Submitted:
30 December 2023
Posted:
03 January 2024
You are already at the latest version
Abstract
Telomeres – special DNA-protein structures at the ends of linear eukaryotic chromosomes define the proliferation potential of cells. Extremely short telomeres promote DNA damage response and cell death to eliminate cells potentially accumulated mutations after multiple divisions. However, telomere elongation is associated with increased proliferative potential of special types of cells, such as stem and germ cells, permanently, and is activated temporally in processes of activation of immune response, regeneration processes. Activation of mechanisms of telomere lengthening coupled with increased proliferation and with requirements of cells in energy and building resourses. To obtain necessary nutrients cells stimulate metabolism by switching of oxidative phosphorylation program to glycolysis. In this review we focused on the interconnection of metabolism program and telomere lengthening mechanisms known for the situation of the programmed activation of proliferation such as germ cells maturation, early embryonic development program and immune response activation. Here, we propose that it is possible to reprogram metabolism in order to regulate the telomere length, proliferative activity of cells that may be important for the development of approaches to regeneration, immune response modulation and cancer therapy and further investigations in this area are necessary to improve the understanding and manipulating of molecular mechanisms used for regulation of proliferation.
Keywords:
telomere
; development
; ALT
; telomerase
; metabolism
; glycolysis
; OXPHOS
1. Introduction
Cell proliferation depends on the availability of intracellular resources and is tightly coordinated with metabolism. To divide cell should accumulate proteins and lipids and replicate the DNA in order to produce daughter cell. Eukaryotic cells could not divide endlessly; the end-replication problem restricts cellular proliferation capacity [1,2]. Telomeres – repetitive sequences located at the end of linear chromosomes provide the defense of genetic information from loss during the replication process and discrimination from internal double-stranded breaks. However, telomeres shorten in each cellular division and once they reach critical length, cells should die [3,4]. This mechanism helps a multicellular organism get rid of cells that may have gathered mutations due to multiple replication rounds. Telomeres shorten and this limits the number of times human cells can divide. This is usually enough for regular cells, but not for special cells like stem and germ cells [5,6]. Tissue regeneration, immune response, and cancer transformation all require increased cell growth. This growth is supported by the lengthening of telomeres and activation of anabolism [7,8,9].
Mainly telomerase – a specialized reverse transcriptase – is activated and elongates telomeres in vertebrates under conditions supporting the increased proliferation [10,11]. The alternative mechanism (ALT-mechanism) through a break-induced replication of telomeric DNA supports telomeres lengthening in some stages of embryonic development and in telomerase-negative cancer cells [12]. The goal of this review to summarize and discuss the information of mechanisms of telomere elongation and its regulation coupling with cellular metabolism program which are more investigated for early development process.
2. Telomeres: structure and regulation
The very ends of linear eukaryotic chromosomes are organized in special DNA-protein structures called telomeres. Telomeres protects linear eukaryotic chromosomes from the loss of genetic information during replication and false recognition as DNA breakage sites. Mammalian telomeric DNA are composed from long double-stranded regions of TTAGGG (Figure 1) repeats several kilobases long which is flanked by a 3’-end single stranded G-overhang [13,14,15]. The length of telomeres shortens in every cell division and serves as molecular clock of the proliferative lifespan of cells. Critically short telomeres activate mechanisms of cellular senescence promoting cell death [16,17]. Telomeres may be elongated by telomerase [18] or through an alternative mechanism based on homologous recombination [19]. The initial telomere length of somatic cells is derived from germline cells [20]. Stem cells need to preserve telomere length so they can regenerate effectively. Cancer cells also use mechanisms to maintain telomere length, which allows them to keep dividing for longer periods. Telomere length is important for the lifespan of cells and organisms. If it is not properly regulated, it can lead to issues with regeneration, premature aging, and cancer progression.
Shelterin complex associates with mammalian telomeres and regulates various aspects of telomere function [21]. Shelterin organizes telomeric regions in different DNA-protein structures (Figure 1), such as telomeric loops (T-loops), facilitating the invasion of ssDNA telomeric 3’-overhang into double-stranded region, regulates recruitment of telomerase for elongation of 3’-end and CTC1-STN1-TEN1 complex (CST) following by telomeric C-strand fill-in by pol a-primase to generate dsDNA telomeric region. Shelterin contains six protein subunits: telomeric repeat binding proteins 1 and 2 (TRF1 and TRF2), TERF1-interacting nuclear factor 2 (TIN2), protection of telomeres protein 1 (POT1), POT1 and TIN2-interacting protein (TPP1) and repressor/activator site-binding protein (RAP1) [22,23,24,25].
