
Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
Phase change refers to the process of maturation and transition from the juvenile to the adult stage. In response to this shift, certain species like chestnut lose the ability to form adventitious roots, thereby hindering the successful micropropagation of adult plants. While auxin is the main hormone involved in adventitious root formation, other hormones, such as ethylene, are also thought to play a role in its induction and development. In this study, experiments were carried out to determine the effects of ethylene on the induction and growth of adventitious roots. The analysis was performed in two types of chestnut microshoots derived from the same tree, a juvenile-like line with high rooting ability derived from basal shoots (P2BS) and a line derived from crown branches (P2CR) with low rooting responses. By means of the application of compounds to modify ethylene content or inhibit its signalling, the potential involvement of this hormone in the in-duction of adventitious roots was analyzed. Our results show that ethylene can modify the rooting competence of mature shoots, while the response in juvenile material was barely affected. To further characterize the molecular reasons underlying this maturation-derived shift in behaviour, specific gene expression analyses were developed. Findings suggest that several mechanisms, including ethylene signalling, auxin transport and epigenetic modifications, relate to the modu-lation of the rooting ability of mature chestnut microshoots and their recalcitrant behaviour.
Keywords:
auxin
; Castanea sativa
; gene expression
; maturation
; recalcitrance
; root induction
1. Introduction
The European sweet chestnut (Castanea sativa) is a profitable and cost-effective woody species in the Mediterranean basin [1,2], highly relevant for biomass, timber and fruit production [3,4]. Chestnut trees have a great ecological value, as they have been related to an increase in plant species richness and they are important carbon sinks aiding to mitigate climate change [5,6]. Besides, chestnut trees are involved in the maintenance of traditional landscapes, significantly contributing to the environmental and cultural heritage [7,8]. Nonetheless, potential applications are hindered by its recalcitrant behaviour, thus driving to the use of biotechnological approaches to overcome this limitation. Selected chestnut genotypes have been successfully micropropagated by using both juvenile and mature material, although acceptable rooting rates have only been attained in juvenile-like material (reviewed in [9]). However, rooting ability is also greatly influenced by the genotype, thus limiting the pool of available material for the vegetative propagation of this species.
Adventitious rooting (AR) is a complex post-embryogenic process through which roots are formed in tissues such as stems, leaves and hypocotyls, from cells not previously determined to form roots. AR is modulated by several internal factors such as mother plant status, the chronological and ontogenetic age, the genetic makeup, the hormonal balance, as well as external conditions like temperature, light or mineral nutrition [10,11,12]. This process is a common occurrence in plant development and can be triggered as a response to different stressors such as flooding, nutrient deficiency, wounding or oxidative stress [13,14]. The formation of adventitious roots (ARs) is a key step in the vegetative propagation of plants. Many species, including several trees, exhibit a recalcitrant behaviour and their ability to form roots decreases dramatically with age as they go through the phase change from the juvenile to the mature stage, with low responses to rooting stimuli [11,15,16]. Reversion of mature-related traits, particularly the improvement of AR, represents a challenge for the propagation of species whose desirable traits are only seen in the mature stage. The lack of rooting or the deficiencies in root architecture reduce the survival rate of plants and hamper the mass propagation of selected genotypes, causing heavy economic losses for the related industries.
AR is divided into three sequential stages, named induction, initiation and expression (outgrowth). In chestnut, the most limiting stage is the induction phase, in which specific cells respond to the stimuli and initiate a root developmental program in root-prone tissues/genotypes, takes place in the first 24-48 hours after the beginning of the induction treatment [9,17]. Every step in the AR process is subject to a dynamic and specific hormone modulation, although auxin is the key hormone in AR [18]. The decline of rooting ability after the transition from the juvenile to the mature stage is linked to changes in auxin homeostasis [11,16], modifying the expression of auxin-responsive genes [19,20]. However, other plant hormones are involved in the regulation of AR. Plant growth regulators such as cytokinins [21] or gibberellins [22] seem to inhibit AR, whereas jasmonates [23,24] or abscisic acid [25] are ARs stimulators in some species. Nonetheless, the role played by different hormones seems to have a species-specific component, while at the same time their function might be dependent on the particular stage of the process.
Ethylene (ET) is a gaseous plant growth regulator involved in different plant developmental processes, including close links to plant aging and phase change [26,27,28]. Moreover, ET has been shown to interact with auxin in primary root development and other rooting processes [29,30]. In chestnut, a transcriptomic analysis revealed that ET signalling is upregulated in mature tissues in response to auxin and wounding when compared to juvenile tissues. This finding suggests that a high concentration of ET in mature tissues could be one of the causes underlying its recalcitrant behaviour [20]. ET biosynthesis and signalling pathways have been properly characterized in plants, leading to the development of specific strategies that allow for their modulation. For instance, ET biosynthesis might be enhanced or reduced by the application of its precursor 1-aminocyclopropane-1-carboxylate (ACC) or aminoethoxyvinylglycine (AVG), respectively. On the other hand, the use of silver ions allows for the inhibition of ET perception (reviewed in [31]). To test the effects of ET on the root system development of chestnut, ACC, silver nitrate (AgNO3) and AVG were applied to the root induction media of juvenile and mature chestnut microshoots. Results obtained prompted us to further characterize the responses in mature chestnut tissues, where specific gene expression related to ET synthesis and signalling, auxin responses and epigenetic processes was evaluated by quantitative PCR. The results suggest a negative effect of ET on the induction of ARs in mature tissues and a link to recalcitrance, which might at least partially exert its action through the interaction with auxin transport and specific epigenetic processes.
2. Results
2.1. Rooting experiments
Ethylene negatively modulates adventitious rooting and worsens root system development in mature chestnut
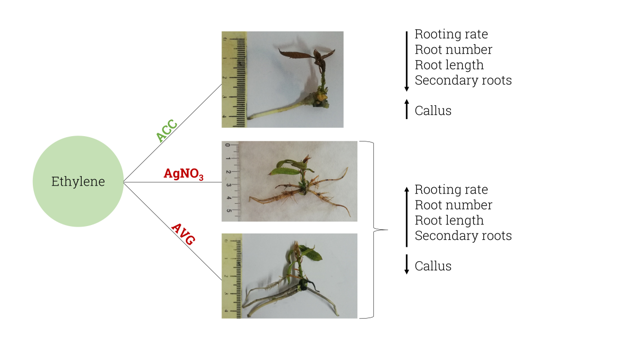
Rooting traits were maintained constant throughout all the experiments in the rooting-competent juvenile line (P2BS, Figure 1 and Figure 2), and the rooting rates were 100% for all treatments. Similarly, the average root number per explant and root length parameters were not significantly affected by the treatments. Root length ranged from 3.3 cm in IBA-treated shoots to 4.5 cm in IBA+AVG treated shoots, with intermediate values for IBA+AgN03 and IBA+ACC treatments (4.3 and 3.7cm, respectively). However, ET perception inhibition seemed to have a slight effect on rooting performance of P2BS shoots, decreasing root number per shoot (Figure 2b) and increasing root length (Figure 2c). On the other hand, AVG treatment promoted lateral root formation, while AgNO3 reduced shoot tip necrosis (Figure 1c,1d).
In contrast, the rooting performance of P2CR shoots was affected by ET modulators (Figure 1 and Figure 2). Shoots induced to root in RIM exhibited a 27% rooting rate, the lowest rooting percentage (18%) was obtained in ACC-treated shoots, while the maximum rooting percentage was achieved in AgNO3- and AVG-treated shoots (44% and 40%, respectively; Figure 2a). Regarding root number per explant, ET attenuation improved this trait more than doubling the data obtained from the IBA and IBA+ACC treatments (Figure 2b). These data highlighted a negative effect of ACC in the number of roots per explant, not only in their ability to root. Moreover, the average root length in IBA+ACC treated shoots was 1.7, while it ranged from 6.6 to 7.9 cm in the rest of the treatments (Figure 2c). Interestingly, P2CR shoots rooted in the presence of silver nitrate or AVG showed a lower amount of callus tissue than those rooted in the other treatments, as well as a marked increase of lateral roots, suggesting a long-lasting effect of ET-inhibition in specific root traits (Figure 1g-h). Altogether these results showed that the presence of ACC seems to negatively modulate the induction and development of ARs in mature shoots, whereas lowering ET action by AgNO3 or AVG has the opposite effect.
