
Submitted:
28 December 2023
Posted:
28 December 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Studies report variable effects of sex hormones on serious diseases. Severe disease and mortality rates in COVID-19 show marked gender differences that may be related to sex hormones. Sex hormones regulate the expression of the viral receptors ACE2 and TMPRSS2, which affect the extent of viral infection and consequently cause variable outcomes. In addition, sex hormones have complex regulatory mechanisms that affect the immune response to viruses. These hormones also affect metabolism through obesity, and severe disease can result from complications such as thrombosis. This review presents the latest research on the regulatory functions of hormones in viral receptors, immune responses, complications as well as their role in COVID-19 progression. It also discusses the therapeutical possibilities of these hormones by reviewing the recent findings of clinical and assay studies.
Keywords:
SARS-CoV-2
; COVID-19
; Hormone
; Gender
; Immune response
1. Introduction
The coronavirus disease 2019 (COVID-19), attributed to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has led to more than 6 million fatalities. (https://coronavirus.jhu.edu/map.html). Studies have shown notable disparities in COVID-19 mortality and critical illness rates between genders. Early reports from the Chinese Center for Disease Control and Prevention (China CDC) indicated a higher mortality rate for males at 2.8% compared to 1.7% for females [1]. Subsequent large population-based studies and meta-analyses have consistently confirmed this result. Although a global meta-analysis involving 3,111,714 reporting cases found no gender disparity in the distribution of COVID-19 diagnoses, men had a significantly increased risk of intensive care unit (ICU) admission (odds ratio, OR = 2.84) and mortality (OR = 1.39) [2]. Moreover, a cohort study involving 18,647 patients revealed that both male gender (adjusted odds ratio, aOR 1.896) and age (aOR 1.065) were linked to heightened risks of ICU admission and all-cause mortality [3]. Intriguingly, observational studies have revealed that specific pregnant women, initially asymptomatic to COVID-19 and testing for SARS-CoV-2 upon admission to obstetric units, experienced worsening symptoms in the immediate postpartum period. It was consistent with the significant drop in hormones seen after giving birth [4,5]. Furthermore, a 2021 study involving 1,902 female COVID-19 patients identified menopause as a notable risk predictor associated with COVID-19 (OR = 1.91 [1.06 ~ 3.46]) [6]. Previous research suggests that gender bias in COVID-19 outcomes becomes more pronounced with age [7]. Aging, childbirth, and menopause bring about substantial alterations in sex hormones, believed to be contributing to the sex bias observed for COVID-19 severity. In line with this hypothesis, there is a relationship among SARS-CoV-2 exposed men with complications of COVID-19 with metabolic and sex hormone imbalances [8,9]. A cohort study demonstrated that testosterone (T), estradiol (E2), and insulin-like growth factor 1 (IGF-1) concentrations in the blood serum, which are regulated by sex hormone signaling, are linked to the disease burden in patients with COVID-19 [10]. In addition, a high proportion of T to E2 in hospitalized COVID-19 participants was associated with favorable outcomes, as shown in a subsequent cohort study of 3,005 patients from four different research centers [11]. Additionally, a study involving patients with gender anxiety disorder revealed that female-to-male patients treated with T had a 3.46 times higher rate of COVID-19 infection than did male-to-female patients treated with estrogen and anti-androgens. This result provides evidence to confirm the assumption that sex hormones might have a role to do with the development of COVID-19 [12]. Indeed, several previous studies have shown that sex hormones play an important role in the development of infectious and immune-related diseases [13,14,15]. Following the COVID-19 outbreak, more researchers have started to focus on the issue. An increasing evidence base has suggested that sex differences in COVID-19 may be due to gonadal hormonal influences, particularly in studies investigating the effects of oestrogen, progesterone and T on COVID-19 progression. However, the results of these studies have not been consistent.
This review presents the latest research on the regulatory functions of hormones in viral receptors, immune response, complications as well as their role in COVID-19 progression (Figure 1). It also discusses the potential therapy of such hormones based on recent findings in the clinic.
2. Sex Hormones and COVID-19
2.1. Androgens and COVID-19 Severity
Current research has demonstrated that baseline androgen values are linked to adverse outcomes in COVID-19. In addition, low levels of T in patients are connected with organ damage and poor prognosis. However, attempts to test antiandrogenic treatments in clinical trials have not yielded positive results. In a prospective study, it was found that severe COVID-19 patients had lower baseline T levels than those with milder symptoms (53ng/dL vs. 151ng/dL) [10]. In addition, a significant positive association between total circulating T levels assessed in several cohorts prior to contact with SARS-CoV-2 infection and the risk of death from COVID-19 was found in an observational study that controlled for covariates such as lifestyle, underlying diseases, and other variables [10]. It provides added support for the hypothesis of an androgen involvement in COVID-19. Early studies during the COVID-19 outbreak suggested a potential connection between male-pattern baldness, driven by increased androgen activity (androgenetic alopecia), and disease severity [16,17]. It implies a potential link between androgen sensitivity and hospitalization and severe symptoms in persons exposed to SARS-CoV-2. Subsequent observational studies found reduced readmission and mortality rates in prostate cancer patients receiving second-generation androgen deprivation therapies (ADTs) and in men with COVID-19 on long-term use of 5-alpha reductase inhibitors (5ARIs) to treat benign prostatic hyperplasia (BPH) [18,19]. However, clinical trial data provide limited evidence to suggest the benefit of anti-androgen therapy for serious disease associated with COVID-19. A clinical trial investigating the androgen receptor blocker enzalutamide in a limited group was prematurely terminated due to a greater rate of hospitalization among the treatment group than among the placebo control group [20]. Additionally, one clinical study of 96 COVID-19 inpatients found no evidence of reduced COVID-19 severity with the use of the antiandrogenic compound degarelix [21]. Furthermore, similar results were demonstrated in an epidemiologic study of 7,894 prostate cancer patients, where ADT had no preventive effect on outcomes among those patients with COVID-19 [20]. Multiple recent observational trials have refuted the link between elevated androgen activity and T replacement therapy and adverse COVID-19 outcomes [22,23]. In fact, several studies have suggested that low serum T levels may indicate organ damage and prognosis in patients infected with SARS-CoV-2 and may characterize the hormonal milieu of critically ill patients [10,24,25,26]. It may be attributed to the significant reduction in T levels caused by contracting SARS-CoV-2 [27]. One potential coherent trial found a significant reduction in T levels in critically ill men compared to mildly ill men. Upon admission, there was a 64.9% decrease in T levels, followed by an 84.1% decrease on three days [10]. In a cohort study published last year, levels of T were lower among severely symptomatic patients with COVID-19 than among mild to moderately symptomatic hospitalized patients (85.1 ng/dl vs. 315 ng/dl), and lower among those requiring ICU care than among non-ICU patients (64 ng/dl vs. 286 ng/dl) [24]. In summary, circulating androgen levels suggest a complex relationship with COVID-19 severity that has not been completely elucidated. More research is needed to elucidate potential effects of androgen on disease outcome [28].
2.2. Estradiol and COVID-19 Severity
E2 has been linked to reduced levels of inflammation in the context of acute infectious disease and play a major rule in modulating both innate and adaptive immunity [13]. Post-menopausal females are at increased hospitalization risk relative to nonmenopausal females, according to a prospective analysis of medical records from 1,902 COVID-19 patients (OR=1.91) [6]. Moreover, anti-Müllerian hormone (AMH) and E2 might be considered possible preventive agents against the severity of COVID-19, showing a negative correlation [6]. Multiple studies have provided evidence that supplemental estrogen confers a preventive effect on COVID-19 in postmenopausal females, with particular emphasis on the effectiveness of E2 [7,29]. A significant population-based study further supports the hypothesis of a preventive effect of estrogen in COVID-19. This study discovered that antiestrogen therapies (AETs) decreased the occurrence of serious illness in patients suffering from hormone-driven cancers (HDCs) [30]. However, it's worth noting that sex hormone values among severely affected women with COVID-19 did not show statistically significant differences compared to female patients with mild disease [9]. This phenomenon can be attributed in part to the complexity of estrogen and COVID-19 interactions, reflecting the interplay among different subtypes of estrogen, age, and reproductive status [31].
2.3. Progesterone and COVID-19 Severity
Progesterone, in addition, serves as a vital humoral steroid hormone with a significant role in maintaining pregnancies in female mammals [32]. Notably, it is also produced and has functions in males. Progesterone possesses the capability to reduce leukocyte activation, mitigate pro-inflammatory mediator production, regulate T-cell differentiation, contribute to neurodevelopment, and yield a wide array of anti-inflammatory effects [33,34,35,36]. Progesterone has been identified as a crucial factor in facilitating rapid recovery from influenza A virus infection, prompting discussions about its potential advantages during instances of immune dysregulation in SARS-CoV-2 infection [37]. Furthermore, regarding progesterone's potential benefits associated with the infection, clinical trial data from 2021 offered additional support for the hypothesis. These trials revealed that subcutaneous delivery of progesterone improved the condition in 20 severely affected male patients diagnosed with SARS-CoV-2 [38]. Moreover, the results suggested that progesterone might have the capacity to modulate Immune dysfunction., mitigate Strong signs, and reduce the likelihood of critical illnesses. However, it's essential to note that the significance of the findings was constrained by the marginally significant p-value and a limited number of samples, which made it difficult to draw conclusions [38]. This hypothesis received added support from experimental studies showing that progesterone, when administered in a dose-dependent manner, could reverse weight loss and ameliorate severe pneumonia in SARS-CoV-2-infected hamsters [39]. Furthermore, current data from China indicates that women who are becoming pregnant are expected to suffer the most serious instances from COVID-19 after childbirth, which coincides with a rapid decline in progesterone levels [40]. According to the U.S. Centers on Disease Control and Prevention (CDC) monitoring database, among 8,207 female pregnancies in early stages of an outbreak, there was an increased chance of serious disease among the pregnant patients, although the risk of death was similar to that of non-pregnant women [41]. However, it's important to note that some small-scale independent reports among female pregnancies exposed to SARS-CoV-2 did not indicate any adverse pregnancy and/or COVID-19 outcomes [42,43]. In summary, the evidence for a direct association of progesterone with adverse outcomes in COVID-19 remains inconclusive at this time.
