
Submitted:
25 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Helicobacter pylori infections are still an urgent and hot topic. This is related to the involvement of H. pylori in human diseases (including gastric diseases, gastric adenocarcinoma, mucosa-associated lymphoid tissue lymphoma, and diabetes), the number of which is constantly increasing. At the same time, agents effective against these bacteria are decreasing due to spreading resistance. Our research focused on extracts of the Rubus genus, commonly present in Central and Eastern Europe and very useful in folk medicine. Results have shown that the extract from Rubus idaeus 'Willamette' shoots, and the extract from R. idaeus 'Poranna Rosa' fruits and R. idaeus 'Laszka' R. occidentalis Litacz' fruits prevent the formation of biofilm by H. pylori on intravenous infusions (they reduce living cells, which was detected using triphenyl tetrazolium chloride (TTC) as a dye) at concentrations of 6.65 and 16.65 mg/mL, respectively. However, the living bacteria in the formed biofilm are affected by the extract from the shoots of R. idaeus 'Willamette' and the fruit of R. idaeus 'Poranna Rosa' at a concentration of 16.65 mg/ml, and the remaining extracts at a concentration of 33.3 mg/ml. The results of the study of interactions of extracts from Rubus sp. and the 2 antibiotics against H. pylori planktonic bacteria showed doxycycline synergism with all tested extracts, but only the extract of R. occidentalis 'Litacz' with levofloxacin. The remaining tested extracts showed indifference with levofloxacin. However, studies on the interaction between antibiotics and extracts of R. idaeus 'Willamette' shoot showed synergy with doxycycline and levofloxacin, additivity with amoxicillin and clarithromycin, and indifference with metronidazole. The tests were carried out on fragments of intravenous infusions with TTC as a dye. The tested extracts may be elements of broadly understood anti-H. pylori therapy.
Keywords:
antibacterial activity
; biofilm
; Helicobacter pylori
; plant extracts
; Rubus sp
; synergism
1. Introduction
Helicobacter pylori is a gram-negative, microaerophilic bacterium [1], that produces catalase, oxidase, and urease [2]. It is estimated that approximately 4.4 billion people have bacteria colonizing their stomach mucosa. H. pylori can cause a series of gastric diseases, including peptic ulcer disease, gastric adenocarcinoma, atrophic gastritis, and mucosa-associated lymphoid tissue lymphoma [3]. Three morphological forms of H. pylori have been described. Spiral forms are viable, culturable, and have a strong colonizing ability; coccoid forms are viable but not culturable (VBNC) and represent persistent forms of this bacterium; and degenerate forms are characterized by the breakdown of the cytoplasmic membrane and are most likely a manifestation of bacterial death [4]. The transition from the spiral to the coccoid form is caused by extreme conditions, e.g. changes in temperature, pH, or the presence of antibiotics [5]. There have been reports for a long time about the possibility of converting H. pylori from the coccoid to the spiral form, stimulated by an enriched environment [6,7].
It is commonly believed that H. pylori infections in humans are related to gastritis [8]. Bacteria in the stomach survive thanks to many virulence factors, including urease, vacuolating cytotoxin A (VacA), and spiral shape, and it is suspected that they also form a bacterial biofilm in the stomach [9]. But there are different routes of infection possible. There have been reports that bacteria have been isolated from dental plaque and drinking water sources [8]. Young et al. [10] reported that the spiral and viable coccoid forms of H. pylori are present in the oral cavity in the form of biofilm [2,8]. It has been proven also that bacteria can form biofilms in water systems and can survive from > 10 days (spiral form) to one year (VBNC coccoidal form) in fresh water [8]. Both places where bacteria are present outside the gastric mucosa are important from the point of view of both the spread of infections from one person to another and recurrent infections in people undergoing antibiotic therapy [8].
Biofilm is a population of bacteria attached to abiotic and biotic surfaces, surrounded on the outside by a polymeric substance. In the case of Helicobacter pylori, biofilm formation begins with a spiral form that attaches to the surface, and then morphological changes of the cells occur, creating various forms: spiral, rod-shaped, curved, coccoid, and filamentous. Finally, all cells transform into a coccoid form that is resistant to environmental factors [11]. Biofilm has tolerance of, or resistance to, antibiotics by expression of efflux pumps (transport proteins with roles in the export of toxic substances, including antibiotics) [1].
First-line antibiotics for the treatment of H. pylori infections are proton pump inhibitor, amoxicillin, and clarithromycin (may be replaced by levofloxacin) or metronidazole. It is recommended that the combination of drugs is used for 10–14 days. After failure, second-line therapy is introduced: proton pump inhibitor, bismuth, metronidazole, and tetracycline [12]; other introduced compounds include vonoprazan and rifabutin [3]. H. pylori resistance to antibiotics in Europe currently remains at a primary resistance level of 21.8% for clarithromycin, 15.8% for levofloxacin, and 38.9% for metronidazole. In the current situation, special attention should be paid to the appropriate level of H. pylori eradication, and alternative, effective methods of eliminating the pathogen should be proposed [13].
In accordance with WHO recommendations, new compounds that act on resistant bacteria are being sought. Extracts from plant raw materials containing secondary metabolites such as flavonoids, terpenes, and saponins offer hope for obtaining compounds with antibacterial properties. Research on plant raw materials to obtain anti-Helicobacter pylori compounds is being carried out by many research teams [11,12,14,15,16,17]. There are several reports regarding the effects of extracts from plants of the Rubus spp. genus on Helicobacter bacteria. Martini et al. [18] studied the effect of extract from Rubus ulmifolius growing in Italian forests on Helicobacter pylori, which showed anti-inflammatory, antiviral, and antimicrobial properties. Park et al. [19] studied the effect of Rubus crataegifolius extract in the treatment of impotence, inflammation, hepatotoxicity, enuresis, allergic diseases, and antibacterial properties. In turn, Krazue Baranowska et al. [20] and Krauze Baranowska et al. [21] studied the influence of raw fruit extracts from 3 cultivar varieties of Rubus idaeus 'Ljulin', 'Veten', and 'Poranna Rosa', one variety of black raspberry – Rubus occidentalis 'Litacz', and R. idaeus 'Willamette' shoot extract on many bacteria, including H. pylori. Raspberries are rich in phenolic compounds, mainly anthocyanins, ellagitannins, and other phenols, such as flavonoids and phenolic acids, including free ellagic acid conjugates named ellagitannins. The characteristic and predominant ellagitannin in Rubus berries is sanguiin H-6. However, the effects of the extracts of the plants mentioned above against Helicobacter pylori biofilm have not yet been investigated.
Some compounds of natural origin and their interactions with antibiotics used in chemotherapy against H. pylori have also been tested [13], but there are no studies on synergism with R. ideaus and R. occidentalis extracts.
In our publication, we present the results of research on the effects of selected fruit and shoot extracts against the biofilm produced by H. pylori. Additionally, we have developed a new model for testing H. pylori biofilm using intravenous infusions, in which we have shown the possibility of creating and examining such a biofilm.
2. Materials and Methods
2.1. Strains, Growth Medium, Antibiotics
Helicobacter pylori reference strain ATCC 43504. Brucella agar plates were used to grow bacteria in microaerobic conditions in GENbag microaer, BioMerieux (Marcy-l'Étoile, France). Brucella broth and brain heart infusion broth (BHI) (Becton Dickinson, Franklin Lakes, NJ, USA) supplemented with 5% horse serum was used in antimicrobial tests with tripticasein soy broth (TSB, Biocorp, Lublin. Polnad). Antibiotics used in this study include the following: clarithromycin (50 mg/mL, KRKA d.d. Novo mesto Slovenia), metronidazole (5 mg/mL, solution for infusion Polpharma, Starogard Gdanski, Poland) levofloxacin (5 mg/mL, solution for infusion, Pharmathen S.A, Pallini, Greece), ciprofloxacin (10 mg/mL, solution for infusion, KRKA d.d. Novo mesto Slovenia), doxycycline (20 mg/mL, solution for infusion, Polfa Tarchomin, Warszawa, Poland), and amoxicillin (100 mg/mL, Tarchomin, Warsaw, Poland). A polypropylene infusion set, sterile, non-pyrogenic, and nontoxic (Magromed, Lublin, Poland) was used. For biofilm studies, we used Brucella and BHI broth supplemented with 5% horse serum (GrasoBiotech, Starogard Gdanski, Poland) with or without 0.1% β-cyclodextrin (Merck Life Science Sp. z. o.o., Poznan, Poland). The colorimetric tests for the estimation of metabolic activity of cells, resazurin, MTT, and TTC (Merck Life Science Sp. z. o.o., Poznan, Poland) markers were used. It is used as an oxidation-reduction indicator in cell viability assays for both aerobic and anaerobic respiration. All the tests were incubated at 37 °C.
2.2. Preparation of Extracts
2.3. Broth Microdilution Assay
Microbroth dilution method assay was performed as per the recommendations of the European Committee on Antimicrobial Susceptibility Testing – EUCAST [22]. The minimal inhibitory concentration (MIC) of extracts and antibiotics was identified, and the results are published in Krauze-Baranowska et al. [20] and [21]. Dry test samples were dissolved in water to a final concentration 512 µg/mL. After dilution in BHI or Brucella broth with 5% horse serum, the final concentration of the extracts used for the testing of antimicrobial activity ranged from 128 to 0.00625 µg/mL. After incubation, 30 µl 0.2% MTT and resazurin were added to the samples, and then visually determination of MIC was performed.
2.4. Microtiter Biofilm Formation
The method was described by Chen et al. [23] with modification. H. pylori bacteria were grown in BHI or Brucella broth supplemented 5% horse serum, at 37°C for 72 hours with shacking. 30 µL of bacteria and 190 µL of broth (BHI or Brucella) were added to 96-well microtiter plates and grown in GENbag microaer without shaking. After 72 hours of incubation, 20 µL 0.2% MTT and 1% TTC were added followed by incubation for a further 2 hours at 37°C. The stained biofilm was washed 3 times with PBS and immobilized by adding 0.5% formalin. The biofilm was dissolved (DMSO from MTT, ethanol from TTC), and the OD (OD 550 nm from MTT and OD 495 from TTC) was measured using a microplate reader (Infinite®200 PRO, Tecan, Männedorf, Switzerland).
2.5. The Effect of Antibiotics on Biofilm Formed on Microtiter
H. pylori bacteria were grown in BHI or Brucella broth supplemented with 5% horse serum for 72 hours at 37°C with shaking. 30 µL bacterial and 190 µL broth (BHI or Brucella) were added to 96-well microtiter plates and grown in GENbag microaer without shacking. After 72 hours of incubation, extracts (33.3, 16.66, 6.65, 3.33 mg/mL) and antibiotics (512, 256, 128, 64, 32, 16, 8, 4, 2, 1 µg/mL) were added, and bacteria were grown in GENbag microaer without shacking. After 72 h of incubation, 20 µL 0.2% MTT was introduced to the studied samples and incubated for 2 hours at 37°C. The stained biofilm was washed 3 times with PBS buffer and immobilized in the 96-well microtiter plates by adding 0.5% formalin. The biofilm was dissolved by DMSO, and OD 550 was measured from a microplate reader (Infinite®200 PRO, Tecan, Männedorf, Switzerland).
2.6. Quantitation of Biofilm Formation on Glass Coverslips
The method was described by Cole et al. [24] with some modification. H. pylori bacteria biofilm was formed on 24 × 24 glass coverslips in 60 mm sterile plastic plates into Brucella broth or BHI broth with 5% horse serum. The media containing 0.1% β-cyclodextrin was incubated in GENbag microaer. Quantitation biofilm bacteria was performed with 1% TTC, 0.2% MTT, and 0.1% crystal violet, according to Bugali et al. [25]. All dye associated with the biofilms was dissolved with 1 mL solvent (MTT and crystal violet in ethanol, TTC in DMSO), and 200 µL of the solution was used to measure the absorbance with a microplate reader to determine the amount of biofilm formation.
