
Submitted:
22 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Transposable elements (TEs) are repetitive elements which make up around 45% of the human genome. A class of TEs known as SINE-VNTR-Alu (SVA) demonstrate the capacity to mobilise throughout the genome, resulting in SVA polymorphisms for presence or absence within the population. Although studies have previously highlighted the involvement of TEs within neurodegenerative diseases, such as Parkinson’s disease and amyotrophic lateral sclerosis (ALS), however the exact mechanism has yet to be identified. In this study we used whole genome sequencing and RNA sequencing data of ALS patients and healthy controls from the New York Genome Center ALS Consortium, to elucidate the influence of reference SVA elements on gene expression genome-wide within central nervous system (CNS) tissues. To investigate this, we applied matrix expression quantitative trait loci analysis and demonstrated that reference SVA insertion polymorphisms can significantly modulate the expression of numerous genes, preferentially in the trans position, and in a tissue-specific manner. We also highlight that SVAs significantly regulate mitochondrial genes as well as genes within the HLA and MAPT loci, previously associated within neurodegenerative disease. In conclusion, this study continues to bring to light the effects of polymorphic SVAs on gene regulation and further highlights the importance of TEs within disease pathology.
Keywords:
SINE-VNTR-Alu
; transposable elements
; whole genome sequencing
; RNA-seq
; quantitative trait loci
; Amyotrophic Lateral Sclerosis
; HLA
; MAPT
1. Introduction
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease, originally termed by the French neurologist Jean-Martin Charcot in 1869 to describe muscular atrophy (amyotrophic) and tissue scarring and hardening of tissue within the lateral spinal cord [1,2]. ALS is the most common form of motor neuron disease (MND) and is characterized by the progressive deterioration of both upper and lower motor neurons within the brain and spinal cord [2,3]. The incidence of which is 1.75 – 3 per 100 000 people per year and an increased incidence of 4 – 8 per 100 000 people per year in the highest ALS risk age group (45 – 75 years old) [4]. Categorised as 2 groups, ALS can present as familial ALS where at least one family member of the affected individual has ALS, accounting for up to 10% of ALS cases, or sporadic ALS (sALS) where the affected individual has no prior family history which accounts for 90 – 95% of cases [5]. The clinical features of typical ALS patients consist of muscle spasticity, atrophy, muscle wasting, weakness and death due to respiratory failure, with the average survival after symptom onset lying between 3-5 years [1,4,6].
Neurodegenerative diseases are complex disorders, involving both environmental and genetic factor interactions. Therefore, attempts to elucidate disease mechanisms and pathogenic genetic variants are essential. Previous research has identified more than 30 genes associated with ALS, highlighting four genes SOD1, TARDBP (TDP-43), C90RF72 and FUS for harbouring pathogenic mutations which cause the greatest number of ALS cases [7,8,9,10]. Although these four genes have been identified as major ALS-associated genes, a 2017 meta-analysis study demonstrated that within European and Asian populations, these genes only contribute to 47.7% and 5.2% of familial and sporadic cases, respectively [11]. Following the identification of these pathogenic mutations, several pathological mechanisms have been implicated in ALS pathogenesis including oxidative stress, mitochondrial dysfunction, axonal transport, inflammation, toxic protein aggregation and RNA metabolism and toxicity [7]. In addition to identified genetic variants only explaining a small fraction of sporadic aetiology, twin studies have highlighted the importance of genetic risk factors within sporadic ALS, estimating a 61% heritability [12]. Therefore, as the exact causation of ALS is still undetermined, a better understanding of disease pathogenesis and identification of genetic biomarkers is essential. To further investigate the missing heritability of ALS, Theunissen et al., proposed structural variations (SVs) as an area of potential significance [13]. SVs are classified as insertions, inversions, deletions and microsatellites, usually of repetitive structure, that are predominantly present within non-coding DNA regions and contribute towards genomic variation [13,14]. Furthermore, SVs have demonstrated the ability to modulate gene expression and have already been implicated in neurodegenerative diseases, such as the C9orf72 repeat expansion in ALS and frontotemporal dementia (FTD) [15,16]. Hence, as 99% of the human genome is non-coding DNA, continued research into these regions is crucial to provide new insights into disease pathogenesis and to identify new potential targets for therapeutics.
Repetitive DNA is a major contributor to structural genomic variation. One form of repetitive DNA is a group of endogenous transposable elements (TEs), which can exist in both a static form and a mobile form. TEs are categorised into two classes known as DNA transposons and retrotransposons, whereby the later class possesses the ability to propagate throughout the genome via a ‘copy-and-paste’ mechanism, involving an RNA intermediate [17]. This results in the insertion of a new retrotransposon copy at a new locus within the host genome. Although, DNA transposons are capable of mobilisation via a ‘cut-and-paste’ mechanism, they are not currently active within the human genome [8,18]. Originally dismissed as “junk” DNA, TEs are known to drive genetic diversity not only by contributing to regulation and evolution of the genome but also by contributing to genetic instability and disease progression [18,19]. Previous research by Prudencio et al., highlighted the potential implication of retrotransposons in ALS through the analysis of repetitive element expression using RNA sequencing data from both healthy controls as well as C9orf72 expansion positive carriers and sporadic ALS patients (C9orf72-negative) [20]. This research revealed that repetitive element expression, including retrotransposons, was significantly increased in ALS patients with the C9orf72 expansion in comparison to C9orf72-negative patients and healthy controls, thus, suggesting retrotransposon involvement in ALS [20]. However, until recently these elements have been largely overlooked in relation to neurodegenerative diseases, even though TEs constitute to around 45% of the human genome [21].
Retrotransposons are further subdivided into two groups dependent on the presence of long terminal repeats (LTRs), known as LTR and non-LTR retrotransposons [18]. SINE-VNTR-Alu (SVA) elements are a member of the non-LTR retrotransposon family, which are typically 0.7 – 4kb in length [22]. SVAs are classified by evolutionary age based on their SINE-R region into sub-family groups A-F, whereby subfamily SVA-F is the youngest in evolutionary history [23]. Full-length SVA elements contain a 5’ CT element, Alu-like region, GC-rich VNTR (variable number tandem repeat), SINE (short interspersed nuclear element)-R domain and a 3’ poly-A tail [23] (Figure 1b). SVAs are major contributors to genetic diversity through a variety of different mechanisms, including acting as transcriptional regulators by providing alternative splice sites, polyadenylation signals and promoters whilst harbouring sites for transcription factor (TF) binding, thus, modulating gene expression [24]. SVAs are not only polymorphic in structure but ongoing mobilisation has resulted in SVAs being polymorphic for presence or absence within the genome and thus are termed retrotransposon insertion polymorphisms (RIPs) [19]. This adds additional layers of complexity to gene expression dynamics, indicating that SVAs could be associated with predisposition to disease [25].
Previous work within our group has shown that seven SVAs polymorphic for their presence or absence were significantly associated with Parkinson’s disease progression and differential gene expression in Parkinson’s disease (PD) patients using the Parkinson’s Progression Markers Initiative (PPMI) cohort [22,26]. This involved an SVA named SVA_67 which is located at the MAPT locus, a locus which has been implicated in neurodegenerative disease risk including PD, FTD and Alzheimer’s disease [27,28,29]. By using a clustered regularly interspaced short palindromic repeats (CRISPR) cell line knock-out model for SVA_67, we demonstrated that this SVA was significantly associated with differential gene expression of three genes at the MAPT locus, associated with neurodegeneration [30].
In this study, we analysed SVAs which are present in the human reference genome identified as RIPs in the cohort analysed, herein termed reference SVA RIPs to investigate differential gene expression patterns associated with ALS. For this analysis, we used whole genome sequencing (WGS) and transcriptomic data obtained from the New York Genome Center (NYGC) ALS Consortium, to elucidate the role of SVA insertion polymorphisms on gene expression in central nervous system (CNS) tissues in healthy controls and ALS patients (Figure 1a). Our analysis demonstrated that reference SVA RIPs regulate single and multiple gene targets genome-wide within CNS tissues and display tissue-specific gene modulation by comparing spinal cord, cerebellum, motor cortex and frontal cortex tissue analyses. Furthermore, we discovered that SVA RIPs influence the expression of genes at loci (HLA and MAPT) previously associated with ALS, highlighting SVA regulation of genes at this locus could be a potential mechanism involved in ALS pathology.
2. Results
2.1. SVA RIPs act as eQTL genome wide in CNS tissues
To determine the impact of reference SVAs RIPs on gene expression profiles, we analysed WGS and transcriptomic data of both ALS patients (n=1544) and healthy controls (n=359) from the ALS Consortium cohort. For this analysis, we initially combined data from different CNS tissues, including motor cortex, frontal cortex, cerebellum, occipital cortex, temporal cortex, hippocampus, and spinal cord tissue (Figure 2).
For eQTL analysis we assessed changes at a gene-based level. We demonstrate that, 14,830 genomic loci were significantly differentially modulated by reference SVA insertion polymorphism (FDR p<0.05). Of these effects, 167 were cis regulated by SVA RIPs, whilst over 98% (14,663 targets) were trans-regulated, where trans-regulated is defined as genes regulated greater than 1Mb away from the SVA site (Figure 3a). This indicates that most regulatory effects observed by these SVA RIPs impact target genes >1Mb away from the SVA element itself, these could be both direct and indirect effects (Figure 1c). In addition, single SVA elements had the capacity to regulate either a single gene or multiple gene targets. Our analysis identified that 92 reference SVA RIPs were responsible for the modulation of the 14,830 genomic loci, with SVA_16 affecting the greatest number of genes (917) (Figure 3b). Of these 92 SVA elements polymorphic for their presence or absence, 26 SVAs affects on more than 200 genes in total, whilst the lowest number of targets for an SVA in this analysis was 6, indicating that all reference SVA RIPs analysed in this study had the ability to modulate expression profiles of multiple gene targets in the CNS (Figure 3b).
2.2. Mitochondrial genes are significantly modulated by SVA RIPs
To further investigate the capacity of reference SVA elements to act as eQTL, we analysed the beta values obtained from the eQTL analysis, to determine the magnitude of the measured effects induced by SVA RIPs. The ten reference SVAs RIPs with the greatest effect size on gene upregulation (highest beta value) and gene downregulation (lowest beta value) and the affected gene targets are displayed in Figure 4a,b. All top ten hits for the highest and lowest beta values are trans-regulatory effects. SVA_55 demonstrated the greatest increase in activation of the MBP gene with a beta coefficient 272,847, whilst SVA_70 demonstrated the greatest repressive effect of the MT-ND1 gene with a beta coefficient of -36441 (FDR p<0.05) (Figure 4a,b). Interestingly, it was noted that the only two SVAs (SVA_55 and SVA_15) were responsible for the top ten hits regarding the effect size for gene activation. Furthermore, although the top hit for effect size for gene repression was SVA_70 modulating MT-ND1 expression, our results also demonstrated that the presence of both SVA_15 and SVA_55 showed the opposite effect by upregulating MT-ND1 expression (Figure 4a,b). To determine the impact of SVA presence on gene modulation in more detail, we examined the top hits for SVAs with the greatest effect on gene upregulation (SVA_55) and downregulation (SVA_70) independently. For MBP, the presence of two copies/alleles (PP genotype) of SVA_55 significantly upregulated MBP gene expression compared to the presence of one copy of SVA_55 (PA genotype) or complete absence (AA genotype) (p=0.0218) (Figure 4c). In comparison to the AA and PA genotypes, individuals with the PP genotype displayed a 9.5-fold and 19.2-fold increase in MBP gene expression, respectively. In contrast, individuals homozygous present for SVA_70 demonstrated significant repression of the MT-ND1 gene in comparison to individuals heterozygous for SVA_70, namely down-regulating gene expression of MT-ND1 by 55% (p=0.0098) (Figure 4d). No statistical significance for differential gene expression between the PP and AA genotype was obtained for MT-ND1 expression using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR p<0.05).
