Submitted:
28 December 2023
Posted:
28 December 2023
You are already at the latest version
Abstract
Geniposide is a major ingredient in the herb gardenia, and this study investigated effects of geniposide on lipopolysaccharide-stimulated macrophages (RAW 264.7). Griess reagent assay, Fluo-4 calcium assay, dihydrorhodamine 123 assay, multiplex cytokine assay, quantitative RT-PCR, and flow cytometry assay were done using RAW 264.7. Data showed that geniposide at concentrations of 10, 25, and 50 μM reduced significantly levels of nitric oxide, intracellular Ca2+, and hydrogen peroxide in lipopolysaccharide-stimulated RAW 264.7. Multiplex cytokine assay represented geniposide at concentrations of 10, 25, and 50 μM meaningfully suppressed levels of IL-6, G-CSF, MCP-1 and MIP-1α in RAW 264.7 provoked by lipopolysaccharide; additionally, geniposide at concentrations of 25 and 50 μM meaningfully suppressed levels of TNF-α, IP-10, GM-CSF, and MIP-1β. Flow cytometry assay represented that geniposide reduces significantly the level of phosphorylated P38 MAPK in RAW 264.7 provoked by lipopolysaccharide. Geniposide meaningfully suppressed lipopolysaccharide-induced transcription of inflammatory target genes such as Chop, Jak2, Fas, c-Jun, c-Fos, Stat3, Nos2, Ptgs2, Gadd34, Asc, Xbp1, Nlrp3, and Par-2. Taken together, geniposide exerts anti-inflammatory effects in lipopolysaccharide-stimulated macrophages via calcium-P38 MAPK pathway.

Keywords:
Geniposide
; Lipopolysaccharide
; Inflammation
; Macrophages
; Cytokine
; Calcium
; P38 MAPK
1. Introduction
Immunity is the essential process for health and human life. Especially, innate immunity protects human body against invading pathogens such as bacteria, virus, and fungus. Inflammation, which is classically known to consist of redness, swelling, heat, pain, and loss of function, is the initial host defense reaction against pathogenic infection [1]. But the uncontrolled inflammation is dangerous to host survival [2] and excessive inflammation sometimes leads to not only acute inflammatory diseases but also chronic inflammatory disorders [3]. P38 MAPK controls macrophage plasticity via promoting the endoplasmic reticulum stress (ERS), unfolded protein responses, and glucose intolerance in the infected host [4]. Atherosclerosis is a chronic inflammatory disease [5] and it is well known that cytokines and chemokines might contribute to the exacerbation of atherosclerosis [5,6]. Thus, the regulation of inflammation has become more and more important for treating inflammatory diseases [7]. Representative cytokines related to inflammatory diseases include interleukin-6 (IL-6). Not only cytokines but also nitric oxide (NO) are important factors involved in the inflammatory response process of macrophages [8]. Tumor necrosis factor-α (TNF-α) is also an important cytokine associated with endotoxemia [9]. Lipopolysaccharide (LPS, endotoxin) can stimulate immune cells like macrophages, which produce cytokines and hydrogen peroxide (H2O2). Monocyte chemoattractant protein-1 (MCP-1) is also one of increased chemokines in endotoxemia. Excessive cytokines in infectious diseases might have a serious outcome in patients. Thus, LPS-induced macrophages is regarded as being useful to study the anti-inflammatory effect of potential therapeutic agents. TNF-α-converting enzyme (TACE) is a sheddase responsible for cleaving membrane TNF and reactive oxygen species (ROS) mediate LPS-induced up-regulation of TACE activity via P38 MAPK activation in monocytes [10]. Many studies reported that ERS-related pathways including C/EBP homologous protein (CHOP) and protease-activated receptor-2 (PAR-2) might be up-regulated through P38 MAPK phosphorylation, which is related with growth arrest and DNA damage-inducible gene 34 (GADD34) [11,12,13,14]. CHOP activation is also linked to X-box binding protein 1 (XBP1). Recently, macrophage inflammatory proteins (MIPs) have been recognized as important in the amplification process of inflammatory phenomena. Interferon inducible protein (IP)-10 is also increased in inflammation. In addition, it is known that not only cytokines and chemokines but also growth factors such as increase in blood levels in inflammatory diseases and worsen the condition.
Geniposide is a major ingredient in the herb gardenia [13]. The herb gardenia, a fruit of Gardenia jasminoides, is widely used in Asia such as Korea, China, India, Japan, Vietnam, Taiwan, and Myanmar to care for inflammatory disorders [15,16,17]. Geniposide is known to suppress inducible nitric oxide synthase and in macrophages provoked by LPS as well as cyclooxygenase-2 [16]. Cyclooxygenase-2 is also reduced by geniposide in macrophages. Cheng et al. suggested geniposide inhibits cytokines levels in RAW 264.7 induced by LPS as well as reduced the plaque size and serum TNF-α level in ApoE-/- mice via microRNA-101/ mitogen-activated protein kinase phosphatase-1/ P38 signaling [18]. Geniposide was reported to reduce atherosclerotic plaques in rabbits via MAPK pathway [19]. Despite such studies, effects of geniposide on ERS-related cytokine production of LPS-provoked macrophages are still unknown.
Figure 1.
Structural formula of geniposide.