TRF1 and TRF2 are homodimers that bind telomeric dsDNA through C-terminal Myb DNA-binding domains. N-terminal TRFH-domains of TRF1 and TRF2 interact with TRFH-binding domain of TIN2 [26,27,28,29,30]. Acidic TRFH-domain of TRF1 recruits poly(ADP-ribose) polymerase tankyrase 1 to inhibit the condensation of telomeric DNA [31]. The TRFH domain of TRF2 is involved in protecting and regulating T-loops [32], Holliday junctions [33], and G4-quadruplexes [34,35]. It also interacts with histones [36] and inhibits DNA double-strand break repair alongside RAP1 [37,38,39].
Heterodimers of POT1-TPP1 recruits to TIN2 through association of TIN2 binding domain of TPP1 with TIN2N (TRFH-like) domain of TIN2 [40,41]. POT1 prefers to bind ssDNA and the addition of TPP1 increases the affinity of POT1 for DNA tenfold. Despite the preference of POT1 and POT1-TPP1 heterodimer for the ssDNA and for capping the 3’-overhang they also bind internal telomeric ssDNA. POT1 interaction with DNA is mediated through two oligonucleotide- or oligosaccharide-binding structural (OB-fold) domains. The OB3 and Holliday junction resolvase-like (HJR) domains of POT1 facilitate interaction with TPP1. The HJR-domain of POT1 may be involved in regulation of organization of T-loop which is proposed to form Holliday junction structures at telomeres [38,42,43].
Shelterin contains 6 subunits, however, in the context of telomeres it demonstrates highly dynamic complex composition. The most abundant complex contains two TRF2 subunits, two RAP1 and one TIN2 subunit. A lot of shelterin complexes consists of two TRF2, two TRF1, two RAP1 and one TIN2 subunits and the minor fraction of shelterin contains all six subunits including TPP1-POT1 heterodimer. Indeed, weak affinity of interactions between shelterin components allows adjusting the composition of the complex to the context of DNA structures and other conditions [44,45,46,47,48].
Shelterin caps the ends of chromosomes and organizes the T-loops structures but the long double stranded region of telomeric DNA is packed with nucleosomes. The telomeric nucleosomes are less stable and more dynamic structures with unwrapping of DNA ends compared to canonical nucleosomes [49,50,51]. Human telomeric chromatin is enriched in histone H3 trimethylated at Lys9 (H3K9me3), the mark of constitutive heterochromatin, which is coexists in telomeres with H3K27me3, mark of facultative heterochromatin. Taken together we can overview telomeric structure at two different scales: higher-order chromatin architecture that regulates telomeres accessibility and more compacted structure of telomeric ends affecting the telomere replication specific mechanisms (Figure 1,2).
Besides T-loops telomeric DNA is involved in formation of another two noncanonical structures: G-rich chain forms G-quadruplexes and transcription of telomeric regions stimulates formation of R-loops (Figure 1). For a long time, telomeric region of chromosome was possessed transcriptionally silent. However, recently the transcription resulting in synthesis of long non-coding RNA named TElomere Repeat containing RNA (TERRA) was reported [52,53]. RNA polymerase II starts from promoters in the subtelomeric region and synthesizes G-rich heterogeneous in length (from 100 nts to more than 9000 nts in mammals) RNA consisting of 5’-UUAGGG-3’ repeats in direction to the end of chromosomes. It is known that highly transcribed regions, and regions where RNA polymerase may be paused, are prone to form R-loops as a three-stranded nucleic acid structure containing RNA:DNA hybrid and a displaced single-stranded DNA [54]. R-loops formation is closely associated with transcription and function in a variety of physiological processes [55]. The formation of R-loops by TERRA and telomeric DNA was proposed at the moment when transcription of telomeres was uncovered and confirmed later. Telomeres contain all features promoting R-loop formation, such as G-rich single-stranded DNA forms a G4-quadruplex, repeated sequence. Increased level of TERRA correlates with accumulation of R-loops at telomeres and it was demonstrated that R-loops level is higher at critically short telomeres [56,57]. Indeed, TERRA R-loops are stabilized at critically short telomeres, interferes with replication that causes replication stress and double-stranded breaks induction [58]. Activated DNA damage response promotes homology-directed repair to elongate telomeres. Interesting, that R-loops may be formed by TERRA not only co-transcriptionally. Their appearance is regulated by homologous recombination protein RAD51 which promotes invasion of TERRA molecules in telomeric dsDNA independently of transcription [59].