2.7. Biofilm Formation on Fragments of an Intravenous Infusion
The bacteria were grown for 72 hours, with shaking, in glass tubes, in 5 mL broth, under paraffin to limit the access to oxygen. The broths used in the studies were as follows: TSB, BHI, Brucella broth, with or without 5% sheep blood, or 5% horse serum, or 0.1% β-cyclodextrin. All tubes contained 2–3 fragments (2 cm) of infusions. After 48 hours of incubation, 1% TTC was added to glass tubes with any type of culture broth and further incubated for up to 72 hours. To another set of culture, after 72 hours, 30 µl of 0.2% MTT was added and incubated at 37°C for 2–3 hours. After incubation the broth was removed from the tubes, and fragments of intravenous infusion were washed 3 times in PBS. Changes in the color of the intravenous infusion into purple (MTT) or red (TTC) revealed the formed biofilm. Fragments of intravenous infusion in the tested broths without bacteria were used as controls and treated as test samples. All dye associated with the biofilm was dissolved with 1 mL DMSO, and 200 µL of the solution was used to measure the absorbance with a microplate reader to determine the amount of biofilm formation.
2.8. Study of the Effect of Extracts on Biofilm Formation of Intravenous Infusion
The bacteria were grown in glass tubes in 5 mL broth under paraffin, to limit the access to oxygen, for 72 hours, with shaking. The H. pylori grew on the BHI broth supplemented with 5% horse serum. All tubes contained 2–3 fragments (2 cm) of infusions and antibiotics: doxycycline, levofloxacin (concentration in the range from 128 to 0.5 µg/mL), amoxicillin, clarithromycin, and metronidazole (concentration in the range from 128 to 32 µg/mL) or extract from Rubus sp. (concentration in the range from 6.66 to 0.8 mg/mL). After 48 hours of incubation 1% TTC was added to glass tubes with any type of culture broth and further incubated for up to 72 hours. To another set of culture, after 72 hours, 30 µl 0.2% MTT was added and incubated for 2–3 hours at 37°C. After incubation the broth was removed from the tubes, and the fragments of intravenous infusion were washed 3 times in PBS. A change in the color of the intravenous infusion into purple (MTT) or red (TTC) showed that the biofilm had formed. Fragments of intravenous infusion in the tested broths without bacteria were used as a negative control and treated as test samples.
2.9. Checkerboard Assays for Planktonic Bacteria
The checkerboard assay method was described by Pillai et al. [26]. In the research BHI broth supplemented with 5% horse serum was used. The antibiotics levofloxacin and doxycycline in the concentration range from 0.5 to 0.016 µg/mL and plant extracts with concentrations ranging from 33.3 to 3.33 mg/mL were tested. Bacteria were grown for 72 hours in microaerobic conditions. After incubation, 30 µl 0.2% MTT and 1% TTC were added to the samples; the reading of the results was carried out using an Infinite 200 PRO Tecan after 2 hours of incubation. After incubation, the turbidity of the samples was visually determined, and the results were read. The MICs of the antibiotic and the test compound were determined and substituted into the formula to calculate the Fractional Inhibitory Concentration Index (FICI):
where MICA and MICB are the MICs of drugs A and B alone, respectively. A and B are the concentrations of the drugs in combinations, respectively. FIC (index) values were interpreted as follows [27]: FIC index ≤ 0.5 indicates synergism; FIC index > 0.5 to < 1.0 shows partial synergism; FIC index = 1.0 indicates addition; FIC index > 1.0 to –4.0 denotes indifference; and FIC index > 4.0 denotes antagonism [28].
FICI = FICA + FICB = A/MICA + B/MICB,
2.10. Checkerboard Assays for Biofilm Formation on Fragments of Intravenous Infusion
The bacteria were grown in glass tubes in 5 mL broth under paraffin, to limit the access of oxygen, for 72 hours with shaking. BHI with 5% horse serum broths were used in the studies. All tubes contained 2–3 fragments (2 cm) of infusions. The following antibiotics were used: amoxicillin, clarithromycin metronidazole, levofloxacin, and doxycycline at concentrations ranging from 128 to 0.125 µg/mL; the plant extract concentrations ranged from 33.3 to 0.8 mg/mL. After 48 hours of incubation 1% TTC was added to glass tubes with culture broth and further incubated for up to 72 hours. After incubation the broth was removed from the tubes, and fragments of intravenous infusion were washed 3 times withs PBS. Changes in the color of the intravenous infusion into red showed the biofilm formed. Fragments of intravenous infusion in the tested broths without bacteria were used as a negative control and treated as test samples. All dye associated with the biofilms was dissolved with 1 mL DMSO, and 200 µL of the solution was used to measure the absorbance with a microplate reader to determine the amount of biofilm formation.
2.11. Scanning Electron Microscope (SEM)
The potential effect of extracts on the biofilm formed on the surface of intravenous infusion was determined with a SEM according to the procedure described by [29]. A scanning electron microscope JSM-6010LA by JEOL (Akishima, Tokyo, Japan), equipped with an energy-dispersive X-ray spectrometer EDX, was used.
2.12. Statistical Analysis
All experiments were performed at least 2 times. The intergroup differences were estimated by one- or two-way analysis of variance using Microsoft Excel 2010. All data were additionally analyzed by STATISTICA ANOVA v. 13.3. The distribution of normality of continuous variables was calculated using the Shapiro-Wilk test. The data are presented as mean and standard deviation (± SD). A p value was considered as statistically significant when it was less than 0.05.
3. Results
3.1. Extracts Preparation and Phytochemical Analysis
3.2. Antimicrobial Activity of Antibiotics and Extracts
Minimum inhibitory concentration (MIC) was defined as the lowest concentration inhibiting bacterial visible growth read visually or using a microtiter reader after change of colors of MTT and resazurin, which reflected the activity of the extracts, as shown in Table 1.
The results showed identical MIC values for the tested antibiotics, despite the different media and dyes used in the assay. The comparison of methods showed that the use of Brucella broth with 5% horse serum and the resazurin gives the weakest signal in the remaining samples. Based on the results, Brucella broth and MTT dye (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) were selected for the next stage of brain heart infusion (BHI) research.
3.3. Microtiter Plate Biofilm Formation. Study of the Effect of Extracts on the Produced Biofilm
H. pylori biofilm detection is mainly limited to checking its level using crystal violet. But the main goal of research should be the killing of bacteria in the biofilm or the prevention of its formation. For the first time (as far as we know), we used the TTC marker and BHI medium with horse serum to study H. pylori biofilm.
A comparison of the obtained results is shown in Figure 1.
The study showed that both media allowed biofilm formation in the wells of the plate within 72 hours. Higher detectability of live bacteria present in biofilms was noted with the MTT dye. The OD value corresponding to the biofilm formed in Brucella medium and the selected MTT is twice as large as the other values. This means that in a 96-well titration plate, in the presence of Brucella broth, H. pylori can form a biofilm with more viable cells than in BHI broth. The sample marked in the Figure 1 as TTC Brucella, gives an OD value 2 times lower than the sample marked as MTT Brucella; these are samples of the same substrate labeled with a different marker. This means that TTC labels live cells less effectively than MTT on the titration plate. The use of both markers in detecting the amount of live biofilm cells in the BHI broth were at a similar level (sample 1 marked as MTT BHI and sample 3 marked as TCC BHI).
To check the effect of extracts on the created biofilm, BHI and Brucella broth with 5% horse serum was used with MTT as the label. The results are shown in Figure 2a,b.
To check the effect of extracts on the created biofilm, BHI and Brucella broth with 5% horse serum and MTT were used as the dye. The results are shown in Figure 2a,b. Analyzing the relationship in Figure 2a, it can be noticed that with the increase in the concentration of the extracts examined, the bactericidal properties increase. The extract from Rubus idaeus ‘Willamette’ shoot at the concentration of 3.33 mg/mL turned out to be the most active, killing 50% of the living cells compared to the control. At the same concentration, the extract from Rubus occidentalis ‘Litacz’ fruits was found to be the least active, reducing living cells only by 10%. The concentration of 16.6 mg/mL extract from Rubus idaeus ‘Willamette’ shoot killed 99% of the H. pylori cells, while the remaining extracts show this activity at a concentration twice as high. BHI medium with the addition of 5% horse serum promotes the maintenance of the formed biofilm, but its level is 2 times lower than in the presence of Brucella medium with the addition of 5% horse serum. At a concentration of 3.33 mg/mL, the most active were the extracts: Rubus idaeus ‘Willamette’ shoot, Rubus idaeus ‘Poranna Rosa’ fruits and Rubus occidentalis ‘Litacz’ fruits, causing a reduction of live bacteria of about 50% in relation to the control. On the other hand, about 80% reduction of live bacteria was demonstrated by R. idaeus ‘Willamette’ shoot, R. idaeus ‘Poranna Rosa’ fruit, and R. idaeus ‘Laszka’ fruit, at a concentration of 33.3 mg/mL. Fruit extracts R. idaeus ‘Poranna Rosa’ and Rubus occidentalis ‘Litacz’ at a concentration of 6.66 mg/mL decreased the number of viable cells in the biofilm formed by about 50% relative to the control.
3.4. Quantitation of Biofilm Formation on Glass Coverslips
The gold standard in H. pylori biofilm research is glass coverslip testing using crystal violet, Brucella broth, and β-cyclodextrin. In our research, we wanted to compare the efficiency of biofilm formation using two different substrates along with the detection of live bacterial cells as an indicator of the amount of biofilm formed. Therefore, determination of the amount of biofilm formed on glass coverslips in BHI and Brucella broth, with the addition of 5% horse serum and in the presence or absence of β-cyclodextrin, using 3 dyes: MTT, CV, and TTC, was performed.
The results are shown in Figure 3.
The level of biofilm formation on glass coverslips in BHI and Brucella medium with 5% horse serum, in the presence or absence of β-cyclodextrin, was evaluated. The dyes used to visualize the biofilm level were MTT, crystal violet, and TTC. The biofilm was grown without shaking for 3 days. The highest level of biofilm (with live cells) was obtained when cultured in BHI and Brucella broth with the addition of 5% horse serum and 0.1% TTC-labelled β-cyclodextrin. We have shown that both media with β-cyclodextrin allow the formation of a biofilm, but only TTC showed how many live bacteria can be found there. The TTC level significantly exceeded that achieved by crystal violet.
3.5. Biofilm Formation on Fragments of an Intravenous Infusion
To demonstrate the possibility of biofilm formation on surfaces made of plastic, experiments in which H. pylori bacteria were cultured in the presence of fragments of intravenous infusions in 5 mL broth under paraffin for 72 hours with shaking were performed. The BHI and Brucella broth, with or without 5% horse serum or 0.1% β-cyclodextrin, were used. The presence of biofilm was checked after 3 and 6 days. The results are presented in Figure 4.
In all samples tested, the bacteria formed a biofilm. Significantly more biofilm was formed in the presence of BHI than in Brucella broth. Incubation of samples for 3 or 6 days resulted only in significant growth of biofilm (live bacteria) in BHI broth with horse serum and β-cyclodextrin (bacteria were detected by TTC). A large amount of biofilm was also detected in BHI medium containing horse serum, in which the number of viable cells was measured using TTC after 3 days of incubation. Comparing the results presented in Figure 3 and Figure 4, it can be concluded that bacteria form biofilms faster on plastic surfaces than on glass surfaces, in the absence of β-cyclodextrin and serum. Biofilm in BHI broth is detected using MTT and TTC at a similar level, but more live cells are detected in the biofilm after 72 hours by TTC.
3.6. Study of the Effect of Extracts on Biofilm Formation on Intravenous Infusion
To study the effect of extracts on the formation of biofilm on fragments of intravenous infusions, BHI broth with the addition of 5% horse serum was used. Samples were incubated in tubes under paraffin at 37°C for 72 hours. MTT was used to detect the biofilm, and the absorbance was measured at 550 nm. The results are shown in Figure 5.
