Submitted:
30 December 2023
Posted:
30 December 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The COVID-19 pandemic, declared in early 2020, is an unprecedented global health crisis, causing over 7.0 million deaths and ongoing challenges. While the pharmaceutical industry expedited vaccine development, mutant SARS-CoV-2 strains remain a major fear. Moreover, concerns regarding the long-term health repercussions of COVID-19-affected individuals persist since individuals affected by mild and moderate to severe SARS-CoV-2 infection experience long-term cardiovascular complications, liver dysfunction, pulmonary afflictions, kidney impairments, and most importantly neurocognitive deficits. In recent studies, we documented pathophysiological changes in various organs following the post-acute infection of mice with murine hepatitis virus-1 (MHV-1), a coronavirus, at both 7 days and 12 months after infection. One part of the body that can be drastically affected by SARS-CoV-2 is the skin. Studies have shown major changes in the skin post-acute SARS CoV-2 infection in humans. However, long-term dermatologic changes post-COVID have never been explored. For the first time, we show several cutaneous findings both at the acute stages and long-term post-infection of mice with MHV-1 coronavirus (a promising experimental model to study acute and long-COVID). Precisely, we found destruction of the epidermal layer, an increase in the number of hair follicles, extensive collagen deposition in the dermal layer, and hyperplasticity of the sebaceous glands at the acute stages, along with thinning of the panniculus carnosus, as well as the adventitial layer, which corresponds well with studies in humans. In contrast, the cutaneous investigation in the long-COVID phase shows the absence of hair follicles from both the epidermal and dermal layers, the destruction of adipose tissues, and the devastation of the epidermal layer. Further, treatment of these mice with a 15 amino acid synthetic peptide, SPIKENET (SPK), which was effective in preventing Spike glycoprotein-1 binding with host receptors, as well as has a potent anti-inflammatory response to severe inflammatory stimulus) restored the loss of hair follicles and re-architected the epidermal and dermal layers. Additionally, destruction in fatty tissue in the infected mice was successfully restored post-treatment with SPK. These findings suggest that SARS-CoV-2 initiates the changes early post-infection, leading to devastating skin alterations in the long term which can be prevented by our newly identified peptide drug SPK.
Keywords:
Dermatologic manifestation
; fibrosis
; inflammation
; long-COVID
; murine hepatitis virus-1.
1. Introduction
The global impact of the COVID-19 pandemic persists, with millions affected and ongoing morbidity and mortality. As of December 2023, deaths have exceeded 7.7 million since the pandemic declaration in early 2020, accompanied by a daily influx of over 39 thousand new cases worldwide [1]. On a positive note, 767 million individuals have successfully recovered from mild to severe SARS-CoV-2 infections out of 775 million affected individuals [1]. Despite concerted global efforts to develop vaccines aimed at reducing mortality, no specific treatment exists to date for the viral infection [1,2,3,4].
Beyond the acute complications of COVID-19, survivors grapple with enduring health issues, encompassing disrupted sleep patterns, osteoporosis, subfertility, exacerbated diabetes, fatigue, and complications affecting various organ systems, including musculoskeletal, cardiovascular, gastrointestinal, pulmonary, neurologic, and urologic [1,2,5,6,7,8]. The intricate interplay of SARS-CoV-2 genetic mutations and the array of documented post-infection complications challenge scientists in pinpointing the exact pathophysiology. Furthermore, the progression of disease pathogenesis, spanning weeks to months, reveals signs of multiple organ dysfunction or failure occurring in one or more episodes of post-acute SARS-CoV-2 infection - referred to as post-acute sequelae of SARS-CoV-2 infection with or without overt symptoms and behavioral changes.
Studies have shown clinical syndrome resembling severe acute respiratory syndrome (SARS), marked by a notably high mortality rate [9,10,11,12,13,14,15,16,17,18] in mice, infected with murine hepatitis virus-1 (MHV-1). These mice exhibit pronounced lung injury with mortality rates ranging from 40% to 60% between days 7 and 12 post-infection [18]. Upon examination at the time of death, the lungs display severe interstitial pneumonitis characterized by interstitial inflammatory reactions, along with substantial infiltrations of lymphocytes and macrophages [16,18]. Additionally, investigations into the livers of MHV-1-infected A/J mice disclose evidence of severe hepatic congestion, mirroring observations in humans infected with SARS-CoV-2 [16,18].
In addition to multi-organ dysfunction, skin changes associated with COVID-19 have been established in acute stages post-infection which encompass a range of manifestations that can vary widely among individuals. One common type is the maculopapular rash, characterized by red macules, and papules. In some cases, individuals with COVID-19 exhibit a vesicular rash, consisting of fluid-filled blisters that can resemble those seen in chickenpox or herpes infections [19,20]. However, this presentation is less common compared to other skin appearances. COVID toes are another distinctive skin change, involving red or purple discoloration of the toes and fingers, often accompanied by swelling and pain [20]. This phenomenon is more frequently observed in younger individuals afflicted with COVID-19 and has been associated with milder cases of illness. Urticaria, or hives, is characterized by raised, itchy welts on the skin and can occur independently or alongside other COVID-19 symptoms [21,22]. Petechiae and purpura, small red or purple spots, and larger discolorations, respectively, may also be present, particularly in severe cases due to coagulation abnormalities associated with the virus or vasculitis [22,23,24,25,26,27,28]. In some cases, a mottled, purplish discoloration known as livedo reticularis was developed, thus forming a net-like pattern on the skin [25]. This is often associated with changes in blood flow and is observed in severe cases, especially in individuals with pre-existing vascular issues [25,28].
Given the common genus shared by MHV severe acute respiratory syndrome coronavirus and SARS-CoV-2, insights gleaned from MHV-1 could potentially provide mechanistic understanding into SARS-CoV-2 infection in humans [1,2,3]. While there are notable similarities between MHV-1 in mice and SARS-CoV-2 in humans, such as certain pathogenic features, differences exist (e.g., there are variations in viral binding receptors ACE2 in SARS-CoV-2 versus carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) in MHV-1 [15,16,18,29]. Additionally, distinctions include the proteolytic cleavage of four crucial amino acids at the S1/S2 site of the SARS-CoV-2 spike protein [29,30,31,32]. Despite these variances, the observed similarities outweigh the differences. Importantly, our identification of pathological and functional changes in the MHV-1 mice model of COVID-19 demonstrates a high degree of comparability to humans afflicted with SARS-CoV-2 infection [15,16,18,29].
In this study, we aimed to investigate the association between acute and long-term dermatological changes in COVID-19. We further examined whether inhibition of viral infection by a newly identified 15-amino-acid synthetic peptide, SPIKENET (SPK, which was effective in preventing Spike glycoprotein-1 binding with host receptors, as well as has a potent anti-inflammatory response to severe inflammatory stimulus), prevents or ameliorates skin alterations.
2. Materials and Methods
2.1. Mice
We used 8-week-old female A/J mice weighing 22–24 grams each. These mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA), and were kept in cages at the University of Miami Miller School of Medicine animal isolation facility. The animals were fed with a standard lab chow diet (Envigo 2918 irradiated, Teklad diet, Dublin, VA, USA) and provided with water ad libitum (autoclaved tap water). The study was performed according to the guidelines of the University of Miami Institutional Animal Care and Use Committee (IACUC protocol number 20-131 LF/Renewed protocol number, 20-162).
2.2. Viral (MHV-1) Inoculation and SPIKENET
Treatment MHV-1 was purchased from the American Type Culture Collection (ATCC, cat# VR 261, Manassas, VA, USA). Mice were split into (1) a healthy control group, (2) an MHV-1 virus-inoculated group, and (3) an MHV-1 virus-inoculated group that is treated with SPIKENET (SPK). MHV-1 viral inoculation was performed as previously described [11,15,16,18,29]. Briefly, groups 2 and 3 have been inoculated intranasally with 5000 PFU MHV-1 and were observed to ensure adequate inhalation of the virus. MHV-1 inoculated mice (group 3) were additionally treated with 5 mg/kg body weight of SPIKENET (SPK) as previously described by us [15,16,18,29].
2.3. Skin Collection and Storage
The skin of the mice was collected, and hair was removed and placed in 10% formalin (7 days post-infection for acute studies and 12 months post-infection for long-term studies). Paraffin embedded sections (processed through Histoscore PELORIS 3 Premium Tissue Processing System, Leica Biosystems Inc., Buffalo Grove, IL, USA), were then cut into 10 µm thick pieces by an ultra-thin semiautomatic microtome (Histoscore auto cut automated rotary microtome, Leica Biosystems Inc., Buffalo Grove, IL, USA).
2.4. Histological staining
Histological staining of the mice skin with Hematoxylin and Eosin (H&E) was performed using the following materials and reagents; formalin-fixed paraffin-embedded mice skin tissue sections, xylene, absolute ethanol, 95% ethanol, 70% ethanol, hematoxylin solution, eosin Y solution, distilled water, microscope slides, and cover slips [30,31]. Initially, deparaffinization was performed through the following steps: the slides were placed in xylene for 5 minutes followed by absolute ethanol for 3 minutes, immersed in 95% ethanol for 2 minutes, then placed in 70% ethanol for 2 minutes [30,31]. The tissue sections were rinsed with distilled water. The hematoxylin staining was done by adding the slides in hematoxylin solution for 5-10 minutes, then rinsing with distilled water to remove excess stain, and differentiating the slides with 1% acid alcohol until sections turned blue. The slides were then rinsed again in distilled water [30,31]. Eosin staining was established by exposing slides to eosin Y solution for 2-3 minutes and then rinsed with distilled water [30,31]. The slides underwent dehydration step through adding them in (70%, 95%, and absolute ethanol), then being placed in xylene for 5 minutes. The slides were then mounted in mounting media and the cover slips were applied [30,31].
3. Results
Diverse histological alterations were discerned during our scrutiny of MHV-1 infection within the murine integument, delving into the acute, long-COVID, and SPK treatments. In the mature murine cohort, the epidermal stratum exhibited a characteristic semblance, showcasing the canonical quartet of epithelial-derived cell strata identified in analogous taxa: the stratum basale, stratum spinosum, stratum granulosum, and the keratinized stratum corneum. The stratum basale, accommodating actively proliferating cells, emerged as the exclusive stratum where mitotic activity attained visibility in unblemished cutaneous tissue. Noteworthy, apparent basophilic granules were identified within the stratum granulosum. Further, a melanin pigmentation manifested within the cells of the stratum basale, as captured in Figure 1A & C.
