29.9 | 10.30 | 2.073 | 22.3 | 7.62 |
| 1 | SNP | 2529315|F|0-56:T>G-56:T>G | -6.163 | 25.8 | 8.82 | -2.073 | 22.3 | 7.62 |
| 1 | SilicoDArT | 7054095 | -6.361 | 27.5 | 9.52 | -2.073 | 22.3 | 7.61 |
| 1 | SilicoDArT | 7051768 | 6.122 | 25.4 | 8.70 | 2.071 | 22.2 | 7.60 |
| 2 | SNP | 4772836|F|0-17:C>A-17:C>A | -6.722 | 30.5 | 11.10 | -2.148 | 23.7 | 8.11 |
| 2 | SNP | 4582743|F|0-42:G>A-42:G>A | -6.073 | 24.6 | 8.42 | -2.135 | 23.3 | 7.97 |
| 2 | SilicoDArT | 9694283 | 5.932 | 23.8 | 8.12 | 2.100 | 22.8 | 7.81 |
| 2 | SilicoDArT | 82349036 | 6.234 | 26.4 | 9.05 | 2.092 | 22.7 | 7.76 |
| 2 | SilicoDArT | 2395963 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 2 | SilicoDArT | 2382023 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 2 | SilicoDArT | 9703016 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 2 | SilicoDArT | 25942787 | 5.764 | 22.3 | 7.63 | 2.088 | 22.5 | 7.69 |
| 2 | SilicoDArT | 9633940 | 6.000 | 24.1 | 8.24 | 2.093 | 22.5 | 7.68 |
| 2 | SilicoDArT | 9718212 | 5.965 | 24.1 | 8.24 | 2.073 | 22.3 | 7.61 |
| 2 | SilicoDArT | 4778784 | -6.347 | 27.4 | 9.40 | -2.072 | 22.3 | 7.61 |
| 2 | SilicoDArT | 29619311 | 5.742 | 22.2 | 7.59 | 2.066 | 22.1 | 7.54 |
| 3 | SNP | 4772456|F|0-52:T>G-52:T>G | -5.802 | 22.5 | 7.68 | -2.227 | 25.6 | 8.77 |
| 3 | SNP | 4585365|F|0-34:T>C-34:T>C | -5.979 | 23.8 | 8.15 | -2.186 | 24.5 | 8.39 |
| 3 | SilicoDArT | 9682713 | -5.718 | 22.0 | 7.52 | -2.138 | 23.7 | 8.11 |
| 3 | SilicoDArT | 9717799 | 5.981 | 24.0 | 8.22 | 2.140 | 23.6 | 8.08 |
| 3 | SilicoDArT | 77157803 | 5.877 | 23.2 | 7.92 | 2.140 | 23.6 | 8.08 |
| 3 | SNP | 2433795|F|0-30:G>C-30:G>C | 5.996 | 24.2 | 8.29 | 2.129 | 23.4 | 8.02 |
| 3 | SilicoDArT | 77158337 | 5.768 | 22.5 | 7.68 | 2.111 | 23.1 | 7.91 |
| 3 | SilicoDArT | 29621917 | -5.998 | 24.4 | 8.36 | -2.101 | 23.0 | 7.85 |
| 3 | SilicoDArT | 5583810 | 6.071 | 24.9 | 8.54 | 2.100 | 22.8 | 7.81 |
| 3 | SNP | 4774080|F|0-65:C>T-65:C>T | -5.868 | 23.3 | 7.95 | -2.092 | 22.7 | 7.76 |
| 3 | SNP | 4774088|F|0-45:G>A-45:G>A | -5.868 | 23.3 | 7.95 | -2.092 | 22.7 | 7.76 |
| 3 | SNP | 7048352|F|0-24:A>G-24:A>G | -5.737 | 22.2 | 7.59 | -2.092 | 22.7 | 7.76 |