The accumulation of R-loops promotes the formation of other noncanonical structures. G-rich strand which is released from dsDNA in the process of R-loops formation forms G4-quadruplex structures that is stimulated by RAD51AP1 binding with R-loops structures [60]. Interesting, that G4 structures prevent dsDNA formation after degradation of TERRA that promotes G-rich strand invasion to form telomeric D-loops that facilitate HDR. Moreover, G4-quadruplex structures are formed by G-rich single stranded 3’-overhang that is generated by telomerase. POT1 shelterin protein is able to disrupt G4 formation regulating by this way the telomere structure and telomerase binding [61,62]. Surprisingly, G4 quadruplex at telomeric 3’-overhang may add telomerase processivity [63,64]. Additionally, structural data show that telomerase has a specific space for binding G4-quadruplexes [65]. The RNA template of telomerase can directly interact with G4-structures, which helps in disrupting them when telomerase moves [66].
Taken together, the diversity of different structures formed by telomeric DNA together and with the assistance of the majority of proteins allows adapting the structure of each telomere to the environment and it is possible to conclude that there are no two identical telomeres in the cell.
3. Mechanisms of telomere length maintenance
In majority cells of vertebrate telomeres shorten in each division cycle because of the end-replication problem and nuclease action. However, telomeres must be maintained in cells with increased proliferation potential, such as stem cells, cancer cells. At the beginning of embryonic development, telomeres elongated very efficiently to obtain the length necessary for many divisions before birth and throughout the life of the organism. There are two basic mechanisms of elongation of telomeres known to be used in cells: elongation of 3’-overhang by telomerase followed by filling the C-rich by DNA polymerase a and alternative mechanism (ALT) based on homology recombination.
3.1. Telomere lengthening by telomerase
Telomerase reverse transcriptase elongates the 3’-overhang of telomeric DNA copying template region of telomerase RNA component [10,11]. Telomeric 3’-overhang is used as primers base pairing of which with 3’-end of template region of TR initiates telomerase catalytic cycle (Figure 1 and Figure 2). RNA-protein interaction inside the telomerase complex causes the 5'-end of the template region to loop out. The formation of this loop facilitates to control the position of the 3’-OH group of telomeric end [67,68]. Binding of DNA-RNA duplex and dNTPs is necessary to initiate conformation changes in active center resulting in elongation-competent structure. After formation of closed active center telomerase adds nucleotides to the 3’-end of primer until it reaches the 5’ boundary of the template region [69]. During synthesis of telomeric repeats 5’ looped out part of template region stretches out and occurs in active site of telomerase displacing 3’ template region and flanking RNA [70]. Displacing the template 3' end destroys this part of the DNA-RNA duplex and helps open the active site for complete strand separation and template release. However, newly synthesized telomeric repeat could retain in TERT while the template translocates. During template translocation its 3’ end may form duplex with 3’ end of DNA which should be captured in active site of telomerase and new telomeric repeat will be added [71]. Alternatively, template RNA and DNA can separate from telomerase. The active site needs to be reestablished for TERT to start a new catalytic cycle.
The interaction with telomere is the first and main step of the catalytic cycle of telomerase. Dynamic structure of telomere and shelterin play an important role in regulating telomeres accessibility for telomerase and telomerase activity. Shelterin can protect telomeres by stimulating T-loops formation and/or end-capping the telomeric 3’-overhang [22]. Alternatively, it can recruit telomerase to telomere and stimulate telomerase processivity in multiple telomere repeats adding [72,73,74]. The TEL-patch (TPP1 glutamate (E) and leucine (L)-rich patch) of the OB-fold of TPP1 directly interacts with the TEN (telomerase essential N-terminal) domain of TERT during the S-phase of cell cycle after genome replication. The process of this interaction is highly dynamic and multiple tentative interactions occur before stable contact will be stabilized through base pairing of telomeric end with template region of telomerase RNA [75,76]. TPP1 should help telomerase designate the telomeric tail and exclude binding with internal regions of telomeres. It was demonstrated that POT1 localizations to telomeres is regulated by its association with TPP1 [40,77]. It was proposed that a preliminary formed TPP1-POT1 heterodimer could transiently interact with TIN2-TRF1-TRF2 complexes. Sliding and hopping along telomeric dsDNA is possibly due to low affinity binding between these two complexes in context of dsDNA. Finally, the TPP1-POT1 complex finds the ssDNA-dsDNA junction where stable protein DNA complex forms, in which TPP1-POT1 interacts with ssDNA and TRF1, TRF2 are bound to the dsDNA. The localization of TPP1-POT1 at the junction of ssDNA-dsDNA triggers the activation of telomerase at the 3’ end [25].