2.2. Analysis of gene expression during AR in rooting-recalcitrant shoots
2.2.1. Ethylene-related genes
To confirm the effect of ET modulators on the expression pattern of ET-related genes during the AR process, expression analysis of CsACS1-like, CsACO1-like, CsRAP2.12, CsERF3, and CsEIN2 genes was performed in the basal parts of P2CR shoots, where the rooting response might take place.
IBA treatment decreased the expression of both ET synthesis-related genes (CsACO1-like, CsACS1-like) at 24h (Figure 3a, 3b). However, the expression of CsACS1-like in IBA-treated shoots at 120h was similar to that in T0 samples, while it remained low for CsACO1-like. AgNO3-mediated inhibition of ET perception of IBA-treated shoots did not affect the expression levels of those genes, suggesting that the treatments (IBA, IBA+AVG and IBA+AgNO3) restrain ET synthesis to a similar degree (Figure 3a, 3b). However, at 120h silver nitrate application had a slight but not significant effect on the expression of CsACS1-like. On the other hand, ACC application upregulated the expression of both genes at 24h, with a sustained induction only for CsACS1-like, while at 120h expression of CsACO1-like in ACC-treated shoots was lower than in the T0 samples (Figure 3b). These results suggest a positive feedback of ACC on its own synthesis, which was active for 120h, and promotion of the conversion of ACC in ET in the short term. Therefore, distinct treatments affected differently the ET biosynthesis pathway, which might be related to the dissimilar phenotypic responses found.
To further characterize ET-related responses, the expression of genes linked to ET signalling was analyzed. CsEIN2, a core gene controlling gene expression in response to ET, showed contrasting results in response to treatments. During the first 120h, expression of CsEIN2 was downregulated by the IBA+AgNO3 treatment, thus showing a block in ET signalling. On the other hand, at 24h ACC treatment induced the expression of CsEIN2 in IBA-treated samples, while at 120h it did not affect the transcription of CsEIN2 since mRNA levels were similar to those detected in control samples (Figure 3c). IBA alone slightly reduced the expression of CsEIN2. Therefore, CsEIN2 expression was responsive to ET-content modifying treatments, particularly at 24h. Two transcription factors involved in ET responses were also analyzed. At 24h, expression of CsERF3 decreased in response to AgNO3 and AVG, while it was scarcely induced by ACC (Figure 3d). In the rest of the samples, no significant shifts were detected in the expression of the gene. These results suggest that CsERF3 expression is determined by ET content in chestnut mature tissues, showing a temporary response that might be related to early events in AR. On the other hand, the expression of CsRAP2.12 showed no significant changes in response to the different treatments. However, all samples showed a lower level of expression than the T0 one, except for the ACC-treated 24h samples (Figure 3e). These results suggest that CsRAP2.12 is modulated by ET at 24h, however, in the rest of the samples and time points subjected to analysis the transcription of the gene was not affected by IBA and ET-related treatments.
2.2.2. Auxin-related genes
Due to the role of auxin in the AR process, expression analysis of specific genes related to this hormone was performed. Regarding CsPIN1, a membrane transporter involved in auxin polar movement, IBA treatment increased the expression of the gene at 24h and 120h, which was expected by the increase of auxin concentration in the tissues. However, no changes in CsPIN1 expression were detected in ACC-treated samples regarding to the control (Figure 4b). Strikingly, the expression level of the gene was dramatically induced by AgNO3 and AVG treatments at 24h, which was maintained upregulated at 120h (Figure 4b). Therefore, the induction of CsPIN1, an auxin transporter gene involved in the generation of hormone gradients, by ET perception inhibition or ET synthesis blocking correlates to the improvement of AR. In the case of CsIAA29, no significant changes in gene expression were detected. However, except for ACC-treated samples at 24h, expression levels were higher than in control samples (Figure 4a), suggesting an auxin-responsive behaviour of the gene that can be restrained in the presence of ET. Indeed, CsIAA29 levels of expression at 24h show a direct link with the AR rates obtained from the three treatments (Figure 2a and Figure 4a). However, the lack of significant differences in expression prevents us from drawing further conclusions. Therefore, improvement of AR in response to ET-perception or synthesis inhibition shows close links with auxin-related processes, specifically transport.
2.2.3. Epigenetic-related genes
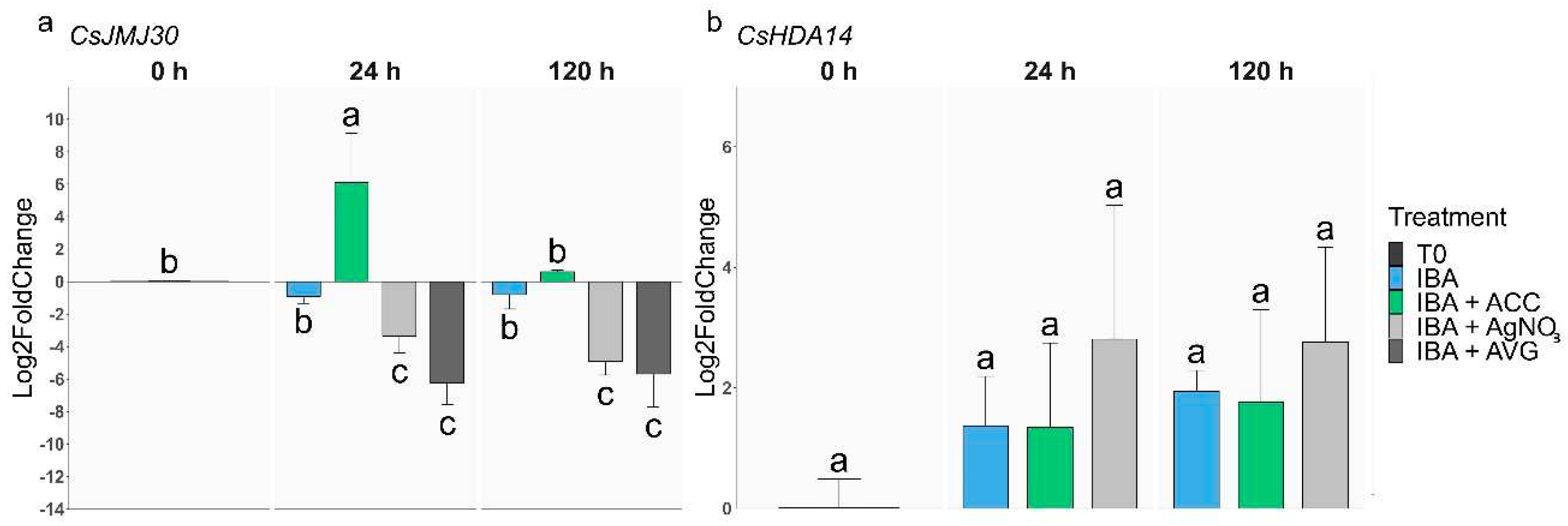
Two genes linked to epigenetic responses were analyzed to infer possible mechanisms of AR induction in mature tissues and its relation to hormone signalling. CsHDA14, a histone deacetylase, showed no significant changes in response to the different treatments, however all samples showed a higher level of expression than the T0 samples (Figure 5b). Moreover, greater levels of expression were detected in AgNO3-treated samples, although as previously mentioned no significant differences were found.
