
Submitted:
29 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
Therapeutic hypothermia (TH) has emerged as a promising approach with neuroprotective effects that can mitigate secondary brain damage and intracranial pressure in the acute phase after successful mechanical thrombectomy. However, the effect of TH on cognitive impairment has yet fully elucidated. In the present study, we demonstrated whether TH can improve cognitive impairment in a transient middle cerebral artery occlusion (tMCAO) mouse model Nine-week-old C57BL/6N mice (male) were randomized to three groups: sham, tMCAO, and tMCAO with TH. One month after model induction, cognitive function test and regional cerebral glucose metabolism were evaluated using the Y-maze test and positron emission tomography with fluorine-18 fluorodeoxyglucose, respectively. tMCAO led to cognitive impairment, which was improved by TH. The cerebral glucose metabolism of the thalamus was significantly recovered in the TH group compared to that in the tMCAO group. These results suggest that TH might have potential as a therapeutic strategy for improving the ischemia/reperfusion-induced cognitive impairment.
Keywords:
Ischemic stroke
; Hypothermia
; Cognition
; Neuroprotection
1. Introduction
Mechanical thrombectomy (MT) has been extensively employed for restoring blood flow following acute occlusion of the intracranial artery [1]. Despite successful MT procedures leading to the restoration of blood flow, there exists the potential for reperfusion injury, which may exacerbate neurological outcomes. Specifically, reperfusion injury can instigate an elevation in reactive oxygen species (ROS), causing direct harm to mitochondria and lipid peroxidation [2]. Furthermore, activation of immune cells accompanies the upregulation of cytokines and chemokines. Consequently, ischemia-reperfusion injury induces chronic neurological dysfunction, notably cognitive impairment, through brain cell death. In the real clinical field, cognitive impairment has been reported in over 80% of patients with ischemic stroke [3,4]. Ettelt et al. reported that 86% of patients with acute large vessel artery occlusion who underwent MT had cognitive impairment three months after onset, even though they had a favorable functional outcome [3]. These findings underscore the importance for clinicians to be vigilant regarding cognitive impairment, which strongly correlates with reduced quality of life, emphasizing the need for swift recanalization efforts.
Body temperature initiates an ascent 4-6 hours post-stroke, with a more pronounced increase in severe cases [5]. Additionally, since the central nervous system is vulnerable to high temperatures, hyperthermia is closely associated with cognitive impairment [6]. Furthermore, patients with higher body temperatures during intra- and post-ischemic periods demonstrated unfavorable outcomes following MT [7]. Specifically, each 1°C increase in temperature significantly elevated the risk of impaired functional independence and mortality [7]. Therefore, therapeutic hypothermia (TH) has emerged as a promising strategy in clinical field to mitigate ischemia-reperfusion injury and achieve favorable neurological outcomes. Nevertheless, the majority of existing TH studies have predominantly focused on mitigating cerebral edema, potentially leading to increased intracranial pressure in the acute phase, there remains a scarcity of investigations exploring the effect of TH on cognitive function.
Considering the aforementioned facts, we hypothesized that TH could ameliorate impaired cognitive function by mitigating ischemia-reperfusion injury. To validate our hypothesis, we investigated the effects of TH on cognition by Y-maze test and evaluate the topographical changes in glucose metabolism using F-18 fluorodeoxyglucose positron emission tomography scan in a mouse model of transient middle cerebral artery occlusion.
2. Materials and Methods
Transient middle cerebral artery occlusion (tMCAO) model
SPF-grade male C57BL/6 mice (9 weeks old) were purchased from Samtako Bio (Osan, South Korea), and were bred in a controlled environment (22°C and 12 hours light/dark cycle, lights on at 6 AM) with free access to water and food. The animals were given an acclimatization period of a week before initiating tMCAO modeling. Following anesthesia induction with 4.0% isoflurane evaporated in an N2O/O2 (70:30) gas mixture, tMCAO or sham surgery was conducted as described in the previous study (Figure 1A) [8]. In brief, a 6-0 nylon monofilament with a silicone-coated tip (0.22 mm; Doccol Co., Sharon, MA, USA) was inserted through the external carotid artery into the right internal carotid artery and advanced to 6 mm from the internal carotid bifurcation site. Subsequently, the surgical site was temporarily covered, and anesthesia was stopped. Following a 60 min of occlusion, the mice were re- anesthetized for removal of the monofilament. The sham-operation group mice underwent an identical procedure to the tMCAO group, excluding the actual MCA occlusion. The success rate of tMCAO modeling in mice reached 93.3%. The specifics of the experimental design used in this study are shown in Figure 1A. The rectal temperature of the mice was carefully regulated at 37 ± 0.5°C during anesthesia, utilizing an infrared lamp and a heat blanket pad. Further, cerebral blood flow (CBF) was monitored throughout the experiment via laser Doppler flowmetry (Omega flow FLO-C1 BV; Omega Wave, Tokyo, Japan). Mice exhibiting a decrease in CBF exceeding 70% compared to the basal level were excluded from the analysis. Approval for all animal experiments was obtained from the Institutional Animal Care and Use Committee of Keimyung University (IACUC No: KM-2019-09R2), and the studies adhered to the principles outlined in the NIH Guide for the Care and Use of Laboratory Animals.