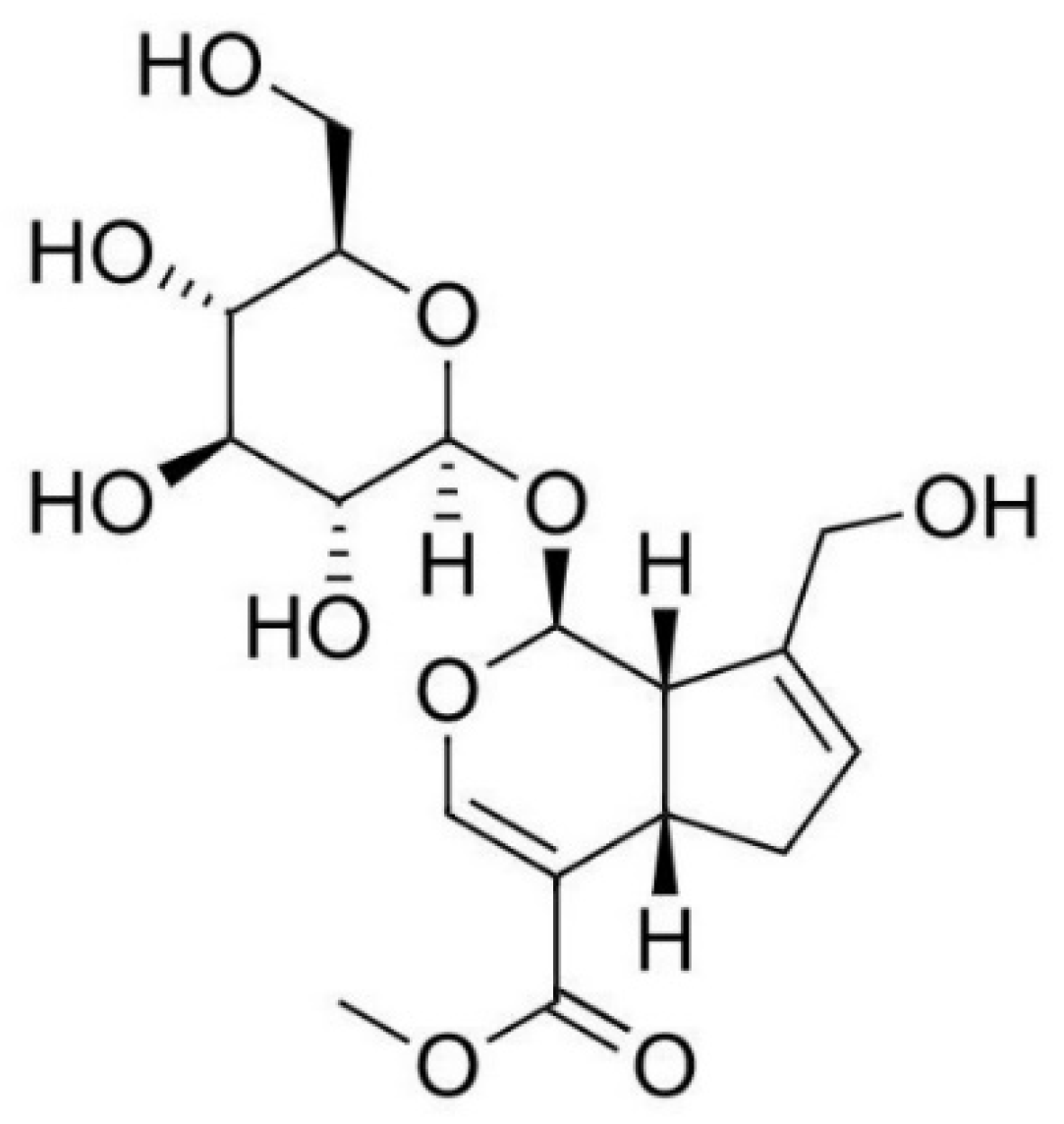
In this study, effects of geniposide on cytokines, H2O2, and NO in LPS-provoked RAW 264.7. In addition, quantitative PCR was carried out to measure mRNA expressions of ERS-related genes such as Chop, Janus kinase 2 (Jak2), first apoptosis signal receptor (Fas), c-Jun, c-Fos, Signal Transducer and Activator of Transcription (Stat)-1, Stat-3, nitric oxide synthase 2 (Nos2), cyclooxygenase-2 (Ptgs2), Gadd34, Apoptosis-associated speck-like protein containing a caspase recruitment domain (Asc), Xbp1, Nlrp3 (cryopyrin), and Par-2 in RAW 264.7. Finally, geniposide has been shown to suppress levels of cytokines, H2O2, and NO in LPS-provoked RAW 264.7 through calcium-P38 MAPK signaling.
2. Materials and Methods
The experiment was conducted with reference to previous valuable reports [20,21], and more details are in the supplementary file.
2.1. Materials
DMEM, FBS, penicillin, streptomycin, phosphate buffered saline (PBS), dimethyl sulfoxide (DMSO), Dihydrorhodamine 123 (DHR123) assay kit, and Fluo-4 NW calcium assay kit were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Geniposide, baicalein, MILLIPLEX MAP Mouse Cytokine/Chemokine Magnetic Bead Panel kit, and Griess reagent were purchased from Millipore (Billerica, MA, USA).
2.2. Methods
2.2.1. Cell culture and cell viability
RAW 264.7 were obtained from the Korea Cell Line Bank (Seoul, Korea). Cells were cultured in DMEM supplemented with 10% FBS containing of penicillin (100 U/mL) and streptomycin (100 µg/mL) at 37°C in a humidified atmosphere of 5% CO2. In order to evaluate cell viability, RAW 264.7 cell lines (passage number 3) were seeded in 96-well plates (1×104 cells/well) and treated with various concentrations of geniposide for 24 h and 48 h [20]. MTT assay was used to measure effects of geniposide on viabilities of RAW 264.7.
