
Submitted:
25 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Grapevine Trunk Diseases (GTDs) are caused by a wide variety of phytopathogenic fungal species that compromise grapevine productivity and wine quality. Each year they lead to major economic losses. Most GTD treatments currently available on the market are chemical-based and environ-mentally harmful. Given that one of the goals proposed by the European Green Deal is to develop sustainable agriculture which does not harm the environment, reduces pesticide use, and pro-motes rural area economies, an alternative to those treatments may be the use of elicitors such as oligosaccharides from fungal organisms. Indeed, many studies have confirmed that oligosaccha-rides play important roles in activating plants' innate immunity. Mannans are one possible type of such oligosaccharides. The present work analysed Vitis vinifera L. (cv. Tempranillo and cv. Airén) leaves and grapes to evaluate their physiological and molecular response to GTDs and how this response might be conditioned using different concentrations of mannans as elicitors. It was found that mannan addition to diseased vines was more positive for the leaves than for the grapes. In general, mannans increased the expression levels of chalcone synthase (CHS1, CHS3), phenylalanine ammonia lyase (PAL), superoxide dismutase (SOD), and polyphenol oxidase (PPO) in healthy leaves of both varieties. In diseased leaves, CHS3 and PAL decreased in both varieties, while CHS1 and PPO increased in Tempranillo but not in Airén. In the grapes, the expression pat-terns of the genes studied varied greatly due to the development of the disease. The mannan treatment seemed to reduce the oxidative stress caused by GTDs, although this response depend-ed on the variety and on the plant's phytosanitary status.
Keywords:
grapevine
; GTD
; mannans
; phenols
; polyphenol oxidase
; superoxide dismutase
; sustainable
* espinosa@unex.es
1. Introduction
Agriculture faces challenges that put its sustainability at risk. These include climate change, phytopathogenic organisms, infectious plant diseases, soil nutrient deficiency, reduced crop yields, ever stricter legislation on using and developing genetically modified crops, and labour shortages [1,2,3]. For years, there has been promotion of agriculture based on the use of chemical inputs (commercial pesticides and fertilizers), crop rotation, precision farming, and traditional plant improvement, and, more recently, their combination with breeding and genetic manipulation techniques, although these last techniques are still only available for a small group of crop species including maize, soybeans, and rice [4]. Furthermore, not only do their volatilization and leaching lead to conventional and chemical fertilizers having a low efficiency but, in the long-run, their excessive use causes environmental pollution (e.g., soil quality degradation, groundwater contamination, and surface-water eutrophication), thereby limiting the goal of sustainable agriculture [5,6]. Given the outlook of climate change and the need to safely and sustainably respond to current and future food demands, it is crucial to develop new approaches that can better protect crops against biotic and abiotic stressors [7,8,9].
Vitis vinifera L is a crop with a worldwide distribution and has a major impact on many countries’ economies. Vineyard products such as wine or table grapes have multiple health benefits [10]. Grapes for example are a rich source of vitamins, minerals, carbohydrates, dietary fibre, and more than 1600 natural plant compounds including catechins, ellagic acid, lutein, quercetin, lycopene, and polyphenols (such as stilbene, flavonols, proanthocyanidins, and anthocyanins) which are very effective in protecting against cardiovascular diseases and other pathologies [11]. The problems facing viticulture today mainly derive from climate change or from grapevine trunk diseases (GTDs) which re-emerged in the late 1990s [12]. These diseases compromise not only vine longevity and productivity but also wine quality [13,14], with the result being economic losses due to increased production costs [15]. Many factors make it difficult to control GTDs. Examples of GTDs are such pathologies as Botryosphaeria dieback, Eutypa dieback, esca, blackfoot disease, excoriosis, and Petri disease or young grapevine decline [14]. Of these, those that most frequently affect mature grapevines are esca, Botryosphaeria dieback, and Eutypa dieback [13].
GTDs are a group of vascular diseases produced by pathogenic fungi, mainly ascomycetes and basidiomycetes, which affect grapevine wood and inhabit the xylem cells in the woody tissue [14,16,17]. When this type of fungus colonizes this tissue, it causes deterioration of the host plant due to the loss of xylem function and the consequent decrease in hydraulic conductivity [18,19,20]. Moreover, pathogenic fungi can produce compounds that activate secondary metabolism, increasing anthocyanin synthesis and key enzymatic reactions (NADPH oxidase and phenylalanine ammonia lyase) or oxidative enzymes such as laccases (a type of PPO) leading to the oxidation of phenols and polyphenols [21,22,23]. New fungal species with this pathogenic capacity continue to be discovered. By 2018 already, 133 species had been identified belonging to 34 different genera [15]. Due to this great variety of pathogenic fungi involved in GTDs, the infection process and the external and internal symptoms can range fairly widely, depending on the pathology being analysed [15,16,17,24,25,26,27], although there is generally necrosis and decay in the wood tissue leading over time to foliar symptoms and the plant’s death [18,19,20]. For instance, plants affected by esca show “tiger stripe” patterns on the leaves, and plants affected by Eutypa dieback have stunted shoots with chlorotic leaves and bunches that mature unevenly [14]. Infection will also depend on the plant’s phenology (young plants are mainly infected in the field while mature vineyards are usually infected through the root system) and on the type of pathogenic fungus (the Botryosphaeriaceae family and Eutypa lata penetrate the plant through pruning wounds), although the same pathogenic fungus can infect through different entry routes (Phaeoacremonium minimo and Phaeomoniella chlamydospora, causal agents of Petri disease, can enter the plant both through the root system and through wounds) [14,16,25,27].
After two decades of unsuccessful attempts to develop chemical control or biocontrol methods to mitigate the effect of GTDs [17], alternatives without the use of pesticides are currently being sought as this is one of the objectives included in the European Green Deal. One such alternative could be the use of elicitors such as carbohydrates, compounds with no toxicity, which can be extracted from renewable sources and contribute to conserving the environment [28].
Many studies have confirmed that oligosaccharides play some important roles in activating plant innate immunity [29]. They are low molecular weight polymers composed of 2 to 20 identical or different monosaccharides linked by glycosidic bonds [30]. This kind of compound derives mainly from plant cellulose or hemicellulose, e.g., pectins, xyloglucans, cellodextrins, etc. [31,32,33], or from fungal cell walls or arthropod exoskeletons, e.g., β-glucan, chitin, and chitosan [32,34,35]. Chitosan oligosaccharides (COS), oligogalacturonides (OGA), and xyloglucan oligosaccharides (XGO) are involved in plant growth and defence responses [36,37]. For example, COS, an analogue of chitin oligosaccharides, may act in plants to elicit their innate immunity to plant diseases. When COS is detected by the plant, the transduction pathway is activated to induce early defence responses [35]. The use of chitosan reduces the incidence of mildew in grapes, with an increase in polyphenols [38] and a reduction in the growth of fungal hyphae that cause GTDs, resulting in improved growth of infected vines [39]. As another example, OGA can be detected by cell-wall-associated receptors such as WAK1 and WAK2, triggering plant immunity through MAPK activation, an oxidative burst, and the accumulation of pathogenesis-related proteins providing the plant with increased resistance to pathogen invasion [40,41,42]. The mannan oligosaccharides (MOS) are a type of saccharide derived from the cell wall of the yeast Saccharomyces cerevisiae. The use of MOS as elicitor could be a valuable strategy for controlling plant disease since they have been reported to induce local defences and systemic resistance to pathogens [29].
Yeast cell-wall extracts contain several compounds (mannoproteins, glucans, and chitin) that can act as elicitors triggering different defence modes in plants [43,44]. They are non-toxic, biodegradable, and ecological in the sense of being environmentally friendly. Treatment with them increases the activity of chitinase, a member of the group of proteins related to pathogenesis, i.e., it is induced by pathogen infection. Plants’ main response to fungus attack is to secrete chitinase because of its strong antifungal potential, degrading the fungus’s cell wall [45]. Application of these extracts in the field can increase the plants’ growth and yield [46] as well as the phenol content, antioxidant activity, and concentration of photosynthetic pigments [47,48].
The use of cell wall extracts, with a large proportion of the compounds they contain being mannans, increases resistance to fungal and bacterial infections in both leaves and fruit [49,50]. In grapes, the anthocyanin and stilbene contents increase [44], while foliar application alters the anthocyanin content but not that of flavonoids [43]. Mannan extracts have the capacity to eliminate ROS [51], and in the vine their application protects against another fungal disease –mildew– modifying the metabolism of phenylpropanoids and phytohormones, enzymes such as SOD and PPO, and the photosynthesis process [52]. With the application of these extracts, there is a physiological alteration involving reduced free and total amino acid contents, and resistance to the disease is induced in cv. Tempranillo [44,53].
By acting as pathogen-associated molecular patterns (PAMPs) that bind and are recognized by pattern recognition receptors (PRRs), the use of oligosaccharides such as mannans can allow the control of diseases in plants. Their recognition triggers defence reactions, both local resistance and acquired systemic resistance. This method may be an alternative to the use of synthetic chemicals to protect the vine.
In this work, the effect was measured of the application of mannans extracted from S. cerevisae on the pigment and phenol content and on the polyphenol oxidase (PPO) and superoxide dismutase (SOD) activities in grapevine leaves and grapes of the Tempranillo and Airén varieties, both healthy and affected by GTD-causing fungi. In general, mannans cause stress in healthy vines, but in diseased vines the effect seems to be favourable, palliating the damage caused by the disease, especially in the Tempranillo variety. Also analysed in this work were the expression patterns of enzymes involved in the synthesis (PAL) and degradation (PPO) of phenols and in the production of flavonoids (chalcone synthase).
2. Materials and Methods
2.1. Plant material and growing conditions
The experiment was carried out in vineyards located in Carrilejo-Manzanares (X:39.022277, Y: -3.388166) and Daimiel (X:39.133228, Y: -3.467667) in the province of Ciudad Real for cv. Tempranillo and cv. Airén (Vitis vinifera L.), respectively. The cv. Tempranillo vineyard (5 ha) had been planted in 2000 with a 2.0 m × 2.9 m frame (1724 vines ha-1), and that of cv. Airén (6 ha) in 1996 with a 2.6 m × 2.6 m frame (1479 vines ha-1). The climate conditions are found in https://crea.uclm.es/siar/datosMeteorologicos.
Healthy vines (H, showing no GTD symptoms) and diseased vines (D, with visible GTD symptoms) were subjected to treatments with mannans obtained from the cell wall of Saccharomyces cerevisae. The experimental design was randomized block with 4 replications (132 trunks per treatment). The mannan doses in each treatment were 0, 1, 2, and 3 mL L-1 (HT0, HT1, HT2, HT3 and DT0, DT1, DT2, DT3, for healthy and diseased vines respectively). A series of 8 foliar applications were applied, spraying 600 L of the mannan broth per ha. For cv. Tempranillo, the first application was at the beginning of flowering (BBCH = 60-62, [54]), and for cv. Airén at fruit set (BBCH = 71, [54]). Further applications were made every 15 days. Leaf and grape samples were taken for the Tempranillo cultivar at the beginning of September (4 September 2020) and for Airén at the end of September (24 September 2020).