3. Sex Hormones and Expression of Viral Receptors
3.1. ACE2 and TMPRSS
The COVID-19 pathogen, SARS-CoV-2, forms a viral envelope with spike glycoproteins (S-proteins) and is a sensory RNA virus [44]. For entry, SARS-CoV-2 relies on surface proteins on the human respiratory tract epithelial cells, particularly transmembrane serine protease 2 (TMPRSS2) as well as angiotensin-converting enzyme 2 (ACE2) [45,46,47] (Figure 1). TMPRSS2 triggers cleavage and activation of the S proteins, resulting in viral-host cell membrane fusing [48]. Intracellular entry is facilitated through ACE2, a type I transmembrane glycoprotein acting as a terminal carboxypeptidase. The two types of proteins differ in their expression levels across several tissues, with the highest expression levels in lung type 1 and type 2 alveolar epithelial cells (AT1 and AT2) [49,50]. TMPRSS2 expression was identified in various lung and bronchial cells, demonstrating higher levels in AT2 cells compared to AT1 cells. ACE2 is normally expressed in the lungs [49], oral mucosa [51], cardiovascular system [52], testes [53], and immune cells [54].
During the course of SARS-CoV-2 disease, excessive levels of ACE2 could facilitate the entry of the virus into the body, potentially leading to organ damage. Experimental studies demonstrate that the removal of respiratory TMPRSS2 affects the primary infection site and the spread of the virus in the airways, ultimately reducing the degree of lung pathology following SARS-CoV and MERS-CoV infection [55]. Various types of TMPRSS2 blockers were found to prevent SARS-CoV-2 entry into the virus in culture [56,57,58]. Research indicates upper airway SARS-CoV-2 transmission involves TMPRSS2 along with closely related proteases, while transmission in human lung may be inhibited by camostat mesylate as well as the metabolite GBPA [59]. During the clinical trial, COVID-19 patients receiving camostat mesylate, a serine protease inhibitor, experienced symptom relief and faster recovery of taste and smell sensation compared to other groups [60]. However, a randomized trial found that adding a hemostat to the usual care regimen did not reduce treatment time, and only a narrow percentage of COVID-19 risk patients who required O2 therapy showed accelerated recovery [61]. Indeed, TMPRSS2 depends on ACE2 for its functional activity in viral fusion. A case-control study determined that the TMPRSS2/ACE2 ratio proved more effective than ACE2 alone in predicting COVID-19 severity [62].
3.2. Sex-Specific Expression of TMPRSS2 and ACE2
ACE2 expression has been found to be increased in men compared to women, both overall and within the lung [50,63]. There are two main causes: sex chromosomes and sex hormones. In females, both X chromosomes are switched off at the end of blastocyst stage to prevent imbalances in genetic expression, but certain genes on the X chromosome, including those encoding ACE2 and angiotensin II receptor 2, escape this silencing [64,65]. Additionally, studies have shown that the SRY family of genes found in the Y sex chromosome of males increases the activation of RAAS components, thereby decreasing the promoter activation of ACE2 [66]. However, studies and experimental models evaluating ACE2 expression across various tissues have yielded conflicting results. Some studies report comparable ACE2 expression levels in various tissues in both women and men [67,68]. In the lung, there is a debate regarding if the levels of TMPRSS2 expression in males are higher than in females, although some research has shown a small significant difference [69,70]. Conversely, other studies have reported a lack of significance in pulmonary phenotypes among the sexes, while indicating a higher level of ACE2 locus in pulmonary and bronchial muscle cells in males [71,72].
3.3. Regulation of TMPRSS2 and ACE2 by Sex Hormones
The intricate interplay between estrogen and ACE2 appears to vary across specific organs or environments (Table 1). Analysis of publicly available genomic data reveals that estrogen amplifies ACE2 expression in thymus cells from mice and adenocarcinoma cells from human lung epithelium [76]. MR antagonists suppress proinflammatory genes while concurrently boosting ACE2 mRNA expression and activity [77]. Elevated concentrations of E2 lead to reduced lung ACE2 expression, whereas lower levels result in an increase [78]. Experimental evidence suggests that estrogen interferes with the glycoprotein and glycol can junctions within ACE2 and SARS-CoV-2, preventing virus insertion peak proteins within alveoli [79]. Conversely, Epithelial cells of the bronchus and smooth muscular tissue cells of respiratory tract cells show a modest downregulation of ACE2 in response to 17β-estradiol [72,73]. Moreover, estrogen appears to diminish ACE2 expression in the myocardium and kidneys in vivo [80]. In comparison, progesterone demonstrates greater efficacy in reducing ACE2 expression in uterine tissue compared to estrogen treatment [81]. Previous animal model studies have reported decreased ACE2 expression and activity with estrogen and estrogen modulators.
In human primary bronchial epithelial cells, estrogen inhibits ACE2 expression without affecting TMPRSS2 expression [73]. In non-pituitary cells, for example, prostate, breast or kidney cells, estrogen may inhibit TMPRSS2 expression [75,82]. Prostate carcinoma containing the TMPRSS2:ERG fusion gene may respond to estrogen signalling[82]. Application of 17β-estradiol treatment on VERO E6 cells results in reduced intracellular SARS-CoV-2 viral load, leading to a decrease in TMPRSS2 mRNA. However, ACE2 mRNA remains unchanged in the same VERO E6 cells [75].
Modulating ACE2 expression is a mechanism through which androgens control viral entry. Androgens raise markers of ACE2 activation, blood plasma renin activation and the expression of angiotensinogen [83]. A recent comprehensive screen of nearly 1,500 drugs approved by the FDA was designed to find compounds that reduce ACE2 in normal cell cultures. The results highlight the androgen signaling pathway as a major regulator of ACE2 levels [8]. Multiple studies have affirmed that T amplifies ACE2 expression. In aging men, declining androgen levels lead to reduced ACE2 expression, while T markedly boosts ACE2 levels in normal male primary airway smooth muscle cells [76]. Treatment with antiandrogen drugs decreases ACE2 expression in cardiomyocytes generated by stem cell-derived cells and protects pulmonary organs from SARS-CoV-2 infection [8]. Androgens and antiandrogens can also regulate ACE2 expression in prostate cancer cell lines [67], corroborated by research on mouse lung tissue [84]. In addition, androgen receptor stimulation enhances ACE2 levels in mouse airway epithelium cells. Exposure to T for 24 hours elevates ACE2 levels on exposed human airway epithelium from both sexes [67,72]. Consequently, a slight downregulation of ACE2 gene levels is found in the lungs of mice treated with the potent anti-androgen enzalutamide [85]. Further, androgen deprivation through antiandrogen treatments, administered via depot or in vitro methods, causes decreased expression of transcripts and proteins of both TMPRSS2 and ACE2 [84].
Androgens influence viral entry by regulating TMPRSS2. Androgens, including T, can enhance TMPRSS2 gene expression, potentially resulting in higher levels in men than in women [69]. Consequently, SARS-CoV-2 can more readily enter target host cells in males than in females [86]. Androgen ligands and androgen receptor promoter binding elements control TMPRSS2 promoter activity in the prostate. TMPRSS2 is high in the prostate tissue and levels increase in response to androgens through targeted androgen receptor expression [87]. Androgen deprivation therapy (ADT) is believed to offer protective effects through preventing the entrance of pseudotyped and live SARS-CoV-2 viruses in prostate cancer cells [67]. In contrast, TMPRSS2 regulation by androgens in lung tissue remains uncertain, unlike in the prostate. Human lung adenocarcinoma cells treated with androgen lines resulted in increased TMPRSS2 expression [88]. Similarly, androgens and anti-androgens regulate TMPRSS2 in various human lung cell lines and mouse lung epithelial cells [70]. Nevertheless, following enzalutamide treatment, TMPRSS2 expression remained unchanged across cancer cell lines, mouse lung and human pulmonary organs [85,89].
4. Sex Hormones and Immune Responses
4.1. Cytokine Storm
SARS-CoV-2 infection triggers the development of innate and adaptive immune responses, often leading to dysregulated innate inflammation and compromised adaptive immunity in critically ill patients. This dysregulation may result in cytokine storm, as demonstrated by a significant increase of pro-inflammatory cytokines in patient serum, including IL-6, IL-1β, IL-2, IL-8, IL-17, G-CSF, GM-CSF, IP10, MCP1, CCL3 and TNF [90,91] (Figure 1). The cytokine storm, in turn, may induce shock, organ damage, respiratory failure, and, in severe cases, multiple organ failure. It is also associated with significant neutrophil and monocyte infiltrates and results in extensive damage to the alveoli, characterized by the formation of hyalinization and alveolar wall thickness. Ultimately, this immune-mediated damage extends to include conditions like splenic atrophy and lymph node necrosis, which have been observed in deceased patients [92].