We next analysed the top ten most significant SVA effects based on FDR p-value. The most significant effects observed (lowest FDR p-value) were SVA_67 acting on the genes MAPK8IP1P2 and ENSG00000285668.1, displaying an FDR p-value of 1.93E-303 (Table 1). Followed by SVA_67 acting on the gene LRRC37A, with an FDR p-value of 5.12E-299 (Table 1). Intriguingly, 6/10 hits for highest significance were cis-specific effects on gene upregulation by SVA_24, SVA_33 and SVA_58 and gene downregulation by SVA_67. This is different to our previous results as the top ten reference SVAs with the greatest effect size on upregulation and downregulation were all trans-regulatory effects. Table 1 also shows that the four most significant trans-specific effects induced by SVA_15, SVA_84, SVA_87, and SVA_93 all influenced the same mitochondrial gene MTND4P24, whereby SVA_84, SVA_87, and SVA_93 increased gene expression and SVA_15 repressed gene expression. This demonstrates that multiple SVA RIPs can act on one gene influencing gene regulation, simultaneously activating and repressing gene expression.
To illustrate the impact of SVA_67 on target gene expression, we investigated the specific influence of SVA_67 allele dosage on transcriptomic profiles of the respective target gene (Figure 5). As an example, this analysis is shown for the genes MAPK8IP1P2 and LRRC37A, whereby gene expression was stratified by SVA_67 genotype (PP, PA, and AA). Analysis using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR<0.05) demonstrated that SVA_67 presence significantly downregulated gene expression of both MAPK8IP1P2 and LRRC37A (p<0.001) (Figure 5). Significant differences were observed between all genotype groups PP vs PA (MAPK8IP1P2 p=7.33E-237, LRRC37A p=1.95E-204), PP vs AA (MAPK8IP1P2 p=4.26E-55, LRRC37A p=1.06E-39), and PA vs AA (MAPK8IP1P2 p=1.33E-07, LRRC37A p=2.13E-12) for both genes (Figure 5). For individuals homozygous absent for SVA_67, a change of 262-fold and 6-fold was observed for gene expression of MAPK8IP1P2 and LRRC37A respectively, in comparison to individuals homozygous present for SVA_67 (Figure 5).
2.3. SVA RIPs demonstrate cis effects on HLA and MAPT loci
To elucidate the impact of cis-acting SVA RIPs, we explored which SVA elements had the largest significant effects on genes regulated in cis, including gene upregulation (positive beta value) and downregulation (negative beta value) using beta values from matrix eQTL analysis (FDR p<0.05) (Figure 6). Here, we highlight that SVA_67 largely influences gene regulation in cis of six different genes, including the upregulation of MAPT and LRRC37A4P and downregulation of MAPK8IP1P2, LRRC37A2, LRRC37A, and KANSL1 (Figure 6a). SVA_67 was responsible for the two greatest increases in gene activation of the MAPT and LRRC37A4P genes with beta coefficients of 1384 and 1101, respectively (Figure 6a). Interestingly, six of the ten most positive hits were effects induced on HLA genes by SVA_24, SVA_25, SVA_27 and SVA_88 (Figure 6a). Furthermore, we demonstrate that SVA_73 had the greatest effect out of all cis-regulatory effects with a beta coefficient of -2843 for the gene FCGBP, indicating a large inhibitory effect of FCGBP gene expression (Figure 6b). FCGBP gene expression is further repressed by SVA_72, the second highest hit in our analysis for the top ten SVAs with the greatest effect size on gene downregulation (Figure 6b).
To determine the influence of cis-acting SVA RIPs on gene regulation, we next analysed gene expression patterns of the top two hits for the largest effects on gene upregulation and downregulation (Figure S1). The presence of either one (PA) or two (PP) SVA_67 alleles significantly upregulated both LRRC37A4P and MAPT gene expression when tested using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR p<0.05) (Figure S1a,b). In comparison to individuals homozygous absent for SVA_67, homozygous present and heterozygous individuals displayed a 447.3-fold (p=1.04E-40) and a 214.4-fold (p=2.55E-38) increase in LRRC37A4P expression and a 1.4-fold (p=2.53E-09) and 1.2-fold (p=2.51E-04) increase in MAPT gene expression, respectively (Figure S1a,b). Analysis of the top two hits for the greatest effect on gene downregulation displayed differing gene expression patterns between the influence of SVA_73 and SVA_72 on FCGBP gene expression (Figure S1c,d). When comparing to the complete absence (AA) of SVA_73, the PA and PP genotype groups displayed a significant 3.7-fold (p=3.61E-02) and 10-fold (p=2.61E-03) increase in FCGBP gene expression (Figure S1c). Although our analysis displayed that the presence of SVA_73 upregulates the FCGBP gene in comparison to AA individuals, FCGBP gene expression is significantly repressed by 63% (p=3.29E-08) in the PP genotype group compared to the PA genotype group (Figure S1c). SVA_72 analysis highlighted that homozygous presence of the SVA_72 allele significantly downregulated FCGBP gene expression by 23% (p=5.74E-03) in comparison to homozygous absence (Figure S1d). PP genotype individuals also displayed a significant reduction of 32% (p=1.85E-05) in FCGBP gene expression in comparison to the PA genotype (Figure S1d). Similar to SVA_73 gene expression analysis, individuals heterozygous for SVA_72 indicated a 1.2-fold (p=4.23E-01) increase in FCGBP expression, however statistical significance was not achieved (Figure S1d).
As a recent genome-wide association study (GWAS) reported the association of the HLA locus with ALS risk [31,32], we investigated the influence that SVA RIPs had on cis regulated HLA genes (Figures S2 and S3). This analysis demonstrated that the top ten hits for greatest effect of SVA RIPs regarding upregulation was for HLA genes. We discovered a common gene expression pattern across all HLA genes analysed, demonstrating that individuals homozygous present for SVA RIPs (SVA_24, SVA_25, SVA_27, SVA_88) significantly upregulate HLA genes in comparison to homozygous absent individuals (p<0.01) (Figures S2 and S3).
2.4. SVA RIPs display tissue-specific modulation if gene expression
To investigate tissue-specific influences of reference SVA polymorphisms, we conducted eQTL analysis on the CNS tissue types spinal cord, cerebellum, motor cortex and frontal cortex individually (Table 2 and Table 3). The tissues temporal cortex, occipital cortex and hippocampus were excluded in the analysis due to the low n numbers. The results tables display the eQTL analysis from each individual tissue to allow determination of the top 40 hits for greatest effect size on gene upregulation (positive beta values) (Table 2) and downregulation (negative beta values) (Table 3). From this analysis, we established that a large proportion of the gene regulation effects were present within spinal cord tissue, displaying 27/40 and 26/40 tissue-specific hits for both gene upregulation and downregulation, respectively (Table 2 and Table 3). Furthermore, our analysis revealed that five SVAs (SVA_55, SVA_15, SVA_37, SVA_85, and SVA_4) were responsible for the 40 most positive beta values and eight SVAs (SVA_90, SVA_5, SVA_87, SVA_93, SVA_30, SVA_70, SVA_84, SVA_91, SVA_16) were responsible for the 40 most negative beta values, all effects of which were in the trans position (Table 2 and Table 3). This data demonstrates the capability of SVAs to modulate the expression of multiple gene targets.
Moreover, the MBP gene located on chromosome 18 and the PLP1 gene located on the X chromosome, were the most significantly upregulated and downregulated genes simultaneously (Table 2 and Table 3). Here, the gene expression of both genes was modulated in spinal cord tissue (Table 2 and Table 3). In addition, the genes MTURN and MOBP, and mitochondrial genes (MTCO1P12, MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND5, MT-CO2, MT-CO3, and MT-CYB) appear multiple times within both gene upregulation and downregulation hits (Table 2 and Table 3). Upon analysis of the whole tissue-specific dataset, the genes MBP, PLP1, MTURN, and MOBP were only regulated within spinal cord tissue, whilst the expression of mitochondrial genes was modulated within spinal cord, cerebellum, and motor cortex while MTCO1P12 gene expression was regulated in all four tissue types (Table 2 and Table 3). Thus, this illustrates that SVAs possess the capacity to influence tissue-specific gene expression.
3. Discussion
In this study, we evaluated the role of reference SVAs polymorphic for their presence in the human genome, to modulate gene expression within CNS tissues of ALS patients and healthy controls. Analysis of the NYGC ALS consortium dataset demonstrated that SVA RIPs significantly regulate gene expression genome-wide and in a tissue-specific manner. This study continues to expand on our previous findings, demonstrating the capability of SVAs to differentially regulate gene expression [24,33]. We have previously illustrated the capacity of SVAs to influence gene expression within Parkinson’s disease, highlighting the role of SVA_67 to modulate genes at the MAPT locus within the PPMI cohort and a CRISPR deletion model [22,26,30]. This analysis further validates our previous research, illustrating the functional capacity of SVA_67 within neurodegenerative disease and potentially expanding the importance of SVA_67 and the MAPT locus to ALS (Figure 5 and Figure 6). Therefore, this study not only emphasizes the correlation between SVA presence or absence and differential gene expression but also the involvement of SVAs within disease pathology.
Using WGS data from the ALS consortium for matrix eQTL analysis, we identified that polymorphic SVA RIPs possess the ability to modify expression of multiple target genes, but also many SVAs can regulate gene expression of a single target. Of these SVA RIPs, a greater proportion of trans regulatory effects were displayed in comparison to cis. Similarly, eQTL studies by both Wang et al. (2017) and Koks et al. (2021) analysing RNA-seq data from the 1000 genome project and the PPMI cohort, respectively, identified the impact of TEs on gene expression. In line with our analysis, these studies highlighted that several TEs simultaneously modulate gene expression of a single gene, an individual TE can regulate the expression of multiple genes, and that a greater number of TEs analysed were in the trans position [33,34]. A potential mechanism for trans-acting eQTLs could be through the binding of CCCTC-binding factor (CTCF) to SVAs. Through the cooperation with protein complex cohesion, CTCF plays a key role in three-dimensional chromatin regulation and chromatin looping, bringing promoters and regulatory elements within close proximity to activate or repress gene expression [35,36]. An additional mechanism could be through indirect transcription factor (TF) mediated associations. This suggests that SVAs influencing the expression of TF could indirectly regulate one or multiple TF gene targets, ultimately activating or repressing TF target gene expression [34].
Upon the examination of beta values from the eQTL analysis, we determined that within the combined analysis of CNS tissues SVA_55 demonstrated the greatest effect on myelin basic protein (MBP) gene upregulation (Figure 4). Following further analysis, we identified that through individual tissue analysis the top hits for gene upregulation and downregulation, were SVAs influencing MBP gene expression (Table 2 and Table 3). However, MBP expression was only significantly modulated within spinal cord tissue. MBP is a key protein involved in the myelination process, whereby myelin sheaths are formed around CNS axons by oligodendrocytes [37,38]. As oligodendrocyte loss and myelin dysfunction has recently been emphasized in neurodegenerative disease, including ALS, it is essential to investigate this relationship between SVAs and MBP differential expression [39,40]. Lorente Pons et al. demonstrated the potential significance of MBP in ALS through post-mortem analysis of both sporadic ALS and C9orf72-related ALS cases, identifying a significant reduction in MBP protein abundance when normalised to proteolipid protein (PLP) in the spinal cord corticospinal tracts in ALS cases in comparison to controls [41]. As MBP mRNA is transported to the myelin compartment by the RNA transport granule and PLP is transported as a protein, this suggests that the reduction in MBP could be due to impaired mRNA transport [41]. Our data suggests that certain SVAs (SVA_90 and SVA_5) act to downregulate MBP gene expression, therefore SVAs could play a role in this mechanism leading to the reduction of MBP protein levels observed in ALS patients compared to controls.