CsJMJ30, a histone lysine-specific demethylase gene, showed to be responsive to the presumed ET increase in the tissues, specifically at 24h after the beginning of the treatment. At 24h, the expression of the gene was upregulated by ACC treatment, while at 120h CsJMJ30 expression levels dropped back to T0 levels. When only IBA was applied to the shoots, no significant changes in CsJMJ30 expression were detected (Figure 5a). ET perception and synthesis inhibition significantly repressed the expression of the gene at both time points, suggesting that AR improvement derived from ET blocking might at least partially exert its effect through the inhibition of this gene.
Overall, results obtained suggest that ET modifying treatments alter the expression of genes related to ET biosynthesis and signalling, with close links to auxin transport and specific epigenetic mechanisms.
3. Discussion
The successful development of ARs requires a change in the fate of specific cells not previously determined to form a meristem that will eventually drive the generation of the new organ [32]. In the case of rooting-competent chestnut microshoots, cells close to the vascular bundles in the stem are able to respond to the rooting stimuli, auxin and wounding, then reprogramming their ongoing genetic pattern and entering a root developmental pathway [9]. Therefore, the plasticity of those rooting-competent cells allows them to modify their gene expression patterns in response to wound stress and external auxin supply and switch their fate to AR founder cells. However, this ability is drastically reduced during maturation, with adult microshoots showing a recalcitrant behaviour that severely hampers their ability to form roots. Despite recent advances, the molecular basis of this recalcitrant behaviour is still poorly understood. Several players are believed to take part in this connection between AR and recalcitrance, particularly hormones, epigenetic mechanisms and their crosstalk, which may underlie developmental plasticity in plants [33]. A recent analysis showed that transcriptomic responses to auxin and wounding vary greatly between the two types of microshoots used in the present study [20]. Among the differences found, mature shoots exhibited an increased response related to ET, including biosynthesis-related genes and ET-responsive transcription factors. To gain deeper insight into the putative role of ET in the modulation of AR in chestnut and its relation to recalcitrance, treatments were designed where the ET content was either increased or decreased, or its perception was blocked. In juvenile shoots, no significant phenotypical changes were detected, suggesting that ET does not influence AR in these shoots. However, wounding is a necessary step for AR induction in cuttings and microshoots [9,10], and injury-related stress induces a temporary increase in ET and jasmonic acid (JA). These two hormones develop an antagonistic relation in which JA stimulates the expression of wound-responsive genes, while ET blocks that expression in order to locally and temporarily restrict the repairing response [34]. However, the lack of ET effect in P2BS shoots suggests that other mechanisms might be active in these tissues to drive tissue repair. On the other hand, ET modulation severely impacted the rooting response of mature shoots, including rooting rates, root number and root length. Therefore, our results suggest that repairing mechanisms and root induction processes might be different according to the ontogenetic state of the tissues.
ET has been shown to present contrasting effects in the formation of ARs in different species. It was described as an AR inhibitor in peach [35], Eucalyptus [36] and Malus x domestica [37]. On the other hand, there are species such as petunia [38], cucumber [39], marigold [40] or woody plants such as Pinus thunbergii [41] or Citrus sinensis [42] in which ET stimulates ARs formation. However, the effect of this hormone in AR has not been analyzed regarding to the ontogenetic state of the tissues in woody species. In Arabidopsis, ET-related signalling was shown to be more active in old leaves, which was linked to a low ability for de-novo root regeneration [43]. Therefore, ET influence on regeneration processes and particularly on AR might be directly connected to the age and development of the plants.
A remarkable effect of ET inhibition in P2BS and P2CR shoots was the ability to induce lateral roots (Figure 1), a trait that improves root system performance. The formation of these lateral roots took place long after the AgNO3 and AVG treatments ended, thus suggesting a lasting effect on microshoots performance. The inhibition of lateral root formation by ET was also described in Arabidopsis [44,45] and tomato [46]. The possible mechanism of this effect in our system is unknown, although a role for auxin transport modulation might be a plausible explanation (see below).
According to the results of the present study, ET signalling is involved in the induction of AR, at least in mature shoots. ET content and signalling modulation were shown to directly influence genes involved in ET synthesis. Particularly, ACC-treated P2CR shoots showed a significant increase in the expression of CsACS1-like and CsACO1-like, and this increase was related to a lower rooting response. Surprisingly, ACC triggered a positive feedback loop in the expression of CsACS1-like, that lasted up to five days after the beginning of the treatment. This gene codes for an enzyme that catalyzes the conversion of S-adenosyl-l-methionine (SAM) into ACC, which is later transformed in ET by the activity of ACO enzymes. Thus, in mature shoots, ACC induced an increase in its own synthesis, whose presence seems to prevent specific developmental processes, at least in this system. In recent years, several reports have suggested that ACC might exert signalling effects on its own, not only due to its role as an ET precursor. Those activities have been related to developmental processes and stress responses, and putative transporters for ACC have been identified [47,48]. However, the ACC effect on the expression of ET-related genes suggest that this might be its main route of action in the system used here.
On the other hand, both IBA, IBA+AVG and IBA+AgNO3 treatments reduced the expression of ET synthesis genes, particularly at 24h, thus indicating that lowering ET synthesis is necessary to induce rooting responses in mature shoots. Expression of CsEIN2, a core component of ET signalling, was modified according to treatments, with greater levels of expression in samples treated with the precursor of ethylene biosynthesis, while lower levels were detected when ET perception was inhibited. EIN2 acts downstream of ET receptors and modulates the activity of ET-Responsive transcription Factors (ERFs), also integrating inputs from other pathways in its expression. Here, CsEIN2 mRNA levels are clearly related to the rooting behaviour of mature shoots, with improved rooting responses by blocking ET signalling and thus decreased CsEIN2 expression. In Arabidopsis, an ACC+IBA treatment reduced the number of ARs, with the antagonistic relation between AtEIN2 and the JA signalling gene AtCOI1 showing to be particularly relevant in response to the IBA induction [49]. Moreover, EIN2 also seems to control ET-related gene expression by inducing histone acetylation, in what is suggested to be a rapid transcriptional regulation process [50]. Our data support this idea of a fast and dynamic control because CsERF3 showed a parallel expression pattern when compared to CsEIN2. However, previous reports suggested a positive role of ERF3 in the formation of ARs in Populus under normal and low-phosphorus conditions, which seems not to match results in our system, and a link to auxin signalling [51,52]. Probably, ET-related variations in the control of gene expression between both species underlie the differences found.
On the other hand, expression of the ET-responsive CsRAP2.12 transcription factor also slightly resembled CsEIN2 expression pattern at 24h. Previously, Valladares et al. [53] analyzed the expression of this gene in chestnut and oak tissues during AR induction. Authors suggested that it might be implicated in the establishment of new developmental programs in an ontogenetic-related mode. This gene belongs to group VII of ERFs, whose activity has been linked to specific responses like low-oxygen conditions or oxidative stress [54]. Here, levels of expression were lower than in T0 samples except for ACC+IBA treatment at 24 h, therefore its expression does not seem to relate to improved rooting responses.
ET and the key AR inducer, auxin, are known to interact in many processes and at different levels. Particularly, ET has been suggested to influence auxin movement through the modulation of the expression of auxin transporters [55]. The activity of these transporters is essential for the generation of hormone gradients in the tissues, which eventually trigger the process of AR [56]. ET-perception and synthesis inhibition dramatically induced the expression of CsPIN1 at 24h, while its induction was much more modest in IBA-treated shoots. Thus, ET seems to block auxin transport in mature chestnut tissues, preventing the establishment of hormone gradients needed for the induction of specific developmental responses. The activity of PIN transporters seems crucial in the early steps of AR for the successful outcome of the process, as seen for example in apple and tea nodal cuttings [57,58]. In other experimental systems ET also seems to influence regeneration processes by influencing auxin distribution, as seen in de novo shoot organogenesis in tamarillo [59].