4.2. Gender Differences in Immune Responses
There are significant sex differences in innate and adaptive immune responses. Typically, immune responses to pathogens are stronger in females compared to males, including female animals. This sexual dimorphism is attributed to various factors. First, females possess a greater abundance of internal defense macrophages, including monocytes, macrophages and dendritic cells [93,94]. Moreover, females exhibit enhanced cytotoxic T-cell activity [95], elevated immunoglobulin levels [96], and increased CD3+ and CD4+ cell counts [97]. Conversely, male COVID-19 patients often have lower lymphocyte counts, elevated neutrophil to leukocyte ratios, and elevated levels of C-reactive plasma protein in serum than women [98]. Additionally, while females typically demonstrate viral infections with stronger cytokine responses, male SARS-CoV-2-infected individuals tend to have elevated proinflammatory cytokines like IL-8, contributing to the poorer prognosis in COVID-19 cases. Furthermore, T-cell activation is generally weaker in males, which correlates with worse outcomes in COVID-19 patients.
4.3. Sex Hormones and the Immune Response
Discrepancies in circulating sex hormone levels may underlie gender-related responses to infections. Progesterone is critical for the release of vital immune response cytokines, including IL-6, TNF-α, and IFN-γ. The hormone also orchestrates immune cell activities and induces potent anti-inflammatory effects [99]. Besides, estrogen upregulates various genes associated with B cell activation and survival, such as CD22, SHP-1, and Bcl-2.119, thus enhancing immune system responses, particularly in B cells [100]. In contrast, T appears to inhibit the recruitment of both eosinophils and neutrophils, interfere with T helper 2 (TH2) CD4+ recruitment and suppress IgE generation [101]. Additionally, T inhibits TH2 and TH17 cell function [102,103] and alters the production of cytokines, including IFNγ [104] resulting in immunosuppressive effects. In summary, these gender-related differences extend from innate to adaptive immunity and are influenced by the interplay of intrinsic and adaptive immune factors, often linked to sex hormone levels.
4.4. Regulation of Disease by sex Hormones Through Immunization
Gender-related disparities in sex hormone-induced immune responses appear to be important for the outcome of COVID-19 therapy (Table 2). Recent research shows that acute respiratory distress syndrome (ARDS) leading to respiratory failure is the leading cause of death in COVID-19 patients [105]. In a 2021 study, males who had reduced T and lymphocyte plasma concentrations on admission to hospital with COVID-19 were more likely to have more severe ARDS and worse condition after admission [106]. Even before the 2020 epidemic, studies had pointed out that low total testosterone (TT) levels, as well as circulating free testosterone (cFT), were related to the emergence of the hyperinflammatory syndrome leading to prolonged hospitalisation after COVID-19 infection [107,108]. Additionally, a retrospective study involving symptomatic men, conducted independently, established a correlation between low serum TT levels and higher estrogen/T ratios, often indicative of systemic inflammation, and a heightened risk of in-hospital mortality [108,109]. Furthermore, two clinical trials, one involving the anti-androgen drug of choice dutasteride, a 5-alpha reductase blocker used to reduce prostate hyperplasia and male baldness in 87 patients, and the other involving, an androgen receptor antagonist, in 236 patients, showed faster viral clear-up, less viral spill-over and lower C-reactive product scores [110,111]. In a clinical evaluation performed in 2022, raloxifene, which is a specific estrogen receptor modulator, increased white blood count and accelerated viral clearance among patients infected with SARS-CoV-2 [112]. Furthermore, there's evidence that progesterone can modulate the sexual dimorphism in SARS-CoV-2 transmission and disease progression, suggesting a therapeutic value of COVID-19. They demonstrated that progesterone can inhibit excessive pro-inflammatory cytokine release and inhibit replication of the virus in the pulmonary tissues of hamsters infected with SARS-CoV-2 [39]. Within pulmonary epithelia, progesterone receptors (PRs) control the production of several proinflammatory cytokines, which also regulate the activation of immune cell s[113,114]. A 2022 animal study Another study, published in the same year, delved deeper into this topic. The authors found that viral infection can induce somatic progesterone through the HPA axis and that progesterone, in turn, can enhance intrinsic antiviral responses in cells and mice through downstream antiviral genes [115].
5. Complications of COVID-19
5.1. Tissue Damage
Severe illness and death from progression of acute viral pneumonia to ARDS are common in COVID-19 patients [116]. These phases encompass exudative and proliferative stages characterized by hyaline membrane changes and microvascular thrombosis, culminating in diffuse alveolar injury [117] (Figure 1). Effective management during these stages is vital for improving patient outcomes and preventing mortality. Specifically, in a lipopolysaccharide model of acute lung damage, males exhibit increased airway hyperreactivity and increased airway obstruction relative to females. Administration of T therapy enhances the inflammatory response in females to levels similar to those observed in males. However, gonadectomy attenuates lung injury in the men rather than the women, indicating androgens maintain inflammatory effects in lipopolysaccharide-induced pulmonary injury [118]. Furthermore, estrogen replacement therapy effectively alleviates the features of pulmonary damage by inhibiting cellular adhesion and inflammatory cytokines, thereby reducing the inflammatory consequences of the disease.
5.2. Cardiovascular System
In the context of COVID-19, patients can experience various extrapulmonary symptoms, making it crucial to promptly diagnose and manage these complications to enhance patient outcomes. Among these complications, over one-third of COVID-19 hospitalized patients encounter cardiovascular issues such as myocardial dysfunction, arrhythmias, acute coronary syndromes, and thrombosis. These complications are highly correlated with mortality and result in direct cardiomyocyte damage, viral-mediated endothelial injury, systemic inflammation and hypoxia [119]. Estrogens exhibit a vasculoprotective effect, which might partly account for the gender disparities in COVID-19 fatalities [120]. Research suggests that 17β-estradiol is protective for ischemic myocardium and reduces infarct size [121]. Through direct membrane signaling, estrogen leads to vasodilation by releasing nitric oxide. Similarly, estrogen receptor signaling preserves the structure and function of endothelial cells by inhibiting the activation of the apoptotic pathway [122]. These findings highlight the cardioprotective effects of estrogen, which could reduce susceptibility to COVID-19-related cardiac injury, endothelial inflammation, and consequent cardiovascular complications [123].
5.3. Visceral Adiposity
Research consistently demonstrates a heightened potential for serious illness and increased mortality among persons with an elevated body mass index (BMI) [124,125,126]. Importantly, adipose tissue expresses ACE2 receptors, and in obese patients with larger fat stores, these receptors are more abundant, intensifying the systemic response to SARS-CoV-2[127]. Conversely, visceral obesity contributes to elevated levels of prothrombotic circulating factors, increasing susceptibility to thrombosis [128]. Fat distribution is also influenced by sex hormones. Knocking down estrogen receptors and reducing estrogen signaling leads to Male and female mouse obesity [129]. In addition, a randomized controlled trial showed the effects of androgen replacement therapy on overall body composition in overweight post-menopausal women, with separate effects on abdominal and visceral fat deposition. It suggests a link between treatments involving anabolic steroids with androgenic properties and an increased accumulation of visceral fat [130]. Notably, the typical distribution of body fat in females has been associated with reduced cardiometabolic risk, reduced systemic activation, and reduced COVID-19 severity. Research suggests a robust correlation between the intensity of COVID-19 and the fat distribution in the abdomen, which is characterized by elevated visceral adiposity (VAT) levels combined with reduced total subcutaneous adiposity (SAT). SAT levels are more likely to increase in females than males and are inversely related to the incidence of severe disease. For each 1-mm increment of SAT thickness, there was a 16% increase in the chance of serious disease [131]. VAT plays a crucial role in the body's defense system, contributing to the production of pro-inflammatory factors such as TNF-α, IL-6 and IL-1β.
It's worth noting that approximately a third of IL-6 is generated by the adipose cells and stroma in adipose tissue [132]. Interestingly, SARS-CoV-2 infection may exacerbate inflammation in VAT. The mesenteric visceral adipose tissue, which surrounds the small intestine, acts as a primary defense against pathogen migration from the intestines to the circulatory system [133]. It's noteworthy that over half of COVID-19 participants with positive fecal SARS-CoV-2 test results have experienced gastroenterological discomfort, reflecting the impact of SARS-CoV-2 in intestinal cells [134]. Single-cell analysis of RNA sequencing data reveals high levels of ACE2 and TMPRSS2 SARS-CoV-2 infectious proteins in intestinal cells [135], suggesting the intestines could potentially serve as an entry point for the virus. When the virus is detected by the intestinal defense mechanism, it can trigger an immune-inflammatory response that spreads to the mesenteric VAT, exacerbating local inflammation. These intricate interactions among sex hormones, immune responses, and immunometabolic factors create an environment that influences COVID-19 progression on multiple levels.
6. Conclusions
In summary, sex hormone level differences contribute in part to the gender variations in SARS-CoV-2 outcomes. Nonetheless, the existing evidence lacks consensus, and the intricate interplay between viruses and sex hormones makes it challenging to determine the precise effect of sexual steroids on neo-coronary remission. The clinical evidence evaluating the impact of sexual hormones on regression of COVID-19 does not support the use of E2 or discontinuation of these agents in patients suffering from COVID-19. Therefore, more prospective trials are required to obtain a full picture of the long-term impact sex hormones have on COVID-19 outcomes. Several pertinent trials have explored the effects of COVID-19 results from sex hormones, including their influence on ACE2 and TMPRSS2 expression, immune responses and complications such as cardiovascular disease. However, these reports yield inconsistent findings, and a thorough exploration of the underlying mechanisms and hypotheses is required, utilizing advanced analytical methods such as multi-omics. Lastly, the impact of sociocultural gender on an individual's life experiences, immunization awareness, and medical conditions indirectly affects COVID-19 outcomes [136,137,138]. Therefore, it is crucial to study both gender-related biological and sociocultural differences to enhance our understanding and management of the pandemic.