Our analysis also demonstrated that SVAs can activate (SVA_37) and repress (SVA_87 and SVA_93) PLP1, a form of PLP. PLP1 has previously been implicated in Pelizaeus-Merzbacher disease (PMD), an X-linked neurodegenerative disease whereby mutations within this gene inhibit CNS myelination [42,43]. In addition, we demonstrate that the myelin-associated oligodendrocyte basic protein (MOBP) gene, the locus of which has been highlighted for ALS risk, is again potentially regulated by multiple SVAs (SVA_5, 15, 37, 55, 84, 85, 87, 91 and 93) only within the spinal cord [31]. Hence, SVA regulation could be a potential mechanism involved in ALS risk at the MOBP locus. Furthermore, our tissue specific analysis displayed that multiple mitochondrial genes (MT-ND1,2,3,4,5, MT-CYB, MT-CO2,3 and pseudogene MTCO1P12) were largely activated and repressed within all four tissues analysed. SVA_30 and SVA_70 displayed the greatest regulatory effects on mitochondrial genes modulating five and four gene targets, respectively. As mitochondrial dysfunction is known to be implicated in other neurodegenerative diseases including PD and Alzheimer’s disease, as well as ALS, SVA regulation resulting in differential expression could be an underlying mechanism involved in disease pathology [44,45].
Upon analysis of CNS tissues from both ALS individuals and healthy controls, seven of the top ten cis-acting reference SVA RIPs imposed the greatest effects on the upregulation of genes at the human leukocyte antigen (HLA) locus (Figure 6A). Four SVAs are responsible for the effects on HLA gene expression, whereby SVA_24 influences the expression of one gene (HLA-A), SVA_25 two genes (HLA-B and HLA-C), SVA_27 two genes (HLA-DRB1 and HLA-DRB5) and SVA_88 one gene (HLA-DQB1). HLA also referred to as the major histocompatibility complex (MHC), acts to regulate both innate and adaptive immunity involved in the human immune response [46]. Since, the involvement of the immune response in neurological disease has been recognised the HLA locus has been highlighted as a region of importance in numerous neurodegenerative disease, including ALS [46]. Various studies have investigated the significance and mechanism of HLA in ALS, demonstrating increased frequencies of HLA-A, HLA-B and HLA-C alleles in ALS cases compared to controls [47,48,49,50]. A recent large-scale GWAS conducted by Van Rheenen et al. has identified the HLA region as a locus significantly associated with ALS, further highlighting the importance of this locus [31]. Previous studies within our group have highlighted the capability of SVAs to modulate HLA gene expression; analysis of whole genome sequencing and transcriptomic data obtained from the whole blood of individuals within the PPMI cohort discovered that SVA_24, SVA_25 and SVA_27 modulate the expression of HLA-A, HLA-B and HLA-C, and HLA-DRB1 and HLA-DRB5, respectively [33,51]. This suggests that the modulation of HLA genes by SVAs could be a common mechanism within neurodegenerative disease.
In conclusion, we show that SVAs demonstrate a significant impact on the expression of individual or numerous genes including those previously associated with neurodegenerative diseases, such as ALS. Ultimately, the ability of SVAs to act as a regulatory domain could highlight the importance of TEs in the missing heritability of neurodegenerative disease. However, due to limitations such as low n numbers for some SVA genotypes and CNS tissue types (occipital cortex, temporal cortex, and hippocampus) further research investigating the involvement of TEs in the pathogenesis of neurodegenerative disease, specifically ALS is crucial. In addition, due to low n numbers in the ALS and control groups, individual group analysis was not possible. Therefore, future experiments investigating the influence of SVAs in ALS patient and control groups individually is required. Furthermore, although this analysis continues to demonstrate the potential role of SVAs in neurodegenerative disease further experiments such as CRISPR are essential to validate SVA-specific influences. For example, we have previously shown that SVA_67 deletion in a CRISPR model resulted in a significant increase in MAPT and LRRC37A gene expression [30].
4. Materials and Methods
4.1. Genotyping reference SVAs polymorphic for their presence/absence and disease association from whole genome sequencing data from ALS Consortium dataset
Whole genome sequencing (WGS) data from the New York Genome Center (NYGC) as part of the ALS consortium dataset, were obtained in cram file format and aligned to hg38. The ALS consortium dataset contains data from individuals diagnosed with ALS spectrum MND, other neurological disorder (Alzheimer’s, Parkinson’s disease etc), other MND and ALS with other neurological disorders, along with non-neurological controls (healthy controls). The structural variant caller Delly2 (https://github.com/dellytools/delly), with default settings, was used to genotype the ALS consortium cohort (4403 individuals) for previously identified reference SVA sites, as outlined in [52]. In total 92 reference SVAs polymorphic for presence/absence [52] were used for matrix eQTL and differential gene expression analysis.
4.2. RNA-seq differential gene expression analysis
To elucidate the influence of SVA RIP genotypes on differential gene expression, RNA-seq data from CNS tissues obtained from the NYGC ALS consortium were analysed (https://www.nygenome.org/als-consortium/). For this analysis both ALS cases and healthy controls were combined. The Salmon quantification tool was used to quantify gene-specific expression levels from previously obtained FASTQ files (https://salmon.readthedocs.io/en/latest/). Salmon-generated quant files were imported using the tximport function in R and raw counts were extracted using the DESeqDataSetFromTximport. The DESeq2 R package was implemented to normalise the raw counts through the median-of-ratios method. DESeq2 was also used to detect statistically significant differential gene expression across SVA genotype groups (AA, PA, PP), whereby P represents SVA presence and A represents SVA absence. Results of the analysis were visualised using the ggplot2 package in R. The Kruskal-Wallis multiple comparison test (non-parametric) was applied to determine statistical significance between genotypes. The Wilcoxon test was used for pairwise comparison to assess significant differences between genotypes and obtained p-values were corrected for multiple comparisons using false discovery rate (FDR).
4.3. eQTL (expression quantitative trait loci) analysis
Matrix eQTL was applied to evaluate the genetic loci (SVA elements) regulating expression of nearby (cis) and distant (trans, >1 Mbp away) genes, using the matrixEQTL package in R. For this analysis, the additive linear model was applied with normalised gene expression levels, chromosomal positions of SVAs (hg38) and covariates (sex and age). Effect size estimates were reported as beta values from matrix eQTL. Obtained p-values were corrected for multiple testing and the threshold was set to 0.05 for FDR corrected p-values.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Boxplots of cis-acting SVA RIPs displaying the top two hits for largest effect size on both gene upregulation and downregulation.; Figure S2: Boxplots of cis-acting SVA RIPs displaying four hits for the greatest effect size on gene upregulation. Figure S3: Boxplots of cis-acting SVA RIPs displaying three hits for the greatest effect size on gene upregulation.
Author Contributions
Conceptualization, L.S.H., A.F. and S.K.; methodology, L.S.H., A.F., A.L.P., and S.K; formal analysis, L.S.H., A.F. and S.K.; writing—original draft preparation, L.S.H.; writing—review and editing, L.S.H., A.F., A.L.P., V.J.B., J.P.Q., and S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Andrzej Wlodarski Memorial Research Fund (A.F. and J.P.Q.), Andrzej Wlodarski Memorial Research PhD scholarship (A.F.), Multiple Sclerosis Western Australia and Perron Institute for Neurological and Translational Science (A.L.P. and S.K.), Darby Rimmer Foundation (V.J.B. and J.P.Q.), and MNDA [ref Quinn/Apr20/875-791] (V.J.B., J.P.Q., A.L.P. and S.K.).
Informed Consent Statement
The WGS and RNA sequencing data from the ALS consortium were provided in a de-identified manner with consent obtained by the participating consortia.
Data Availability Statement
The sequencing (RNA and WGS) data analysed in this study from the ALS consortium were obtained upon application to the New York Genome Center, and data requests can be made by completing a genetic data request form at ALSData@nygenome.org. Additional data from this study will be made available upon reasonable request.
