
Submitted:
27 December 2023
Posted:
28 December 2023
You are already at the latest version
Abstract
Eryngium foetidum L., a biennial herb with diverse applications in food and traditional medicine, holds economic and pharmacological significance. Given its growing commercial interest, implementing biotechnological approaches like plant tissue culture is vital for sustainable propagation and metabolite production. In this study, we investigated the in vitro photoautotrophic potential of E. foetidum, examining growth, chlorophyll a fluorescence, photosynthetic pigments, and anatomical features under different sucrose concentrations and gas exchange rates. Acclimatization and survival rates of plants after ex vitro transfer were also assessed. E. foetidum exhibited robust growth in both photoautotrophic and photomixotrophic conditions, with natural ventilation significantly enhancing plant development. Chlorophyll a fluorescence and photosynthetic performance were influenced by sucrose and gas exchange, highlighting the importance of these factors in plant micropropagation. Moreover, the species demonstrated remarkable plasticity during acclimatization, with high survival rates and rapid inflorescence emission. The research provides valuable insights into optimizing in vitro cultivation conditions for E. foetidum, emphasizing the potential for large-scale clonal propagation and exploring secondary metabolites. The observed phenotypic plasticity underscores the adaptability of the species to diverse environments. These biotechnological strategies open avenues for future studies, including the application of elicitors for enhanced secondary metabolite production.
Keywords:
culantro
; gas exchange rate
; natural ventilation
; plant tissue culture
; sucrose supplementation
; wild coriander
1. Introduction
Eryngium foetidum L. (Apiaceae family), commonly known as culantro or wild coriander, is a biennial herb with an erect growth habit, featuring a rosette of long spatulate spiny-toothed leaves and reaching a height of 10–50 cm. It is commonly found in tropical and subtropical regions [1,2]. This species serves a dual purpose, functioning both as a food source (unconventional seasoning vegetable) and a traditional medicine in Latin America, Central and Pacific Islands, Africa, and Asia [3]. The leaves of E. foetidum are notably rich in phosphorus, potassium, and dietary micronutrients, making them suitable for potential use in food fortification [4,5,6]. In the realm of medicine, E. foetidum is recognized for its high bioactive and antioxidant value. It is utilized in the treatment of diseases related to the gastrointestinal tract and exhibits antibacterial, analgesic, anti-inflammatory, anthelmintic, anticonvulsant, and anticancer properties [7,8,9].
The pharmacological importance attributed to E. foetidum is particularly notable due to the various classes of compounds produced in its secondary metabolism, including aldehydes, carotenoids, phenols, and anthraquinones [7]. Furthermore, it is important to note that essential oils are found in the leaves, including molecules such as (2E)-2-dodecenal, trans-2-tetradecenal, 1-dodecanal, 1-decanal, tetradecanal, γ-terpinene, mesitylene, ρ-cymene, α-pinene, 1-undecanal, carotol, and 1-tetradecene [10]. Collectively, this phytochemical profile adds economic value to international trade and in the cosmetic, pharmaceutical, and food industries [7,8,9,10,11].
Due to the increasing commercial interest in E. foetidum, it becomes essential to implement biotechnological approaches, such as plant tissue culture, for the rational propagation of the species. This is crucial to mitigate the risk of genetic erosion resulting from the disorderly exploitation of the population [12]. Furthermore, apart from being a biotechnological strategy, plant tissue culture offers the potential for controlled production of metabolites of interest, in contrast to field conditions. It also enables the exploration of complex biosynthetic pathways that have not been achievable artificially until now [13,14].
Plant tissue culture serves as a platform for plant cloning under aseptic conditions and environmental control in the laboratory [15]. Traditionally, in vitro multiplication relies on three nutritional types of cultures: heterotrophic, where there are no photosynthetically active organs, and the carbon source is solely the carbohydrate supplied in the medium; photomixotrophic, where photosynthetically active organs use the carbohydrate in the medium, along with the consumption of CO2 in the flask headspace; and photoautotrophic (or sugar–free medium), where photosynthetically active organs rely on CO2 in the flask headspace as the carbon source [16,17]. The latter two cultivation systems, photoautotrophic and photomixotrophic, promote in vitro plant growth compared to the first (heterotrophic), which is more suitable for callus cultivation [16].
While photoautotrophic cultivation requires the use of gas-permeable films to enhance gas exchange between the flask headspace and the external environment, these films can also be employed in photomixotrophic (sugar-provided medium) cultivation to improve the ventilation rate of the culture flask [17,18,19]. Investigating the impact of natural ventilation on both photoautotrophy and photomixotrophy in the in vitro cultivation of E. foetidum plants will offer valuable insights into the morphophysiological characteristics influencing the growth and development of plants in these systems. This research can contribute new knowledge for implementing strategies applicable to clonal propagation on a large scale and in the in vitro germplasm banks.
Considering the information presented above, our objective was to assess the in vitro photoautotrophic potential of E. foetidum. In this study, we initially characterized the growth and development of E. foetidum plants under in vitro photomixotrophic conditions (with- and without- natural ventilation) and photoautotrophic systems. Subsequently, we examined the acclimatization of these plants. Our research provides novel insights into how E. foetidum responds to photomixotrophy and photoautotrophy, demonstrating its adaptability to different carbon sources.
2. Materials and Methods
2.1. Plant material and experimental design
The Eryngium foetidum plants utilized in the experiments were sourced from the Germplasm Bank at the Tissue Culture Laboratory (LCT), State University of Maranhão, MA, Brazil (2°34′00″ S and 44°12′00″ W). These plants are regularly maintained in 350-mL transparent glass flasks, each containing 60 mL of Murashige and Skoog (MS) medium with vitamins [20] (PhytoTechnology®, Lenexa, KS, USA), 3% sucrose (w/v; Dinâmica® Química Contemporânea Ltda, São Paulo, SP, Brazil), and solidified with 5.5 g L-1 agar (Dinâmica® Química Contemporânea Ltda, São Paulo, SP, Brazil). The maintenance conditions include a temperature of 25 ±2 °C, an irradiance of 85 μmol m-2 s-1 (provided by four tubular white LED lamps, T8, 9 W, Avant, São Paulo, SP, Brazil), and a 16-hour photoperiod.
Explants (approximately 2.0–2.5 cm) were inoculated into 350-mL transparent glass flasks containing 60 mL of MS culture medium [20] (PhytoTechnology®, Lenexa, KS, USA), following the conditions employed in vitro cultivation in the Germplasm Bank, with sucrose concentrations of 0 and 30 g L−1. Two explants were inoculated per flask. We employed two flask-sealing systems, resulting in different gas exchange rates [21]: polypropylene lids without membranes (14 μL L−1 s−1 CO2 exchange rate) and polypropylene lids with two 10 mm holes covered by microporous tape membranes, as proposed by Saldanha et al. [18] (25 μL L−1 s−1 CO2 exchange rate) (Figure 1A).
The flasks were maintained for 45 days in a growth room under an irradiance of 85 μmol m−2 s−1, a 16-hour photoperiod, and a temperature of 25±2 °C. The design was completely randomized in a 2 × 2 factorial [sucrose concentrations (0 and 30 g L-1) and gas exchange rate (14 and 25 μL L−1 s−1 CO2)], consisting of twelve replications for each treatment (Figure 1B). The experiments were repeated twice.
2.2. Chlorophyll a fluorescence
Chlorophyll fluorescence measurements were conducted on the third fully expanded leaf from the apex. The following parameters: initial fluorescence (F0), maximum fluorescence (Fm), variable fluorescence (Fv), PSII maximum quantum yield (Fv/Fm), total number of active reaction centers per absorption (RC/ABS), maximum primary efficiency of PSII (Fv/F0), and performance index (PI), were assessed using a portable non-modulated fluorimeter (Pocket PEA, Hansatech Instrument Ltd., King’s Lynn, Norfolk, UK). The instrument was calibrated to the leaf under dark conditions for 30 min using a leaf-clip system (Hansatech Instrument Ltd., King’s Lynn, Norfolk, UK). This calibration ensured the complete opening of reaction centers with minimal heat loss and the complete oxidation of QA [22].