Author Contributions
Conceptualization: H.X., X.L., J.L. and C.Z.: conceptualization, investigation, formal analysis, writing—original draft; X.L., G.L. and L.Y.: conceptualization, supervision, writing—reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32201152, 81925019, U1705281, U22A20333), the Major State Basic Research Development Program of China (2017YFA0205201), the Fundamental Research Funds for the Central Universities (20720190088, 20720200019), the Program for New Century Excellent Talents in University, China (NCET-13-0502), Shenzhen Natural Science Foundation (JCYJ20220530143407016). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
All data is referred to in the references.
Conflicts of Interest
The authors declare no conflict of interest.
References
- The Novel Coronavirus Pneumonia Emergency Response Epidemiology Team, n. The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19) - China, 2020. China CDC weekly 2020, 2, 113–122. [Google Scholar] [CrossRef]
- Peckham, H.; de Gruijter, N.M.; Raine, C.; Radziszewska, A.; Ciurtin, C.; Wedderburn, L.R.; Rosser, E.C.; Webb, K.; Deakin, C.T. Male sex identified by global COVID-19 meta-analysis as a risk factor for death and ITU admission. Nature Communications 2020, 11, 6317. [Google Scholar] [CrossRef]
- Cardoso, F.S.; Papoila, A.L.; Machado, R.S.; Fidalgo, P. Age, sex, and comorbidities predict ICU admission or mortality in cases with SARS-CoV2 infection: A population-based cohort study. Critical Care (London, England) 2020, 24, 465. [Google Scholar] [CrossRef]
- Breslin, N.; Baptiste, C.; Miller, R.; Fuchs, K.; Goffman, D.; Gyamfi-Bannerman, C.; D'Alton, M. Coronavirus disease 2019 in pregnancy: Early lessons. American journal of obstetrics & gynecology MFM 2020, 2, 100111. [Google Scholar] [CrossRef]
- Breslin, N.; Baptiste, C.; Gyamfi-Bannerman, C.; Miller, R.; Martinez, R.; Bernstein, K.; Ring, L.; Landau, R.; Purisch, S.; Friedman, A.M.; et al. Coronavirus disease 2019 infection among asymptomatic and symptomatic pregnant women: Two weeks of confirmed presentations to an affiliated pair of New York City hospitals. American journal of obstetrics & gynecology MFM 2020, 2, 100118. [Google Scholar] [CrossRef]
- Ding, T.; Zhang, J.; Wang, T.; Cui, P.; Chen, Z.; Jiang, J.; Zhou, S.; Dai, J.; Wang, B.; Yuan, S.; et al. Potential Influence of Menstrual Status and Sex Hormones on Female Severe Acute Respiratory Syndrome Coronavirus 2 Infection: A Cross-sectional Multicenter Study in Wuhan, China. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America 2021, 72, e240–e248. [Google Scholar] [CrossRef]
- Seeland, U.; Coluzzi, F.; Simmaco, M.; Mura, C.; Bourne, P.E.; Heiland, M.; Preissner, R.; Preissner, S. Evidence for treatment with estradiol for women with SARS-CoV-2 infection. BMC medicine 2020, 18, 369. [Google Scholar] [CrossRef] [PubMed]
- Samuel, R.M.; Majd, H.; Richter, M.N.; Ghazizadeh, Z.; Zekavat, S.M.; Navickas, A.; Ramirez, J.T.; Asgharian, H.; Simoneau, C.R.; Bonser, L.R.; et al. Androgen Signaling Regulates SARS-CoV-2 Receptor Levels and Is Associated with Severe COVID-19 Symptoms in Men. Cell Stem Cell 2020, 27, 876–889.e812. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Schaumburg, B.; Mueller, Z.; Parplys, A.; Jarczak, D.; Roedl, K.; Nierhaus, A.; de Heer, G.; Grensemann, J.; Schneider, B.; et al. High estradiol and low testosterone levels are associated with critical illness in male but not in female COVID-19 patients: A retrospective cohort study. Emerging Microbes & Infections 2021, 10, 1807–1818. [Google Scholar] [CrossRef]
- Dhindsa, S.; Zhang, N.; McPhaul, M.J.; Wu, Z.; Ghoshal, A.K.; Erlich, E.C.; Mani, K.; Randolph, G.J.; Edwards, J.R.; Mudd, P.A.; et al. Association of Circulating Sex Hormones With Inflammation and Disease Severity in Patients With COVID-19. JAMA network open 2021, 4, e2111398. [Google Scholar] [CrossRef] [PubMed]
- Gebhard, C.E.; Hamouda, N.; Gebert, P.; Regitz-Zagrosek, V.; Gebhard, C.; Investigators, C. Sex versus gender-related characteristics: Which predicts clinical outcomes of acute COVID-19? Intensive Care Medicine 2022, 48, 1652–1655. [Google Scholar] [CrossRef]
- Durcan, E.; Turan, S.; Bircan, B.E.; Yaylamaz, S.; Demirel, O.; Demir, A.N.; Sulu, C.; Kara, Z.; Sahin, S.; Taze, S.S.; et al. TransCOVID: Does Gender-Affirming Hormone Therapy Play a Role in Contracting COVID-19? Journal of Sex & Marital Therapy 2022, 48, 415–426. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nature Reviews. Immunology 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Li, C.-L.; Li, C.-Y.; Lin, Y.-Y.; Ho, M.-C.; Chen, D.-S.; Chen, P.-J.; Yeh, S.-H. Androgen Receptor Enhances Hepatic Telomerase Reverse Transcriptase Gene Transcription After Hepatitis B Virus Integration or Point Mutation in Promoter Region. Hepatology (Baltimore, Md.) 2019, 69, 498–512. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-J.; Yeh, S.-H.; Liu, W.-H.; Lin, C.-C.; Huang, H.-C.; Chen, C.-L.; Chen, D.-S.; Chen, P.-J. Androgen pathway stimulates MicroRNA-216a transcription to suppress the tumor suppressor in lung cancer-1 gene in early hepatocarcinogenesis. Hepatology 2012, 56, 632–643. [Google Scholar] [CrossRef]
- A preliminary observation: Male pattern hair loss among hospitalized COVID-19 patients in Spain - A potential clue to the role of androgens in COVID-19 severity - PubMed. Available online: (accessed on.
- Wambier, C.G.; Vaño-Galván, S.; McCoy, J.; Gomez-Zubiaur, A.; Herrera, S.; Hermosa-Gelbard, Á.; Moreno-Arrones, O.M.; Jiménez-Gómez, N.; González-Cantero, A.; Fonda-Pascual, P.; et al. Androgenetic alopecia present in the majority of patients hospitalized with COVID-19: The "Gabrin sign". Journal of the American Academy of Dermatology 2020, 83, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.G.; Zhong, X.; Liaw, B.; Tremblay, D.; Tsao, C.-K.; Galsky, M.D.; Oh, W.K. Does androgen deprivation therapy protect against severe complications from COVID-19? Annals of Oncology: Official Journal of the European Society for Medical Oncology 2020, 31, 1419–1420. [Google Scholar] [CrossRef]
- Lazzeri, M.; Duga, S.; Azzolini, E.; Fasulo, V.; Buffi, N.; Saita, A.; Lughezzani, G.; Paraboschi, E.M.; Hurle, R.; Nobili, A.; et al. Impact of chronic exposure to 5-alpha reductase inhibitors on the risk of hospitalization for COVID-19: A case-control study in male population from two COVID-19 regional centers of Lombardy, Italy. Minerva Urology and Nephrology 2022, 74, 77–84. [Google Scholar] [CrossRef] [PubMed]
- A Phase 2 Trial of the Effect of Antiandrogen Therapy on COVID-19 Outcome: No Evidence of Benefit, Supported by Epidemiology and In Vitro Data - PubMed. Available online: (accessed on.