2.2. Photosynthetic pigment content
Leaf discs were taken from fresh leaves and incubated in methanol (0.0125 g mL-1) for 24 h in darkness at room temperature. The chlorophyll a, chlorophyll b, and carotenoid contents were determined spectrophotometrically by measuring A666, A653, and A470. The total chlorophyll and carotenoid contents were calculated in accordance with Wellburn [55]. The results are expressed as µg pigment g-1 FW (fresh weight).
2.3. Phenol content
Total phenols, flavonoids, and phenylpropanoid glycosides (PPG) were extracted from 1 g fresh leaves or grapes by homogenizing in methanol, chloroform, and 1% NaCl (5:5:2.5 mL), filtering, and centrifuging at 3200 g for 10 min. Total phenols were determined spectrophotometrically at A765 with the Folin-Ciocalteu reagent [56], expressing the result as μg caffeic acid g-1 FW. Total flavonoid content was measured at A415 [57], expressing the result as μg of rutin g-1 FW. Phenylpropanoid glycosides were determined at A525 [58], expressing the result as μg of verbascoside g-1 FW.
2.4. Grape anthocyanin content
The anthocyanin content was measured in cv. Tempranillo grapes by the differential pH method. This is based on the structural transformation that anthocyanins undergo with changing pH (coloured at pH 1.0, colourless at pH 4.5) [59]. For this, 2.5 g of grapes were homogenized in 2.5 mL of a 0.1 M ethanol:HCl solution (85:15% v:v), followed by centrifuging at 4000 g for 10 min, diluting the supernatant in two different buffers (0.025 M KCl, pH 1.0, and 0.4 M sodium acetate, pH 4.5), and incubating for 30 min at room temperature. The absorbance at 520–700 nm was measured. The anthocyanin content is expressed as mg of malvidin 3-glucoside (a major grape pigment) g-1 FW, in accordance with the equation given by MohdMaidin et al. [60].
2.5. PPO and SOD activities
To measure the PPO activity, leaves (0.5 g mL-1) or grapes (1.5 g mL-1) were homogenized in phosphate buffer (100 mM, pH 6.5) with 1% PVPP, centrifuged at 19 000 g for 30 min at 4 °C, and the supernatant used for the enzyme determination. The protein content was determined by the Bradford method [61]. PPO activity was determined by measuring A420 at 25 °C in a medium containing the enzyme extract, 100 mM phosphate buffer, and 0.1 M catechol [62].
For SOD activity, fresh leaves (0.5 g mL-1) or grapes (1.5 g mL-1) were homogenized at 4 °C in 50 mM pH 6.0 phosphate buffer, 1 mM EDTA, 0.5 mM PMSF (phenylmethylsulfonyl fluoride), 1 mM β-mercaptoethanol, 1 g L-1 PVPP. The homogenate was filtered and centrifuged at 39 000 g for 30 min at 4 °C, and the supernatant used for the enzyme determination. The protein content was determined by the Bradford method [61]. Superoxide dismutase (SOD) activity was determined at A560 in a medium containing 50 mM phosphate buffer pH 7.8, 0.1 mM EDTA, 1.3 μM riboflavin, 13 mM methionine, and 63 μM 4-nitro blue tetrazolium (NBT) [63].
2.6. RNA extraction and cDNA transformation
Plant material (fresh leaves or grapes) was frozen in liquid nitrogen and stored at -80 °C until RNA extraction. The RNA was extracted and purified using the Spectrum Plant Total RNA kit (Sigma-Aldrich®) and RNase-Free DNase (Qiagen® No 79254). The RNA quantity and quality of each resulting sample was determined using an Eppendorf D30 biophotometer (Eppendorf, Germany). The Applied Biosystems® High Capacity cDNA Reverse Transcription Kit was used for reverse transcription of 1 to 2 µg samples of purified RNA. The integrity of the extracted RNA was assessed by gel electrophoresis using a 1.5% agarose gel with 1x TAE (Tris-acetate-EDTA) buffer and ethidium bromide as intercalating agent (0.075%), loading 2.5 µL of RNA from each sample with 10 µL of RNase-free water and 2 µL of loading buffer.
2.7. qRT-PCR
The real-time amplification was monitored with SYBR Green (Thermo Fisher Scientific) on a QuantStudio 1 amplification and detection instrument (Applied Biosystems, Thermo Fisher Scientific R). VATP16 (V-type proton ATPase) and actin1 were used as housekeeping genes [64,65]. Previous studies have qualified both as being genes with a stable expression pattern under biotic stress conditions [66]. The primers used for grapevine cDNA amplification are given in Table 1.
2.8. Statistical analysis
The means ± SE of each measurement are presented in each figure or table. At least 10 replicates obtained from three independent experiments were analysed. For each measurement, a Shapiro–Wilk normality test was performed (since n<50) followed by a one-way ANOVA parametric test. The values for which there are significant differences, i.e., where p≤0.05, are marked with different letters. All statistical analyses were performed with Microsoft Excel 365 (Microsoft Inc., Alburquerque, NM, USA) and R version 2.9.2 (2009-08-24; Copyright (C) 2009 The R Foundation for Statistical Computing; ISBN 3-900051-07-0).
3. Results and Discussion
3.1. Photosynthetic pigment content
The photosynthetic pigment contents of leaves of both varieties were determined (Table 2). In healthy cv. Tempranillo leaves, there were decreases in both chlorophyll a and chlorophyll b content in response to the application of mannans. These decreases increased with dose. In diseased leaves, the mannan application led to a decrease in the content of both chlorophylls relative to the healthy leaf control values (HT0), but no such significant effect relative to the diseased leaf control values (DT0). In healthy cv. Airén leaves, the application of the different mannan treatments did not alter the chlorophyll content but, as was the case for cv. Tempranillo, in diseased leaves, there were significant decreases in both chlorophylls with respect to the control values (DT0). In particular, there were slight dose-dependent decreases in chlorophyll a while the chlorophyll b content remained unaltered. For the same given treatment, diseased leaves showed lower chlorophyll a and b contents.
The chlorophyll a/b ratio was calculated since its increase may indicate stress in plants (Table 2). For cv. Tempranillo, in healthy leaves the mannan application led to an increase in this ratio, and it was also significantly higher in diseased leaves relative to healthy leaves (HT0). Mannan application induced a decrease in the a/b ratio in diseased leaves only for T3D. For cv. Airén, in healthy leaves neither HT1 nor HT2 affected this ratio, with only HT3 leading to a significant decrease. In diseased leaves, this ratio was higher than in the healthy leaves (DT0), and remained unaffected by the mannan treatments. The increase in the chlorophyll a/b ratio would indicate reduced thylakoid appression which, according to Zhou et al. [67], might contribute to the dissipation of excess energy that would imply an adaptation to stress. Thus, in cv. Tempranillo both the disease and the mannans would cause stress in the photosynthetic apparatus. Only the highest mannan concentration showed a positive effect on the leaves of diseased vines. In cv. Airén only the disease showed a negative effect, while mannans had practically no effect except for HT3 in which they had a protective effect.
With respect to the carotenoid content, in Tempranillo (Table 2), slight oscillations with little significance were observed in both healthy and diseased leaves due to the effect of mannan application. Neither were there changes in the carotenoid content in HT0 and DT0. One observes that the ratio between carotenoids and total chlorophylls increased significantly in healthy leaves when mannans were applied. In diseased leaves, the values were higher than in healthy leaves, and at DT3 the mannan application led to their significant reduction. In cv. Airén, the carotenoid content of healthy leaves was unaffected by mannan treatments. For diseased leaves, the carotenoid content was similar in DT0 to that in HT0, and the mannan treatments DT2 and DT3 significantly reduced their carotenoid contents. With respect to the ratio of carotenoids to total chlorophyll, mannan application led to no significant differences in either healthy or diseased leaves, but the disease increased this ratio relative to the healthy leaf case. This increased carotenoid content could be indicative of these compounds’ protective role because of their antioxidant activity. This response of an increased carotenoid-to-chlorophyll ratio was observed in cv. Tempranillo in response both to mannans and to the disease, but in cv. Airén only in response to the disease.
In general, similar results regarding photosynthetic pigment content had been obtained by Petit et al. [68] who describe a decrease in chlorophylls a and b in leaves of cv. Chardonnay affected by tinder while carotenoids seem to be unaltered and may have, according to those workers, a protective role for plant membranes against fungi in both diseased and healthy leaves. Those authors report no differences in the chlorophyll a/b ratio between healthy and diseased leaves, apparently because the two chlorophylls are damaged in the same way. García et al. [23] describe similar results for cv. Tempranillo, with a decrease in chlorophyll content in vines affected by tinder, and no variation due to the effect of tinder in the carotenoids in the vines harvested in August but an increased carotenoid/chlorophyll ratio. Also for cv. Tempranillo, Martin et al. [69] describe decreases in diseased leaves in chlorophylls and in carotenoids. Santos et al. [70] also observed decreased chlorophyll a and b contents in infected vines, with an increased chlorophyll a/b ratio, the behaviour depending on the variety studied. García et al. [23] observed a decrease in both chlorophylls in cv. Tempranillo affected by tinder, with increases in the chlorophyll a/b and the carotenoid/chlorophyll ratios despite the lower carotenoid content.
Mannan treatment negatively affected the chlorophyll content of healthy cv. Tempranillo vines, but diseased vines were unaffected. In Airén, mannans did not alter the pigment content in healthy vines but led to decreased chlorophyll a content in diseased vines. These results do not coincide with those of Dawood et al. [48] who observed increases in all photosynthetic pigments following the application of yeast extract to soybean plants. De Miccolis-Angelini et al. [52] describe the application of yeast extract to grapevines as having a strong enhancing effect on the expression of genes that encode enzymes related to photosynthesis processes.
3.2. Phenol content
Total phenol, total flavonoid, and total phenylpropanoid glycoside (PPG) contents were determined in leaves and grapes of healthy and diseased vines of both varieties, as also were the effects on these contents of the different mannan treatments used. The three groups of effects were similar.
For cv. Tempranillo (Figure 1A,C,E), in healthy leaves the mannans reduced the total contents of all three groups -Phenol, flavonoid and PPG-. In diseased leaves, however, relative to DT0 there were significant increases in these compounds in DT2 but hardly any differences in DT1 and DT3. As evidenced by the comparison of DT0 with HT0, the disease itself did lead to a decrease in the content of these compounds. For cv. Airén (Figure 1B,D,F), the different treatments had little effect on these compounds’ contents. Only the total phenol content in healthy leaves for the dose rate of HT1 showed a slight, but significant, increase. There were no differences in this response between healthy and diseased leaves. The total content of these phenols was unaffected by the disease.