Interestingly, POT1 has controversial action in order to regulate telomerase attraction. When it is bound at the internal regions of ssDNA overhang it stimulates telomerase association and activity, however, in case of the POT1 interaction with the very end of telomere it will block telomerase binding and inhibit telomere lengthening [78]. Moreover, POT1 in coordination with TERRA and hnRNPA1 [79] removes from telomeric ssDNA replication protein A (RPA) which inhibits telomerase activity [80]. POT1 binding is necessary to protect telomeric ssDNA [81,82] and its association with TPP1 stimulates telomerase processivity [83].
Telomere extension by telomerase is terminated by heterotrimeric CTC1-STN1-TEN1 (CST) complex [84] (Figure 1). CST is an RPA-like ssDNA-binding protein with specificity to short (<30 nucleotides) G-rich sequences [85,86]. It prefers to interact with ssDNA-dsDNA junctions, specialized structures at DNA replication and breakage sites [87,88]. It can also bind non-telomeric sequences within longer ssDNA (>30 nucleotides). Structural data revealed one telomeric ssDNA OB-domain and five additional OB-domains without sequence specificity [89]. CST also interacts and unfolds G4 DNA structures, helping to restore stalled replication forks at telomeres. Besides action at stalled replication fork at telomeres and genome-wide CST inhibits telomere 3’overhang elongation by telomerase [90]. The binding and unfolding of G4 DNA structures by CST at newly synthesized G-rich strand may lead to the prevention of telomerase translocation [65]. CST is recruited to the telomeres by TPP1 during the late S/G2 phase of the cell cycle, where it competes with telomerase [84]. However, the mechanism of CST action at telomeres in telomerase regulating manner is not clear. It is proposed that CST unfolds G4 DNA structures and inhibits telomerase translocation, however, TPP1-POT1 can passively disrupt G4 with opposite regulatory effect on telomerase activity [62]. It was proposed that TPP1-POT1 and CST work at different stages of telomerase catalytic cycle [25]. TPP1-POT1 stimulates the interaction of telomerase with 3’overhang and CST acts when telomerase should translocate along newly synthesized telomeric DNA. However, CST terminates elongation of telomeres by telomerase and recruits pol a-primase to fill in telomeric C-strand [84]. Pol a-primase demonstrates two activities: it synthesizes RNA primer, which is used for DNA polymerization when switching of action from RNA to DNA synthesis occurs. CST stimulates both activity of pol a-primase, but details of his action are still unclear [91,92,93].
3.2. ALTernative mechanism of telomere lengthening
In most cases, telomeres are elongated by telomerase expression which should be reactivated [21] by mutations in promoter of TERT gene or by genomic rearrangement upstream of TERT promoter [94,95,96]. However, TERT may be permanently repressed in some tumours and in order to survive cells activate DNA repair pathways, particularly homologous recombination (HR) to maintain telomere length by the ALT mechanism (Figure 3) [19]. The ALT pathway is associated with inactivation of a-thalassemia/mental retardation syndrome, X-linked (ATRX) and death domain-associated protein (DAXX) [97]. ATRX and DAXX are the components of a multifunctional chromatin-remodelling histone chaperone complex that is responsible for the replication-independent localization of histone H3.3 at telomeres and pericentromeric chromatin [98,99,100]. ATRX interacts with DNA methyltransferase 1 (DNMT1). Mutations of lysine 27 (K27M) in histones H3.1 and H3.3 have been identified in some cases of glioblastomas [101]. Moreover, mutations in isocitrate dehydrogenase (IDH1) [102], which regulates the production of a-ketoglutarate that is required for the enzymatic demethylation of histones and DNA were found in ALT-tumours (low-grade astrocytoma and multiform glioblastoma) [103]. All mutations which are associated with ALT-mechanism of telomeres maintenance promote considerable remodification and expansion of telomeric chromatin which becomes acceptable for HDR mechanisms.