Acknowledgments
We would like to acknowledge the ALS Consortium/Target ALS Human Postmortem Tissue Core, New York Genome Center for Genomics of Neurodegenerative Disease, Amyotrophic Lateral Sclerosis Association and TOW Foundation for the RNA-sequencing sequencing data used in this publication. This work was supported by resources provided by the Pawsey Supercomputing Centre with funding from the Australian Government and the Government of Western Australia.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Zarei, S.; Carr, K.; Reiley, L.; Diaz, K.; Guerra, O.; Altamirano, P.; Pagani, W.; Lodin, D.; Orozco, G.; Chinea, A. A Comprehensive Review of Amyotrophic Lateral Sclerosis. Surg Neurol Int 2015, 6, 171. [Google Scholar] [CrossRef] [PubMed]
- Grad, L.I.; Rouleau, G.A.; Ravits, J.; Cashman, N.R. Clinical Spectrum of Amyotrophic Lateral Sclerosis (ALS). Cold Spring Harb Perspect Med 2017, 7, a024117. [Google Scholar] [CrossRef]
- Rowland, L.P.; Shneider, N.A. Amyotrophic Lateral Sclerosis. N Engl J Med 2001, 344, 1688–1700. [Google Scholar] [CrossRef]
- Masrori, P.; Van Damme, P. Amyotrophic Lateral Sclerosis: A Clinical Review. Euro J of Neurology 2020, 27, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. ALS Genetics, Mechanisms, and Therapeutics: Where Are We Now? Front. Neurosci. 2019, 13, 1310. [Google Scholar] [CrossRef]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N Engl J Med 2017, 377, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Van Es, M.A.; Hardiman, O.; Chio, A.; Al-Chalabi, A.; Pasterkamp, R.J.; Veldink, J.H.; Van Den Berg, L.H. Amyotrophic Lateral Sclerosis. The Lancet 2017, 390, 2084–2098. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.L.; Schumann, G.G.; Breen, G.; Bubb, V.J.; Al-Chalabi, A.; Quinn, J.P. Retrotransposons in the Development and Progression of Amyotrophic Lateral Sclerosis. J Neurol Neurosurg Psychiatry 2019, 90, 284–293. [Google Scholar] [CrossRef]
- Gregory, J.M.; Elliott, E.; McDade, K.; Bak, T.; Pal, S.; Chandran, S.; Abrahams, S.; Smith, C. Neuronal Clusterin Expression Is Associated with Cognitive Protection in Amyotrophic Lateral Sclerosis. Neuropathology Appl Neurobio 2020, 46, 255–263. [Google Scholar] [CrossRef]
- Suzuki, N.; Nishiyama, A.; Warita, H.; Aoki, M. Genetics of Amyotrophic Lateral Sclerosis: Seeking Therapeutic Targets in the Era of Gene Therapy. J Hum Genet 2023, 68, 131–152. [Google Scholar] [CrossRef]
- Zou, Z.-Y.; Zhou, Z.-R.; Che, C.-H.; Liu, C.-Y.; He, R.-L.; Huang, H.-P. Genetic Epidemiology of Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. J Neurol Neurosurg Psychiatry 2017, 88, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Al-Chalabi, A.; Fang, F.; Hanby, M.F.; Leigh, P.N.; Shaw, C.E.; Ye, W.; Rijsdijk, F. An Estimate of Amyotrophic Lateral Sclerosis Heritability Using Twin Data. Journal of Neurology, Neurosurgery & Psychiatry 2010, 81, 1324–1326. [Google Scholar] [CrossRef]
- Theunissen, F.; Flynn, L.L.; Anderton, R.S.; Mastaglia, F.; Pytte, J.; Jiang, L.; Hodgetts, S.; Burns, D.K.; Saunders, A.; Fletcher, S.; et al. Structural Variants May Be a Source of Missing Heritability in sALS. Front. Neurosci. 2020, 14, 47. [Google Scholar] [CrossRef] [PubMed]
- Roses, A.D.; Akkari, P.A.; Chiba-Falek, O.; Lutz, M.W.; Gottschalk, W.K.; Saunders, A.M.; Saul, B.; Sundseth, S.; Burns, D. Structural Variants Can Be More Informative for Disease Diagnostics, Prognostics and Translation than Current SNP Mapping and Exon Sequencing. Expert Opinion on Drug Metabolism & Toxicology 2016, 12, 135–147. [Google Scholar] [CrossRef]
- GTEx Consortium; Chiang, C. ; Scott, A.J.; Davis, J.R.; Tsang, E.K.; Li, X.; Kim, Y.; Hadzic, T.; Damani, F.N.; Ganel, L.; et al. The Impact of Structural Variation on Human Gene Expression. Nat Genet 2017, 49, 692–699. [Google Scholar] [CrossRef]
- Liscic, R.M. Als and Ftd: Insights into the Disease Mechanisms and Therapeutic Targets. European Journal of Pharmacology 2017, 817, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Elbarbary, R.A.; Lucas, B.A.; Maquat, L.E. Retrotransposons as Regulators of Gene Expression. Science 2016, 351, aac7247. [Google Scholar] [CrossRef]
- Ayarpadikannan, S.; Kim, H.-S. The Impact of Transposable Elements in Genome Evolution and Genetic Instability and Their Implications in Various Diseases. Genomics Inform 2014, 12, 98. [Google Scholar] [CrossRef]
- Gianfrancesco, O.; Geary, B.; Savage, A.L.; Billingsley, K.J.; Bubb, V.J.; Quinn, J.P. The Role of SINE-VNTR-Alu (SVA) Retrotransposons in Shaping the Human Genome. IJMS 2019, 20, 5977. [Google Scholar] [CrossRef]
- Prudencio, M.; Gonzales, P.K.; Cook, C.N.; Gendron, T.F.; Daughrity, L.M.; Song, Y.; Ebbert, M.T.W.; Van Blitterswijk, M.; Zhang, Y.-J.; Jansen-West, K.; et al. Repetitive Element Transcripts Are Elevated in the Brain of C9orf72 ALS/FTLD Patients. Human Molecular Genetics 2017, 26, 3421–3431. [Google Scholar] [CrossRef]
- Hancks, D.C.; Kazazian, H.H. Active Human Retrotransposons: Variation and Disease. Current Opinion in Genetics & Development 2012, 22, 191–203. [Google Scholar] [CrossRef]
- Fröhlich, A.; Pfaff, A.L.; Bubb, V.J.; Koks, S.; Quinn, J.P. Characterisation of the Function of a SINE-VNTR-Alu Retrotransposon to Modulate Isoform Expression at the MAPT Locus. Front. Mol. Neurosci. 2022, 15, 815695. [Google Scholar] [CrossRef]
- Wang, H.; Xing, J.; Grover, D.; Hedges, D.J.; Han, K.; Walker, J.A.; Batzer, M.A. SVA Elements: A Hominid-Specific Retroposon Family. Journal of Molecular Biology 2005, 354, 994–1007. [Google Scholar] [CrossRef]
- Savage, A.L.; Bubb, V.J.; Breen, G.; Quinn, J.P. Characterisation of the Potential Function of SVA Retrotransposons to Modulate Gene Expression Patterns. BMC Evol Biol 2013, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.P.; Bubb, V.J. SVA Retrotransposons as Modulators of Gene Expression. Mobile Genetic Elements 2014, 4, e32102. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, A.L.; Bubb, V.J.; Quinn, J.P.; Koks, S. Reference SVA Insertion Polymorphisms Are Associated with Parkinson’s Disease Progression and Differential Gene Expression. npj Parkinsons Dis. 2021, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Verpillat, P.; Camuzat, A.; Hannequin, D.; Thomas-Anterion, C.; Puel, M.; Belliard, S.; Dubois, B.; Didic, M.; Michel, B.-F.; Lacomblez, L.; et al. Association Between the Extended Tau Haplotype and Frontotemporal Dementia. Arch Neurol 2002, 59, 935. [Google Scholar] [CrossRef] [PubMed]
- Wider, C.; Vilariño-Güell, C.; Jasinska-Myga, B.; Heckman, M.G.; Soto-Ortolaza, A.I.; Cobb, S.A.; Aasly, J.O.; Gibson, J.M.; Lynch, T.; Uitti, R.J.; et al. Association of the MAPT Locus with Parkinson’s Disease. Euro J of Neurology 2010, 17, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Juan, P.; Moreno, S.; De Rojas, I.; Hernández, I.; Valero, S.; Alegret, M.; Montrreal, L.; García González, P.; Lage, C.; López-García, S.; et al. The MAPT H1 Haplotype Is a Risk Factor for Alzheimer’s Disease in APOE Ε4 Non-Carriers. Front. Aging Neurosci. 2019, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, A.; Hughes, L.S.; Middlehurst, B.; Pfaff, A.L.; Bubb, V.J.; Koks, S.; Quinn, J.P. CRISPR Deletion of a SINE-VNTR-Alu (SVA_67) Retrotransposon Demonstrates Its Ability to Differentially Modulate Gene Expression at the MAPT Locus. Front. Neurol. 2023, 14, 1273036. [Google Scholar] [CrossRef]
- Van Rheenen, W.; Van Der Spek, R.A.A.; Bakker, M.K.; Van Vugt, J.J.F.A.; Hop, P.J.; Zwamborn, R.A.J.; De Klein, N.; Westra, H.-J.; Bakker, O.B.; Deelen, P.; et al. Common and Rare Variant Association Analyses in Amyotrophic Lateral Sclerosis Identify 15 Risk Loci with Distinct Genetic Architectures and Neuron-Specific Biology. Nat Genet 2021, 53, 1636–1648. [Google Scholar] [CrossRef]
- Nona, R.J.; Greer, J.M.; Henderson, R.D.; McCombe, P.A. HLA and Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. Amyotrophic Lateral Sclerosis and Frontotemporal Degeneration 2023, 24, 24–32. [Google Scholar] [CrossRef]
- Koks, S.; Pfaff, A.L.; Bubb, V.J.; Quinn, J.P. Expression Quantitative Trait Loci (eQTLs) Associated with Retrotransposons Demonstrate Their Modulatory Effect on the Transcriptome. IJMS 2021, 22, 6319. [Google Scholar] [CrossRef]
- Wang, L.; Rishishwar, L.; Mariño-Ramírez, L.; Jordan, I.K. Human Population-Specific Gene Expression and Transcriptional Network Modification with Polymorphic Transposable Elements. Nucleic Acids Res 2017, gkw1286. [Google Scholar] [CrossRef]
- Pugacheva, E.M.; Teplyakov, E.; Wu, Q.; Li, J.; Chen, C.; Meng, C.; Liu, J.; Robinson, S.; Loukinov, D.; Boukaba, A.; et al. The Cancer-Associated CTCFL/BORIS Protein Targets Multiple Classes of Genomic Repeats, with a Distinct Binding and Functional Preference for Humanoid-Specific SVA Transposable Elements. Epigenetics & Chromatin 2016, 9, 35. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, J.; Cao, C. CTCF and Its Partners: Shaper of 3D Genome during Development. Genes 2022, 13, 1383. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Snaidero, N.; Pähler, G.; Frey, S.; Sánchez, P.; Zweckstetter, M.; Janshoff, A.; Schneider, A.; Weil, M.-T.; Schaap, I.A.T.; et al. Myelin Membrane Assembly Is Driven by a Phase Transition of Myelin Basic Proteins Into a Cohesive Protein Meshwork. PLoS Biol 2013, 11, e1001577. [Google Scholar] [CrossRef]
- Kang, S.H.; Li, Y.; Fukaya, M.; Lorenzini, I.; Cleveland, D.W.; Ostrow, L.W.; Rothstein, J.D.; Bergles, D.E. Degeneration and Impaired Regeneration of Gray Matter Oligodendrocytes in Amyotrophic Lateral Sclerosis. Nat Neurosci 2013, 16, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Boccazzi, M.; Fumagalli, M. Oligodendrocyte Dysfunction in Amyotrophic Lateral Sclerosis: Mechanisms and Therapeutic Perspectives. Cells 2021, 10, 565. [Google Scholar] [CrossRef] [PubMed]
- Lubetzki, C.; Zalc, B.; Williams, A.; Stadelmann, C.; Stankoff, B. Remyelination in Multiple Sclerosis: From Basic Science to Clinical Translation. The Lancet Neurology 2020, 19, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Lorente Pons, A.; Higginbottom, A.; Cooper-Knock, J.; Alrafiah, A.; Alofi, E.; Kirby, J.; Shaw, P.J.; Wood, J.D.; Highley, J.R. Oligodendrocyte Pathology Exceeds Axonal Pathology in White Matter in Human Amyotrophic Lateral Sclerosis. The Journal of Pathology 2020, 251, 262–271. [Google Scholar] [CrossRef]
- Hübner, C.A.; Orth, U.; Senning, A.; Steglich, C.; Kohlschütter, A.; Korinthenberg, R.; Gal, A. Seventeen Novel PLP1 Mutations in Patients with Pelizaeus-Merzbacher Disease: MUTATIONS IN BRIEF. Hum. Mutat. 2005, 25, 321–322. [Google Scholar] [CrossRef]
- Cloake, N.; Yan, J.; Aminian, A.; Pender, M.; Greer, J. PLP1 Mutations in Patients with Multiple Sclerosis: Identification of a New Mutation and Potential Pathogenicity of the Mutations. JCM 2018, 7, 342. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Rivera, Y.E.; Perez-Morales, J.; Santiago, Y.M.; Gonzalez, V.M.; Morales, L.; Cabrera-Rios, M.; Isaza, C.E. A Selection of Important Genes and Their Correlated Behavior in Alzheimer’s Disease. JAD 2018, 65, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, E.; Musich, P.R.; Lin, F. Mitochondrial Dysfunction in Neurodegenerative Diseases and the Potential Countermeasure. CNS Neurosci Ther 2019, 25, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Misra, M.K.; Damotte, V.; Hollenbach, J.A. The Immunogenetics of Neurological Disease. Immunology 2018, 153, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Antel, J.P.; Arnason, B.G.W.; Fuller, T.C.; Lehrich, J.R. Histocompatibility Typing in Amyotrophic Lateral Sclerosis. Archives of Neurology 1976, 33, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Behan, P.; Durward, W.; Dick, H. HISTOCOMPATIBILITY ANTIGENS ASSOCIATED WITH MOTOR-NEURONE DISEASE. The Lancet 1976, 308, 803. [Google Scholar] [CrossRef] [PubMed]
- Jokelainen, M.; Tiilikainen, A.; Lapinleimu, K. Polio Antibodies and HLA Antigens in Amyotrophic Lateral Sclerosis. Tissue Antigens 1977, 10, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Kott, E.; Livni, E.; Zamir, R.; Kuritzky, A. Cell-Mediated Immunity to Polio and HLA Antigens in Amyotrophic Lateral Sclerosis. Neurology 1979, 29, 1040–1040. [Google Scholar] [CrossRef]
- Kulski, J.K.; Pfaff, A.L.; Marney, L.D.; Fröhlich, A.; Bubb, V.J.; Quinn, J.P.; Koks, S. Regulation of Expression Quantitative Trait Loci by SVA Retrotransposons within the Major Histocompatibility Complex. Exp Biol Med (Maywood) 2023, 15353702231209411. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, A.L.; Bubb, V.J.; Quinn, J.P.; Koks, S. A Genome-Wide Screen for the Exonisation of Reference SINE-VNTR-Alus and Their Expression in CNS Tissues of Individuals with Amyotrophic Lateral Sclerosis. IJMS 2023, 24, 11548. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
General study overview, structure of full-length SVA element and both cis- and trans-acting mechanisms. (a) This study incorporated whole genome sequencing and transcriptomic data from the New York Genome Centre ALS Consortium cohort, to investigate the ability of SVA retrotransposon insertion polymorphisms (RIPs) to act as expression quantitative trait loci (eQTL) within central nervous system (CNS) tissues. (b) Schematic of SVA structure, displaying a full-length SVA element consisting of a 5’ CT rich hexamer repeat, Alu-like region, variable number tandem repeat (VNTR), SINE (short interspersed nuclear element)-R domain and a 3’ poly-A tail. (c) The mechanism by which SVAs implement a cis or trans regulatory effect. Cis-regulatory effects are defined as effects observed by elements (SVA RIPs) which act to modulate the expression of genes less than 1 Mb away from the element site, whilst trans regulatory effects are defined as effects observed by elements which act to modulate the expression of genes greater than 1 Mb away from the element site.