The cutaneous histopathology displays a pronouncedly compromised epidermal stratum characterized by akeratosis, papillomatosis, and instances of apoptotic events among the epidermal cells. Furthermore, we found conspicuous features encompassing hyperplasticity in sebaceous gland development and pronounced deposition of collagens within the lower epidermal and dermal strata. A discernible augmentation in the number of anagenic hair follicles is observed concomitant with heightened sebaceous gland development. Noteworthy, the attenuation of the panniculus carnosus and adventitial layer coupled with dermal thickening attributable to the extensive destruction/obliteration of the fatty tissue layer and architectural modification throughout the skin strata were observed (Figure 1B, D & Figure 2D). Moreover, we detected a complete abolition of select epidermal regions (Figure 2B), as compared to control mice that were not inoculated with the virus (Figure 2A). Additionally, the presence of bullae dispersed within the dermal layer of the infected mice as illustrated in figure 2D, in stark contrast to the normative cutaneous morphology identified in non-infected mice, depicted in Figure 2C.
We next examined potential cutaneous alterations in the long-COVID phase, precisely 12 months post-MHV-1 infection. The histopathological paradigm is illustrated in Figure 3B, showing pronounced architectural perturbation, extensive attenuation of the epidermal stratum, keratinocyte loss, diverse apoptotic bodies, and the conspicuous absence of hair follicles within the dermal stratum (Figure 3B & Figure 4B). Comparative analysis with the control group reveals a discernible thinning of the panniculus carnosus in the long-COVID cohort (Figure 3A & Figure 4A). Concurrently, we observed adipose tissue degeneration/fibrosis concomitant with the obliteration of the adventitial layer (Figure 3B), phenomena reminiscent of those observed during the acute phase infection.
Remarkably, treatment of the MHV-1 infected mice with the synthetic peptide SPIKENET (SPK) exhibited substantial amelioration of cutaneous changes discerned in long-COVID. Notably, SPK intervention effectively reinstated the morphogenesis of hair follicles (Figure 3D) in stark contrast to the untreated control group (Figure 3C). Fascinatingly, this therapeutic intervention demonstrated the restoration of architectural integrity not only in the adipose tissue but also across the dermal and epidermal strata, encompassing the sebaceous glands (Figure 4D) juxtaposed to the untreated control mice (Figure 4C). Additionally, the sebaceous glands exhibit conspicuous dispersion along the length of hair follicles, accompanied by a discernible augmentation in the thickening of the panniculus carnosus (Figure 3D), which is notably absent in the untreated control group (Figure 3C).
In Figure 5 (B & C), diverse stages of hair follicle degeneration are evident long-term post-infection, as juxtaposed with the normal, healthy mice illustrated in Figure 5A. The application of SPK in infected mice exhibited the restoration of hair follicles across distinct stages, along with the regeneration of adipose tissue. Figure 6, portraying acutely infected mice (A-D), revealed microthrombosis (indicated by black and brown arrows in A & B), dermal edema (depicted by the blue arrow in A), keratinocyte apoptosis (highlighted by the green arrow in B), apoptotic cells surrounding a dying hair follicle encircled by edema, and Merkel cells surrounded by edema (marked by red/blue and black arrows in C). Furthermore, in D, observations included edema around hair follicles, thrombotic vessels, a group of melanocytes, some of which are undergoing degeneration, and intra-keratinocyte vacuolization (represented by red, black, blue, and orange arrows, respectively). In Figure 7, notable alterations were observed. Subfigure A exhibits the enlargement of sebaceous cells with intracellular vacuolization (denoted by the black arrow), while the red arrow indicates the presence of an eosinophil. Subfigure B depicts bullae, and Subfigure D reveals a nest of inflammation (indicated by the blue arrow), and thrombotic vessels (marked by the black arrow), with the red arrow highlighting melanocyte intracellular edema. Conversely, the green arrow points to the presence of edema.
4. Discussion
The histopathological changes in acute and long-term post-COVID have not been investigated so far. Several cutaneous abnormalities were detected during our investigation in a mice model of acute and long-COVID. In the acute phase of the disease, we observed destruction of the epidermal layer, an increase in the number of hair follicles, extensive deposition of collagen in the dermal layer, and hyperplasticity of the sebaceous glands. Further, the investigation disclosed the thinning of the panniculus carnosus and the adventitial layer. In contrast, the long-term post-COVID cutaneous investigation showed the absence of hair follicles from the epidermal and dermal layers, destruction of adipose tissues, and obliteration of the epidermal layer. Further, the treatment of infected mice with SPK reversed these cutaneous abnormalities. Precisely, the drug was able to restore the number of hair follicles and re-architecture the epidermal and dermal layers. Moreover, adipose tissue was restored in treated mice, which strongly suggests the possibility of long-term cutaneous defects in humans with COVID-19, and SPK can be used as a s potential therapeutic agent.
Since the onset of the COVID-19 pandemic in 2020, a comprehensive examination of its clinical manifestations has unveiled diverse and often enigmatic multiorgan pathological changes and complications. Our investigations have particularly focused on elucidating the potential alterations in the skin, revealing a spectrum of pathological changes in a well-established mouse model of COVID-19. Over time, it is anticipated that various dermatological disorders may emerge as sequelae of COVID-19. These potential disorders encompass but are not limited to alopecia, vitiligo, skin malignancy, accelerated skin wrinkling, pemphigus, and atopic dermatitis, among others. The etiology of these dermatological manifestations is likely multifactorial, involving intricate interactions between the virus and the host’s genes. While the virus itself may act as a potential trigger, its role in directly contributing to genetic mutations that lead to these disorders remains an area of ongoing investigation. It is essential to recognize the complexity of the SARS-CoV-2 virus and its potential mechanistic involvement in the development of dermatological disorders.
The International League of Dermatological Societies (ILDS), in collaboration with the American Academy of Dermatology (AAD), has meticulously documented a compendium of 2500 instances emanating from diverse regions, totaling up to 72 countries [32]. Predominant dermatological manifestations correlated with COVID-19 were concluded. These include: (1) exanthematous rash (20%), (2) pernio-like acral lesions (19%), (3) urticaria (15%), (4) varicella-like eruption (12%), (5) papulosquamous rash (10%), and (6) retiform purpura (7%) [33,34]. Additionally, sporadic occurrences of multisystem inflammatory syndrome in children (MIS-C) have been ascribed to COVID-19 infections, with documented instances in the scientific literature [33,34]. It is noteworthy that MIS-C exhibits similarities to Kawasaki Disease, albeit differentiated by distinctive clinical characteristics, encompassing the mean age of onset, cardiovascular sequelae, and pertinent laboratory parameters [33,34,35].
During the acute phase of MHV-1 infection, a conspicuous augmentation in the numerical density of hair follicles was distinctly observed in comparison to the control group. This phenomenon can be explicated by reference to the established pathophysiological mechanisms operative in non-COVID-19 cases. Principal factors contributing to this phenomenon encompass heightened androgen receptor expression and elevated levels of insulin-like growth factor-1 (IGF-1) [36,37,38]. Androgenic influence assumes a pivotal role in this context, acting as an inhibitory hormone within the scalp, while exerting a stimulatory effect in other anatomical regions [37]. Remarkable is the indispensable involvement of androgen-metabolizing enzymes, namely aromatase, 17 beta-hydroxysteroid dehydrogenase (17 beta-HSD), and 5-alpha reductase (types I and II), in the intricate orchestration of hair follicle development [37,38]. Concurrently, an array of hormones actively participates in fostering the developmental trajectory of hair follicles. These include estradiol, progesterone, prolactin, thyroid hormones (T3 and T4), melatonin, corticotropin-releasing hormone (CRH), adrenocorticotropic hormone (ACTH), cortisol, and thyroid-releasing hormone [37,38,39,40,41,42].
However, there exist inhibitory hormones that exert a regulatory influence on hair follicle development, such as galanin (predominantly localized in the peripheral and central nervous systems, with documented effects including the reduction of matric keratinocyte proliferation, shortening of the anagen phase, and attenuation of hair shaft extension) and transforming growth factor-beta (TGF-beta) [43].
In the prolonged course of COVID-19, a marked and complete abatement of hair follicle numbers was identified, which was characterized by severe obliteration and diminished follicular presence. This deleterious phenomenon is thought to be attributable to multiple factors, including the direct impact of the virus, the influence of transforming growth factor-beta (TGF-beta) [44], and alterations in the hormonal milieu [45]. Notably, our experimental intervention involving the utilization of our proprietary therapeutic agent (SPK) yielded a remarkable resurgence in hair follicle numbers. This promising outcome suggests the potential efficacy of SPK in ameliorating or reversing alopecia induced by COVID-19. Consequently, this innovative medication holds promise as a prospective therapeutic modality for the treatment of COVID-19-associated alopecia in the future.
Diverse viruses exhibit an adept capacity to infiltrate the integumentary system, specifically establishing residence within the basal layer of the epidermis. Among the notable viral entities engaging in this modus operandi are herpes simplex, vaccinia virus, molluscum contagiosum, varicella-zoster, human papillomavirus, polyomavirus, human immunodeficiency virus (HIV), Zika virus, West Nile virus, Dengue virus, Chikungunya virus, and an array of other pathogens [46,47,48,49]. These viruses strategically exploit the epidermal stratum as a conducive environment for their replication processes [48], with keratinocytes emerging as the principal cellular cohort for such proliferative endeavors [48]. The epidermis, a proliferative substrate, becomes the locus for the intricate orchestration of viral replication, thereby facilitating the propagation and dissemination of these viral agents [49,50].
Shortly following entry, the toll-like receptor-2 (TLR) promptly engages, initiating a cascade that amplifies the activation of the nuclear factor-kappa B (NF-κB) pathway [51]. This orchestrated activation event precipitates the induction of an array of pro-inflammatory cytokines, notably interleukins (IL-1, IL-6, and IL-12), tumor necrotic factor-alpha (TNF-alpha), and chemokines, such as chemokine ligand-2 [49,52]. Concomitantly, the production of additional inflammatory constituents, including interferons and histamine, ensues, thereby fostering the recruitment of inflammatory cells to the affected site [52]. This heightened recruitment of inflammatory cells while constituting an immune response, paradoxically begets consequential deleterious effects. The ensuing influx of inflammatory cells inflicts severe and undesirable injury upon the cellular constituents of the epidermal layer [53]. This injurious process is observable in both acute and long-term manifestations of COVID-19, wherein a conspicuous and consequential detriment to the structural integrity of the epidermal layer is evident. The observed severe damage underscores the complex interplay between viral-induced immune responses and the resultant pathological consequences within the epidermal microenvironment.