2.3. Determination of photosynthetic pigments
Pigments were extracted from leaf discs (5 mm in diameter), which were obtained from the third fully expanded leaf from the apex. The leaf discs were immersed in 5 mL of dimethyl sulfoxide (DMSO; Isofar® Ltda, Duque de Caxias, RJ, Brazil) and left in the dark for 48 h [23]. Absorbance measurements of the samples at 665 nm, 649 nm, and 480 nm were taken using a UV-visible spectrophotometer (mono-beam) (model UV-M51; BEL Engineering Company, Monza, Italy) in a 10-mm quartz cuvette. The calculation of chlorophyll a, b, and carotenoids followed the method outlined by Wellburn [24].
2.4. Quantification of photosynthetic performance
Gas exchange and quantification of the in vitro photosynthetic rate were conducted following the procedures outlined by Castro et al. [25]. A CO2 gas analyzer (Model SBA-5; PP Systems International, Amesbury, MA, USA) and LoggerLite 1.9.3 software (Vernier Software & Technology Caliper, Beaverton, OR, USA) were used for these measurements. Gas exchange was determined by calculating the difference between the reference CO2 and the CO2 levels of plants exposed to atmospheric air. This calculation was based on the leaf area per plant (cm2).
2.5. Growth analyzes
To analyze the growth and development of plants grown in vitro for 45 d, various parameters were assessed, including shoot length (cm), root length (cm), base diameter of the rosette (mm), leaf area (cm2), number of leaves, shoot dry mass (g), root dry mass (g), and shoot-to-root dry mass ratio. The leaf area was determined following the method outlined by Silva et al. [26]. To determine the dry mass, the samples were subjected to 60 °C in an oven with forced air circulation (AmbiKontrol, Arujá, SP, Brazil) until a constant weight was reached.
2.6. Leaf anatomical analysis
For an anatomical structural characterization of plants grown in vitro for 45 days, samples of the third pair of expanded leaves of the median region were fixed in FAA solution (formalin: acetic acid: 50 % ethyl alcohol in 1:1:18 ratio) [27]. After fixation, the samples were dehydrated in increasing ethanolic series (10, 20, 30, 40, 50, 60, 70, 80, 85, 90, and 95%) and included in acrylic resin (Historesin; Leica Instruments, Jena, Germany). Cross-sections of 6-μm thickness were obtained on a semi-automated rotary microtome (model MRP2015, Lupetec Tecnologia Aplicada, São Carlos, SP, Brazil) and stained with toluidine blue (pH 3.2) [28]. Slides were mounted and images were captured using a light microscope (model B20T; Bioptika, Colombo, PR, Brazil) with the U-photo system, coupled to a digital photographic camera (model CMOS-5.0; Bioptika, Colombo, PR, Brazil) and computer with Capture V2.1 software.
2.7. Ex vitro acclimatization
The culture medium was removed from the root systems of 45-day-old E. foetidum plants grown in cultivation systems by gently rinsing them under running water. The plants of each in vitro treatment were then transplanted into 350-mL polystyrene pots (model KP-350 TR; Cristalcopo Descartaveis S/A, Içara, SC, Brazil) containing commercial substrate (Carolina soil®, Santa Cruz do Sul, RS, Brazil). The plants were grown in a greenhouse with an average temperature of 28.5 ± 2°C; Humidity of 80 ±5%; and natural light conditions with an intensity of ~1050 µmol photons m-2 s-1.
2.8. Statistical analyzes
The data were statistically analyzed by two-way analysis of variance (ANOVA), with the means compared by Tukey’s test. All statistical analyses were performed using the software Genes [29].
3. Results
3.1. Gas exchange rates improve chlorophyll a fluorescence and photosynthetic rate in E. foetidum grown in vitro
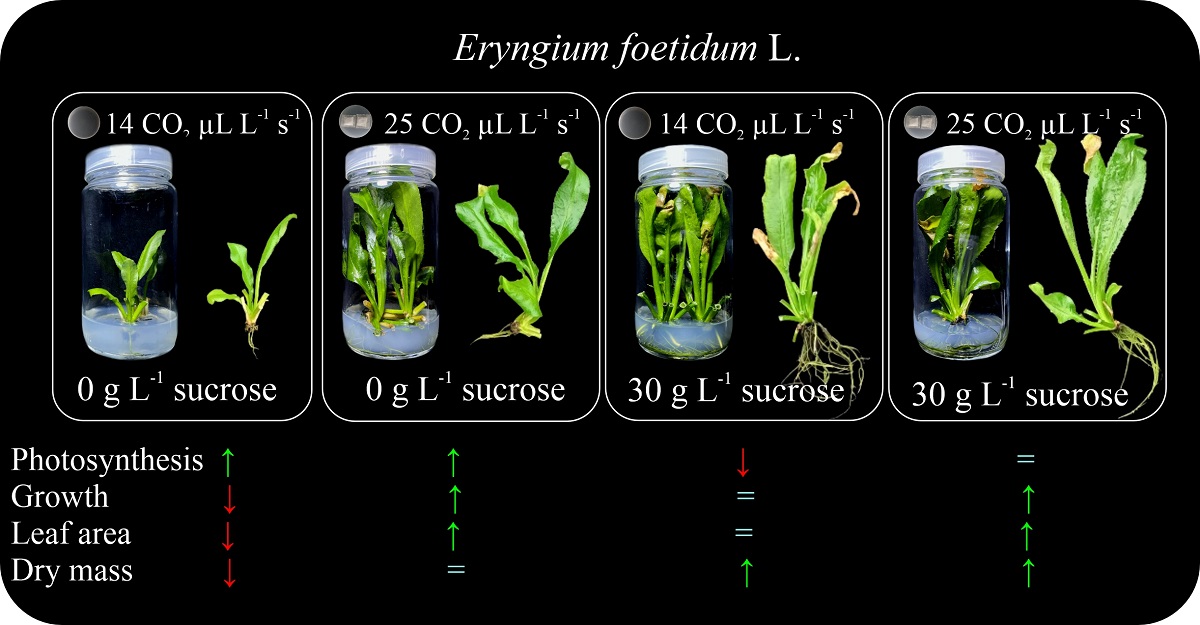
Our results indicate that E. foetidum plants exhibit phenotypic traits indicative of robust growth and development under photoautotrophic conditions (0 g L−1 sucrose and 25 μL L−1 s−1 CO2) and photomixotrophic conditions (30 g L−1 sucrose) both without (14 μL L−1 s−1 CO2; lower gas exchange rate) and with (25 μL L−1 s−1 CO2; higher gas exchange rate) natural ventilation. However, plants under 0 g L−1 sucrose and 14 μL L−1 s−1 CO2 were negatively impacted, likely due to the limiting condition of carbon availability — considered as the control treatment—(Figure 2).
E. foetidum plants grown under 0 g L−1 sucrose and 14 μL L−1 s−1 CO2 displayed a significantly higher initial fluorescence compared to the other treatments (45% higher) (Figure 3A). In parallel, there were no significant differences in the maximum fluorescence for all treatments (ranged from 2112 to 2250) (Figure 3B). Plants under a higher gas exchange rate (25 μL L−1 s−1 CO2), regardless of sucrose concentration, displayed better results for variable fluorescence, total number of active reaction centers per absorption, maximum primary efficiency of PSII, and performance index (Figures 3C; 3E–G). Lastly, only plants under the limiting condition of in vitro cultivation (0 g L−1 sucrose and 14 μL L−1 s−1 CO2) exhibited an average of 0.61 for PS II maximum quantum yield (Figure 3C), which indicates that they are facing stress.