Figure 5 shows that extracts at a concentration of 3.33 mg/mL reduce the amount of living H. pylori cells in the formed biofilm. Strong inhibitory properties can be observed at a concentration of 6.66 mg/mL for Rubus idaeus ‘Willamette’ shoot, while other extracts from fruits inhibit biofilm formation from a concentration of 16.65 mg/mL.
3.7. Study of the Effect of Antibiotics or Extracts on Biofilm Formed in Intravenous Infusions
To check the effect of extracts on the biofilm formed on fragments of intravenous infusions, the technique for research on biofilm formation after 144 h was used. The results are shown in Figure 6.
All extracts at concentrations of 33.3 and 16.65 mg/mL kill most of the bacterial cells in the 144-h biofilm formed by Helicobacter pylori. Lower concentrations (6.66 and 3.33 mg/mL) reduce the living bacterial cells in the biofilm by approximately 40%. Only the extract from Rubus idaeus "Laszka" fruits in concentrations of 3.33 and 6.66 causes a reduction of living cells by 20% compared to the control.
3.8. Checkerboard Arrays on Planktonic Cells and Biofilm Formation
Nowadays, antibiotic therapy of infection caused by H. pylori more often ends in failure due to the bacteria's resistance to antibiotics. It seems logical to check what interactions occur between antibiotics and the tested extracts.
The results for the study of interactions on planktonic bacteria are presented in Table 2.
We used all extracts and only the 2 most active antibiotics against our H. pylori strain (Table 2). All the tested extracts showed synergism with doxycycline, and only the extract of Rubus occidentalis 'Litacz' showed synergism with levofloxacin. The remaining extracts tested showed indifference with levofloxacin.
To compare the results obtained for planktonic bacteria, we tested the interactions of antibiotics and the extract, as shown in Table 3. We conducted the research on fragments of intravenous infusions.
Among the tested extracts, only the extract from Rubus idaeus 'Willamette' shoot (the most active) (Table 3) in combination with doxycycline and levofloxacin showed synergy. With amoxicillin it showed additivity and with clarithromycin and metronidazole, indifference. Similar studies should be carried out with the remaining extracts.
3.9. Scanning electron microscope (SEM)
The potential effect of extracts on the biofilm of an intravenous infusion using SEM. Photos taken using this technique are presented in Figure 7.
Figure 7a shows a clean sterile surface of the intravenous infusion. Figure 7b,c show a H. pylori biofilm on the intravenous infusion surface with coccoid cells, characteristic of a mature biofilm. Figure 7d shows the surface of an intravenous infusion incubated with a mixture of bacteria and extract (inhibition of biofilm formation).
Figure 7e,f show the remains of bacterial cells exposed to extract from Rubus idaeus 'Willamette' shoot at a concentration of 6.66 mg/mL. The presence of mature H. pylori biofilm visible in the photos (Figure 7b,c) confirms the TTC absorbance (OD 550) of H. pylori biofilm control and the presence of live bacterial cells in the biofilm after 72 hours, measured using TTC. However, the remains of bacterial cells constituting the biofilm visible in Figure 7d,e are the effect of the extract. The effect shown in Figure 7d,e confirms zero (close to zero) TTC absorbance in samples where the bacteria were killed by the tested extracts, in this case on the example of the extract from Rubus idaeus 'Willamette' shoot a concentration of 6.66 mg/mL.
4. Discussion
In combating H. pylori in the population, it is more important to detect the bacteria and kill them quickly, rather than inhibiting their growth. Therefore, research on the activity of compounds against H. pylori should include the detection of live bacteria.
Various teams studying H. pylori use media to test the activity of different compounds. Tests using the microdilution method (MIC determination) allowed us to demonstrate that Brucella broth with 5% horse serum and resazurin gives the weakest signal in the remaining samples. While the use of brain heart infusion (BHI), Brucella broth, and MTT dye gives clear test results. Further studies allowed us to demonstrate that the Brucella broth medium produces a higher percentage of live bacteria in the biofilm formed on the microtiter plate, which was detected using the MTT marker.
For the colorimetric tests for the estimation of metabolic activity of cells, resazurin and MTT markers were used. NAD(P)H-dependent cellular oxidoreductase enzymes can, under certain conditions, reflect the number of viable cells present. These enzymes are capable of reducing the tetrazolium dye 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to its insoluble purple-colored formazan. Resazurin (7-hydroxy-3H-phenoxazin-3-one 10-oxide) is a blue dye, itself weakly fluorescent until it is irreversibly reduced to the pink colored and highly red fluorescent resorufin. It is used as an oxidation-reduction indicator in cell viability assays for both aerobic and anaerobic respiration.
Chatterjee et al. [30] tested extracts of raspberry, cranberry, elderberry, strawberry, bilberry, blueberry, and OptiBerry ® (a blend of the 6 berries) on H. pylori strain ATCC 49503. They used tryptic soy broth (TSA) supplemented with 5% defibrinated sheep blood. All extracts at the 0.5 and 1% concentration inhibited growth of bacteria by > 70%.
Goodman et al. [31] analyzed commercially available black raspberry (Rubus occidentalis; BRB), blackberry (Rubus fruticosus; BB), and red raspberry (Rubus idaeus; RRB). H. pylori PMSS1 was cultured on Brucella broth supplemented with 10% FBS, and it was visualized with a proprietary redox-sensitive tetrazolium dye (2,3,5-triphenyl tetrazolium chloride [TTC]). Assays were performed in 96-well plates using an OmniLog (Biolog, Hayward, CA, USA) plate reader-incubator. All berry preparations tested had antibacterial activity with complete inhibition of H. pylori growth at a concentration of about 4%.
Martini et al. [18] determined the activity of plant leaves from Rubus ulmifolius. The studies were performed in microtiter plates using Brucella broth–bovine fetal serum, and H. pylori G21 and 10K strains were tested. They showed that all the isolated polyphenols (ellagic acid, gallic acid, quercetin) and extracts had antibacterial activity against both of the H. pylori strains. The MBC values of the extract, after 24 h of exposure, against H. pylori G21 and 10K strains were 1200 µg/mL and 1500 µg/mL, respectively. In turn, after 48 h of exposure, the MBC values were 134 µg/mL and 270 µg/mL, respectively.
Park et al. [19] determined minimum inhibitory concentrations using Brucella media containing 10% FBS in the Petri plates. Agar plates contained 0, 10, 50, 75, 100, 150, and 200 μg/mL of plant extract: Rubus crataegifolius (RF, Family: Rosaceae) or Ulmus macrocarpa (UL, Family: Ulmaceae). Tests were performed on 2 reference strains of Helicobacter pylori (ATCC: 43504, SS1) and 11 clinical isolates. RF extract inhibited growth of H. pylori at 150 μg/mL, whereas UL showed MIC50 at 200 μg/mL.
Nakase et al. [32] tested modified Gingyo-san, several Chinese herbal medicines, and Kampo medicines using Mueller-Hinton broth (CAMHB) supplemented with 10% inactivated FBS, and 25 µg/mL and 10 µg/mL of Ca2+ and Mg2+, respectively. Incubation was performed in microaerobic conditions at 37°C for 5 days. The MIC values of the tested extracts against the H. pylori strains were in the range of 512 to > 8192 µg/mL.
Trung et al. [33] tested the effects of naringenin (NRG), luteolin (LUT), myricetin (MCT), and protocatechuic acid (PCA) identified in the Hibiscus rosa sinensis flower against 2 reference strains and 5 clinical isolates of Helicobacter pylori. MIC was determined by serial dilutions of concentrations of the test compounds in 10% NBS (newborn bovine serum)-supplemented Brucella broth in sterile 96-well plates using resazurin. The MIC values for NRG, LUT, MCT, and PCA were as follows: 100, 125, 150, and 250 mg/L, respectively.
Krzyżak et al. [9], testing the MICs of myricetin (MYR), used flat-bottomed, ventilated 12-well microtiter plates. MYR comprised a series of dilutions, which were made in brain heart infusion broth supplemented with 7% fetal calf serum. The MIC was taken to be the lowest concentration in which no microbial growth was observed visually after a 3-day microaerophilic incubation at 37°C. The MIC for both tested strains (H. pylori J99 and Tx30a) were determined in a concentration of 160 µg/mL. The compound was dissolved in DMSO, hence its good penetration through bacterial membranes. The viability of H. pylori was examined by staining bacterial cells with the LIVE/DEAD kit and analyzing by fluorescence microscopy.
Chen et al. [23] determined the MIC values of clarithromycin (CLR), amoxicillin (AMX), and rhamnolipid (RHL) by broth dilution assay using BHI broth containing 2% FBS in 96-well microtiter plates. Samples were incubated at 37°C for 48 h in microaerophilic conditions with 200 rpm shaking, and then the OD600nm was measured by microplate reader. The obtained MIC values were as follows: MICs90 was 0.96 μg/mL for CLR, 0.06 μg/mL for AMX, while the MIC90 of RHL was 307.2 μg/mL.
For minimum inhibitory concentration (MIC) determination according to Sposito et al. [34] MHB supplemented with 50% BFS was added to each well of a 96-well microplate; H. pylori ATCC 43504 was used for the tests. The essential oil, ethanolic extract, fractions, sub-fractions, casearin J, and hydrolyzed casearin J from C. sylvestris leaves were tested. For visual reading of the bacterial strains, 0.01% of resazurin aqueous solution was added to each microplate well. The MIC of the ethanol extract and fractions showed MICs ranging from 62.5 to 1000 µg/mL. The ethanol extract was fractionated into 63 subfractions, casearins were detected in fractions marked SF20, SF13, SF46, and SF17, and the range of MIC values was from 250 to > 1000 µg/mL. When tested as a standard, Cas J (casearin J) showed weak antibacterial activity, and the essential oil itself showed an effect of 125 µg/mL.
Research on the H. pylori biofilm mainly includes tests performed on titration plates or coverslips using Brucella broth fetal calf serum (FCS). First, we wanted to investigate which substrate and dyes would more effectively determine live cells in biofilm grown on titration plates. BHI and Brucella broth with 5% horse serum were used, and the dyes were MTT and TTC. The use of TTC as a dye was due to scientific findings [35].
Triphenyl tetrazolium chloride (2,3,5-triphenyl-2H-tetrazolium chloride) (TTC) is a redox indicator commonly used in biochemical experiments, especially to indicate cellular respiration (for example, to check for the viability of seeds). The white compound is enzymatically reduced to red TPF (1,3,5-triphenylformazan) in living cells due to the activity of various dehydrogenases (enzymes important in the oxidation of organic compounds and thus cellular metabolism).
Our research, including testing in a 96-well titration plate in the presence of Brucella and BHI broth, and using TTC and MTT as dyes, showed that H. pylori can form a biofilm with more viable cells in Brucella than in BHI broth with the addition of 5% horse serum (Figure 1). Studies on the effect of extracts on the formation of biofilm by H. pylori in a 96-well titration plate demonstrated that the extract form R. idaeus ‘Willamette’ shoot at concentrations of 3.33 mg/mL and 16.6 mg/mL were the most active. While the remaining extracts showed lower activities, depending on the medium used (BHI and Brucella broth with 5% horse serum was used with MTT) relative to the control (Figure 2a,b). Our studies using coverslips for biofilm formation by H. pylori demonstrated that the highest level of live cells in biofilm is obtained when cultured in BHI and Brucella broth with the addition of 5% horse serum and 0.1% TTC-labelled β-cyclodextrin. Moreover, the obtained results showed that both media with β-cyclodextrin allow the formation of a biofilm, but only TTC shows how many live bacteria can be found there. The TTC level significantly exceeds that achieved by crystal violet.
Studies showing that H. pylori can form a biofilm on plastics (titration plates, pipes with treated water, on tissues) allow us to assume that bacterial cells may behave similarly if they encounter plastics in higher organisms, such as stents, prostheses, or central insertions. An additional element that helps create these formations is the presence of tissue fluids, including serum and the nutrients it contains. This led us to propose a different research model for H. pylori biofilms.