Therapeutic hypothermia (TH)
TH was conducted as described in the previous paper [9]. Briefly, a 70% alcohol spray was applied to the entire mouse body while rigorously controlling the core temperature within the range of 32-34°C. This hypothermic state was sustained for 2 h following reperfusion, after which the body temperature was gradually returned to 37°C [10,11]. Throughout the TH process, the temperature was tightly regulated using a DAS-7007R (BMDS, Seaford, DE) body temperature feedback system and an IPTT-300 transponder (BMDS, Seaford, DE).
Neurological function test
Global neurological deficits were quantified according to the Bederson score [12]. Briefly, the scope was based on the following information: 0 point: no deficit; 1 point: forelimb weakness; 2 points: circling to affected side; 3 points: partial paralysis on affected side; and 4 points: no spontaneous motor activity.
Cognitive function test
Cognitive function assessment was conducted through the Y-maze test, utilizing a maze featuring three arms constructed from white plexiglass. Each arm measures 40 cm in length, 12 cm height, and 4 cm width, with the arms positioned at 120° to each other. During 8 min testing period, mice were placed on one arm of the device and allowed free movement through the maze. Change behavior (%) was calculated as the ratio of actual change to possible change [13]. Video recordings of each trial were captured using a Smart video tracking system (Panlab-Harvard apparatus, MA, USA).
F-18 fluorodeoxyglucose (FDG) PET
To evaluate cerebral glucose metabolism, sham (n=9), tMCAO (n=11), and tMCAO+TH (n=12) group underwent an F-18 FDG PET at 1 month after tMCAO reperfusion, using the Triumph II PET/CT system (Lab-PET8; Gamma Medica-Ideas, Waukesha, WI, USA). Before the PET scan, the mice were fasted for 12 hours. They were sedated with 2.0% isoflurane in N2O/O2 (70:30) and injected intravenously with a dose of approximately 7.4 MBq of F-18 FDG into the tail vein. The mice underwent PET scanning approximately 30 min after F-18 FDG injection to acquire whole-brain images; the PET scan lasted for 5 min. The acquired data were assumed to indicate cerebral glucose metabolism. For spatiotemporal quantification of the cerebral glucose metabolism, a volume-of interest (VOI) analysis was performed for each scan with the use of the PMOD software package (PMOD Technologies, Ltd., Zurich, Switzerland) in conjunction with mouse brain template and atlas, as previously described, with some modifications [14,15]. PMOD was used to transform each of the mouse brain PET datasets to the appropriate space, and mouse brain atlas was automatically applied to measure the F-18 FDG uptake to acquire standardized F-18 FDG uptake values within defined subregions of mouse brain. The mouse brain VOI atlas was used in an iterative fashion with the standard brain model to further optimize the fusion of the experimental data. The regional standardized F-18 FDG uptake values ratio (SUVR) was calculated by dividing the standardized F-18 FDG uptake value for the individual target region by that for the corresponding region in the left cerebral hemisphere.
Cell culture
HT22 mouse hippocampal neuronal cells were grown in Dulbecco’s modified Eagle’s medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (Gibco/Thermo Fisher Scientific, Waltham, MA, USA) and 1% penicillin-streptomycin (Invitrogen/Thermo Fisher Scientific, Carlsbad, CA, USA). The cells were maintained in a humidified CO2 incubator (Sanyo, Osaka, Japan) at 37℃ and 5% CO2/95% air atmosphere and were sub-cultured when the confluence reached approximately 80% using a 0.05% trypsin-EDTA sodium salt solution (Welgene, Gyeongsan, South Korea).
In vitro hypoxia/reoxygenation (H/R) model
The HT22 cells were seeded into a 6-well plate at a density of 1 × 105 cells/well and cultured in DMEM supplemented with 10% fetal bovine serum and 1%penicillin in a 5% CO2 humid atmosphere at 37℃. After 24 hours, the H/R group of cells was cultured in a serum-free and glucose-free medium in a chamber with 0.5% O2 and 5% CO2 at 37℃ for 6 hours. Glucose (final concentration 11 mM) was added to hypoxia medium and the cells were cultured at 37℃ in a humidified 5% CO2 incubator for an additional 24 hours. In the therapeutic hypothermia group, cells were cultured at 32℃ following the hypoxia challenge.
Western blotting
The HT22 cells were collected and prepared for western blotting. Briefly, cellular proteins were extracted using RIPA buffer (ELPIS-BIOTECH, Daejeon, South Korea). The protein preparations were separated on sodium dodecyl sulfate polyacrylamide gel and transferred to polyvinylidene fluoride membranes (Merck Millipore Corp., Billerica, MA, USA). The proteins on the membranes were allowed to bind to primary antibodies against PARP, cleaved caspase-3 or α-tubulin and the appropriate secondary antibodies conjugated with horseradish peroxidase. Protein bands were then visualized using SuperSignal™ West Pico Chemiluminescent Substrate (Pierce, Cheshire, United Kingdom), digitalized using FUSIONSOLO5 (KOREA BIOMICS, Seoul, South Korea).
Statistical analysis
Statistical analyses were performed using SPSS software, version 27.0 (SPSS Inc., Chicago, IL, USA). Statistical differences between the means of two groups were tested by Student’s t-test. The results were considered significant at p values of less than 0.05. Statistical differences were indicated by asterisks.