Substantial evidence has been amassed, attesting to the discernible presence of SARS-CoV-2 within the skin, substantiated by the observable loss of adipose tissue during both acute and protracted phases of infection. Notably, the adipose tissue undergoes a conspicuous and seemingly complete disappearance, as delineated above. This peculiar manifestation is ascribed to the virus’s proclivity for extrapulmonary adipose tissue targeting, as Thangavel et al. elucidated in a murine model [4]. The investigative work by Thangavel et al. not only underscored the virus’s predilection for adipose tissue but also shed light on the infiltration of inflammatory cells, particularly macrophages, within the afflicted adipose tissue [54].
In light of these revelatory findings, our research endeavors were extended to ascertain the potential therapeutic effects of SPK. Strikingly, as illustrated in Figure 4 above, the outcomes of our investigations revealed a noteworthy restoration of both the epidermal layer and the adipose tissue layer following SPK administration. This unforeseen recuperation implies a promising role for SPK in mitigating the deleterious effects on both cutaneous components inflicted by SARS-CoV-2 infection, suggesting a potential avenue for therapeutic intervention in the context of skin manifestations associated with COVID-19.
The notable deposition of collagen emerges because of the inflammatory processes induced by the viral infection [55]. Within the intricate milieu of the extracellular matrix (ECM), a myriad of constituent elements contributes to its structural complexity. These include various types of collagens (I-XXIII), elastic fibers (comprising elastin, fibrillin-1, and fibrillin-2), glycoproteins (such as laminin, fibronectin, nidogen, entactin, periostin, tenascin-C, and thrombospondin-1), proteoglycans (encompassing biglycan, decorin, perlecan, versican, lumican, fibromodulin, and agrin), galactosaminoglycan and glycosaminoglycan (including heparan sulfate, dermatan sulfate, and hyaluronan) [56,57,58,59,60,61,62,63,64,65]. This comprehensive array of ECM components underscores the intricate and dynamic nature of the tissue microenvironment, where the interplay of diverse molecular constituents contributes to the structural integrity and functionality of tissues. The observed collagen deposition, linked to the viral-induced inflammatory response, underscores the complexity of the molecular landscape orchestrating tissue remodeling and repair in the context of viral infections.
The extracellular matrix (ECM) stands as a perpetually modulated, evolutionarily conserved, highly adaptable, and dynamically transformative scaffold, orchestrating controlled processes of metamorphosis and restructuring [63]. Within the integumentary system, the skin’s layers, comprising the epidermis and dermis, are demarcated by the basement membrane. This intricate amalgamation of ECM proteins and fibers assumes the pivotal role of bridging the epithelium to the subjacent tissue, thereby establishing a structural and functional continuum [65,66]. Within the interstitial expanse of the dermis, a diverse array of cells, including fibroblasts and immune/vascular cells, reside proximate to the ECM, providing a supportive milieu and ensconcing these cells in a manner conducive to intimate interactions [67]. This dynamic cellular-ECM interplay contributes to the maintenance of tissue homeostasis and function. However, any aberrations in the ECM, particularly those induced by pathological agents such as bacterial or viral infections, can exert discernible effects on the dermal layer. Such alterations manifest through the recruitment of various immune cells, thereby instigating a cascade of events that influence the structural and physiological aspects of the dermal microenvironment.
In our experimental model, a conspicuous augmentation in the abundance of the extracellular matrix (ECM) was noted, with discernible evidence of heightened collagen deposition observable in Figure 1 and Figure 2. This substantial increase in ECM can be ascribed to incremental alterations occurring in both fibroblasts and keratinocytes, which emerge as principal catalysts driving the remodeling processes within the ECM. Notably, our earlier investigations have revealed a significant elevation of transforming growth factor-beta (TGF-β) within the renal tissue, as referenced [49]. TGF-β, conventionally latent in the extracellular matrix, undergoes activation in response to various stimuli, including tissue injury or inflammatory cues [49,68]. Upon activation, TGF-β engages with its cell surface receptors, predominantly serine/threonine kinase receptors recognized as TGF-β receptors [68]. This activation of TGF-β and subsequent interaction with its receptors constitute pivotal molecular events contributing to the orchestrated modulation of the ECM [69]. The intricate interplay between these cellular and molecular constituents underscores the multifaceted nature of ECM dynamics and the regulatory mechanisms governing tissue remodeling in response to various stimuli.
TGF-β initiates an intracellular signaling cascade, involving proteins like Smad proteins that transduce the TGF-β signal from the cell membrane to the nucleus [70,71,72]. In the nucleus, Smad proteins, along with other transcription factors, regulate the expression of genes involved in collagen synthesis, including genes that encode various types of collagens, the principal component of the extracellular matrix [70,71,72]. As a result of these signaling pathways, fibroblasts, the primary cells responsible for collagen production, are stimulated to synthesize and release increased amounts of collagen [70,71,72]. The excess collagen is then deposited in the extracellular matrix, leading to remodeling and alterations in the structure of the skin.
Consequently, it is plausible that the virus can elicit an augmented secretion of transforming growth factor-beta (TGF-β), either directly or indirectly, by activating periostin in fibroblasts and basal keratinocytes during both the acute and protracted phases of COVID-19 [49,73]. Remarkably, our investigations have delved into the impact of SPK on skin tissue remodeling, revealing discernible outcomes such as a reduction in collagen accumulation and fibrosis within the dermal layer. Noteworthy is the consistent observation that TGF-β levels were attenuated in our prior renal model upon employing SPK [49]. This confluence of findings suggests a potential modulatory effect of SPK on the TGF-β signaling pathway, mitigating its downstream effects on collagen deposition and fibrosis. The multifaceted influence of SPK on distinct tissue types underscores its therapeutic potential in attenuating pathological alterations associated with COVID-19, implicating SPK as a promising candidate for interventions aimed at mitigating tissue remodeling and fibrosis in diverse anatomical contexts.
5. Conclusions
In summary, our findings suggest that various skin changes occur during the acute phase of SARS-CoV-2 infection, potentially leading to continuous dermatological alterations in long-COVID. These observations are evident in all skin layers, with hair follicle destruction in the long-term phase. Furthermore, the use of SPK demonstrates the potential to restore normal skin architecture.
Author Contributions
HH: MJP, and ARJ conceived the study; HH, MJP, ARJ, RR, AF, MA, and PC performed the study, as well as wrote the paper.
Funding
This study was supported by the University of Miami Miller School of Medicine Department Obstetrics, Gynecology and Reproductive Sciences, Muriel, Murray & Robert Smith Foundation COVID-19 Research Fund, University of Miami Miller School of Medicine Team Science Award, and the Charles M. Vallee Foundation.
Institutional Review Board Statement
The animal study protocol was approved by the University of Miami Institutional Animal Care and Use Committee (IACUC protocol number 20–162 approved on 9/12/2023).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to the University of Miami Miller School of Medicine’s privacy policy.
Acknowledgments
We thank Anis Ahmad, Department of Radiation Oncology, University of Miami Miller School of Medicine for his assistance in picture scanning.
Conflicts of Interest
Paidas is a Scientific Advisory Board Member of BioIncept, LLC with stock options. The other authors have no conflict of interest or competing interest to declare.
References
- World Health Organization. Who Coronavirus (COVID-19) Dashboard. World Health Organization. Available online: https://covid19.who.int/ (accessed on 20 July 2023).
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms, and recommendations. Nat. Rev. Microbiol. 2023, 3, 133–146. [CrossRef]
- Sullivan, M.K.; Lees, J.S.; Drake, T.M.; Docherty, A.B.; Oates, G.; Hardwick, H.E. Acute Kidney Injury in Patients Hospitalized With COVID-19 From the ISARIC WHO CCP-UK Study: A Prospective, Multicentre Cohort Study Michael. Nephrol. Dial. Transplant. 2021, 27708, 1–19. [CrossRef]
- Mortaz, E.; Tabarsi, P.; Jamaati, H.; Dalil Roofchayee, N.; Dezfuli, N.K.; Hashemian, S.M.; Moniri, A.; Marjani, M.; Malekmohammad, M.; Mansouri, D.; et al. Increased Serum Levels of Soluble TNF-α Receptor Is Associated with ICU Mortality in COVID-19 Patients. Front. Immunol. 2021, 12, 592727. [CrossRef]
- Schiffl, H.; Lang, S.M. Long-term interplay between COVID-19 and chronic kidney disease. Int. Urol. Nephrol. 2023, 55, 1977–1984. [CrossRef]
- Kudose, S.; Batal, I.; Santoriello, D.; Xu, K.; Barasch, J.; Peleg, Y.; Canetta, P.; Ratner, L.E.; Marasa, M.; Gharavi, A.G.; Stokes MB, Markowitz GS, D’Agati VD. Kidney Biopsy Findings in Patients with COVID-19. J. Am. Soc. Nephrol. 2020, 31, 1959–1968. [CrossRef]
- Zamoner, W.; Santos, C.A.D.S.; Magalhães, L.E.; de Oliveira, P.G.S.; Balbi, A.L.; Ponce, D. Acute Kidney Injury in COVID-19: 90 Days of the Pandemic in a Brazilian Public Hospital. Front. Med. 2021, 8, 622577. [CrossRef]
- Davis H.E.; McCorkell L.; Vogel J.M.; Topol E.J. Long COVID: major findings, mechanisms and recommendations. Nat Rev Microbiol. 2023, 3, 133-146. [CrossRef]
- Sharma, P.; Uppal, N.N.; Wanchoo, R.; Shah, H.H.; Yang, Y.; Parikh, R.; Khanin, Y.; Madireddy, V.; Larsen, C.P.; Jhaveri, K.D.; Bijol V. COVID-19-associated kidney injury: A case series of kidney biopsy findings. J. Am. Soc. Nephrol. 2020, 31, 1948–1958. [CrossRef]
- Körner, R.W.; Majjouti, M.; Alcazar, M.A.A.; Mahabir, E. Of Mice and cMHV and Mouse Models as a Translational Approach to Understand SARS-CoV-2. Viruses 2020, 12, 880.