Studies on biofilm formation on the intravenous infusion fragments showed that the highest number of living bacteria in the biofilm, incubated for 3 days (6 days) in BHI broth with horse serum and β-cyclodextrin, were detected using TTC indicator. A large number of live bacteria were also detected in BHI medium containing horse serum, in which TTC was used after 3 days of incubation. Biofilm in BHI broth was detected using MTT and TTC at a similar level. However, more live cells were detected in the biofilm after 72 hours by TTC.
Our research on the influence of extracts on the formation of biofilm showed that concentrations of 6.66 and 16.65 mg/mL of the extracts from the shoots and fruit of R. idaeus 'Willamette', respectively, inhibit the formation of biofilm on fragments of the intravenous infusion. However, all extracts at concentrations of 33.3 and 16.65 mg/mL reduce the number of viable bacteria in the 144-hour biofilm formed by H. pylori. Based on these results, we tested the effect of Rubus sp. fruit and shoot extracts on biofilm formation and on the biofilm already formed by H. pylori. The research results were confirmed by SEM photos, in which the destructive effect of the R. idaeus 'Willamette' shoot extract on the biofilm was clearly visible. Our results from studies of interactions of extracts from Rubus sp. and only the 2 most active antibiotics on planktonic bacteria H. pylori showed synergism of all extracts with doxycycline, and only the extract of R. occidentalis 'Litacz' with levofloxacin. The remaining extracts tested showed indifference with levofloxacin. However, studies on the interaction of antibiotics and extracts of R. idaeus 'Willamette' shoot were carried out on intravenous infusion fragments and showed synergism with doxycycline and levofloxacin, additivity with amoxicillin, and indifference with clarithromycin and metronidazole.
Windham et al. [36] studied biofilm formation by H. pylori DSM-1 using various substrates: BB (Brucella broth) supplemented with 10% FBS (fetal bovine serum) and 10 µg/mL vancomycin, brain heart infusion medium supplemented with 0.1% β-cyclodextrin and 10 µg/mL vancomycin, and BB supplemented with 2% FBS, 10 µg/mL vancomycin, and 0.2% glucose. They tested different surfaces, and standard 24-well tissue culture-treated plates were replaced by either untreated or poly-D-lysine-treated 24-well plates. The studies were performed for the time points T24, T48, and T72. The researchers used crystal violet to test the number of bacteria in biofilm. They did not notice any differences in the formation of biofilm according to the tested strain after using different media with different additives. They indicated also that their observation of biofilm formation is not an artifact of medium choice.
Cole et al. [24] compared the number of bacteria in biofilm of 16 H. pylori mutants on coverslips using 3 methods. The first method included staining with carbolic fuchsin and observation under a light microscope, the second method was staining with the Live/Dead BacLight kit (Molecular Probes), and the third method was scanning electron microscopy (SEM). They checked the efficiency of biofilm formation of mutants (mutations in several genes, including the quorum-sensing ones) on a 12-unit microtiter using BHI broth containing 0.1% β-cyclodextrin (5 days of incubation) or 5% heat-inactivated (56°C, 20 min) fetal calf serum (at 3 to 4 days of incubation) with or without gentle shaking. They detected bacteria in the biofilm using MTT. The researchers found, in preliminary experiments, that incubation of H. pylori with MTT gave higher values and less background absorbance than other tetrazolium reagents in BHI broth.
Trung et al. [33] employed biofilm inhibition assay in 96-well plates on 10% NBS (newborn bovine serum)-supplemented Brucella broth by the crystal violet assay. Naringenin showed a reduction of H. pylori biofilm formation by 85.9 and 52.7% at concentrations of 50 mg/L and 25 mg/L, respectively. The use of myricetin resulted in a reduction in biofilm formation by 39.5% at a concentration of 75 mg/L, while luteolin and protocatechuic acid showed no effect.
Krzyżak et al. [9] used the following conditions to create biofilm: a BHI broth with 2% fetal calf serum, sterile 96-well polystyrene microtiter plates, a microaerophilic atmosphere, and 3 days incubation at 37°C with shaking (50 rpm). The crystal violet method was used to assess biofilm formation. The authors showed that exposure to 1/8 × MIC and 1/4 × MIC of myricetin resulted in a 50% and 70% reduction in the biofilm formation of H. pylori Tx30a and 60% and 70% decrease for H. pylori J99, respectively. The viability of H. pylori was examined by staining bacterial cells with the LIVE/DEAD kit and analysis by fluorescence microscopy.
Results obtained by Spósito et al. [34] showed that the incorporated and non-incorporated ethanolic extract of C. sylvestris leaf, in the nanostructured lipid system, inhibited H. pylori biofilm formation at the assayed concentrations, except for the extract incorporated at 78.12 μg/mL. At lower concentrations, the incorporated ethanol extract had better activity than the unincorporated extract (625, 312.5, and 156.2 µg/mL) and inhibited a higher percentage of biofilm at the same concentration. Although C. sylvestris extracts inhibited H. pylori biofilm formation, none was superior to amoxicillin, which inhibited a greater percentage of biofilm at a lower concentration.
Krzyżek et al. [11] assumed that the formation of a biofilm by H. pylori depends on the flow speed of the medium (which is inversely proportional to adhesion), surface charge, and pH of the solution in which the biofilm is formed. The authors stated that the minimum serum concentration necessary for biofilm formation is 2%, and the maximum amount of biofilm to be tested in culture is one cultured for 72 hours; extending the incubation time may even result in its reduction.
Hathroubi et al. [37] investigated the activity of amoxicillin, clarithromycin, and tetracycline on H. pylori wild type, clinical strains, and several mutants using Brucella broth (Difco) containing 10% fetal bovine serum (BB10; Gibco/BRL). Bacteria were grown under microaerobic conditions (10% CO2, 5% O2, 85% N2) at 37°C. Biofilms were incubated for 2 days on a sterile 96-well polystyrene microtiter plate and then marked by crystal violet. To determine the number of viable cells in planktonic cultures, H. pylori samples incubated overnight or for 2 or 3 days were plated for enumeration by counting CFU.
Chen et al. [23] showed that clarithromycin could inhibit approximately 90% of H. pylori biofilm formation at MIC concentrations of 0.03 µg/mL and 0.48 µg/mL (1/2 MIC). Amoxicillin can inhibit less than 50% of biofilm formation at 0.03 µg/mL (1/2 MIC), and RHL can inhibit more than 90% of biofilm formation at 1/2MIC – 76.8 µg/mL. The inhibitory effect of clarithromycin and amoxicillin on biofilm formation increases with increasing rhamnolipid concentration up to 76.8 μg/mlL. Chen et al. [23] studied the synergy of 2 antibacterial agents using 96-well microtiter plates as a checkerboard. After incubation, OD600nm measurement was performed, and a synergy effect between CLR, AMX, and RHL in inhibiting the growth of H. pylori planktonic cells was found, i.e., an inhibitory fraction concentration (FIC). The determined FIC values for CLR and AMX, RHL and CLR were 0.75, indicating an additive effect. However, the combination of RHL and AMX did not show a stronger effect than the individual compounds alone, with an FIC value of 1.0. Chen et al. [23], testing H. pylori biofilm formation, performed incubation in 48-well microtiter plates at 37°C for 24 h, in static microaerophilic conditions. Biofilm biomass was evaluated by crystal violet test and LIVE/DEAD BacLight Bacterial Viability kits (Invitrogen, USA) were used to evaluate the viability and architecture of the biofilm. Active concentrations towards the formed biofilm were 61.44 μg/mL for clarithromycin (CLR) and 122.88 μg/mL for amoxicillin (AMX); CLR and AMX could eradicate about 50–60% of H. pylori biofilm. CLR + AMX eradicated mature biofilm. The rhamnolipid (RHL) alone eradicated mature H. pylori biofilm at a concentration of 1228.8 μg/mL. Moreover, RHL in combination with AMX eliminated over 90% of H. pylori biofilm, which was better than the combination of RHL + CLR + AMX. Lansoprazole (used as PPI) + RHL + AMX resulted in a 95% reduction of H. pylori biofilm.
Nakase et al. [32] studied the interactions of modified Gingyo-san and clarithromycin, amoxicillin, metronidazole, minocycline, and sitafloxacin against reference (ATCC 700392) and 2 clinical strains of planktonic H. pylori at pH 7.0 and 5.0. Two types of interactions were found: additive and indifferent; the pH slightly influenced the changes in interactions.
Krzyżek et al. [9] investigated interaction activity of myricetin (MYR) with the classically used antibiotics amoxicillin, clarithromycin, tetracycline, metronidazole, and levofloxacin. They used the checkerboard assay. Three selected concentrations of MYR (1/4 ×, 1/8 ×, and 1/16 × MIC) and antibiotics at the range of concentrations 0.0015 to 16 μg/mL were incubated in 1 mL of BHI broth with 7% serum at 37°C for 72 h, in microaerophilic conditions, with shaking (100 rpm). Their research results showed that 1/4 × MIC of MYR is able to decrease the MICs of all tested antibiotics by 4–16 times its potential to synergistically enhance the action of the antibiotics ( FICI = 0.31–0.5). The 1/8 × MIC of MYR also increased the activity of all tested antibiotics; most of the interactions were additive (FICI = 0.625). The 1/16 × MIC of MYR additively increased only the activity of amoxicillin, metronidazole, and levofloxacin (FICI = 0.56), but not clarithromycin and tetracycline (FICI = 2.0). The viability of H. pylori was examined by staining bacterial cells with the LIVE/DEAD kit and analyzing them by fluorescence microscopy.
Park et al. [19] studied the interaction of Rubus crataegifolius (RF) and Ulmus macrocarpa (UL) extracts in concentrations of 150 and 200 µg/mL, respectively, using the in vitro method from dilutions in agar. UL extract did not show any inhibition at 150 μg/mL, RF or UL did not show anti-H. pylori effect at 50 or 75 μg/mL, while RF in combination with UL at 75 and 50 μg/ml showed a strong synergistic effect. Combination of RF and UL at 75 μg/mL each showed complete inhibition of H. pylori colonization. All 11 H. pylori clinical strains and 2 reference strains were tested for the synergistic effect of RF and UL on amoxicillin, clarithromycin, and omeprazole and showed similar inhibitory effects on H. pylori colony formation. Park et al. [19] studied an extract from R. crataegifolius that contained polyphenols including sanguine, coreanoside-F1, nigaichigoside, gallic acid, and ellagic acid. According to the authors, the strong anti-H. pylori properties of the extract were indicated by the presence of ellagic acid (14.2 mg/g of dry extract) and catechin (30.5 mg/g of dry extract).
Krzyżek et al. [11] cited that the presence of H. pylori in the biofilm in coccoid forms only indicates a mature form of the biofilm created by this bacteria. This form results in physiological changes in the bacteria, including tolerance to variable environmental factors. Transformation may lead to the development of a viable but non-culturable (VBNC) phenotype by spherical H. pylori forms, but still metabolically active.
Martini et al. [18] studied the effect of the extract from R. ulmifolius and the isolated compounds ellagic acid, gallic rutin (quercetin-3-rutinoside), and kaempferol — effective anti-H. pylori agents. Rutin (quercetin-3-rutinoside) is known to possess antimicrobial activity against some gram-positive and gram-negative bacteria, and ellagic acid kills H. pylori by inhibiting arylamine N-acetyltransferase activity. Kaempferol induces a significant decrease in the number of colonies of H. pylori in gerbils’ stomachs after oral treatment. They hypothesize that the target of the antibacterial action of R. ulmifolius and its extracts, which are polyphenols, could be one or more bacterial ion pumps. To support such conjecture, they compared the structure of the gastric proton pump with those of H. pylori ion pumps and found significant linear homologies. The researchers suggested that blackberry polyphenols kill H. pylori because of inactivation of its ion pumps, i.e., enzymes that regulate the flux of copper and metal cations through membranes. The authors noted that the extract appears to be much less effective after 24 h of exposure than the isolated compounds. This behavior, which is more evident after 24 h of incubation, may be explained by the low concentration of single compounds in the extract and the presence of other constituents that may lack antibacterial activity.
