
Submitted:
19 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Plant growth and productivity are hindered by abiotic stresses. Crop productivity and the ecosystem are suffering due to climate change and harmful farming practices. We urgently need to use environment-friendly techniques like Arbuscular Mychorriza Fungi (AMF) to improve crop productivity. It is a well-established fact that AMF are effective bio-fertilizers. It is believed that AMF injections can increase plant resilience to stressors like heat,metals, salt, drought, and extreme temperatures. AMF aids in enhancing tolerance mechanisms and safeguarding essential metabolic pathways. AMF, as natural root symbionts, give plants important inorganic nutrients, helping them grow better, produce more, and acclimatize even under stress. AMF acts as a bio-fertilizer, boosting plants’ ability to adjust to different environments. Thus, researchers need to conduct further studies on how AMF promotes crop quality and productivity. This review describes the current understanding of AMF’s impact on host plants at their different growth stage, their benefits, applications, and the significance of plant nutrient relationships with AMF.
Keywords:
Arbuscular mycorrhizal fungi
; Plant-growth
; Abiotic-factors
; Stress-tolerance
; Mineral utilization
; Mychorriza application
1.0. Introduction
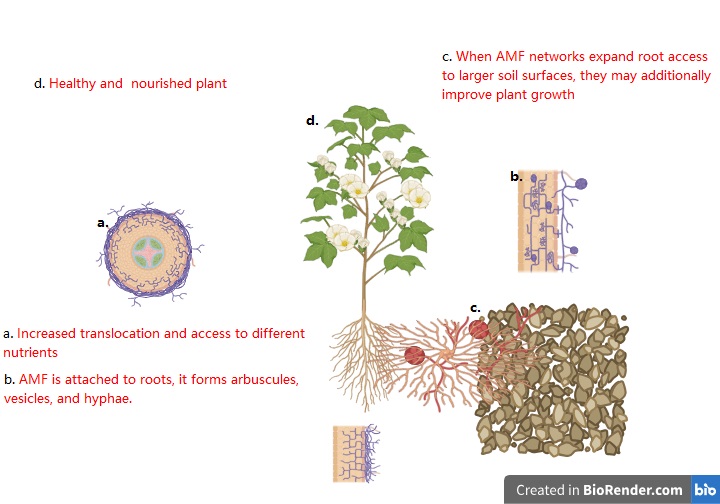
Friendly symbiotic communication between arbuscular mycorrhizal fungi (AMF) aids in growing plant crop under stressful conditions (Orine et al., 2022, Bhantana et al., 2023, Zou et al., 2023). AMF act as facilitators in the plant host, resulting into enhanced photosynthetic activities, plant development, and prolific gaseous exchange ability in plants (Birhane et al., 2012, Sun et al., 2013, Yang et al., 2021). It has been shown to promote adaptation to some stressful conditions which includes salinity-stress, drought-stress, heavy-metal-contamination, acidic and alkaline stress, temperature stress, and diseases are as a result of fungal-plant symbiotic relationships (Rodriguez et al., 2008; Sun et al., 2013, Ahanger et al., 2014; Salam et al., 2017, Zou et al., 2023). Symbiotic relationships with AMF can be formed between over 90% of most plant species, which includes ferns, legumes, tubers, bryophytes, and plants that are flowering (Zhu et al., 2010a; Ahanger et al., 2014). When AMF is attached to roots, it forms arbuscules, vesicles, and hyphae; when it is free-living in the the root system, it stays spores and hyphae. When AMF networks expand root access to larger soil surfaces, they may additionally improve plant growth (Bowles et al., 2016, Yang et al., 2021, Orine et al., 2022). Increased translocation and access to different nutrients (nourishment uptake occurs directly via transporters, which is located and occur within the root epidermis and root hair; nourishment are moved indirectly using fungal transporter systems that activate in the extra-radical mycelium (ERM) associated with the maturing fungus before being transferred to the intra-radical mycelium (IRM) in arbuscular mycorrhizal connections or the hartig net in EM interactions (Rouphael et al., 2015, Yang et al., 2021) (Figure 1) are additional mechanisms that AMF can improve nutritional activity in plants.
Plant health is eventually promoted by AMF's influence on the structural and textural status of the soil (Sun et al., 2013; Zou et al., 2016; Thirkell et al., 2017; Yang et al., 2021; Orine et al., 2022). AMF fungal formed hyphae contribute to the soil's increased nutrient content by hastening the break-down of organic materials (Paterson et al., 2016;Yang et al., 2021; Bhantana et al., 2023). By promoting flow from the tips of the roots to the main roots, mycorrhizal fungi can affect how CO2 is fixed in plants. These mushrooms offer amazing and practical advantages to all kinds of plants. It is a widely and well established fact that AMF-fungi improves nutrient uptakes, especially of phosphorus, and biomass accumulation of several crops in soils having very low phosphorus levels. AM-fungi support plants in thriving in harsh conditions. The majority of publications point out that mycorrhizal inoculation promotes plant development in soil contaminated with metals. Whether you plant flowers, veggies, trees, your plants will look noticeably better after adding them to the soil. Both indoor and outdoor plants benefit equally from the treatment, and its effects build up over time (Bucking et al.,2012; Zou et al.,2023).
This study review iterates on the application of AMF as biofertilizers for plant husbandry, their part and function as regulators of plant development and growth, and the processes by which they are able to improve plant growth under stressful conditions and mitigate nutrient’s intake.
2.0. History of Arbuscular Mycorrhizal Fungi
AMF is a natural soil resident bacteria that improves plant nutrient uptake and stress tolerance (Sun et al., 2013, Sun et al., 2018 ). Maturing species of AMF belong to subphylums Glocomycota, phylum Myxomycota (Spatafora et al., 2016). AMF is divided into four orders and 25 rubrics: Videlicetales, Glomerales, Paleosporeales, Paraglomerales, and Hypomorphiae (Redecker et al., 2013). Living as obligate bio-trophs they feed on stored plant products (photosynthetic) and lipids (Bago et al., 2000;Sun et al., 2013; Jiang et al., 2017).
AMF promotes plant growths by perfecting the infiltration of soil water, substrates and also available nutrients from the soils and protecting against fungal pathogens. These are important endo-symbionts because they help businesses grow and maintain healthy ecosystems.They role they play is highly important role in achieving sustainable crop improvements (Gianinazzi et al., 2010; Bhantana et al., 2023).
2.1. The process of Mycorrhizae Colonization in Plants
The development of these fungi on plants occurs through three main pathways: spores, colonized root fragments, or vegetative hyphae. Propagules is the term used for the latter two sources. Propagules are typically the main unit of measurement for mycorrhizae products on the label. The availability of mycorrhizza fungi near the plant’s roots is a prerequisite. Exudates are fluids formed from plant's root(s) as they extend directly into the surrounding soil. Upon sensing the presence of these fluids, the nearby fungi start advancing towards the roots.
Once they arrive, they feed on the exudates and establish a colony. The root thrive by feeding the fungi, while the fungi grows along with the plant’s root system. The mutual relationship continues and is strengthen as the mycorrhizae expands within the root system. Most Potting soil/compost mixes and other composed soil blends contain little or no mycorrhizae, but can be added before or during planting. Once the fungi is introduced there is quick colonization which can be hastened between AMF and plants by mixing it into surrounding soils, adding it during transplanting by wetting the seedlings or using it to coat seed before planting (Bucking et al. 2012).
2.2. The traits of AMF Synmbiosis
For 400 million years (Selosse et al., 2015; Bhantana et al., 2023), it is widely known for a fact that AMF-plants associations have a highly beneficial relationship. The establishment of such links occur as stepwise series of biological procedures, leading to highly beneficious effects within natural agricultural soil ecosystems (Sun et al., 2013;Van der Heijden et al., 2015). These symbiotic associations with AMF helps with regulating the growths and developments of plants within the ecosystem. Mycelial networks formed by fungi extend beneath the root structures of the plant and promoting nutrient uptakes which are sometimes unavailable. Common mycorrhizal network (CMN) can be formed by fungal mycelium which colonize the root systems of several plants, from different species. The term CMN can described as the fundamental components within a terrestrial eco-system having major effects on the various plant communities, even in invasive plant(s) (Pringle et al., 2009) with a well developed fungal-enhanced transportation of nitrogen (N) and phosphorus (P) plant(s) (Yang et al., 2021). In addition, rhizospheric nutrients are transferred from AMF fungi to the roots of plants, giving rise to other associated effects, allowing AMF to improve plants tolerance in response(s) to biotic and abiotic factor(s) (Plassard and Dell, 2010;Sun et al., 2013;Yang et al., 2021). AMF’s are also able to boost the character of soil encouraging enhanced plant development, improves their tolerance and elevate the morpho-physiological trait(s) of plant(s) under regular and also in stressful environmental conditions or surroundings (Alqarawi et al., 2014 a&b, Hashem et al., 2015). The community of the terrestrial flora depend soly on AMFs as natural growth regulators. Using AMF as bio-inoculant(s) is endorsed by researchers as a relevant bio-fertilizer for sustained crop productivity (Barrow, 2012; Navarro et al., 2014). Syamsiyah et al. (2018) showed that soils when treated using AMF produce a constant mass with much higher extra-radical hyphal mycelium when compared with non-AMF treated soil(s). AMF glomalin-related protein(s) (GRSP) found in the soil is believed to regulate water contents in soil(s) experiencing or subjected to various abiotic stress types (Wu et al., 2014; Navarro et al., 2014; Orine et al., 2022), they regulate water movements between the soil(s) and plant(s), and promote plant development(s). Glomalin with other closely associated compounds protects the soil from undergoing desiccation as it enhances water-holding-capacities in the soils (Sharma et al., 2017; Zou et al.,2023). Other growth-related function(s), leaf water potential, such as the stomatal conductances, relative water contents (RWC), CO2 assimilation and PSII efficiency are strongly impacted by AMF inoculations (He et al., 2017; Yang et al., 2021). When above-ground organs and tissues are altered, AMF can aid in improving tolerance to water stress (Bárzana et al., 2012; Chandrasekaran et al., 2019). With AMF inoculations, accumulations of dry-matter and water uptake(s) are enhanced, improving plant’s levels of tolerance to stress conditions like salinity and also drought. Organic farm practises for growth promotion(s) with yield maximization(s) could greatly profit from utilizing AMF to promote plant growths in various biological ecosystem(s) (Figure 2).
3.0. AMF AND PLANT DEVELOPMENT
3.1. AMF and Mineral Nutrient Utilization
Excessive and uncensored land-use has left deadly impact(s) on its total bio-diversity, affecting the normal functioning within the ecosystem (Smith and Read, 1997; Balliu et al., 2015; Wagg et al., 2015; Jiang et al., 2017). The transfers of nutrients (major and minor) is a primary role of the AMF-Plant symbiotic relationship (Nouri et al., 2015; Luginbuehl et al., 2017). AMF colonizations are mostly believed to be important in the process(es) of uptake of nutrients in plant(s). Inoculation of AMF has proven severally to boost the concentrations of macro and micro-nutrient(s) substantially, leads to increase(s) in the production(s) of photosynthate and increase(d) plant weight and biomass (Chen et al., 2017; Mitra et al., 2019). AMFs are able to increase uptake(s) of inorganic nutrient in most plant types, particularly phosphorus (Smith et al., 2003; Nell et al., 2010; Jiang et al., 2017). Plants are able to receive more nutrients from nutrient-depleted soils when AMFs are present (Kayama and Yamanaka, 2014). Plant-AMF associations enhances phyto-availability of micronutrient(s) including copper and zinc (Smith & Read, 1997), while the absorbing capacity of host roots is increased by AMF (Bisleski, 1973). AMF-tomato association resulted in increased leaf total areas, nitrogen, calcium, potassium, and also phosphorus content, indicating boosted plant’s growth rates (Balliu et al., 2015; Li et al., 2016). AMF’s symbiotic association with root(s) allow for increased availability of essential nutrient(s) to the host plant(s) and ultimately providing mineral nutrient(s) in return, like, N, Ca, K, Zn, P and S. They are able to give nutritional supports for plant(s) under poor condition(s) within the root cell(s). AMF produces fungal-like structures known as arbuscules, that assists in the exchange process for inorganic mineral(s) and compound(s) of phosphorus carbon e.t.c, ultimately with an impart of considerable vigour to host plant(s) (Chen et al., 2017; Prasad et al., 2017; Bhantana et al., 2023).