- De Albuquerque, N.; Baig, E.; Ma, X.; Zhang, J.; He, W.; Rowe, A.; Habal, M.; Liu, M.; Shalev, I.; Downey, G.P.; Gorczynski R.; Butany J.; Leibowitz J.; Weiss S.R.; McGilvray I.D.; Phillips M.J.; Fish E.N.; Levy G.A. Murine hepatitis virus strain 1 produces a clinically relevant model of severe acute respiratory syndrome in A/J mice. J. Virol. 2006, 80, 10382–10394. [CrossRef]
- Gong, H.H.; Worley, M.J.; Carver, K.A.; Goldstein, D.R.; Deng, J.C. Neutrophils drive pulmonary vascular leakage in MHV-1 infection of susceptible A/J mice. Front. Immunol. 2023, 13, 1089064. [CrossRef]
- Cox, G.; Gonzalez, A.J.; Ijezie, E.C.; Rodriguez, A.; Miller, C.R.; Van Leuven, J.T.; Miura, T.A. Priming With Rhinovirus Protects Mice Against a Lethal Pulmonary Coronavirus Infection. Front. Immunol. 2022, 13, 886611. [CrossRef]
- Gain, C.; Song, S.; Angtuaco, T.; Satta, S.; Kelesidis, T. The role of oxidative stress in the pathogenesis of infections with coronaviruses. Front. Microbiol. 2023, 13, 1111930. [CrossRef]
- Paidas, M.J.; Cosio, D.S.; Ali, S.; Kenyon, N.S.; Jayakumar, A.R. Long-Term Sequelae of COVID-19 in Experimental Mice. Mol. Neurobiol. 2022, 59, 5970–5986. [CrossRef]
- Paidas, M.J.; Sampath, N.; Schindler, E.A.; Cosio, D.S.; Ndubizu, C.O.; Shamaladevi, N.; Kwal, J.; Rodriguez, S.; Ahmad, A.; Kenyon, N.S.; Jayakumar A.R. Mechanism of Multi-Organ Injury in Experimental COVID-19 and Its Inhibition by a Small Molecule Peptide. Front. Pharmacol. 2022, 30, 864798. [CrossRef]
- Caldera-Crespo, L.A.; Paidas, M.J.; Roy, S.; Schulman, C.I.; Kenyon, N.S.; Daunert, S.; Jayakumar, A.R. Experimental Models of COVID-19. Front. Cell Infect. Microbiol. 2022, 11, 792584. [CrossRef]
- Paidas, M.J.; Mohamed, A.B.; Norenberg, M.D.; Saad, A.; Barry, A.F.; Colon, C.; Kenyon, N.S.; Jayakumar, A.R. Multi-Organ Histopathological Changes in a Mouse Hepatitis Virus Model of COVID-19. Viruses 2021, 13, 1703. [CrossRef]
- Gottlieb, M.; Long, B. Dermatologic manifestations and complications of COVID-19. The American journal of emergency medicine. 2020, 9, 1715-1721. [CrossRef]
- Gao, J. C.; Huang, A.; Desai, A.; Safai, B.; Marmon, S. “COVID toes”: A true viral phenomenon or a diagnosis without a leg to stand on?. JAAD international. 2022, 9, 1-6. [CrossRef]
- Muntean, I. A.; Pintea, I.; Bocsan, I. C.; Dobrican, C. T.; Deleanu, D. COVID-19 disease leading to chronic spontaneous urticaria exacerbation: a Romanian retrospective study. In Healthcare. 2021, 9, 1144. [CrossRef]
- Sameni, F.; Hajikhani, B.; Yaslianifard, S.; Goudarzi, M.; Owlia, P.; Nasiri, M. J.; Dadashi, M. COVID-19 and skin manifestations: an overview of case reports/case series and meta-analysis of prevalence studies. Frontiers in Medicine. 2020, 7, 573188. [CrossRef]
- Becker, R.C. COVID-19-associated vasculitis and vasculopathy. Journal of thrombosis and thrombolysis. 2020, 50, 499-511. [CrossRef]
- Iba, T.; Connors, J. M.; Levy, J. H. The coagulopathy, endotheliopathy, and vasculitis of COVID-19. Inflammation Research. 2020, 69, 1181-1189. [CrossRef]
- Sahara, T.; Yokota, K. Livedo reticularis associated with COVID-19. Internal Medicine. 2022, 3, 441-441. [CrossRef]
- Jaimes, J. A.; Millet, J. K.; Whittaker, G. R. Proteolytic cleavage of the SARS-CoV-2 spike protein and the role of the novel S1/S2 site. IScience.2020, 6, 22-50. [CrossRef]
- Rajan, M.B.; Kumar, M.P.; Bhardwaj, A. The trend of cutaneous lesions during COVID-19 pandemic: lessons from a meta-analysis and systematic review. International Journal of Dermatology. 2020, 11, 1358-1370. [CrossRef]
- Tan, S.W.; Tam, Y.C.; Oh, C.C. Skin manifestations of COVID-19: a worldwide review. JAAD international. 2020, 2, 119-133. [CrossRef]
- Ramamoorthy, R.; Hussain H.; Ravelo, N.; Sriramajayam, K.; Di Gregorio, D.M.; Paulrasu K.; Chen, P.; Young, K.; Masciarella, A.D.; Jayakumar, A.R.; Paidas, M.J. Kidney Damage in Long COVID: Studies in Experimental Mice. Biology (Basel). 2023 30,1070. [CrossRef]
- Bancroft, J. D.; Gamble, M. Theory and Practice of Histological Techniques (6th ed.). Churchill Livingstone, Elsevier, China, 2008.
- Kiernan, J.A. Histological and Histochemical Methods: Theory and Practice (4th ed.). Cold Spring Harbor Laboratory Press, 2008.
- Strahan, A.G.; Lubov, J.E.; Prasad, S.; Fox, L.P.; McMahon, D.E.; Singh, R.; Rosenbach, M.; Desai, S.R.; Lim, H.W.; Thiers, B.H.; Hruza, G.J.; French, L.E.; Freeman, E.E. The impact of the American Academy of Dermatology/International League of Dermatological Societies COVID-19 Registry during the pandemic: 2500 cases across 72 countries. J Am Acad Dermatol. 2023, 89, e225-e227. [CrossRef]
- Jiang, L.; Tang, K.; Irfan, O.; Li, X.; Zhang, E.; Bhutta, Z. Epidemiology, clinical features, and outcomes of multisystem inflammatory syndrome in children (MIS-C) and adolescents—a live systematic review and meta-analysis. Current pediatrics reports. 2022, 2, 19-30. [CrossRef]
- Rafferty, M.S.; Burrows, H.; Joseph, J.P.; Leveille, J.; Nihtianova, S.; Amirian, E.S. Multisystem inflammatory syndrome in children (MIS-C) and the coronavirus pandemic: Current knowledge and implications for public health. J Infect Public Health. 2021, 4, 484-494. [CrossRef]
- Roe, K. A. viral infection explanation for Kawasaki disease in general and for COVID-19 virus-related Kawasaki disease symptoms. Inflammopharmacology. 2020, 28, 1219-1222. [CrossRef]
- Horesh, E.J.; Chéret, J.; Paus, R. Growth Hormone and the Human Hair Follicle. Int J Mol Sci. 2021, 8,13205. [CrossRef]
- Trüeb, R.M. Further Clinical Evidence for the Effect of IGF-1 on Hair Growth and Alopecia. Skin Appendage Disord. 2018, 4, 90-95. [CrossRef]
- Sawaya, M.E. Steroid chemistry and hormone controls during the hair follicle cycle. Ann N Y Acad Sci. 1991, 26, 642:376-83; discussion 383-4. [CrossRef]
- Hamada, K.; Thornton, M.J.; Laing, I.; Messenger, A.G.; Randall, V.A. The metabolism of testosterone by dermal papilla cells cultured from human pubic and axillary hair follicles concurs with hair growth in 5 alpha-reductase deficiency. J Invest Dermatol. 1996, 106,1017-22. [CrossRef]
- Babadjouni, A.; Reddy, M.; Zhang, R.; Raffi, J.; Phong, C.; Mesinkovska, N. Melatonin and the Human Hair Follicle. J Drugs Dermatol. 2023, 22, 260-264. [CrossRef]
- Heidelbaugh, J.J. Endocrinology Update: Hirsutism. FP Essent. 2016, 451, 17-24.
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal Effects on Hair Follicles. Int J Mol Sci. 2020, 28, 5342. [CrossRef]
- Holub, B.S.; Kloepper, J.E.; Tóth, B.I.; Bíro, T.; Kofler, B.; Paus, R. The neuropeptide galanin is a novel inhibitor of human hair growth. Br J Dermatol. 2012, 167, 10-6. [CrossRef]
- Zhu, H.L.; Gao, Y.H.; Yang, J.Q.; Li, J.B.; Gao, J. Serenoa repens extracts promote hair regeneration and repair of hair loss mouse models by activating TGF-β and mitochondrial signaling pathway. Eur Rev Med Pharmacol Sci. 2018, 22, 4000-4008.
- Randall, V.A.; Thornton, M.J.; Messenger, A.G.; Hibberts, N.A.; Loudon, A.S.; Brinklow, B.R. Hormones and hair growth: variations in androgen receptor content of dermal papilla cells cultured from human and red deer (Cervus elaphus) hair follicles. J Invest Dermatol. 1993, 101, 114S-120S. [CrossRef]
- Lei, V.; Petty, A.J.; Atwater, A.R.; Wolfe, S.A.; MacLeod, A.S. Skin Viral Infections: Host Antiviral Innate Immunity and Viral Immune Evasion. Front Immunol. 2020, 11, 593901. [CrossRef]
- De La Cruz, N.C.; Möckel, M.; Wirtz, L.; Knebel-Mörsdorf, D. Ex vivo Human Skin Infection with Herpes Simplex Virus 1. Bio Protoc. 2022, 12, e4411. [CrossRef]
- Knöpfel, N.; Noguera-Morel, L.; Latour, I.; Torrelo, A. Viral exanthems in children: A great imitator. Clin Dermatol. 2019, 37, 213-226. [CrossRef]
- Hussain, H.; Paidas, M.J.; Fadel, A.; Ramamoorthy, R.; Garcia, E.; Saadoon, Z.F.; Casmartino, E.; Mendez, L.; Williams, E.A.; Ruiz-Cordero, R.; Jayakumar, A.R. Prior viral infection determines the mode and severity of monkeypox virus. Int J Infect Dis. 2023, 131, 95-99. [CrossRef]
- Cunningham, A.L.; Carbone, F.; Geijtenbeek, T.B. Langerhans cells and viral immunity. Eur J Immunol. 2008, 38, 2377-85. [CrossRef]
- Hasannejad, H.; Takahashi, R.; Kimishima, M.; Hayakawa, K.; Shiohara, T. Selective impairment of Toll-like receptor 2-mediated proinflammatory cytokine production by monocytes from patients with atopic dermatitis. J Allergy Clin Immunol. 2007, 120, 69-75. [CrossRef]
- Kawamura, T.; Ogawa, Y.; Aoki, R.; Shimada, S. Innate and intrinsic antiviral immunity in skin. J Dermatol Sci. 2014, 75, 159-66. [CrossRef]
- Magro, C.; Crowson, A.N.; Franks, L.; Schaffer, P.R.; Whelan, P.; Nuovo, G. The histologic and molecular correlates of COVID-19 vaccine-induced changes in the skin. Clin Dermatol. 2021, 39, 966-984.