In healthy cv. Tempranillo grapes (Figure 2A,C,E), the mannans caused a decrease in the content of these compounds, the decrease being greater in HT1 and HT2 in the case of the PPGs. There were no differences between HT3 and HT0. In grapes from diseased vines, there were few differences between treatments, only worthy of note being significant increases in phenols and PPGs for DT2 relative to DT0. In the absence of mannans, there were no differences between the contents of these compounds as a consequence of the disease (HT0 and DT0). In healthy cv. Airén grapes (Figure 2B,D,F), mannans strongly reduced the content of all these compounds. The response was somewhat different in grapes from diseased vines, with a decrease in DT1 relative to DT0 in the case of the PPGs, while in DT2 and DT3 the contents were similar to the control, DT0. The disease itself modifies the total content of these compounds, as can be seen in the comparison of DT0 in HT0, with reductions in the content of all three groups – phenols, flavonoids, and PPGs.
In both healthy and diseased cv. Tempranillo grapes, mannan application reduced the content of anthocyanin, the main flavonoid associated with red wine’s colour and astringency (Figure 2G). This content was high for both HT0 and DT0. In HT1, the decrease was slight and non-significant (6%), but more pronounced and significant in HT2 and HT3. In diseased grapes, the decrease in anthocyanin content was strong, with the DT1, DT2, and DT3 values being similar to each other, and all lower than DT0. The disease itself did not alter the grape anthocyanin content since the HT0 and DT0 values were very similar.
For cv. Tempranillo, in leaves and grapes of healthy vines, the addition of mannans negatively affected their phenol content. On the contrary, in diseased leaves, treatments DT1 and DT2 showed recovery of the content of these compounds to values similar to those of healthy vines. Thus, in grapes from mannan-treated vines, the disease caused an increase in the total phenol content, and in grapes from both healthy and diseased vines, the mannans caused a decrease in their anthocyanin content. For cv. Airén however, the mannan treatments had little effect in leaves, whether healthy or diseased, and in grapes led to a decrease in the healthy case, but hardly any effect in the diseased case.
The response of the different phenols does not, in general, coincide with the effects described by other workers for different elicitors. Thus, Sak et al. [71], applying MeJA or P. chlamydospora in vine callus cultures, describe an increase in the production of polyphenols, this effect being much greater with the fungal elicitor. García-Pastor et al. [72] also describe increased ripening, harvest, total phenols, and particularly anthocyanins following the application of low concentrations of MeJA to cv. Magenta and Crimson grapes. An increase in phenols has also been observed [73] in grape cell cultures elicited with two fungi (E. lata and T. atroviride). However, Sgherri et al. [74] describe a decrease in phenols in Ramonda serbica in response to water stress, possibly because these compounds would play an important role against the oxidation caused by the stress, which coincides with our results.
Decreased phenol content due to the effect of tinder has already been described by other workers [23,75]. This could be due to the involvement of phenols in defence reactions, being derived for the synthesis of compounds such as lignin [76]. However, Calzarano et al. [77] and Lima et al. [78] detected an increase in polyphenols in diseased vines. In general, the use of yeast in plants seems to cause an increase in phenol content [47,48,52].
With respect to grape anthocyanin content, Lorrain et al. [75] describe a slight decrease in cv. Cabernet Sauvignon grapes from vines affected by tinder. García et al. [23], however, indicate that tinder does not affect the anthocyanin content in cv. Tempranillo grapes, which results that coincide with those of the present work in DT0. The application of mannans did affect the anthocyanin content causing it to decrease, possibly due to an effect on the route of synthesis of the flavonoids themselves responsible for the synthesis of tannins and anthocyanins, giving rise to a lower content of phenols.
3.3. Polyphenol oxidase (PPO) and Superoxide dismutase (SOD) activities
Figure 3 and Figure 4 show the PPO and SOD activities in leaves and grapes, respectively. With respect to the cv. Tempranillo leaves (Figure 3A), in the healthy leaf case, the addition of mannans increased the PPO activity but not in a dose-dependent manner (HT1 and HT3 were similar, although the values were surprisingly low in HT2). In the diseased leaf case, the addition of mannans also increased this activity, with that of DT0 being lower than that of HT0, evidence of a clear negative effect of the disease. With respect to the cv. Airén leaves, in the healthy case (Figure 3B), PPO activity increased markedly in HT1 and HT2 (a dose-dependent increase), to then decrease in HT3, although with activity levels that were still higher than the control, HT0. In diseased leaves, only DT1 led to increased PPO activity compared with DT0, with DT2 and DT3 leaves presenting values similar to DT0. The disease (DT0) induced increased PPO activity relative to the healthy case (HT0), although with there being great variability in these increases.
The SOD activity in cv. Tempranillo leaves (Figure 3C) increased after the application of mannan to healthy vines. In leaves from diseased vines, mannans seemed to increase this activity slightly, although in some cases without significance. Likewise for cv. Airén, the healthy leaves (Figure 3D) showed SOD activity in response to the increase in the amount of mannans, as was the case for PPO activity. In HT3, although the activity level was higher than that of HT0, there was a decrease with respect to the lower mannan concentrations. In diseased leaves, DT1 and DT2 increased the SOD activity, but not significantly. Comparison of the SOD activity values in HT0 and DT0 leaves shows that disease induced an increase in this activity through increasing the amount of H2O2 by dismutation and elimination of the O2- produced in response to the stress caused by the disease.
For cv. Tempranillo grapes (Figure 4A), in the healthy case, the mannan application led to a slight and non-significant increase in PPO activity. On the contrary, in the case of diseased grapes, it led to decreased PPO activity, especially T3D. The disease itself induced increased PPO activity (DT0 vs HT0). For cv. Airén grapes (Figure 4B, the mannans induced increased PPO activity in healthy grapes. Their application also increased PPO activity in diseased grapes (DT1 vs DT0) but, because of the great variability in the response, this increase was not significant. For DT2 and DT3, no significant differences were observed relative to DT0. In the absence of mannans, diseased grapes (DT0) presented greater PPO activity than healthy grapes (HT0).
In healthy cv. Tempranillo grapes (Figure 4C), an increase in SOD activity was observed following the addition of the first two doses of mannans (191% and 153% for HT1 and HT2, respectively). For HT3, there was no significant difference relative to HT0. In grapes from diseased vines, no significant differences were observed between DT1, DT2, and DT3 and the DT0 control. In cv. Airén grapes (Figure 4D), the SOD activity increased slightly, but not significantly, for those from healthy vines in response to the mannan application. For those from diseased vines, while the first doses of mannan affected this activity little, DT3 caused significant activation. Comparing the levels of SOD activity in healthy and diseased grapes of both varieties without application of mannans (HT0 vs DT0), one can say that disease caused a significant increase in SOD activity.
Previous studies have described increased PPO activity in grapevines affected by tinder [23,79] and, as in this case, in grapes from diseased vines [23,80]. Rusjan et al. [81] describe an increase in PPO activity in vine leaves of cv. Chardonnay as a consequence of Bois Noir infection. Shi et al. [82] in Cabernet Sauvignon describe an increase in PPO activity due to Colomerus vitis infection, although this activation decreases as the infection develops. Similarly, our results show that a decrease in PPO activity occurs in leaves due to the effect of the disease, which may be due to the disease being in an advanced stage of development.
The addition of mannans to healthy vines caused an increase in PPO activity, which would indicate the development of stress. Singh et al. [83] describe a similar response after the application of chitosan to two grapevine varieties, with an increase in the expression of PPO genes in both leaves and grapes. The effect of mannan application to diseased vines was variable. Thus, DT1 increased PPO activity in the leaves of both varieties but, in the grapes, decreased it in cv. Tempranillo and left it unchanged in cv. Airén. With respect to the effect of the disease with no mannan application, in leaves there was either no variation (Airén) or it was very slight (Tempranillo), while in grapes of both varieties there was an increase.
In practically all the cases that have been reported previously, the effect of mannans in healthy vines is to increase SOD activity as an indicator of stress. This increase allows excess ROS to be controlled and eliminated [51]. Disease and other stressors induce SOD activation in grapevine leaves [84,85].
The finding that in Airén leaves and Tempranillo grapes SOD activity increased at lower mannan concentrations and decreased at higher concentrations could be compared with the results of Shi et al. [82] for Cabernet Sauvignon in which the increased SOD activity in response to Colomerus vitis infection declined in more advanced stages of the infection.
3.4. Gene expression
The expression patterns of the chalcone synthase (CHS1, CHS3), polyphenol ammonia-lyase (PAL), PPO, and SOD genes were determined in leaves and grapes of both varieties with the different treatments. Figure 5 shows the healthy leaf results. In Tempranillo (Figure 5A), there was a very significant increase in the expression patterns of all the enzymes analysed with the HT2 treatment when compared with the untreated case. There was also a significant increase in the CHS1 pattern in HT1 relative to the control plant, while in HT3 there was no difference with respect to HT0. CHS3 in HT1 was expressed less than the control (HT0), and in HT3 there was practically no expression. The PAL expression also decreased in HT1 and HT3. However, the PPO and SOD expressions were activated in all treatments, although the difference between HT1 and HT0 was not significant. In Airén (Figure 5B), there were significant increases in the expression patterns of all the enzymes analysed with all treatments, especially in HT2 compared with HT0.
This increase in the PPO and SOD activities was also observed in the Airén variety, as has been described above (Figure 3B,D). Again, the increase was greatest in HT2. There was not such an obvious relationship between expression and activity in cv. Tempranillo (Figure 3A,C), although the mannan treatments generally activated these activities in most cases, even if just slightly.
Analysing the changes in the expression level of CHS1 and CHS3, PAL, PPO, and SOD of healthy vines (H) as against diseased vines (D) without and with mannan treatments, it is clear that, both in Tempranillo (Figure 5C) and in Airén (Figure 5D), the infection causes a significant increase in the expression pattern of all the enzymes studied when compared to the leaves of healthy vines. In Tempranillo leaves, there is no correspondence between the expression and the measured activities. The disease causes a decrease in PPO activity (Figure 3A), while SOD activity is not affected (Figure 3C). However, in Airén leaves from diseased vines a correspondence is observed between the increases in activity and expression, both for PPO activity (Figure 3B), although it does not show significance, and for SOD activity (Figure 3D).
In Tempranillo, treatment with the highest dose of mannans (DT3) always mitigated the increase in expression caused by the infection in all the enzymes measured. With the treatments DT1 and DT2, the results differed according to the enzyme. In Airén, the foliar treatment with mannans seemed capable of alleviating the stress caused by GTDs. In particular, the expression level of the enzymes evaluated in treated GTD-affected vines was still significantly higher than that of healthy vines, but lower than that found in untreated diseased vines. The mitigating effect of mannans was not seen in the enzymatic activities measured in diseased leaves of cv. Tempranillo: PPO increased due to the effect of mannans (Figure 3A) and SOD was unaltered (Figure 3C). Similar results were obtained in Airén (Figure 3B,D).