The photosynthetic rate showed that E. foetidum plants in the absence of sucrose exhibited better performance compared to treatments with the presence of this carbohydrate in the culture medium (3–8 times in sucrose-free with 14 and 25 μL L−1 s−1 CO2) (Figure 3H). Notably, plants under cultivation with sucrose and low gas exchange (30 g L−1 sucrose and 25 μL L−1 s−1 CO2) did not show photosynthetic carbon assimilation when their gas exchange was analyzed (Figure 3H). Collectively, the data indicate that sucrose negatively affects photosynthetic performance, but is mitigated when using a higher gas exchange rate (25 μL L−1 s−1 CO2).
3.2. Sucrose concentrations and gas exchange rates do not modulate pigment concentration in E. foetidum grown in vitro
E. foetidum plants grown under photoautotrophic (0 g L−1 sucrose and 25 μL L−1 s−1 CO2), photomixotrophic (0 g L−1 sucrose and 14 μL L−1 s−1 CO2) and photomixotrophic with ventilation natural (0 g L−1 sucrose and 14 μL L−1 s−1 CO2) did not differ statistically from each other for the variables chlorophyll a (ranged from 37 to 49 µg cm−2), total chlorophyll (ranged from 45 to 57 µg cm−2), carotenoids (ranged from 10 to 12 µg cm−2) and total chlorophyll/carotenoid ratio (∼5 µg cm−2); but these three cultivation systems differed significantly for cultivation under 0 g L−1 sucrose and 14 μL L−1 s−1 CO2 (Figure 4A,D–F). Chlorophyll b concentrations and the chlorophyll a/b ratio did not show significant differences between all treatments, with averages of 8 and 6 µg cm−2, respectively (Figure 4B,C).
3.3. Sucrose concentrations and gas exchange rates affect the in vitro growth and development of E. foetidum
E. foetidum plants grown under a higher gas exchange rate (25 μL L−1 s−1 CO2) displayed an increase in shoot size of 74% and 11% under concentrations of 0 and 30 g L−1 sucrose, respectively (Figure 5A). Furthermore, in this same condition of greater gas exchange, there was a significant increase in root length by 382% and 39% at concentrations of 0 and 30 g L−1 sucrose, respectively (Figure 5B). Plants under a lower gas exchange rate (14 μL L−1 s−1 CO2) displayed larger rosette base diameter compared to the treatment with a higher gas exchange rate (25 μL L−1 s−1 CO2) under the concentration of 30 g L−1 sucrose (10% larger diameter); while at the concentration of 0 g L−1 sucrose, the opposite occurred, an expected result since this treatment (0 g L−1 and 14 μL L−1 s−1 CO2) is an extremely limiting cultivation condition for growth and plant development (Figure 5C).
Interestingly, plants grown in a photoautotrophic system (0 g L−1 sucrose and 25 μL L−1 s−1 CO2) displayed as good an increase in leaf area as the photomixotrophic with natural ventilation (30 g L−1 sucrose and 25 μL L−1 s−1 CO2; it ranged from 43 to 59 cm2), both treatments being significantly superior concerning the system with the lowest gas exchange rate (14 μL L−1 s−1 CO2) (Figure 5D). However, these differences in leaf area did not follow for the variable number of leaves, in which the plants under 0 g L−1 sucrose and 14 μL L−1 s−1 CO2 and the plants under 30 g L−1 sucrose and 14 or 25 μL L−1 s−1 CO2 displayed number of leaves with close values (∼5 leaves) (Figure 5E).
Plants grown under a higher concentration of sucrose displayed greater accumulation of dry mass in the shoot and root, independent of the gas exchange rate (Figures 5F and 5G). Concerning the shoot-to-root dry mass ratio, it was observed that in plants under 0 g L−1 sucrose, there was a greater carbon allocation in the shoot, while plants under 30 g L−1 sucrose displayed a more balanced carbon allocation between the shoot and root (Figure 5H).
3.4. Sucrose concentrations and gas exchange rates affect leaf anatomy
E. foetidum plants displayed the same leaf histological organization in all in vitro cultivation conditions accessed. The anatomy of the midrib consists of a uniseriate and amphistomatic epidermis, subepidermal collenchyma on the adaxial and abaxial surfaces; secretory canal and idioblasts distributed throughout the fundamental parenchyma; vascular bundles and fundamental parenchyma distributed throughout the central cavity (Figure 6). However, natural ventilation affected the histological differentiation of leaves compared to treatments with less gas exchange, leading to the formation of thicker and larger leaves (Figure 6).
3.5. Ex vitro acclimatization of E. foetidum grown under different sucrose concentrations and gas exchange rates
Plants showed interesting plasticity to face the acclimatization phase (Figure 7A). E. foetidum plants grown under photoautotrophic (0 g L−1 sucrose and 25 μL L−1 s−1 CO2), photomixotrophic (30 g L-1 sucrose) without- (14 μL L−1 s−1 CO2) and with- (25 μL L−1 s−1 CO2) natural ventilation displayed 100% survival, while only the limiting (0 g L−1 sucrose and 14 μL L−1 s−1 CO2) displayed a low survival (∼33%) (Figure 7B). An interesting fact was the emission of inflorescence in plants grown in these photoautotrophy and photomixotrophy treatments (Figure 7C).
4. Discussion
Plants coordinate their growth and development with carbon availability and source in vitro [30,31,32]. Understanding how sucrose levels (carbon source provided in the cultivation medium) and natural ventilation (carbon source provided in the flask headspace) can affect plant micropropagation offers new perspectives for producing plants with improved morphophysiological characteristics [19]. Our results here demonstrate that E. foetidum plants exhibit in vitro photoautotrophic potential. However, the addition of sucrose in the photomixotrophic cultivation system increased the accumulation of plant dry mass.
Firstly, it is worth highlighting that the E. foetidum plants in limiting cultivation (14 μL L−1 s−1 CO2 and 0 g L−1 sucrose) were drastically affected in their physiological entirety, reducing growth and affecting plant development. This response was expected, since there is a strong limitation in the supply of carbon to plants, both in the cultivation medium due to only organic components making carbon available in small quantities (e.g., vitamins and myoinositol), and in the headspace of the flask due to the use of rigid lids making it difficult for CO2 to diffusion [16,19]. Although this cultivation condition is an obstacle, the explants did not die, which may indicate the high phenotypic plasticity of E. foetidum to face adverse environments. In contrast, the other three in vitro culture systems exemplified by photoautotrophic and photomixotrophic with- and without- natural ventilation provided better responses at the plant morphophysiological level.
In the photomixotrophic system with a lower gas exchange rate (or without natural ventilation), the microenvironment of the flask is the factor with the greatest influence on the growth and development of plants, since carbon is made available through the supply of sucrose in the medium cultivation [33,34]. Undeniably, when using sealing the bottles (rigid lids or without membranes) provides high humidity inside them, greater accumulation of the phytohormone ethylene, reduced gas exchange, and low CO2 concentration [36,37,38,39]. These factors lead to the emergence of plants with morphophysiological disorders such as leaf formation displaying mesophyll with large intercellular spaces, poorly developed vascular system, lack of epicuticular wax formation, and non-functional stomata [16,18,19,40]. Still, in this system, the accumulation of ethylene is a factor that can strongly affect the growth and development of plants [36].
Ethylene affects several processes in plant growth and development, in which its synthesis increases as the concentration of its precursor increases, suggesting that 1-aminocyclopropane-1-carboxylic acid can act as a signaling molecule in plants [41]. Depending on the plant species and type of flask sealing, there may be a high accumulation of ethylene in the flask headspace, causing a lower photosynthetic rate due to inducing leaf senescence pathways and, consequently, lower plant growth [36,42,43]. In the current study, although some symptoms of leaf chlorosis were seen in plants under systems with a lower gas exchange rate (data not shown), there was no leaf abscission; thus, we believe that ethylene may indeed harm plants, but not to the level of being decisive for lower photosynthetic performance. Probably, the sucrose supplied in the cultivation medium is the main inhibitor of photosynthesis in these systems without natural ventilation [19,44].