- Thangavel, H.; Dhanyalayam, D.; Lizardo, K.; Oswal, N.; Dolgov, E.; Perlin, D.S.; Nagajyothi, J.F. Susceptibility of Fat Tissue to SARS-CoV-2 Infection in Female hACE2 Mouse Model. Int J Mol Sci. 2023, 24, 1314. [CrossRef]
- Hofman, P.; Copin, M.C.; Tauziede-Espariat, A.; Adle-Biassette, H.; Fortarezza, F.; Passeron, T.; Salmon, I.; Calabrese, F. Les lésions histologiques associées à l’infection par le SARS-CoV-2 [Histopathological features due to the SARS-CoV-2]. Ann Pathol. 2021, 41, 9-22. [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am J Pathol. 2006, 168, 1861-8. [CrossRef]
- Schmidtchen, A.; Carlstedt, I.; Malmström, A.; Fransson, L.A. Inventory of human skin fibroblast proteoglycans. Identification of multiple heparan and chondroitin/dermatan sulphate proteoglycans. Biochem J. 1990, 265, 289-300. [CrossRef]
- Lee, D.H.; Oh, J.H.; Chung, J.H. Glycosaminoglycan and proteoglycan in skin aging. J Dermatol Sci. 2016, 83, 174-81. [CrossRef]
- Metze, D.; Bhardwaj, R.; Amann, U.; Eades-Perner, A.M.; Neumaier, M.; Wagener, C.; Jantscheff, P.; Grunert, F.; Luger, T.A. Glycoproteins of the carcinoembryonic antigen (CEA) family are expressed in sweat and sebaceous glands of human fetal and adult skin. J Invest Dermatol. 1996, 106, 64-9. [CrossRef]
- Makihara, H.; Hidaka, M.; Sakai, Y.; Horie, Y.; Mitsui, H.; Ohashi, K.; Goshima, Y.; Akase, T. Reduction and fragmentation of elastic fibers in the skin of obese mice is associated with altered mRNA expression levels of fibrillin-1 and neprilysin. Connect Tissue Res. 2017, 58, 479-486. [CrossRef]
- Li, H.; Zhao, Y.; Zhou, L.; Hu, J. Cutaneous, skin histopathological manifestations and relationship to COVID-19 infection patients. Dermatol Ther. 2020, 33, e14157. [CrossRef]
- Huang, J.; Heng, S.; Zhang, W.; Liu, Y.; Xia, T.; Ji, C.; Zhang, L.J. Dermal extracellular matrix molecules in skin development, homeostasis, wound regeneration and diseases. Semin Cell Dev Biol. 2022, 128, 137-144. [CrossRef]
- Talbott, H.E.; Mascharak, S.; Griffin, M.; Wan, D.C.; Longaker, M.T. Wound healing, fibroblast heterogeneity, and fibrosis. Cell Stem Cell. 2022, 29,1161-1180. [CrossRef]
- Pfisterer, K.; Shaw, L.E.; Symmank, D.; Weninger, W. The Extracellular Matrix in Skin Inflammation and Infection. Front Cell Dev Biol. 2021, 6, 682414. [CrossRef]
- Daraban Bocaneti, F.; Altamura, G.; Corteggio, A.; Tanase, O.I.; Dascalu, M.A.; Pasca, S.A.; Hritcu, O.; Mares, M.; Borzacchiello, G. Expression of matrix metalloproteinases (MMPs)-2/-7/-9/-14 and tissue inhibitors of MMPs (TIMPs)-1/-2 in bovine cutaneous fibropapillomas associated with BPV-2 infection. Front Vet Sci. 2022, 28, 1063580. [CrossRef]
- Alpantaki, K.; Zafiropoulos, A.; Tseliou, M.; Vasarmidi, E.; Sourvinos, G. Herpes simplex virus type-1 infection affects the expression of extracellular matrix components in human nucleus pulposus cells. Virus Res. 2019, 259, 0-17. [CrossRef]
- Moretti, L.; Stalfort, J.; Barker, T.H.; Abebayehu, D. The interplay of fibroblasts, the extracellular matrix, and inflammation in scar formation. J Biol Chem. 2022, 298, 101530. [CrossRef]
- Chen, J.; Chen, J.K.; Nagai, K.; Plieth, D.; Tan, M.; Lee, T.C.; Threadgill, D.W.; Neilson, E.G.; Harris, R.C. EGFR signaling promotes TGFβ-dependent renal fibrosis. J. Am. Soc. Nephrol. 2012, 23, 215–224. [CrossRef]
- Sisto, M.; Ribatti, D.; Lisi, S. Organ Fibrosis and Autoimmunity; The Role of Inflammation in TGFβ-Dependent EMT. Biomolecules 2021, 11, 310. [CrossRef]
- Bao, M.; Feng, Q.; Zou, L.; Huang, J.; Zhu, C.; Xia, W. Endoplasmic reticulum stress promotes endometrial fibrosis through the TGF-β/SMAD pathway. Reproduction 2023, 165, 171–182. [CrossRef]
- Guo, Y.; Tian, G.; Chen, X.; Hou, Y.; Zhang, X.; Xue, X.; Zhao, L.; Wu, Y. GL-V9 ameliorates liver fibrosis by inhibiting TGF-β/smad pathway. Exp. Cell Res. 2023, 425, 113521. [CrossRef]
- Zhang, T.; He, X.; Caldwell, L.; Goru, S.K.; Ulloa Severino, L.; Tolosa, M.F.; Misra, P.S.; McEvoy, C.M.; Christova, T.; Liu, Y.; Atin C.; Zhang J.; Hu C.; Vukosa N.; Chen X.; Krizova A.; Kirpalani A.; Gregorieff A.; Ni R.; Chan, K.; Gill, M.K.; Attisano, L.; Wrana, J.L.; Yuen, D.A. NUAK1 promotes organ fibrosis via YAP and TGF-β/SMAD signaling. Sci. Transl. Med. 2022, 14, eaaz4028. [CrossRef]
- Laloglu, E.; Alay, H. Role of transforming growth factor-beta 1 and connective tissue growth factor levels in coronavirus disease-2019-related lung Injury: a prospective, observational, cohort study. Rev Soc Bras Med Trop. 2022, 55, e06152021. [CrossRef]
Figure 1.
Acute cutaneous changes in mice infected with MHV-1 (7 days post infection). There is thinning of the epidermis (red arrow in B) with the absence of stratum corneum (dark blue arrow), papillomatosis (black arrow), and dying keratinocyte (light blue arrow) in MHV-1 infected mice. In contrast, the epidermal layer is preserved in healthy mice (A, asterisk). The adipose tissue is replaced by fibrosis and collagen deposition in MHV-1 infected mice (black asterisk in D), as compared to the control (C, asterisk). Additionally, extensive collagen deposition is observed in MHV-1 infected mice (D, blue asterisk), and the number of hair follicles is increased, as compared to the controls (A & C). A and B = 66x; C and D = 23x in Figs 1 and 2 below, (n=16).
Figure 1.
Acute cutaneous changes in mice infected with MHV-1 (7 days post infection). There is thinning of the epidermis (red arrow in B) with the absence of stratum corneum (dark blue arrow), papillomatosis (black arrow), and dying keratinocyte (light blue arrow) in MHV-1 infected mice. In contrast, the epidermal layer is preserved in healthy mice (A, asterisk). The adipose tissue is replaced by fibrosis and collagen deposition in MHV-1 infected mice (black asterisk in D), as compared to the control (C, asterisk). Additionally, extensive collagen deposition is observed in MHV-1 infected mice (D, blue asterisk), and the number of hair follicles is increased, as compared to the controls (A & C). A and B = 66x; C and D = 23x in Figs 1 and 2 below, (n=16).