3. Results
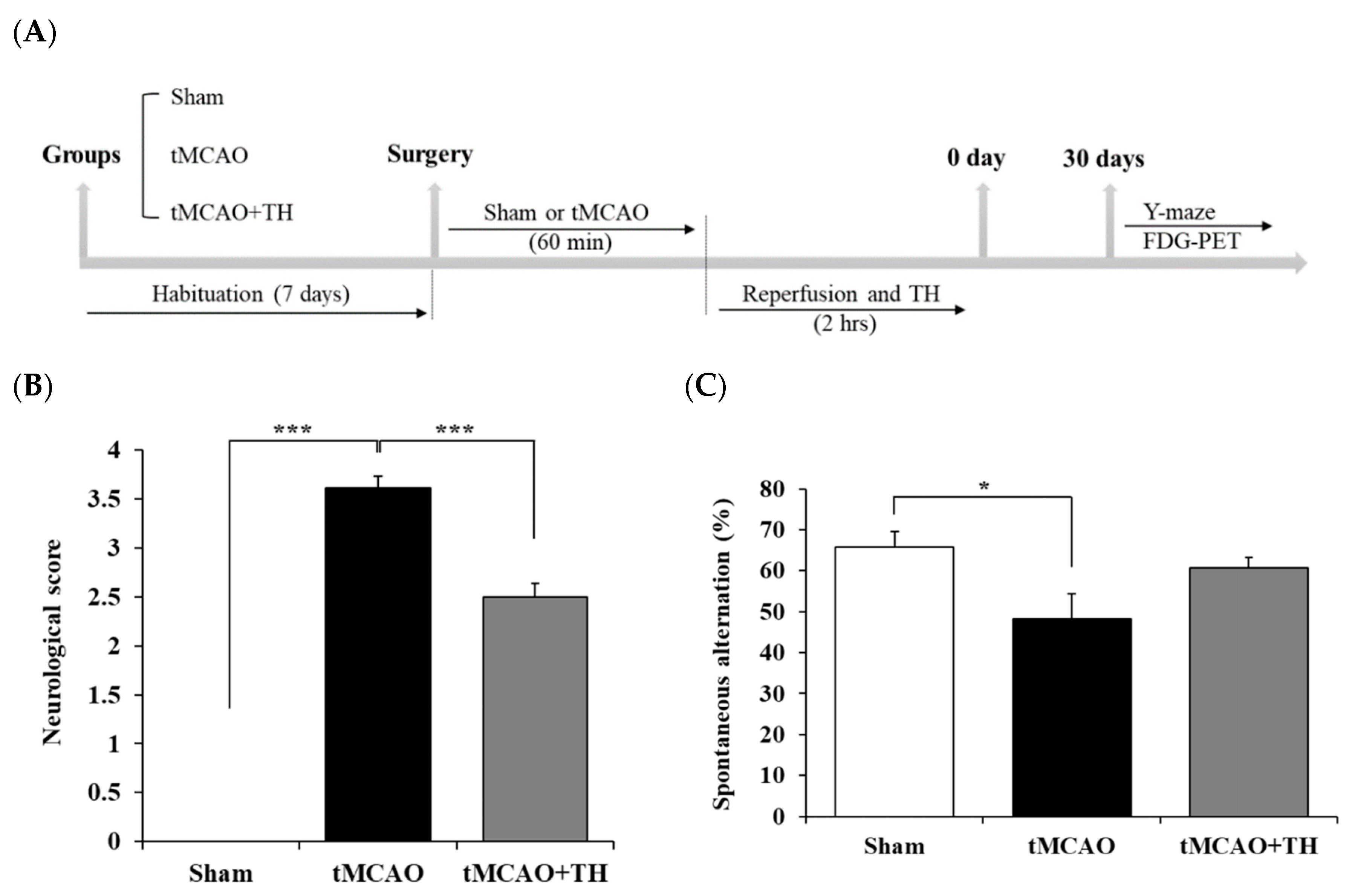
3.1. Hypothermia improved functional outcome and cognitive function
To evaluate the effect of TH on functional outcome in the tMCAO group, we used the Bederson neurological scoring system. Functional outcome was decreased in the tMCAO group and was improved in the tMCAO+TH group (Figure 1B). Next, to investigate the effect of TH on short-term memory, we conducted Y-maze behavior tests in tMCAO mouse model. The results showed that the percentage of spontaneous change was significantly decreased after tMCAO compared to sham controls (Figure 1C). TH increased the percentage in spontaneous changes compared with tMCAO without TH, although the change was not statistically significant (Figure 1C). These data suggest that TH tended to improve spatial motor memory reduced by tMCAO.
Figure 1.
Comparison of neurological score and spatial working memory among sham, tMCAO, and tMCAO+TH groups. (A) Experimental schedule for FDG-PET and behavioral study using tMCAo mouse model. (B) Neurological score test was performed after surgery. (C) Y-maze test was performed at 30 days after surgery. Data were expressed as mean ± SEM and statistical analysis was performed by student’s t-test. (*p < 0.05, ***p < 0.001).
Figure 1.
Comparison of neurological score and spatial working memory among sham, tMCAO, and tMCAO+TH groups. (A) Experimental schedule for FDG-PET and behavioral study using tMCAo mouse model. (B) Neurological score test was performed after surgery. (C) Y-maze test was performed at 30 days after surgery. Data were expressed as mean ± SEM and statistical analysis was performed by student’s t-test. (*p < 0.05, ***p < 0.001).