Alternative lengthening of telomeres is carried out by proteins which participate in homology recombination mechanism. Telomeric DNA synthesis in ALT-cells involves both intra- and inter-telomeric recombination and replication. ALT-cells are characterized with several hallmarks at telomeres: specialized ALT-associated promyelocytic leukemia foci (APBs), heterogeneous telomere length, abundant extrachromosomal telomeric sequences (ECTS) such as C-circles, and high-level of telomere sister chromatid exchange (T-SCE) [12,104,105]. APBs are unique specific to ALT-cells nuclear structures containing the PML-protein and telomeric DNA. In APBs clusters of telomeres and shelterin associate with recombinase RAD51, ssDNA-binding replication protein (RPA) [106], DNA resection MRE11-RAD50-NBS1 (MRN) complex [107], Bloom helicase (BLM) [108] and other HR accessory factors such as FANCM, BRCA1 [109], BRCA2, RAD51AP1 [110] and RAD52 [104]. There are two distinct types of ALT-mechanism of telomere lengthening were considered: RAD51-dependent and -independent [12].
The structural features of telomeric GC-rich sequences with G4 structures and R-loops formed with the help of TERRA hamper the replication process of these regions and leads to increased level of replication fork stalling [111]. DNA damage sensor kinase, ataxia-telangiectasia and Rad3-related (ATR) detects replisome collisions and recruits the RPA to single-stranded telomeric DNA [112]. Stalled replication forks are subsequently repaired and restarted by remodeling enzymes such as ATP-dependent strand annealing helicase SMARCAL1 [113], and/or DNA-dependent ATPase/translocase subunit of Fanconi anemia core complex (FANCM) [109,114,115]. FANCM also resolves aberrant R-loops which may be accumulated due to transcription replication conflicts at telomeric regions [116]. Hybrid RNA-DNA duplexes between TERRA and telomeric DNA are regulated by RNaseH with DNA/RNA-helicases, including RTEL1, FANCM, PIF1 [58,109,114,116,117]. SMARCAL1 and FANCM activity is important for replication fork reversal and repair. Depletion or disruption of SMARCAL1 and FANCM result in increased number and size of APBs [113,114,115,118].
After stabilization and repair replication fork may be restored by RAD51-dependent HR. RAD51 forms the presynaptic filaments at ssDNA and probes for identical telomeric sequences as a template for error-free repair [119]. Search of homologous template by RAD51 is coregulated and stimulated by BRCA1, BRCA2, RAD51AP1 and SMC5/6 heterodimer [110,119,120]. Surprisingly, meiotic-specific HOP2-MND1 heterodimer is involved in regulation of HR at telomeric in ALT-cells [119]. The alignment of homologous DNA strands can be stimulated by nuclear F-actin filaments. This allows the presynaptic filament to invade and form a synaptic complex, displacing the DNA strand in the D-loop [121]. Loading of terminal 3’-hydroxyl group promoted by PCNA and RFC to prime and initiate semi-conservative replication by Polh which is replaced by Pold for more processive synthesis [122]. Effective telomeric DNA synthesis requires the effective branch migration and resolving of D-loops and Holliday junctions which are stimulated by translocase activity of FANCM and unwinding and resolving of decatenates activity of BLM-TOP3A-RMI1/2 (BTR) complex [123,124,125]. The activity of BTR complex is necessary to suppress t-SCE events that preserves the original orientation of telomeric DNA strands [126].
In case when stalled replication fork is not reversed BLM helicase facilitates EXO1-DNA2 dependent resection of DNA in RAD51-independent HDR pathway [127]. RAD52 may be recruited to stabilize these HR intermediates and restrict resection, as suggested by studies [128,129]. RAD52 facilitates intrachromosomal pairing of telomeric DNA sequences of proximal sister chromatids and D-loop formation which are necessary to initiate break-induced replication (BIR)-related DNA synthesis at ALT telomeres [12,130]. These mechanisms occur during G2 and M phases of the cell cycle in contrast with RAD51-dependent mechanism which is realized during the S phase.
If stalled replication forks are not resolved by the HR or HDR pathways prior to entering mitosis, they should be eliminated. SLX1-SLX4, MUS81-EME1, and XPF-ERCC1 (SMX) complex creates double-strand breaks. PCNA-RFC-Pold can utilize these breaks for DNA synthesis [131]. RAD52 stimulates mitotic DNA synthesis (MiDaS) at chromosome arms and telomeres [132,133,134] which is initiated by SLX4 and SLX4IPand results in unprocessive abortive telomere synthesis, telomere cleavage and genome instability [130].