2.2.2. Levels of H2O2, NO, and Ca2+ in cells
The production of H2O2 in RAW 264.7 (1x104 cells/well) were measured by dihydrorhodamine (DHR) 123 assay after 24 h and 48 h treatment [21]. NO level in RAW 264.7 (1x104 cells/well) were measured via Griess reagent assay after 24 h treatment [20]. Ca2+ release in RAW 264.7 (1x105 cells/well) was measured with Fluo-4 NW Calcium Assay Kits (Thermo Fisher Scientific, Waltham, MA, USA) after 18 h treatment [20]. Baicalein (25 µM) was used as a positive control.
2.2.3. Levels of inflammatory cytokines
RAW 264.7 were incubated in 96-well plates (1 x 104 cells/well) with various concentrations of geniposide and LPS for 24 h. After 24 h incubation, cytokines released from treated cells were measured in cell culture supernatants using a Luminex assay based on xMAP technology with Bio-Plex 200 suspension array system (Bio-Rad, Hercules, CA, USA) [20]. The following cytokine productions were analyzed: IL-6, TNF-α MCP-1, IP-10, MIP-1α, MIP-1β, MIP-2, granulocyte colony-stimulating factor (G-CSF) and granulocyte-macrophage colony-stimulating factor (GM-CSF), and lipopolysaccharide-induced CXC chemokine (LIX; CXCL5). Baicalein (25 µM) was used as a positive control.
2.2.4. Quantitative Real Time RT-PCR for inflammatory genes
The transcription of inflammatory genes such as Chop, Jak2, Fas, c-Jun, c-Fos, Stat-1, Stat-3, Nos2, Ptgs2, Gadd34, Asc, Xbp1, Nlrp3, and Par-2 was evaluated with quantitative real time RT-PCR using Bio-Rad CFX 96 Real-Time PCR Detection System (Bio-Rad) [20]. Baicalein (25 µM) was used as a positive control.
2.2.5. Phosphorylation of P38 MAPK
Phosphorylation of P38 MAPK was evaluated via flow cytometry using an Attune NxT flow cytometer (Thermo Fisher Scientific) [20]. RAW 264.7 were incubated in 6-well plates (3 x 105 cells/well) with various concentrations of geniposide and LPS. After 18 h incubation, RAW 264.7 were stained with 5 µg/mL of phospho-P38 MAPK (T180/Y182) Antibody (eBioscience 17-9078-42, Life Technologies Corporation, Carlsbad, CA, USA). A serial gating strategy used forward scatter versus side scatter plots, forward scatter versus viability stain plots, and the target antibody expression plots. Details for startup, proper calibration and operation of the Attune can be found in the Attune User Guide (https://assets.thermofisher.com/TFS-Assets/LSG/manuals/100024235_AttuneNxT_HW_UG.pdf). For analysis of raw data, Attune NxT software (Thermo Fisher Scientific) was used. Mouse IgG2b kappa Isotype Control (eBioscience 12-4732-81) was used to confirm the specificity of phospho-p38 MAPK Antibody. Baicalein (25 µM) was used as a positive control.
2.2.6. Statistical analysis
The results shown are summarized from three independent experiments and represent the mean ± SD. After testing for the normality of data, significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test or Kruskal-Wallis test, as appropriate. The program GraphPad Prism 6.0 software (GraphPad Software, San Diego, CA, USA) was used for statistics.
3. Results
3.1. Effect of geniposide on cell viability
Exposure to geniposide at concentrations of 10, 25, and 50 µM did not decrease the viability of RAW 264.7 after 24 h and 48 h treatment. Cell viabilities in RAW 264.7 treated for 24 h with geniposide at concentrations of 10, 25, and 50 µM were 112.75 ± 6.78%, 120.37 ± 3.22%, and 126.31 ± 3.25% of the normal group (media only) respectively (Figure 2A); for 48 h treatment, cell viabilities were 104.87 ± 5.32%, 109.26 ± 3.75%, and 112.44 ± 2.45% of the normal group, respectively (Figure 2B).
3.2. Effect of geniposide on the level of hydrogen peroxide, NO, and Ca2+
Geniposide significantly inhibited the production of hydrogen peroxide in LPS-stimulated RAW 264.7 for 24 h and 48 h treatment (Figure 3A and Figure 3B). Geniposide significantly inhibited NO levels in LPS-stimulated RAW 264.7 (IC50: 135.9 µM) (Figure 3C). Geniposide also significantly inhibited Ca2+ release in LPS-stimulated RAW 264.7 (IC50: 503.5 µM) (Figure 3D). Baicalein, a flavonoid and aglycon hydrolyzed from baicalin, also inhibited hydrogen peroxide generation, NO production, and Ca2+ release in LPS-stimulated RAW 264.7. Considering the inhibitory effect of baicalein, it is estimated that the inhibitory effect of geniposide on Ca2+ release in LPS-stimulated RAW 264.7 is also related to the anti-inflammatory effect of geniposide.
3.3. Effect of geniposide on cytokines production
Geniposide significantly decreased the production of IL-6, TNF-α, G-CSF, GM-CSF, IP-10, LIX, MCP-1, MIP-1α, and MIP-1β in LPS-stimulated RAW 264.7 (Figure 4).