| 3 | SNP | 5586725|F|0-40:C>A-40:C>A | -5.761 | 22.2 | 7.60 | -2.099 | 22.7 | 7.75 |
| 3 | SilicoDArT | 82349016 | 6.017 | 24.5 | 8.38 | 2.074 | 22.3 | 7.60 |
| 3 | SilicoDArT | 4768318 | -5.845 | 23.1 | 7.90 | -2.067 | 22.2 | 7.58 |
| 3 | SilicoDArT | 34685358 | 6.096 | 25.2 | 8.64 | 2.066 | 22.2 | 7.57 |
| 3 | SNP | 2403483|F|0-62:C>A-62:C>A | -5.881 | 23.4 | 8.00 | -2.066 | 22.2 | 7.57 |
| 3 | SNP | 4771426|F|0-54:A>G-54:A>G | 6.129 | 25.3 | 8.66 | 2.072 | 22.1 | 7.55 |
| 3 | SNP | 4592970|F|0-41:T>C-41:T>C | -5.924 | 23.6 | 8.08 | -2.067 | 22.1 | 7.54 |
| 3 | SNP | 4586493|F|0-10:T>C-10:T>C | -5.734 | 22.1 | 7.56 | -2.063 | 22.0 | 7.52 |
| 4 | SilicoDArT | 70092308 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 4 | SilicoDArT | 9680684 | 5.823 | 22.9 | 7.83 | 2.092 | 22.7 | 7.76 |
| 4 | SilicoDArT | 25001071 | 5.854 | 23.1 | 7.90 | 2.078 | 22.4 | 7.64 |
| 4 | SilicoDArT | 25942566 | 5.747 | 22.2 | 7.60 | 2.078 | 22.4 | 7.64 |
| 4 | SilicoDArT | 25004669 | -6.261 | 26.3 | 9.05 | -2.078 | 22.1 | 7.56 |
| 5 | SNP | 4583014|F|0-63:A>G-63:A>G | -5.998 | 24.0 | 8.20 | -2.235 | 25.6 | 8.80 |
| 5 | SNP | 2536415|F|0-25:C>T-25:C>T | -5.926 | 23.6 | 8.09 | -2.187 | 24.8 | 8.48 |
| 5 | SilicoDArT | 4578971 | -5.881 | 23.2 | 7.93 | -2.177 | 24.5 | 8.38 |
| 5 | SilicoDArT | 2401113 | -6.205 | 26.0 | 8.92 | -2.168 | 24.3 | 8.33 |
| 5 | SilicoDArT | 29628894 | 6.294 | 26.8 | 9.22 | 2.148 | 23.9 | 8.17 |
| 5 | SilicoDArT | 2432042 | -5.722 | 22.0 | 7.51 | -2.128 | 23.4 | 8.01 |
| 5 | SilicoDArT | 2499631 | -6.401 | 27.7 | 9.52 | -2.108 | 23.0 | 7.85 |
| 5 | SilicoDArT | 9690308 | 6.326 | 27.2 | 9.40 | 2.092 | 22.7 | 7.76 |
| 5 | SilicoDArT | 9681187 | 6.145 | 25.6 | 8.77 | 2.092 | 22.7 | 7.76 |
| 5 | SilicoDArT | 2539842 | 6.016 | 24.5 | 8.38 | 2.081 | 22.4 | 7.66 |
| 5 | SilicoDArT | 4776114 | 6.062 | 24.8 | 8.51 | 2.081 | 22.4 | 7.66 |
| 5 | SilicoDArT | 4779143 | 6.465 | 28.5 | 9.70 | 2.066 | 22.2 | 7.57 |
| 5 | SilicoDArT | 2532640 | 6.355 | 27.5 | 9.52 | 2.066 | 22.2 | 7.57 |
| 6 | SNP | 4771464|F|0-65:C>T-65:C>T | -6.251 | 25.7 | 8.82 | -2.309 | 27.0 | 9.30 |
| 6 | SilicoDArT | 5587687 | 5.989 | 24.1 | 8.24 | 2.158 | 24.0 | 8.22 |