Studies conducted by Krazue Baranowska et al. [20,21] on the planktonic form of H. pylori showed the activity of ellagic acid against the cells of these bacteria (MIC 0.125 mg/mL). In contrast, dimeric sanguine H-6 had no antimicrobial activity against H. pylori. Researchers cited the view that among hydrolysable tannins only monomeric forms are the most effective and promising H. pylori inhibitors. The inhibiting effect of raspberry fruit extracts against H. pylori was on the same level for all the analyzed varieties (MIC 8 mg/mL). The inhibitory activity of the fruit extracts towards H. pylori may be connected with their content of free ellagic acid. However, the extract from R. occidentalis ‘Litacz’ was found to contain about 5 times more anthocyanins than the fruit extracts of red ‘Ljulin’ and ‘Veten’. In turn, in the fruits of R. occidentalis ‘Litacz’, R. idaeus ‘Ljulin’ and ‘Veten’ the cyanidin glycosides dominated. The fruits of ‘Litacz’, ‘Poranna Rosa’, and ‘Ljulin’ have similar levels of sanguiin H-6, but the ‘Veten’ has 3 to 5 times higher concentrations of sanguiin H-6. A similar situation is the content of free ellagic acid in fruits extracts from ‘Litacz’‘Poranna Rosa’, and 'Ljulin' varieties, in which the discussed compound occurs at the same level, and in 'Veten' it was approximately 2 times higher. The extract from R. idaeus ‘Willamette’ shoot contained about 5% sanguiin H-6 and about 1% free ellagic acid [21]. According to the authors, ellagic acid is mentioned among the compounds that might contribute to the antimicrobial activity of raspberries. It has been proposed that by binding to the bacterial membrane, ellagic acid can destabilize its structure, thus disrupting cell functionality. The authors cited research to support this theory, in which alcoholic beverages enriched with raspberries had a greater potential against H. pylori than pure alcohol. The authors suggested that this effect is due to raspberry extracts damaging bacterial cell membranes and thus making them more sensitive to alcohol. The researchers concluded that the raspberry shoot extract (MIC for H. pylori – 7.4 mg/mL) is a prospective source of sanguiin H-6 and ellagic acid; these 2 components are believed to be responsible for its antimicrobial properties.
In the presented studies, the extract from R. idaeus ‘Willamette’ shoots transpired to be more active in preventing biofilm formation on intravenous infusions compared to fruit extracts. Perhaps antibacterial activity was demonstrated not only by sanguiin H-6 and free ellagic acid, but also by isoquercetin, quercetin 3-Oglucuronide, epicatechin, and catechin contained in this extract. However, on the already formed biofilm on fragments of the intravenous infusion, the following agents had the best effect; for the fruit extracts from R. idaeus 'Poranna Rosa' and extract from R. idaeus 'Willamette' shoots, the activity was at the same level. In this case, in our opinion, the main role against H. pylori biofilm was played by sanguiin H-6 and free ellagic acid, which are the only biologically active compounds detected so far in fruit extracts from ‘Poranna Rosa’, and the extract from the shoot contains a high concentration of them. It would be worth examining the mechanism of action of both compounds on bacteria.
There are various theories cited by Krauze-Baranowska et al. [20] regarding the absorption of anthocyanins from the gastrointestinal tract, suggest that they are poorly absorbed and excreted unmetabolized. Ellagitannins are reported not to be absorbed in the intestine, but instead are metabolized by colonic microflora to form urolithins. Therefore, the authors believe it is difficult to predict and estimate whether the compounds will be able to reach the infection sites at a desired concentration.
Krauze-Baranowska et al. [21] proposed the use of raspberry extracts, e.g. in the form of lozenges, to prevent or limit infections with various pathogens of infectious diseases of the respiratory and gastrointestinal tract. An advantage that should be added here is wide access to the fruits of various varieties of raspberry plants consumed as part of the daily diet in various forms. But it should also be noted that the current active compounds isolated and identified in extracts from raspberry fruit, leaves, and shoots may provide a starting point for the synthesis of derivatives with much greater antibacterial properties. The synergism of extracts form Rubus sp. fruit with doxycycline and R. idaeus 'Willamette' shoot with doxycycline and levofloxacin, as well as the additivity of amoxicillin, which we have demonstrated, may provide a basis for the use of extracts in combination with antibiotics in clinical trials to combat H. pylori infections. In our research, we have shown that H. pylori can form a biofilm on medical materials used in the treatment of patients. These results suggest that the presence of artificial materials in the human body, e.g., stents, may make it possible to colonize them with H. pylori. Additionally, the use of treated water, used, e.g., for artificial respiration in hospitals, in which H. pylori has already been detected, in our opinion, expands the ways of spreading H. pylori infections. Our demonstration of the activity of the tested extracts from Rubus sp. against H. pylori biofilm on fragments of infusions may provide a new option for covering artificial surfaces used in hospital treatments with compounds of natural origin.
5. Conclusions
In our experiments, we have shown that the best medium for examining H. pylori biofilm in 96-well titration plates is the Brucella broth medium, in which the metabolic activity of cells is detected by MTT. The extract from R. idaeus 'Poranna Rosa' fruits and R. idaeus 'Willamette' shoots proved be the most active. However, we considered the BHI medium supplemented with 5% bovine serum medium, which contains the highest percentage of live bacteria after 72 hours of incubation, detected using the TTC marker, as the best medium for testing fragments of intravenous infusions as a model where H. pylori forms a biofilm. In this model, we showed that the extract from R. idaeus 'Morning Rosa' fruits and R. idaeus 'Willamette' shoots are the most active in biofilm formation, and R. idaeus 'Willamette' shoot extract is active against bacteria in the biofilm formed by H. pylori. In the results of the study of interactions on planktonic H. pylori bacteria our extracts from Rubus sp. and only the 2 most active antibiotics showed synergism with doxycycline, and only the extract of Rubus occidentalis 'Litacz' showed synergism with levofloxacin. The remaining tested extracts showed indifference with levofloxacin. However, studies on the interaction of antibiotics and extracts of Rubus idaeus 'Willamette' shoot were carried out on fragments of intravenous infusions. The results were confirmed using SEM microscopy.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, R.H.; methodology, R.H,U.M. and M.K.B.; software, R.H.; validation, R.H. and K.T.; formal analysis, R.H.; investigation, R.H. and U.M.; resources, R.H..; data curation, R.H.; writing—original draft preparation, R.H.; writing—review and editing K.T., U.M. and M.K.B.; visualization, R.H.; supervision, K.T.,U.M. and M.K.B. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to many thanks to: dr M.Kula and prof dr hab M. Krauze-Baranowska for providing the tested extracts used in the research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wilkinson, D.; Alsharaf, L.; Thompson1, S.; Paulin, A.; Takor, R.; Robinson, A.Z.K.; Thomas, J.; McVicker, G.; Winter, J. Characterization of a Helicobacter pylori strain with high biofilm-forming ability. Journal of Medical Microbiology 2023, 2023, 72. [Google Scholar] [CrossRef]
- Mao, X.; Jakubovics, N.S.; Bächle, M.; Buchalla, W.; Hiller, K.A.; Maisch, T.; Hellwig, E.; Kirschneck, C.; Gessner, A.; Al-Ahmad, A.; Cieplik, F. Colonization of Helicobacter pylori in the oral cavity – an endless controversy? Critical Reviews in Microbiology 2021, 47, 612–629. [Google Scholar] [CrossRef]
- Lai, Y.; Wei, W.; Du, Y.; Gao, J.; Li, Z. Biomaterials for Helicobacter pylori therapy: therapeutic potential and future perspectives. Gut Microbes 2022, 14, e2120747, (22 pages.. [Google Scholar] [CrossRef]
- Krzyżek, P.; Gościniak, G. A Aproposed role for diffusible signal factors in the biofilm formation and morphological transformation of Helicobacter pylori. Turk J Gastroenterol 2018, 29(1), 7–13. [Google Scholar] [CrossRef]
- Krzyżek, P.; Biernat, M.M.; Gościniak, G. Intensive formation of coccoid forms as a feature strongly associated with highly pathogenic Helicobacter pylori strains. Folia Microbiologica 2019, 64, 273–281. [Google Scholar] [CrossRef]
- Kurokawa, M.; Nukina, M.; Nakanishi, H.; Tomita, S.; Tamura, T.; Shimoyama, T. Resuscitation from the viable but nonculturable state of Helicobacter pylori. Kansenshogaku Zasshi. The Journal of the Japanese Association for Infectious Diseases 1999, 73, 9–15. [Google Scholar]
- Azevedo, N.F.; Almeida, C.; Cerqueira, L.; Dias, S.; Keevil, C.W.; Vieira, M.J. Coccoid form of Helicobacter pylori as a morphological manifestation of cell adaptation to the environment. Appl. Environ Microbiol. 2007, 73, 3423–7. [Google Scholar] [CrossRef]
- Percival, S.L.; Suleman, L. Biofilms and Helicobacter pylori : Dissemination and persistence within the environment and host. World J Gastrointest Pathophysiol 2014, 5(3), 122–132. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Paluch, E.; Karwańska, M.; Wieliczko, A.; Gościniak, G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. Int. J. Mol. Sci., 2021, 22, 2695. [Google Scholar] [CrossRef] [PubMed]
- Young, K.A.; Allaker, R.P.; Hardie, J.M. Morphological analysis of Helicobacter pylori from gastric biopsies and dental plaque by scanning electron microscopy. Oral Microbiol Immunol. 2001, 16, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P.; Grande, R.; Migdał, P.; Paluch, E.; Gożciniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020a, 9, 1062. [Google Scholar] [CrossRef]
- Roszczenko-Jasińska, P.; Wojtyś, M.I.; Jagusztyn-Krynicka, E.K. Helicobacter pylori treatment in the post-antibiotics era—searching for new drug targets. Applied Microbiology and Biotechnology, 2020, 104, 9891–9905. [Google Scholar] [CrossRef]
- Krzyżek, P.; Paluch, E.; Gościniak, G. Synergistic Therapies as a Promising Option for the Treatment of Antibiotic-Resistant Helicobacter pylori. Antibiotics 2020b, 9, 658. [Google Scholar] [CrossRef]
- Vale, F.F.; Oleastro, M. Overview of the Phytomedicine Approaches against Helicobacter pylori. World J. Gastroenterol. 2014, 20, 5594–5609. [Google Scholar] [CrossRef]
- Safavi, M.; Shams-Ardakani, M.; Foroumadi, A. Medicinal Plants in the Treatment of Helicobacter pylori Infections. Pharm. Biol. 2015, 53, 939–960. [Google Scholar] [CrossRef]
- Ardalani, H.; Hadipanah, A.; Sahebkar, A. Medicinal Plants in the Treatment of Peptic Ulcer Disease: A Review. Mini. Rev. Med. Chem. 2019, 20, 662–702. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Yin, F.; Wang, S.; Zhao, A.; Li, Y.; Liu, Y. Helicobacter pylori Biofilm-Related Drug Resistance and New Developments in Its Anti-Biofilm Agents. Infection and Drug Resistance 2022, 15, 1561–1571. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; D’Addario, C.; Colacevich, A.; Focardi, S.; Borghini, F.;Santucci, A.; Figura, N.; Rossi, C. Antimicrobial activity against Helicobacter pylori strains and antioxidant properties of blackberry leaves (Rubus ulmifolius) and isolated compounds. International Journal of Antimicrobial Agents, 2009, 34, 50–59. [Google Scholar] [CrossRef]
- Park, J.U.; Cho, J.S.; Kim, J.S.; Kim, H.K.; Jo, Y.H.; Rahman, M.A.A.; Lee, Y.I. Synergistic Effect of Rubus crataegifolius and Ulmus macrocarpa Against Helicobacter pylori Clinical Isolates and Gastritis. Front. Pharmacol., 2020, 11, 4. [Google Scholar] [CrossRef]
- Krauze-Baranowska, M.; Majdan, M.; Hałasa, R.; Głód, D.; Kula, M.; Fecka, I.; Orzeł, A. The antimicrobial activity of fruits from some cultivar varieties of Rubus idaeus and Rubus occidentalis. Food Funct. 2014, 5, 2536–41. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M.; Głód, D.; Kula, M.; Majdan, M.; Hałasa, R.; Matkowski, A.; Kozłowska, W.; Kawiak, A. Chemical composition and biological activity of Rubus idaeus shoots – a traditional herbal remedy of Eastern Europe. Complementary and Alternative Medicine, 2014; 14, 480. Available online: https://www.biomedcentral.com/1472-6882/14/480.