Figure 1.
General study overview, structure of full-length SVA element and both cis- and trans-acting mechanisms. (a) This study incorporated whole genome sequencing and transcriptomic data from the New York Genome Centre ALS Consortium cohort, to investigate the ability of SVA retrotransposon insertion polymorphisms (RIPs) to act as expression quantitative trait loci (eQTL) within central nervous system (CNS) tissues. (b) Schematic of SVA structure, displaying a full-length SVA element consisting of a 5’ CT rich hexamer repeat, Alu-like region, variable number tandem repeat (VNTR), SINE (short interspersed nuclear element)-R domain and a 3’ poly-A tail. (c) The mechanism by which SVAs implement a cis or trans regulatory effect. Cis-regulatory effects are defined as effects observed by elements (SVA RIPs) which act to modulate the expression of genes less than 1 Mb away from the element site, whilst trans regulatory effects are defined as effects observed by elements which act to modulate the expression of genes greater than 1 Mb away from the element site.
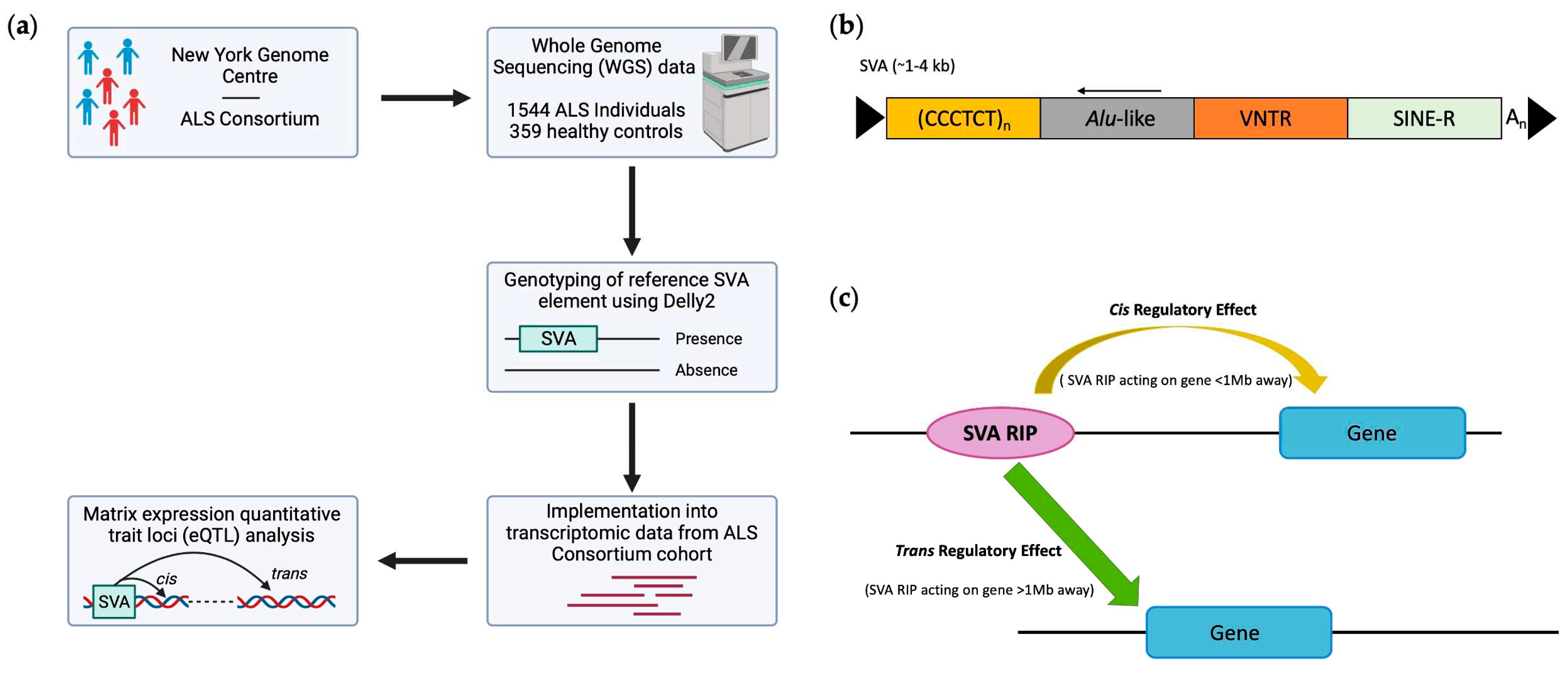
Figure 2.
Composition of tissues used in this study. For this study, CNS tissue data from healthy controls (CO) and ALS patients combined (n=1903) composed of spinal cord (n=710), motor cortex (n=440), frontal cortex (n=335), cerebellum (n=240), occipital cortex (n=57), temporal cortex (n=58) and hippocampus (n=63) were included for our analysis.
Figure 2.
Composition of tissues used in this study. For this study, CNS tissue data from healthy controls (CO) and ALS patients combined (n=1903) composed of spinal cord (n=710), motor cortex (n=440), frontal cortex (n=335), cerebellum (n=240), occipital cortex (n=57), temporal cortex (n=58) and hippocampus (n=63) were included for our analysis.
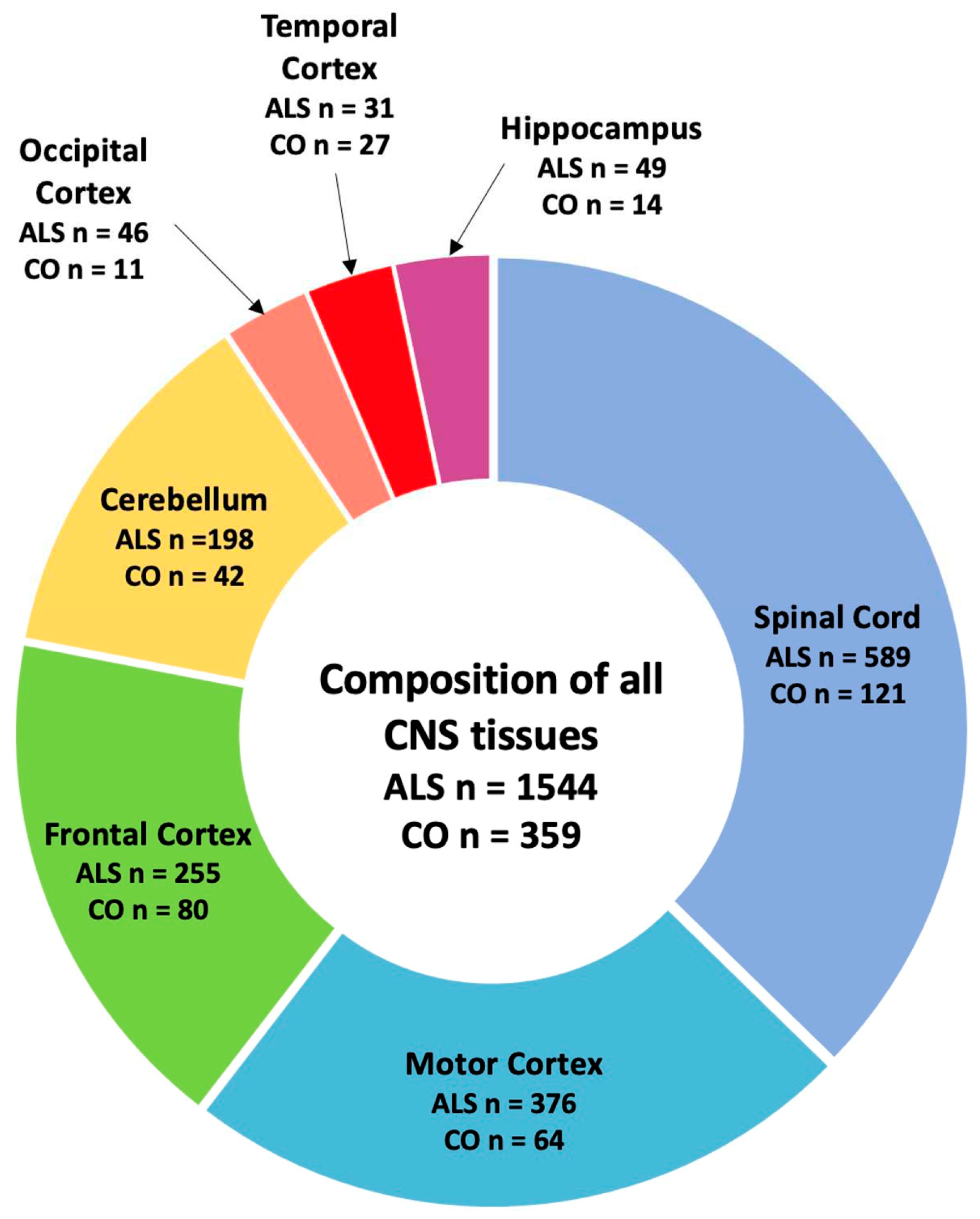
Figure 3.
Overview of the number of genomic loci affected by SVA polymorphism following matrix eQTL analysis of all CNS tissues. (a) Pie chart representing the composition of all significant differentially regulated genetic loci (n=14830), displaying the number of cis-regulatory (n=167) and trans-regulatory (n=14663) effects exhibited by SVAs within all CNS tissues. (b) Bar chart displaying reference SVAs and the number of genome wide gene targets. Each of the 92 analysed SVAs had a significant impact on multiple targets, with the lowest number of targets for one SVA being 6 (FDR p<0.05). Only SVAs affecting more than 200 targets are displayed (n=26). For this analysis data from ALS individuals and healthy controls were combined.
Figure 3.
Overview of the number of genomic loci affected by SVA polymorphism following matrix eQTL analysis of all CNS tissues. (a) Pie chart representing the composition of all significant differentially regulated genetic loci (n=14830), displaying the number of cis-regulatory (n=167) and trans-regulatory (n=14663) effects exhibited by SVAs within all CNS tissues. (b) Bar chart displaying reference SVAs and the number of genome wide gene targets. Each of the 92 analysed SVAs had a significant impact on multiple targets, with the lowest number of targets for one SVA being 6 (FDR p<0.05). Only SVAs affecting more than 200 targets are displayed (n=26). For this analysis data from ALS individuals and healthy controls were combined.