The expression patterns of these enzymes in grapes from healthy vines of both varieties with and without mannan treatment differed from those obtained in leaves. In Tempranillo (Figure 6A), all the enzymes measured with the HT1 and HT3 treatments showed increased expression with respect to HT0 with one exception being that there was no significant difference between HT1 and HT0 in the expression of CHS3. With HT2 however, there was a decrease in CHS1 expression relative to the control, but no such difference for CHS3, PAL, PPO, or SOD. In Airén (Figure 6B), each enzyme had a different response pattern to the different mannan doses. Thus, CHS1 had increased expression in all treatments, but to a greater extent as the amount of mannans provided was lower. CHS expression decreased slightly in HT1, but increased in HT2 and HT3. PAL expression increased slightly in HT1, decreased in HT2 (to close to the HT0 value), and increased considerably in HT3. PPO expression increased in HT1 but decreased in HT2 and HT3 relative to HT0. SOD expression was practically unaffected in HT1 and HT3, but decreased in HT2.
Comparing the PPO and SOD expression results with those of these enzymes’ activities described above, one observes that in healthy Tempranillo grapes the PPO activity (Figure 4A) was not significantly affected by mannans, but the expression was. The SOD activity (Figure 4C) showed a closer relationship with what was obtained in gene expression, with in general there being activation in both cases. In Airén, the maximum PPO expression was in HT1, decreasing in HT2 and HT3. The activation of PPO activity by mannans (Figure 4B) was similar in all treatments. Neither the activity nor the expression of SOD (Figure 4D) were affected by mannan treatment.
Figure 6C,D shows the expression of the enzymes CHS1 and 3, PAL, PPO, and SOD in grapes from GTD-affected vines with and without mannan application versus grapes from healthy vines. For Tempranillo (Figure 6C), the expression was greater in diseased vines, with or without mannan application, than in control vines. In this variety, the DT1 expression values were similar to those of untreated diseased vines except for PPO where DT1 expression fell significantly. DT2 decreased the expression of all the enzymes studied, although without reaching the levels of healthy grapes. In DT3, the results depended on the enzyme: the expression levels of CHS1, PAL, and SOD fell compared with the diseased grapes, the CHS3 expression levels were similar to those of diseased grapes, DT0, and there was a considerable increase in the case of PPO. In Airén (Figure 6D), the expression levels in infected grapes did not increase in all cases relative to those of healthy grapes. Instead, the said levels decreased for PPO and SOD. The results obtained with mannan treatments were very variable. Thus, DT1 increased CHS3, PAL, and SOD expression relative to untreated diseased grapes (DT0), decreased that of CHS1, and left that of PPO unchanged. Regarding DT2 relative to DT0, there was a decrease in the expression of CHS1, CHS3, and PAL, no change in that of PPO, and an increase in that of SOD. Finally, relative to DT1 and DT2, DT3 induced increased expression except for CHS3 whose expression declined considerably, and, relative to DT0, it decreased the expression of CHS3, increased that of PAL, PPO, and SOD, and left that of CHS1 unchanged.
The expression of PPO in Tempranillo grapes is comparable with the results obtained for the activity of this enzyme (Figure 4A). An increase in activity, as in expression, was observed in diseased DT0 grapes relative to healthy HT0 ones. Treatment with mannans lessened both this increase and the expression in DT1 and DT2. Only DT3 caused a decrease in activity, while the gene expression was very high. In Airén grapes, there was little parallelism between PPO expression and activity (Figure 4B). The disease caused an increase in activity that did not correspond to an increase in expression relative to healthy grapes. The DT1 and DT2 mannan treatments affected neither the expression nor the activity with respect to DT0, while DT3 induced an increase in PPO expression but not in activity.
Regarding the SOD expression and activity of Tempranillo grapes (Figure 4C), the infection caused an increase in both compared with healthy grapes. Mannan treatment did not change the activity compared with untreated diseased grapes, but it did cause a decrease in expression in DT2 and DT3 (DT1 was unaffected). In Airén, while the infection caused activation of SOD activity (Figure 4D), this was not due to increased expression of the enzyme. Mannan treatment increased both expression and activity relative to untreated diseased grapes.
These expression results indicate that, with just a few exceptions, mannans cause increased expression of the enzymes measured in both leaves and grapes of healthy vines. Furthermore, the expression of these enzymes is greater in diseased vines than in healthy ones, and a consequence of the addition of mannans to diseased vines is in most cases to lessen the increase in the expression of these genes.
Coherent with these results, De Miccolis et al. [52] indicate that a yeast derivative (cerevisane) is effective against mildew in grapevines, and produces an increase in various genes including those, such as PAL and SOD, which encode proteins related to pathogenesis. Chitosan also has been found to activate PAL, PPO, and SOD genes in two grapevine varieties in both their leaves and grapes [83,86].
4. Conclusions
Healthy vines (H) of both varieties respond to mannan treatment in a stress-like manner. Thus, in cv. Tempranillo, for both leaves and grapes all the parameters measured are affected (chlorophylls, phenols, CHS1, CHS3, PAL, PPO and SOD expression and SOD and PPO activities), at least with some mannan treatment. Neither the PPO activity of grapes nor the carotenoid content of leaves are affected. In cv. Airén leaves, these treatments do not affect the phenols or photosynthetic pigment contents, but they do affect CHS1, CHS3, PAL, PPO and SOD expression, as well as PPO and SOD activity. On the contrary, in the grapes, phenol content, PPO activity, and the expression of CHS1, CHS3, PAL, and PPO are altered, but not that of SOD.
For diseased vines (D), mannan addition is more positive in leaves than in grapes. Of the varieties, the greater response was observed in cv. Tempranillo. Thus, in its leaves, mannans increased the phenol content, PPO activity, and CHS1 and PPO expression, with decreases in CHS3, PAL, and SOD expression. In its grapes, the phenol (but not the anthocyanin) content increased, PPO activity decreased, and the expression of all enzymes declined (at least, with some mannan treatments). For cv. Airén, in its leaves, mannans led to decreased chlorophyll a and carotenoids, increased PPO activity, and decreased expression of CHS3, PAL, and PPO, while CHS1 and SOD were unaffected. In its grapes, increased SOD activity was observed, with the expression patterns of the genes studied being very variable between treatments, possibly due to the degree of affectation of the grapes as an effect of the development of the disease.
In summary, mannan treatment may represent one form of preventing or mitigating the attack of pathogenic fungi that trigger GTDs, although this treatment will depend on such factors as the cultivar involved and the vines’ phytosanitary status (healthy vs diseased). Also, in the varieties studied, due to the degree of affectation of the grapes because of the development of the disease, it was hard to establish a correlation between the fruit and the leaf organ in the activity and gene expression results of enzymes involved in antioxidant action.
Author Contributions
Conceptualization, A.O., F.E. and I.G.; methodology, A.O., F.E. and I.G.; software, A.O.; validation, A.O., J.A.G., J.M., F.E. and I.G.; formal analysis, A.O., F.E. and I.G.; investigation, A.O., J.A.G., J.M., F.E. and I.G.; data curation, A.O.; writing—original draft preparation, A.O., F.E. and I.G.; writing—review and editing, A.O., F.E. and I.G.; funding acquisition, F.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was made possible thanks to the Junta de Extremadura (GR21112) and the “Bodegas Yuntero, S.C.” (074/19).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank “Bodegas Yuntero, S.C.” for the application and monitoring of the treatments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abhilash, P.; Tripathi, V.; Edrisi, S.A.; Dubey, R.K.; Bakshi, M.; Dubey, P.K.; Singh, H.; Ebbs, S.D. Sustainability of crop production from polluted lands. Energy Ecol Environ. 2016, 1, 54–65. [Google Scholar] [CrossRef]
- Pandey, G. Challenges and future prospects of agri-nanotechnology for sustainable agriculture in India. Environ Technol Innov. 2018, 11, 299–307. [Google Scholar] [CrossRef]
- Acharya, A.; Pal, P.K. Agriculture nanotechnology: translating research outcome to field applications by influencing environmental sustainability. NanoImpact. 2020, 19, 100232. [Google Scholar] [CrossRef]