- EUCAST, Clinical breakpoints - breakpoints and guidance, Available online https://www.eucast.org/clinical_breakpoints accessed on 2 January 2020. 2 January.
- Chen, X.; Li, P.; Shen, Y.; Zou, Y.; Yuan, G.; Hu, H. Rhamnolipid-involved antibiotics combinations improve the eradication of Helicobacter pylori biofilm in vitro: A comparison with conventional triple therapy. Microbial Pathogenesis 2019, 131, 112–119. [Google Scholar] [CrossRef]
- Cole, S.P.; Harwood, J.; Lee, R.; She, R.; Guiney, D.G. Characterization of Monospecies Biofilm Formation by Helicobacter pylori. J.Bacter. 2004, 186, 3124–3132. [Google Scholar] [CrossRef] [PubMed]
- Bugli, F.; Palmieri, V.; Torelli, R.; Papi, M.; De Spirito, M.; Cacaci, M.; Galgano, S.; Masucci, L.; Paroni Sterbini, F.; Vella, A.; Graffeo, R.; Posteraro, B.; Sanguinetti, M. In vitro effect of clarithromycina and alginate lyase aginst Helicobacter pylori biofilm. Biotechnol Prog. 2016, 32, 1584–1591. [Google Scholar] [CrossRef]
- Pillai, S.K.; Moellering, R.C., Jr.; Eliopoulos, G.M. Antimicrobial Combination. In Antibiotics in Laboratory Medicine, 5th Edition; Victor Lorain, Ed.; Publisher: Lippincott Williams & Wilkins: USA, Philadelphia, 2005; Chapter 9; pp. 374–440. [Google Scholar]
- Turecka, K.; Chylewska, A.; Kawiak, A.; Waleron, K.F. Antifungal Activity and Mechanism of Action of the Co(III) Coordination Complexes With Diamine Chelate Ligands Against Reference and Clinical Strains of Candida spp. Front. Microbiol. 2018, 9, 1594. [Google Scholar] [CrossRef] [PubMed]
- Teethaisong, Y.; Chueakwon, P.; Poolpol, K.; Ayamuang, I.; Suknasang, S.; Apinundecha, C.; Eumkeb, G. Stephania suberosa Forman extract synergistically inhibits ampicillin- and vancomycin-resistant Enterococcus faecium. Saudi Journal of Biological Sciences 2023, 30, 103557. [Google Scholar] [CrossRef]
- Turecka, K.; Chylewska, A.; Rychłowski, M.; Zakrzewska, J.; Waleron, K. Antibacterial Activity of Co(III) Complexes with Diamine Chelate Ligands against a Broad Spectrum of Bacteria with a DNA Interaction Mechanism. Pharmaceutics 2021, 13, 946. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Yasmin, T.; Bagchi, D.; Stohs, J.S. Inhibition of Helicobacter pylori in vitro by various berry extracts, with enhanced susceptibility to clarithromycin. Molecular and Cellular Biochemistry 2004, 265, 19–26. [Google Scholar] [CrossRef]
- Goodman, C.; Lyon, K.N.; Scotto, A.; Smith, C.; Sebrell, T.A.; Gentry, A.B.; Bala, G.; Stoner, G.D.; Bimczok, D. A High-Throughput Metabolic Microarray Assay Reveals Antibacterial Effects of Black and Red Raspberries and Blackberries against Helicobacter pylori Infection. Antibiotics 2021, 10, 845. [Google Scholar] [CrossRef]
- Nakase, K.; Nakanishi-Yamasaki, M.; Harada, K.; Koizumi, J.; Yamada, T.; Ikoshi, H.; Nakaminami, H.; Noguchi, N. Antimicrobial activity and additive effect of the modified Gingyo-san with antimicrobials against Helicobacter pylori. J. Infect. Chemiother. 2021, 27, 957–961. [Google Scholar] [CrossRef]
- Trung, H.T.; Huynh, H.T.T.; Thuy, L.N.T.; Minh, H.N.V.; Nguyen, M.-N.T.; Thi, M.N.L. Growth-Inhibiting, Bactericidal, Antibiofilm, and Urease Inhibitory Activities of Hibiscus rosa sinensis L. Flower Constituents toward Antibiotic Sensitive- and Resistant-Strains of Helicobacter pylori ACS Omega 2020, 5, 20080–20089. [Google Scholar] [CrossRef]
- Spósito, L.; Odab, F.B.; Vieiraa, J.H.; Carvalhob, F.A.; dos Santos Ramosa, M.A.; de Castrob, R.C.; Crevelinc, E.J.; Crottic, A.E.M.; Santosb, A.G.; da Silvad, P.B.; Chorillid, M.; Bauab, T.M. In vitro and in vivo anti-Helicobacter pylori activity of Casearia sylvestris leaf derivatives. Journal of Ethnopharmacology, 2019, 233, 1–12. [Google Scholar] [CrossRef]
- Brown, H.L.; van Vliet, A.H.M.; Betts, R.P.; Reuter, M. Tetrazolium reduction allows assessment of biofilm formation by Campylobacter jejuni in a food matrix model. J. Appl. Microbiol. 2013, 115(5), 1212–21. [Google Scholar] [CrossRef]
- Windham, I.H.; Servetas, S.L.; Whitmire, J.M.; Pletzer, D.; Hancock, R.E.W.; Merrell, D.S. Helicobacter pylori Biofilm Formation Is Differentially Affected by Common Culture Conditions, and Proteins Play a Central Role in the Biofilm Matrix. Applied and Environmental Microbiology 2018, 84, e00391-18. [Google Scholar] [CrossRef]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Cells are Metabolically Distinct, Express Flagella, and Antibiotic Tolerant. bioRxiv 2019. [Google Scholar] [CrossRef]
Figure 1.
The level of viable cells in a biofilm formed in 2 different media (BHI and Brucella) labeled with 2 dyes (TTC and MTT). The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 1.
The level of viable cells in a biofilm formed in 2 different media (BHI and Brucella) labeled with 2 dyes (TTC and MTT). The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
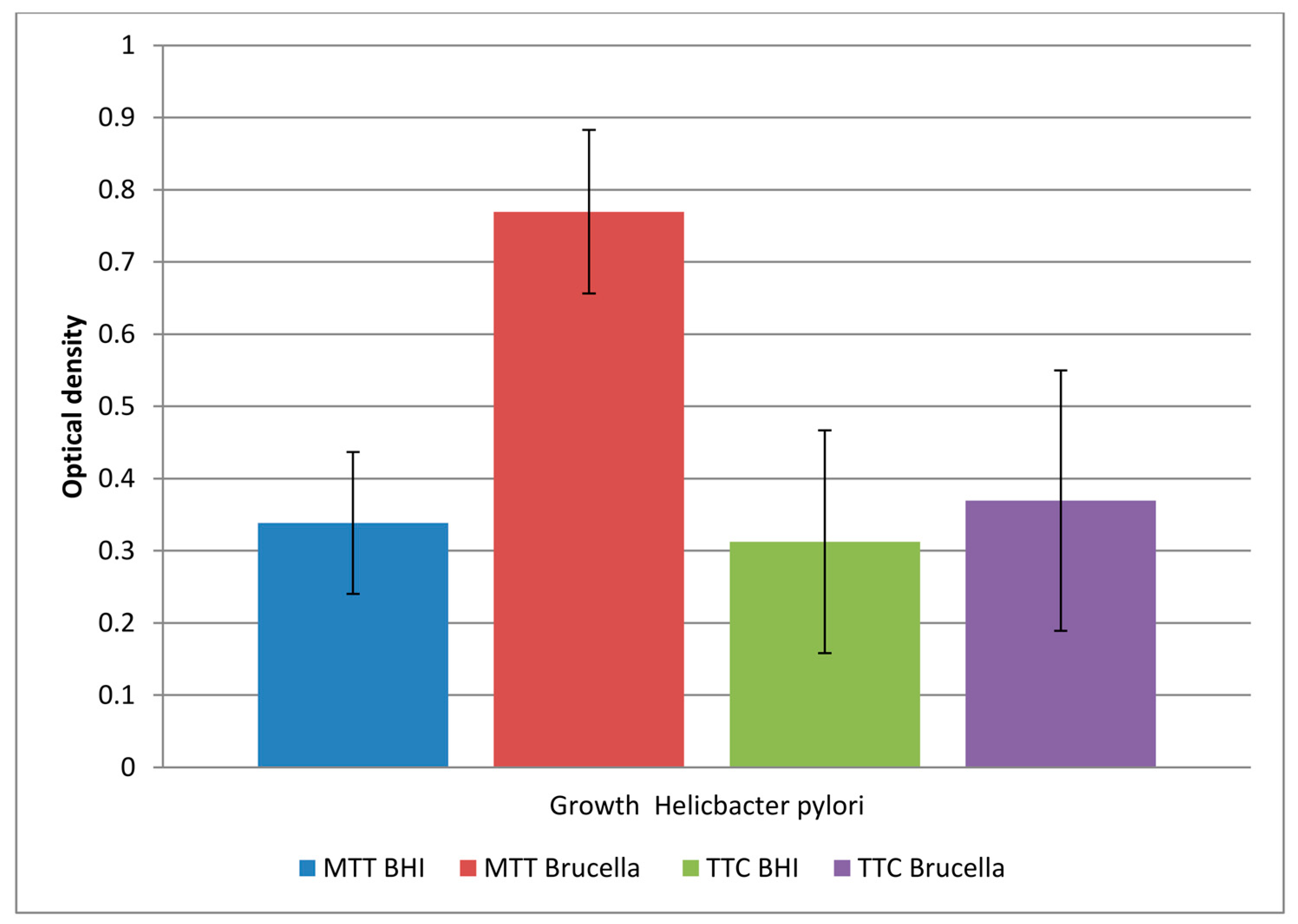
Figure 2.
Examination of the activity of plant extracts on the created biofilm by Helicobacter pylori in a 96-well titration plate in Brucella (a) and BHI media with 5% horse serum (b). MTT was used as the marker. Measurements were made at OD 550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 2.