4. Metabolism
4.1. An overview of glucose metabolism
In somatic cells, telomeres shorten with each cellular division because of end-replication problem when endless RNA-primer is degraded and gap cannot be filled with the canonical mechanism working with intrachromosomal gaps. Nuclease processing further shortens telomeres. Critically short telomeres lose their protective properties and are discriminated as double stranded breaks. Cells activate senescence program and induce chromothripsis and polyploidization [135,136,137] and after crisis should be eliminated by authophagy [138]. Cancer cells activate telomere maintenance mechanisms to achieve replicative immortality.
Telomere maintenance mechanisms are not only activated during cancer transformation, but also during certain physiological stages and processes that require an increase in cell proliferation rate [139,140]. Proliferation rate activation requires the increased level of nutrients for synthesis of components necessary for building new cells. Moreover, it is becoming clear that metabolic pathways can also play modulatory or instructive roles in the regulation of cellular programs, which can be summarized as metabolic signaling functions [141,142].
The majority of cells in normal physiological status use the Oxidative Phosphorylation (OxPhos) as main metaboliс program. However, in cases of increased proliferation rate and deficiency of oxygen OxPhos is switched to the glycolysis program, which generates less molecules of ATP per round in comparison with OxPhos, however, the turning rate is increased and moreover during glycolysis cell obtains other compounds necessary for synthesis of nucleotides, aminoacids and lipids (Figure 4) [141,142,143]. Intermediate metabolites affect downstream biochemical reactions and protein modifications, such as protein acetylation, glycosylation, and methylation [144].
It is well known the regulation of metabolism in time and space, at the intercellular and tissue level, subcellular level. The metabolism is regulated during cell cycle progression. Glycolysis is activated during the G1/S transition, while mitochondrial respiration increases during the G2/M transition [145,146,147,148]. The reduced OXPHOS activity at the stage of DNA replication should minimize the risk of oxidative damage of DNA by ROS produced by mitochondria. Moreover, acetyl-CoA the main substrate for energy production by mitochondria, is also used for the acetylation of histones promoting the epigenetic regulation of gene expression and its level should be enhanced during DNA synthesis and epigenetic marks establish [149]. The epigenetic state of chromatin is also regulated by the level of other metabolic intermediates. The level of a-ketoglutarate is important for the maintain the methylation of DNA and histones regulating the expression activity of the genome [150].
It is interesting that the cell cycle phase when telomerase elongates telomeres is characterized by glycolytic metabolism program and, moreover, cells with increased proliferation rate where telomeres should be elongated also reduce mitochondrial OXPHOS activity and enhance the glycolysis to obtain resources necessary for synthesis of compounds for new cells.
The influence of short telomeres on the metabolism was reviewed several years ago [151]. Here we want to summarize and discuss the role of metabolism in activation of mechanisms of telomere lengthening. Cancer cells activate glycolysis to get the nutrients needed for making new cells. In transformation and cancer progression, the cells' environment changes. For example, it was demonstrated that at the first steps of tumour growth cells are in the conditions of hypoxia, but further development promotes the vascular growth and level of oxygenation reverses to the normal. Moreover, cancer cells activate the glycolysis by the mechanism known as Warburg effect, but OxPhos mechanism is hyperactivated by tumour cells during cancer progression [152]. Cancer cells multiply faster because of changes in certain mechanisms that lengthen telomeres. These changes can be caused by mutations in promoter regions [94,95,96] or a decrease in repair processes. This leads to the activation of a mechanism called ALT, which is based on a type of DNA recombination [19].
This review aims to discuss how telomere lengthening is regulated during processes like development and immune response activation. In these cases, the activation of telomere lengthening should be transitional and switched off after a short period of telomere elongation. We will focus on the mechanisms of switching the metabolism program and telomere lengthening mechanisms during development and T-cell activation because both these topics are investigated much better than the other examples of short-term activation of telomere lengthening coupled with increased proliferation.
4.2. Metabolism and telomere lengthening during early development and in T-cell activation
Mammalian organisms start to develop from fertilization, where two specialized cells, the spermatozoa and the oocyte, fuse together to form a totipotent 1-cell zygote. Further dividing coupled with differentiation of obtained new cells give rise the whole embryo and extra-embryonic tissues, such as a placenta. Successful development of new organism requires proper reorganization of chromatin restricted in time with the coordination of stage of development. Proper telomere length is crucial for organizing the genome of eukaryotic organisms. It enables cells to divide multiple times during development. During development, both mechanisms of telomere lengthening are used at different stages.