| 6 | SilicoDArT | 4770044 | -6.073 | 24.6 | 8.42 | -2.160 | 23.9 | 8.17 |
| 6 | SilicoDArT | 5588627 | 5.855 | 23.1 | 7.90 | 2.119 | 23.3 | 7.96 |
| 6 | SNP | 4770865|F|0-46:C>T-46:C>T | -6.028 | 24.6 | 8.41 | -2.101 | 22.9 | 7.81 |
| 6 | SilicoDArT | 2530960 | 5.794 | 22.6 | 7.73 | 2.098 | 22.8 | 7.79 |
| 6 | SilicoDArT | 9698143 | 6.459 | 28.2 | 9.70 | 2.084 | 22.4 | 7.67 |
| 6 | SNP | 4764810|F|0-20:T>C-20:T>C | -6.424 | 27.2 | 9.40 | -2.107 | 22.3 | 7.63 |
| 6 | SilicoDArT | 4775726 | 6.185 | 26.0 | 8.92 | 2.073 | 22.3 | 7.63 |
| 6 | SilicoDArT | 24029725 | 5.850 | 23.1 | 7.90 | 2.073 | 22.3 | 7.61 |
| 6 | SNP | 25947704|F|0-38:C>T-38:C>T | -6.129 | 25.4 | 8.72 | -2.061 | 22.0 | 7.51 |
| 7 | SNP | 67225764|F|0-56:C>G-56:C>G | -6.228 | 26.3 | 9.05 | -2.073 | 22.3 | 7.61 |
| 7 | SNP | 2523212|F|0-36:C>T-36:C>T | -5.778 | 22.5 | 7.70 | -2.072 | 22.3 | 7.61 |
| 8 | SilicoDArT | 9694332 | 5.899 | 23.5 | 8.03 | 2.081 | 22.4 | 7.66 |
| 8 | SilicoDArT | 9698173 | 5.991 | 24.3 | 8.33 | 2.066 | 22.2 | 7.57 |
| 8 | SilicoDArT | 4766257 | 5.708 | 22.0 | 7.52 | 2.060 | 22.0 | 7.52 |
| 9 | SNP | 2475427|F|0-37:A>G-37:A>G | -6.020 | 24.2 | 8.27 | -2.195 | 24.7 | 8.46 |
| 9 | SilicoDArT | 77157588 | 5.888 | 22.8 | 7.81 | 2.156 | 23.6 | 8.06 |
| 9 | SilicoDArT | 4584767 | -5.958 | 23.6 | 8.09 | -2.140 | 23.4 | 8.01 |
| 9 | SilicoDArT | 7049648 | -6.051 | 24.7 | 8.46 | -2.083 | 22.4 | 7.66 |
| 9 | SilicoDArT | 2384252 | 5.712 | 22.0 | 7.52 | 2.072 | 22.3 | 7.61 |
| 9 | SilicoDArT | 25947915 | 5.811 | 22.5 | 7.71 | 2.082 | 22.2 | 7.60 |
| 9 | SilicoDArT | 73750965 | 5.912 | 23.5 | 8.05 | 2.065 | 22.0 | 7.52 |
| 10 | SNP | 9667431|F|0-38:G>C-38:G>C | 6.403 | 27.8 | 9.52 | 2.120 | 23.3 | 7.96 |
| 10 | SilicoDArT | 2575390 | 5.817 | 22.9 | 7.82 | 2.085 | 22.6 | 7.72 |
| 10 | SilicoDArT | 9669953 | -5.857 | 22.8 | 7.80 | -2.095 | 22.4 | 7.66 |
| 10 | SilicoDArT | 5584917 | 6.512 | 28.9 | 10.00 | 2.073 | 22.3 | 7.62 |
| 10 | SilicoDArT | 2408858 | -5.816 | 22.6 | 7.72 | -2.084 | 22.3 | 7.61 |
| 10 | SilicoDArT | 7054499 | 6.179 | 25.9 | 8.89 | 2.066 | 22.2 | 7.57 |
| 10 | SilicoDArT | 25002574 | 6.455 | 28.2 | 9.70 | 2.070 | 22.1 | 7.56 |
1 Percentage variance accounted for
Table 6.