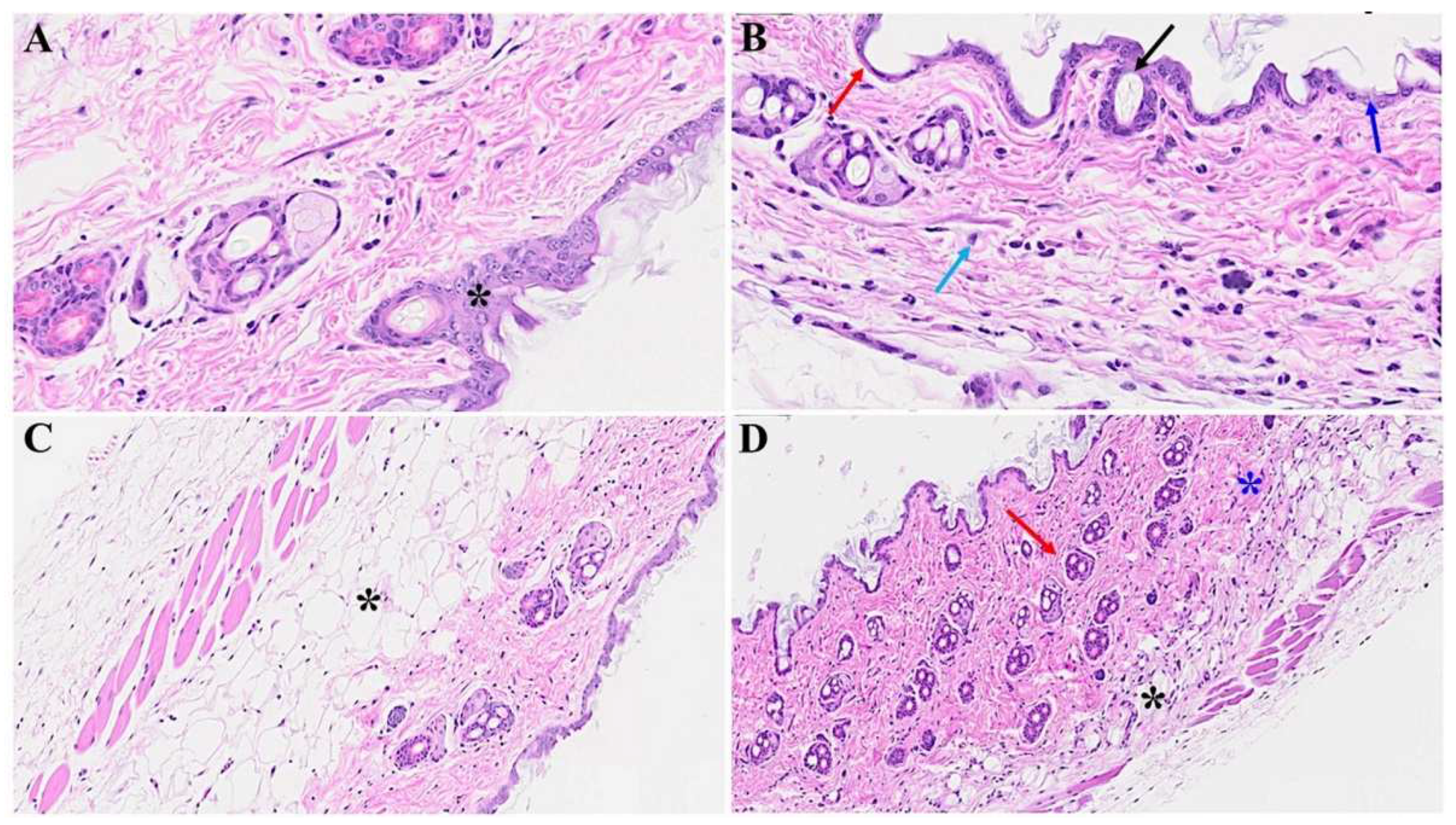
Figure 2.
Acute skin alteration in MHV-1 infection (7 days post-infection). There is an area of absence of epidermis noted in MHV-1 infected mice (red arrow in B), as compared to controls (asterisk in A). Additionally, there are various forms of dermal edema (blue arrows in B), and hyperplasticity of the sebaceous gland (black arrow in B). Further, there are various bullae and thinning of panniculus carnosus in MHV-1 infected mice (red arrow), (n=16).
Figure 2.
Acute skin alteration in MHV-1 infection (7 days post-infection). There is an area of absence of epidermis noted in MHV-1 infected mice (red arrow in B), as compared to controls (asterisk in A). Additionally, there are various forms of dermal edema (blue arrows in B), and hyperplasticity of the sebaceous gland (black arrow in B). Further, there are various bullae and thinning of panniculus carnosus in MHV-1 infected mice (red arrow), (n=16).
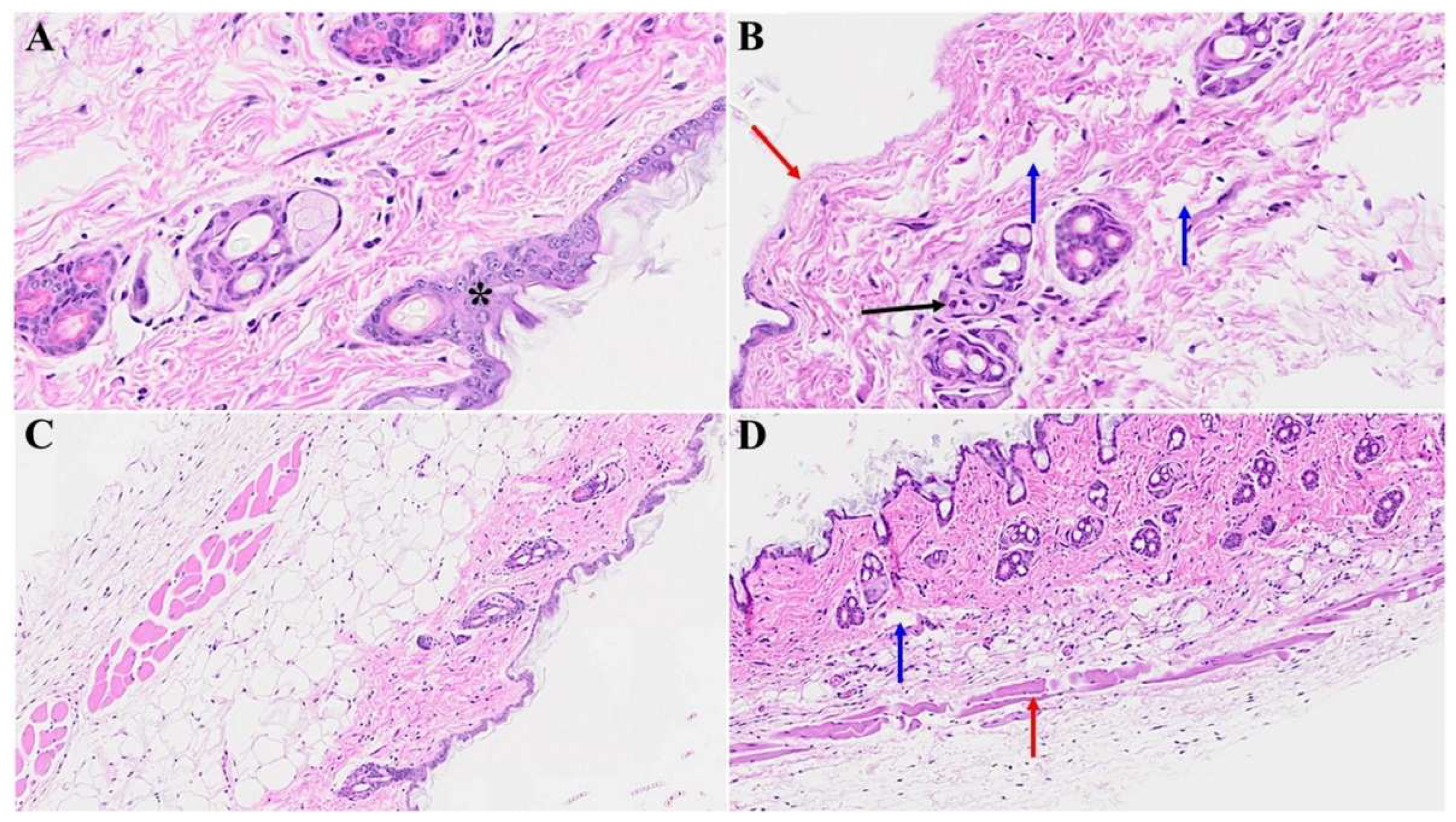
Figure 3.
Skin changes long-term post-MHV-1 infection in mice (1-year post-infection). There is a destruction of the adipose tissue in MHV-1-infected mice (black arrow), and an absence of the hair follicles in a dermal layer in mice 1-year post-infection (B), as compared to healthy control mice (A). Treatment of MHV-1 infected mice with SPK (5 mg/kg, as previously described by us, Ref 15) reversed the hair follicle loss (D). The destruction of the epidermal layer, dermal, and adipose tissue was also restored by SPK (black, blue, and red arrows in D), respectively. A and B = 23x; C and D = 66x in Figs 3 and 4 below, (n=6).
Figure 3.
Skin changes long-term post-MHV-1 infection in mice (1-year post-infection). There is a destruction of the adipose tissue in MHV-1-infected mice (black arrow), and an absence of the hair follicles in a dermal layer in mice 1-year post-infection (B), as compared to healthy control mice (A). Treatment of MHV-1 infected mice with SPK (5 mg/kg, as previously described by us, Ref 15) reversed the hair follicle loss (D). The destruction of the epidermal layer, dermal, and adipose tissue was also restored by SPK (black, blue, and red arrows in D), respectively. A and B = 23x; C and D = 66x in Figs 3 and 4 below, (n=6).
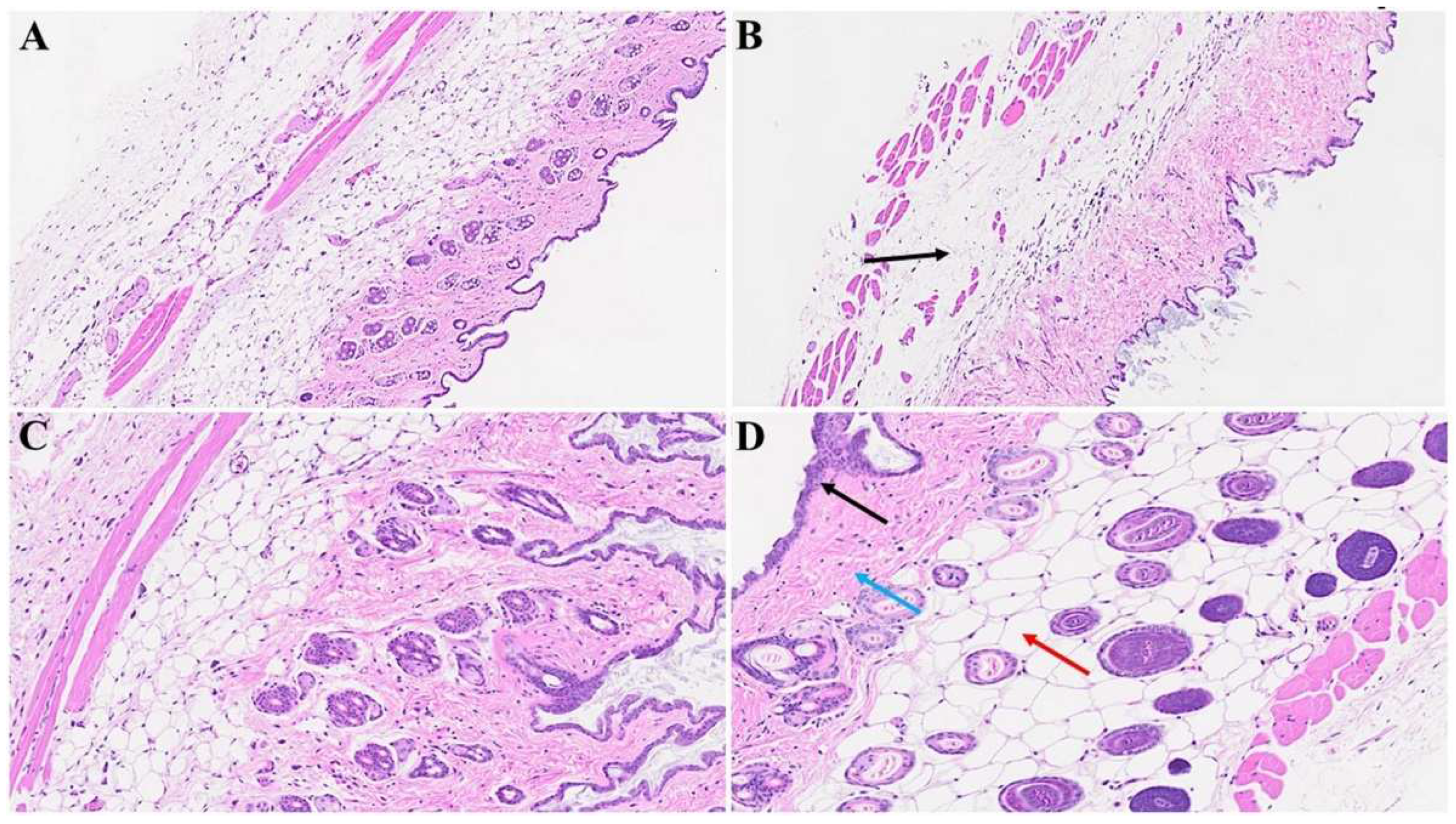
Figure 4.