Moreover, they are able to boost the phosphorus contents in the root(s) and shoot(s) system(s) (Chen et al., 2017; Al-Hmoud and Al-Momany, 2017; Orine et al., 2022), even with limited phosphorus, the mycorrhizal association(s) enhances the supply of phosphorus to the colonized root(s) of host plant(s) (Bucher, 2007; Chen et al., 2017). The Pi uptake rate(s) in maize plants colonized with AMF showed a significant improvement (Garcés-Ruiz, 2017). Enhanced activities (especially photosynthetic) and several leaf function(s) have been shown to occur with improvements in growth rates of the AMF inoculations that is laterally connected to the uptake(s) of P, N, and also carbon, that migrate toward(s) root(s) and promoting the developing tuber(s). It has been shown that AMF maintains P and N uptake(s) mainly by helping with plant’s developments at high and low P level(s) using different irrigation methods (Liu et al., 2014; Chen et al., 2017; Liu et al., 2018). Pelargonium graveolens L. under limited water stress also showed increased nitrogen, phosphorus, and iron concentrations with mycorrhizal symbiosis, while Pistachio plants inoculated with AMF had higher levels of P, Zn, K and also Mn with drought related stress(es) (Bagheri et al., 2012). The Gomez-Bellot et al. (2015) study showed increased P, K and Ca levels in Euonymus japonica under salt related stress when AMF was present. In addition, AMF inoculation was able to improve P and N content(s) in the plant Chrysanthemum morifolium cell tissue(s) (Wang et al., 2018) with higher seedlings weight(s) as a result of improved water-contents and also P found intercellulaly, CO2, and also N content(s) in Leymus chinensis (Jixiang et al., 2017).
3.2. AMF use Bio-fertilizers
Bio-fertilizer(s) are a natural mixture used to enhance soil fertility they are beneficial for soil- health for growth and development of plants (Sadhana, 2014). Two decades, of research studies have iterated the numerous benefits that AMF gives for soil-health and also crop production. AMF may be the future key to replacing inorganic fertilizers effectively decreasing the amount of chemical fertilizers needed and released into the environment (Ortas, 2012). AMF are able to stimulate reduction of root biomass in plants while enhancing increased nutrient uptake using a larger surface area and extending deeply into the pores of the soil. The persistent use of chemicals in farm practises has given rise to several problems for soil, plants, and human health. AMF can help to reduce chemical fertilizer use by as much as 50% giving rise to better eco-friendly farm practises, this may depend on the different types of plant species and stress/environmental conditions.
4.0. STRESS DUE TO BOTH ABIOTIC AND BIOTIC FACTORS.
4.1. Drought
Stress from lack of water impedes plant life in several way(s); for example, restricted water to plant root(s) decreases the rate at which transpiration occurs inducing oxidative stress (Impa et al., 2012; Sun et al., 2013; Hasanuzzaman et al., 2013; Zou et al., 2023). Stress leaves a damaging effect(s) on drought affected plant(s) affecting the enzymatic activities, ion uptakes, and also nutrient assimilations (Ahanger and Agarwal, 2017; Jixiang et al., 2017; Ahanger et al., 2017a). Strong evidences have been shown of drought-stress alleviations with AMF applied in varied crops like wheat, soybean, barley, strawberries, maize-crop and also onion (Mena-Violante et al., 2006; Jixiang et al., 2017; Yooyongwech et al., 2016; Moradtalab et al., 2019; Zou et al., 2023 ). Plants ability to tolerate drought maybe as a result of the large volumes of soil exploration by associations of roots-extra-radical hyphae of the fungi (Orfanoudakis et al., 2010; Gianinazzi et al., 2010; Ruiz-Lozano et al., 2015; Gutjahr and Paszkowski, 2013; Zhang et al., 2016; Yang et al., 2021; Zou et al., 2023).
This kind of symbiosis and association(s) are believed and have shown abilities to control diverse plants and physio-biochemical processe(s) which include increased osmotic adjustments (Kubikova et al., 200; Orine et al., 2022; Zou et al., 2023), regulating the stomata through controlled ABA metabolism(s) (Duan et al., 1996), promoted accumulations of prolines (Ruiz-Sánchez et al., 2010; Gianinazzi et al., 2010; Sun et al., 2013; Yooyongwech et al., 2013), or enhanced glutathione levels (Rani, 2016). The mutalistic relationships of varied plants with AMF amilerates for improved efficiency and root sizes, biomass and leaf-area indexes under drastic drought (Al-Karaki et al., 2004; Ludwig-Müller, 2010; Gholamhoseini et al., 2013; Orine et al., 2022; Zou et al., 2023), showing that AMF interactions with plant(s) can enable them cope with adverse environmental conditions (Ruiz-Lozano, 2003). Moreso, Morte et al., 2000 and Mena-violante et al., 2006 report that AMF symbiosis result(s) in enhanced gaseous exchange, retention of water in leaf, transpiration rate and stomatal conductance. Recent research has indicated that ABA responses are helpful in liquid conduction through the stomata and similar physiologically related processes (Ludwig-Müller, 2010). In addition, Li et al. (2019) found that the anti-oxidant system is elevated in Hemarthria altissima and Leymus chinensis plant species, resulting in enhanced growth and photosynthesis.
4.2. Salinity
Saline-soils are a major environmental problem that pose serious threats to global food security. Salinity related-stress cause reduced plant growth via impacting vegetative development(s) and net rate(s) of assimilation, which result in lower yield and productivity (Hasanuzzaman et al., 2013; Talaat and Shawky, 2014; Ahanger et al., 2017a) and reactive oxygen species are also formed (Ahmad et al., 2010; Ahanger et al., 2017b). Several investigations are currently being done for the discovery of suitable ways to promote crop production in soils that are affected by salt to ameliorate the negative impacts, by using AMF with caution (Ahanger et al., 2018; Santander et al., 2019). According to research reports (Abdel Latef and Chaoxing, 2014; Talaat and Shawky, 2014; Ahanger et al., 2018), AMF is effectively improving the growths and yields of plants subjected to salinity stress. In the findings of El-Nashar; 2017 Antirrhinum majus plants have experienced improvements in leaf-water potential, growth-rate and water-use efficiency. Some physiological characteristics such as photosynthetic rates, conductance in the stomata, and leaf water retentions can benefit from AMF symbiosis with plants that are cultivated under saline conditions. Mycorrhizal had a significant effect in alleviating the adverse impacts of salinity conditions on photosynthesis (Sheng et al., 2011). AMF inoculation was effective in improving the photosynthesis rate, other gas exchanges, leaf chlorophyll contents, and water-use efficiencies of Ocimum basilicum L. when under high salinity. The growth traits of Allium sativum plants, such as the index of the leaf-area and plant biomass, have improved in salinity conditions in the past (Borde et al., 2010). Under moderate saline condition(s), mycorrhiza inoculations led to an impressive increase(s) in fresh and also dry weight(s), as well as an increase in nitrogen concentration in shoot and root, as determined by Wang et al. (2018).
4.3. High and low temperatures
AMF interactions greatly influences plant rhizospheric responses with changes such as increasing soil temperatures, leading to positive yield and production increase which are sustainable (Bunn et al., 2009; Sun et al., 2013; Yang et al., 2021). Heat related stress have significant effects on plant growths and also developments, their influences include i) losing plant’s vigor in addition to inhibiting time of seed-germinations, ii) retarding plant growth-rates, iii) decreasing biomass values, iv) leaf and reproductive organs wilts and burns , v) abscissing and senescencing of plant leaf, vi) spoilage and loss of fruit colour, vii) yield reduction and cell deaths (Wahid et al., 2007; Bunn et al., 2009; Hasanuzzaman et al., 2013) and viii) elevated oxidative stress(es). AMF-inoculated plants generally grow better when subjected to heat stress (Gavito et al., 2005;Sun et al., 2013;Yang et al., 2021). The study by Maya and Matsubara (2013) report the close association(s) of AMF (Glomus fasciculatum) in plant(s) leads to positive change(s) in growths and yields under the higher temperatures (Figure 3).
Photosynthesis is well documented as a highly sensitive to temperature stress and is usually first affected and then other related symptoms of stress become visible (Berry and Björkman 1980). When photosynthesis is limited, plant(s) growth is reduced under lower or higher temperature(s) (Wahid et al. 2007), with AMF, stimulation of photosynthetic performances that are different from tthe non-AM plants that are visibly expressed (Augé 2001; Zhu et al., 2011).Temperature related stress in AMF inoculated plants show higher Pn than in non-AMF plant crops (Ruotsalainen and Kytöviita 2004; Wahid et al. 2007; Zhu et al. 2010a, 2015), having more capacity to assimilate CO2. However, Wu and Zou (2010) report no impact with AM inoculation on Pn in Citrus (tangerine) seedlings when grown at low temperatures. Stress leads to decreased chlorophyll concentrations, because chlorophyll biosynthesis is adversely affected or increased chlorophyll degradation rates, as reported by many authors. The work of Paradis et al. (1995), report higher chlorophyll concentrations in AMF inoculated plants than plants without AMF at regular temperatures, showing that temperature related stress rarely interferes or tampers with light-harvesting and chlorophyll-synthesis in AMF and alleviating damages to mesophylls chloroplasts, improving photosynthetic-efficiency than in non-AMF plants (Evelin et al., 2009). On the contrary, the study of Charest et al. (1993) report that AMF-maize plant(s) produced lower chlorophyll concentrations, but more carotenoid content. AMF take crucial roles to protect the photosynthetic apparatus acting as aids to light harvesting pigments (Young et.). Zhu et. al. (2011) also observed AMF- maize plant(s) had more carotenoid concentrations than the plants having no AMF showing that AMF colonization helps with stabilizing the lipid related phase(s) within the thylakoid membrane(s), providing photo-protections for cell structure(s) and also photosynthetic related apparatus(es) (Karim et al. 1999; Wahid et al. 2007; Orine et al., 2022; Zou et al., 2023).
AMF increases plant ability for tolerance under cold temperatures (Birhane et al., 2012; Chen et al., 2013; Liu et al., 2013; Yang et al., 2021), giving rise to better growths than in non-AMF experimental plants ( Abdel Latef and Chaoxing, 2011b;Sun et al., 2013; Chen et al., 2013; Liu et al., 2013; Yang et al., 2021). AMF supported plants are able to easily combat cold stress, resulting in eventually improved plant developments (Gamalero et al., 2009; Birhane et al., 2012; Zou et al., 2023). In addition, AMF helps with the retention of moisture inside its host-plant (Zhu et al., 2010a), increasing secondary produced metabolites that strengthens the plant’s natural immunity system, enhancing water-conservation capacity and water-use efficiency (Zhu et al., 2010b) and increases protein contents aiding the plants to combat cold environments (Abdel Latef and Chaoxing, 2011b) (Figure 3). AMF Symbiotic relationships forges good water-plant relationship(s) and osmotic adjustments (Zhu et al., 2012; Zou et al., 2023). AMF promotes the process of synthesizing chlorophyll with significant improvements in metabolite(s) concentration(s) in plant(s) growing under cold stress condition(s) (Zhu et al., 2010a; Sun et al., 2013; Yang et al., 2021). AMF also helped in altering protein contents in tomato(es) and other vegetable(s) subjected cold-stresed environments (Abdel Latef and Chaoxing, 2011b).