In response to the establishment of auxin gradients, specific related gene expression is triggered. CsIAA29 resembled the expression pattern of CsPIN1 at 24h, when AR induction is taking place. Aux/IAA proteins are auxin-responsive and work as repressors of Auxin Responsive Factors, modulating their activity [60]. Therefore, CsIAA29 might be involved in AR induction playing a role in the fine-tuning of the auxin signalling events, at least in chestnut. Arabidopsis AtIAA29 has been suggested to be involved in the modulation of root system development [61], and ET was shown to target this gene through the EIN3 proteins, which work downstream of EIN2 [62]. Therefore, ET negatively influences AR induction in mature chestnut shoots by putatively preventing the establishment of auxin gradients and impeding further related signalling.
As previously mentioned, EIN2 influences histone acetylation. P2CR rooting might also be blocked through epigenetic mechanisms governed by ET, and the present results support this hypothesis. However, CsHDA14, a histone deacetylase that reduces the accessibility of the transcriptional machinery to DNA, showed no significant differences among treatments despite a higher level of expression in IBA+AgNO3 treated samples. On the other hand, the expression of CsJMJ30 clearly resembled the ET signalling status of the tissues, with expression being up-regulated in ACC-treated shoots and down-regulated by the inhibition of ET perception or synthesis. JMJ30 is a histone demethylase that generally acts in conjunction with its paralog JMJ32. In Arabidopsis, AtJMJ30 has been shown to lead to callus formation by inducing the genes AtLBD16 and AtLBD29 [63]. Moreover, it seems to control root elongation in response to abscisic acid [64]. In a previous report, abscisic acid-related gene expression was found to be more active in mature than in juvenile shoots of chestnut [20], thus suggesting that CsJMJ30 might be integrating different cues into its expression that eventually relate to the recalcitrant behaviour of P2CR shoots, as low levels of activity of this demethylase might be increasing the accessibility of the transcriptional machinery to genes whose expression is needed for the development of ARs.
4. Materials and Methods
4.1. Plant material and culture conditions
Microshoots of P2BS and P2CR lines established in vitro from basal shoots and crown branches, respectively, of an 80-year-old C. sativa Mill. tree [65] were used in this study. Shoot cultures were grown in GD [66] culture medium supplemented with 0.1 mg L-1 benzyladenine, 30 g L-1 of sucrose and 7 g L-1 of Bacto Agar as gelling agent. Culture medium pH was adjusted to 5.6 – 5.7 and autoclaved at 121°C for 20 minutes. Every 4 weeks, the basal callus from well-developed shoots was removed and the shoots were used for a new multiplication cycle by sub-culturing them on fresh culture medium, or for rooting experiments.
4.2. Rooting experiments
At the end of the proliferation cycle, microshoots devoided from the callus and basal leaves were used in rooting experiments. Root induction medium (RIM) consisted of GD medium with 1/3 of macronutrients concentration, 30 g L-1 of sucrose, 7 g L-1 of Bacto Agar and 25 µM Indole Butyric Acid (IBA). For rooting experiments, RIM-incubated microshoots for five days under dark conditions were transferred to the IBA-free RIM medium (REM, root expression medium) for 25 days and cultured under a 16h light/8h dark photoperiod with a light intensity of 40 µmol m-2 s-1 provided by cool-white fluorescent lamps.
To test the effect of ET on adventitious rooting, the induction of roots was carried out in: i) RIM, ii) RIM supplemented with 30 µM 1-aminocyclopropane-1-carboxylic acid (ACC), iii) RIM supplemented with 30 µM silver nitrate (AgNO3), and iv) RIM supplemented with 30 µM aminoethoxyvinylglycine (AVG). Rooting experiments were carried out with 6 explants in each oval Microbox container (OV80+OVD80 with white filter, Microbox, Deinze, Belgium), three containers per replicate and three repeats per treatment (6 explants per replication x 3 replications x 3 repeats = 54 explants per treatment). At the end of the rooting cycle, the percentage of rooted shoots, root number per explant and root length were recorded. Normality of data was tested by Shapiro-Wilk test of normality and the homogeneity of variance was analyzed by Levene’s test. Then, the data were analyzed by ANOVA or Kruskal-Wallis test and with a post-hoc comparison, HSD Tukey and Dunn test respectively. Those analyses were performed in R software [67].
4.3. RNA extraction and qPCR analysis
Plant material from P2CR shoots was collected at the end of the multiplication cycle (T0), and 24 hours and 120 hours after the initiation of rooting experiments (IBA, IBA+ACC, IBA+AgNO3, IBA+AVG). The basal parts (1 cm) of microshoots were cut off, immediately frozen in liquid nitrogen, and stored at -80 ºC until use. Samples were homogenised with liquid N2 and total RNA was extracted using Quick-RNATM Miniprep Kit from ©Zymo Research according to the manufacturer’s instructions. The concentration and quality of total RNA were assessed using a ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, NC, USA).
Synthesis of cDNA was performed from 1 µg of total RNA, using the NZY First-Strand cDNA Synthesis Flexible Pack kit following the manufacturer’s instructions. qPCR Primers were designed with the Primer Designing Tool Primer-BLAST based on the sequences of Quercus suber and Castanea mollissima. Genes whose expression was analyzed by qPCR include Castanea sativa ACC Synthase 1-like (CsACS1-like), ACC oxidase 1-like (CsACO1-like), Ethylene Insensitive 2 (CsEIN2), Ethylene Responsive Factor 3 (CsERF3), auxin transporter CsPIN1, IAA29 (CsIAA29), Histone deacetylase 14 (CsHDA14) and Jumonji 30 (CsJMJ30). For the analysis of the Ethylene Responsive Factor CsRAP2.12 and the reference genes for qPCR validation Actin-2 (CsACT2) and Elongation factor 1 (CsELF-1), sequences from a previous analysis were used [53]. The primer sequences are shown in Supplementary File 1 (S1). Three biological replicates were included for each qPCR analysis, using NZYSpeedy qPCR Green Master Mix (2x), following the provided instructions and with the samples diluted 10 times. The relative gene expression value was calculated according to the 2−∆∆CT method [68]. Results are expressed as relative values, using as reference the expression of the genes in the Control samples (T0), harvested at the end of the multiplication cycle and prior to any treatment. Data normality and homoscedasticity were tested by Shapiro-Wilk and by Levene’s tests, respectively. Then, ANOVA or Kruskal-Wallis tests were performed, and HSD Tukey or Dunn tests were used as post hoc comparison tests. Those analyses were performed in R software [67].
5. Conclusions
The rooting experiments carried out in this study revealed the effect of ET on AR in chestnut before and after phase change. Results showed a negative role of ET on the induction of ARs in mature chestnut-derived tissues but not in juvenile-like tissues, revealing that ET activity depends on the ontogenetic state of plant material. Molecular analysis revealed the existence of feedback-regulatory mechanisms of ACC on its own synthesis. ET perception inhibition by AVG and AgNO3 treatments seems to influence root development in three related ways: direct ET response inhibition through CsERF3 and CsEIN2 repression, auxin transport modification through CsPIN1 enhancement and epigenetic changes driven by CsJMJ30. More research is needed to elucidate the specific mode of action of AVG, AgNO3 and ET on the activity of CsJMJ30 and CsPIN1 to regulate root development and their implication in the aging processes of chestnut. However, the results provided in this study open new potential approaches to understand recalcitrance in chestnut and to improve protocols for its vegetative propagation. In addition, they allow for a better comprehension of the acquisition of rooting competence and its relationship with maturation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. List of primer sequences used in this study (Table S1).
Author Contributions
C.S., R.C-C., J.M.V.: Conceptualization. R.C-C., M.N.: Investigation. R.C-C., M.N., S.C.: Methodology. R.C-C, J.M.V., J.C.: Formal Analysis. R.C-C., M.N.: Project administration. J.C. C.S.: Resources. J.C. C.S.: Supervision. R.C-C.: Visualization R.C-C.: Writing – original draft. C.S, S.C., J.M.V.: Writing – review & editing. C.S., J.C.: Funding acquisition.