3.2. Cerebral glucose metabolism
To investigate the effect of TH on cerebral glucose metabolism, F-18 FDG PET was performed in the sham, tMCAO, and tMCAO+TH groups. Regional SUVRs were determined using VOI analysis (Figure 2). At 1month after the tMCAO surgery, the SUVRs of the right hippocampus, left striatum, right striatum, left thalamus, right thalamus, and central gray were significantly lower in the tMCAO group than those in the sham group, and left thalamus was significantly increased in the tMCAO+TH group compared to tMCAO group (Table 1). However, there were no significant differences in the SUVR in other regions of the brain.
3.3. Therapeutic hypothermia attenuated H/R-induced neuronal apoptosis
To verify the effect of TH on cognitive impairment in tMCAO model, we established in vitro H/R model. Following the H/R challenge, the viability of neuronal cells was decreased, and PARP cleavage and cleaved-caspase-3 were increased (Figure 3). However, TH restored the decreased neuronal viability and the expression levels of increased PARP cleavage and cleaved-caspase-3 induced by H/R.
4. Discussion
Ischemic stroke has been identified as a precipitating factor in selective neuronal degeneration and neuroinflammation, leading to consequential cognitive impairment [16] . Cognitive deficits are present in over 70% of stroke survivors [17]. These deficits encompass not only impairments specific to the stroke lesion site, such as aphasia or memory deficits, but also extend to those arising from strategic infarcts in the hippocampus, thalamus, and key cortical regions [18]. Additionally, it encompasses deficits that may have anteceded the onset of the stroke.
Mild to moderate hypothermia (31-34°C) during or after cerebral ischemia emerges as a potent neuroprotective strategy [19-25]. This therapeutic approach not only concurrently inhibits multiple mechanisms of brain cell death but also decelerates metabolic processes, effectively limiting tissue damage [23,26–28]. These cumulative or synergistic benefits of hypothermia offer promising clinical effects for stroke patients.
Cerebral glucose hypometabolism constitutes a major feature of dementia [29]. The hypometabolism pattern identified through F-18 FDG PET has predictive value for progression from baseline cognition to mild cognitive impairment [30]. Beyond its role as a biomarker, cerebral glucose hypometabolism significantly contributes to the pathophysiology of Alzheimer’s disease [31,32]. Human clinical trials aimed at restoring cerebral bioenergetic pathways have yielded promising early results [31]. Our study showed that tMCAO decreased glucose metabolism and improved after TH in the thalamus. Several studies reported that cognitive dysfunction subsequent to acute stroke event is often attributed to direct injury to brain structures crucial for cognitive function such as the thalamus or basal ganglia [17,33,34] which was consistent with our results. The Y-maze test also shown that decreased short-term memory in tMCAO group is recovered after TH. Considered together, TH might provide a potential adjuvant therapeutic option for vascular stroke. We demonstrated that TH can be effectively induced at the tMCAO affected injured regions, resulting in neuro-regenerative effects and behavior recovery.
Our study has several limitations that warrant consideration. Firstly, the assessment of TH effect on cognitive impairment was conducted in relative healthy young male mice. The extrapolation of these findings to actual patients may encounter variations, as clinical populations often include individuals with advanced atherosclerosis and elderly patients with have experienced ischemic stroke. Secondly, the therapeutic impact of hypothermia on cognitive impairment may exhibit variability contingent on the degree of ischemic-reperfusion injury. The extent of the injury following successful recanalization is influenced by factors such as the duration of occlusion, underlying diseases, and collateral circulation. Thus, additional studies are essential to investigate the cognitive function recovery effects of TH in mouse models representing atherosclerosis or elderly populations. Thirdly, our study did not comprehensively assess damage to brain structures and isoflurane concentration, both of which could potentially influence cognitive function. For example, the thalamus is a part of the integral neuronal network responsible for cognition [35]. Moreover, recent study suggested that cognitive impairment ensues through the fronto–parieto– cerebellar–thalamic loop [36]. Thalamic injury has been demonstrated to cause a decline in word retrieval, particularly in relation to responses from the posterior supplementary motor area. In this study, a concentration of 4.0% isoflurane was utilized, potentially influencing cellular damage and thereby impacting cognitive function [37,38]. Finally, we did not investigate neuroinflammation in detail. Neuroinflammation, either by itself or through interactions with Aβ protein and tau proteins, has the potential to impair cognitive function. Considering these findings together, further studies are needed to examining thalamic damage, isoflurane concentration, and neuroinflammation following TH in a model of ischemic-reperfusion injury.
5. Conclusions
Our finding suggests that TH after tMCAO in mice improves the impairment of cognitive function, without causing concomitant long-term sensorimotor deficits. It also shows that mice could be a suitable alternative to rats for studying the effects of potential stroke therapies on cognitive outcomes. Further studies are needed to elucidate the mechanisms which induce tMCAO ischemia and TH treatment and will investigate the link between the candidate and the brain function.
Author Contributions
H.W.K., and J-H.H. conceptualized and designed the study. J.S.L., M-N.P. acquired and analysed the experimental data. HS.L., H.S.B. and J.K.K discussed analyses and interpreted the data. J.SLJ., and S.K. wrote the original draft. H.W.K. and J-H.H. reviewed and revised the manuscript. H.W.K. and J-H.H. supervised and financially supported the study.