At increasing higher temperatures (HT) deleterious effects on plant growth were noticed but the protective and helpful role(s) of Arbuscular mycorrhizal fungi (AMF) when plant(s) were cultivated under stress conditions, were particularly able to enable the experimental plant(s) to survive (Mohammadi et al., 2011). Under high-temperature pepper inoculated with Glomus isolate and G. intraradices mixtures had enhanced growths relative to non-AM controls, in-spite of the reduction in amounts of AM colonization showing lesser transferred fungal P to the plants (Martin et al., 2004).
4.4. Heavy Metals
AMF are commonly believed to assist plant to establish themselves in soils with heavy-metals contaminants, as the have the potentials to strengthen the defence systems of the AMF colonized plants promoting growths and also development(s). Accumulations of heavy-metal(s) in staple food crops, fruits, veggies, and soils lead to several health hazard(s) (Liu et al., 2013; Yousaf et al., 2016). Aguilera et al. (2014) revealed that AMF association(s) with wheat significantly increases nutrient uptakes when subjected to aluminum stress. While shoot-root growth(s), leaf and cell death in crops planted on soil(s) enriched with Zn and Cd were highly suppressed (Moghadam, 2016) as also seen in repeated studies which have explored the effects of AMFs on metal-accumulation in plant(s) (Souza et al., 2012). The heavy metal(s) deactivated in the fungal hyphae (both external and internal types) (Ouziad et al., 2005), which are able to collect and fix such heavy metals within their cell walls to be stored within vacuoles or chelated along with other substance(s) that immobilizes them inside the cell cytoplasmic walls (Punamiya et al., 2010) significantly reducing the plants metal-toxicity. AMF are highly effective on plant developments and growths especially in stress-induced conditions and aids in increasing the physiological and morphological processes which enhance plant(s) biomass (root and shoot) and uptakes of immobilized nutrient(s) including Zn, Cu and P ultimately reducing the metal toxicity in plant(s) under these harsh condition(s) (Kanwal et al., 2015; Miransari, 2017). Additionally, metal dilution in plant tissues may result from increase(d) growths or chelations within the rhizosphere (Kapoor et al., 2013; Sun et al., 2013; Audet, 2014; Bhantana et al., 2023). According to the study by Andrade and Silveira (2008) and Garg and Chandel (2012), AMF is said to bind Zn and Cd within the cell wall(s) of mantle hyphae and of cortical cells, limiting uptakes and improving yield, growth, and nutritional status.
Numerous metals that plants acquire from within the rhizosphere are carried through the root zone(s) to aerial portions of the plant can be interrupted by mycorrhizae (Dong et al., 2008; Kapoor et al., 2013; Li et al., 2015). For mycelial of various AMFs, there is a high metal absorption and cation-exchange capacities (Takács and Vörös, 2003). As demonstrated in Lolium perenne found in severely contaminated soils with varying contaminants such as Ni, Cd, and Zn, metals, the presence of non-adapted AMF fixes the pollution within soil(s) and reduces uptakse and accumulations of heavy metals (Takács and Vörös, 2003). It is believed that AMF regulates the absorption and build-up of a few essential inorganic nutrients.
There have been reports of elevated uptake of Si in plants with mycorrhiza, such as Glycine max and Zea mays (Yost and Fox, 1982 and Clark and Zeto, 2000). According to Hammer et al. (2011), Si was substantially absorbed by Rhizophagus irregularis propagated spores and formed hyphae and then transported towards the host root. It is highly important to iterate that AMF may also be used to address inadequate Cd toxicity and mobility by increasing the pH of the soil (Shen et al., 2006), supplementing Cd inside extra-radical mycelium walls (Janouškova and Pavlíková, 2010), attaching Cd to the glycoprotein glomalin. AMF was very effective in lowering rice's Cd levels, particularly in the cell walls and vacuole(s), which resulted in Cd detoxifications (Li et al., 2016a).
According to the report of Wang et al. (2012), the altered chemical form(s) of Cd in many plant tissues could have influenced alfalfa's (Medicago sativa L.) increased Cd tolerance through AMF-mediated means. The AMF plays a role for a number of processes, including the immobilization and limitation of metal compound(s), the precipitations of polyphosphate granule(s) in soil, the adsorptions to the chitin of cell walls of fungi, and the chelations of heavy metal(s) inside the fungus (Figure 2).
4.5. AMF and Combined Abiotic Stresses
It's commonly acknowledged that AMF can assist to reduce a number of stress(es) or combination(s) of stresses, including as heavy metals, temperature, salinity, nutrients, and drought. For instance, plants exposed to both salinity and drought produce more reactive oxygen species, which are sometimes extremely harmful to plant(s) (Bauddh and Singh, 2012). Enzymes such as catalases (CAT), peroxidases (POD), glutathione reductases (GR) and superoxide dismutases (SOD) are extensively used in the process detoxification of the reactive oxygen species (ROS) (Ahanger & Agarwal, 2017). Moreover, tomato(es) plants with which Scolecobasidium constrictum was inoculated with exhibited enhanced biomass output, stomatal conductance, leaf water relations and Fv/Fm in comparison with the un-inoculated plants when drought and salinity were applied simultaneously (Duc et al., 2018). Therefore, according to several studies (Abdel Latef, 2011; Abdel Latef and Chaoxing, 2011b; Abdel Latef and Chaoxing, 2014), AMF are essential for enhancing plant development and yield outputs under stress. There are extremely few research reports in the literature that show how AMF helps to mitigate the combined impacts of two or more stresses. Through a number of mechanisms, including enhanced photosynthetic rate, accumulation of osmoprotectants, uptakes and accumulations of mineral nutrients, up-regulation of antioxidant enzymatic activities, and changes to the rhizospheric ecosystems, AMF’s symbiosis shields plants against wide ranges of abiotic stress(es) (Bárzana et al., 2015; Calvo-Polanco et al., 2016; Yin et al., 2016). Several research (Augé et al., 2014; Lehmann et al., 2014; Lehmann and Rillig, 2015) have demonstrated increased nutritional qualities of AMF plant(s) when subjected to osmotic stressed-environments as a result of salinity or deficit watering. It's possible that tolerance mechanisms are similar because of coupled stress adaptations mediated by AMF. It is suggested that similar mechanisms generated by varied stresses may include AMF-mediated change(s) in the phytohormone profiles, mineral absorptions and assimilations, build-ups of suitable secondary metabolite(s) and osmolytes, and up-regulations of antioxidant(s) system(s). Certain mechanisms, such as the formation of phytochelatins, the sequestration and compartmentation of harmful ions, and protein expression, can be unique and vary significantly depending on the type of stress and AMF species involved. Osmotic stress tolerance can be significantly enhanced by altering root characteristics such as hydraulic conductivities (Evelin et al., 2009).the research of Zhang et al. (2018) found that via modifying gas-exchange characteristics and the concentrations of several important metabolites, the AMF insulated castor beans against saline stress. The aforementioned qualities of AMF have potentials for improving the nutraceutical quality of crops and may have substantial agronomic implications for the management and the productions of many prospective crops. To fully understand how AMF works to offset the impacts of combined, more research is necessary.
5.0. AMF AND PLANT GROWTH
5.1. Germination
Plants are often stimulated by mycorrhizal fungi to decrease biomass of roots while also increasing nutrient(s) uptake capacities by reaching far beyond the root’s surfaces and multiplying in soil pores enhancing germination. In a study using two sampling periods, there was high significant differences in seed germinations rate per treatment, but not in the final sampling period. Inoculated plants that initiated flowers and pods had a higher number of inoculated plants than controls. Mycorrhizae's positive effects on plant growth can be attributed to decreased germination periods and boosted plant developments, as shown by the findings of this study. This earlier development is not known to have a physiological explanation. Plant and mycorrhizae exudates are known to have complex chemical interactions that are able to impact on the numerous aspects of plant developments. Plant strigolactones serve as signal compounds for the initial stages of Arbuscular mycorrhizae colonizations, stimulating fungal metabolism and branching, while also promoting seed germination. According to Fusconi (2014), AM is associated with an increase in the production of auxins and cytokines, which stimulate plant growth. It's unclear if these chemical interaction(s) are responsible for the earlier germinations and reproductive development(s) of rapid Growth, which necessitates more experimental scrutiny. This study suggests that adding mycorrhizae can speed up the colonizations and propagations of desired plants in a natural environments. Mycorrhizae can significantly impact habitat restoration efforts due to their ability to accelerate plant development and germination. Agriculture could be positively impacted by having shorter crop cycles. More studies may explore the effects of mycorrhizae on colonizations and propagations of desired vegetations in non-greenhouse experiments Gutowski, Valeria (2015) "The resulting effects of mycorrhizae on seed developments, germinations, and reproductive yields with rapid growth,"(Binti et al., 2023). The interactive resulting effects of light, mycorrhizal status and exogenous carbon, on germinations and growths were investigated in-vitro with use of axenic agar microcosms for three geophytic orchid species and one tropical epiphyte. The geophytic species strongly depend on their associated mycorrhiza for growth and cannot be replaced by exogenous sucrose, while on the other hand the epiphytic species can. Exogenous sucrose in the dark resulted in the epiphytic species achieving 95% of more mycorrhiza seedling volumes. Germination was made possible by the strong interaction between mycorrhiza status and light exposure. In the absence of mycorrhiza, light can hinder or severely reduce the growth of terrestrial orchids (Sameera, 2019). Mycorrhiza's role in the increase in phytohormones and enzyme activities supporting seed germinations and growths of orchids.
5.2. AMF and Plant Yield
The quality of crops is enhanced by beneficial rhizosphere microorganisms, which improve the nutrient status as described above. AMF-colonized strawberry showed an increase in secondary metabolites, which led to improved antioxidant properties (Castellanos-Morales et al., 2010). AMF's ability to affect and produce carotenoids and volatile compounds can improve the dietary quality of crops (Hart et al., 2015). The quality of the tomatoes was observed to be beneficial by Bona et al. (2017) due to the beneficial effects of AMF. Zeng et al. (2014) found that Glomus versiforme caused a rise in sugars, vitamin C, organic acids, flavonoids, and minerals, which resulted in improved citrus fruit quality. Chlorophyll, anthocyanins, carotenoids, total soluble phenolics, tocopherols and various mineral nutrient(s) are increased in mycorrhizal symbiosis, as demonstrated by Baslam et al. (2011). AMFs have been used for large-scaled farm cultivations of potatoes (Hijri, 2016), maize-crop (Sabia et al., 2015), and yam-crop (Lu et al., 2015) demonstrating their significant potentials to increased crop yield. By enhancing the biosynthesis of important phytochemical(s) in edible plant(s), AMF can makes those plants more suitable for the chain that produces healthy food (Sbrana et al., 2014; Rouphael et al., 2015). It has generally been suggested that maintaining the pH of the soil could help AMF mitigate abiotic stress and preserve its horticultural value (Rouphael et al., 2015). Furthermore, AMF can be extremely important in enhancing plants' ability to withstand harsh conditions. Mycorrhizae on boosting production and growth in nine horticultural plants at two distinct P fertilization rates in a field setting (Ronsheim, 2012). According to Ortas (2010), the findings of the field experiment demonstrated that mycorrhiza inoculation greatly improved fruit yield and cucumber seedling survival. The studies of Gao et al. (2020) found a statistically significance of 28.54% increase in yields in cotton between inoculated and non-inoculated control plots.