- Nickols, N.G.; Mi, Z.; DeMatt, E.; Biswas, K.; Clise, C.E.; Huggins, J.T.; Maraka, S.; Ambrogini, E.; Mirsaeidi, M.S.; Levin, E.R.; et al. Effect of Androgen Suppression on Clinical Outcomes in Hospitalized Men With COVID-19: The HITCH Randomized Clinical Trial. JAMA network open 2022, 5, e227852. [Google Scholar] [CrossRef]
- Rambhatla, A.; Bronkema, C.J.; Corsi, N.; Keeley, J.; Sood, A.; Affas, Z.; Dabaja, A.A.; Rogers, C.G.; Liroff, S.A.; Abdollah, F. COVID-19 Infection in Men on Testosterone Replacement Therapy. The Journal of Sexual Medicine 2021, 18, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.B.O.; Leal, F.; Argenton, J.L.P.; Carvalheira, J.B.C. Impact of androgen deprivation therapy on mortality of prostate cancer patients with COVID-19: A propensity score-based analysis. Infectious Agents and Cancer 2021, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Cinislioglu, A.E.; Cinislioglu, N.; Demirdogen, S.O.; Sam, E.; Akkas, F.; Altay, M.S.; Utlu, M.; Sen, I.A.; Yildirim, F.; Kartal, S.; et al. The relationship of serum testosterone levels with the clinical course and prognosis of COVID-19 disease in male patients: A prospective study. Andrology 2022, 10, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Rastrelli, G.; Di Stasi, V.; Inglese, F.; Beccaria, M.; Garuti, M.; Di Costanzo, D.; Spreafico, F.; Greco, G.F.; Cervi, G.; Pecoriello, A.; et al. Low testosterone levels predict clinical adverse outcomes in SARS-CoV-2 pneumonia patients. Andrology 2021, 9, 88–98. [Google Scholar] [CrossRef]
- Camici, M.; Zuppi, P.; Lorenzini, P.; Scarnecchia, L.; Pinnetti, C.; Cicalini, S.; Nicastri, E.; Petrosillo, N.; Palmieri, F.; D'Offizi, G.; et al. Role of testosterone in SARS-CoV-2 infection: A key pathogenic factor and a biomarker for severe pneumonia. International journal of infectious diseases: IJID: Official publication of the International Society for Infectious Diseases 2021, 108, 244–251. [Google Scholar] [CrossRef]
- Li, C.; Ye, Z.; Zhang, A.J.X.; Chan, J.F.W.; Song, W.; Liu, F.; Chen, Y.; Kwan, M.Y.W.; Lee, A.C.Y.; Zhao, Y.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection by Intranasal or Intratesticular Route Induces Testicular Damage. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America 2022, 75, e974–e990. [Google Scholar] [CrossRef] [PubMed]
- Pozzilli, P.; Lenzi, A. Commentary: Testosterone, a key hormone in the context of COVID-19 pandemic. Metabolism: Clinical and Experimental 2020, 108, 154252. [Google Scholar] [CrossRef]
- Costeira, R.; Lee, K.A.; Murray, B.; Christiansen, C.; Castillo-Fernandez, J.; Ni Lochlainn, M.; Capdevila Pujol, J.; Macfarlane, H.; Kenny, L.C.; Buchan, I.; et al. Estrogen and COVID-19 symptoms: Associations in women from the COVID Symptom Study. PLoS ONE 2021, 16, e0257051. [Google Scholar] [CrossRef]
- Montopoli, M.; Zorzi, M.; Cocetta, V.; Prayer-Galetti, T.; Guzzinati, S.; Bovo, E.; Rugge, M.; Calcinotto, A. Clinical outcome of SARS-CoV-2 infection in breast and ovarian cancer patients who underwent antiestrogenic therapy. Annals of Oncology: Official Journal of the European Society for Medical Oncology 2021, 32, 676–677. [Google Scholar] [CrossRef]
- Baker, M.E. What are the physiological estrogens? Steroids 2013, 78, 337–340. [Google Scholar] [CrossRef]
- Lange, C.A.; Gioeli, D.; Hammes, S.R.; Marker, P.C. Integration of rapid signaling events with steroid hormone receptor action in breast and prostate cancer. Annual Review of Physiology 2007, 69, 171–199. [Google Scholar] [CrossRef]
- Bashour, N.M.; Wray, S. Progesterone directly and rapidly inhibits GnRH neuronal activity via progesterone receptor membrane component 1. Endocrinology 2012, 153, 4457–4469. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ulrich, B.; Cho, J.; Park, J.; Kim, C.H. Progesterone promotes differentiation of human cord blood fetal T cells into T regulatory cells but suppresses their differentiation into Th17 cells. Journal of Immunology (Baltimore, Md.: 1950) 2011, 187, 1778–1787. [Google Scholar] [CrossRef] [PubMed]
- Bereshchenko, O.; Bruscoli, S.; Riccardi, C. Glucocorticoids, Sex Hormones, and Immunity. Frontiers in Immunology 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Goddard, L.M.; Ton, A.N.; Org, T.; Mikkola, H.K.A.; Iruela-Arispe, M.L. Selective suppression of endothelial cytokine production by progesterone receptor. Vascular Pharmacology 2013, 59, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.B. COVID-19 and Progesterone: Part 1. SARS-CoV-2, Progesterone and its potential clinical use. Endocrine and Metabolic Science 2021, 5, 100109. [Google Scholar] [CrossRef]
- Ghandehari, S.; Matusov, Y.; Pepkowitz, S.; Stein, D.; Kaderi, T.; Narayanan, D.; Hwang, J.; Chang, S.; Goodman, R.; Ghandehari, H.; et al. Progesterone in Addition to Standard of Care vs Standard of Care Alone in the Treatment of Men Hospitalized With Moderate to Severe COVID-19: A Randomized, Controlled Pilot Trial. Chest 2021, 160, 74–84. [Google Scholar] [CrossRef]
- Yuan, L.; Zhu, H.; Wu, K.; Zhou, M.; Ma, J.; Chen, R.; Tang, Q.; Cheng, T.; Guan, Y.; Xia, N. Female sex hormone, progesterone, ameliorates the severity of SARS-CoV-2-caused pneumonia in the Syrian hamster model. Signal Transduction and Targeted Therapy 2022, 7, 47. [Google Scholar] [CrossRef]
- Chen, L.; Li, Q.; Zheng, D.; Jiang, H.; Wei, Y.; Zou, L.; Feng, L.; Xiong, G.; Sun, G.; Wang, H.; et al. Clinical Characteristics of Pregnant Women with Covid-19 in Wuhan, China. The New England Journal of Medicine 2020, 382, e100. [Google Scholar] [CrossRef]
- Ellington, S.; Strid, P.; Tong, V.T.; Woodworth, K.; Galang, R.R.; Zambrano, L.D.; Nahabedian, J.; Anderson, K.; Gilboa, S.M. Characteristics of Women of Reproductive Age with Laboratory-Confirmed SARS-CoV-2 Infection by Pregnancy Status - United States, January 22-June 7, 2020. MMWR. Morbidity and mortality weekly report 2020, 69, 769–775. [Google Scholar] [CrossRef]
- Tang, M.; Zhang, H.; Cai, J. Coronavirus disease 2019 in pregnancy. International journal of infectious diseases: IJID: Official publication of the International Society for Infectious Diseases 2021, 105, 721. [CrossRef]
- Knight, M.; Bunch, K.; Vousden, N.; Morris, E.; Simpson, N.; Gale, C.; O'Brien, P.; Quigley, M.; Brocklehurst, P.; Kurinczuk, J.J.; et al. Characteristics and outcomes of pregnant women admitted to hospital with confirmed SARS-CoV-2 infection in UK: National population based cohort study. BMJ (Clinical research ed.) 2020, 369, m2107. [Google Scholar] [CrossRef]
- Leach, D.A.; Brooke, G.N.; Bevan, C.L. Roles of steroid receptors in the lung and COVID-19. Essays in Biochemistry 2021, 65, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Shulla, A.; Heald-Sargent, T.; Subramanya, G.; Zhao, J.; Perlman, S.; Gallagher, T. A transmembrane serine protease is linked to the severe acute respiratory syndrome coronavirus receptor and activates virus entry. Journal of Virology 2011, 85, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Science China. Life Sciences 2020, 63, 457–460. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e286. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. The Journal of Pathology 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-Cell RNA Expression Profiling of ACE2, the Receptor of SARS-CoV-2. American Journal of Respiratory and Critical Care Medicine 2020, 202, 756–759. [Google Scholar] [CrossRef]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. International Journal of Oral Science 2020, 12, 8. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Chen, M.; Feng, Y.; Xiong, C. The ACE2 expression in human heart indicates new potential mechanism of heart injury among patients infected with SARS-CoV-2. Cardiovascular Research 2020, 116, 1097–1100. [Google Scholar] [CrossRef]
- Douglas, G.C.; O'Bryan, M.K.; Hedger, M.P.; Lee, D.K.L.; Yarski, M.A.; Smith, A.I.; Lew, R.A. The novel angiotensin-converting enzyme (ACE) homolog, ACE2, is selectively expressed by adult Leydig cells of the testis. Endocrinology 2004, 145, 4703–4711. [Google Scholar] [CrossRef]
- Bernstein, K.E.; Khan, Z.; Giani, J.F.; Cao, D.-Y.; Bernstein, E.A.; Shen, X.Z. Angiotensin-converting enzyme in innate and adaptive immunity. Nature Reviews. Nephrology 2018, 14, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Iwata-Yoshikawa, N.; Okamura, T.; Shimizu, Y.; Hasegawa, H.; Takeda, M.; Nagata, N. TMPRSS2 Contributes to Virus Spread and Immunopathology in the Airways of Murine Models after Coronavirus Infection. Journal of Virology 2019, 93, e01815–01818. [Google Scholar] [CrossRef]
- Hoffmann, M.; Hofmann-Winkler, H.; Smith, J.C.; Krüger, N.; Arora, P.; Sørensen, L.K.; Søgaard, O.S.; Hasselstrøm, J.B.; Winkler, M.; Hempel, T.; et al. Camostat mesylate inhibits SARS-CoV-2 activation by TMPRSS2-related proteases and its metabolite GBPA exerts antiviral activity. EBioMedicine 2021, 65, 103255. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Schroeder, S.; Kleine-Weber, H.; Müller, M.A.; Drosten, C.; Pöhlmann, S. Nafamostat Mesylate Blocks Activation of SARS-CoV-2: New Treatment Option for COVID-19. Antimicrobial Agents and Chemotherapy 2020, 64, e00754–00720. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Meyerholz, D.K.; Bartlett, J.A.; McCray, P.B. The TMPRSS2 Inhibitor Nafamostat Reduces SARS-CoV-2 Pulmonary Infection in Mouse Models of COVID-19. mBio 2021, 12, e0097021. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, M.; Damalanka, V.C.; Tartell, M.A.; Chung, D.H.; Lourenço, A.L.; Pwee, D.; Mayer Bridwell, A.E.; Hoffmann, M.; Voss, J.; Karmakar, P.; et al. A novel class of TMPRSS2 inhibitors potently block SARS-CoV-2 and MERS-CoV viral entry and protect human epithelial lung cells. Proceedings of the National Academy of Sciences of the United States of America 2021, 118, e2108728118. [Google Scholar] [CrossRef]
- A Phase 2 Randomized, Double-Blind, Placebo-controlled Trial of Oral Camostat Mesylate for Early Treatment of COVID-19 Outpatients Showed Shorter Illness Course and Attenuation of Loss of Smell and Taste - PubMed. Available online: (accessed on.