Characteristics and location of markers significantly related to plant resistance to Fusarium stalk rot.
Table 6.
Characteristics and location of markers significantly related to plant resistance to Fusarium stalk rot.
| Marker | Marker Type | Chromosome | Marker Location | Candidate Genes |
|---|---|---|---|---|
| 4772836 | SNP | Chr 2 | 1002990 | serine/threonine-protein kinase bsk3 |
| 9626410 | SilicoDArT | Chr 1 | 147389719 | 45560 bp at 5' side: uncharacterized protein loc100276421 isoform x1 74882 bp at 3' side: ubiquitin carboxyl-terminal hydrolase 15 isoform x1 |
| 5584917 | SilicoDArT | Chr 10 | 137279593 | uncharacterized protein loc103641914 isoform x1 uncharacterized protein loc103641914 isoform x2 |
| 157434 | SilicoDArT | Chr 2 | 210848643 | 14188 bp at 5' side: formin-like protein 11 23892 bp at 3' side: uncharacterized protein loc100273879 |
| 9698143 | SilicoDArT | Chr 6 | 168628814 | 440 bp at 5' side: uncharacterized protein loc100273604 86823 bp at 3' side: uncharacterized protein loc100382826 precursor |
| 2499631 | SilicoDArT | Chr 5 | 223354035 | 33662 bp at 5' side: putative protein of unknown function (duf640) domain fami 10443 bp at 3' side: nac domain-containing protein 92 |
| 21699135 | SilicoDArT | Chr 5 | 148291902 | 3967 bp at 5' side: uncharacterized protein loc103627005 11771 bp at 3' side: uncharacterized protein loc100279673 precursor |
| 7054095 | SilicoDArT | Chr 1 | 192290825 | 1016 bp at 5' side: abc transporter c family member 10 24169 bp at 3' side: probable lipoxygenase 8, chloroplastic |
| 4779143 | SilicoDArT | Chr 5 | 32830342 | 20659 bp at 5' side: putative disease resistance protein rga4 517 bp at 3' side: uncharacterized protein loc101027116 precursor |
| 4765764 | SNP | Chr 5 | 213770951 | histidine kinase 1 |
Table 7.
Sequences of designed primers used to identify newly selected markers significantly associated with plant Fusarium stalk rot resistance.
Table 7.
Sequences of designed primers used to identify newly selected markers significantly associated with plant Fusarium stalk rot resistance.
| Marker | Primer sequence | Melting temperature (°C) | Product size (bp) | |
|---|---|---|---|---|
| Forward | Reverse | |||
| 4772836 | GGTGGTTTTACCCCCTGCAG | TATTTGCAGGCCCTTGACCT | 59 | 281 |
| 9626410 | AGCAATTTCTCCAGAGTCTGATG | CATGCATTTTTCTGCATTGGGC | 61 | 50 |
| 5584917 | TATTGAAGAGAGATATGATAATCGCTGCAG | GTTCAAATAACTCGCAAAAGACTCG | 58 | 78 |
| 157434 | CGGACCGTATTACCCGGTTA | AATTTCCGCGGTACCGAGGC | 55 | 270 |
| 9698143 | ACCGTGGCTAATCCGGTTAT | GGCATTCCGGGTAATCCGTT | 59 | 350 |
| 2499631 | CGGTTCCAATTGGGATTACC | CCTGGACCGGCTTTACAATC | 61 | 145 |
| 21699135 | CCGATACTGCATGCTCTGCG | CCTCTGTTTGGCGTAGGTGA | 59 | 64 |
| 7054095 | GTCGACGACGAACCCTGCAG | CCAATATCCGGCGGACAGAC | 61 | 55 |
| 4779143 | AATACCCTGGGTCCGGTAA | TTACCGGGTCCAACCTGGC | 58 | 180 |
| 4765764 | TTTTTTCCTTCTTGCTGCAG | CCTCGTTCTGTGAACTGGAA | 61 | 263 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