5.3. Fight pathogens
Numerous plants associate with fungus to produce what are known as mycorrhizae, which allows the plants to obtain soil nutrients and shields them from toxins and disease. In addition, mycorrhizae aid in shielding plants from soil-borne diseases such as mold and pests. AMF can enhance the manufacturing of beneficious phytochemical(s) in edible plant(s), to make them more suitable and adapted for the chains of production of healthful food (Sbrana et al., 2014; Rouphael et al., 2015; Orine et al., 2022; Bhantana et al., 2023). According to reports, AMF may mitigate abiotic stress by maintaining the pH within the soil, preserving its horticultural values (Rouphael et al., 2015). Plants that form mycorrhizae—associations with fungi retrieve nutrients from the surrounding soils and are shielded from toxins and disease. They help plants avoid being affected by bugs, mold, and harmful pathogens living in the soil.
5.4. Photosynthesis
It is known that (AM) boosts nitrogen fixation and photosynthesis in plants. Cotton with AMF symbiosis demonstrated in increased growth, photosynthesis, number of balls for each plant, and cotton ripeness (Gao et al., 2020). Given that chlorophyll molecules efficiently absorb nitrogen, the amount of chlorophyll in plants colonized by AMF rises (De Andrade et al., 2015),the mycorrhiza arcs has become well known as agents accelerates photosynthesis and nitrogen fixation in plants. Nonetheless, in cold-stressed cucumber seedlings, AM has a negligible effect on leaf chlorophyll (Bulgarelli et al., 2020). AM exerts zero impact on gas exchange or PSII fluorescence in guar (Cyamopsis tetragonoloba (L.). In order to produce gum, photosynthesis was boosted in legumes, namely in Bambara groundnut (El-Sawah et al., 2021). This demonstrates that AMF can contribute significantly to improved photosynthesis.
5.5. Water use efficiency
Arbuscular mycorrhizal (AM) fungus modify the water interactions of plants during droughts, increasing the plants' tolerance to the dry spells. Using a randomized greenhouse experiment, Birhane et al. (2012) studied the impactive effects of the symbiotic relationship of plants with AM and precipitation pattern on the growth, transpiration, nutritional status, and mycorrhizal responsiveness in seedlings of Boswellia plants. Plants that mycorrhizal Mycorrhizal plants possess decreased rates of transpiration than non-mycorrhizal plants, which optimizes water usage efficiency.
5.6. Soil Enzymatic activity
A meta-analysis of multiple research revealed that distinct AMF types impact soil enzyme activity, even though the precise magnitude of these consequences has not been reported yet. Neutral pH and lower accessible phosphorus are necessary for AMF's optimum influence on soil enzymatic activity. An increased soil enzymatic activities may result from AMF's encouragement of plant growth. The employment of AMF inoculation in agriculture has advantages. Arbuscular mycorrhizal fungi possess a positive effect on the soil enzyme activities and boost plant biomass, in accordance to findings published by Mingsen et al. (2019). The percentage of root colonization or plant growth proved to be associated with endoxyloglucanase activity in mycelia connected to root development in plants.
As reported by Arriagada et al. (2012), introducing mycorrhiza to G. claroideum + T. versicolor resulted in a substantial rise in the rate of activity of the dehydrogenase enzyme, but introducing plants with G. claroideum alone showed a significant boost in β-glucosidase activity in blueberries. Cellulase and xylanase activity were considerably higher in grassland orchid fungus. While the fungi from orchids with epiphytic growth favor starch and arginine, the fungi from terrestrial orchids utilized carboxymethyl cellulose with greater efficiency. The extracellular enzyme output of fungi from grassland and epiphytic orchid species differ, according to the current research, indicating that they have become accustomed to various growing environments. In order to mitigate the harmful effects caused by temperature-stress causes to maize plants, the AM fungus could decrease the amount of lipid in membranes. Enzymes like superoxide catalase, dismutase and peroxidase appeared more active in the roots and leaves of plants after colonizations with AMF.
The investigations by Chen et al. (2013), Liu et al. (2013), and Birhane et al. (2012) suggests that AMF is capable of improving plant immunity to cold stress. The vast majority of research indicates that crops treated with AMF at colder temperatures grow better than plants treated without AMF (Zhu et al., 2010b). AMF helps plants fight off cold stress, a process that in turn fosters optimized plant developments (Gamalero et al., 2009; Birhane et al., 2012; Sun et al., 2013, Yang et al., 2021). It also helps the plants that are being treated retain water (Zhu et al., 2010a), boosts secondary metabolism in plants which boosts their immune system, and raises protein content in plants to help them fight off the cold (Chaoxing and Abdul Latef, 2011b). Improvements in the interactions between plants and water are credited to the presence of AMF associations in plants under stress (Zhu et al., 2010a; Abdel Latef and Chaoxing, 2011b). Additionally, it has documented how AMF affects tomato and other vegetable protein levels when exposed to cold stress conditions (Abdel Latef and Chaoxing, 2011b). High temperatures on the other hand exerted a detrimental effects on plant growths (HT). Previous studies has proven the highly protective role of Arbicular mycorrhizal fungi (AMF) in stressful environments, ( Mohammadi et al., 2011).
Furthermore, AMF treated plants exhibit greater productions of salicylic acid, jasmonic acid, as well as a number of additional significant inorganic nutrients. For example, under salt-stress conditions, the AMF-treated Cucumis sativus plants had greater concentrations of total P, Ca2+, N, Mg2+, and K+ than the uninoculated plants (Hashem et al., 2018). In saline conditions, mycorrhizal-inoculated capsicum annuum exhibited greater levels of chlorophyll in leaf , as well as greater Mg2+ and N absorption, but poorer Na+ transportions (Cekic et al., 2012). Furthermore, using lettuce, Santander et al. (2019)in ther work, showed that the mycorrhizal plants exhibited more biomass, improved N uptake, increased proline synthesis and obvious variations in ionic relations, notably less retention of Na+, than those in than those in non-mycorrhizal plants under stressed conditions. Crucial growth regulators can have their levels efficiently regulated by AMF inoculation. In accordance with studies by Talaat and Shawky (2014) and Hameed et al. (2014), AMF increases cytokinin quantity, which causes a notable photosynthetic transport during salinity stress. Growth promotion during salinity stress is triggered by an alteration in the polyamine pool, which is mediated by AMF (Kapoor et al., 2013). Increased strigolactone in AMF-treated plants substantially decreased the consequences of different saline effects on lettuce plants (Aroca et al., 2013). Crop colonization with AMF may minimize oxidative stress by suppressed lipid membrane peroxidations under salinity-stressed conditions (Abdel Latef and Chaoxing, 2014; Talaat and Shawky, 2014). Additionally, it was discovered that AMF inoculation accelerated the accumulations of multiple organic acids, which resulted to increases the process of osmoregulation in plants nurtured in salinity-laden conditions. Sheng et al. (2011), for instance, observed that maize plants planted in salty soil had higher synthesis/accumulations of certain particular organic acids, and that AMF promoted increased productions of betaine, validating what indirect roles are played by AMF in vegetative osmoregulations under salinity stressed environments.
The findings suggest that the AM fungus can mitigate the detrimental impacts of temperature stress on maize plants by limiting membrane permeability and lipid peroxidation while augmenting the build-up of osmotic equilibrium chemicals and antioxidant enzyme activity. AMF invasion in plants additionally, affects the activities of catalases, peroxidases and superoxide dismutase in plant leaves and roots have been boosted by the biofertilizer. By raising the production of the enzymes such as phosphatase, protease, dehydrogenase and invertase, treatments have enhanced soil microbial activity (El-Sawah et al., 2021).
6.0. Future prospects and conclusions
Several investigations have previously established the beneficial impact of AMF in enhancing plant growth in stressed situations. In order to comprehend the symbiotic relationships that formed and exists between AMF and a range of plants in stressful conditions, the updates of information addressing the beneficial roles of AMF has been presented in a logical manner in this review. It recently came to light that plants to which AMF was inoculated can effectively and efficiently combat a wide range of environmental stressed situations, like drought, saltiness, cold-stress, nutrient-stress and severe temperatures. AMF have primarily been discussed as beneficial organisms for the absorption of nutrient(s) from soil which contributes to enhancing the per hectare yields of a several numerous crops (such as cereals, legumes, tubers), horticultural plants and also vegetables. Promoting the use of AMF is essential to the sustainability of contemporary global agricultural systems. It is indisputable that applying AMF to improve agriculture may assist in fostering bio-healthy agriculture through lowering the volume of synthetic fertilizers as well as other chemicals utilized. AMF-mediated crop cultivation and yield enhancement can help agricultural plants meet the world's growing population's consumption needs. Environmentally friendly technologies will become increasingly popular and highly encouraged. Subsequent investigations need to be focused on pinpointing genetic components and genetic products that drive the regulation of AMF mediated developments and growth in high-stress environments. Identification of the key physiological and also the metabolic pathways when under various environmental conditions, as well as specific plant hosts and AMF specific protein factors controlling symbiotic interaction, can be can be highly interesting for future researches. Understanding the modifications that AMF causes in the tolerance methods, mechanisms and the communication between them that is set off to control plant performance can help to increase crop productivities. AMF activities needs to be researched at all levels in order to investigate its function in natural environments as bio-fertilizer(s) for sustainable agricultural output.
Proteomics strategies maybe significance in expanding the comprehension of the molecular foundations of AMF plant resistance to temperature-stressed conditions and for managing it. Across nature, AMF fungus can be found; they aren't restricted to their host plants. Nevertheless, temperature-stressed circumstances determine how successful AM symbiosis is. In order to inoculate plants that are acclimated to temperature-stress, it is highly necessary that future studies evaluate isolates of native and potential temperature-stress-tolerant AMF species. A significant consideration is the current situation of the global climate. The change scenarios anticipate that temperatures will climb and that extreme temperatures may occur more frequently, heightening the likelihood that crops will be exposed to unfavorable temperatures. Multiple investigations have shown that this strategy works better for agriculture in the future than trying to adjust to unfavorable environmental conditions. There are still difficulties with AM application in agriculture, such as the need to cultivate AM fungi on a big scale and intense competition from other micro-organisms in the soil environments.
Author Contributions
All enlisted authors have contributed to this review study by editing, providing information and graphical presentation for this research.
Funding
There is no funding report for this study.
Conflict of Interest Statement
All enlisted authors declare that there are no potential conflict of interest.