Examination of the activity of plant extracts on the created biofilm by Helicobacter pylori in a 96-well titration plate in Brucella (a) and BHI media with 5% horse serum (b). MTT was used as the marker. Measurements were made at OD 550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
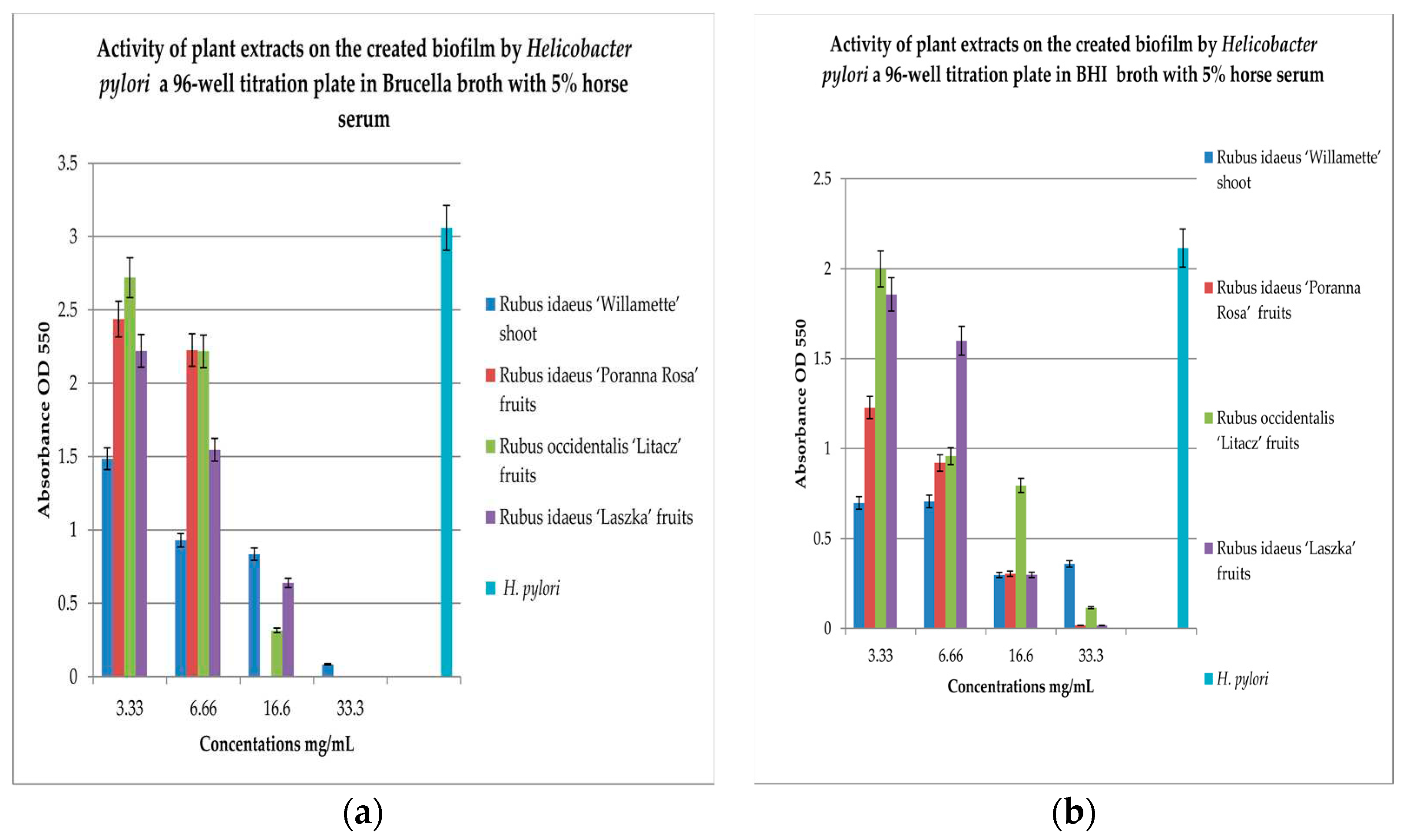
Figure 3.
Determination of the amount of biofilm formed on glass coverslips in BHI and Brucella broth, with the addition of 5% horse serum and in the presence or absence of β-cyclodextrin, using 3 dyes: MTT, CV, and TTC. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 3.
Determination of the amount of biofilm formed on glass coverslips in BHI and Brucella broth, with the addition of 5% horse serum and in the presence or absence of β-cyclodextrin, using 3 dyes: MTT, CV, and TTC. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
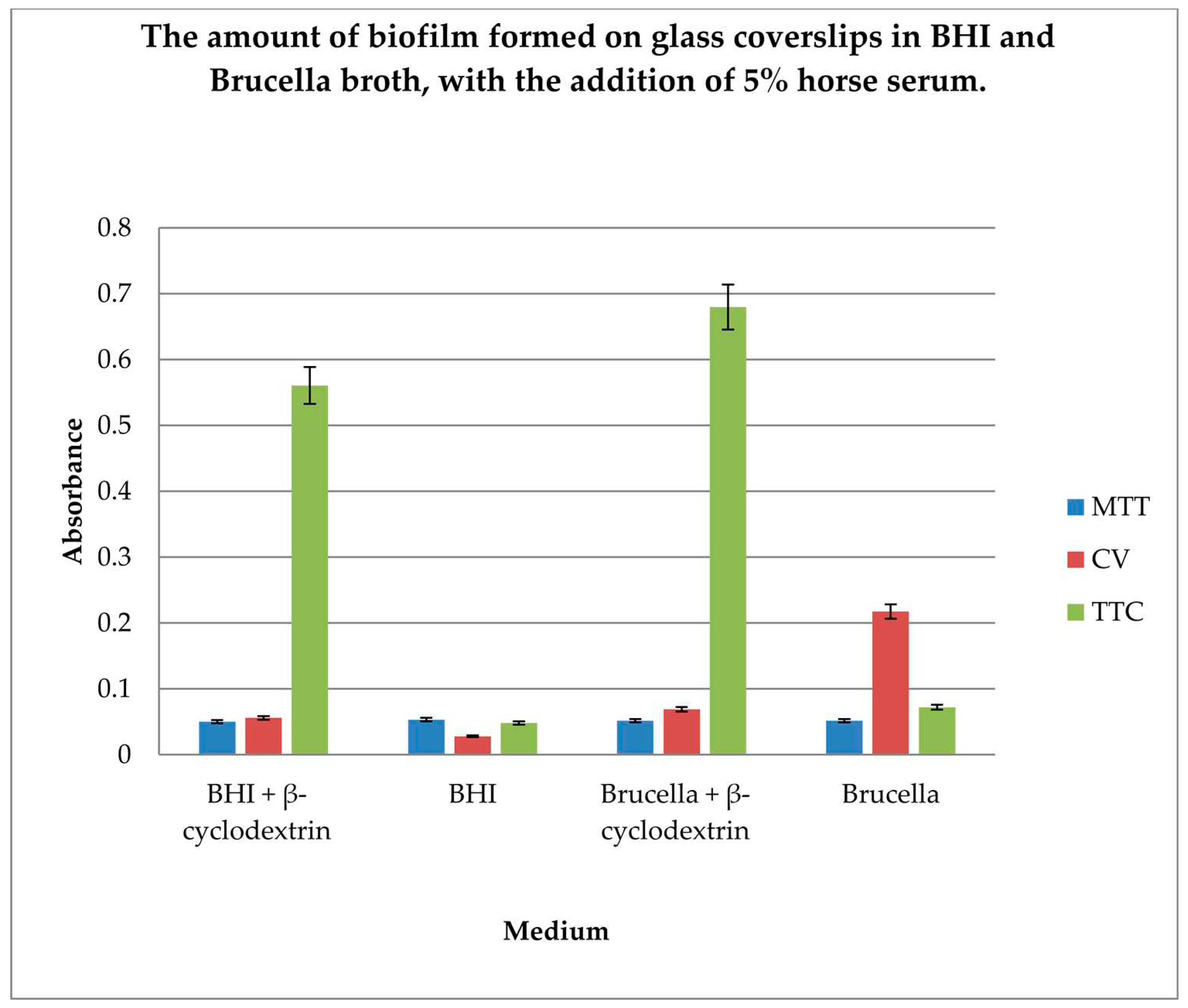
Figure 4.
Determination of the amount of biofilm formed on fragments of an intravenous infusion. BHI and Brucella broth, with the addition of 5% horse serum and in the presence or absence of β-cyclodextrin, MTT, and TTC markers, were used. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 4.
Determination of the amount of biofilm formed on fragments of an intravenous infusion. BHI and Brucella broth, with the addition of 5% horse serum and in the presence or absence of β-cyclodextrin, MTT, and TTC markers, were used. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
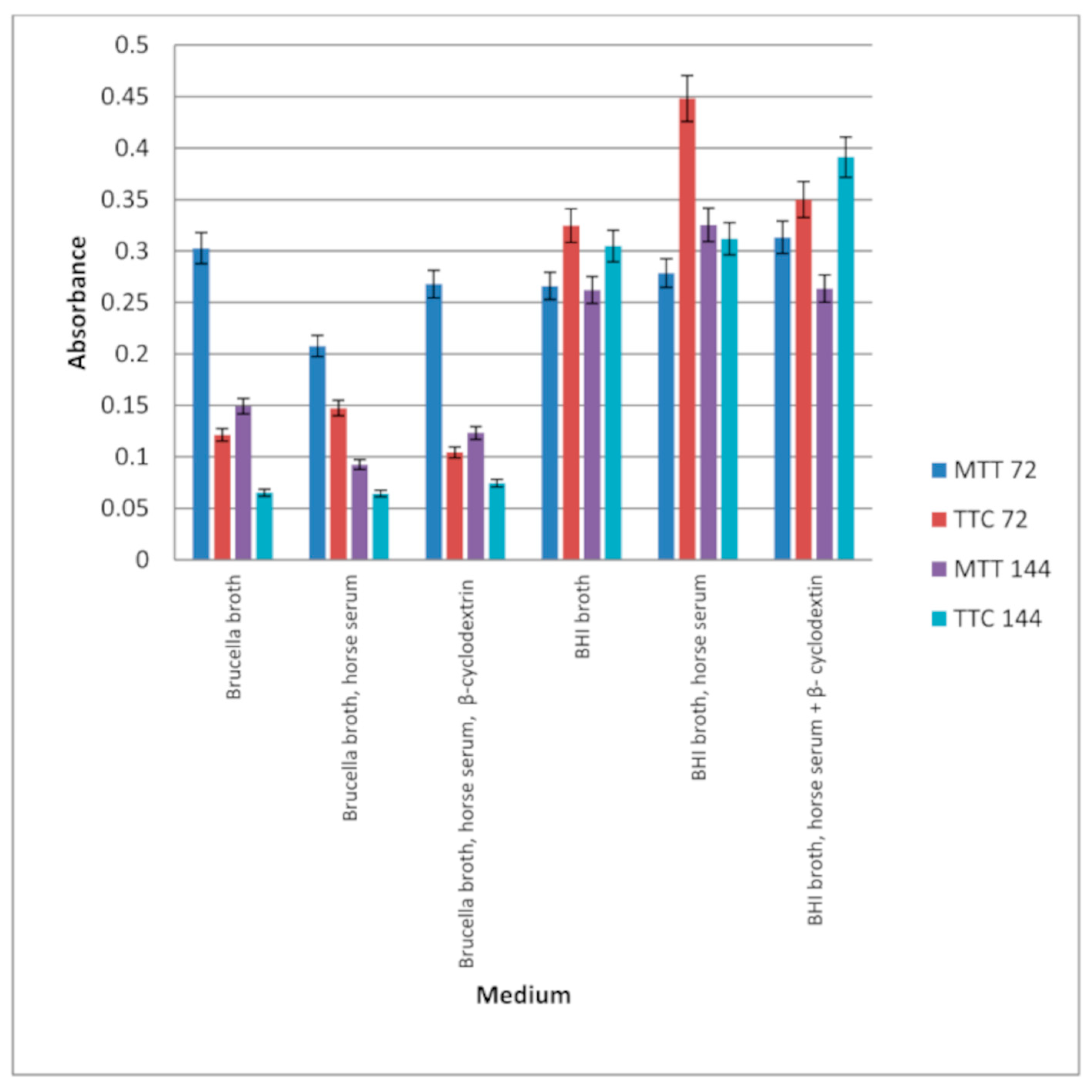
Figure 5.
Study of the effect of extracts on the formation of biofilm by Helicobacter pylori on fragments of intravenous infusions in BHI broth with the addition of 5% horse serum. The determination was carried out by TTC at absorbance OD550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 5.