Funding
This research was funded by the National Research Foundation of Korea (NRF) Grant funded by the Korea Government (MSIP) (No. 2017R1C1B507687314).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Blanc R, Escalard S, Baharvadhat H, Desilles JP, Boisseau W, Fahed R; et al. Recent advances in devices for mechanical thrombectomy. Expert Rev Med Devices. 2020, 17, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Mizuma A, You JS, Yenari MA. Targeting Reperfusion Injury in the Age of Mechanical Thrombectomy. Stroke. 2018, 49, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Ettelt P, Maier IL, Schnieder M, Bahr M, Behme D, Psychogios MN; et al. Bridging therapy is associated with improved cognitive function after large vessel occlusion stroke - an analysis of the German Stroke Registry. Neurol Res Pract. 2020, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Sun JH, Tan L, Yu JT. Post-stroke cognitive impairment: Epidemiology, mechanisms and management. Ann Transl Med. 2014, 2, 80. [Google Scholar]
- Boysen G, Christensen H. Stroke severity determines body temperature in acute stroke. Stroke. 2001, 32, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Walter EJ, Carraretto M. The neurological and cognitive consequences of hyperthermia. Crit Care. 2016, 20, 199. [Google Scholar] [CrossRef] [PubMed]
- Diprose WK, Liem B, Wang MTM, Sutcliffe JA, Brew S, Caldwell JR; et al. Impact of Body Temperature Before and After Endovascular Thrombectomy for Large Vessel Occlusion Stroke. Stroke. 2020, 51, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Sawada M, Alkayed NJ, Goto S, Crain BJ, Traystman RJ, Shaivitz A; et al. Estrogen receptor antagonist ICI182, 780 exacerbates ischemic injury in female mouse. Journal of Cerebral Blood Flow & Metabolism. 2000, 20, 112–118. [Google Scholar]
- Lee H, Park M-N, Lim JS, Baek HS, Kim S, Kim HW; et al. The Effect of Hypothermia on Cognitive Impairment and Anxiety-like Behavior in a Cardiac Arrest Mouse Model. Quant BioSci. 2023:9-16.
- Maier CM, Ahern Kv, Cheng ML, Lee JE, Yenari MA, Steinberg GK. Optimal depth and duration of mild hypothermia in a focal model of transient cerebral ischemia: Effects on neurologic outcome, infarct size, apoptosis, and inflammation. Stroke. 1998, 29, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Tang Y, Liu X, Zhao J, Tan X, Liu B, Zhang G; et al. Hypothermia-induced ischemic tolerance is associated with Drp1 inhibition in cerebral ischemia-reperfusion injury of mice. Brain Research. 2016, 1646, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hattori K, Lee H, Hurn PD, Crain BJ, Traystman RJ, DeVries AC. Cognitive deficits after focal cerebral ischemia in mice. Stroke. 2000, 31, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Choi M, Lee Y, Cho S-H. Angelica tenuissima Nakai ameliorates cognitive impairment and promotes neurogenesis in mouse model of Alzheimer’s disease. Chinese journal of integrative medicine. 2018, 24, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Park J-H, Hong J-H, Lee S-W, Ji HD, Jung J-A, Yoon K-W; et al. The effect of chronic cerebral hypoperfusion on the pathology of Alzheimer's disease: A positron emission tomography study in rats. Scientific reports. 2019, 9, 14102. [Google Scholar] [CrossRef] [PubMed]
- Casteels C, Vunckx K, Aelvoet S-A, Baekelandt V, Bormans G, Van Laere K; et al. Construction and evaluation of quantitative small-animal PET probabilistic atlases for [18F] FDG and [18F] FECT functional mapping of the mouse brain. PLoS ONE. 2013;8:e65286.
- Thiel A, Cechetto DF, Heiss W-D, Hachinski V, Whitehead SN. Amyloid burden, neuroinflammation, and links to cognitive decline after ischemic stroke. Stroke. 2014, 45, 2825–2829. [Google Scholar] [CrossRef]
- Rost NS, Brodtmann A, Pase MP, van Veluw SJ, Biffi A, Duering M; et al. Post-stroke cognitive impairment and dementia. Circulation Research. 2022, 130, 1252–1271. [Google Scholar] [CrossRef] [PubMed]
- Dichgans M, Leys D. Vascular cognitive impairment. Circulation research. 2017, 120, 573–591. [Google Scholar] [CrossRef]
- Fisher M, Feuerstein G, Howells DW, Hurn PD, Kent TA, Savitz SI; et al. Update of the stroke therapy academic industry roundtable preclinical recommendations. Stroke. 2009, 40, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Kallmünzer B, Kollmar R. Temperature management in stroke–an unsolved, but important topic. Cerebrovascular Diseases. 2011, 31, 532–543. [Google Scholar] [CrossRef]
- Faridar A, Bershad EM, Emiru T, Iaizzo PA, Suarez JI, Divani AA. Therapeutic hypothermia in stroke and traumatic brain injury. Frontiers in neurology. 2011, 2, 80. [Google Scholar]
- Tveita T, Sieck GC. Physiological impact of hypothermia: The good, the bad, and the ugly. Physiology. 2022, 37, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Tahir RA, Pabaney AH. Therapeutic hypothermia and ischemic stroke: A literature review. Surgical Neurology International. 2016;7:S381.