Funding
This work was supported by the Xunta de Galicia through the project IN607A 2021/06 (Spain) and by Center for Functional Ecology—Science for People and the Planet (UIDB/04004/2020), financed by FCT/MCTES through national funds (PIDDAC, Portugal).
Acknowledgments
This manuscript is based upon work from COST Action COPYTREE CA21157, supported by COST (European Cooperation in Science and Technology).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martín, M.A.; Mattioni, C.; Molina, J.R.; Alvarez, J.B.; Cherubini, M.; Herrera, M.A.; Villani, F.; Martín, L.M. Landscape genetic structure of chestnut (Castanea sativa Mill.) in Spain. Tree Genet. Genomes 2012, 8, 127–136. [CrossRef]
- Braga, N.; Rodrigues, F.; P.P. Oliveira, M.B. Castanea sativa by-products: a review on added value and sustainable application. Nat. Prod. Res. 2015, 29, 1–18. [CrossRef]
- Vega, A.; Dieste, A.; Guaita, M.; Majada, J.; Baño, V. Modelling of the mechanical properties of Castanea sativa Mill. structural timber by a combination of non-destructive variables and visual grading parameters. Eur. J. Wood Wood Prod. 2012, 70, 839–844. [CrossRef]
- Nunes, L.; Gower, S.; Monteiro, M.; Lopes, D.; Rego, F. Growth dynamics and productivity of pure and mixed Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco plantations in northern Portugal. iForest - Biogeosciences For. 2014, 7, 92–102. [CrossRef]
- Guitián, J.; Guitián, P.; Magrach, A.; Docampo, C.; Domínguez, P.; Guitián, L. Effect of management and spatial characteristics on plant species richness of Castanea sativa Mill. woodlots in the NW Iberian Peninsula. J. For. Res. 2012, 17, 98–104. [CrossRef]
- Castaño-Santamaría, J.; Barrio-Anta, M.; Álvarez-Álvarez, P. Potential above ground biomass production and total tree carbon sequestration in the major forest species in NW Spain. Int. For. Rev. 2013, 15, 273–289. [CrossRef]
- Pardo-de-Santayana, M.; Tardío, J.; Blanco, E.; Carvalho, A.M.; Lastra, J.J.; San Miguel, E.; Morales, R. Traditional knowledge of wild edible plants used in the northwest of the Iberian Peninsula (Spain and Portugal): a comparative study. J. Ethnobiol. Ethnomed. 2007, 3, 27. [CrossRef]
- Martínez, S.; Ramil, P.; Chuvieco, E. Monitoring loss of biodiversity in cultural landscapes. New methodology based on satellite data. Landsc. Urban Plan. 2010, 94, 127–140. [CrossRef]
- Vielba; Vidal, N.; San-José, M.C.; Rico, S.; Sánchez, C.; José, M.C.S.; Rico, S.; Sánchez, C. Recent Advances in Adventitious Root Formation in Chestnut. Plants 2020, 9, 1543. [CrossRef]
- da Costa, C.T.; de Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.S.; Fett-Neto, A.G. When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133. [CrossRef]
- Rasmussen, A.; Hosseini, S.A.; Hajirezaei, M.-R.; Druege, U.; Geelen, D. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. J. Exp. Bot. 2015, 66, 1437–1452. [CrossRef]
- Christiaens, A.; Gobin, B.; Van Huylenbroeck, J.; Van Labeke, M.-C. Adventitious rooting of Chrysanthemum is stimulated by a low red:far-red ratio. J. Plant Physiol. 2019, 236, 117–123. [CrossRef]
- Steffens, B.; Rasmussen, A. The Physiology of Adventitious Roots. Plant Physiol. 2016, 170, 603–617. [CrossRef]
- Zhang, Y.; Xiao, Z.; Zhan, C.; Liu, M.; Xia, W.; Wang, N. Comprehensive analysis of dynamic gene expression and investigation of the roles of hydrogen peroxide during adventitious rooting in poplar. BMC Plant Biol. 2019, 19, 99. [CrossRef]
- Husen, A.; Pal, M. Variation in Shoot Anatomy and Rooting Behaviour of Stem Cuttings in Relation to Age of Donor Plants in Teak (Tectona grandis Linn. f.). New For. 2006, 31, 57–73. [CrossRef]
- Aumond, M.L.; de Araujo, A.T.; de Oliveira Junkes, C.F.; de Almeida, M.R.; Matsuura, H.N.; de Costa, F.; Fett-Neto, A.G. Events Associated with Early Age-Related Decline in Adventitious Rooting Competence of Eucalyptus globulus Labill. Front. Plant Sci. 2017, 8. [CrossRef]
- Ballester, A.; San-José, M.C.; Vidal, N.; Fernández-Lorenzo, J.L.; Vieitez, A.M. Anatomical and Biochemical Events during in vitro Rooting of Microcuttings from Juvenile and Mature Phases of Chestnut. Ann. Bot. 1999, 83, 619–629. [CrossRef]
- de Almeida, M.R.; de Bastiani, D.; Gaeta, M.L.; de Araújo Mariath, J.E.; de Costa, F.; Retallick, J.; Nolan, L.; Tai, H.H.; Strömvik, M. V.; Fett-Neto, A.G. Comparative transcriptional analysis provides new insights into the molecular basis of adventitious rooting recalcitrance in Eucalyptus. Plant Sci. 2015, 239, 155–165. [CrossRef]
- Vielba, J.M.; Varas, E.; Rico, S.; Covelo, P.; Sánchez, C. Auxin-mediated expression of a GH3 gene in relation to ontogenic state in Chestnut. Trees 2016, 30, 2237–2252. [CrossRef]
- Vielba, J.M.; Rico, S.; Sevgin, N.; Castro-Camba, R.; Covelo, P.; Vidal, N.; Sanchez, C.; Sánchez, C. Transcriptomics Analysis Reveals a Putative Role for Hormone Signaling and MADS-Box Genes in Mature Chestnut Shoots Rooting Recalcitrance. Plants (Basel) 2022, 11, 3486. [CrossRef]
- Mao, J.; Zhang, D.; Meng, Y.; Li, K.; Wang, H.; Han, M. Inhibition of adventitious root development in apple rootstocks by cytokinin is based on its suppression of adventitious root primordia formation. Physiol. Plant. 2019, 166, 663–676. [CrossRef]
- Castro-Camba, R.; Sánchez, C.; Vidal, N.; Vielba, J.M. Plant Development and Crop Yield: The Role of Gibberellins. Plants 2022, 11, 2650. [CrossRef]
- Lischweski, S.; Muchow, A.; Guthörl, D.; Hause, B. Jasmonates act positively in adventitious root formation in petunia cuttings. BMC Plant Biol. 2015, 15, 229. [CrossRef]
- Fattorini, L.; Hause, B.; Gutierrez, L.; Veloccia, A.; Della Rovere, F.; Piacentini, D.; Falasca, G.; Altamura, M.M. Jasmonate promotes auxin-induced adventitious rooting in dark-grown Arabidopsis thaliana seedlings and stem thin cell layers by a cross-talk with ethylene signalling and a modulation of xylogenesis. BMC Plant Biol. 2018, 18, 182. [CrossRef]
- Li, C.; Bian, B.; Gong, T.; Liao, W. Comparative proteomic analysis of key proteins during abscisic acid-hydrogen peroxide-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Physiol. 2018, 229, 185–194. [CrossRef]
- Haffner, V.; Enjalric, F.; Lardet, L.; Carron, M. Maturation of woody plants: a review of metabolic and genomic aspects. Ann. des Sci. For. 1991, 48, 615–630. [CrossRef]
- Schaller, G.E. Ethylene and the regulation of plant development. BMC Biol. 2012, 10, 9. [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front. Plant Sci. 2017, 08. [CrossRef]
- Druege, U.; Franken, P.; Hajirezaei, M.R. Plant Hormone Homeostasis, Signaling, and Function during Adventitious Root Formation in Cuttings. Front. Plant Sci. 2016, 7, 381. [CrossRef]
- Qin, H.; He, L.; Huang, R. The Coordination of Ethylene and Other Hormones in Primary Root Development. Front. Plant Sci. 2019, 10. [CrossRef]
- Neves, M.; Correia, S.; Cavaleiro, C.; Canhoto, J. Modulation of Organogenesis and Somatic Embryogenesis by Ethylene: An Overview. Plants 2021, 10, 1208. [CrossRef]
- Gonin; Bergougnoux; Nguyen; Gantet; Champion What Makes Adventitious Roots? Plants 2019, 8, 240. [CrossRef]