Sucrose supplementation in the culture medium affects carbon assimilation by photosynthesis done activity reduction of the ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) enzyme [44] since that active carboxylation sites of the Rubisco are blocked with phosphorylated sugars [45]. To face these morphophysiological disorders, there is the photoautotrophic system, which improves the environment in the flask headspace in line with the elimination of the carbon source (carbohydrate; e.g., sucrose) [16,40,42]. Despite this, we cannot consider the sucrose supplemented in the cultivation medium to be a deleterious factor in its entirety. Here, sucrose supplemented in the in vitro culture medium increased the absolute values of plant dry weight.
Previous studies with Solanum tuberosum plants grown under 3% sucrose estimated that CO2 assimilation contributed about 40% of the dry mass, while the other 60% is from sucrose absorption of culture medium [33]. This may be the explanation for the greater biomass of E. foetidum plants in a photomixotrophic with- and without- natural ventilation compared to photoautotrophic, as, often, these plants grown in a photoautotrophic system may not be achieving maximum photosynthesis performance due to extrinsic factors such as quantity (irradiance) and quality (spectral) of light, in addition to the low concentration of CO2 in the headspace of the bottle. Based on this, CO2 enrichment – or forced ventilation – has been applied to plant tissue culture, see [30,31,32,46].
Considering the chlorophyll a fluorescence and photosynthesis of E. foetidum plants, it appears that the response of CO2 assimilation by photosynthesis was not likely associated with photochemical limitations between cultivation systems. Instead, it suggests potential obstacles in terms of biochemical reactions to CO2 fixation. The utilization of natural ventilation in the in vitro cultivation of E. foetidum plants (both photoautotrophic and photomixotrophic with natural ventilation) significantly improved fluorescence aspects in correlation with the photosynthetic rate, compared to photomixotrophic cultivation without natural ventilation.
Photoautotrophic or photomixotrophic with natural ventilation systems also provided greater development of the leaf area of E. foetidum, in addition to altering the cellular differentiation of the leaves. These results corroborate findings observed in other species, for example, Vernonia condensata [19], Pfaffia glomerata [40], Aechmea blanchetiana [47], Lippia dulcis [48], and Brassavola tuberculata [49], in which natural ventilation led to the formation of thicker and larger leaves. These response patterns are important in photosynthetic performance; however, even better, natural ventilation also leads to the formation of functional stomata even in the in vitro cultivation, capable of better controlling gas exchange and evapotranspiration during the acclimatization phase [16,42].
E. foetidum plants grown in vitro displayed an excellent survival percentage, indicating that not only the photoautotrophic system but also photomixotrophy without and with natural ventilation are effective in the micropropagation of the species. An interesting fact was the rapid emission of inflorescences in plants. We do not know for sure whether the cultivation conditions caused an effective stress capable of leading to this or whether it could be related to the vegetative propagation that favored the rapid emission of inflorescences in the photoautotrophic and photomixotrophic treatments with- and without- natural ventilation.
The pioneering results of this research, based on biotechnological strategies, open avenues for future studies. These studies could explore the application of elicitors to develop methods promoting the exploration of secondary metabolites in the medicinal context of the species. The spectacular phenotypic plasticity observed in the in vitro cultivation of the species, as demonstrated here, emphasizes the potential for further advancements in this field.
5. Conclusions
Eryngium foetidum plants exhibit in vitro photoautotrophic potential. Simultaneously, the supplementation of sucrose, along with natural ventilation, resulted in the highest increase in plant biomass. The findings of this research could be applied to the mass propagation of this species, aligning with the exploration of secondary metabolites, particularly those of interest in the phytochemical industry, such as essential oils.
Author Contributions
Conceptualization, S.H.S.F. and M.V.M.P.; methodology, S.H.S.F. and D.R.S.; formal analysis, D.R.S., M.N.B.S., M.C.R.S., M.C.L., V.K.O.S., G.L.A., A.M.R.-R., and A.S.L.; investigation, D.R.S., M.N.B.S., M.C.R.S., M.C.L., V.K.O.S., G.L.A., A.M.R.-R., and A.S.L.; resources, T.R.C., F.A.M.M.A.F., F.O.R., and T.M.F.; data curation, S.H.S.F., D.R.S., and G.L.A.; writing—original draft preparation, S.H.S.F., D.S.B. and D.R.S.; writing—review and editing, S.H.S.F.; D.S.B.; M.V.M.P.; T.R.C., F.A.M.M.A.F., F.O.R., and T.M.F.; supervision, S.H.S.F. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Data Availability Statement
All the data supporting the findings of this study are included in this article.
Acknowledgments
The authors thank the Universidade Estadual do Maranhão (UEMA) and the Fundação de Amparo à Pesquisa e ao Desenvolvimento Científico e Tecnológico do Maranhão (FAPEMA), MA, Brazil.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Castro, T.F.D; Carneiro, W.F.; Reichel, T.; Fabem, S.L.; Machado, M.R.F; Souza, K.K.C; Resende, L.V.; Murgas, L.D.S. The toxicological effects of Eryngium foetidum extracts on zebrafish embryos and larvae depend on the type of extract, dose, and exposure time. Toxicol Res 2022, 11(5), 891–899. [CrossRef]
- Lucas, D.B.; Cardozo, A.L. Eryngium in Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Available online: https://floradobrasil.jbrj.gov.br/FB15529 (accessed on 05 December 2023).
- POWO – Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 30 November 2023).
- Singh, S.; Singh, D.R.; Banu, S.; Salim, K.M. Determination of bioactives and antioxidant activity in Eryngium foetidum L.: a traditional culinary and medicinal herb. Proc Natl Acad Sci, India, Sect B Biol Sci 2013, 83, 453–460. [CrossRef]
- Anju, T.; Rai, N.K.S.; Uthirchamkavu, I.; Sreedharan, S.; Ndhlala, A.R.; Singh, P.; Kumar, A. Analysis of nutritional and antioxidant potential of three traditional leafy vegetables for food security and human wellbeing. S Afr J Bot 2022, 145, 99–110. [CrossRef]
- Leitão, D.d.S.T.C.; Barbosa-Carvalho, A.P.P.; de Siqueira, F.C.; Sousa, R.P.e.; Lopes, A.S.; Chisté, R.C. Extracts of Eryngium foetidum Leaves from the Amazonia Were Efficient Scavengers of ROS and RNS. Antioxidants 2023, 12, 1112. [CrossRef]
- Rodrigues, T.L.M.; Silva, M.E.P.; Gurgel, E.S.C.; Oliveira, M.S.; Lucas, F.C.A. Eryngium foetidum L. (Apiaceae): a literature review of traditional uses, chemical composition, and pharmacological activities. Evid Based Complement Alternat Med 2022, 14, 2896895. [CrossRef]
- Paw, M.; Gogoi, R.; Sarma, N.; Saikia, S.; Chanda, S.K.; Lekhak, H.; Lal, M. Anti-microbial, anti-oxidant, anti-diabetic study of leaf essential oil of Eryngium foetidum L. along with the chemical profiling collected from North East India. J Essent Oil-Bear 2022, 26, 814–829. [CrossRef]
- Zhang, X.; Chen, J.; Zhou, S.; Zhao, H. Ethanol extract of Eryngium foetidum leaves induces mitochondrial associated apoptosis via ROS generation in human gastric cancer cells. Nutr Cancer 2022, 74, 2996–3006. [CrossRef]
- Rodrigues, T.L.M. Desempenho fisiológico e perfil químico do óleo essencial. Orientadora: Eloisa Helena de Aguiar Andrade. 2019. 60 f. Dissertação (Mestrado em Ciências Biológicas/ Botânica Tropical) – Universidade Federal Rural da Amazônia/ Museu Paraense Emílio Goeldi, Belém, 2019. Available online: http://repositorio.ufra.edu.br/jspui/handle/123456789/812 (accessed on 10 December 2023).