Data revealed that geniposide exhibited IC50 values of 1454, 310.3, 1289, 65.55, 128.6, 925.8, 91.08, 846.2, 1949, and 2569 µM for IL-6, TNF-α, G-CSF, GM-CSF, IP-10, LIX, MCP-1, MIP-1α, MIP-1β, and MIP-2, respectively. Data suggest that geniposide exhibits anti-inflammatory effects in LPS-stimulated RAW 264.7 by reducing various cytokines, leading to amelioration of the hyper-inflammatory syndrome, known as cytokine storm, caused by endotoxemia. The already known inhibitory effect of baicalein on cytokine productions could also be confirmed.
3.6. Effect of geniposide on inflammatory target genes expressions
Geniposide significantly the transcription of Chop, Jak2, Fas, c-Jun, c-Fos, Stat-3, Nos2, Ptgs2, Gadd34, Asc, Xbp1, Nlrp3, and Par-2 genes (Figure 5 and Figure 6). Data mean that geniposide on inhibits the production of inflammatory mediators via ERS/CHOP-related pathway. The already known inhibitory effect of baicalein on expressions of ERS-related genes could also be confirmed.
3.7. Effect of geniposide on P38 MAPK phosphorylation
Geniposide meaningfully decreased the level of phosphorylated P38 MAPK in LPS-stimulated RAW 264.7 (Figure 7). These data indicate that geniposide exerts anti-inflammatory effect on LPS-provoked RAW 264.7 via calcium-P38 MAPK pathway. The already known inhibitory effect of baicalein on P38 activation could also be confirmed.
4. Discussion
Research on natural products for treating various inflammatory diseases continues [21]. Geniposide is a major constituent of the herb gardenia, which is known to treat inflammatory disorder, fever, hypertension, edema, jaundice, and dysphoria for centuries [13,14,22]. Since geniposide is a major component of gardenia, if the inhibitory effect of geniposide on infectious inflammatory reactions is revealed in more detail, it will help develop anti-inflammatory substances using gardenia or develop new treatments to prevent worsening inflammatory diseases caused by infection.
Shi et al. have already reported that geniposide inhibits nitric oxide synthase in LPS-provoked macrophages as well as cyclooxygenase-2 [14]. Interesting reports have been made about anti-asthmatic property of geniposide [23], anti-tumor activity of geniposide [24], anti-angiogenic activity of the herb gardenia [25], and anti-inflammatory activity of geniposide in oxygen/glucose-deprived rat microglial cells [26]. Liu et al. have reported that geniposide inhibits IL-6 and IL-8 production in LPS-induced human umbilical vein endothelial cells by blocking the P38 and ERK 1/2 signaling [27]. But the complete mechanism for bioactivity of geniposide still remains unresolved.
LPS can stimulate immune cells like macrophages to secrete various inflammatory factors including NO, cytokines, prostaglandins, and reactive oxygen species. Excessive production of inflammatory mediators in infectious diseases might have a serious outcome in patients. The production of large amounts of inflammatory factors from activated macrophages is associated with exacerbation of sepsis [14,28]. Thus, LPS-provoked macrophages is regarded as being useful to search a new anti-inflammatory agent.
Although many studies for evaluating pharmacological activities of geniposide have been reported, effects of geniposide on cytokines secreted from LPS-provoked macrophages have not been fully reported till now. Therefore, effects of geniposide on ERS-related cytokine production in LPS-provoked RAW 264.7 was investigated in this study.
The uncontrolled inflammation can develop into acute inflammatory diseases and further chronic inflammatory diseases such as diabetes [29]. Sepsis is a representative disease concerned with the uncontrolled inflammation and the hyper-inflammatory response (cytokine storm). Hu et al. have reported that sepsis is a systemic response that results from a harmful host response to infection [30].
Sun and Bhatia reported that MIP-1α, MCP-1, and MIP-2 are elevated after acute pancreatitis induced by cerulein in mouse model and they also suggested that MIP-1α, MCP-1, and MIP-2 have a role in the pathogenesis of acute pancreatitis [31]. IL-6, an important factor of B-cell maturation and autoantibody production, induces the articular and systemic symptoms of rheumatoid arthritis [32]. MCP-1 is hypersecreted in airway hyper reactivity and chronic airway inflammatory diseases such as asthma [33]. The exaggerated allergic inflammation with the airway hyper-responsiveness can be accompanied by an increase in G-CSF and KC [34]. GM-CSF and IL-5 have clinical importance in the pathophysiology of allergy and asthma [35]. During lung inflammation, levels of MCP-1, IP-10, G-CSF, and GM-CSF are increased in bronchoalveolar fluid [36]. In the current study, geniposide at concentrations of 10, 25, and 50 μM meaningfully suppressed the production of IL-6, G-CSF, MCP-1 and MIP-1α in LPS-stimulated RAW 264.7 and geniposide at concentrations of 25 and 50 μM meaningfully suppressed levels of TNF-alpha, IP-10, GM-CSF, and MIP-1β. These experimental results mean that various hyper-inflammatory phenomena caused by endotoxins can be alleviated by geniposide. More research will be needed on which inflammatory diseases geniposide will have a valid effect.
Interestingly, the current data showed that geniposide significantly inhibits levels of intracellular Ca2+, NO, and hydrogen peroxide in LPS-provoked RAW 264.7; geniposide decreased significantly the transcription of Chop, Jak2, Fas, c-Jun, c-Fos, Stat-3, Nos2, Ptgs2, Gadd34, Asc, Xbp1, Nlrp3, and Par-2 genes as well as P38 MAPK phosphorylation. LPS stimulation is well known to makes RAW 264.7 produce cytokines and ROS via P38 MAPK activation [37]. Because ROS-mediate up-regulation of TACE activity in LPS-stimulated monocytes might be progressed via P38 MAPK activation [10], inhibitory effects of geniposide on ROS production and expressions of ERS-related genes in LPS-stimulated RAW 264.7 could be interpreted as that geniposide inhibits cytokine production, such as TNF-α and MCP-1, through regulation of ERS-related P38 signaling (Figure 8).