- Maury, S.; Sow, M.D.; Le Gac, A.L.; Genitoni, J.; Lafon-Placette, C.; Mozgova, I. Phytohormone and Chromatin Crosstalk: The Missing Link For Developmental Plasticity? Front Plant Sci 2019, 10, 395. [CrossRef]
- Moore, B.M.; Lee, Y.S.; Wang, P.; Azodi, C.; Grotewold, E.; Shiu, S.-H. Modeling temporal and hormonal regulation of plant transcriptional response to wounding. Plant Cell 2022, 34, 867–888. [CrossRef]
- Park, S.-H.; Elhiti, M.; Wang, H.; Xu, A.; Brown, D.; Wang, A. Adventitious root formation of in vitro peach shoots is regulated by auxin and ethylene. Sci. Hortic. (Amsterdam). 2017, 226, 250–260. [CrossRef]
- Kilkenn, A.J.; Wallace, H.M.; Walton, D.A.; Adkins, M.F.; Trueman, S.J. Improved Root Formation in Eucalypt Cuttings Following Combined Auxin and Anti-ethylene Treatments. J. Plant Sci. 2012, 7, 138–153. [CrossRef]
- Ma, J.-H.; Yao, J.-L.; Cohen, D.; Morris, B. Ethylene inhibitors enhance in vitro root formation from apple shoot cultures. Plant Cell Rep. 1998, 17, 211–214. [CrossRef]
- Druege, U.; Franken, P.; Lischewski, S.; Ahkami, A.H.; Zerche, S.; Hause, B.; Hajirezaei, M.R. Transcriptomic analysis reveals ethylene as stimulator and auxin as regulator of adventitious root formation in petunia cuttings. Front. Plant Sci. 2014, 5. [CrossRef]
- Deng, Y.; Wang, C.; Zhang, M.; Wei, L.; Liao, W. Identification of Key Genes during Ethylene-Induced Adventitious Root Development in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2022, 23, 12981. [CrossRef]
- Jin, X.; Liao, W.-B.; Yu, J.-H.; Ren, P.-J.; Dawuda, M.M.; Wang, M.; Niu, L.-J.; Li, X.-P.; Xu, X.-T. Nitric Oxide Is Involved in Ethylene-Induced Adventitious Rooting in Marigold. Can. J. Plant Sci. 2017, CJPS-2016-0156. [CrossRef]
- Mori, Y.; Miyahara, F.; Tsutsumi, Y.; Kondo, R. Effects of combinational treatment with ethephon and indole-3-butyric acid on adventitious rooting of Pinus thunbergii cuttings. Plant Growth Regul. 2011, 63, 271–278. [CrossRef]
- Mendes, A.F.S.; Cidade, L.C.; Otoni, W.C.; Soares-Filho, W.S.; Costa, M.G.C. Role of auxins, polyamines and ethylene in root formation and growth in sweet orange. Biol. Plant. 2011, 55, 375–378. [CrossRef]
- Li, H.; Yao, L.; Sun, L.; Zhu, Z. ETHYLENE INSENSITIVE 3 suppresses plants de novo root regeneration from leaf explants and mediates age-regulated regeneration decline. Development 2020. [CrossRef]
- Negi, S.; Ivanchenko, M.G.; Muday, G.K. Ethylene regulates lateral root formation and auxin transport in Arabidopsis thaliana. Plant J. 2008, 55, 175–187. [CrossRef]
- Park, C.H.; Roh, J.; Youn, J.H.; Son, S.H.; Park, J.H.; Kim, S.Y.; Kim, T.W.; Kim, S.K. Arabidopsis ACC oxidase 1 coordinated by multiple signals mediates ethylene biosynthesis and is involved in root development. Mol. Cells 2018, 41, 923–932. [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [CrossRef]
- Polko, J.K.; Kieber, J.J. 1-Aminocyclopropane 1-Carboxylic Acid and Its Emerging Role as an Ethylene-Independent Growth Regulator. Front. Plant Sci. 2019, 10. [CrossRef]
- Li; Mou, W.; Van de Poel, B.; Chang, C. Something old, something new: Conservation of the ethylene precursor 1-amino-cyclopropane-1-carboxylic acid as a signaling molecule. Curr. Opin. Plant Biol. 2022, 65, 102116. [CrossRef]
- Betti, C.; Della Rovere, F.; Ronzan, M.; Fattorini, L. EIN2 and COI1 control the antagonism between ethylene and jasmonate in adventitious rooting of Arabidopsis thaliana thin cell layers. Plant Cell, Tissue Organ Cult. 2019, 138, 41–51. [CrossRef]
- Zhang; Wang, L.; Qi, B.; Zhao, B.; Ko, E.E.; Riggan, N.D.; Chin, K.; Qiao, H. EIN2 mediates direct regulation of histone acetylation in the ethylene response. Proc. Natl. Acad. Sci. 2017, 114, 10274–10279. [CrossRef]
- Trupiano, D.; Yordanov, Y.; Regan, S.; Meilan, R.; Tschaplinski, T.; Scippa, G.S.; Busov, V. Identification, characterization of an AP2/ERF transcription factor that promotes adventitious, lateral root formation in Populus. Planta 2013, 238, 271–282. [CrossRef]
- Wang; Pak, S.; Yang, J.; Wu, Y.; Li, W.; Feng, H.; Yang, J.; Wei, H.; Li, C. Two high hierarchical regulators, PuMYB40 and PuWRKY75, control the low phosphorus driven adventitious root formation in Populus ussuriensis. Plant Biotechnol. J. 2022, 20, 1561–1577. [CrossRef]
- Valladares, S.; Varas, E.; Vielba, J.M.; Vidal, N.; Codesido, V.; Castro, R.; Sanchez, C. Expression of a Rap2.12 like-1 ERF gene during adventitious rooting of chestnut and oak microshoots. Isr. J. Plant Sci. 2020, 67, 69–82. [CrossRef]
- Papdi, C.; Pérez-Salamó, I.; Joseph, M.P.; Giuntoli, B.; Bögre, L.; Koncz, C.; Szabados, L. The low oxygen, oxidative and osmotic stress responses synergistically act through the ethylene response factor VII genes RAP2.12, RAP2.2 and RAP2.3. Plant J. 2015, 82, 772–784. [CrossRef]
- Veloccia, A.; Fattorini, L.; Della Rovere, F.; Sofo, A.; D’Angeli, S.; Betti, C.; Falasca, G.; Altamura, M.M. Ethylene and auxin interaction in the control of adventitious rooting in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6445–6458. [CrossRef]
- Li Molecular Bases for the Regulation of Adventitious Root Generation in Plants. Front. Plant Sci. 2021, 12. [CrossRef]
- Guan, L.; Li, Y.; Huang, K.; Cheng, Z.-M. Auxin regulation and MdPIN expression during adventitious root initiation in apple cuttings. Hortic. Res. 2020, 7, 143. [CrossRef]
- Hu; Liu, X.; Xuan, W.; Mei, H.; Li, J.; Chen, X.; Zhao, Z.; Zhao, Y.; Jeyaraj, A.; Periakaruppan, R.; et al. Genome-wide identification and characterization of PIN-FORMED (PIN) and PIN-LIKES (PILS) gene family reveals their role in adventitious root development in tea nodal cutting (Camellia Sinensis). Int. J. Biol. Macromol. 2023, 229, 791–802. [CrossRef]
- Neves, M.; Correia, S.; Canhoto, J. Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav. Plants 2023, 12, 1854. [CrossRef]
- Li; Xie, Z.-Z.; Hu, C.-G.; Zhang, J.-Z.; Li, S.-B.; Xie, Z.-Z.; Hu, C.-G.; Zhang, J.-Z.; Li; Xie, Z.-Z.; et al. A Review of Auxin Response Factors (ARFs) in Plants. Front. Plant Sci. 2016, 7, 47. [CrossRef]
- Tang, Y.; Wang, L.; Qu, Z.; Huang, C.; Zhao, T.; Li, Y.; Zhang, C. BSISTER transcription factors directly binds to the promoter of IAA19 and IAA29 genes to up-regulate gene expression and promote the root development. Plant Sci. 2022, 321, 111324. [CrossRef]
- Chang, K.N.; Zhong, S.; Weirauch, M.T.; Hon, G.; Pelizzola, M.; Li, H.; Huang, S.C.; Schmitz, R.J.; Urich, M.A.; Kuo, D.; et al. Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis. Elife 2013, 2. [CrossRef]
- Lee, K.; Park, O.; Seo, P.J. JMJ30-mediated demethylation of H3K9me3 drives tissue identity changes to promote callus formation in Arabidopsis. Plant J. 2018, 95, 961–975. [CrossRef]
- Wu, J.; Yamaguchi, N.; Ito, T. Histone demethylases control root elongation in response to stress-signaling hormone abscisic acid. Plant Signal. Behav. 2019, 14, 1604019. [CrossRef]
- Sanchez, M.C.; Vieitez, A.M. In vitro morphogenetic competence of basal sprouts and crown branches of mature chestnut. Tree Physiol. 1991, 8, 59–70. [CrossRef]
- Gresshoff, P.M.; Doy, C.H. Development and differentiation of haploid Lycopersicon esculentum (tomato). Planta 1972, 107, 161–170. [CrossRef]
- R Core Team R: A language and environment for statistical computing. 2021.