- Devi, P.B.; Deb, P.; Singh, H.B. Promotion, utilization, and commercial cultivation of local spices with special reference to eryngo (Eryngium foetidum L.) as a measure for livelihood improvement towards achieving the goal of sustainable development in the indo-burma biodiversity hotspot: a case study from manipur, North-East India. In: Sharma, S.; Kuniyal, J.C.; Chand, P.; Singh, P. (eds) Climate change adaptation, risk management and sustainable practices in the Himalaya. Cham: Springer International Publishing, United States, 2023; pp. 253–266. [CrossRef]
- Rajamohan, S. Harbouring the potential of medicinal and aromatic plants of India: novel biotechnological approach and extraction technologies. In: Máthé, Á.; Khan, I.A. (eds) Medicinal and aromatic plants of India Vol. 1. Medicinal and aromatic plants of the world, vol 8. Cham: Springer International Publishing, United States, 2022; pp. 323–339. [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J Med Plants Res 2009, 3(13), 1222–1239. [CrossRef]
- Fazili, M.A.; Bashir, I.; Ahmad, M.; Yaqoob, U.; Geelani, S.N. In vitro strategies for the enhancement of secondary metabolite production in plants: a review. Bull Natl Res Cent 2022, 46, 35. [CrossRef]
- Nhut, D.T. General information: some aspects of plant tissue culture. In: Nhut, D.T., Tung, H.T., YEUNG, E.CT. (eds) Plant Tissue Culture: new techniques and application in horticultural species of tropical region. Singapore: Springer Singapore, Singapore, 2022; pp. 1–23. [CrossRef]
- Kozai, T., Kubota, C. Concepts, definitions, ventilation methods, advantages and disadvantages. In: Kozai, T., Afreen, F., Zobayed, S. (eds) Photoautotrophic (sugar–free medium) Micropropagation as a New Micropropagation and Transplant Production System. Springer: Dordrecht, Dordrecht, 2005, pp. 19–30. [CrossRef]
- Pires, H.P.; Felipe, S.H.S.; Pinheiro, M.V.M.; Paula Alves, A.; Alves, G.L.; Catunda, F.E.A.; Figueiredo, F.A.M.M.A.; Reis, F.O.; Ferraz, T.M.; Corrêa, T.R. Natural ventilation and sucrose concentrations in the in vitro culture system affect the acclimatization of "Perola" pineapple plants under different substrates. Aust J Crop Sci 2023, 17(1), 90–98. [CrossRef]
- Saldanha, C.W.; Otoni, C.G.; Azevedo, J.L.F.; Dias, L.L.C.; Rêgo, M.M.; Otoni, W.C. A low-cost alternative membrane system that promotes growth in nodal cultures of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell Tiss Organ Cult 2012, 110, 413–422. [CrossRef]
- Fortini, E.A.; Batista, D.S.; Mamedes-Rodrigues, T.C.; Felipe, S.H.S.; Correia, L.N.F.; Chagas, K.; Silva, P.O.; Rocha, D.I.; Otoni, W.C. Gas exchange rates and sucrose concentrations affect plant growth and production of flavonoids in Vernonia condensata grown in vitro. Plant Cell Tiss Organ Cult 2021, 144, 593–605. [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 1962, 15, 473–497. [CrossRef]
- Batista, D.S.; Castro, K.M.; Ribeiro, D.M.; Caixeta, E.T.; Oliveira Santos, M.; Viccini, L.F.; Otoni, W. C. Ethylene responses and ACC oxidase gene expression in Lippia alba (Verbenaceae) chemotypes with varying ploidy levels. In Vitro Cell Dev Biol -Plant 2017, 53, 278–284. [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.L.E.G.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: a review of current instrumentation. Funct Ecol 1989, 3(4), 497-514. [CrossRef]
- Santos, R.P.; Ferreira Da Cruz, A.C.; Iarema, L.; Kuki, K.N.; Campos Otoni, W. Protocolo para extração de pigmentos foliares em porta-enxertos de videira micropropagados. Rev Ceres 2008, 55, 356–364.
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J Plant Physiol 1994, 144(3), 307-313. [CrossRef]
- Castro, K.M.; Batista, D.S.; Fortini, E.A.; Silva, T.D.; Felipe, S.H.S.; Fernandes, A.M.; Sousa, R.M.J.; Nascimento, L.S.Q., Campos, V.R.; Grazul, R.M.; Viccini, L.F.; Otoni, W.C. Photoperiod modulates growth, morphoanatomy, and linalool content in Lippia alba L. (Verbenaceae) cultured in vitro. Plant Cell Tiss Organ Cult 2019, 139, 139–153. [CrossRef]
- Silva, T.D.; Batista, D.S.; Fortini, E.A.; Castro, K.M.; Felipe, S.H.S.; Fernandes, A.M.; Sousa, R.M.J.; Chagas, K.; Silva, J.V.S.; Correia, L.N.F.; Farias, L.M.; Leite, J.P.V.; Rocha, D.I.; Otoni, W.C. Blue and red light affects morphogenesis and 20-hydroxyecdisone content of in vitro Pfaffia glomerata accessions. J Photochem Photobiol B, Biol 2020, 203, 111761. [CrossRef]
- Johansen, D.A. Plant microtechnique. Mac Graw-Hill Book Co., Inc., New York, 1940; 523 pp.
- O’Brien, T.P.; McCully, M.E. The study of plant structure: principles and selected methods; Termarcarphi Pty. Ltd.: Melbourne, Australia, 1981.
- Cruz, C.D. Genes: a software package for analysis in experimental statistics and quantitative genetics. Acta Sci Agron 2013, 35, 271-276. [CrossRef]
- Ferreira, P.R.B.; Cruz, A.C.F.; Batista, D.S.; Nery, L.A.; Andrade, I.G.; Rocha, D. I.; Felipe, S.H.S.; Koehler, A.D.; Nunes-Nesi, A. CO2 enrichment and supporting material impact the primary metabolism and 20-hydroxyecdysone levels in Brazilian ginseng grown under photoautotrophy. Plant Cell Tiss Organ Cult 2019, 139, 77–89. [CrossRef]
- Louback, E.; Batista, D.S.; Pereira, T.A.R.; Mamedes-Rodrigues, T.C.; Silva, T.D.; Felipe, S.H.S.; Rocha, D.I.; Steinmacher, D.A.; Otoni, W.C. CO2 enrichment leads to altered cell wall composition in plants of Pfaffia glomerata (Spreng.) Pedersen (Amaranthaceae). Plant Cell Tiss Organ Cult 2021, 145, 603–613. [CrossRef]
- Luis, S.J.; Jabín, B.J. CO2–enriched air in a temporary immersion system induces photomixotrophism during in vitro multiplication in vanilla. Plant Cell Tiss Organ Cult 2023, 155, 29–39. [CrossRef]
- Wolf, S.; Kalman-Rotem, N.; Yakir, D.; Zrv, M. Autotrophic and heterotrophic carbon assimilation of in vitro grown potato (Solanum tuberosum L) plants. J Plant Physiol 1998, 153(5–6), 574–580. [CrossRef]
- Shin, KS.; Park, SY.; Paek, KY. Sugar metabolism, photosynthesis, and growth of in vitro plantlets of Doritaenopsis under controlled microenvironmental conditions. In Vitro Cell Dev Biol -Plant 2013, 49, 445–454. [CrossRef]