- Zhuravel, S.V.; Khmelnitskiy, O.K.; Burlaka, O.O.; Gritsan, A.I.; Goloshchekin, B.M.; Kim, S.; Hong, K.Y. Nafamostat in hospitalized patients with moderate to severe COVID-19 pneumonia: A randomised Phase II clinical trial. EClinicalMedicine 2021, 41, 101169. [Google Scholar] [CrossRef]
- Rossi, Á.D.; de Araújo, J.L.F.; de Almeida, T.B.; Ribeiro-Alves, M.; de Almeida Velozo, C.; Almeida, J.M.d.; de Carvalho Leitão, I.; Ferreira, S.N.; da Silva Oliveira, J.; Alves, H.J.; et al. Association between ACE2 and TMPRSS2 nasopharyngeal expression and COVID-19 respiratory distress. Scientific Reports 2021, 11, 9658. [Google Scholar] [CrossRef]
- Chappell, M.C.; Marshall, A.C.; Alzayadneh, E.M.; Shaltout, H.A.; Diz, D.I. Update on the Angiotensin converting enzyme 2-Angiotensin (1-7)-MAS receptor axis: Fetal programing, sex differences, and intracellular pathways. Frontiers in Endocrinology 2014, 4, 201. [Google Scholar] [CrossRef]
- Berletch, J.B.; Yang, F.; Xu, J.; Carrel, L.; Disteche, C.M. Genes that escape from X inactivation. Human Genetics 2011, 130, 237–245. [Google Scholar] [CrossRef]
- Tukiainen, T.; Villani, A.-C.; Yen, A.; Rivas, M.A.; Marshall, J.L.; Satija, R.; Aguirre, M.; Gauthier, L.; Fleharty, M.; Kirby, A.; et al. Landscape of X chromosome inactivation across human tissues. Nature 2017, 550, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Milsted, A.; Underwood, A.C.; Dunmire, J.; DelPuerto, H.L.; Martins, A.S.; Ely, D.L.; Turner, M.E. Regulation of multiple renin-angiotensin system genes by Sry. Journal of Hypertension 2010, 28, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, X.-M.; Mannan, R.; Pitchiaya, S.; Zhang, Y.; Wotring, J.W.; Xiao, L.; Robinson, D.R.; Wu, Y.-M.; Tien, J.C.-Y.; et al. Targeting transcriptional regulation of SARS-CoV-2 entry factors ACE2 and TMPRSS2. Proceedings of the National Academy of Sciences of the United States of America 2021, 118, e2021450118. [Google Scholar] [CrossRef]
- Baughn, L.B.; Sharma, N.; Elhaik, E.; Sekulic, A.; Bryce, A.H.; Fonseca, R. Targeting TMPRSS2 in SARS-CoV-2 Infection. Mayo Clinic Proceedings 2020, 95, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging 2020, 12, 10087–10098. [Google Scholar] [CrossRef] [PubMed]
- Leach, D.A.; Mohr, A.; Giotis, E.S.; Cil, E.; Isac, A.M.; Yates, L.L.; Barclay, W.S.; Zwacka, R.M.; Bevan, C.L.; Brooke, G.N. The antiandrogen enzalutamide downregulates TMPRSS2 and reduces cellular entry of SARS-CoV-2 in human lung cells. Nature Communications 2021, 12, 4068. [Google Scholar] [CrossRef]
- Song, H.; Seddighzadeh, B.; Cooperberg, M.R.; Huang, F.W. Expression of ACE2, the SARS-CoV-2 Receptor, and TMPRSS2 in Prostate Epithelial Cells. European Urology 2020, 78, 296–298. [Google Scholar] [CrossRef]
- Kalidhindi, R.S.R.; Borkar, N.A.; Ambhore, N.S.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Sex steroids skew ACE2 expression in human airway: A contributing factor to sex differences in COVID-19? American Journal of Physiology. Lung Cellular and Molecular Physiology 2020, 319, L843–L847. [Google Scholar] [CrossRef]
- Stelzig, K.E.; Canepa-Escaro, F.; Schiliro, M.; Berdnikovs, S.; Prakash, Y.S.; Chiarella, S.E. Estrogen regulates the expression of SARS-CoV-2 receptor ACE2 in differentiated airway epithelial cells. American Journal of Physiology. Lung Cellular and Molecular Physiology 2020, 318, L1280–L1281. [Google Scholar] [CrossRef]
- Liu, J.; Ji, H.; Zheng, W.; Wu, X.; Zhu, J.J.; Arnold, A.P.; Sandberg, K. Sex differences in renal angiotensin converting enzyme 2 (ACE2) activity are 17β-oestradiol-dependent and sex chromosome-independent. Biology of Sex Differences 2010, 1, 6. [Google Scholar] [CrossRef]
- Lemes, R.M.R.; Costa, A.J.; Bartolomeo, C.S.; Bassani, T.B.; Nishino, M.S.; Pereira, G.J.d.S.; Smaili, S.S.; Maciel, R.M.d.B.; Braconi, C.T.; da Cruz, E.F.; et al. 17β-estradiol reduces SARS-CoV-2 infection in vitro. Physiological Reports 2021, 9, e14707. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Q.; Xia, X.; Liu, K.; Yu, Z.; Tao, W.; Gong, W.; Han, J.-D.J. Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell 2020, 19, e13168. [Google Scholar] [CrossRef]
- Keidar, S.; Gamliel-Lazarovich, A.; Kaplan, M.; Pavlotzky, E.; Hamoud, S.; Hayek, T.; Karry, R.; Abassi, Z. Mineralocorticoid receptor blocker increases angiotensin-converting enzyme 2 activity in congestive heart failure patients. Circulation Research 2005, 97, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jessup, J.A.; Zhao, Z.; Da Silva, J.; Lin, M.; MacNamara, L.M.; Ahmad, S.; Chappell, M.C.; Ferrario, C.M.; Groban, L. Characterization of the cardiac renin angiotensin system in oophorectomized and estrogen-replete mRen2.Lewis rats. PLoS ONE 2013, 8, e76992. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Pineda, J.A.; Albaghdadi, M.; Jiang, W.; Vera-Lopez, K.J.; Nieto-Montesinos, R.; Alvarez, K.L.F.; Davila Del-Carpio, G.; Gómez, B.; Lindsay, M.E.; Malhotra, R.; et al. Structural and Functional Analysis of Female Sex Hormones against SARS-CoV-2 Cell Entry. International Journal of Molecular Sciences 2021, 22, 11508. [Google Scholar] [CrossRef]
- Fischer, M.; Baessler, A.; Schunkert, H. Renin angiotensin system and gender differences in the cardiovascular system. Cardiovascular Research 2002, 53, 672–677. [Google Scholar] [CrossRef]
- Chadchan, S.B.; Popli, P.; Maurya, V.K.; Kommagani, R. The SARS-CoV-2 receptor, angiotensin-converting enzyme 2, is required for human endometrial stromal cell decidualization†. Biology of Reproduction 2021, 104, 336–343. [Google Scholar] [CrossRef]
- Setlur, S.R.; Mertz, K.D.; Hoshida, Y.; Demichelis, F.; Lupien, M.; Perner, S.; Sboner, A.; Pawitan, Y.; Andrén, O.; Johnson, L.A.; et al. Estrogen-dependent signaling in a molecularly distinct subclass of aggressive prostate cancer. Journal of the National Cancer Institute 2008, 100, 815–825. [Google Scholar] [CrossRef]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension (Dallas, Tex.: 1979) 2001, 37, 1199–1208. [Google Scholar] [CrossRef]
- Targeting androgen regulation of TMPRSS2 and ACE2 as a therapeutic strategy to combat COVID-19 - PubMed. Available online: (accessed on.
- Baratchian, M.; McManus, J.M.; Berk, M.P.; Nakamura, F.; Mukhopadhyay, S.; Xu, W.; Erzurum, S.; Drazba, J.; Peterson, J.; Klein, E.A.; et al. Androgen regulation of pulmonary AR, TMPRSS2 and ACE2 with implications for sex-discordant COVID-19 outcomes. Scientific Reports 2021, 11, 11130. [Google Scholar] [CrossRef]
- Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer - PubMed. Available online: (accessed on.
- Lucas, J.M.; Heinlein, C.; Kim, T.; Hernandez, S.A.; Malik, M.S.; True, L.D.; Morrissey, C.; Corey, E.; Montgomery, B.; Mostaghel, E.; et al. The androgen-regulated protease TMPRSS2 activates a proteolytic cascade involving components of the tumor microenvironment and promotes prostate cancer metastasis. Cancer Discovery 2014, 4, 1310–1325. [Google Scholar] [CrossRef]
- Mikkonen, L.; Pihlajamaa, P.; Sahu, B.; Zhang, F.-P.; Jänne, O.A. Androgen receptor and androgen-dependent gene expression in lung. Molecular and Cellular Endocrinology 2010, 317, 14–24. [Google Scholar] [CrossRef]
- Distinct mechanisms for TMPRSS2 expression explain organ-specific inhibition of SARS-CoV-2 infection by enzalutamide - PubMed. Available online: (accessed on.