References
- Abdel Latef, A. A., Chaoxing, H. (2011a). Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hort. 127, 228–233. [CrossRef]
- Abdel Latef, A. A., Chaoxing, H. (2011b). Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress. Acta Physiol. Plant. 33, 1217–1225. [CrossRef]
- Abdel Latef, A. A., Chaoxing, H. J. (2014)a. Does the inoculation with Glomus mosseae improve salt tolerance in pepper plants? Plant Grow. Regul. 33, 644–653. [CrossRef]
- Aguilera, P., Pablo, C., Fernando, B., Fritz, O. (2014). Diversity of arbuscular mycorrhizal fungi associated with Triticum aestivum L. plants growing in an andosol with high aluminum level. Agri. Eco. Environ. 186, 178–184. [CrossRef]
- Ahanger, M. A., Alyemeni, M. N., Wijaya, L., Alamri, S. A., Alam, P., Ashraf, M., et al. (2018). Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate–glutathione cycle and glyoxalase system. PLoS One 13 (9), e0202–e0175. [CrossRef]
- Ahanger, M. A., Agarwal, R. M. (2017). Potassium up-regulates antioxidant metabolism and alleviates growth inhibition under water and osmotic stress in wheat (Triticum aestivum L.). Protoplasma 254 (4), 1471–1486. [CrossRef]
- Ahanger, M. A., Tittal, M., Mir, R. A., Agarwal, R. M. (2017a). Alleviation of water and osmotic stress-induced changes in nitrogen metabolizing enzymes in Triticum aestivum L. cultivars by potassium. Protoplasma 254 (5), 1953–1963. [CrossRef]
- Ahanger, M. A., Tomar, N. S., Tittal, M., Argal, S., Agarwal, R. M. (2017b). Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Biol. Plants. 23 (4), 731–744. [CrossRef]
- Ahanger, M. A., Tyagi, S. R., Wani, M. R., Ahmad, P. (2014). “Drought tolerance: role of organic osmolytes, growth regulators, and mineral nutrients,” in Physiological mechanisms and adaptation strategies in plants under changing environment, vol. 1 . Eds. Ahmad, P., Wani, MR (New York, NY: Springer), 25–55. [CrossRef]
- Ait-El-Mokhtar, M., Laouane, R. B., Anli, M., Boutasknit, A., Wahbi, S., Meddich, A. (2019). Use of mycorrhizal fungi in improving tolerance of the date palm (Phoenix dactylifera L.) seedlings to salt stress. Sci. Hori. 253, 429–438. [CrossRef]
- Al-Hmoud, G., Al-Momany, A. (2017). Effect of four mycorrhizal products on squash plant growth and its effect on physiological plant elements. Adv. Crop. Sci. Tech. 5, 260. [CrossRef]
- Ali, N., Masood, S., Mukhtar, T., Kamran, M. A., Rafique, M., Munis, M. F. H., et al. (2015). Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake of Linum usitatissimum in association with Glomus intraradices. Environ. Monitor. Assess. 187 (6), 311. [CrossRef]
- Al-Karaki, G., Mcmichael, B., Zak, J. (2004). Field response of wheat to arbuscular mycorrhizal fungi and drought stress. Mychorrhiza 14, 263–269. [CrossRef]
- Alqarawi, A. A., Abd-Allah, E. F., Hashem, A. (2014a). Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J. Plant. Interact. 9 (1), 802–810. [CrossRef]
- Amiri, R., Ali, N., Nematollah, E., Mohammad, R. S. (2017). Nutritional status, essential oil changes and water-use efficiency of rose geranium in response to arbuscular mycorrhizal fungi and water deficiency stress. Symbiosis 73, 15–25. [CrossRef]
- Amiri, R., Nikbakht, A., Etemadi, N. (2015). Alleviation of drought stress on rose geranium Pelargonium graveolen L Herit. in terms of antioxidant activity and secondary metabolites by mycorrhizal inoculation. Sci. Hort. 197, 373–380. [CrossRef]
- Arriagada C., Manquel D., Cornejo P., Soto J. , Sampedro I., Ocampo J. (2012). Effects of the co-inoculation with saprobe and mycorrhizal fungi on Vaccinium corymbosum growth and some soil enzymatic activities. Journal of Soil Science and Plant Nutrition. 12: 287-298. http://dx.doi.org/10.4067/S0718-95162012000200008. [CrossRef]
- Audet, P. (2014). “Arbuscular mycorrhizal fungi and metal phytoremediation: ecophysiological complementarity in relation to environmental stress,” in Emerging technologies and management of crop stress tolerance. Eds. Ahmad, P., Rasool, S. (San Diego: Academic Press), 133–160. [CrossRef]
- Augé, R. M., Toler, H. D., Saxton, A. M. (2014). Arbuscular mycorrhizal symbiosis and osmotic adjustment in response to NaCl stress: a meta-analysis. Front. Plant. Sci. 5, 562. [CrossRef]
- Bagheri, V., Shamshiri, M. H., Shirani, H., Roosta, H. (2012). Nutrient uptake and distribution in mycorrhizal pistachio seedlings under drought stress. J. Agric. Sci. Technol. 14, 1591–1604. [CrossRef]
- Balliu, A., Sallaku, G., Rewald, B. (2015). AMF Inoculation enhances growth and improves the nutrient uptake rates of transplanted, salt-stressed tomato seedlings. Sustainability 7, 15967–15981. [CrossRef]
- Barrow, C. J. (2012). Biochar potential for countering land degradation and for improving agriculture. App. Geogr. 34, 21–28. [CrossRef]
- Bárzana, G., Aroca, R., Ruiz-Lozano, J. M. (2015). Localized and nonlocalized effects of arbuscular mycorrhizal symbiosis on accumulation of osmolytes and aquaporins and on antioxidant systems in maize plants subjected to total or partial root drying. Plant Cell Environ. 38, 1613–1627. [CrossRef]
- Baslam, M., Garmendia, I., Goicoechea, N. (2011). Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse grown lettuce. J. Agric. Food Chem. 59, 5504–C5515. [CrossRef]
- Bati, C. B., Santilli, E., Lombardo, L. (2015). Effect of arbuscular mycorrhizal fungi on growth and on micronutrient and macronutrient uptake and allocation in olive plantlets growing under high total Mn levels. Mycorrhiza 25 (2), 97–108. [CrossRef]
- Battini, F., Grønlund, M., Agnolucci, M., Giovannetti, M., Jakobsen, I. (2017). Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria. Sci. Rep. 7, 4686. [CrossRef]
- Bauddh, K., Singh, R. P. (2012). Growth: tolerance efficiency and phytoremediation potential of Ricinus communis (L.) and Brassica juncea (L.) in salinity and drought affected cadmium contaminated soil. Ecotoxicol. Environ. Saf. 85, 13–22. [CrossRef]
- Bhantana, P., Rana, M.S., Sun, Xc. et al. Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 84, 19–37 (2021). [CrossRef]
- Birhane, E., Sterck, F., Fetene, M., Bongers, F., Kuyper, T. (2012). Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 169, 895–904. [CrossRef]
- Bona, E., Cantamessa, S., Massa, N., Manassero, P., Marsano, F., Copetta, A., et al. (2017). Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: a field study. Mycorrhiza 27, 1–C11. [CrossRef]
- Borde, M., Dudhane, M., Jite, P. K. (2010). AM fungi influences the photosynthetic activity, growth and antioxidant enzymes in Allium sativum L. under salinity condition. Not. Sci. Biol. 2, 64–71. [CrossRef]
- Bowles, T. M., Barrios-Masias, F. H., Carlisle, E. A., Cavagnaro, T. R., Jackson, L. E. (2016). Effects of arbuscular mycorrhizae on tomato yield, nutrient uptake, water relations, and soil carbon dynamics under deficit irrigation in field conditions. Sci. Total Environ. 566, 1223–1234. [CrossRef]
- Birhane, E., Sterck, F.J., Fetene, M. et al. Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 169, 895–904 (2012). [CrossRef]
- Binti T. et al. Important Role of Mycorrhiza for Seed Germination and Growth of Dendrobium Orchids. Journal of Tropical Biodiversity and Biotechnology, [S.l.], v. 6, n. 2, p. jtbb60805, june 2021. ISSN 2540-9581. Available at: <https://jurnal.ugm.ac.id/jtbb/article/view/60805>. Date accessed: 29 june 2023. /*doi:http://dx.doi.org/10.22146/jtbb.60805. */ doi:. [CrossRef]
- 35. Bucking H, Liepold E, Ambilwade P (2012) The role of the mycorrhizal symbiosis in nutrient uptake of plants and the regulatory mechanisms underlying these transport processes. World â€TM s largest Science, Technology and Medicine Open Access book publisher. Intech Open.
- Bulgarelli R. G., Marcos F. C. C., Ribeiro R. V., López de Andrade S. A. (2017). Mycorrhizae enhance nitrogen fixation and photosynthesis in phosphorus-starved soybean (Glycine max L. Merrill), Environmental and Experimental Botany pp 140 : 26-33. [CrossRef]
- Bucher, M. (2007). Functional biology of plant phosphate uptake at root and mycorrhizae interfaces. New Phytol. 173, 11–26. [CrossRef]
- Bucking, H., Kafle, A. (2015). Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: current knowledge and research gaps. Agronomy 5, 587–612. [CrossRef]
- Bunn, R., Lekberg, Y., Zabinski, C. (2009). Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants. Ecology 90 (5), 1378–1388. [CrossRef]
- Calvo-Polanco, M., Sanchez-Romera, B., Aroca, R., Asins, M. J., Declerck, S., Dodd, I. C., et al. (2016). Exploring the use of recombinant inbred lines in combination with beneficial microbial inoculants (AM fungus and PGPR) to improve drought stress tolerance in tomato. Environ. Exp. Bot. 131, 47–57. [CrossRef]
- Castellanos-Morales, V., Villegas, J., Wendelin, S., Vierheiling, H., Eder, R., Cardenas-Navarro, R. (2010). Root colonization by the arbuscular mycorrhizal fungus Glomus intraradices alters the quality of strawberry fruit (Fragaria ananassa Duch.) at different nitrogen levels. J. Sci. Food Agric. 90, 1774–1782. [CrossRef]
- Cekic, F. O., Unyayar, S., Ortas, I. (2012). Effects of arbuscular mycorrhizal inoculation on biochemical parameters in capsicum annuum grown under long term salt stress. Turk. J. Bot. 36, 63–72. [CrossRef]
- Chandrasekaran, M., Chanratana, M., Kim, K., Seshadri, S., Sa, T. (2019). Impact of arbuscular mycorrhizal fungi on photosynthesis, water status, and gas exchange of plants under salt stress—a meta-analysis. Front. Plant Sci. 10, 457. [CrossRef]
- Chen, S., Jin, W., Liu, A., Zhang, S., Liu, D., Wang, F., et al. (2013). Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress. Sci. Hort. 160, 222–229. [CrossRef]
- Chen, S., Zhao, H., Zou, C., Li, Y., Chen, Y., Wang, Z., et al. (2017). Combined Inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front. Microbiol. 8, 25–16. [CrossRef]
- Clark, R. B., Zeto, S. K. (2000). Mineral acquisition by arbuscular mycorrhizal plants. J. Plant Nutr. 23, 867–902. [CrossRef]
- De Andrade, S. A. L., Domingues, A. P., Mazzafera, P. (2015). Photosynthesis is induced in rice plants that associate with arbuscular mycorrhizal fungi and are grown under arsenate and arsenite stress. Chemosphere 134, 141–149. [CrossRef]
- Dong, Y., Zhu, Y. G., Smith, F. A., Wang, Y., Chen, B. (2008). Arbuscular mycorrhiza enhanced arsenic resistance of both white clover Trifolium repens L. and ryegrass Lolium perenne L. plants in an arsenic-contaminated soil. Environ. Pollut. 155, 174–181. [CrossRef]
- Duan, X., Neuman, D. S., Reiber, J. M., Green, C. D., Arnold M., Saxton, A. M., et al. (1996). Mycorrhizal influence on hydraulic and hormonal factors implicated in the control of stomatal conductance during drought. J. Exp. Bot. 47 (303), 1541–1550. [CrossRef]
- Duc, N. H., Csintalan, Z., Posta, K. (2018). Arbuscular mycorrhizal fungi mitigate negative effects of combined drought and heat stress on tomato plants. Plant Physiol. Biochem. 132, 297–307. [CrossRef]
- Elhindi, K. M., El-Din, S. A., Elgorban, A. M. (2017). The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 24, 170–179. [CrossRef]
- EL-Nashar, Y. I. (2017). Response of snapdragon Antirrhinum majus L. to blended water irrigation and arbuscular mycorrhizal fungi inoculation: uptake of minerals and leaf water relations. Photosynthetica 55 (2), 201–209. [CrossRef]
- El-Sawah AM, El-Keblawy A, Ali DFI, Ibrahim HM, El-Sheikh MA, Sharma A, Alhaj Hamoud Y, Shaghaleh H, Brestic M, Skalicky M, et al. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture. 2021; 11(3):194. [CrossRef]
- Evelin, H., Giri, B., Kapoor, R. (2012). Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 22, 203–217. [CrossRef]
- Evelin, H., Kapoor, R., Giri, B. (2009). Arbuscular mycorrhizal fungi in alleviation of salt stress: a review. Ann. Bot. 104, 1263–1281. [CrossRef]
- Gamalero, E., Lingua, G., Berta, G., Glick, B. R. (2009). Beneficial role of plant growth promoting bacteria and arbuscular mycorrhizal fungi on plant responses to heavy metal stress. Can. J. Microbiol. 55, 501–514. 245. [CrossRef]
- Garcés-Ruiz, M., Calonne-Salmon, M., Plouznikoff, K., Misson, C., Navarrete-Mier, M., Cranenbrouck, S., et al. (2017). Dynamics of short-term phosphorus uptake by intact mycorrhizal and non-mycorrhizal maize plants grown in a circulatory semi-hydroponic cultivation system. Front. Plant Sci. 8, 1471. [CrossRef]
- Garg, N., Chandel, S. (2012). Role of arbuscular mycorrhizal (AM) fungi on growth, cadmium uptake, osmolyte, and phytochelatin synthesis in Cajanus cajan (L.) Millsp. under NaCl and Cd stresses. J. Plant Growth Regul. 31 (3), 292–308. [CrossRef]
- Gao, X., Guo, H., Zhang, Q. et al. Arbuscular mycorrhizal fungi (AMF) enhanced the growth, yield, fiber quality and phosphorus regulation in upland cotton (Gossypium hirsutum L.). Sci Rep 10, 2084 (2020). [CrossRef]
- Garcia-Garrido JM, Tribak M, Rejon-Palomares A, Ocampo JA, Garcia-Romera I. Hydrolytic enzymes and ability of arbuscular mycorrhizal fungi to colonize roots. J Exp Bot. 2000 Aug;51(349):1443-8. PMID: 10944158.