- Alves, J.P.; Pinheiro, M.V.M.; Corrêa, T.R.; Alves, G.L.; Marinho, T.R.S.; Batista, D.S.; Figueiredo, F.A.M.M.A.; Reis, F.O.; Ferraz, T.M.; Campostrini, E. Morphophysiology of Ananas comosus during in vitro photomixotrophic growth and ex vitro acclimatization. In Vitro Cell Dev Biol -Plant 2023, 59, 106–120. [CrossRef]
- Vinh, B.V.T.; Tung, H.T.; Mai, N.T.N.; Khai, H.D.; Luan, V.Q.; Phong, T.H.; Phong, T.N.P.; Nhut, D.T. Enhanced efficient micropropagation and reduced abnormal phenomena in Phyllanthus amarus plantlets cultured on medium containing silver nanoparticles. S Afr J Bot 2023, 163, 217–225. [CrossRef]
- Askari, N.; Aliniaeifard, S.; Visser, R.G.F. Low CO2 Levels are detrimental for in vitro plantlets through disturbance of photosynthetic functionality and accumulation of reactive oxygen species. Horticulturae 2022, 8, 44. [CrossRef]
- Silva, L.M.; Carvalho, V.S.; Generoso, A.L.; Miranda, D.P.; Costa Júnior, O.D.; Simioni, P.F.; Santana, D.B.; Cunha, M.; Oliveira, J.G.; Viana, A.P. Micropropagation of interspecific hybrids of Vitis spp. in microenvironments with different gas exchanges. Sci Hortic 2022, 305, 111413. [CrossRef]
- Carrari-Santos, R.; Vettorazzi, R.G.; Pinto, V.B.; Pinto, V.B.; Sena, E.O.A.; Oliveira, J.G.; Campostrini, E.; Silveira, V.; Santa-Catarina, C. Microporous membrane and culture medium affect in vitro seedling development of Dalbergia nigra (Vell.) Ex Benth. (Fabaceae) by modulation of the protein profile and accumulation of ethylene and CO2. Plant Cell Tiss Organ Cult 2023, 153, 559–576. [CrossRef]
- Iarema, L.; Cruz, A.C.F.; Saldanha, C.W.; Dias, L.L.C.; Vieira, R.F.; Oliveira, E.J.; Otoni, W.C. Photoautotrophic propagation of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell Tiss Organ Cult 2012, 110, 227–238. [CrossRef]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, Issue suppl_1, S131–S151. [CrossRef]
- Kubota, C. Concepts and background of photoautotrophic micropropagation. In: Morohoshi N, Komamine A (eds) Molecular breeding of woody plants. Elsevier, Amsterdam, 2001; pp. 325–334. [CrossRef]
- Cournac, L.; Dimon, B.; Carrier, P.; Lohou, A.; Chagvardieff, P. Growth and photosynthetic characteristics of Solanum tuberosum plantlets cultivated in vitro in different conditions of aeration, sucrose supply, and CO2 enrichment. Plant Physiol 1991, 97 (1), 112–117. [CrossRef]
- Desjardins, Y.; Hdider, C.; Riek, J. Carbon nutrition in vitro — regulation and manipulation of carbon assimilation in micropropagated systems. In: Aitken-Christie, J., Kozai, T., Smith, M.A.L. (eds) Automation and environmental control in plant tissue culture. Springer: Dordrecht, 1995; pp. 441–471. [CrossRef]
- Mayak, S.; Tirosh, T.; Ilan, A.; Duvdevani, A.; Khayat, E. Growth and development of pineapple (Ananas comosus L.) plantlets cultured in vitro at enriched and ambient CO2 environments. In International Symposium on Biotechnology of Tropical and Subtropical Species Part 2 1998, 461 (pp. 225-230). [CrossRef]
- Marques, I.; Fernandes, I.; Paulo, O.S.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. A transcriptomic approach to understanding the combined impacts of supra-optimal temperatures and CO2 revealed different responses in the polyploid Coffea arabica and its diploid progenitor C. canephora. Int J Mol Sci 2021, 22, 3125. [CrossRef]
- Martins, J.P.R.; Almeida-Rodrigues, L.C.; Santos, E.R.; Gontijo, A.B.P.L.; Falqueto, A.R. Impacts of photoautotrophic, photomixotrophic, and heterotrophic conditions on the anatomy and photosystem II of in vitro-propagated Aechmea blanchetiana (Baker) L.B. Sm. (Bromeliaceae). In Vitro Cell.Dev.Biol.-Plant 2020, 56, 350–361. [CrossRef]
- Rocha, T.T.; Araújo, D.X.; Silva, A.M.; Oliveira, J.P.V.; Carvalho, A.A.; Gavilanes, M.L.; Bertolucci, S.K.V.; Alves, E.A.; Pinto, J.E.B.P. Morphoanatomy and changes in antioxidant defense associated with the natural ventilation system of micropropagated Lippia dulcis plantlets. Plant Cell Tiss Organ Cult 2022, 151, 467–481. [CrossRef]
- Soares, J.S.; Ramos, J.C.M.; Sorgato, J.C.; Ribeiro, L.M.; Reis, L.C. Brassavola tuberculata Hook.: in vitro growth and ex vitro establishment as a function of the micropropagation system and sucrose. Braz J Biol 2023, 83, e270892. [CrossRef]
Figure 1.
Schematic representation of the experimental design. (A) Polypropylene lids without a membrane (14 μL L−1 s−1 CO2 exchange rate) and with two membranes (25 μL L−1 s−1 CO2 exchange rate). (B) Explants of Eryngium foetidum at the beginning of in vitro culture. Further details can be found in the Materials and Methods section. Scale bars: 1A = 1 cm and 1B = 5 cm.
Figure 1.
Schematic representation of the experimental design. (A) Polypropylene lids without a membrane (14 μL L−1 s−1 CO2 exchange rate) and with two membranes (25 μL L−1 s−1 CO2 exchange rate). (B) Explants of Eryngium foetidum at the beginning of in vitro culture. Further details can be found in the Materials and Methods section. Scale bars: 1A = 1 cm and 1B = 5 cm.

Figure 2.
Forty-five-day-old Eryngium foetidum plants under different sucrose concentrations and gas exchange rates in vitro. Scale bars: 2 cm.
Figure 2.
Forty-five-day-old Eryngium foetidum plants under different sucrose concentrations and gas exchange rates in vitro. Scale bars: 2 cm.

Figure 3.
Chlorophyll fluorescence parameters and photosynthetic rate in Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rates). F0: initial fluorescence; Fm: maximum fluorescence; Fv: variable fluorescence; Fv/Fm: PS II maximum quantum yield; RC/ABS: total number of active reaction centers per absorption; Fv/F0: maximum primary efficiency of PSII; PI: performance index; and A: photosynthetic rate. Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 5).
Figure 3.
Chlorophyll fluorescence parameters and photosynthetic rate in Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rates). F0: initial fluorescence; Fm: maximum fluorescence; Fv: variable fluorescence; Fv/Fm: PS II maximum quantum yield; RC/ABS: total number of active reaction centers per absorption; Fv/F0: maximum primary efficiency of PSII; PI: performance index; and A: photosynthetic rate. Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 5).