- Safdar, M.; Kim, W.; Park, S.; Gwon, Y.; Kim, Y.O.; Kim, J. Engineering plants with carbon nanotubes: a sustainable agriculture approach. J. Nanobiotechnology. 2022, 20(1), 1–30. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, P.; Dapkekar, A.; Oak, M.D.; Paknikar, K.M.; Rajwade, J.M. Zinc complexed chitosan/TPP nanoparticles: a promising micronutrient nanocarrier suited for foliar application. Carbohydr Polym. 2017, 165, 394–401. [Google Scholar] [CrossRef] [PubMed]
- FAO. The future of food and agriculture—Trends and challenges. Annual Report. Rome 2017. [Google Scholar]
- Panpatte, D.G.; Jhala, Y.K.; Shelat, H.N.; Vyas, R.V. Nanoparticles: the next generation technology for sustainable agriculture. In Microbial inoculants in sustainable agricultural productivity. Vol. 2. Functional application; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: New Delhi, 2016; pp. 289–300. [Google Scholar] [CrossRef]
- Kah, M.; Tufenkji, N.; White, J.C. Nano-enabled strategies to enhance crop nutrition and protection. Nat Nanotechnol. 2019, 14, 532–40. [Google Scholar] [CrossRef] [PubMed]
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and challenges for nanotechnology in the agri-tech revolution. Nat Nanotechnol. 2019, 14, 517–22. [Google Scholar] [CrossRef]
- Kgang, I.E.; Klein, A.; Husselmann, L.; Nkomo, A.; Mathabe, P. M.; Belay, Z.A.; Caleb, O.J. Bioassays and proteomics as early detection tools in postharvest management of table grapes (Vitis vinifera L.) diseases–A Review. Food Biosci, 2023; 102645. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Grapes and human health: A perspective. J. Agric. Food Chem. 2008, 56, 6777–6784. [Google Scholar] [CrossRef]
- Mesguida, O.; Haidar, R.; Yacoub, A.; Dreux-Zigha, A.; Berthon, J.Y.; Guyoneaud, R.; Attard, E.; Rey, P. Microbial Biological Control of Fungi Associated with Grapevine Trunk Diseases: A Review of Strain Diversity, Modes of Action, and Advantages and Limits of Current Strategies. J. Fungi, 2023; 9, 638. [Google Scholar] [CrossRef]
- Guerin-Dubrana, L.; Fontaine, F.; Mugnai, L. Grapevine Trunk disease in European and Mediterranean vineyards: Occurrence, distribution and associated disease-affecting cultural factors. Phytopathol. Mediterr. 2019, 58, 49–71. [Google Scholar] [CrossRef]
- Cobos, R.; Ibañez, A.; Diez-Galán, A.; Calvo-Peña, C.; Ghoreshizadeh, S.; Coque, J.J.R. The grapevine microbiome to the rescue: implications for the biocontrol of trunk diseases. Plants. 2022, 11(7), 840. [Google Scholar] [CrossRef] [PubMed]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R.; Urbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases with Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed]
- Pouzoulet, J.; Pivovaroff, A.L.; Santiago, L.S.; Rolshausen, P.E. Can vessel dimension explain tolerance toward fungal vascular wilt diseases in woody plants? lessons from dutch elm disease and Esca disease in grapevine. Front. Plant Sci. 2014, 5, 253. [Google Scholar] [CrossRef] [PubMed]
- Ouadi, L.; Bruez, E.; Bastien, S.; Yacoub, A.; Coppin, C.; Guérin-Dubrana, L.; Fontaine, F.; Domec, J.C.; Rey, P. Sap flow disruption in grapevine is the early signal predicting the structural, functional, and genetic responses to Esca disease. Front. Plant Sci. 2021, 12, 695846. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gonzalez, G.; Sebestyen, D.; Petit, E.; Jellison, J.; Mugnai, L.; Lee, N.; Farine, S.; Bertsch, C.; Goodell, B. Oxygen Radical-Generating Metabolites Secreted by Eutypa and Esca Fungal Consortia: Understanding the Mechanisms Behind Grapevine Wood Deterioration and Pathogenesis. Front. Plant Sci. 2022, 13, 921961. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Amat, A.; Solano, F. A pluripotent polyphenol oxidase from the melanogenic marine Alteromonas sp shares catalytic capabilities of tyrosinases and laccases. Biochem. Biophys. Res. Commun. 1997, 240, 787–792. [Google Scholar] [CrossRef]
- Abou-Mansour, E.; Polier, J.; Pezet, R. Tabacchi, R. Purification and partial characterisation of a 60 KDa laccase from Fomitiporia mediterranea. Phytopathol. Mediterr. 2009, 48, 447–453. [Google Scholar]
- García, J.A.; Garrido, I.; Ortega, A.; Del Moral, J.; Llerena, J.L.; Espinosa, F. Physiological and Molecular Responses of Vitis vinifera cv. Tempranillo Affected by Esca Disease. Antioxidants. 2022, 11(9), 1720. [Google Scholar] [CrossRef]
- Aroca, A.; Gramaje, D.; Armengol, J.; García-Jiménez, J.; Raposo, R. Evaluation of the grapevine nursery propagation process as a source of Phaeoacremonium spp. and Phaeomoniella chlamydospora and occurrence of trunk disease pathogens in rootstock mothervines in Spain. Eur. J. Plant Pathol. 2010, 126, 165–174. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J. Fungal Trunk Pathogens in the Grapevine Propagation Process: Potential Inoculum Sources, Detection, Identification, and Management Strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef] [PubMed]
- Agustí-Brisach, C.; Gramaje, D.; García-Jiménez, J.; Armengol, J. Detection of black-foot disease pathogens in the grapevine nursery propagation process in Spain. Eur. J. Plant Pathol. 2013, 137, 103–112. [Google Scholar] [CrossRef]
- Luque, J.; Elena, G.; Garcia-Figueres, F.; Reyes, J.; Barrios, G.; Legorburu, F.J. Natural infections of pruning wounds by fungal trunk pathogens in mature grapevines in Catalonia (Northeast Spain). Aust. J. Grape Wine Res. 2014, 20, 134–143. [Google Scholar] [CrossRef]
- Trouvelot, S.; Héloir, M.C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed]
- Zang, H.; Xie, S.; Zhu, B.; Yang, X.; Gu, C.; Hu, B. , GAo, T.; Chen, Y.; Gao, X. Mannan oligosaccharides trigger multiple defence responses in rice and tobacco as a novel danger-associated molecular pattern. Mol. Plant Pathol. 1067. [Google Scholar] [CrossRef]
- Liu, N.; Shen, H.; Zhang, F.; Liu, X.; Xiao, Q.; Jiang, Q.; Tan, B.; Ma, X. Applications and prospects of functional oligosaccharides in pig nutrition: a review. Anim. Nutr. 2023, 13, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Laere, V.; Hartemink, R.; Bosveld, M.; Schols, H.; Voragen, A. Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J. Agr. Food Chem. 2000, 48, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M; Pugin, A. Laminarin elicitors defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef]
- Paulert, R.; Ebbinghaus, D.; Urlass, C.; Moerschbacher, B.M. Priming of the oxidative burst in rice and wheat cell cultures by ulvan, a polysaccharide from green macroalgae, and enhanced resistance against powdery mildew in wheat and barley plants. Plant Pathol. 2010, 59, 634–642. [Google Scholar] [CrossRef]
- Li, Y.; Yin, H.; Wang, Q.; Zhao, X.M.; Du, Y.G.; Li, F.L. Oligochitosan-induced Brassica napus L. production of NO and H2O2 and their physiological function. Carbohydr. Polym. 2009, 75, 612–617. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, X.M; Du, Y.G. Oligochitosan: a plant diseases vaccine. A review. Carbohyd. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Kaida, R.; Sugawara, S.; Negoro, K.; Maki, H.; Hayashi, T.; Kaneko, T.S. Acceleration of cell growth by xyloglucan oligosaccharides in suspension-cultured tobacco cells. Mol. Plant. 2010, 3, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Pontiggia, D.; Raggi, S.; Cheng, Z.; Scaloni, F.; Ferrari, S.; Ausubel, F.; Cervone, F.; Lorenzo, G. Plant immunity triggered by engineered in vivo release of oligogalacturonides, damage-associated molecular patterns. Proc. Natl. Acad. Sci. USA, 2015, 112, 5533–5538. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Di Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Nascimento, T.; Rego, C.; Oliveira, H. Potential use of chitosan in the control of grapevine trunk diseases. Phytopathol. Mediterr. 2007, 46, 218–224. [Google Scholar] [CrossRef]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; Lorenzo, G.D.; Ferrari, S.; Ausubel, F.; Dewdney, J. Activation of defense response pathways by OGs and Flg22 elicitors in Arabidopsis seedlings. Mol. Plant. 2008, 1, 423–445. [Google Scholar] [CrossRef]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; Lorenzo, G.D. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc. Natl. Acad. Sci. USA, 2010, 107, 9452–9457. [Google Scholar] [CrossRef]
- Galletti, R.; Ferrari, S.; Lorenzo, G. Arabidopsis MPK3 and MPK6 play different roles in basal and oligogalacturonide- or flagellin-induced resistance against Botrytis cinerea. Plant Physiol. 2011, 157, 804–814. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A review of the use of biostimulants in the vineyard for improved grape and wine quality: effects on prevention of grapevine diseases. J Sci Food Agric. 2019, 99(3), 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Preety. Purification and properties of plant chitinases: A review. J. Food Biochem. 2019, 43, e12762. [Google Scholar] [CrossRef]
- Shehata, S.; Fawzy, Z.; El-Ramady, H. Response of Cucumber Plants to Foliar Application of Chitosan and Yeast under Greenhouse Conditions. Aust. J. Basic Appl. Sci. 2012, 4, 63–71. [Google Scholar]
- Yan, Q.; Shi, M.; Ng, J.; Wu, J. Elicitor-induced rosmarinic acid accumulation and secondary metabolism enzyme activities in Salvia miltiorrhiza hairy roots. Plant Sci. 2006, 170, 853–858. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Lethy, S.R.; Sadak, M. Role of methanol and yeast in improving growth, yield, nutritive value and antioxidants of soybean. World Appl. Sci. J. 2013, 26, 6–14. [Google Scholar] [CrossRef]