- Gavito, M. E., Olsson, P. A., Rouhier, H., Medinapeñafiel, A., Jakobsen, I., Bago, A. (2005). Temperature constraints on the growth and functioning of root organ cultures with arbuscular mycorrhizal fungi. New Phytol. 168, 179–188. [CrossRef]
- Gholamhoseini, M., Ghalavand, A., Dolatabadian, A., Jamshidi, E., Khodaei-Joghan, A. (2013). Effects of arbuscular mycorrhizal inoculation on growth, yield, nutrient uptake and irrigation water productivity of sunflowers grown under drought stress. Agric. Water Manag. 117, 106–114. [CrossRef]
- Gianinazzi, S., Golotte, A., Binet, M. N., Van Tuinen, D., Redecker, D., Wipf, D. (2010). Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 20, 519–530. [CrossRef]
- Gomez-Bellot, M. J., Ortuño, M. F., Nortes, P. A., Vicente-Sánchez, J., Bañón, S., Sánchez Blanco, M. J. (2015). Mycorrhizal euonymus plants and reclaimed water: biomass, water status and nutritional responses. Sci. Hort. 186, 61–69. [CrossRef]
- Gutjahr, C., Paszkowski, U. (2013). Multiple control levels of root system remodeling in arbuscular mycorrhizal symbiosis. Front. Plant Sci. 4, 204. [CrossRef]
- Hameed, A., Dilfuza, E., Abd-Allah, E. F., Hashem, A., Kumar, A., Ahmad, P. (2014). “Salinity stress and arbuscular mycorrhizal symbiosis in plants,” in Use of microbes for the alleviation of soil stresses, vol. 1. Ed. Miransari, M. (NY: Springer Science+Business Media), 139–159. [CrossRef]
- Hammer, E. C., Nasr, H., Pallon, J., Olsson, P. A., Wallander, H. (2011). Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 21 (2), 117–129. [CrossRef]
- Hart, M., Ehret, D. L., Krumbein, A., Leung, C., Murch, S., Turi, C., et al. (2015). Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 25, 359–376. [CrossRef]
- Hasanuzzaman, M., Gill, S. S., Fujita, M. (2013). “Physiological role of nitric oxide in plants grown under adverse environmental conditions,” in Plant acclimation to environmental stress. Eds. Tuteja, N., Gill, S. S. (NY: Springer Science+Business Media), 269–322. [CrossRef]
- Hashem, A., Abd_Allah, E. F., Alqarawi, A. A., Aldubise, A., Egamberdieva, D. (2015). Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J. Plant Interact. 10 (1), 230–242. [CrossRef]
- Hashem, A., Alqarawi, A. A., Radhakrishnan, R., Al-Arjani, A. F., Aldehaish, H. A., Egamberdieva, D., et al. (2018). Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 25 (6), 1102–1114. [CrossRef]
- He, F., Sheng, M., Tang, M. (2017). Effects of Rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in Robinia pseudoacacia L. under drought Stress. Front. Plant Sci. 8, 183. [CrossRef]
- Hijri, M. (2016). Analysis of a large dataset form field mycorrhizal inoculation trials on potato showed highly significant increase in yield. Mycorrhiza 2, 209–214. [CrossRef]
- Impa, S. M., Nadaradjan, S., Jagadish, S. V. K. (2012). “Drought stress induced reactive oxygen species and anti-oxidants in plants,” in Abiotic stress responses in plants: metabolism, productivity and sustainability. Eds. Ahmad, P., Prasad, M. N. V. (LLC: Springer Science+ Business Media), 131–147. [CrossRef]
- Janouškova, M., Pavlíková, D. (2010). Cadmium immobilization in the rhizosphere of arbuscular mycorrhizal plants by the fungal extraradical mycelium. Plant Soil 332, 511–520. [CrossRef]
- Jiang, Y. N., Wang, W. X., Xie, Q. J., Liu, N., Liu, L. X., Wang, D. P., et al. (2017). Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 356, 1172–1175. [CrossRef]
- Jixiang, L., Yingnan, W., Shengnan, S., Chunsheng, M., Xiufeng, Y. (2017). Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition. Sci. Total Environ. 576, 234–241. [CrossRef]
- Kanwal, S., Bano, A., Malik, R. N. (2015). Effects of arbuscular mycorrhizal fungi on metals uptake, physiological and biochemical response of Medicago sativa L. with increasing Zn and Cd concentrations in soil. Am. J. Plant Sci. 6, 2906–2923. [CrossRef]
- Kapoor, R., Evelin, H., Mathur, P., Giri, B. (2013). “Arbuscular mycorrhiza: approaches for abiotic stress tolerance in crop plants for sustainable agriculture,” in Plant acclimation to environmental stress. Eds. Tuteja, N., Gill, S. S. (LLC: Springer Science+Business Media), 359–401. [CrossRef]
- Kayama, M., Yamanaka, T. (2014). Growth characteristics of ectomycorrhizal seedlings of Quercus glauca, Quercus salicina, and Castanopsis cuspidata planted on acidic soil. Trees 28, 569–583. [CrossRef]
- Kubikova, E., Moore, J. L., Ownlew, B. H., Mullen, M. D., Augé, R. M. (2001). Mycorrhizal impact on osmotic adjustment in Ocimum basilicum during a lethal drying episode. J. Plant Physiol. 158, 1227–1230. [CrossRef]
- Lehmann, A., Rillig, M. C. (2015). Arbuscular mycorrhizal contribution to copper, manganese and iron nutrient concentrations in crops—a meta-analysis. Soil Biol. Biochem. 81, 147–158. [CrossRef]
- Lehmann, A., Veresoglou, S. D., Leifheit, E. F., Rillig, M. C. (2014). Arbuscular mycorrhizal influence on zinc nutrition in crop plants: a meta-analysis. Soil Biol. Biochem. 69, 123–131. [CrossRef]
- Li, H., Chen, X. W., Wong, M. H. (2015). Arbuscular mycorrhizal fungi reduced the ratios of inorganic/organic arsenic in rice grains. Chemosphere 145, 224–230. [CrossRef]
- Li, H., Luo, N., Zhang, L. J., Zhao, H. M., Li, Y. W., Cai, Q. Y., et al. (2016a). Do arbuscular mycorrhizal fungi affect cadmium uptake kinetics, subcellular distribution and chemical forms in rice? Sci. Total Environ. 571, 1183–1190. [CrossRef]
- Li, J., Meng, B., Chai, H., Yang, X., Song, W., Li, S., et al. (2019). Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 10, 499. [CrossRef]
- Li, X., Zeng, R., Liao, H. (2016b). Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 58, 193–202. [CrossRef]
- Liu, C., Ravnskov, S., Liu, F., Rubæk, G. H., Andersen, M. N. (2018). Arbuscular mycorrhizal fungi alleviate abiotic stresses in potato plants caused by low phosphorus and deficit irrigation/partial root-zone drying. J. Agric. Sci. 156, 46–58. [CrossRef]
- Liu, X., Song, Q., Tang, Y., Li, W., Xu, J., Wu, J., et al. (2013). Human health risk assessment of heavy metals in soil–vegetable system: a multi-medium analysis. Sci. Total. Environ. 463–464, 530–540. [CrossRef]
- Ludwig-Müller, J. (2010). “Hormonal responses in host plants triggered by arbuscular mycorrhizal fungi,” in Arbuscular mycorrhizas: Physiology and function. Eds. Koltai, H., Kapulnik, Y. (Dordrecht: Springer), 169–190. [CrossRef]
- Luginbuehl, L. H., Menard, G. N., Kurup, S., Van Erp, H., Radhakrishnan, G. V., Breakspear, A., et al. (2017). Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 356, 1175–1178. [CrossRef]
- 92. Martin CA, Stutz JC (2004). Interactive effects of temperature and arbuscular mycorrhizal fungi on growth, P uptake and root respiration of Capsicum annuum L. Mycorrhiza. 14:241-4. [CrossRef] [PubMed]
- Maya, M. A., Matsubara, Y. (2013). Influence of arbuscular mycorrhiza on the growth and antioxidative activity in Cyclamen under heat stress. Mycorrhiza 23 (5), 381–390. [CrossRef]
- Mena-Violante, H. G., Ocampo-Jimenez, O., Dendooven, L., Martinez-Soto, G., Gonzalez-Castafeda, J., Davies, F. T., et al. (2006). Arbuscular mycorrhizal fungi enhance fruit growth and quality of chile ancho Capsicum annuum L. cv San Luis plants exposed to drought. Mycorrhiza 16, 261–267. [CrossRef]
- Miransari, M. (2017). “Arbuscular mycorrhizal fungi and heavy metal tolerance in plants,” in Arbuscular mycorrhizas and stress tolerance of plants. Ed. Wu, Q. S. (Singapore: Springer Nature), 174–161. [CrossRef]
- Mitra, D., Navendra, U., Panneerselvam, U., Ansuman, S., Ganeshamurthy, A. N., Divya, J. (2019). Role of mycorrhiza and its associated bacteria on plant growth promotion and nutrient management in sustainable agriculture. Int. J. Life Sci. Appl. Sci. 1, 1–10.
- 97. Mingsen Qin, Qi Zhang, Jianbin Pan, Shengjing Jiang, Yongjun Liu, Ali Bahadur, Zhenling Peng, Yue Yang, Huyuan Feng (2019). Hydrolytic enzymes and ability of arbuscular mycorrhizal fungi to colonize roots. European Journal of Soil Science. 71: 84-92. [CrossRef]
- Moghadam, H. R. T. (2016). Application of super absorbent polymer and ascorbic acid to mitigate deleterious effects of cadmium in wheat. Pesqui. Agropecu. Trop. 6 (1), 9–18. [CrossRef]
- Moradtalab, N., Roghieh, H., Nasser, A., Tobias, E. H., Günter, N. (2019). Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 9, 41. [CrossRef]
- Morte, A., Lovisolo, C., Schubert, A. (2000). Effect of drought stress on growth and water relations of the mycorrhizal association Helianthemum almeriense–Terfezia claveryi. Mycorrhiza 10, 115–119. [CrossRef]
- Mohammadi K., Shiva K. ,Yousef S., Gholamreza H. (2011). A Review: Beneficial Effects of the Mycorrhizal Fungi for Plant Growth. J. Appl. Environ. Biol. Sci., 1:310-319.