CHOP (GADD153) in stressed cells turns out to be activated by P38 MAPK [38]. LPS causes an ERS with the overexpression of Chop [39]. During inflammation, a characteristic phenomenon occurs in which intracellular calcium concentration increases while ER calcium storage decreases [40]. CHOP-amplified cytoplasmic calcium is known to activate ROS generation via Camk2a activation [41]. ERS-induced calcium release might be related with Camk2a and Stat1 activation in macrophages [42]. As well as CHOP, GADD34 [11,12], XBP1 [13], and PAR-2 [14] might also be up-regulated through P38 MAPK phosphorylation in ERS reaction [11,12,13,14]. Asc and Nlrp3 play an important role in the inflammasome complex activation caused by infectious pathogen and in infection-induced macrophage activation [43]. Interestingly, the induction of the ASC and Nlrp3 inflammasome plays an intermediate role in a series of inflammatory cascade from ERS to pyroptosis, which begins with ROS production and subsequently results in cellular injury [44]. In this study, it was not possible to determine whether XBP1 directly activates the transcription factor activity of CHOP. Since oxidative stress and ERS can cause each other, additional investigation is needed to determine whether geniposide suppresses ERS through regulation of oxidative stress, or whether it suppresses oxidative stress through regulation of ERS. These results mean that geniposide could modulate M1-type polarization in LPS-stimulated macrophages, which might be induced with ERS in the bacterial infection [45]. Further studies need to verify effects of geniposide on the conversion of M1 to M2 macrophages. Finally, geniposide significantly inhibited levels of NO, ROS, IL-6, TNF-α, G-CSF, GM-CSF, IP-10, MCP-1, MIP-1α, and MIP-1β in LPS-stimulated RAW 264.7 via calcium-P38 MAPK pathway. The shortfall of this study was the inability to use primary macrophages. Another shortcoming of this study is the inability to confirm protein levels such as CHOP, STAT-3, PTGS2, NOS2, GADD34, XBP1, PAR-2, Asc, and Nlrp3 as well as the phosphorylation of STATs. More research is needed to elucidate the medicinal benefits of geniposide for inflammatory disorders.
5. Conclusions
Geniposide significantly inhibited levels of Ca2+, NO, H2O2, IL-6, TNF-α, G-CSF, GM-CSF, IP-10, MCP-1, MIP-1α, MIP-1β, and phosphorylated P38 MAPK as well as mRNA expressions of Chop, Jak2, Fas, c-Jun, c-Fos, Stat3, Nos2, Ptgs2, Gadd34, Asc, Xbp1, Nlrp3, and Par-2. in LPS-stimulated RAW 264.7, which means that anti-inflammatory activity of geniposide in macrophages takes place through calcium-P38 MAPK signaling pathway.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea, funded by the Ministry of Science, ICT and Future Planning (2017R1A2B4004933).
Acknowledgments
The author thanks Ji-Young Lee and Young-Jin Kim (College of Korean Medicine, Gachon University) for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage Polarization in Inflammatory Diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Lariccia, V.; Magi, S.; Serfilippi, T.; Toujani, M.; Gratteri, S.; Amoroso, S. Challenges and Opportunities from Targeting Inflammatory Responses to SARS-CoV-2 Infection: A Narrative Review. J. Clin. Med. 2020, 9. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine Storm and Leukocyte Changes in Mild Versus Severe SARS-CoV-2 Infection: Review of 3939 COVID-19 Patients in China and Emerging Pathogenesis and Therapy Concepts. J. Leukoc. Biol. 2020, 108, 17–41. [Google Scholar] [CrossRef]
- Roy, R.K.; Sharma, U.; Wasson, M.K.; Jain, A.; Hassan, M.I.; Prakash, H. Macrophage Activation Syndrome and COVID 19: Impact of MAPK Driven Immune-Epigenetic Programming by SARS-Cov-2. Front. Immunol. 2021, 12, 763313. [Google Scholar] [CrossRef] [PubMed]
- Bacchiega, B.C.; Bacchiega, A.B.; Usnayo, M.J.; Bedirian, R.; Singh, G.; Pinheiro, G.D. Interleukin 6 Inhibition and Coronary Artery Disease in a High-Risk Population: A Prospective Community-Based Clinical Study. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Little, P.J.; Xu, S.; Kamato, D. Curcumin Inhibits Lysophosphatidic Acid Mediated MCP-1 Expression Via Blocking ROCK Signalling. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Schett, G.; Neurath, M.F. Resolution of Chronic Inflammatory Disease: Universal and Tissue-Specific Concepts. Nat. Commun. 2018, 9, 3261-018-05800-6. [Google Scholar] [CrossRef]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A Guiding Map for Inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, Y.; Zhou, R.; Li, Y.; Gao, Y.; Tu, D.; Wilson, B.; Song, S.; Feng, J.; Hong, J.S.; et al. A Novel Role of NLRP3-Generated IL-1beta in the Acute-Chronic Transition of Peripheral Lipopolysaccharide-Elicited Neuroinflammation: Implications for Sepsis-Associated Neurodegeneration. J. Neuroinflammation 2020, 17, 64-020-1728-5. [Google Scholar] [CrossRef]