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [CrossRef]
Figure 1.
Phenotypes of rooted chestnut plantlets. P2BS shoots are shown in the upper panel after treatment with a) IBA, b) IBA+ACC, c) IBA+AVG and d) IBA + AgNO3. P2CR shoots are shown in the lower panel after treatment with e) IBA, f) IBA+ACC, g) IBA+AVG and h) IBA+AgNO3.
Figure 1.
Phenotypes of rooted chestnut plantlets. P2BS shoots are shown in the upper panel after treatment with a) IBA, b) IBA+ACC, c) IBA+AVG and d) IBA + AgNO3. P2CR shoots are shown in the lower panel after treatment with e) IBA, f) IBA+ACC, g) IBA+AVG and h) IBA+AgNO3.

Figure 2.
Rooting rates of microshoots from lines P2CR and P2BS. Shoots were treated with IBA 25 µM (RIM), supplemented with ACC 30 µM (RIM+ACC), with AVG 30 µM (RIM+AVG) or with AgN03 30 µM (RIM+AgNO3). Different letters indicate significant differences in each trait (p ≤ 0.05). (a) Rooting rate; (b) Root number; (c) Root length.
Figure 2.
Rooting rates of microshoots from lines P2CR and P2BS. Shoots were treated with IBA 25 µM (RIM), supplemented with ACC 30 µM (RIM+ACC), with AVG 30 µM (RIM+AVG) or with AgN03 30 µM (RIM+AgNO3). Different letters indicate significant differences in each trait (p ≤ 0.05). (a) Rooting rate; (b) Root number; (c) Root length.
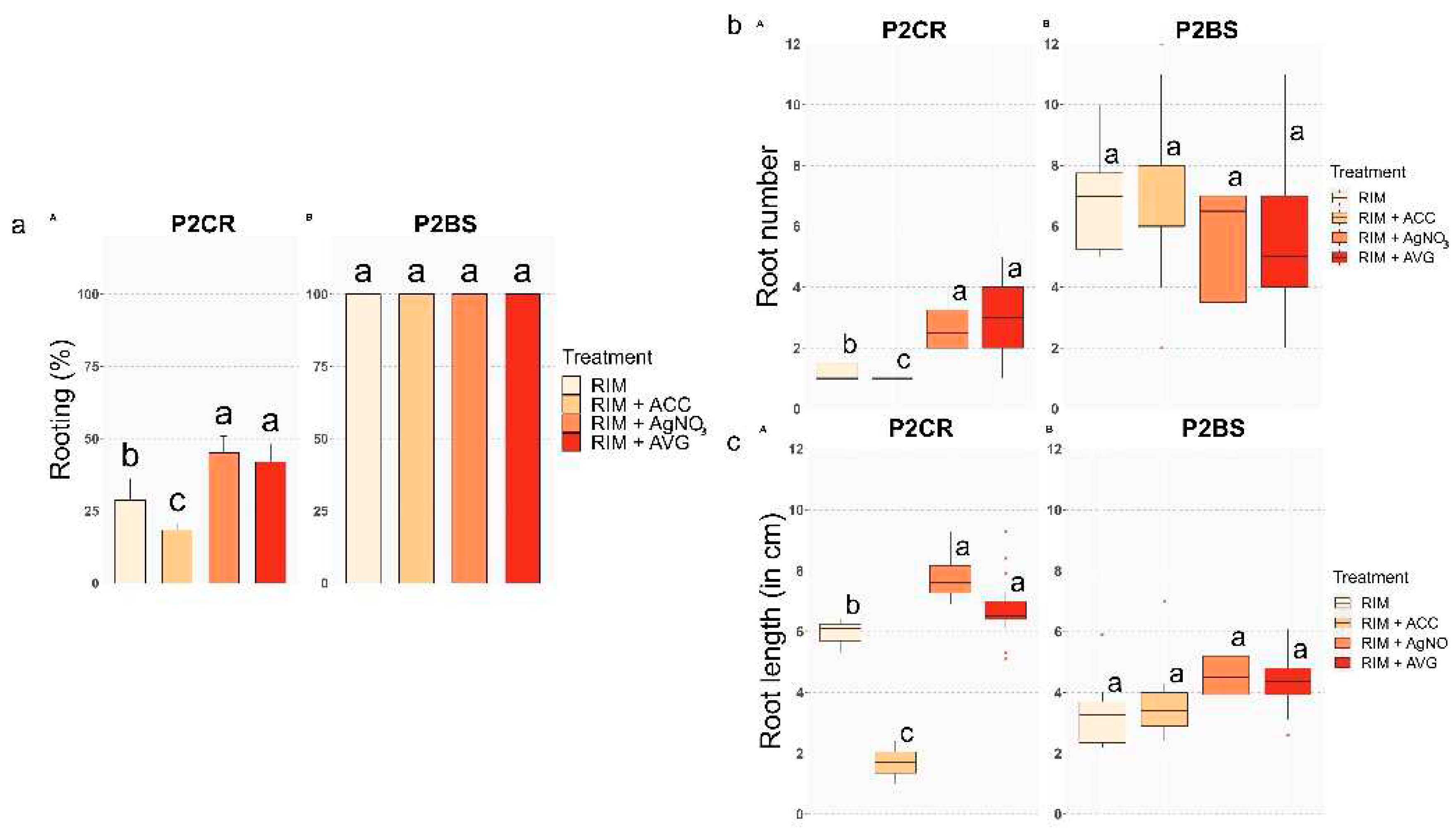
Figure 3.
qPCR expression analysis of genes related to ethylene synthesis and signalling. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsACS1-like; (b) CsACO1-like; (c) CsEIN2; (d) CsERF3; (e) CsRAP2.12. All data were normalized to the expression of the genes in the T0 samples. For each gene, different letters indicate statistical differences (p<0.05).