Gametogenesis represents a pivotal phase preceding embryonic development, wherein germline cells, having colonized the gonadal tissue, engage in successive processes of active proliferation and meiotic division, leading to substantial cellular specialization. In the field of telomere biology, a prevailing consensus asserts that germ cells prevent the attrition of telomeres through the active expression of telomerase, and then, as embryonic development ensues, telomeres undergo gradual reduction with each cycle of DNA replication [20]. Nonetheless, there exists a significant disparity in telomerase activity between oogenesis and spermatogenesis. Sperm cells go through a maturation process that increases their telomerase activity. This leads to the sperm cells having long telomeres when they are fully matured [153]. It was observed that late-generations of mice lacking telomerase RNA (mTR-/-) have unfunctional germ cells. Male germ cells, which require strong telomerase activity for their efficient maintenance, are depleted in these animals. The majority of research findings have consistently indicated that men experiencing idiopathic infertility tend to exhibit shorter telomere lengths in comparison to their fertile counterparts [154]. Telomere length is positively correlated with sperm count, motility, and the ratio of high-quality and transferable embryos [155]. However, it is negatively associated with sperm DNA fragmentation [156,157]. Abundant evidence underscores the significant role of reactive oxygen species (ROS)-mediated sperm damage as a primary contributing factor to infertility among patients [158]. Sperm, due to their relatively limited antioxidant defense mechanisms, are particularly vulnerable to elevated oxidative stress levels. Oxidative stress can be risky for sperm DNA and RNA, and can also affect the integrity of telomeres and telomerase activity. This can lead to shorter telomeres [159]. As counterintuitive as it might appear, several studies have demonstrated an increase in telomerase activity and telomere length in human spermatozoa with advancing age [160]. Furthermore, the significance of this issue is heightened by findings from a mouse study which demonstrated that spermatozoa telomeres serve as a template for telomerase-independent telomere lengthening, leading to persistent differences in telomere length after birth [161]. Consequently, contemporary research places considerable emphasis on telomere length as a highly promising marker for evaluating male reproductive biology [162].
In contrast to the general trend of "greater potency equating longer telomeres," the female germline in many mammalian species stands as an exception. Notably, telomeres within mouse and human oocytes are among the shortest observed throughout the body, and they exhibit a low to absent level of telomerase activity [20,163]. The divergence in telomerase activity between oogenesis and spermatogenesis manifests with varying degrees across distinct mammalian species. A noteworthy illustration can be found in the Tasmanian devil and other marsupials, where an exceptional degree of telomere length dimorphism in gametes has been elucidated. This phenomenon involves a distinctive pattern: all telomeres in spermatozoa undergo hyperelongation, whereas those in oocytes experience hypershortening [164].
Nonetheless, in the case of mice, humans, and many other mammals, oocyte telomere length is delicately balanced and serves as an indicator of their quality. Telomerase activity measurements conducted through in vitro TRAP assays have unveiled a distinctive pattern: a peak in pre-ovulation oocytes followed by a subsequent decline in mature oocytes [165,166]. The resultant telomere length is directly associated with the quality of the oocyte and, in turn, influences its developmental potential. Oocytes with shortened telomeres resulting from telomerase-null mice exhibit a striking pattern of failure, occurring during both fertilization and the early cleavage embryonic stages [167]. This observation supports the hypothesis that low telomerase activity may serve as a selective mechanism, favoring the successful fertilization of egg cells that have already attained adequate telomere length during oogenesis [168]. This is predominantly because telomere attrition substantially promotes genomic instability through mechanisms like non-homologous end joining, ultimately resulting in conditions such as aneuploidy, mosaicism, and the emergence of copy number variants [137]. The shortening of telomeres in telomerase-null mice has been linked to several detrimental outcomes in oocyte development. This includes the formation of abnormal meiotic spindles [167], the arrest and fragmentation of embryos [169], a decrease in chiasmata and synapsis [170], and ultimately, infertility. Furthermore, oocyte telomere length has been associated with various challenges in in vitro fertilization (IVF) cycles, such as failed IVF cycles, embryo fragmentation [169], and blastocyst aneuploidy [171]. These findings underscore the critical role of telomere length in oocyte health and reproductive success. The diminishing ovarian reserve that occurs in tandem with advancing female age leads to a concomitant telomere depletion, which holds a central role in the process of oocyte aging. This is underscored by the observation of telomere shortening in oocytes obtained from females of reproductive age. The shortening of telomeres in oocytes could have served as an evolutionary selective mechanism actual during historical periods when the risks associated with grand-multiparity and advanced maternal age led to pronounced maternal mortality rates [172]. The age-related telomere shortening is primarily attributed to the enduring negative consequences of reduced telomerase activity and heightened exposure to reactive oxygen species (ROS) [173]. Research findings have demonstrated that the application of antioxidants enhances the quality of oocytes obtained from older females, including an increase in both telomerase activity and telomere length [174].