- Xue D, Huang Z-G, Smith KE, Buchan AM. Immediate or delayed mild hypothermia prevents focal cerebral infarction. Brain research. 1992, 587, 66–72. [Google Scholar] [CrossRef]
- Barber PA, Hoyte L, Colbourne F, Buchan AM. Temperature-regulated model of focal ischemia in the mouse: A study with histopathological and behavioral outcomes. Stroke. 2004, 35, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Yenari MA, Hemmen TM. Therapeutic hypothermia for brain ischemia: Where have we come and where do we go? Stroke. 2010;41:S72-S4.
- Wu D, Chen J, Zhang X, Ilagan R, Ding Y, Ji X. Selective therapeutic cooling: To maximize benefits and minimize side effects related to hypothermia. Journal of Cerebral Blood Flow & Metabolism. 2022, 42, 213–215. [Google Scholar]
- Zhang S, Zhang X, Zhong H, Li X, Wu Y, Ju J; et al. Hypothermia evoked by stimulation of medial preoptic nucleus protects the brain in a mouse model of ischaemia. Nature Communications. 2022, 13, 6890. [Google Scholar] [CrossRef] [PubMed]
- Butterfield DA, Halliwell B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat Rev Neurosci. 2019, 20, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Kato T, Inui Y, Nakamura A, Ito K. Brain fluorodeoxyglucose (FDG) PET in dementia. Ageing research reviews. 2016, 30, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kellar D, Craft S. Brain insulin resistance in Alzheimer's disease and related disorders: Mechanisms and therapeutic approaches. The Lancet Neurology. 2020, 19, 758–766. [Google Scholar] [CrossRef] [PubMed]
- An Y, Varma VR, Varma S, Casanova R, Dammer E, Pletnikova O; et al. Evidence for brain glucose dysregulation in Alzheimer's disease. Alzheimer's & dementia. 2018, 14, 318–329. [Google Scholar]
- Weaver NA, Kuijf HJ, Aben HP, Abrigo J, Bae H-J, Barbay M; et al. Strategic infarct locations for post-stroke cognitive impairment: A pooled analysis of individual patient data from 12 acute ischaemic stroke cohorts. The Lancet Neurology. 2021, 20, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao L, Biesbroek JM, Shi L, Liu W, Kuijf HJ, Chu WW; et al. Strategic infarct location for post-stroke cognitive impairment: A multivariate lesion-symptom mapping study. Journal of Cerebral Blood Flow & Metabolism. 2018, 38, 1299–1311. [Google Scholar]
- Karussis D, Leker R, Abramsky O. Cognitive dysfunction following thalamic stroke: A study of 16 cases and review of the literature. Journal of the neurological sciences. 2000, 172, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, S. Cognitive and linguistic dysfunction after thalamic stroke and recovery process: Possible mechanism. AIMS neuroscience. 2022, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Borgstedt L, Bratke S, Blobner M, Pötzl C, Ulm B, Jungwirth B; et al. Isoflurane has no effect on cognitive or behavioral performance in a mouse model of early-stage Alzheimer's disease. Frontiers in Neuroscience. 2022;16.
- Lin D, Zuo Z. Isoflurane induces hippocampal cell injury and cognitive impairments in adult rats. Neuropharmacology. 2011, 61, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Cerebral glucose metabolism measured by F-18 FDG PET. (A) Regional standardized F-18 FDG uptake values (SUVRs). (B) A sham-operated mouse showing no abnormal glucose metabolism. (C) tMCAO mouse showing a decreased glucose metabolism of the hemisphere on the right side. (D) tMCAO+TH mouse showing no change of glucose metabolism on the right side compared to left side. TH upregulated glucose metabolism in tMCAO induced ischemia brain.
Figure 2.
Cerebral glucose metabolism measured by F-18 FDG PET. (A) Regional standardized F-18 FDG uptake values (SUVRs). (B) A sham-operated mouse showing no abnormal glucose metabolism. (C) tMCAO mouse showing a decreased glucose metabolism of the hemisphere on the right side. (D) tMCAO+TH mouse showing no change of glucose metabolism on the right side compared to left side. TH upregulated glucose metabolism in tMCAO induced ischemia brain.