- Narusaka, M.; Minami, T.; Iwabuchi, C.; Hamasaki, T.; Takasaki, S.; Kawamura, K.; Narusaka, Y. Yeast cell wall extract induces disease resistance against bacterial and fungal pathogens in Arabidopsis thaliana and Brassica crop. PLoS One. 2015, 10(1), e0115864. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Lin, M.; Fu, D.; Yang, J.; Huang, Y.; Zheng, X.; Yu, T. Yeast cell wall induces disease resistance against Penicillium expansum in pear fruit and the possible mechanisms involved. Food Chem. 2018, 241, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, Y.; Huang, G. Preparation and antioxidant activities of important traditional plant polysaccharides. Int. J. Biol. Macromol. 2018, 111, 780–786. [Google Scholar] [CrossRef]
- De Miccolis Angelini, R.M.; Rotolo, C.; Gerin, D.; Abate, D.; Pollastro, S.; Faretra, F. Global transcriptome analysis and differentially expressed genes in grapevine after application of the yeast-derived defense inducer cerevisane. Pest Manag. Sci. 2019, 75(7), 2020–2033. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Portu, J.; Santamaría, P.; López, R.; Garde-Cerdán, T. Effects on grape amino acid concentration through foliar application of three different elicitors. Food Res Int. 2017, 99(Pt 1), 688–692. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Blei-Holder, H.; Klose, R.; Meier, U.; Weber, E. Phänologische Entwicklungsstadien der Weinrebe (Vitis vinifera L. ssp. vinifera). Codierung und Beschreibung nach der erweiterten BBCH-Skala. Vitic. Enol. Sci 1994, 49, 66–70. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Journal Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Singleton, V.L.; Salgues, M.; Zaya, J.; Troudsale, E. Caftaric acid disappearance and conversion to products of enzymatic oxidation in grape must and wine. Am. J. Enol. Vitic. 1985, 36, 50–56. [Google Scholar] [CrossRef]
- Kim, D.; Jeong, S.W.; Leo, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Gálvez, M.; Martín-Cordero, C.; Houghton, P.J.; Ayuso, M.J. Antioxidant activity of methanol extracts obtained from Plantago species. J. Agric. Food Chem. 2008, 53, 1927–1933. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Wrolstadm, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1-F1.2.13. [CrossRef]
- MohdMaidin, N.; Oruna-Concha, M.J.; Jauregi, P. Surfactant TWEEN20 provides stabilisation effect on anthocyanins extracted from red grape pomace. Food Chem. 2019, 271, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Walker, J. Polarographic and Spectrophotometric Assay of Diphenol Oxidases (Polyphenol Oxidase). In Current Protocols in Food Analytical Chemistry; Wiley, 2001; pp. C4.1.1–C4.1.15. [Google Scholar] [CrossRef]
- Beaucham, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Pilati, S.; Perazzolli, M.; Malossini, A.; Cestaro, A.; Demattè, L.; Fontana, P.; Dal Ri, A.; Viola, R.; Velasco, R.; Moser, C. Genome-wide transcriptional analysis of grapevine berry ripening reveals a set of genes similarly modulated during three seasons and the occurrence of an oxidative burst at vèraison. BMC Genomics 2007, 8, 428. [Google Scholar] [CrossRef]
- Gamm, M.; He´loir, M.C.; Kelloniemi, J.; Poinssot, B.; Wendehenne, D.; Adrian, M. Identification of reference genes suitable for qRT-PCR in grapevine and application for the study of the expression of genes involved in pterostilbene synthesis. Mol. Genet. Genom. 2011, 285, 273–285. [Google Scholar] [CrossRef]
- Joseph, J.T.; Poolakkalody, N.J.; Shah, J.M. Plant reference genes for development and stress response studies. J. Biosci. 2018, 43, 173–187. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, C.; Zhu, P.; Liu, F. Effects of Antimony Stress on Photosynthesis and Growth of Acorus calamus. Front Plant Sci. 2018, 9, 579. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.N.; Vaillant, N.; Boulay, M.; Clément, C.; Fontaine, F. Alteration of Photosynthesis in Grapevines Affected by Esca. Phytopathology. 2006, 96, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Fontaine, F.; Castaño, F.J.; Songy, A.; Roda, R.; Vallet, J.; Ferrer-Gallego, R. Specific profile of Tempranillo grapevines related to Esca-leaf symptoms and climate conditions. Plant Physiol. Biochem. 2019, 135, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Fragoeiro, S.; Phillips, A. Physiological response of grapevine cultivars and a rootstock to infection with Phaeoacremonium and Phaeomoniella isolates: an in vitro approach using plants and calluses. Sci. Hortic. 2005, 103, 187–198. [Google Scholar] [CrossRef]
- Sák, M.; Dokupilová, I.; Mihálik, D.; Lakatosová, J.; Gubisová, M.; Kraic, J. Elicitation Phenolic Compounds in Cell Culture of Vitis vinifera L. by Phaeomoniella chlamydospora. Nova Biotechnol. et Chim. 2014, 13, 162–171. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Serrano, M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Zapata, P. Methyl jasmonate effects on table grape ripening, vine yield, berry quality and bioactive compounds depend on applied concentration. Sci. Hortic. 2019, 247, 380–389. [Google Scholar] [CrossRef]
- Mutawila, C.; Stander, C.; Halleen, F.; Vivier, M.A.; Mostert, L. Response of Vitis vinifera cell cultures to Eutypa lata and Trichoderma atroviride culture filtrates: expression of defence-related genes and phenotypes. Protoplasma. 2017, 254, 863–879. [Google Scholar] [CrossRef] [PubMed]
- Sgherri, C.; Stevanovic, B.; Navari-Izzo, F. Role of phenolics in the antioxidative status of the resurrection plant Ramonda serbica during dehydration and rehydration. Physiol. Plant. 2004, 122, 478–485. [Google Scholar] [CrossRef]
- Lorrain, B.; Ky, I.; Pasquier, G.; Jourdes, M.; Dubrana, L.G.; Gény, L.; Rey, P.; Donéche, B.; Teissedre, P.L. Effect of Esca disease on the phenolic and sensory attributes of Cabernet Sauvignon grapes, musts and wines. Aust. J. Grape Wine Res. 2012, 18, 64–72. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Vidigal, P.; Amâncio, S. Oxidative stress homeostasis in grapevine (Vitis vinifera, L.). Front. Environ. Sci. 2015, 3, 20. [Google Scholar] [CrossRef]
- Calzarano, F.; Seghetti, L.; Del Carlo, M.; Cichelli, A. Effect of esca on the quality of berries, musts and wines. Phytopathol. Mediterr. 2004, 43, 125–135. [Google Scholar] [CrossRef]
- Lima, M.R.M.; Felgueiras, M.L.; Cunha, A.; Chicau, G.; Ferreres, F.; Dias, A.C.P. Differential phenolic production in leaves of Vitis vinifera cv. Alvarinho affected with esca disease. Plant Physiol. Biochem. 2017, 112, 45–52. [Google Scholar] [CrossRef]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Mercier, L.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Clément, C.; Fontaine, F. Physiological Changes in Green Stems of Vitis vinifera L. cv. Chardonnay in Response to Esca Proper and Apoplexy Revealed by Proteomic and Transcriptomic Analyses. J. Proteome Res. 2012, 11, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, G.; Lapaillerie, D.; Vilain, S.; Dupuy, J.W.; Lomenech, A.M.; Clavero, S.; Gény, L.; Bonneu, M.; Teissedre, P.L.; Donèche, B. Impact of foliar symptoms of “Esca proper” on proteins related to defense and oxidative stress of grape skins during ripening. Proteomics. 2013, 13, 108–118. [Google Scholar] [CrossRef]
- Rusjan, D.; Halbwirth, H.; Stich, K.; Mikulic-Petkovsek, M.; Veberic, R. Biochemical response of grapevine variety ‘Chardonnay’ (Vitis vinifera L.) to infection with grapevine yellows (Bois noir). Eur. J. Plant Pathol. 2012, 134, 231–237. [Google Scholar] [CrossRef]
- Shi, W.; He, W.; Zhang, Z.; Sun, J.; Zhu, C.; Liu, Z.; Xu, Y.; Zhao, B. Study on the Resistance of ‘Cabernet Sauvignon’ Grapevine with Different Rootstocks to Colomerus vitis. Sustainability 2022, 14, 15193. [Google Scholar] [CrossRef]
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ’Touriga Franca’ and ’Tinto Cão’) Tissues. Antioxidants (Basel) 2019, 8, 525. [Google Scholar] [CrossRef] [PubMed]
- Ozden, M.; Demirel, U.; Kahraman, A. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Hortic. 2009, 119, 163–168. [Google Scholar] [CrossRef]
- Sgherri, C.; Ranieri, A.; Quartacci, M.F. Antioxidative responses in Vitis vinifera infected by grapevine fanleaf virus. J Plant Physiol. 2013, 170(2), 121–128. [Google Scholar] [CrossRef]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306, polluted lands. Energy Ecol Environ. 2016, 1, 54–65. DOI: 10.1007/s40974-016-0007-x. [Google Scholar] [CrossRef]
Figure 1.
Total phenol (A,B) flavonoid (C,D) and PPGs (E,F) in leaves of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 1.
Total phenol (A,B) flavonoid (C,D) and PPGs (E,F) in leaves of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
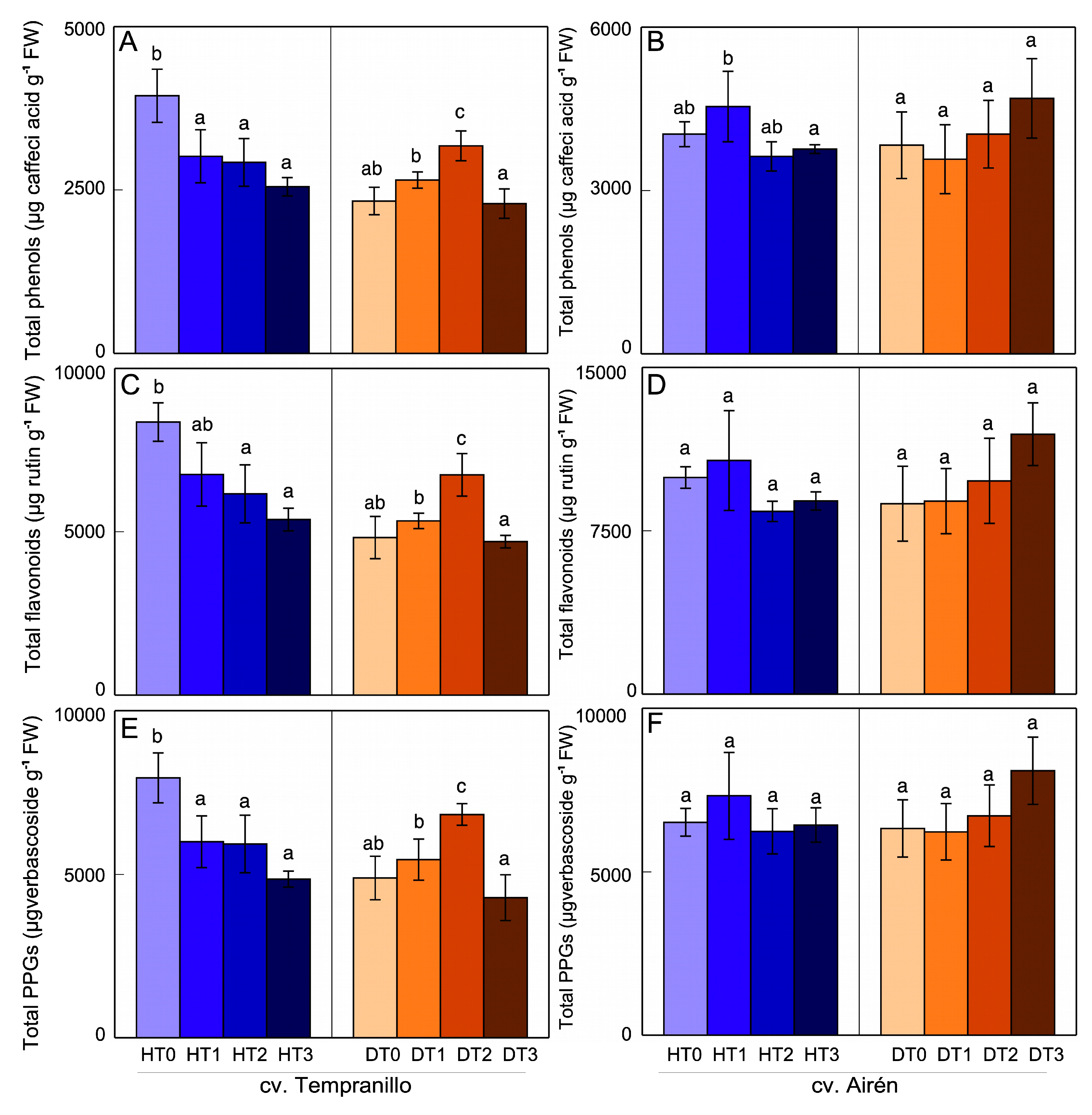
Figure 2.
Total phenol (A,B) flavonoid (C,D), PPGs (E,F) and anthocyanin content (G) in grapes of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 2.
Total phenol (A,B) flavonoid (C,D), PPGs (E,F) and anthocyanin content (G) in grapes of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).

Figure 3.
PPO (A,B) and SOD (C,D) activities in leaves of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 3.