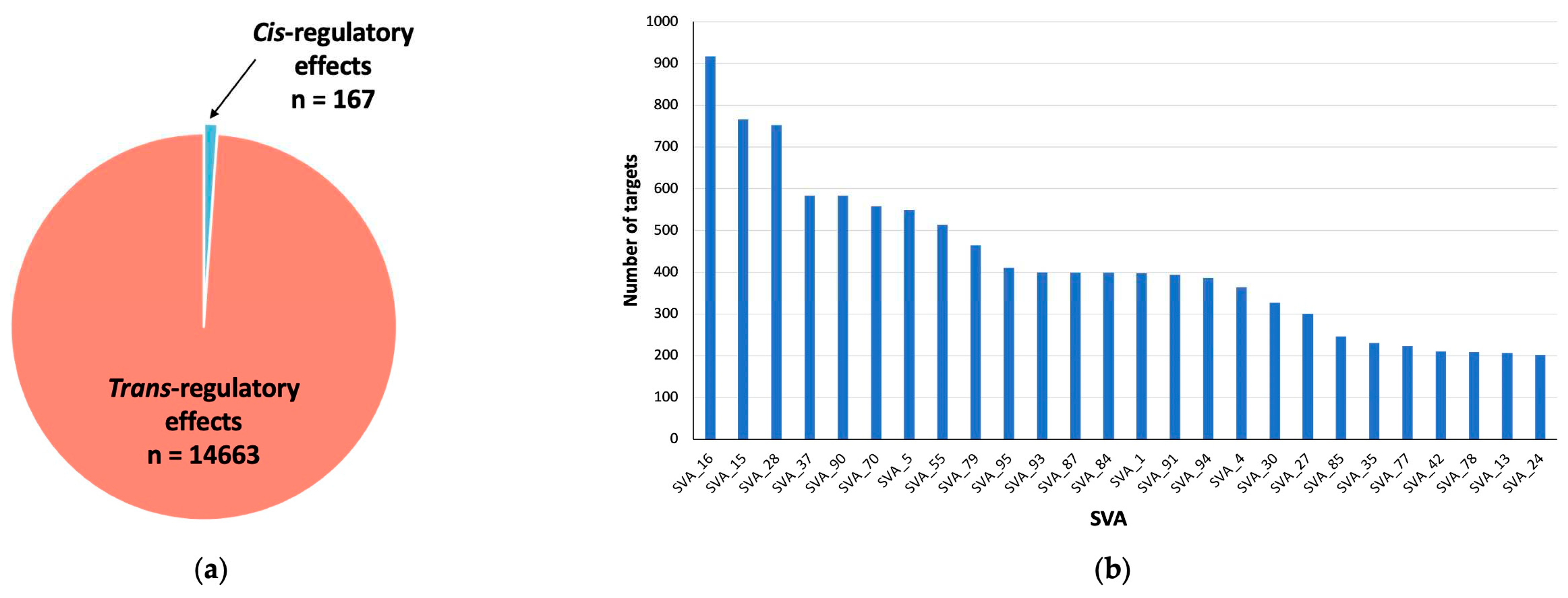
Figure 4.
Reference SVA RIP elements with the greatest effect size from matrix eQTL analysis of all CNS tissues. (a/b) Clustered bar chart showing the top ten reference SVA RIPs across all CNS tissues with the greatest effect size on gene upregulation (positive beta values) (a) and gene downregulation (negative beta values) (b) from eQTL analysis. SVA_55 demonstrated the greatest increase in activation of the MBP gene with a beta coefficient of 272,847, whilst SVA_70 demonstrated the greatest repressive effect on the MT-ND1 gene with a beta coefficient of -36441. (c) Boxplot of SVA_55 indicating MBP gene expression stratified by SVA_55 genotype. Genotypes PP (n=2), PA (n=20) and AA (n=1731). Significant differences in MBP gene expression were observed between the PP and PA group (p=0.0218) and PP and AA group (p=0.0218). Subjects with the PP genotype displayed a 9.5-fold and 19.2-fold increase in MBP gene expression in comparison to subjects with AA and PA genotypes respectively. (d) Boxplot of SVA_70 displaying MT-ND1 gene expression, stratified by SVA_70 genotype. Genotypes PP (n=1581), PA (n=159) and AA (n=13). A significant repression in MT-ND1 gene expression of 55% was observed between the PP and PA subject group (p=0.0098). No statistical significance was obtained for differences between the PP and PA subject groups. For both boxplots the significance of gene expression changes between groups was determined using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR p<0.05). PP, PA, and AA groups represent when there are two copies of the SVA present, one copy of the SVA present and the complete absence of the SVA, respectively. * p<0.05, ** p<0.01.
Figure 4.
Reference SVA RIP elements with the greatest effect size from matrix eQTL analysis of all CNS tissues. (a/b) Clustered bar chart showing the top ten reference SVA RIPs across all CNS tissues with the greatest effect size on gene upregulation (positive beta values) (a) and gene downregulation (negative beta values) (b) from eQTL analysis. SVA_55 demonstrated the greatest increase in activation of the MBP gene with a beta coefficient of 272,847, whilst SVA_70 demonstrated the greatest repressive effect on the MT-ND1 gene with a beta coefficient of -36441. (c) Boxplot of SVA_55 indicating MBP gene expression stratified by SVA_55 genotype. Genotypes PP (n=2), PA (n=20) and AA (n=1731). Significant differences in MBP gene expression were observed between the PP and PA group (p=0.0218) and PP and AA group (p=0.0218). Subjects with the PP genotype displayed a 9.5-fold and 19.2-fold increase in MBP gene expression in comparison to subjects with AA and PA genotypes respectively. (d) Boxplot of SVA_70 displaying MT-ND1 gene expression, stratified by SVA_70 genotype. Genotypes PP (n=1581), PA (n=159) and AA (n=13). A significant repression in MT-ND1 gene expression of 55% was observed between the PP and PA subject group (p=0.0098). No statistical significance was obtained for differences between the PP and PA subject groups. For both boxplots the significance of gene expression changes between groups was determined using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR p<0.05). PP, PA, and AA groups represent when there are two copies of the SVA present, one copy of the SVA present and the complete absence of the SVA, respectively. * p<0.05, ** p<0.01.
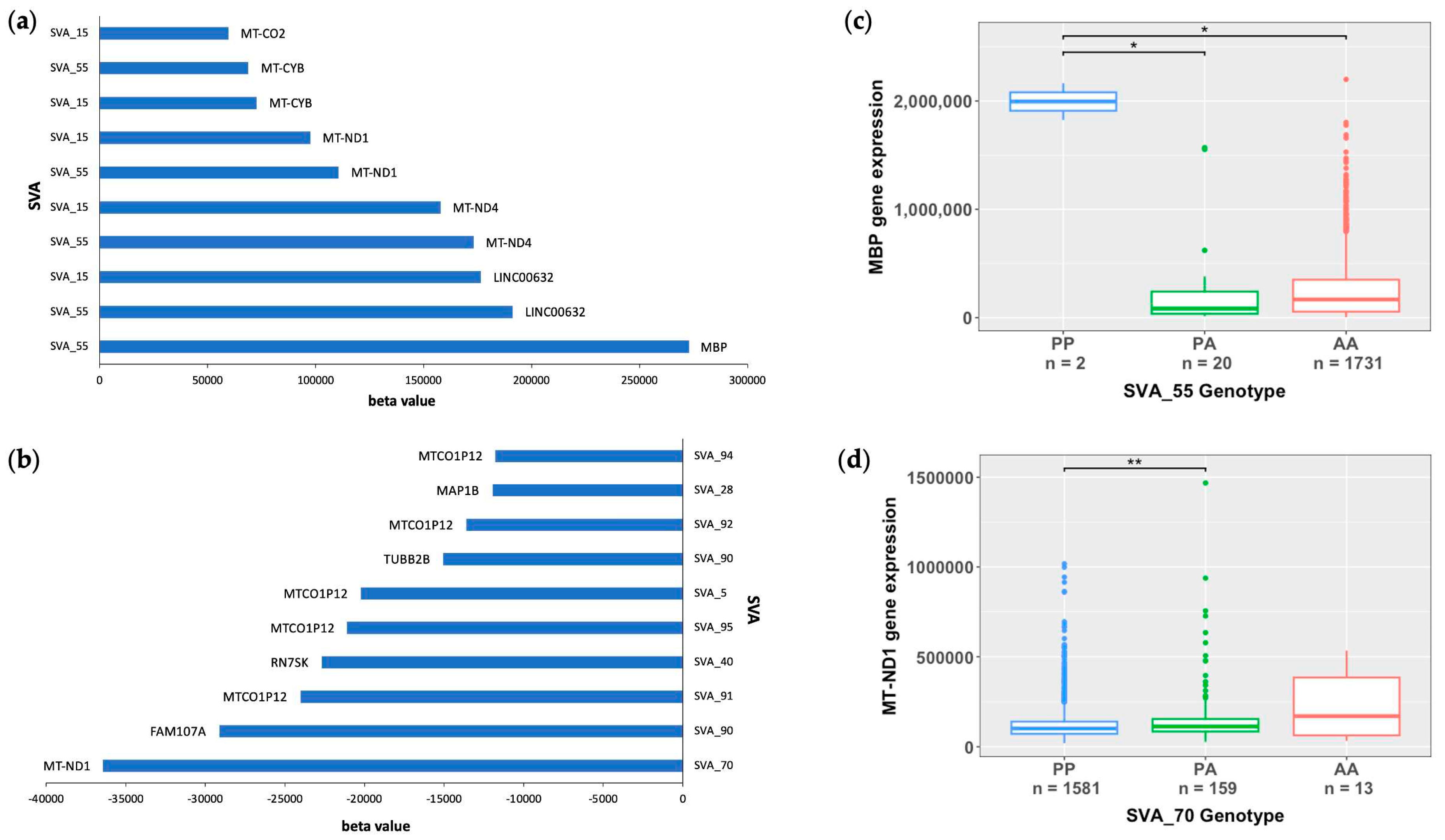
Figure 5.
Boxplots of the two of the most significant SVA_67 interactions with MAPK8IP1P2 and LRRC37A obtained from matrix eQTL analysis. Both significant effects are cis-regulatory effects. Datapoints from both ALS individuals and healthy controls across all CNS tissues were combined and the significance of gene expression changes between groups was determined using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR<0.05). (a) Boxplot showing the association of SVA_67 genotype with MAPK8IP1P2 gene expression. Significant differences were observed between all groups for MAPK8IP1P2 expression, namely PP and PA (p=7.33E-237), PP and AA (p=4.26E-55) and PA and AA (p=1.33E-07). (b) Boxplot showing the association of SVA_67 genotype with LRRC37A gene expression. Significant differences were observed between all groups for LRRC37A gene expression, PP and PA (p=1.95E-204), namely PP and AA (p=1.06E-39) and PA and AA (p=2.13E-12). Fold change in expression of 262-fold and 6-fold for MAPK8IP1P2 and LRRC37A, respectively, was observed for individuals with AA genotype in comparison to PP genotype. PP (n=1183), PA (n=500) and AA (n=63). *** p<0.001.
Figure 5.