Cutaneous abnormalities in Long-COVID. We found extensive destruction of the epidermis, dermal, and adipose tissue in MHV-1-infected mice (1-year post-infection, B). We also identified severe dermal edema and demolition of the skin architecture (B), as well as dying hair follicles (black arrow in C), and distortion of the panniculus carnosus architecture (red arrow, in C) in these mice, as compared to the healthy controls (A). These cutaneous abnormalities were restored when MHV-1-infected mice were treated with 5 mg/kg SPK (D), (n=6).
Figure 4.
Cutaneous abnormalities in Long-COVID. We found extensive destruction of the epidermis, dermal, and adipose tissue in MHV-1-infected mice (1-year post-infection, B). We also identified severe dermal edema and demolition of the skin architecture (B), as well as dying hair follicles (black arrow in C), and distortion of the panniculus carnosus architecture (red arrow, in C) in these mice, as compared to the healthy controls (A). These cutaneous abnormalities were restored when MHV-1-infected mice were treated with 5 mg/kg SPK (D), (n=6).
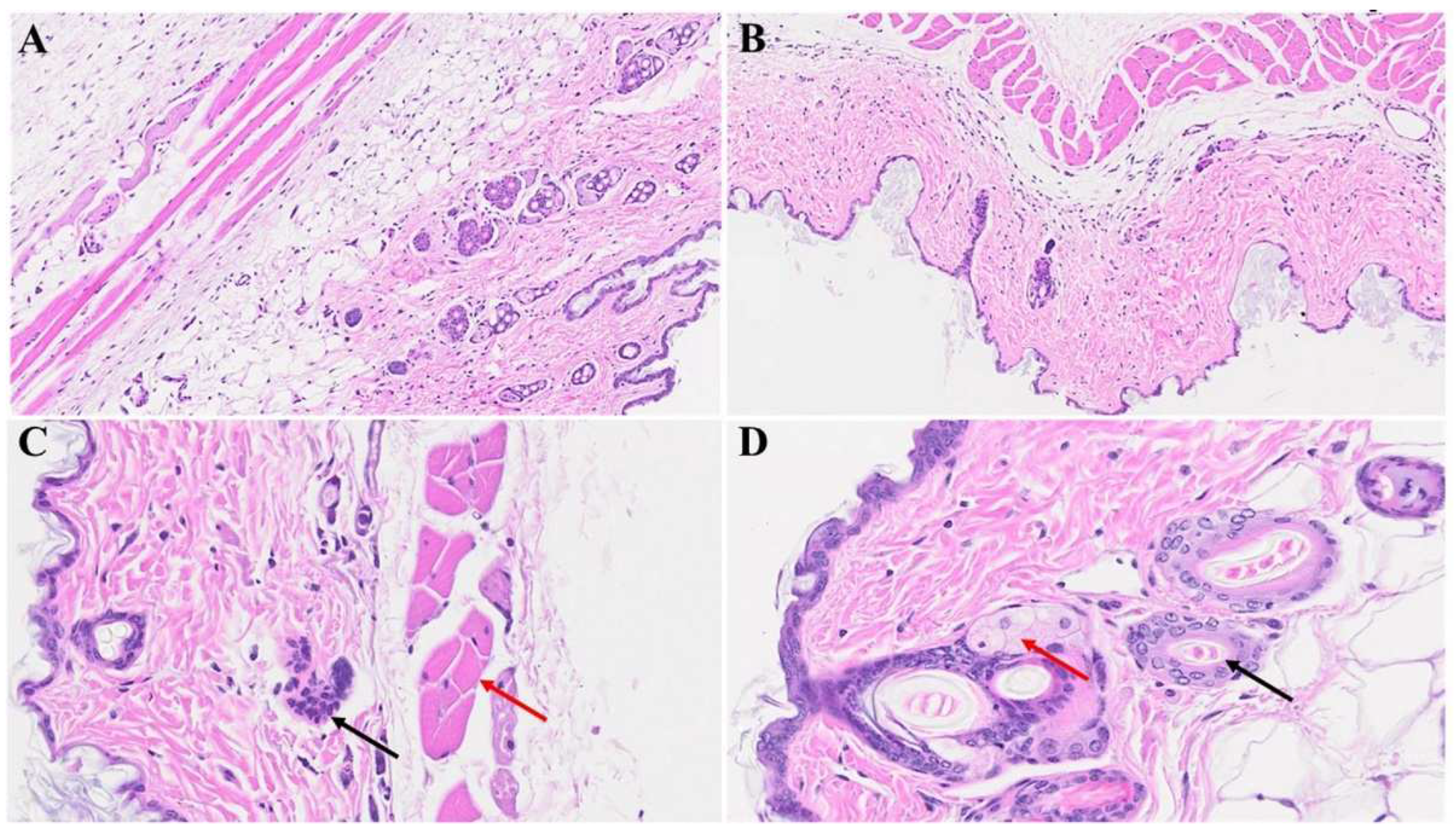
Figure 5.
Skin changes long-term post-MHV-1 infection in mice. Dying hair follicles with surrounding inflammatory cells (red arrows in B & C), as well as the destruction of skin architecture were observed 1-year post-MHV-1 infection, as compared to controls (A). These skin aberrations were, however, not observed in MHV-1-infected mice that were treated with SPK (5 mg/kg). A-D = 23x, (n=6).
Figure 5.
Skin changes long-term post-MHV-1 infection in mice. Dying hair follicles with surrounding inflammatory cells (red arrows in B & C), as well as the destruction of skin architecture were observed 1-year post-MHV-1 infection, as compared to controls (A). These skin aberrations were, however, not observed in MHV-1-infected mice that were treated with SPK (5 mg/kg). A-D = 23x, (n=6).
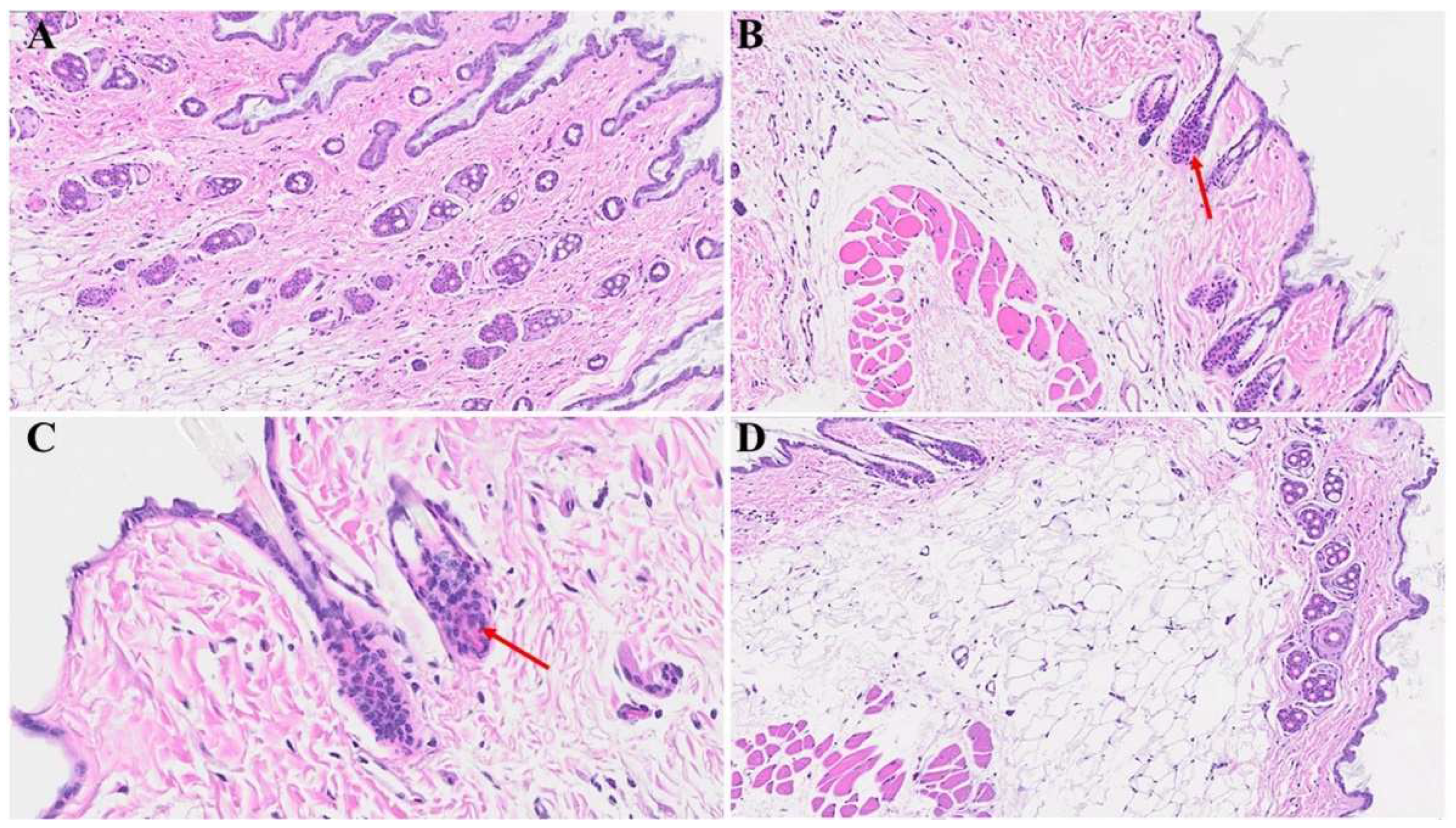
Figure 6.