- Navarro, J. M., Perez-Tornero, O., Morte, A. (2014). Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the root stock salt tolerance. J. Plant Physiol. 171 (1), 76–85. [CrossRef]
- Nell, M., Wawrosch, C., Steinkellner, S., Vierheilig, H., Kopp, B., Lössl, A. (2010). Root colonization by symbiotic arbuscular mycorrhizal fungi increases sesquiterpenic acid concentrations in Valeriana officinalis L. Planta Med. 76, 393–398. [CrossRef]
- Nouri, E., Breuillinsessoms, F., Feller, U., Reinhardt, D. (2015). Phosphorus and nitrogen regulate arbuscular mycorrhizal symbiosis in Petunia hybrid. PLoS One 9, e90–841. [CrossRef]
- Orfanoudakis, M., Wheeler, C. T., Hooker, J. E. (2010). Both the arbuscular mycorrhizal fungus Gigaspora rosea and Frankia increase root system branching and reduce root hair frequency in Alnus glutinosa. Mycorrhiza 20, 117–126. [CrossRef]
- Orine D, Defossez E, Vergara F,Uthe H, van Dam NM, Rasmann S. Arbuscular mycorrhizal fungi prevent the negative effect of drought and modulatethe growth-defence trade-off in tomato plants. J Sustain Agric Environ. 2022;1:177–190. [CrossRef]
- OrtasI. (2010). Effect of mycorrhiza application on plant growth and nutrient uptake in cucumber production under field conditions. Spanish Journal of Agricultural Research, 8(S1), 116-122. [CrossRef]
- Ortas, I. (2012). The effect of mycorrhizal fungal inoculation on plant yield, nutrient uptake and inoculation effectiveness under long-term field conditions. Field Crops Res. 125, 35–48. [CrossRef]
- Ouziad, F., Hildebrandt, U., Schmelzer, E., Bothe, H. (2005). Differential gene expressions in arbuscular mycorrhizal-colonized tomato grown under heavy metal stress. J. Plant Physiol. 162, 634–649. [CrossRef]
- Paterson, E., Sim, A., Davidson, J., Daniell, T. J. (2016). Arbuscular mycorrhizal hyphae promote priming of native soil organic matter mineralization. Plant Soil. 408, 243–C254. [CrossRef]
- Plassard, C., Dell, B. (2010). Phosphorus nutrition of mycorrhizal trees. Tree Physiol. 30, 1129–1139. [CrossRef]
- Prasad, R., Bhola, D., Akdi, K., Cruz, C., Sairam, K. V. S. S., Tuteja, N., et al. (2017). Introduction to mycorrhiza: historical development,” in Mycorrhiza. Eds. Varma, A., Prasad, R., Tuteja, N. (Cham: Springer), 1–7. [CrossRef]
- Pringle, A., Bever, J. D., Gardes, M., Parrent, J. L., Rillig, M. C., Klironomos, J. N. (2009). Mycorrhizal symbioses and plant invasions. Ann. Rev. Ecol. Evol. Syst. 40, 699–715. [CrossRef]
- Punamiya, P., Datta, R., Sarkar, D., Barber, S., Patel, M., Da, P. (2010). Symbiotic role of Glomus mosseae in phytoextraction of lead in vetiver grass Chrysopogon zizanioides L. J. Hazard. Mater. 177, 465–474. [CrossRef]
- Rani, B. (2016) Effect of arbuscular mycorrhiza fungi on biochemical parameters in wheat Triticum aestivum L. under drought conditions. Doctoral dissertation, CCSHAU, Hisar.
- Rodriguez, R. J., Henson, J., Van Volkenburgh, E., Hoy, M., Wright, L., Beckwith, F., et al. (2008). Stress tolerance in plants via habitat-adapted symbiosis. Int. Soc. Microb. Ecol. 2, 404–416. [CrossRef]
- Ronsheim M. L. "The Effect of Mycorrhizae on Plant Growth and Reproduction Varies with Soil Phosphorus and Developmental Stage," The American Midland Naturalist 167(1), 28-39, (1 January 2012). [CrossRef]
- Rouphael, Y., Franken, P., Schneider, C., Schwarz, D., Giovannetti, M., Agnolucci, M. (2015). Arbuscular mycorrhizal fungi act as bio-stimulants in horticultural crops. Sci. Hort. 196, 91–108. [CrossRef]
- Ruiz-Lozano, J. M. (2003). Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. Mycorrhiza 13, 309–317. [CrossRef]
- Ruiz-Lozano, J. M., Aroca, R., Zamarreño, Á.M., Molina, S., Andreo-Jiménez, B., Porcel, R., et al. (2015). Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. Plant Cell Environ. 39 (2), 441–452. [CrossRef]
- Ruiz-Sánchez, M., Aroca, R., Muñoz, Y., Polón, R., Ruiz-Lozano, J. M. (2010). The arbuscular mycorrhizal symbiosis enhances the photosynthetic efficiency and the antioxidative response of rice plants subjected to drought stress. J. Plant Physiol. 167, 862–869. [CrossRef]
- Sabia, E., Claps, S., Morone, G., Bruno, A., Sepe, L., Aleandri, R. (2015). Field inoculation of arbuscular mycorrhiza on maize (Zea mays L.) under low inputs: preliminary study on quantitative and qualitative aspects. Italian J. Agron. 10, 30–33. [CrossRef]
- Sadhana, B. (2014). Arbuscular mycorrhizal fungi (AMF) as a biofertilizers—a review. Int. J. Curr. Microbiol. App. Sci. 3 (4), 384–400.
- Salam, E. A., Alatar, A., El-Sheikh, M. A. (2017). Inoculation with arbuscular mycorrhizal fungi alleviates harmful effects of drought stress on damask rose. Saudi J. Biol. Sci. 25 (8), 1772–1780. [CrossRef]
- Sameera A. Alghamdi (2019). Influence of mycorrhizal fungi on seed germination and growth in terrestrial and epiphytic orchids. Saudi Journal of Biological Sciences. 26: 495-502, . [CrossRef]
- Santander, C., Sanhueza, M., Olave, J., Borie, F., Valentine, C., Cornejo, P. (2019). Arbuscular mycorrhizal colonization promotes the tolerance to salt stress in lettuce plants through an efficient modification of ionic balance. J. Soil Sci. Plant Nutr. 19 (2), 321–331. [CrossRef]
- Sbrana, C., Avio, L., Giovannetti, M. (2014). Beneficial mycorrhizal symbionts affecting the production of health-promoting phytochemicals. Electrophoresis 35, 1535–1546. [CrossRef]
- Selosse, M. A., Strullu-Derrien, C., Martin, F. M., Kamoun, S., Kenrick, P. (2015). Plants, fungi and oomycetes: a 400-million years affair that shapes the biosphere. New Phytol. 206, 501–506. [CrossRef]
- Sharma, S., Prasad, R., Varma, A., Sharma, A. K. (2017). Glycoprotein associated with Funneliformis coronatum, Gigaspora margarita and Acaulospora scrobiculata suppress the plant pathogens in vitro. Asian J. Plant Pathol. 11 (4), 192–202. [CrossRef]
- Shen, H., Christie, P., Li, X. (2006). Uptake of zinc, cadmium and phosphorus by arbuscular mycorrhizal maize (Zea mays, L.) from a low available phosphorus calcareous soil spiked with zinc and cadmium. Environ. Geochem. Health 28, 111. [CrossRef]
- Sheng, M., Tang, M., Zhang, F., Huang, Y. (2011). Influence of arbuscular mycorrhiza on organic solutes in maize leaves under salt stress. Mycorrhiza 21, 423–430. [CrossRef]
- Smith, S. E., Read, D. J. (1997). Mycorrhizal symbiosis. San Diego: Academic Press, 607.
- Smith, S., Read, D. (2008). Mycorrhiza symbiosis, 3rd Ed. San Diego, CA: Academic Press.
- Smith, S. E., Smith, F. A., Jakobsen, I. (2003). Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 133, 16–20. [CrossRef]
- Souza, L. A., Andrade, S. A. L., Souza, S. C. R., Schiavinato, M. A. (2012). Evaluation of mycorrhizal influence on the development and phytoremediation potential of Canavalia gladiata in Pb contaminated soils. Int. J. Phytorem. 15, 465–476. [CrossRef]
- 136. Sun X F, Su Y Y, Zhang Y, et al. Diversity of arbuscular mycorrhizal fungal spore communities and its relations to plants under increased temperature and precipitation in a natural grassland. Chin Sci Bull, 2013, 58: 41094119, . [CrossRef]
- Takács, T., Vörös, I. (2003). Effect of metal non-adapted arbuscular mycorrhizal fungi on Cd, Ni and Zn uptake by ryegrass. Acta Agron. Hung. 51, 347–354.
- Talaat, N. B., Shawky, B. T. (2014). Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 98, 20–31. [CrossRef]
- Thirkell, T. J., Charters, M. D., Elliott, A. J., Sait, S. M., Field, K. J. (2017). Are mycorrhizal fungi our sustainable saviours considerations for achieving food security. J. Ecol. 105, 921–929. [CrossRef]
- Turrini, A., Bedini, A., Loor, M. B., Santini, G., Sbrana, C., Giovannetti, M., et al. (2018). Local diversity of native arbuscular mycorrhizal symbionts differentially affects growth and nutrition of three crop plant species. Biol. Fertil. Soils 54, 203–217. [CrossRef]
- Van der Heijden, M. G., Martin, F. M., Selosse, M. A., Sanders, I. R. (2015). Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol. 205, 1406–1423. [CrossRef]
- Wagg, C., Barendregt, C., Jansa, J., Heijden, M. G. A. (2015). Complementarity in both plant and mycorrhizal fungal communities are not necessarily increased by diversity in the other. J. Ecol. 103, 1233–1244. [CrossRef]
- Wahid, A., Gelani, S., Ashraf, M., Foolad, M. R. (2007). Heat tolerance in plants: an overview. Environ. Exp. Bot. 61, 199–223. [CrossRef]
- Wang, Y., Jing, H., Gao, Y. (2012). Arbuscular mycorrhizal colonization alters subcellular distribution and chemical forms of cadmium in Medicago sativa L. and resists cadmium toxicity. PLoS One 7, 3161–3164. [CrossRef]
- Wang, Y., Wang, M., Li, Y., Wu, A., Huang, J. (2018). Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS One 13 (4), e0196408. [CrossRef]
- Wu, Z., McGrouther, K., Huang, J., Wu, P., Wu, W., Wang, H. (2014). Decomposition and the contribution of glomalin-related soil protein (GRSP) in heavy metal sequestration: field experiment. Soil Biol. Biochem. 68, 283–290. [CrossRef]
- Yang, S., Li, F., Malhi, S. S., Wang, P., Dongrang, S., Wang, J. (2004). Long term fertilization effects on crop yield and nitrate nitrogen accumulation in soil in Northwestern China. Agron. J. 96, 1039–1049. [CrossRef]
- Yang X, Mariotte P, Guo J, Hautier Y, Zhang T. Suppression of arbuscular mycorrhizal fungi decreases the temporal stability of community productivity under elevated temperature and nitrogen addition in a temperate meadow. Sci Total Environ. 2021 Mar 25;762:143137. [CrossRef] [PubMed]
- Yin, N., Zhang, Z., Wang, L., Qian, K. (2016). Variations in organic carbon, aggregation, and enzyme activities of gangue-fly ash-reconstructed soils with sludge and arbuscular mycorrhizal fungi during 6-year reclamation. Envi. Sci. Pollut. Res. 23 (17), 17840–17849. [CrossRef]
- Yooyongwech, S., Phaukinsang, N., Cha-Um, S., Supaibulwatana, K. (2013). Arbuscular mycorrhiza improved growth performance in Macadamia tetraphylla L. grown under water deficit stress involves soluble sugar and proline accumulation. Plant Growth Regul. 69, 285–293. [CrossRef]
- Yooyongwech, S., Samphumphuang, T., Tisarum, R., Theerawitaya, C., Chaum, S. (2016). Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline. Sci Hort. 198, 107–117. [CrossRef]
- Yost, R. S., Fox, R. L. (1982). Influence of mycorrhizae on the mineral contents of cowpea and soybean grown in an oxisol. Agron. J. 74 (3), 475–481. [CrossRef]
- Yousaf, B., Liu, G., Wang, R., Imtiaz, M., Zia-ur-Rehman, M., Munir, M. A. M., et al. (2016). Bioavailability evaluation, uptake of heavy metals and potential health risks via dietary exposure in urban-industrial areas. Environ. Sci. Pollut. Res. 23, 22443–22453. [CrossRef]
- Zeng, L., JianFu, L., JianFu, L., MingYuan, W. (2014). Effects of arbuscular mycorrhizal (AM) fungi on citrus quality under nature conditions. Southwest China J. Agric. Sci. 27, 2101–2105. [CrossRef]
- Zhang, F., Jia-Dong, H. E., Qiu-Dan, N. I., Qiang-Sheng, W. U., Zou, Y. N. (2018a). Enhancement of drought tolerance in trifoliate orange by mycorrhiza: changes in root sucrose and proline metabolisms. Not. Bot. Horti. Agrobot. Cluj-Napoca 46, 270. [CrossRef]
- Zhang, T., Hub, Y., Zhang, K., Tian, C., Gu, J. (2018b). Arbuscular mycorrhizal fungi improve plant growth of Ricinus communis by altering photosynthetic properties and increasing pigments under drought and salt stress. Ind. Crop. Prod. 117, 13–19. [CrossRef]
- .