Study of the effect of extracts on the formation of biofilm by Helicobacter pylori on fragments of intravenous infusions in BHI broth with the addition of 5% horse serum. The determination was carried out by TTC at absorbance OD550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
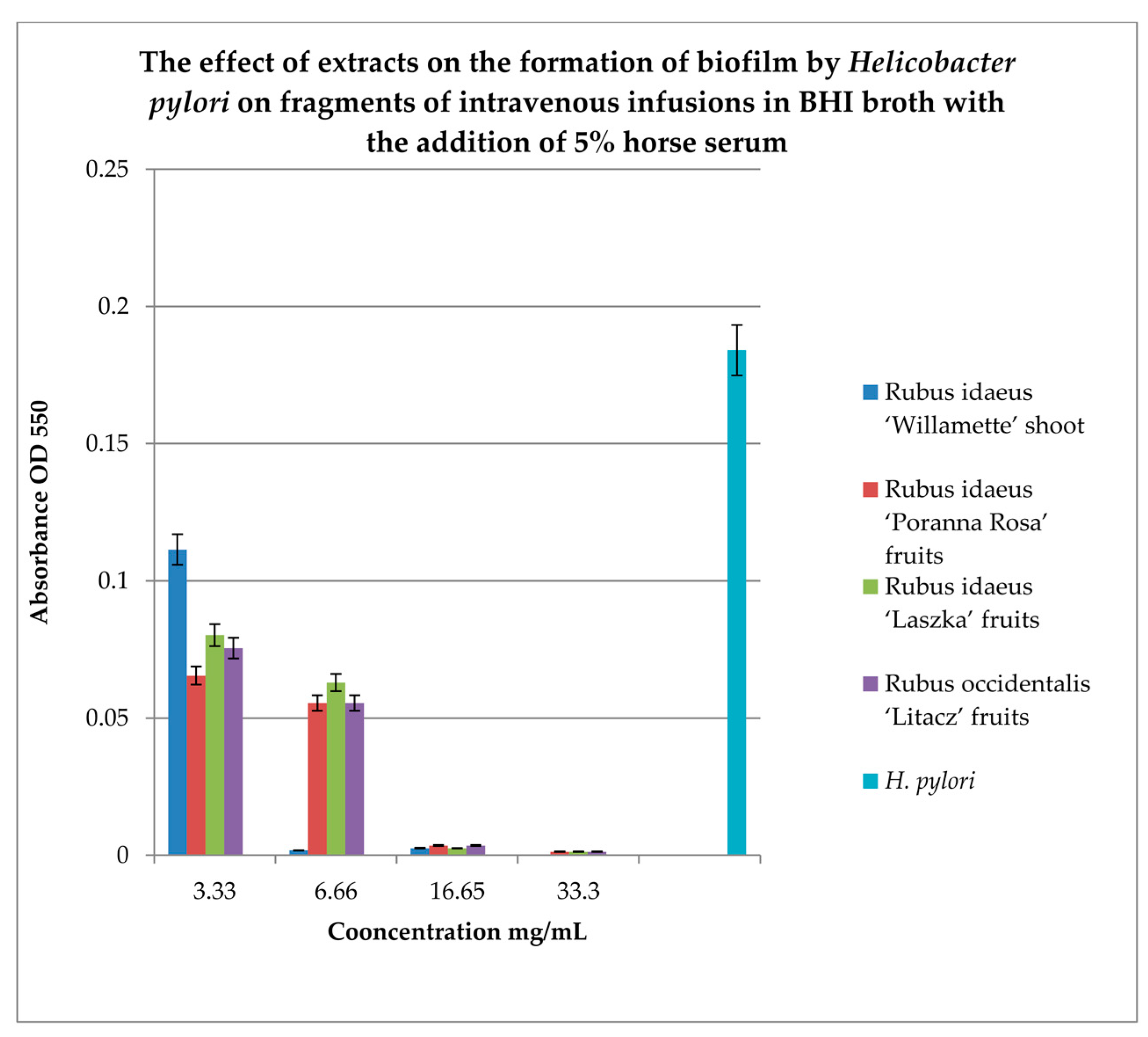
Figure 6.
Examination of the effect of extracts on Helicobacter pylori biofilm formed on fragments of intravenous infusions in BHI broth, including 5% horse serum. The determination was carried out by TTC at OD550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
Figure 6.
Examination of the effect of extracts on Helicobacter pylori biofilm formed on fragments of intravenous infusions in BHI broth, including 5% horse serum. The determination was carried out by TTC at OD550. The results are presented as mean values ± standard deviation (±SD) from 3 independent experiments. Error bars represent standard deviation. p < 0.05 was considered as statistically significant.
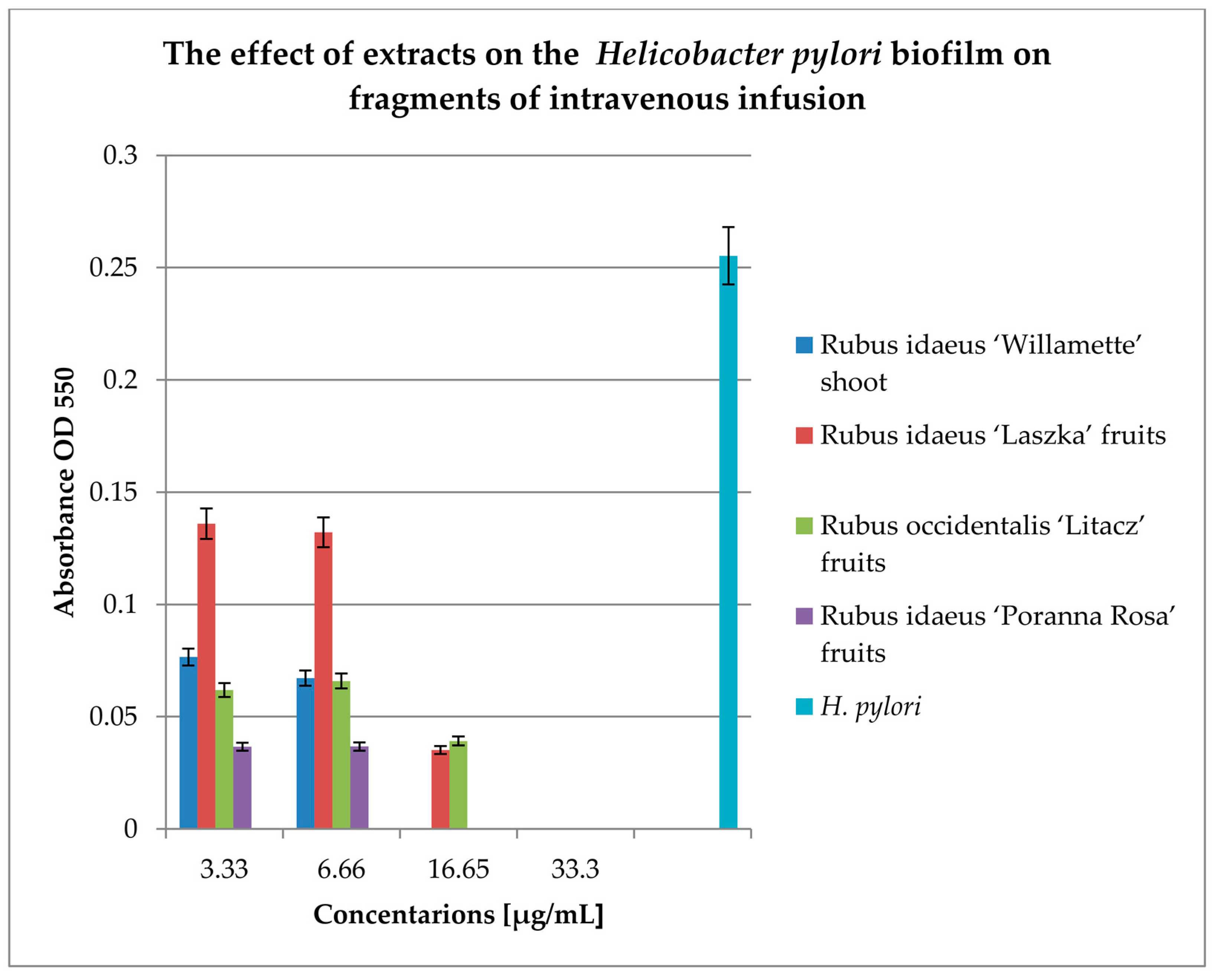
Figure 7.
Photo showing the following: (a) Sterile surface of the intravenous infusion. (b), (c) The biofilm of H.pylori on surface of the intravenous infusion. (d) The effect of Rubus idaeus 'Willamette' shoot extract at a concentration of 6.66 mg/mL on forming biofilm. (e), (f) The effect of Rubus idaeus 'Willamette' shoot extract at a concentration of 6.66 mg/mL on bacteria in formed biofilm.
Figure 7.
Photo showing the following: (a) Sterile surface of the intravenous infusion. (b), (c) The biofilm of H.pylori on surface of the intravenous infusion. (d) The effect of Rubus idaeus 'Willamette' shoot extract at a concentration of 6.66 mg/mL on forming biofilm. (e), (f) The effect of Rubus idaeus 'Willamette' shoot extract at a concentration of 6.66 mg/mL on bacteria in formed biofilm.



Table 1.
MIC values of antibiotics and Rubus sp. extracts [mg/mL].
| Antibiotics | MIC | |
|---|---|---|
| BHI broth with 5% horses serum |
Brucella broth with 5% horses serum |
|
| amoxicillin | 0.0160±0.01 | 0.016±0.01 |
| clarithromycin | >0.128 | >0.128 |
| doxycycline | <0.000625 | <0.000625 |
| levofloxacin | 0.000125±0.01 | 0.000125±0.01 |
| metronidazole | >0.128 | >0.128 |
| Extracts | ||
|
Rubus occidentalis ‘Litacz’ fruits |
7.8±0.50 | 7.8±0.50 |
|
Rubus idaeus ‘Laszka’ fruits |
7.5±0.50 | 7.5±0.50 |
|
Rubus idaeus ‘Poranna Rosa’ fruits |
7.2±0.50 | 7.2±0.50 |
| Rubus idaeus ‘Willamette’ shoot | 7.4±0.50 | 7.4±0.50 |
| Ellagic acid | 0.125±0.05 | 0.125±0.05 |
The results are presented as mean values ± standard deviation (±SD).
Table 2.
Interactions of Rubus spp. extracts with antibiotics on planktonic cells.
| Antibiotics | MIC antibiotic alone [µg/mL] |
MIC antibiotic combination [µg/mL] | Extracts | MIC extracts alone [mg/mL] |
MIC extracts combination [mg/mL] |
FICI | Outcome |
|---|---|---|---|---|---|---|---|
| Levofloxacin | 0.5 | 0.016 | Rubus idaeus ‘Willamette’ shoot | 33.3 | 33.3 | 1.032 | Indifference |
| Doxycycline | 0.125 | 0.016 | 3.33 | 0.228 | Synergy | ||
| Levofloxacin | 0.5 | 0.016 |
Rubus idaeus ‘Poranna Rosa’ fruits |
16.65 | 16.65 | 1.032 | Indifference |
| Doxycycline | 0.125 | 0.016 | 3.33 | 0.328 | Synergy | ||
| Levofloxacin | 0.5 | 0.016 |
Rubus occidentalis ‘Litacz’ fruits |
33.3 | 16.65 | 0.532 | Synergy |
| Doxycycline | 0.0625 | 0.016 | 3.33 | 0.356 | Synergy | ||
| Levofloxacin | 0.25 | 0.016 |
Rubus idaeus ‘Laszka’ fruits |
33.3 | 33.3 | 1.064 | Indifference |
| Doxycycline | 0.125 | 0.016 | 3.33 | 0.228 | Synergy |
Table 3.
Interactions of Rubus idaeus ‘Willamette’ shoot extracts with antibiotics on biofilm formed in intravenous infusions.
Table 3.
Interactions of Rubus idaeus ‘Willamette’ shoot extracts with antibiotics on biofilm formed in intravenous infusions.
| MIC antibiotic alone [µg/mL] |
MIC antibiotic combination [µg/mL] | MIC Rubus idaeus ‘Willamette’ shoot [mg/mL] |
MIC extracts combination [mg/mL] |
FICI | Outcome | |
|---|---|---|---|---|---|---|
| Amoxicillin | 128 | 32 | 6.66 | 3.33 | 1 | Addition |
| Clarithromycin | 128 | 128 | 6.66 | 2 | Indifference | |
| Metronidazole | 128 | 128 | 3.33 | 1.5 | Indifference | |
| Levofloxacin | 0.5 | 0.25 | 0.8 | 0.62 | Synergy | |
| Doxycycline | 0.25 | 0.125 | 0.8 | 0.62 | Synergy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