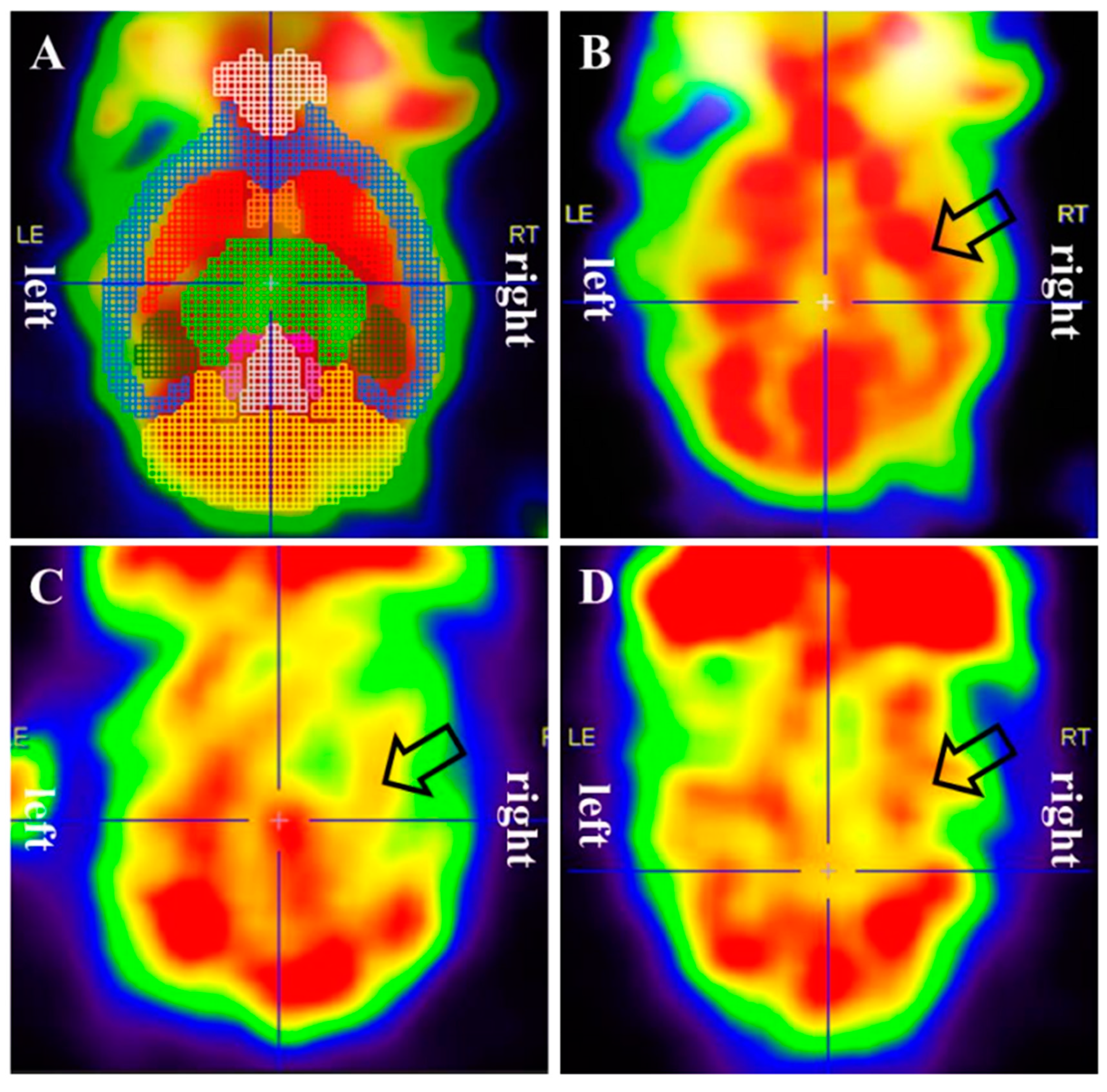
Figure 3.
Therapeutic hypothermia attenuated H/R-induced neuronal apoptosis by modulating the expression of cleaved-PARP and cleaved-caspase3. (A) The experimental scheme for the in vitro hypoxia/reoxygenation model is presented. (B) Cell viability assay was performed after H/R with therapeutic hypothermia (TH) or without. (C) The expression levels of apoptotic protein markers were assessed using western blotting. Data were expressed as mean ± SEM and statistical analysis was conducted using the student’s t-test. (***; compared to normoxia, p < 0.001, ##; compared to H/R, p < 0.01).
Figure 3.
Therapeutic hypothermia attenuated H/R-induced neuronal apoptosis by modulating the expression of cleaved-PARP and cleaved-caspase3. (A) The experimental scheme for the in vitro hypoxia/reoxygenation model is presented. (B) Cell viability assay was performed after H/R with therapeutic hypothermia (TH) or without. (C) The expression levels of apoptotic protein markers were assessed using western blotting. Data were expressed as mean ± SEM and statistical analysis was conducted using the student’s t-test. (***; compared to normoxia, p < 0.001, ##; compared to H/R, p < 0.01).
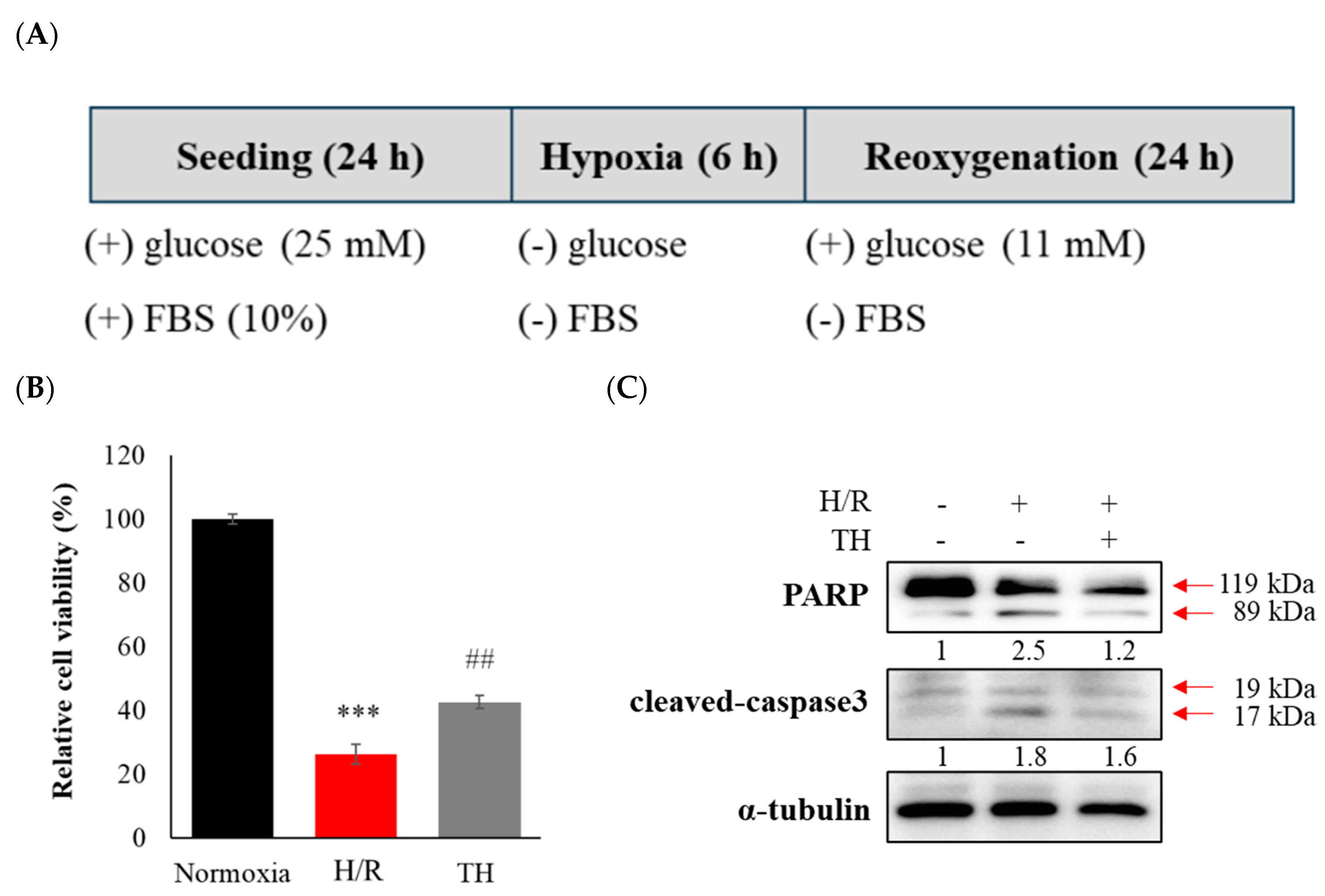

Table 1.
Comparison of regional cerebral glucose metabolism after tMCAO surgery.
| Regions | Side | Mean (standard deviation) | p-value | |||||
| Sham | tMCAO | tMCAO+TH | Sham vstMCAO | Sham vstMCAO+TH | tMCAO vstMCAO+TH | |||
| Amygdala | left | 0.828 (0.081) | 0.904 (0.064) | 0.890 (0.066) | 0.115 | 0.213 | 1.000 | |
| right | 0.883 (0.107) | 0.856 (0.087) | 0.904 (0.088) | 1.000 | 1.000 | 0.883 | ||