Boxplots of the two of the most significant SVA_67 interactions with MAPK8IP1P2 and LRRC37A obtained from matrix eQTL analysis. Both significant effects are cis-regulatory effects. Datapoints from both ALS individuals and healthy controls across all CNS tissues were combined and the significance of gene expression changes between groups was determined using the Wilcoxon pairwise comparison with FDR adjusted p-values (FDR<0.05). (a) Boxplot showing the association of SVA_67 genotype with MAPK8IP1P2 gene expression. Significant differences were observed between all groups for MAPK8IP1P2 expression, namely PP and PA (p=7.33E-237), PP and AA (p=4.26E-55) and PA and AA (p=1.33E-07). (b) Boxplot showing the association of SVA_67 genotype with LRRC37A gene expression. Significant differences were observed between all groups for LRRC37A gene expression, PP and PA (p=1.95E-204), namely PP and AA (p=1.06E-39) and PA and AA (p=2.13E-12). Fold change in expression of 262-fold and 6-fold for MAPK8IP1P2 and LRRC37A, respectively, was observed for individuals with AA genotype in comparison to PP genotype. PP (n=1183), PA (n=500) and AA (n=63). *** p<0.001.
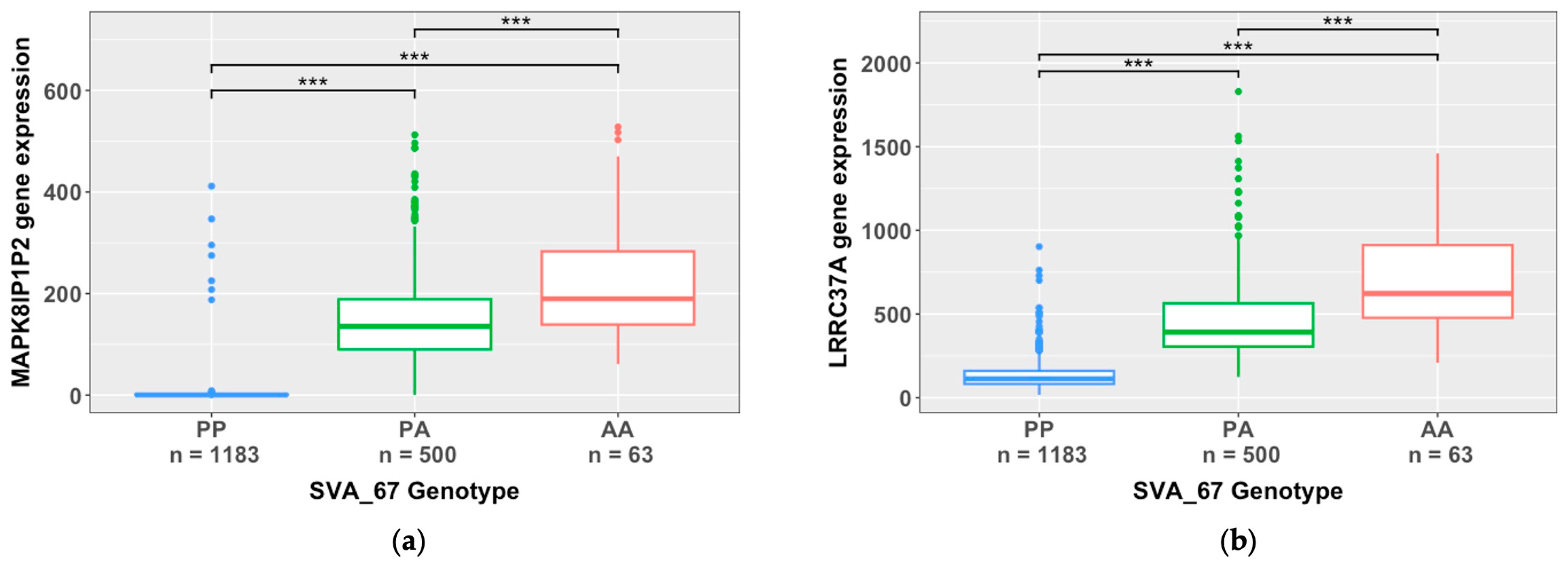
Figure 6.
Top ten cis-acting reference SVA elements with the greatest effects on gene upregulation and downregulation in CNS tissues. (a) Clustered bar chart displaying ten reference SVA RIPs with the greatest cis-regulatory effects on gene activation (most positive beta values) from matrix eQTL analysis (FDR p<0.05). SVA_67 is responsible for the two greatest increases in activation of the genes LRRC37A4P and MAPT displaying beta coefficients of 1384 and 1101, respectively. SVA_24 (HLA-A), SVA_25 (HLA-C and HLA-B), SVA_27 (HLA-DRB1, HLA-DRB5 and HLA-DQB1) and SVA_88 (HLA-DPA1) showed to be responsible for the large increases in activation of a series of HLA genes. (b) Clustered bar chart displaying ten reference SVA RIPs with the greatest cis-regulatory effects on gene downregulation (most negative beta values) from matrix eQTL analysis (FDR p<0.05). The two greatest repressive effects were on the FCGBP gene, regulated by both SVA_73 and SVA_72 demonstrating a beta coefficient of -2843 and -668, respectively. SVA_67 was responsible for four of these effects, by showing a downregulating effect, for the genes KANSL1, LRRC37A, LRRC37A2 and MAPK8IP1P2. This analysis combined ALS individuals and healthy controls datapoints combined.
Figure 6.
Top ten cis-acting reference SVA elements with the greatest effects on gene upregulation and downregulation in CNS tissues. (a) Clustered bar chart displaying ten reference SVA RIPs with the greatest cis-regulatory effects on gene activation (most positive beta values) from matrix eQTL analysis (FDR p<0.05). SVA_67 is responsible for the two greatest increases in activation of the genes LRRC37A4P and MAPT displaying beta coefficients of 1384 and 1101, respectively. SVA_24 (HLA-A), SVA_25 (HLA-C and HLA-B), SVA_27 (HLA-DRB1, HLA-DRB5 and HLA-DQB1) and SVA_88 (HLA-DPA1) showed to be responsible for the large increases in activation of a series of HLA genes. (b) Clustered bar chart displaying ten reference SVA RIPs with the greatest cis-regulatory effects on gene downregulation (most negative beta values) from matrix eQTL analysis (FDR p<0.05). The two greatest repressive effects were on the FCGBP gene, regulated by both SVA_73 and SVA_72 demonstrating a beta coefficient of -2843 and -668, respectively. SVA_67 was responsible for four of these effects, by showing a downregulating effect, for the genes KANSL1, LRRC37A, LRRC37A2 and MAPK8IP1P2. This analysis combined ALS individuals and healthy controls datapoints combined.
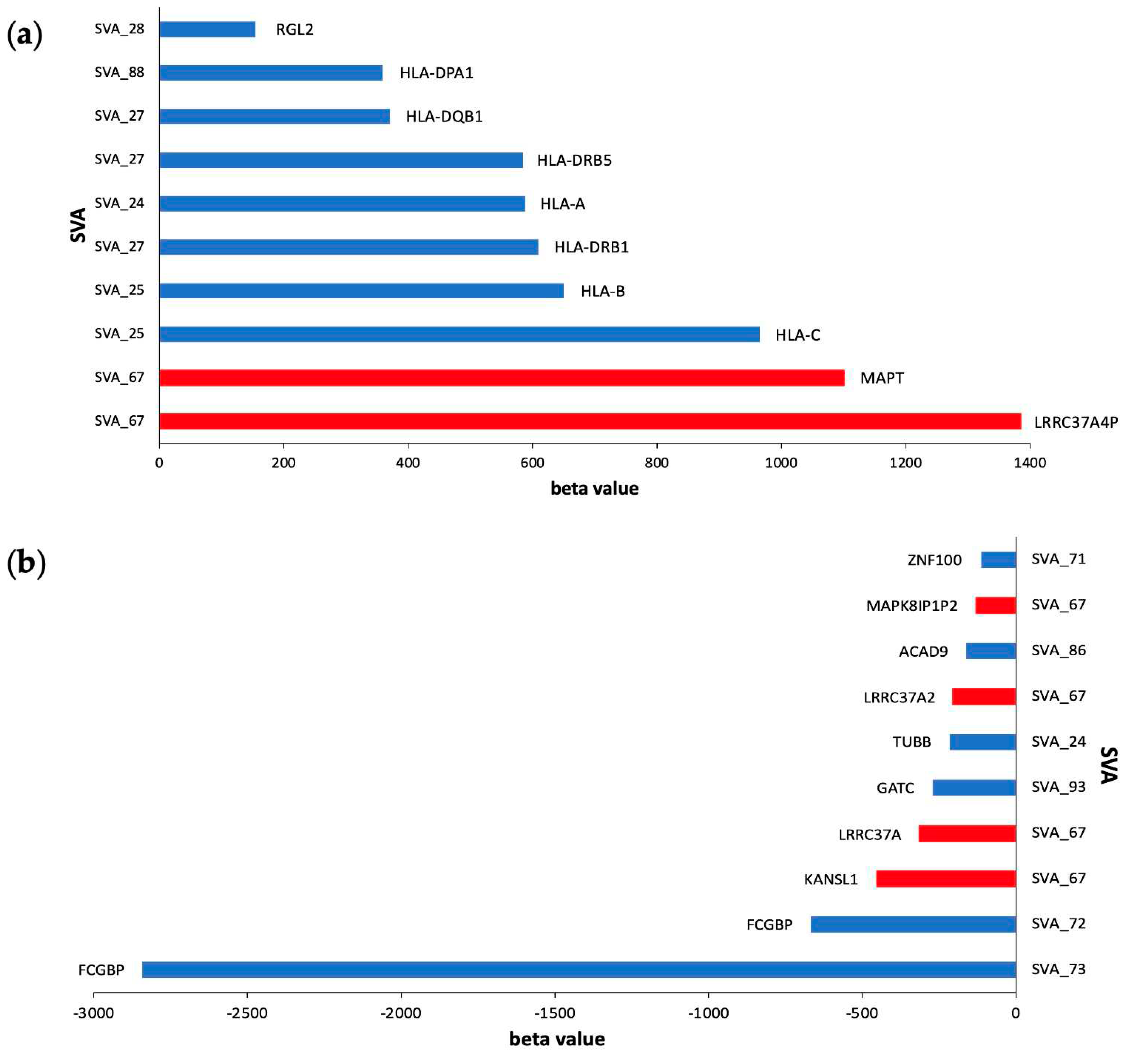

Table 1.
Top ten most significant reference SVA RIP effects from matrix eQTL analysis of all CNS tissues. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
Table 1.
Top ten most significant reference SVA RIP effects from matrix eQTL analysis of all CNS tissues. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
| SVA | beta value | False Discovery Rate (FDR) | Target gene | cis/trans effect |
|---|---|---|---|---|
| SVA_67 | -131.2 | 1.93E-303 | MAPK8IP1P2 | cis |
| SVA_67 | -5.2 | 1.93E-303 | ENSG00000285668.1 | cis |
| SVA_67 | -315.1 | 5.12E-299 | LRRC37A | cis |
| SVA_87 | -126.2 | 3.02E-224 | MTND4P24 | trans |
| SVA_93 | -126.2 | 4.22E-224 | MTND4P24 | trans |
| SVA_84 | -63.1 | 4.22E-224 | MTND4P24 | trans |
| SVA_58 | 4.1 | 3.89E-211 | LLPH-DT | cis |
| SVA_24 | 12.3 | 5.06E-201 | HLA-K | cis |
| SVA_15 | 59.6 | 3.13E-189 | MTND4P24 | trans |
| SVA_33 | 48.4 | 8.84E-188 | ZFAND2A-DT | cis |
Table 2.
Top 40 significant reference SVA RIPs with the greatest effects on gene upregulation (most positive beta values) from tissue specific matrix eQTL analysis. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
Table 2.