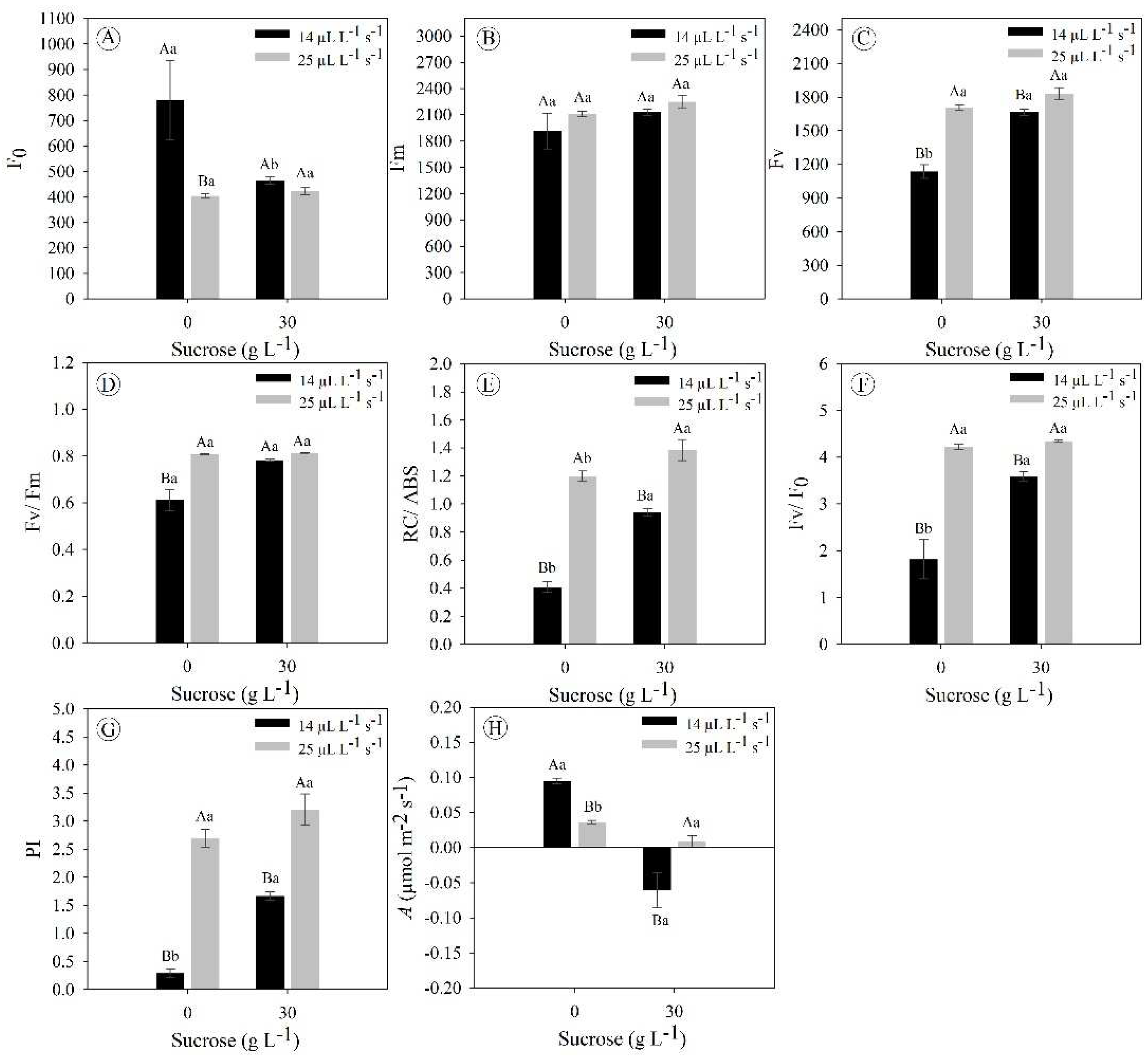
Figure 4.
Concentration of photosynthetic pigments in Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L−1) and gas exchange levels (14 and 25 μL L−1 s−1 CO2 exchange rates). Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 5).
Figure 4.
Concentration of photosynthetic pigments in Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L−1) and gas exchange levels (14 and 25 μL L−1 s−1 CO2 exchange rates). Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 5).
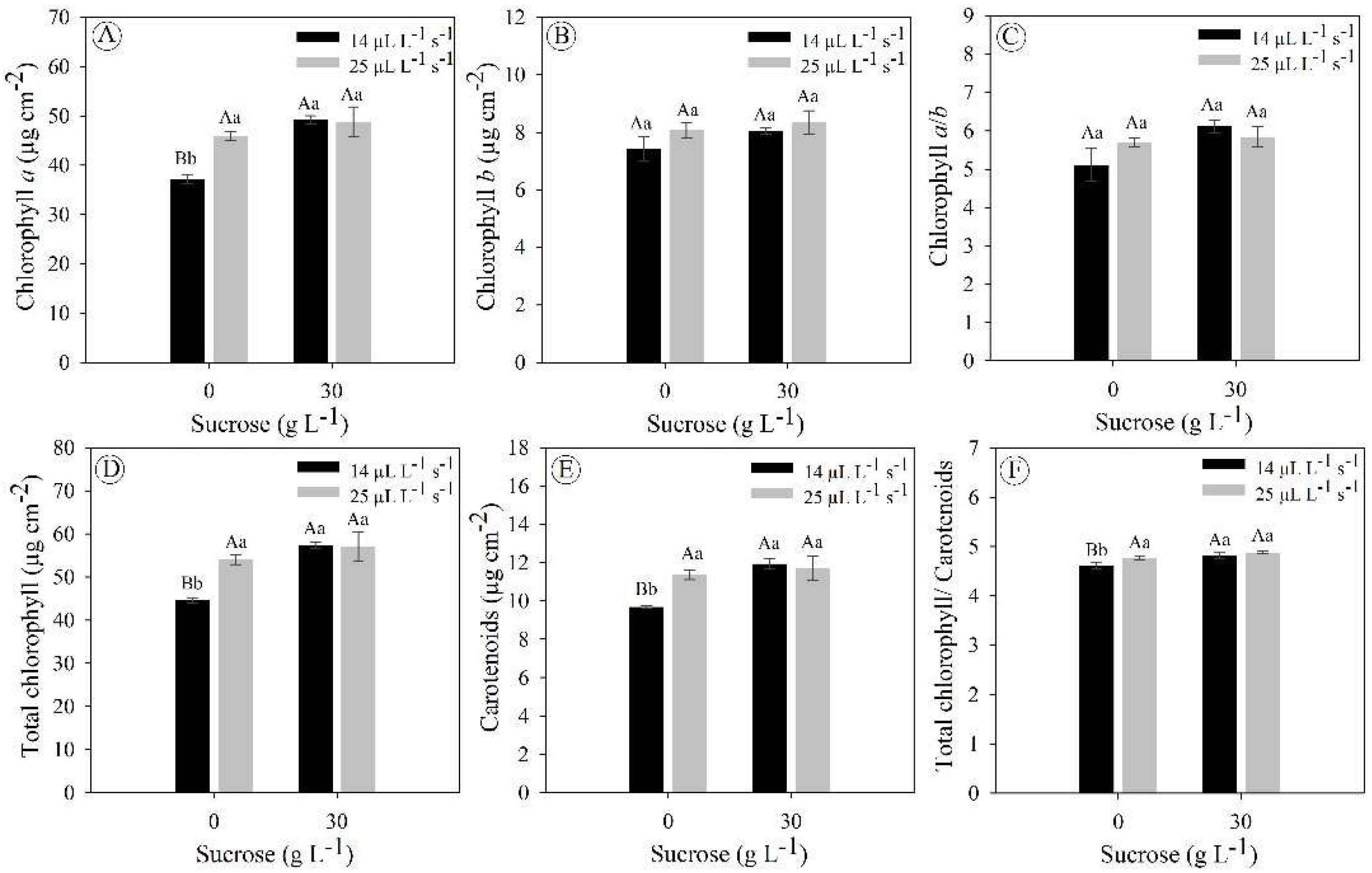
Figure 5.
Growth parameters of Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 7).
Figure 5.
Growth parameters of Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). The bars represent the standard error (n = 7).