- Scott, A.J.; O’Dea, K.P.; O’Callaghan, D.; Williams, L.; Dokpesi, J.O.; Tatton, L.; Handy, J.M.; Hogg, P.J.; Takata, M. Reactive Oxygen Species and p38 Mitogen-Activated Protein Kinase Mediate Tumor Necrosis Factor Alpha-Converting Enzyme (TACE/ADAM-17) Activation in Primary Human Monocytes. J. Biol. Chem. 2011, 286, 35466–35476. [Google Scholar] [CrossRef]
- Emadali, A.; Nguyen, D.T.; Rochon, C.; Tzimas, G.N.; Metrakos, P.P.; Chevet, E. Distinct Endoplasmic Reticulum Stress Responses are Triggered during Human Liver Transplantation. J. Pathol. 2005, 207, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Oh-Hashi, K.; Maruyama, W.; Isobe, K. Peroxynitrite Induces GADD34, 45, and 153 VIA p38 MAPK in Human Neuroblastoma SH-SY5Y Cells. Free Radic. Biol. Med. 2001, 30, 213–221. [Google Scholar] [CrossRef]
- Zhou, C.M.; Luo, L.M.; Lin, P.; Pu, Q.; Wang, B.; Qin, S.; Wu, Q.; Yu, X.J.; Wu, M. Annexin A2 Regulates Unfolded Protein Response Via IRE1-XBP1 Axis in Macrophages during P. Aeruginosa Infection. J. Leukoc. Biol. 2021, 110, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Boileau, C.; Amiable, N.; Martel-Pelletier, J.; Fahmi, H.; Duval, N.; Pelletier, J.P. Activation of Proteinase-Activated Receptor 2 in Human Osteoarthritic Cartilage Upregulates Catabolic and Proinflammatory Pathways Capable of Inducing Cartilage Degradation: A Basic Science Study. Arthritis Res. Ther. 2007, 9, R121. [Google Scholar] [CrossRef]
- Koo, H.J.; Lim, K.H.; Jung, H.J.; Park, E.H. Anti-Inflammatory Evaluation of Gardenia Extract, Geniposide and Genipin. J. Ethnopharmacol. 2006, 103, 496–500. [Google Scholar] [CrossRef]
- Shi, Q.; Cao, J.; Fang, L.; Zhao, H.; Liu, Z.; Ran, J.; Zheng, X.; Li, X.; Zhou, Y.; Ge, D. et al. Geniposide Suppresses LPS-Induced Nitric Oxide, PGE2 and Inflammatory Cytokine by Downregulating NF-kappaB, MAPK and AP-1 Signaling Pathways in Macrophages. Int. Immunopharmacol. 2014, 20, 298–306. [Google Scholar] [CrossRef]
- Xiaofeng, Y.; Qinren, C.; Jingping, H.; Xiao, C.; Miaomiao, W.; Xiangru, F.; Xianxing, X.; Meixia, H.; Jing, L.; Jingyuan, W.; et al. Geniposide, an Iridoid Glucoside Derived from Gardenia Jasminoides, Protects Against Lipopolysaccharide-Induced Acute Lung Injury in Mice. Planta Med. 2012, 78, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Zhou, F.; Xu, Y.; Liu, X.; Zhang, Y.; Gu, M.; Su, Z.; Zhao, D.; Zhang, L.; Jia, Y. Geniposide Regulates the miR-101/MKP-1/p38 Pathway and Alleviates Atherosclerosis Inflammatory Injury in ApoE(-/-) Mice. Immunobiology 2019, 224, 296–306. [Google Scholar] [CrossRef]
- Jin, Z.; Li, J.; Pi, J.; Chu, Q.; Wei, W.; Du, Z.; Qing, L.; Zhao, X.; Wu, W. Geniposide Alleviates Atherosclerosis by Regulating Macrophage Polarization Via the FOS/MAPK Signaling Pathway. Biomed. Pharmacother. 2020, 125, 110015. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, J.Y.; Kim, Y.J.; Kim, H.J.; Park, W. Rubi Fructus Water Extract Alleviates LPS-Stimulated Macrophage Activation Via an ERS-Induced Calcium/CHOP Signaling Pathway. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Hsieh, S.C.; Fang, S.H.; Rao, Y.K.; Tzeng, Y.M. Inhibition of Pro-Inflammatory Mediators and Tumor Cell Proliferation by Anisomeles Indica Extracts. J. Ethnopharmacol. 2008, 118, 65–70. [Google Scholar] [CrossRef]
- Tseng, T.H.; Chu, C.Y.; Huang, J.M.; Shiow, S.J.; Wang, C.J. Crocetin Protects Against Oxidative Damage in Rat Primary Hepatocytes. Cancer Lett. 1995, 97, 61–67. [Google Scholar] [CrossRef]
- Liaw, J.; Chao, Y.C. Effect of in Vitro and in Vivo Aerosolized Treatment with Geniposide on Tracheal Permeability in Ovalbumin-Induced Guinea Pigs. Eur. J. Pharmacol. 2001, 433, 115–121. [Google Scholar] [CrossRef]
- Lee, M.J.; Hsu, J.D.; Wang, C.J. Inhibition of 12-O-Tetradecanoylphorbol-13-Acetate-Caused Tumor Promotion in Benzoa.Pyrene-Initiated CD-1 Mouse Skin by Geniposide. Anticancer Res. 1995, 15, 411–416. [Google Scholar]
- Park, E.H.; Joo, M.H.; Kim, S.H.; Lim, C.J. Antiangiogenic Activity of Gardenia Jasminoides Fruit. Phytother. Res. 2003, 17, 961–962. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hou, J.; Zhang, P.; Li, D.; Zhang, C.; Liu, J. Geniposide Reduces Inflammatory Responses of Oxygen-Glucose Deprived Rat Microglial Cells Via Inhibition of the TLR4 Signaling Pathway. Neurochem. Res. 2012, 37, 2235–2248. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; He, J.L.; Li, W.M.; Yang, Z.; Wang, Y.X.; Yin, J.; Du, Y.G.; Yu, C. Geniposide Inhibits Interleukin-6 and Interleukin-8 Production in Lipopolysaccharide-Induced Human Umbilical Vein Endothelial Cells by Blocking p38 and ERK1/2 Signaling Pathways. Inflamm. Res. 2010, 59, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A. Management of Sepsis. N. Engl. J. Med. 2006, 355, 1699–1713. [Google Scholar] [CrossRef]