We noticed macrothrombusi’s (black arrow in A), and various dermal edema (blue arrow in A). While there was an apoptotic keratinocyte (green arrow) and microthrombi (brown arrows) 1-year post-MHV-1 infection (B). We also detected Merkel cells with mild intracellular edema (black arrow in C), as well as intra-sebaceous gland vacuolization (brown arrow in D), and dying melanocytes (blue arrow in D) in MHV-1-infected mice 1-year post-infection. A-D = 23x, (n=6).
Figure 6.
We noticed macrothrombusi’s (black arrow in A), and various dermal edema (blue arrow in A). While there was an apoptotic keratinocyte (green arrow) and microthrombi (brown arrows) 1-year post-MHV-1 infection (B). We also detected Merkel cells with mild intracellular edema (black arrow in C), as well as intra-sebaceous gland vacuolization (brown arrow in D), and dying melanocytes (blue arrow in D) in MHV-1-infected mice 1-year post-infection. A-D = 23x, (n=6).
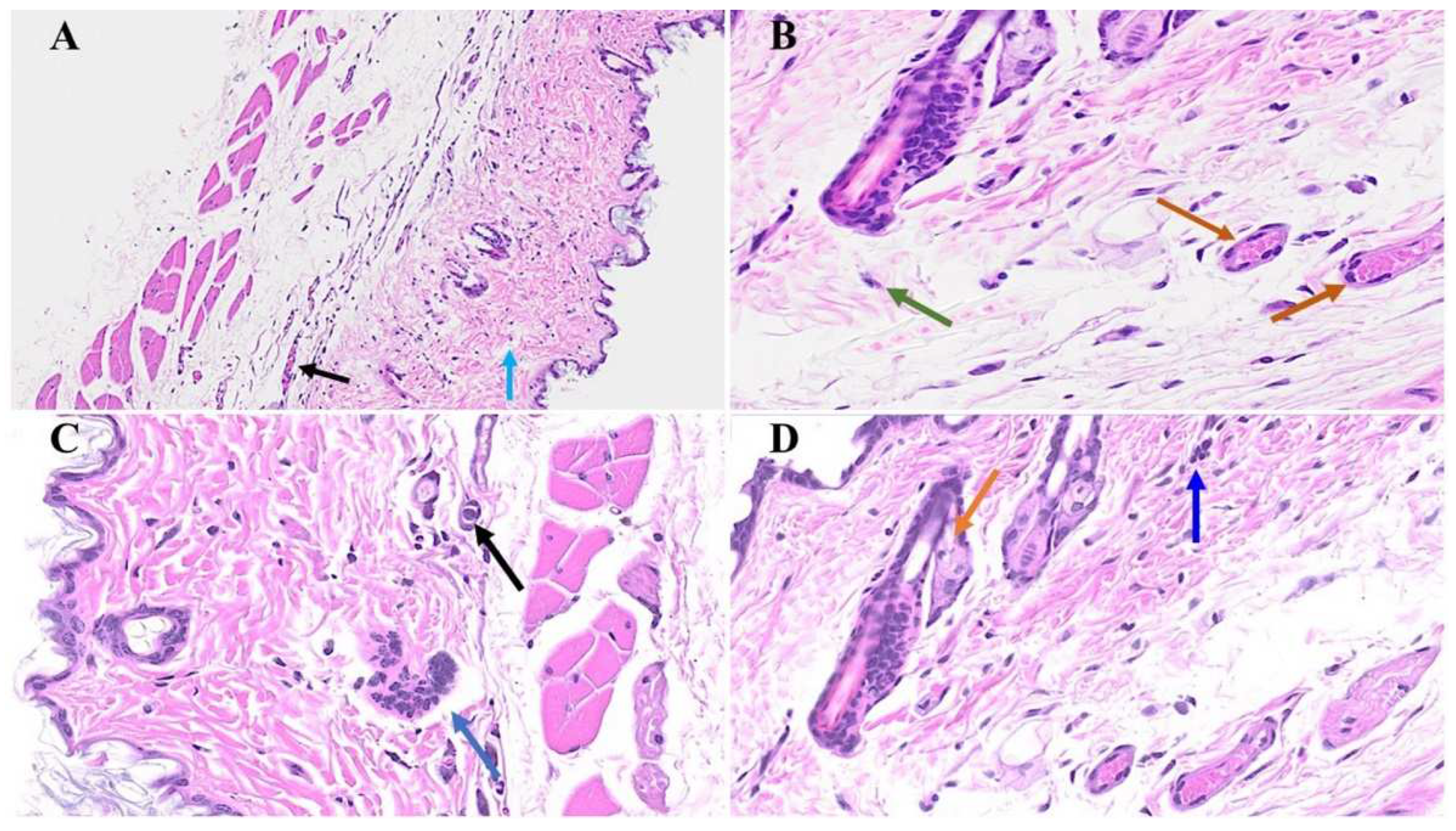
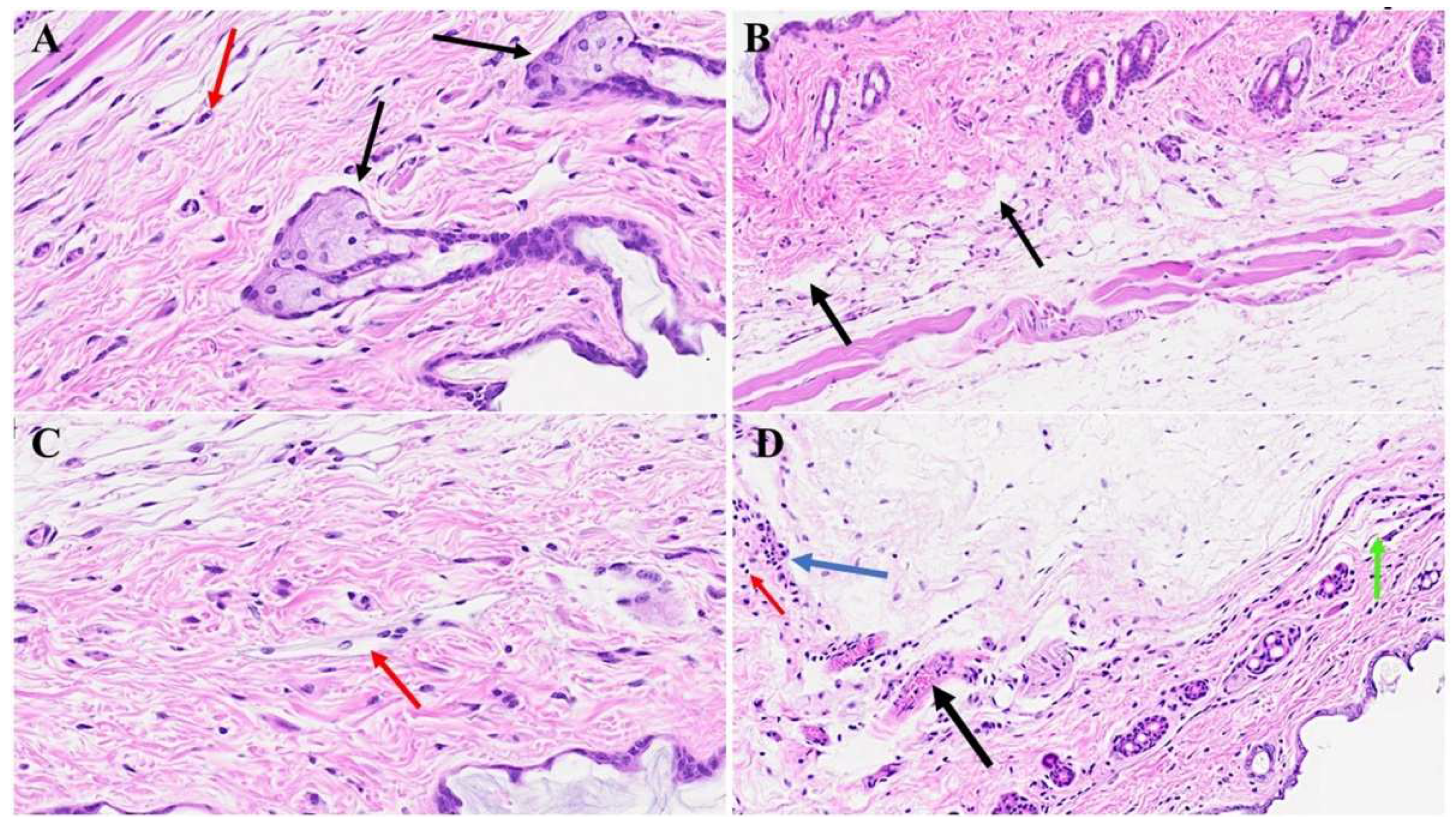
Figure 7.
Skin changes long-term post-MHV-1 infection in mice. MHV-1-infected mice (1-year post-infection) show the presence of eosinophil (red arrow in A) and hyperplasia of sebaceous glands (black arrows in A). There is also dermal bulla filled with inflammatory cells (red arrow), as well as scattered inflammatory cells, along with dermal edema in these mice (C). We also found a nest of inflammation (blue arrow), edematous melanocyte (red arrow), thrombosis (black arrow), and intradermal edema (green arrow) (D) in MHV-1 infected mice A-D = 23x, (n=6).
Figure 7.
Skin changes long-term post-MHV-1 infection in mice. MHV-1-infected mice (1-year post-infection) show the presence of eosinophil (red arrow in A) and hyperplasia of sebaceous glands (black arrows in A). There is also dermal bulla filled with inflammatory cells (red arrow), as well as scattered inflammatory cells, along with dermal edema in these mice (C). We also found a nest of inflammation (blue arrow), edematous melanocyte (red arrow), thrombosis (black arrow), and intradermal edema (green arrow) (D) in MHV-1 infected mice A-D = 23x, (n=6).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