| Cortex | left | 0.960 (0.036) | 0.970 (0.043) | 0.943 (0.035) | 1.000 | 1.000 | 0.438 | |
| right | 0.998 (0.085) | 0.948 (0.107) | 0.942 (0.094) | 0.867 | 0.624 | 1.000 | ||
| Hippocampus | left | 1.141 (0.070) | 1.062 (0.079) | 1.151 (0.079) | 0.134 | 1.000 | 0.065 | |
| right | 1.187 (0.109) | 1.062 (0.078) | 1.162 (0.102) | 0.044* | 1.000 | 0.127 | ||
| Midbrain | left | 1.280 (0.131) | 1.175 (0.086) | 1.260 (0.108) | 0.193 | 1.000 | 0.362 | |
| right | 1.282 (0.152) | 1.143 (0.081) | 1.243 (0.135) | 0.106 | 1.000 | 0.345 | ||
| Striatum | left | 1.165 (0.078) | 1.082 (0.031) | 1.133 (0.069) | 0.040* | 0.887 | 0.307 | |
| right | 1.191 (0.116) | 1.074 (0.052) | 1.145 (0.091) | 0.043* | 0.845 | 0.345 | ||
| Thalamus | left | 1.224 (0.086) | 1.102 (0.049) | 1.213 (0.079) | 0.007** | 1.000 | 0.012* | |
| right | 1.280 (0.136) | 1.135 (0.067) | 1.248 (0.113) | 0.038* | 1.000 | 0.127 | ||
| Inferior colliculi | left | 1.336 (0.165) | 1.205 (0.074) | 0.265 (0.084) | 0.083 | 0.570 | 0.850 | |
| right | 1.390 (0.209) | 1.222 (0.083) | 1.331 (0.124) | 0.089 | 1.000 | 0.412 | ||
| Superior colliculi | 1.318 (0.152) | 1.182 (0.053) | 1.261 (0.099) | 0.056 | 0.827 | 0.438 | ||
| Basal forebrain | 1.046 (0.041) | 1.031 (0.037) | 1.006 (0.042) | 1.000 | 0.126 | 0.602 | ||
| Central gray | 1.434 (0.177) | 1.237 (0.079) | 0.328 (0.138) | 0.023* | 0.325 | 0.551 | ||
| Hypothalamus | 1.078 (0.086) | 1.043 (0.058) | 1.083 (0.053) | 0.899 | 1.000 | 0.675 | ||
| Olfactory bulb | 1.377 (0.135) | 1.321 (0.173) | 1.248 (0.142) | 1.000 | 0.220 | 0.944 | ||
Asterisks indicate statistical significance. *p < 0.05, **p < 0.01; All values are presented as mean (standard deviation).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