PPO (A,B) and SOD (C,D) activities in leaves of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
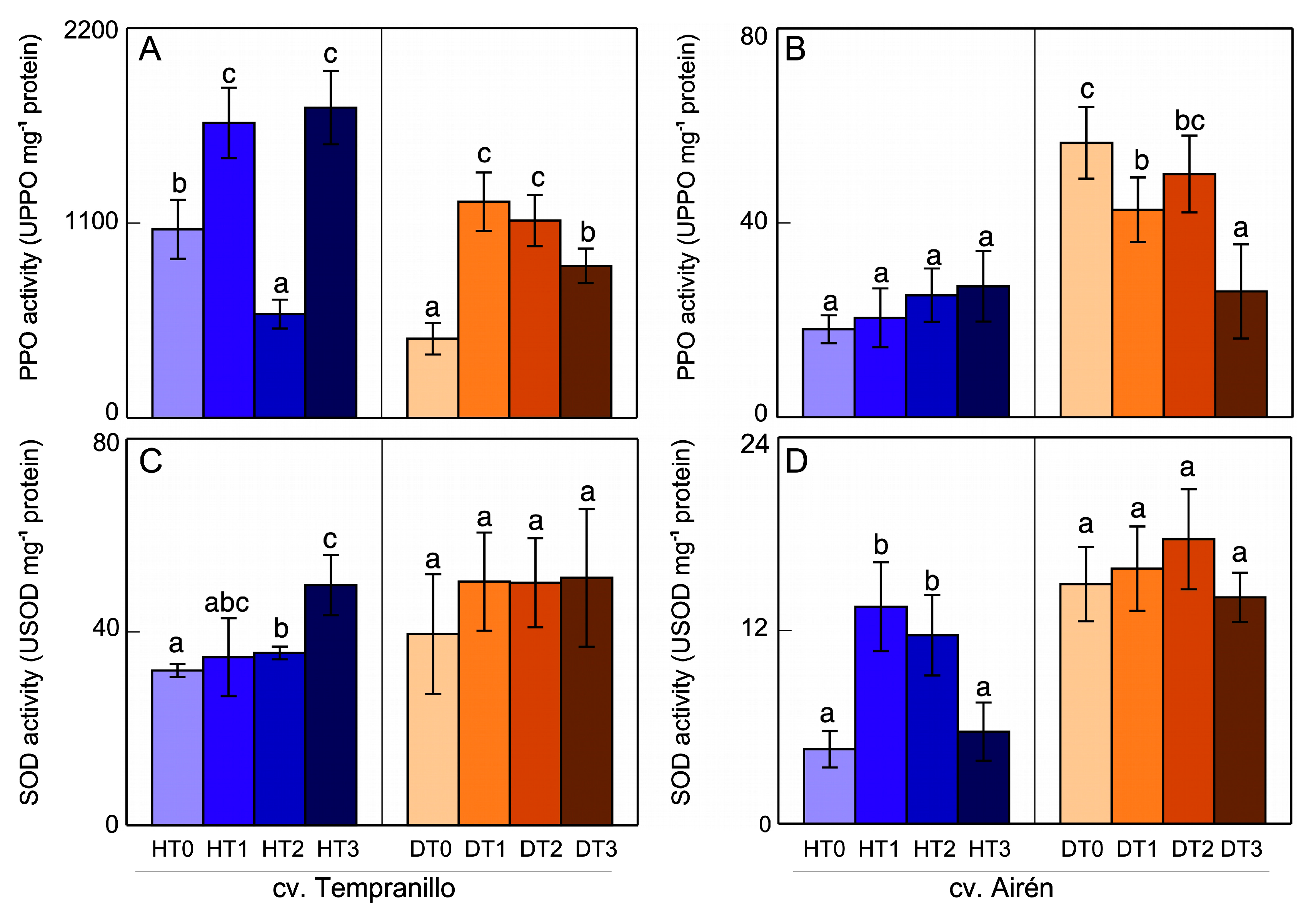
Figure 4.
PPO (A,B) and SOD (C,D) activities in grapes of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 4.
PPO (A,B) and SOD (C,D) activities in grapes of the two grapevine varieties healthy (H) and diseased (D), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (bars of each grapevine cultivar marked with the same letter do not differ significantly according to a parametric test of one-way ANOVA).
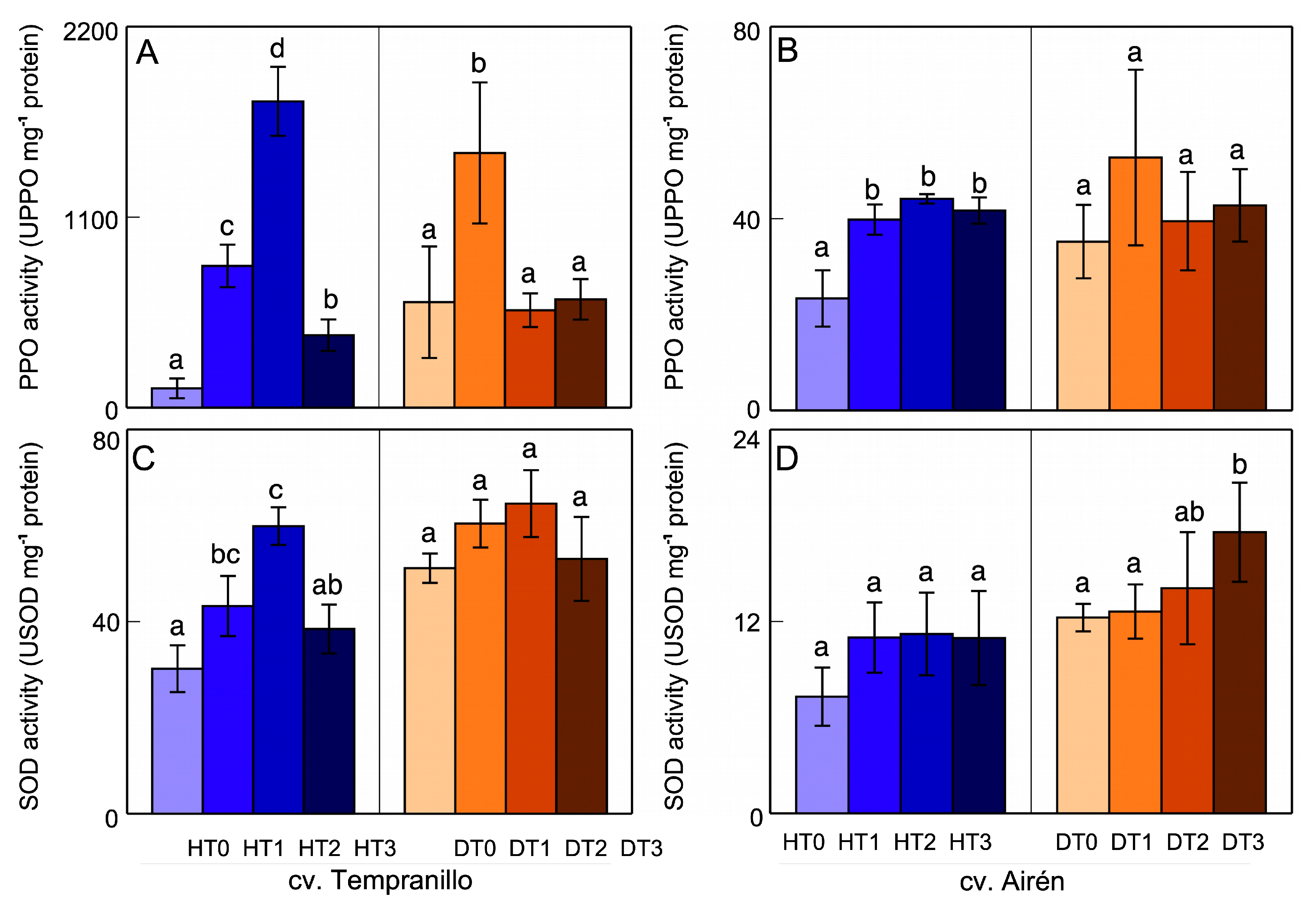
Figure 5.
(A,B) Transcript accumulation in leaves of the two grapevine varieties healthy (H), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). (C,D) Transcript accumulation in leaves of the two grapevine varieties healthy without treatment of manans (HT0) and diseased treated with the different doses of manans used. Quantitative PCR in CHS1, CHS3, PAL, PPO and SOD (the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 5.
(A,B) Transcript accumulation in leaves of the two grapevine varieties healthy (H), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). (C,D) Transcript accumulation in leaves of the two grapevine varieties healthy without treatment of manans (HT0) and diseased treated with the different doses of manans used. Quantitative PCR in CHS1, CHS3, PAL, PPO and SOD (the same letter do not differ significantly according to a parametric test of one-way ANOVA).
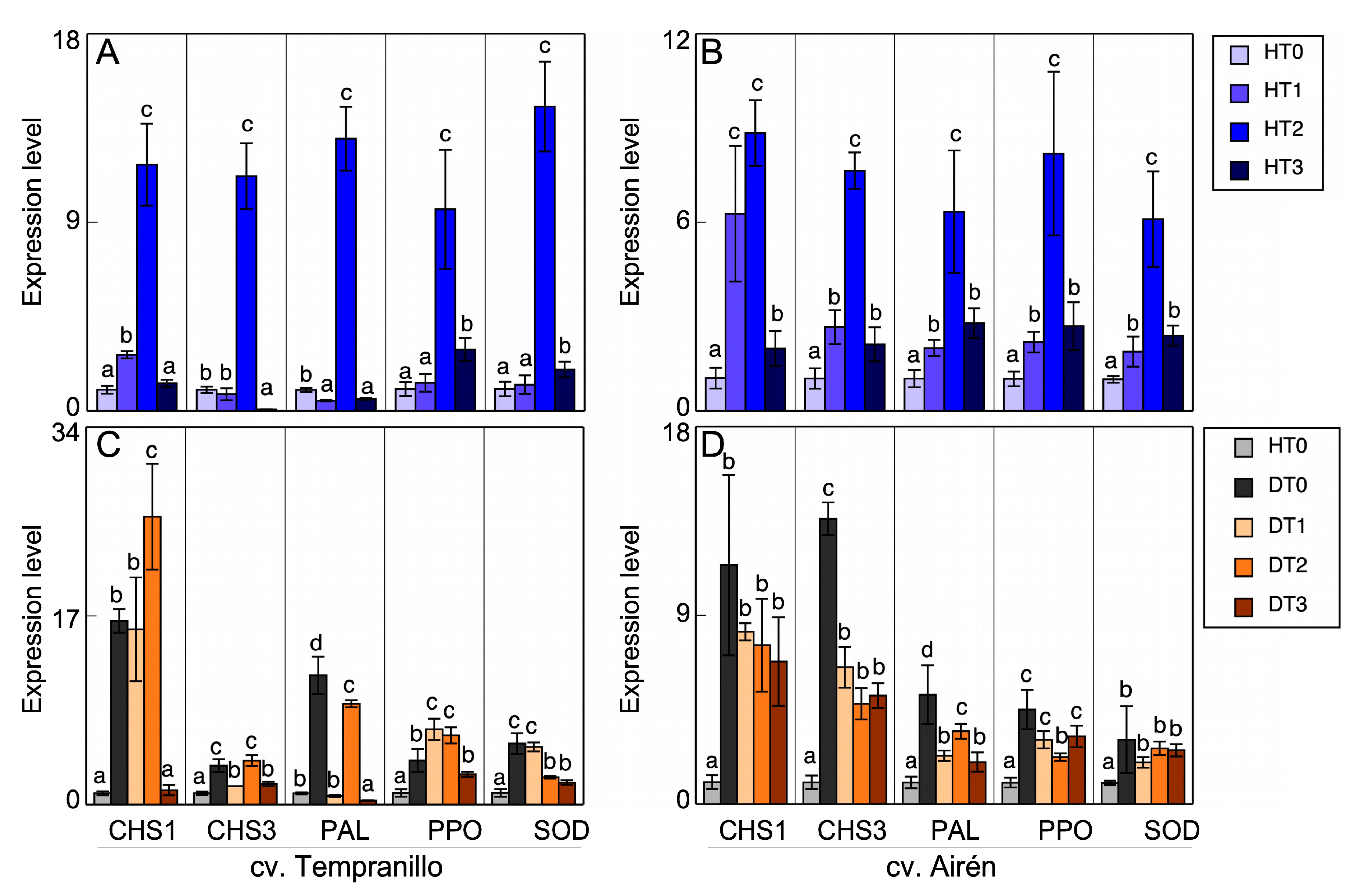
Figure 6.