- Spite, M.; Norling, L.V.; Summers, L.; Yang, R.; Cooper, D.; Petasis, N.A.; Flower, R.J.; Perretti, M.; Serhan, C.N. Resolvin D2 is a Potent Regulator of Leukocytes and Controls Microbial Sepsis. Nature 2009, 461, 1287–1291. [Google Scholar] [CrossRef]
- Hu, Z.; Murakami, T.; Suzuki, K.; Tamura, H.; Kuwahara-Arai, K.; Iba, T.; Nagaoka, I. Antimicrobial Cathelicidin Peptide LL-37 Inhibits the LPS/ATP-Induced Pyroptosis of Macrophages by Dual Mechanism. PLoS One 2014, 9, e85765. [Google Scholar] [CrossRef]
- Sun, J.; Bhatia, M. Blockade of Neurokinin-1 Receptor Attenuates CC and CXC Chemokine Production in Experimental Acute Pancreatitis and Associated Lung Injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G143–53. [Google Scholar] [CrossRef] [PubMed]
- Dayer, J.M.; Choy, E. Therapeutic Targets in Rheumatoid Arthritis: The Interleukin-6 Receptor. Rheumatology (Oxford) 2010, 49, 15–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, C.B.; Cardell, L.O. Long-Term Exposure to IL-1beta Enhances Toll-IL-1 Receptor-Mediated Inflammatory Signaling in Murine Airway Hyperresponsiveness. Eur. Cytokine Netw. 2009, 20, 148–156. [Google Scholar] [PubMed]
- Sheller, J.R.; Polosukhin, V.V.; Mitchell, D.; Cheng, D.S.; Peebles, R.S.; Blackwell, T.S. Nuclear Factor Kappa B Induction in Airway Epithelium Increases Lung Inflammation in Allergen-Challenged Mice. Exp. Lung Res. 2009, 35, 883–895. [Google Scholar] [CrossRef]
- Brahmachari, S.; Pahan, K. Role of Cytokine p40 Family in Multiple Sclerosis. Minerva Med. 2008, 99, 105–118. [Google Scholar]
- Srivastava, M.; Jung, S.; Wilhelm, J.; Fink, L.; Buhling, F.; Welte, T.; Bohle, R.M.; Seeger, W.; Lohmeyer, J.; Maus, U.A. The Inflammatory Versus Constitutive Trafficking of Mononuclear Phagocytes into the Alveolar Space of Mice is Associated with Drastic Changes in their Gene Expression Profiles. J. Immunol. 2005, 175, 1884–1893. [Google Scholar] [CrossRef]
- Liu, H.; Xu, R.; Feng, L.; Guo, W.; Cao, N.; Qian, C.; Teng, P.; Wang, L.; Wu, X.; Sun, Y. et al. A Novel Chromone Derivative with Anti-Inflammatory Property Via Inhibition of ROS-Dependent Activation of TRAF6-ASK1-p38 Pathway. PLoS One 2012, 7, e37168. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Z.; Ron, D. Stress-Induced Phosphorylation and Activation of the Transcription Factor CHOP (GADD153) by p38 MAP Kinase. Science 1996, 272, 1347–1349. [Google Scholar] [CrossRef]
- Endo, M.; Mori, M.; Akira, S.; Gotoh, T. C/EBP Homologous Protein (CHOP) is Crucial for the Induction of Caspase-11 and the Pathogenesis of Lipopolysaccharide-Induced Inflammation. J. Immunol. 2006, 176, 6245–6253. [Google Scholar] [CrossRef]
- Stout, B.A.; Melendez, K.; Seagrave, J.; Holtzman, M.J.; Wilson, B.; Xiang, J.; Tesfaigzi, Y. STAT1 Activation Causes Translocation of Bax to the Endoplasmic Reticulum during the Resolution of Airway Mucous Cell Hyperplasia by IFN-Gamma. J. Immunol. 2007, 178, 8107–8116. [Google Scholar] [CrossRef]
- Tabas, I.; Seimon, T.; Timmins, J.; Li, G.; Lim, W. Macrophage Apoptosis in Advanced Atherosclerosis. Ann. N. Y. Acad. Sci. 2009, 1173 Suppl 1, E40-5. [Google Scholar] [CrossRef]
- Timmins, J.M.; Ozcan, L.; Seimon, T.A.; Li, G.; Malagelada, C.; Backs, J.; Backs, T.; Bassel-Duby, R.; Olson, E.N.; Anderson, M.E.; et al. Calcium/calmodulin-Dependent Protein Kinase II Links ERS with Fas and Mitochondrial Apoptosis Pathways. J. Clin. Invest. 2009, 119, 2925–2941. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.B.; Dorfleutner, A.; Rojanasakul, Y.; Stehlik, C. Activation of Inflammasomes Requires Intracellular Redistribution of the Apoptotic Speck-Like Protein Containing a Caspase Recruitment Domain. J. Immunol. 2009, 182, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- Menu, P.; Mayor, A.; Zhou, R.; Tardivel, A.; Ichijo, H.; Mori, K.; Tschopp, J. ER Stress Activates the NLRP3 Inflammasome Via an UPR-Independent Pathway. Cell. Death Dis. 2012, 3, e261. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Yi, M.H.; Choi, J.A.; Lee, J.; Han, J.Y.; Jo, S.H.; Oh, S.M.; Cho, H.J.; Kim, D.W.; Kang, M.W.; et al. Roles of Endoplasmic Reticulum Stress-Mediated Apoptosis in M1-Polarized Macrophages during Mycobacterial Infections. Sci. Rep. 2016, 6, 37211. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Effect of geniposide on cell viability for 24 h (A) and 48 h (B) treatment in RAW 264.7. Nor, normal group (media only). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. Values are mean ± SD of three independent experiments (n=9). Significant differences were examined using Kruskal-Wallis test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor.