Figure 3.
qPCR expression analysis of genes related to ethylene synthesis and signalling. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsACS1-like; (b) CsACO1-like; (c) CsEIN2; (d) CsERF3; (e) CsRAP2.12. All data were normalized to the expression of the genes in the T0 samples. For each gene, different letters indicate statistical differences (p<0.05).
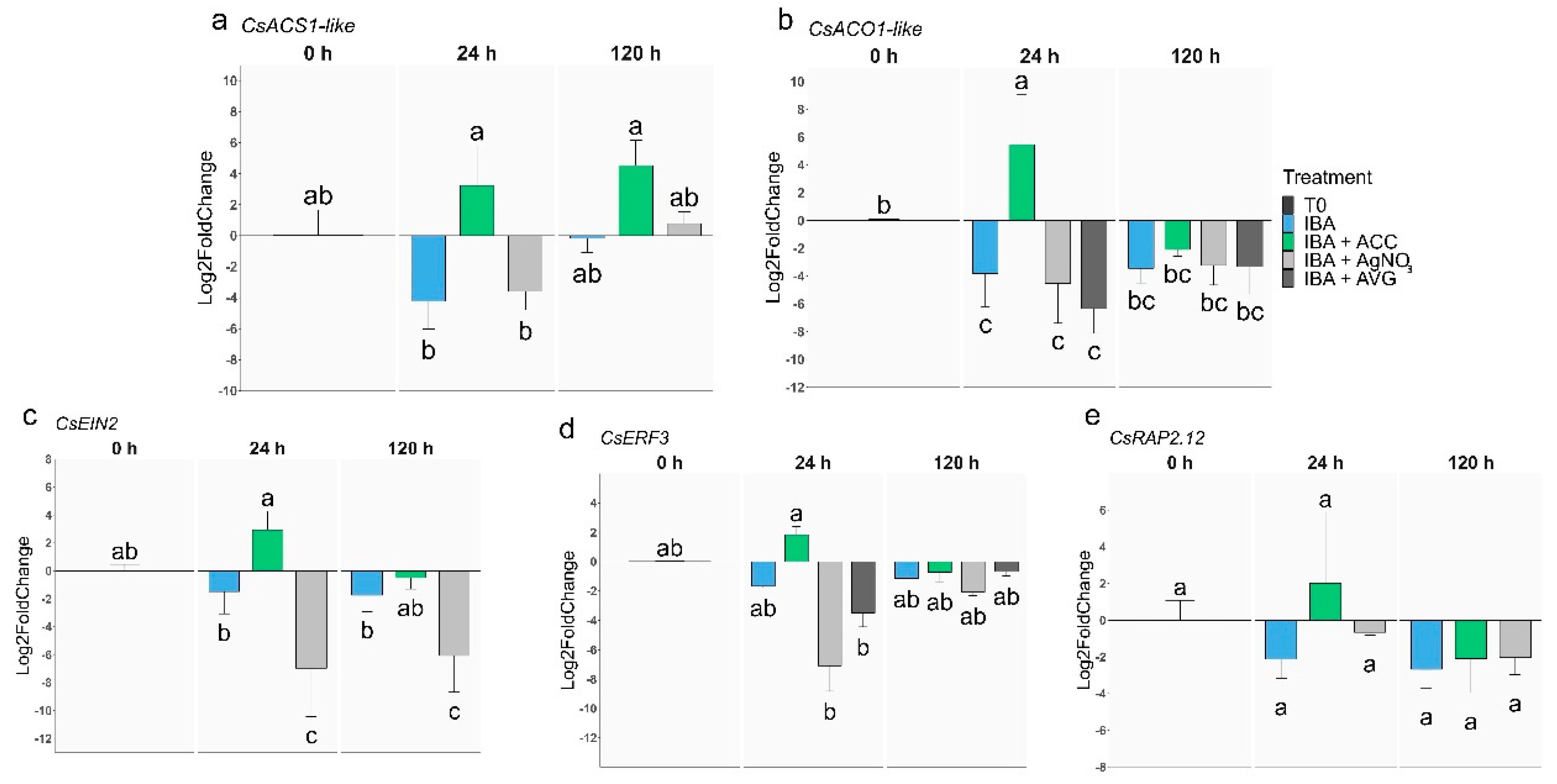
Figure 4.
qPCR expression analysis of genes related to auxin transport and signalling. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsIAA29; (b) CsPIN1. All data were normalized to the expression of T0 sample. For each gene, different letters indicate statistical differences (p<0.05).
Figure 4.
qPCR expression analysis of genes related to auxin transport and signalling. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsIAA29; (b) CsPIN1. All data were normalized to the expression of T0 sample. For each gene, different letters indicate statistical differences (p<0.05).
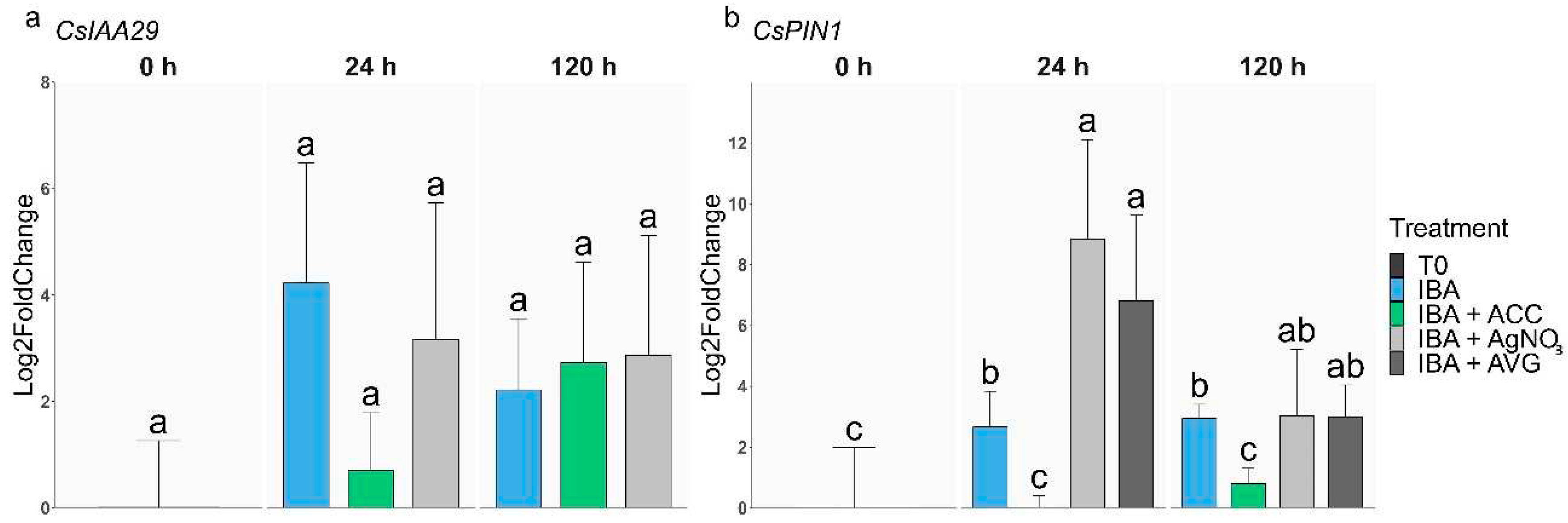
Figure 5.
qPCR expression analysis of genes related to epigenetics processes. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsJMJ30; (b) CsHDA14. All data were normalized to the expression of T0 sample. For each gene, different letters indicate statistical differences (p<0.05).
Figure 5.
qPCR expression analysis of genes related to epigenetics processes. Samples from P2CR shoots were subjected to different treatments: IBA 25 µM (IBA); IBA 25 µM supplemented with ACC 30 µM (IBA+ACC), with AVG 30 µM (IBA+AVG) or with AgNO3 30 µM (IBA+AgNO3) and collected 24h and 120h after the beginning of the treatments. (a) CsJMJ30; (b) CsHDA14. All data were normalized to the expression of T0 sample. For each gene, different letters indicate statistical differences (p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