- Bischof, E.; Wolfe, J.; Klein, S.L. Clinical trials for COVID-19 should include sex as a variable. The Journal of Clinical Investigation 2020, 130, 3350–3352. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. The Lancet. Respiratory Medicine 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nature Reviews. Immunology 2020, 20, 269–270. [Google Scholar] [CrossRef]
- Xia, H.-J.; Zhang, G.-H.; Wang, R.-R.; Zheng, Y.-T. The influence of age and sex on the cell counts of peripheral blood leukocyte subpopulations in Chinese rhesus macaques. Cellular & Molecular Immunology 2009, 6, 433–440. [Google Scholar] [CrossRef]
- Melgert, B.N.; Oriss, T.B.; Qi, Z.; Dixon-McCarthy, B.; Geerlings, M.; Hylkema, M.N.; Ray, A. Macrophages: Regulators of sex differences in asthma? American Journal of Respiratory Cell and Molecular Biology 2010, 42, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Hewagama, A.; Patel, D.; Yarlagadda, S.; Strickland, F.M.; Richardson, B.C. Stronger inflammatory/cytotoxic T-cell response in women identified by microarray analysis. Genes and Immunity 2009, 10, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, M.; McClellan, B.; Allansmith, M. Influence of sex in immunoglobulin levels. Nature 1967, 214, 1224–1225. [Google Scholar] [CrossRef] [PubMed]
- Amadori, A.; Zamarchi, R.; De Silvestro, G.; Forza, G.; Cavatton, G.; Danieli, G.A.; Clementi, M.; Chieco-Bianchi, L. Genetic control of the CD4/CD8 T-cell ratio in humans. Nature Medicine 1995, 1, 1279–1283. [Google Scholar] [CrossRef]
- Meng, Y.; Wu, P.; Lu, W.; Liu, K.; Ma, K.; Huang, L.; Cai, J.; Zhang, H.; Qin, Y.; Sun, H.; et al. Sex-specific clinical characteristics and prognosis of coronavirus disease-19 infection in Wuhan, China: A retrospective study of 168 severe patients. PLoS pathogens 2020, 16, e1008520. [Google Scholar] [CrossRef] [PubMed]
- Verthelyi, D.; Klinman, D.M. Sex hormone levels correlate with the activity of cytokine-secreting cells in vivo. Immunology 2000, 100, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, C.M.; Cleary, J.; Dagtas, A.S.; Moussai, D.; Diamond, B. Estrogen alters thresholds for B cell apoptosis and activation. The Journal of Clinical Investigation 2002, 109, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Fuseini, H.; Yung, J.A.; Cephus, J.Y.; Zhang, J.; Goleniewska, K.; Polosukhin, V.V.; Peebles, R.S.; Newcomb, D.C. Testosterone Decreases House Dust Mite-Induced Type 2 and IL-17A-Mediated Airway Inflammation. Journal of Immunology (Baltimore, Md.: 1950) 2018, 201, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Kadihasanoglu, M.; Aktas, S.; Yardimci, E.; Aral, H.; Kadioglu, A. SARS-CoV-2 Pneumonia Affects Male Reproductive Hormone Levels: A Prospective, Cohort Study. The Journal of Sexual Medicine 2021, 18, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Trigunaite, A.; Dimo, J.; Jørgensen, T.N. Suppressive effects of androgens on the immune system. Cellular Immunology 2015, 294, 87–94. [Google Scholar] [CrossRef]
- Kadel, S.; Kovats, S. Sex Hormones Regulate Innate Immune Cells and Promote Sex Differences in Respiratory Virus Infection. Frontiers in Immunology 2018, 9, 1653. [Google Scholar] [CrossRef]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Medicine 2020, 46, 846–848. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet (London, England) 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Correction to: Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Medicine 2020, 46, 1294–1297. [Google Scholar] [CrossRef]
- Pagano, M.T.; Peruzzu, D.; Busani, L.; Pierdominici, M.; Ruggieri, A.; Antinori, A.; D'Offizi, G.; Petrosillo, N.; Palmieri, F.; Piselli, P.; et al. Predicting respiratory failure in patients infected by SARS-CoV-2 by admission sex-specific biomarkers. Biology of Sex Differences 2021, 12, 63. [Google Scholar] [CrossRef]
- Infante, M.; Pieri, M.; Lupisella, S.; D'Amore, L.; Bernardini, S.; Fabbri, A.; Iannetta, M.; Andreoni, M.; Morello, M. Low testosterone levels and high estradiol to testosterone ratio are associated with hyperinflammatory state and mortality in hospitalized men with COVID-19. European Review for Medical and Pharmacological Sciences 2021, 25, 5889–5903. [Google Scholar] [CrossRef]
- Cadegiani, F.A.; McCoy, J.; Gustavo Wambier, C.; Goren, A. Early Antiandrogen Therapy With Dutasteride Reduces Viral Shedding, Inflammatory Responses, and Time-to-Remission in Males With COVID-19: A Randomized, Double-Blind, Placebo-Controlled Interventional Trial (EAT-DUTA AndroCoV Trial - Biochemical). Cureus 2021, 13, e13047. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F.A.; McCoy, J.; Gustavo Wambier, C.; Vaño-Galván, S.; Shapiro, J.; Tosti, A.; Zimerman, R.A.; Goren, A. Proxalutamide Significantly Accelerates Viral Clearance and Reduces Time to Clinical Remission in Patients with Mild to Moderate COVID-19: Results from a Randomized, Double-Blinded, Placebo-Controlled Trial. Cureus 2021, 13, e13492. [Google Scholar] [CrossRef] [PubMed]
- Nicastri, E.; Marinangeli, F.; Pivetta, E.; Torri, E.; Reggiani, F.; Fiorentino, G.; Scorzolini, L.; Vettori, S.; Marsiglia, C.; Gavioli, E.M.; et al. A phase 2 randomized, double-blinded, placebo-controlled, multicenter trial evaluating the efficacy and safety of raloxifene for patients with mild to moderate COVID-19. EClinicalMedicine 2022, 48, 101450. [Google Scholar] [CrossRef]
- Woolf, P.D.; Hamill, R.W.; McDonald, J.V.; Lee, L.A.; Kelly, M. Transient hypogonadotropic hypogonadism caused by critical illness. The Journal of Clinical Endocrinology and Metabolism 1985, 60, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Hall, O.J.; Klein, S.L. Progesterone-based compounds affect immune responses and susceptibility to infections at diverse mucosal sites. Mucosal Immunology 2017, 10, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Hua, D.; Li, J.-P.; Zhang, X.-N.; Bai, L.; Cao, L.-B.; Guo, Y.; Zhang, M.; Dong, J.-Z.; Liang, X.-W.; et al. Modulation of innate immune response to viruses including SARS-CoV-2 by progesterone. Signal Transduction and Targeted Therapy 2022, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, G.; Tonetti, T.; Protti, A.; Langer, T.; Girardis, M.; Bellani, G.; Laffey, J.; Carrafiello, G.; Carsana, L.; Rizzuto, C.; et al. Pathophysiology of COVID-19-associated acute respiratory distress syndrome: A multicentre prospective observational study. The Lancet. Respiratory Medicine 2020, 8, 1201–1208. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. The New England Journal of Medicine 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Card, J.W.; Carey, M.A.; Bradbury, J.A.; DeGraff, L.M.; Morgan, D.L.; Moorman, M.P.; Flake, G.P.; Zeldin, D.C. Gender differences in murine airway responsiveness and lipopolysaccharide-induced inflammation. Journal of Immunology (Baltimore, Md.: 1950) 2006, 177, 621–630. [Google Scholar] [CrossRef]
- Diaz, G.A.; Parsons, G.T.; Gering, S.K.; Meier, A.R.; Hutchinson, I.V.; Robicsek, A. Myocarditis and Pericarditis After Vaccination for COVID-19. JAMA 2021, 326, 1210–1212. [Google Scholar] [CrossRef]
- da Silva, J.S.; Montagnoli, T.L.; Rocha, B.S.; Tacco, M.L.C.A.; Marinho, S.C.P.; Zapata-Sudo, G. Estrogen Receptors: Therapeutic Perspectives for the Treatment of Cardiac Dysfunction after Myocardial Infarction. International Journal of Molecular Sciences 2021, 22, 525. [Google Scholar] [CrossRef]
- Hale, S.L.; Birnbaum, Y.; Kloner, R.A. beta-Estradiol, but not alpha-estradiol, reduced myocardial necrosis in rabbits after ischemia and reperfusion. American Heart Journal 1996, 132, 258–262. [Google Scholar] [CrossRef]
- Manavathi, B.; Kumar, R. Steering estrogen signals from the plasma membrane to the nucleus: Two sides of the coin. Journal of Cellular Physiology 2006, 207, 594–604. [Google Scholar] [CrossRef]
- Gupta, A.; Madhavan, M.V.; Sehgal, K.; Nair, N.; Mahajan, S.; Sehrawat, T.S.; Bikdeli, B.; Ahluwalia, N.; Ausiello, J.C.; Wan, E.Y.; et al. Extrapulmonary manifestations of COVID-19. Nature Medicine 2020, 26, 1017–1032. [Google Scholar] [CrossRef]
- Ryan, D.H.; Ravussin, E.; Heymsfield, S. COVID 19 and the Patient with Obesity - The Editors Speak Out. Obesity (Silver Spring, Md.) 2020, 28, 847. [Google Scholar] [CrossRef] [PubMed]
- Dietz, W.; Santos-Burgoa, C. Obesity and its Implications for COVID-19 Mortality. Obesity (Silver Spring, Md.) 2020, 28, 1005. [Google Scholar] [CrossRef]
- Petrilli, C.M.; Jones, S.A.; Yang, J.; Rajagopalan, H.; O'Donnell, L.; Chernyak, Y.; Tobin, K.A.; Cerfolio, R.J.; Francois, F.; Horwitz, L.I. Factors associated with hospital admission and critical illness among 5279 people with coronavirus disease 2019 in New York City: Prospective cohort study. BMJ (Clinical research ed.) 2020, 369, m1966. [Google Scholar] [CrossRef] [PubMed]
- Kassir, R. Risk of COVID-19 for patients with obesity. Obesity Reviews: An Official Journal of the International Association for the Study of Obesity 2020, 21, e13034. [CrossRef]
- Giagulli, V.A.; Castellana, M.; Pelusi, C.; Triggiani, V. Androgens, Body Composition, and Their Metabolism Based on Sex. Frontiers of Hormone Research 2019, 53, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proceedings of the National Academy of Sciences of the United States of America 2000, 97, 12729–12734. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, J.C.; Bray, G.A.; Bourgeois, M.O.; Macchiavelli, R.; Rood, J.C.; Greeson, C.; Partington, C. Exogenous androgens influence body composition and regional body fat distribution in obese postmenopausal women--a clinical research center study. The Journal of Clinical Endocrinology and Metabolism 1996, 81, 2198–2203. [Google Scholar] [CrossRef] [PubMed]
- Battisti, S.; Pedone, C.; Napoli, N.; Russo, E.; Agnoletti, V.; Nigra, S.G.; Dengo, C.; Mughetti, M.; Conte, C.; Pozzilli, P.; et al. Computed Tomography Highlights Increased Visceral Adiposity Associated With Critical Illness in COVID-19. Diabetes Care 2020, 43, e129–e130. [Google Scholar] [CrossRef]
- Fain, J.N.; Madan, A.K.; Hiler, M.L.; Cheema, P.; Bahouth, S.W. Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology 2004, 145, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Nutrition, the visceral immune system, and the evolutionary origins of pathogenic obesity. Proceedings of the National Academy of Sciences of the United States of America 2019, 116, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Guo, C.; Tang, L.; Hong, Z.; Zhou, J.; Dong, X.; Yin, H.; Xiao, Q.; Tang, Y.; Qu, X.; et al. Prolonged presence of SARS-CoV-2 viral RNA in faecal samples. The Lancet. Gastroenterology & Hepatology 2020, 5, 434–435. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nature Medicine 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Galasso, V.; Pons, V.; Profeta, P.; Becher, M.; Brouard, S.; Foucault, M. Gender differences in COVID-19 attitudes and behavior: Panel evidence from eight countries. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 27285–27291. [Google Scholar] [CrossRef]
- The Lancet, n. The gendered dimensions of COVID-19. Lancet (London, England) 2020, 395, 1168. [Google Scholar] [CrossRef]
- Tadiri, C.P.; Gisinger, T.; Kautzy-Willer, A.; Kublickiene, K.; Herrero, M.T.; Raparelli, V.; Pilote, L.; Norris, C.M.; Consortium, G.-F. The influence of sex and gender domains on COVID-19 cases and mortality. CMAJ: Canadian Medical Association journal = journal de l'Association medicale canadienne 2020, 192, E1041–E1045. [Google Scholar] [CrossRef]
Figure 1.