Figure 2.
Effect of geniposide on cell viability for 24 h (A) and 48 h (B) treatment in RAW 264.7. Nor, normal group (media only). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. Values are mean ± SD of three independent experiments (n=9). Significant differences were examined using Kruskal-Wallis test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor.

Figure 3.
Effect of geniposide on hydrogen peroxide production for 24 h (A) and 48 h (B) treatment in lipopolysaccharide (LPS)-stimulated RAW 264.7 as well as NO production for 24 h treatment (C) and calcium release for 18 h treatment (D). Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.
Figure 3.
Effect of geniposide on hydrogen peroxide production for 24 h (A) and 48 h (B) treatment in lipopolysaccharide (LPS)-stimulated RAW 264.7 as well as NO production for 24 h treatment (C) and calcium release for 18 h treatment (D). Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.

Figure 4.
Effect of geniposide on the production of IL-6 (A), TNF-α (B), G-CSF (C), GM-CSF (D), IP-10 (E), MCP-1 (F), MIP-1α (G), MIP-1β (H), MIP-2 (I), and LIX (J) in LPS-stimulated RAW 264.7. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.
Figure 4.
Effect of geniposide on the production of IL-6 (A), TNF-α (B), G-CSF (C), GM-CSF (D), IP-10 (E), MCP-1 (F), MIP-1α (G), MIP-1β (H), MIP-2 (I), and LIX (J) in LPS-stimulated RAW 264.7. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.

Figure 5.
Effect of geniposide on the mRNA expression of Chop (A), Jak2 (B), Fas (C), c-Jun (D), c-Fos (E), and Stat-1 (F) in LPS-stimulated RAW 264.7. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. # p < 0.05 vs. Nor; ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.
Figure 5.
Effect of geniposide on the mRNA expression of Chop (A), Jak2 (B), Fas (C), c-Jun (D), c-Fos (E), and Stat-1 (F) in LPS-stimulated RAW 264.7. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. # p < 0.05 vs. Nor; ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.

Figure 6.
Effect of geniposide on the mRNA expression of Nos2 (A), Ptgs2 (B), Gadd34 (C), Asc (D), Xbp1 (E), Stat-3 (F), Nlrp3 (G), and Par-2 (H) in LPS-stimulated RAW 264.7. Nor means the normal group treated with media only. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. # p < 0.05 vs. Nor; ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.
Figure 6.
Effect of geniposide on the mRNA expression of Nos2 (A), Ptgs2 (B), Gadd34 (C), Asc (D), Xbp1 (E), Stat-3 (F), Nlrp3 (G), and Par-2 (H) in LPS-stimulated RAW 264.7. Nor means the normal group treated with media only. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone). GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. BA denotes baicalein (25 µM). Values are mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. # p < 0.05 vs. Nor; ## p < 0.01 vs. Nor; ### p < 0.001 vs. Nor; * p < 0.05 vs. Con; ** p < 0.01 vs. Con; *** p < 0.001 vs. Con.

Figure 7.
Effects of geniposide on the level of phosphorylated P38 MAPK in lipopolysaccharide (LPS)-stimulated RAW 264.7. The level of phosphorylated P38 MAPK was measured with Flow Cytometric Analysis. Values are the mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone); BA, baicalein (25 µM); p-P38, phosphorylated P38 MAPK. GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. # p < 0.05 vs. Nor; *** p < 0.001 vs. Con; ** p < 0.01 vs. Con.
Figure 7.
Effects of geniposide on the level of phosphorylated P38 MAPK in lipopolysaccharide (LPS)-stimulated RAW 264.7. The level of phosphorylated P38 MAPK was measured with Flow Cytometric Analysis. Values are the mean ± SD of three independent experiments. Significant differences were examined using one-way analysis of variance test followed by Tukey’s multiple comparison test. Nor, normal group (media only); Con, control group (1 µg/mL of LPS alone); BA, baicalein (25 µM); p-P38, phosphorylated P38 MAPK. GN10, GN25, and GN50 indicate 10, 25, and 50 µM of geniposide, respectively. # p < 0.05 vs. Nor; *** p < 0.001 vs. Con; ** p < 0.01 vs. Con.

Figure 8.
A diagram for effects of geniposide on lipopolysaccharide-provoked RAW 264.7 through calcium-P38 MAPK signaling.
Figure 8.
A diagram for effects of geniposide on lipopolysaccharide-provoked RAW 264.7 through calcium-P38 MAPK signaling.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