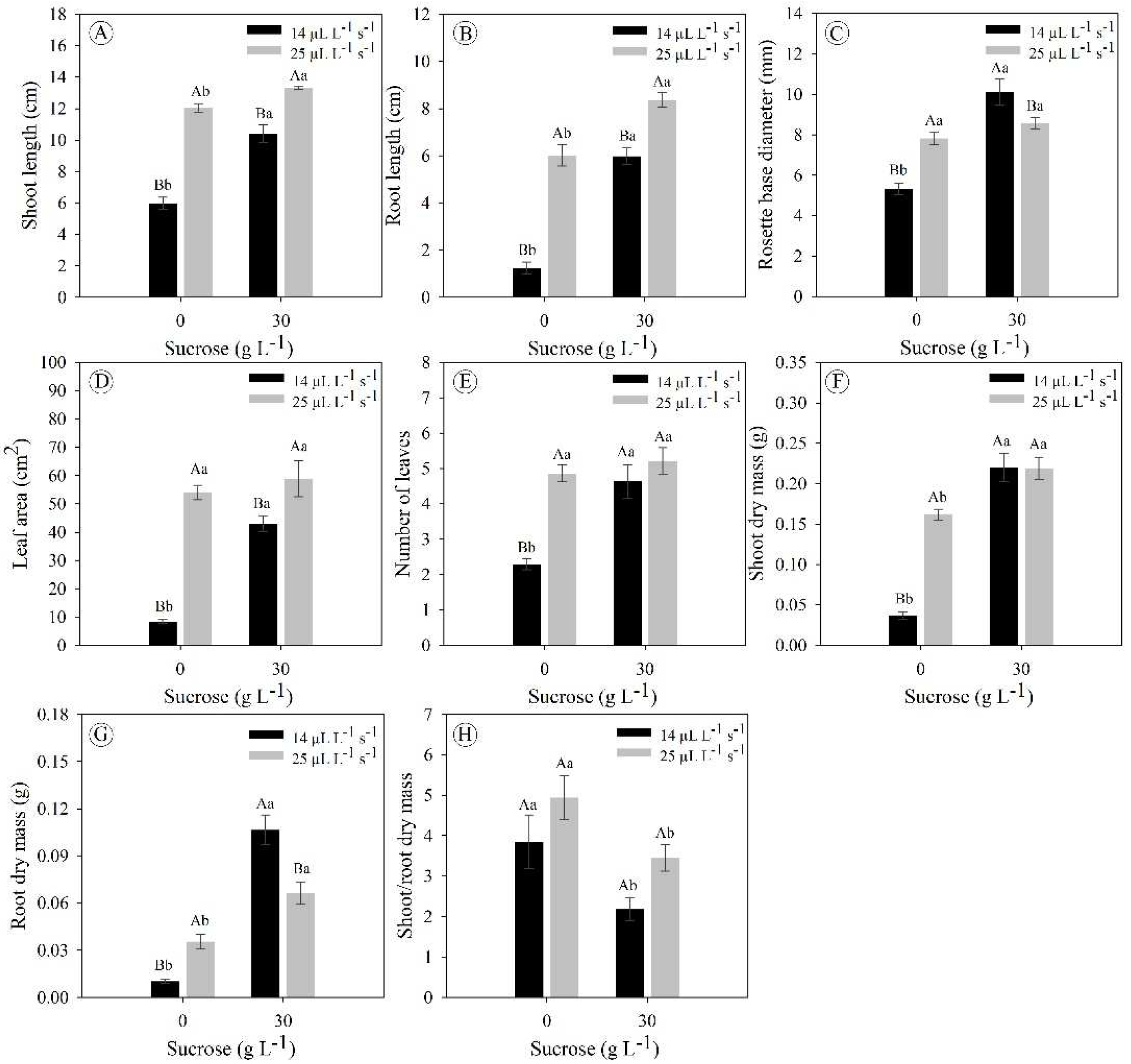
Figure 6.
Leaves of Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). Ab: abaxial epidermis; Ad: adaxial epidermis; Cc: central cavity; Co: collenchyma; Ep: epidermis; Gp: ground parenchyma; Id: idioblast; Sc: secretory canal; and Vb: vascular bundle. Scale bars: 300 μm.
Figure 6.
Leaves of Eryngium foetidum after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). Ab: abaxial epidermis; Ad: adaxial epidermis; Cc: central cavity; Co: collenchyma; Ep: epidermis; Gp: ground parenchyma; Id: idioblast; Sc: secretory canal; and Vb: vascular bundle. Scale bars: 300 μm.
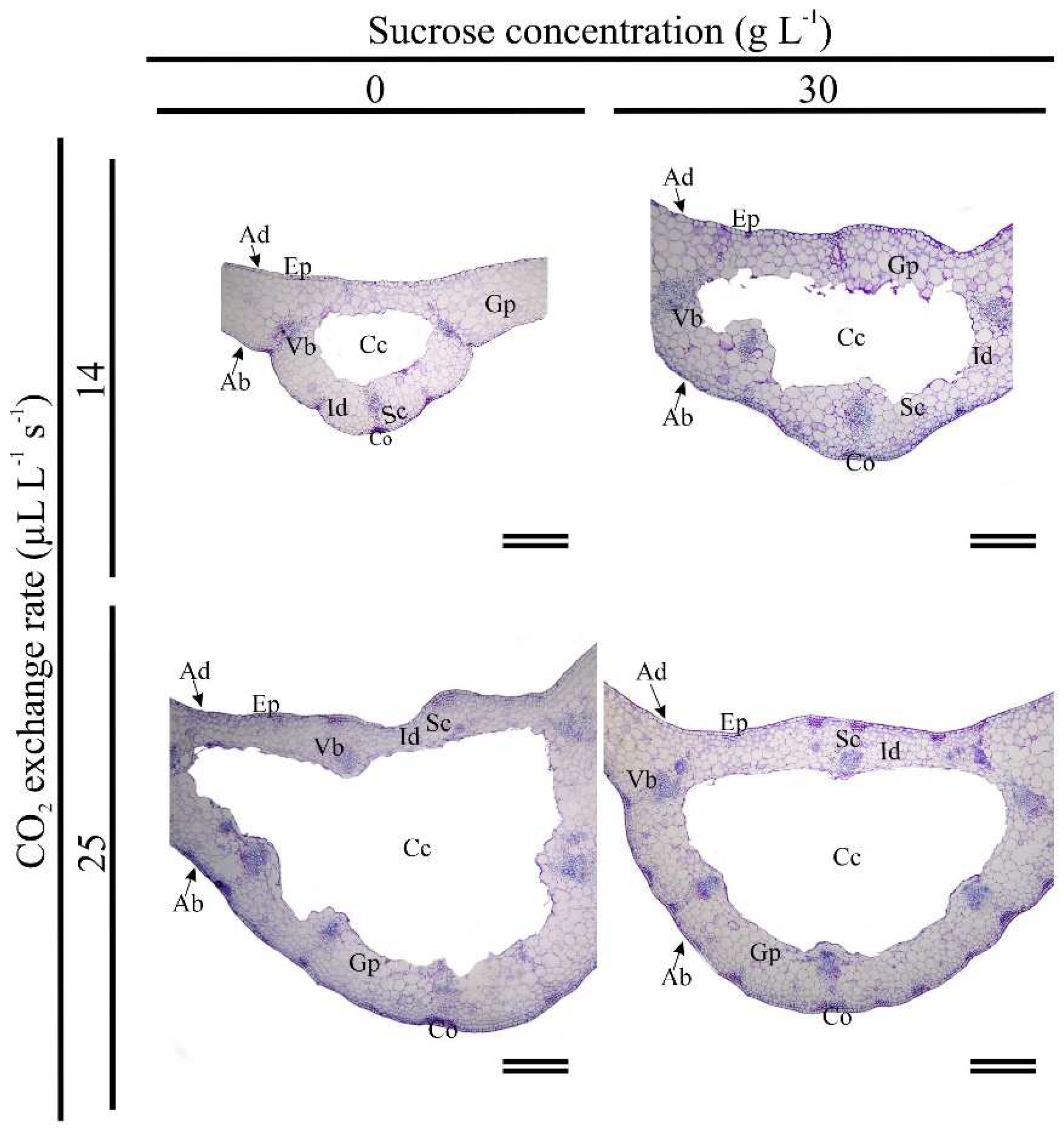
Figure 7.
Acclimatization of Eryngium foetidum plants after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). (A) Sixty-five-day-old Eryngium foetidum plants during acclimatization; (B) Survival after 60 days of ex vitro cultivation; and (C) Inflorescence emission after 60 days of ex vitro cultivation. Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). Scale bars: 7A = 5 cm and 7B–C = the bars represent the standard error (n = 6).
Figure 7.
Acclimatization of Eryngium foetidum plants after 45 days of in vitro cultivation under different sucrose concentrations (0 and 30 g L-1) and gas exchange levels (14 and 25 μL L-1 s-1 CO2 exchange rate). (A) Sixty-five-day-old Eryngium foetidum plants during acclimatization; (B) Survival after 60 days of ex vitro cultivation; and (C) Inflorescence emission after 60 days of ex vitro cultivation. Uppercase letters compare the means of plants under different CO2 exchange rates at the same sucrose concentration, while lowercase letters compare plants under the same CO2 exchange rate at different sucrose concentrations (Tukey’s test; P ≤ 0.05). Scale bars: 7A = 5 cm and 7B–C = the bars represent the standard error (n = 6).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