Effects of sex hormones on the course of severe COVID-19. (a) Mechanism of viral invasion (b) Immune system re-sponse. (c) Regulation and direction of severe disease course a-l by sex hormones. TMPRSS2, transmembrane protease serine 2; ACE2, angiotensin-converting enzyme 2; ARDS, acute respiratory distress syndrome; VAT, visceral adiposity.
Figure 1.
Effects of sex hormones on the course of severe COVID-19. (a) Mechanism of viral invasion (b) Immune system re-sponse. (c) Regulation and direction of severe disease course a-l by sex hormones. TMPRSS2, transmembrane protease serine 2; ACE2, angiotensin-converting enzyme 2; ARDS, acute respiratory distress syndrome; VAT, visceral adiposity.
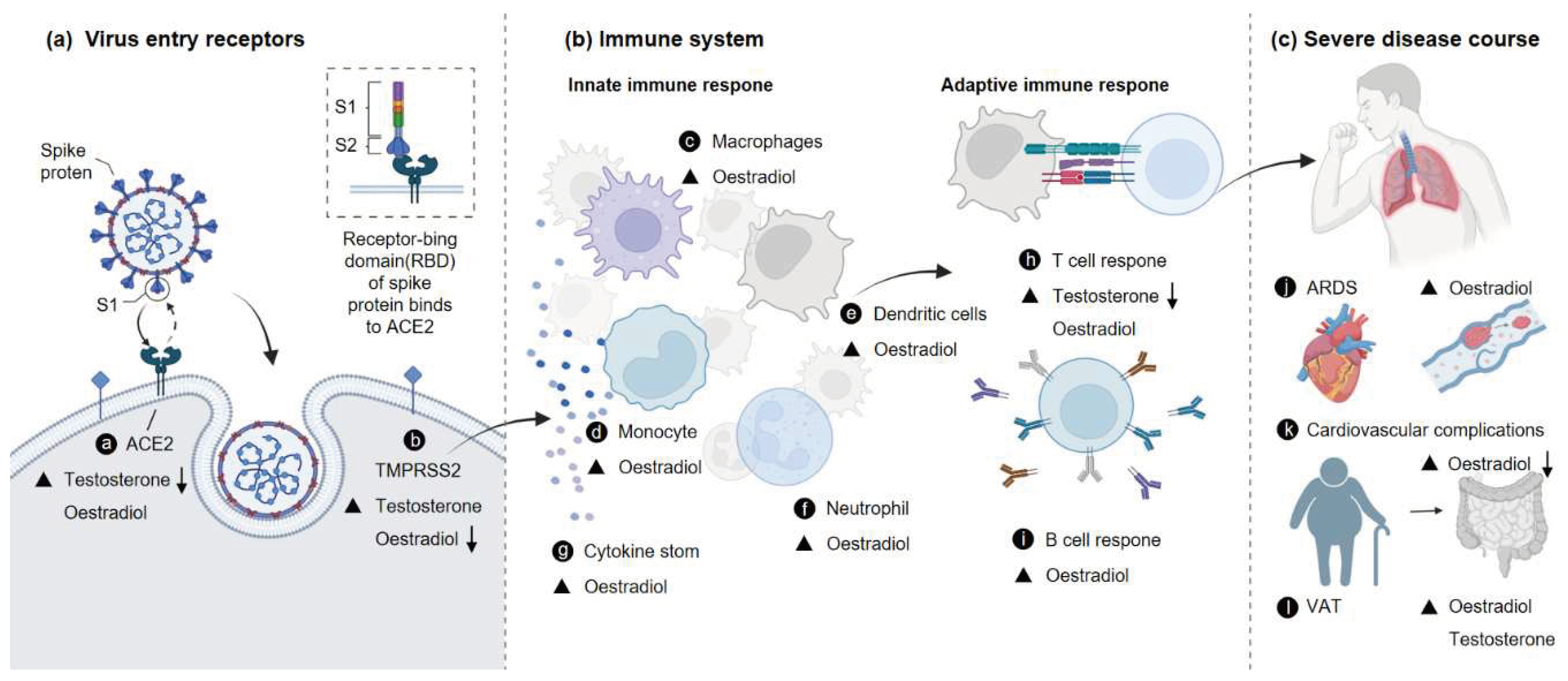

Table 1.
Regulation of TMPRSS2 and ACE2 by sex hormones.
| Viral Receptor |
Sex Hormone /Study Drug |
Species | Primary Findings | Sample | Refs. |
|---|---|---|---|---|---|
| ACE2 | Antiandrogen enzalutamide | Human | Decreases ACE2 expression | Heart cells and pulmonary tissue derived from human embryonic stem cells (hESC) | [8] |
| Testosterone | Human | Upregulation of ACE2 gene in men and women | Airway smooth muscle cells | [62] | |
| Estrogen | Human | Reduces ACE2, but not substantially | Airway smooth muscle cells | ||
| Antiandrogen enzalutamide | Human | Reducing TMPRSS2 expression | Human lung cells | [84] | |
| Estrogen | Mouse | Increases ACE2 expression | Thymus | [76] | |
| Testosterone | Mouse | Increase ACE2 expression | Kidney | ||
| Antagonist enzalutamide | Mouse | Moderately suppressible | Lung | [85] | |
| TMPRSS2 | Antagonist enzalutamide | Mouse | Did not decrease pulmonary TMPRSS2 | Lung | |
| Antiandrogen enzalutamide | Mouse | Reduced TMPRSS2 levels | Airway epithelial cells | [70] | |
| Leuprolide or Estradiol | Human | In males treated with leuprolide or estradiol, TMPRSS2 levels were markedly lower. | Epithelial cells | [87] | |
| ACE2 and TMPRSS2 | Androgen | Mouse | Decreased Tmprss2 and Ace2 expression in lung epithelial cells. | Lung epithelial cells | [67,72] |
| Castration | Mouse | Reduced levels of ACE2 and TMPRSS2 in lung, seminal vesicles and small intestine; ACE2 upregulated in kidney tissue, but not TMPRSS2 | Systemic | [84] |
Table 2.
Regulation of COVID-19 by sex hormones through immunization.
| Research | STUDY DRUG | Species | Primary Findings | Refs. |
|---|---|---|---|---|
| Observational Research | Testosterone | Male | Men with lower plasma levels of Testosterone were more likely to have more severe ARDS and to experience a worsening of their status after hospitalization. | [106] |
| Exploratory retrospective study | Testosterone | Male | Plasma angiotensin 1-7 levels and neutrophil count were predictors of ARDS outcome only in women, while plasma Testosterone levels and lymphocyte count were only indicative in men. | [108] |
| Exploratory retrospective study | Estradiol | Female | Estrogen plasma concentrations were positively correlated with pulmonary function in COVID-19 women and negative correlated with pulmonary function in COVID-19 men. | |
| Exploratory retrospective study | Estradiol/Testosterone ratio | Male/Female | The non-survivors had a significantly higher median value for the Estradiol/Testosterone ratio. | [104] |
| Randomized controlled trial | Raloxifene | Male/Female | Increased white blood cell counts and accelerated viral clearing. | [112] |
| Experimental research | Progesterone | Hamsters | Inhibit proinflammatory cytokine overproduction and viral replication in lung. | [39] |
| Experimental research | Progesterone | Mice | Triggers downstream antiviral genes, stimulating cellular as well as mouse innate antiviral response. | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