(A,B) Transcript accumulation in grapes of the two grapevine varieties healthy (H), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). (C,D) Transcript accumulation in grapes of the two grapevine varieties healthy without treatment of manans (HT0) and diseased treated with the different doses of manans used. Quantitative PCR in CHS1, CHS3, PAL, PPO and SOD (the same letter do not differ significantly according to a parametric test of one-way ANOVA).
Figure 6.
(A,B) Transcript accumulation in grapes of the two grapevine varieties healthy (H), treated with different doses of mannans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3). (C,D) Transcript accumulation in grapes of the two grapevine varieties healthy without treatment of manans (HT0) and diseased treated with the different doses of manans used. Quantitative PCR in CHS1, CHS3, PAL, PPO and SOD (the same letter do not differ significantly according to a parametric test of one-way ANOVA).
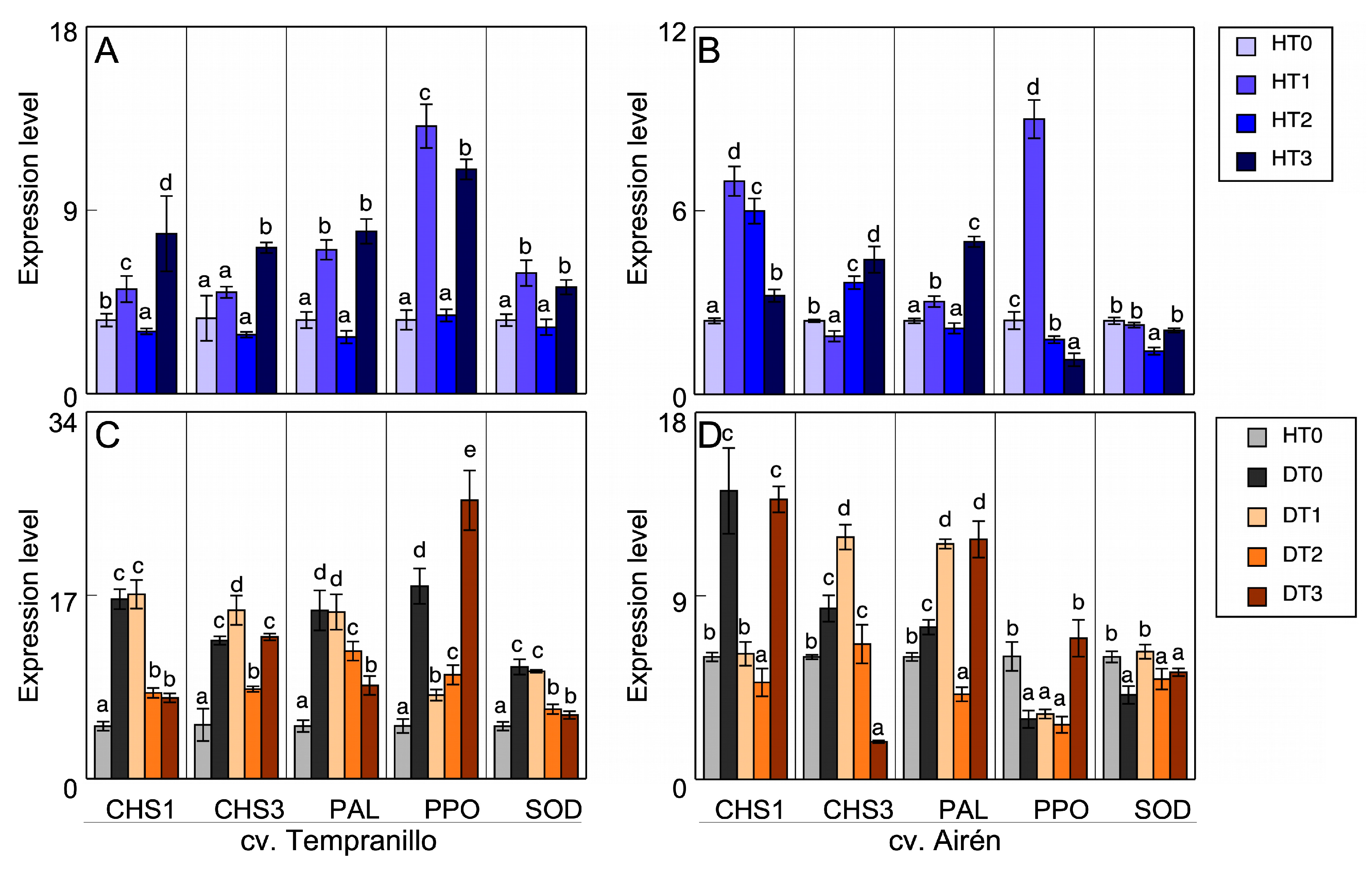

Table 1.
Primers used for amplification of grapevine cDNAs.
| Gene | F/R | Sequence 5´-3´ | Information | Size (bp) |
|---|---|---|---|---|
| SOD | F | CTGCGGGTTGGTGTTCTAAT | superoxide dismutase, chloroplastic/cytosolic VIT_02s0025g04830 | 156 |
| R | TTCCCATATGGTGGTTCCAT | |||
| PAL | F | ACAACAATGGACTGCCATCA | Phenylalanine ammonia lyase VIT_16s0039g01300 | 192 |
| R | GGAGGAGATTAAGCCCAAGG | |||
| PPO | F | GGCTTTTCTTCCCTTTCCAC | V. vinifera polyphenol oxidase, chloroplastic-like (LOC100261681), misc_RNA | 205 |
| R | ATTACAGTCGGAGGCAGGTG | |||
| Actine 1 | F | ACTGCTGAACGGGAAATTGT | V. vinifera actin 2 (act2) mRNA chensunanActin2-S1 AF369525 | 189 |
| R | AGTCCTCTTCCAGCCATCT | |||
| CHS1 | F | AGCCAGTGAAGCAGGTAGCC | chalcone synthase (AB015872) | 155 |
| R | GTGATCCGGAAGTAGTAAT | |||
| CHS3 | F | GTTTCGGACCAGGGCTCACT | chalcone synthase 3 (AB066274) | 93 |
| R | GGCAAGTAAAGTGGAAACAG | |||
| VATP16 | F | CTTCTCCTGTATGGGAGCTG | V-type proton ATPase chensunan16 kDa proteolipid subunit | 112 |
| R | CCATAACAACTGGTACAATCGAC |
Table 2.
Efect of treatment of manans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3) on the chlorophyll a and b, and total chlorophyll content, chlorophyll a/b ratio, total carotenoids and carotenoid/chlorophyll ratio in leaves of cv. Tempranillo and Airen grapevines, healthy (H) and diseased (D). Chlorophylls and carotenoids are expressed in µg g-1 FW. The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (the same letters of each grapevine cultivar or H and D indicate do not differ significantly).
Table 2.
Efect of treatment of manans (0 mg L-1: T0, 1 mg L-1: T1, 2 mg L-1: T2, and 3 mg L-1: T3) on the chlorophyll a and b, and total chlorophyll content, chlorophyll a/b ratio, total carotenoids and carotenoid/chlorophyll ratio in leaves of cv. Tempranillo and Airen grapevines, healthy (H) and diseased (D). Chlorophylls and carotenoids are expressed in µg g-1 FW. The data are means ±SD from, at least, 4 independent experiments, each carried out in triplicate (the same letters of each grapevine cultivar or H and D indicate do not differ significantly).
| cv | Treatment | Chl a | Chl b | Chl a+b | Chl a/b | Carotenoids | Car/Chl |
|---|---|---|---|---|---|---|---|
 |
HT0 | 1723.4±52.9c | 1138.6±87.1c | 2861.9±128.7c | 1.52±0.08a | 100.0±13.4ab | 0.030±0.008a |
| HT1 | 1560.5±67.0b | 909.1±98.7b | 2469.6±165.5b | 1.73±0.11b | 107.2±16.0ab | 0.044±0.009ab | |
| HT2 | 1448.4±19.3b | 784.4±31.2ab | 2232.8±49.7b | 1.85±0.04b | 123.0±2.8b | 0.055±0.001b | |
| HT3 | 1219.8±74.2a | 700.6±73.3a | 1920.4±144.3a | 1.75±0.09b | 95.2±7.4a | 0.050±0.007b | |
| DT0 | 996.9±68.6a | 439.7±20.6a | 1520.2±104.8a | 2.00±0.10bc | 116.2±4.9ab | 0.078±0.006b | |
| DT1 | 1134.7±120.5a | 589.2±68.5a | 1724.0±187.2a | 1.93±0.06b | 125.3±12.0ab | 0.074±0.007b | |
| DT2 | 973.2±211.4a | 485.1±118.9a | 1456.5±331.5a | 2.02±0.07c | 130.0±12.5b | 0.090±0.019b | |
| DT3 | 1020.4±168.1a | 627.6±120.6a | 1647.9±273.8a | 1.67±0.19a | 107.2±11.2a | 0.053±0.007a | |
 |
HT0 | 1330.4±70.1a | 668.8±26.3a | 1999.2±95.8a | 1.99±0.03b | 117.6±17.6a | 0.060±0.012a |
| HT1 | 1275.0±244.3a | 663.6±163.2a | 1938.6±405.9a | 1.94±0.11b | 118.6±8.8a | 0.064±0.014a | |
| HT2 | 1066.9±92.4a | 536.3±49.6a | 1603.2±127.1a | 2.00±0.17b | 114.4±19.5a | 0.071±0.010a | |
| HT3 | 1248.4±208.0a | 747.8±129.0a | 1996.2±331.8a | 1.67±0.10a | 84.5±21.7a | 0.044±0.011a | |
| DT0 | 868.8±0.8c | 405.6±12.4a | 1274.4±12.0b | 2.14±0.06a | 111.8±11.7 b | 0.089±0.011a | |
| DT1 | 680.8±85.8b | 303.0±38.3a | 983.8±123.5a | 2.18±0.14a | 115.8±20.6 b | 0.112±0.023a | |
| DT2 | 647.8±115. b | 314.6±70.7a | 962.4±183.8a | 2.08±0.13a | 83.2±12.6 a | 0.088±0.016a | |
| DT3 | 509.2±111.5a | 293.4±112.8a | 802.6±216.1a | 2.01±0.25a | 75.5±11.6 a | 0.109±0.042a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