Top 40 significant reference SVA RIPs with the greatest effects on gene upregulation (most positive beta values) from tissue specific matrix eQTL analysis. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
| SVA | Gene ID | FDR p-value | Beta value | Gene | Chr | Cis/trans | Tissue |
|---|---|---|---|---|---|---|---|
| SVA_55 | ENSG00000197971.16 | 5.99E-10 | 638804.662 | MBP | 18 | trans | Spinal Cord |
| SVA_15 | ENSG00000197971.16 | 3.59E-07 | 620026.724 | MBP | 18 | trans | Spinal Cord |
| SVA_37 | ENSG00000197971.16 | 8.76E-04 | 585791.502 | MBP | 18 | trans | Spinal Cord |
| SVA_85 | ENSG00000197971.16 | 2.08E-03 | 560691.911 | MBP | 18 | trans | Spinal Cord |
| SVA_37 | ENSG00000123560.14 | 3.72E-12 | 168441.474 | PLP1 | X | trans | Spinal Cord |
| SVA_55 | ENSG00000198888.2 | 1.47E-05 | 153947.476 | MT-ND1 | MT | trans | Spinal Cord |
| SVA_55 | ENSG00000203930.12 | 5.97E-04 | 148616.177 | LINC00632 | X | trans | Motor Cortex |
| SVA_15 | ENSG00000198888.2 | 1.62E-03 | 142961.051 | MT-ND1 | MT | trans | Spinal Cord |
| SVA_55 | ENSG00000203930.12 | 1.25E-16 | 115977.491 | LINC00632 | X | trans | Spinal Cord |
| SVA_15 | ENSG00000203930.12 | 1.86E-08 | 96910.7666 | LINC00632 | X | trans | Spinal Cord |
| SVA_55 | ENSG00000180354.16 | 1.34E-22 | 90266.4311 | MTURN | 7 | trans | Spinal Cord |
| SVA_15 | ENSG00000123560.14 | 3.43E-03 | 80380.5623 | PLP1 | X | trans | Spinal Cord |
| SVA_85 | ENSG00000203930.12 | 4.76E-03 | 78457.5282 | LINC00632 | X | trans | Spinal Cord |
| SVA_15 | ENSG00000180354.16 | 1.86E-12 | 77839.9088 | MTURN | 7 | trans | Spinal Cord |
| SVA_55 | ENSG00000198712.1 | 3.04E-03 | 76318.8326 | MT-CO2 | MT | trans | Spinal Cord |
| SVA_85 | ENSG00000180354.16 | 1.35E-05 | 67900.9382 | MTURN | 7 | trans | Spinal Cord |
| SVA_85 | ENSG00000237973.1 | 4.10E-33 | 66702.2584 | MTCO1P12 | 1 | trans | Motor Cortex |
| SVA_37 | ENSG00000168314.18 | 3.50E-09 | 65555.4973 | MOBP | 3 | trans | Spinal Cord |
| SVA_85 | ENSG00000237973.1 | 1.01E-32 | 53997.5748 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_15 | ENSG00000168314.18 | 5.02E-09 | 53362.8451 | MOBP | 3 | trans | Spinal Cord |
| SVA_55 | ENSG00000237973.1 | 3.61E-22 | 52048.0183 | MTCO1P12 | 1 | trans | Motor Cortex |
| SVA_55 | ENSG00000168314.18 | 1.38E-09 | 49216.1425 | MOBP | 3 | trans | Spinal Cord |
| SVA_15 | ENSG00000237973.1 | 2.83E-26 | 48449.3666 | MTCO1P12 | 1 | trans | Motor Cortex |
| SVA_85 | ENSG00000168314.18 | 3.76E-04 | 47350.2242 | MOBP | 3 | trans | Spinal Cord |
| SVA_85 | ENSG00000237973.1 | 8.82E-42 | 39521.0532 | MTCO1P12 | 1 | trans | Spinal Cord |
| SVA_55 | ENSG00000064787.13 | 1.62E-30 | 36504.4209 | BCAS1 | 20 | trans | Spinal Cord |
| SVA_37 | ENSG00000091513.16 | 1.65E-06 | 35838.5557 | TF | 3 | trans | Spinal Cord |
| SVA_55 | ENSG00000237973.1 | 1.49E-59 | 33728.2747 | MTCO1P12 | 1 | trans | Spinal Cord |
| SVA_4 | ENSG00000237973.1 | 3.59E-17 | 32837.5825 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_37 | ENSG00000237973.1 | 1.52E-23 | 32783.3654 | MTCO1P12 | 1 | trans | Motor Cortex |
| SVA_37 | ENSG00000237973.1 | 1.72E-31 | 32728.512 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_15 | ENSG00000237973.1 | 8.40E-41 | 32193.3558 | MTCO1P12 | 1 | trans | Spinal Cord |
| SVA_37 | ENSG00000099194.6 | 2.82E-04 | 31333.7253 | SCD | 10 | trans | Spinal Cord |
| SVA_85 | ENSG00000064787.13 | 3.78E-10 | 30888.7058 | BCAS1 | 20 | trans | Spinal Cord |
| SVA_15 | ENSG00000237973.1 | 3.46E-14 | 30602.307 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_15 | ENSG00000064787.13 | 1.97E-15 | 30356.5865 | BCAS1 | 20 | trans | Spinal Cord |
| SVA_37 | ENSG00000237973.1 | 1.28E-06 | 30025.949 | MTCO1P12 | 1 | trans | Cerebellum |
| SVA_15 | ENSG00000237973.1 | 5.29E-13 | 29472.617 | MTCO1P12 | 1 | trans | Cerebellum |
| SVA_55 | ENSG00000237973.1 | 2.00E-08 | 27693.2393 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_4 | ENSG00000237973.1 | 2.56E-09 | 27128.1241 | MTCO1P12 | 1 | trans | Motor Cortex |
Table 3.
Top 40 significant reference SVA RIPs with the greatest effects on gene downregulation (most negative beta values) from tissue specific matrix eQTL analysis. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
Table 3.
Top 40 significant reference SVA RIPs with the greatest effects on gene downregulation (most negative beta values) from tissue specific matrix eQTL analysis. This analysis included the combination of both cis and trans effects as well as datapoints from both ALS individuals and healthy controls.
| SVA | Gene ID | FDR p-value | Beta value | Gene | Chr | Cis/trans | Tissue |
|---|---|---|---|---|---|---|---|
| SVA_90 | ENSG00000197971.16 | 3.28E-03 | -1337611.1 | MBP | 18 | trans | Spinal Cord |
| SVA_5 | ENSG00000197971.16 | 3.06E-03 | -351064.11 | MBP | 18 | trans | Spinal Cord |
| SVA_87 | ENSG00000123560.14 | 1.98E-07 | -193423.77 | PLP1 | X | trans | Spinal Cord |
| SVA_93 | ENSG00000123560.14 | 2.05E-07 | -193266.83 | PLP1 | X | trans | Spinal Cord |
| SVA_30 | ENSG00000198888.2 | 8.47E-03 | -143159.48 | MT-ND1 | MT | trans | Cerebellum |
| SVA_70 | ENSG00000198886.2 | 1.029E-02 | -133464.74 | MT-ND4 | MT | trans | Cerebellum |
| SVA_30 | ENSG00000198763.3 | 7.85E-04 | -131182.45 | MT-ND2 | MT | trans | Motor Cortex |
| SVA_30 | ENSG00000198938.2 | 6.86E-03 | -111339.82 | MT-CO3 | MT | trans | Cerebellum |
| SVA_30 | ENSG00000198727.2 | 5.66E-03 | -104147.35 | MT-CYB | MT | trans | Cerebellum |
| SVA_84 | ENSG00000123560.14 | 2.11E-07 | -96556.036 | PLP1 | X | trans | Spinal Cord |
| SVA_30 | ENSG00000198786.2 | 7.25E-03 | -94294.133 | MT-ND5 | MT | trans | Motor Cortex |
| SVA_70 | ENSG00000198763.3 | 1.29E-02 | -82929.066 | MT-ND2 | MT | trans | Cerebellum |
| SVA_90 | ENSG00000259001.3 | 6.14E-03 | -73740.939 | ENSG00000259001 | 14 | trans | Spinal Cord |
| SVA_91 | ENSG00000203930.12 | 2.02E-03 | -71147.071 | LINC00632 | X | trans | Spinal Cord |
| SVA_87 | ENSG00000168314.18 | 3.60E-04 | -67162.942 | MOBP | 3 | trans | Spinal Cord |
| SVA_93 | ENSG00000168314.18 | 3.67E-04 | -67112.229 | MOBP | 3 | trans | Spinal Cord |
| SVA_91 | ENSG00000180354.16 | 2.99E-05 | -57438.97 | MTURN | 7 | trans | Spinal Cord |
| SVA_90 | ENSG00000168309.18 | 4.26E-03 | -48145.904 | FAM107A | 3 | trans | Spinal Cord |
| SVA_5 | ENSG00000180354.16 | 1.55E-05 | -43141.656 | MTURN | 7 | trans | Spinal Cord |
| SVA_70 | ENSG00000198712.1 | 6.03E-03 | -43049.326 | MT-CO2 | MT | trans | Cerebellum |
| SVA_90 | ENSG00000168309.18 | 2.45E-03 | -40637.548 | FAM107A | 3 | trans | Motor Cortex |
| SVA_91 | ENSG00000168314.18 | 5.48E-04 | -40457.332 | MOBP | 3 | trans | Spinal Cord |
| SVA_90 | ENSG00000177575.13 | 2.99E-19 | -36552.585 | CD163 | 12 | trans | Spinal Cord |
| SVA_84 | ENSG00000168314.18 | 3.83E-04 | -33494.891 | MOBP | 3 | trans | Spinal Cord |
| SVA_90 | ENSG00000087086.15 | 1.64E-08 | -32820.222 | FTL | 19 | trans | Motor Cortex |
| SVA_5 | ENSG00000168314.18 | 1.72E-04 | -31165.253 | MOBP | 3 | trans | Spinal Cord |
| SVA_16 | ENSG00000123560.14 | 8.97E-07 | -31144.963 | PLP1 | X | trans | Spinal Cord |
| SVA_91 | ENSG00000237973.1 | 6.16E-30 | -29834.037 | MTCO1P12 | 1 | trans | Spinal Cord |
| SVA_87 | ENSG00000173786.17 | 5.88E-04 | -28556.333 | CNP | 17 | trans | Spinal Cord |
| SVA_93 | ENSG00000173786.17 | 5.98E-04 | -28536.883 | CNP | 17 | trans | Spinal Cord |
| SVA_90 | ENSG00000137285.11 | 1.48E-40 | -27173.64 | TUBB2B | 6 | trans | Motor Cortex |
| SVA_5 | ENSG00000237973.1 | 3.92E-10 | -24995.474 | MTCO1P12 | 1 | trans | Motor Cortex |
| SVA_5 | ENSG00000237973.1 | 8.77E-13 | -24871.705 | MTCO1P12 | 1 | trans | Frontal Cortex |
| SVA_91 | ENSG00000064787.13 | 5.47E-07 | -22960.148 | BCAS1 | 20 | trans | Spinal Cord |
| SVA_93 | ENSG00000198840.2 | 3.12E-03 | -22049.981 | MT-ND3 | MT | trans | Spinal Cord |
| SVA_87 | ENSG00000198840.2 | 3.13E-03 | -22046.442 | MT-ND3 | MT | trans | Spinal Cord |
| SVA_90 | ENSG00000164733.22 | 1.39E-04 | -21828.547 | CTSB | 8 | trans | Spinal Cord |
| SVA_90 | ENSG00000079215.15 | 1.69E-05 | -21020.491 | SLC1A3 | 5 | trans | Motor Cortex |
| SVA_5 | ENSG00000237973.1 | 8.40E-24 | -19907.219 | MTCO1P12 | 1 | trans | Spinal Cord |
| SVA_87 | ENSG00000136541.15 | 1.45E-10 | -19551.323 | ERMN | 2 | trans | Spinal Cord |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