- 7878.
- 159. Zhu X, Song F, Xu H. Influence of arbuscular mycorrhiza on lipid peroxidation and antioxidant enzyme activity of maize plants under temperature stress. Mycorrhiza. 2010 Jun;20(5):325-32. [CrossRef] [PubMed]
- Zhu, X., Song, F., Liu, S., Liu, T., Zhou, X. (2012). Arbuscular mycorrhizae improve photosynthesis and water status of Zea mays L. under drought stress. Plant Soil Environ. 58, 186–191. [CrossRef]
- Zhu, X. C., Song, F. B., Xu, H. W. (2010a). Arbuscular mycorrhizae improve low temperature stress in maize via alterations in host water status and photosynthesis. Plant Soil. 331, 129–137. [CrossRef]
- Zhu, X. C., Song, F. B., Liu, S. Q., Liu, F. L. (2016). Arbuscular mycorrhiza improve growth, nitrogen uptake, and nitrogen use efficiency in wheat grown under elevated CO2. Mycorrhiza 26, 133–140. [CrossRef]
- Zhu, X. C., Song, F. B., Xu, H. W. (2010b). Effects of arbuscular mycorrhizal fungi on photosynthetic characteristics of maize under low temperature stress. Acta Ecol. Sin. 21, 470–475. [CrossRef]
- Zou, Y. N., Srivastava, A. K., Wu, Q. S. (2016). Glomalin: a potential soil conditioner for perennial fruits. Int. J. Agric. Biol. 18, 293–297. [CrossRef]
- Zou, YN., Qin, QY., Ma, WY. et al. Metabolomics reveals arbuscular mycorrhizal fungi-mediated tolerance of walnut to soil drought. BMC Plant Biol 23, 118 (2023). [CrossRef]
Figure 1.
Process via which AMF and plants develop symbiosis.
Figure 2.
Diagrammatic representation of functions of mychorriza in regulating various processes in the environment and plant growth promotion under abiotic stress conditions.
Figure 2.
Diagrammatic representation of functions of mychorriza in regulating various processes in the environment and plant growth promotion under abiotic stress conditions.
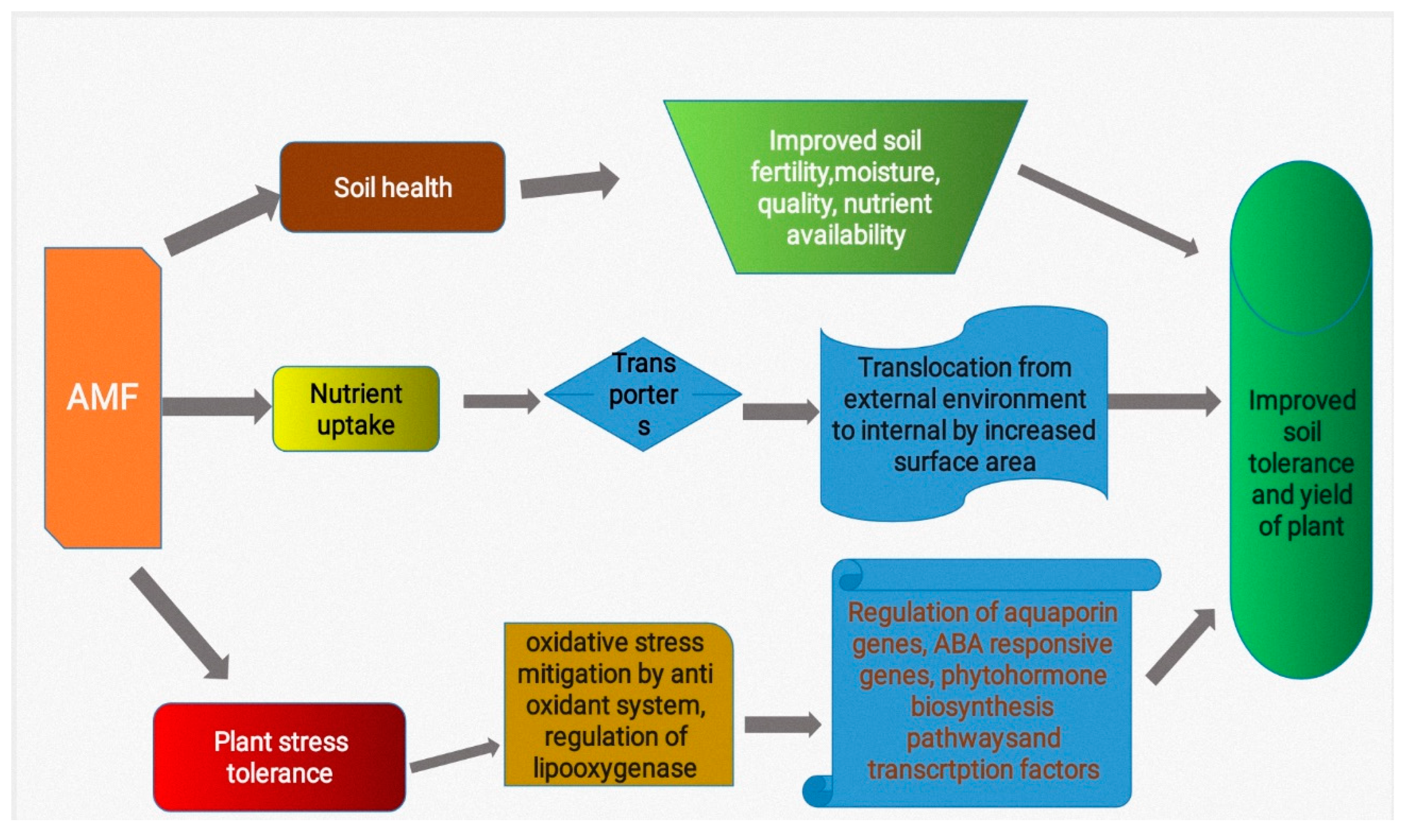
Figure 3.
AMF inoculation and how it mitigates temperature stress in plants.
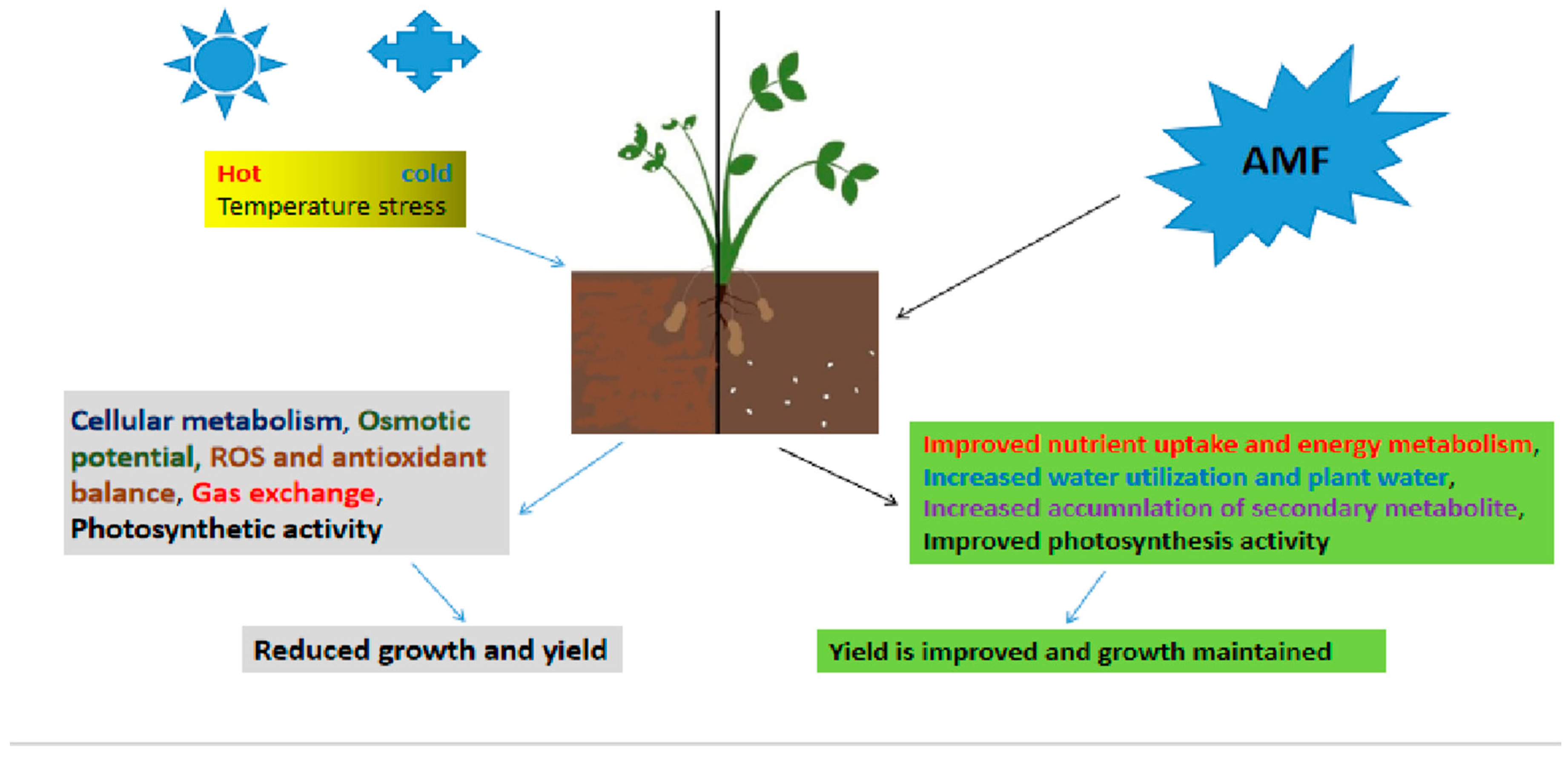
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