The process of oogenesis occurs in close collaboration with somatic follicular cells, specifically granulosa cells. These cells not only provide support for oocyte maturation but also play a crucial role in the synthesis of estrogen. During the period of follicular growth, granulosa cells exhibit robust proliferative activity and possess elevated levels of telomerase activity [175,176,177,178]. The length of telomeres in granulosa cells plays a pivotal role in regulating the normal progression of folliculogenesis and overall ovarian function. Reducing telomerase activity in granulosa cells has been associated with an elevated rate of apoptosis and an increase in the number of atretic (degenerating) follicles [176,178]. In granulosa cells, the activity of telomerase and the length of telomeres are regulated by estrogen levels [179,180]. High concentrations of estradiol-17β have been shown to significantly increase the telomere length of granulosa cells cultured in vitro [181]. Conversely, the withdrawal of estrogen consistently results in reduced telomerase activity, which may lead to telomere shortening in granulosa cells, subsequently contributing to follicular atresia [178,182]. Short telomere length and the absence of telomerase activity in the granulosa cells of women have been associated with occult ovarian insufficiency [183]. Indeed, short telomere length has been reported in young patients who exhibit a low ovarian response to hormonal stimulation [184]. This observation highlights the potential significance of telomere length as a predictor or indicator of ovarian responsiveness and reproductive health in individuals of reproductive age. During ovulation, a subset of the follicular cells that envelop the oocyte accompanies it as part of the cumulus-oocyte complex. These cumulus cells can be conveniently obtained alongside oocytes, presenting significant diagnostic potential when employing assisted reproductive technologies. The relative telomere length tends to be greater in cumulus cells derived from good-quality embryos as compared to cumulus cells associated with embryos of lesser quality [185]. This finding highlights the utility of assessing telomere length in cumulus cells as an effective means of evaluating embryo quality in the context of assisted reproduction.
The early stages of mammalian development involve a dynamic regulation of telomere length in embryonic cells, as observed in various mammalian species, including mice, rats, cows, and humans (Figure 5). Telomere length tends to increase during preimplantation development, reaching its peak at the blastocyst stage [186,187]. Remarkably, this elongation process occurs even in the absence of telomerase activity during the early cleavage stages, spanning from 2-4 cells up to the morula stage. Telomeres undergo elongation through an alternative lengthening mechanism (ALT-like mechanism) during these early developmental stages. The recombination mediated telomere lengthening at the early cleavage stage is driven by the telomeric chromatin reorganization due to H3K9 demethylation by KDM4 and Zscan4. The maintenance of Zscan4 activity in early embryos and 2-cell-like embryonic stem cells is facilitated by Dcaf11 (Ddb1- and Cul4-associated factor 11) [188]. Moreover, the transcription of TERRA is activated during early cleavage stages confirming the open conformation of telomeric chromatin. TERRA accumulation at telomeres promotes the ATRX recruitment followed by attraction of HP1 and compactization of the telomeres at the morula stage, accompanied by inhibition of ALT-like mechanism telomere elongation and activation of telomerase to provide the telomere lengthening at blastocyst stage [189].
Interestingly, that zygotic gene activation, which occurs at the early cleavage stage, is supported by the regulation of metabolic program. During the early cleavage stage of development, the mitochondrial enzymes responsible for the production of acetyl-CoA and aKG are transiently localized to the nucleus where they impact the epigenetic histone modification to promote the chromatin opening. It was demonstrated that mitochondrial pyruvate dehydrogenase complex is phosphorylated and inhibited in cleavage-stage embryos that results in decreased activity of respiratory chain and OxPhos, but nuclear PDH remains unphosphorylated and active that influences on the epigenetic regulation of genome expression. Pyruvate metabolism supports development of embryo upon fertilization and glucose catabolism starts to be activated at the eight-cell stage and should be associated with inhibition of ALT-like telomere lengthening and activation of telomerase which elongates telomeres up to blastocyst stage and is inactivated during cell differentiation [190].
At the blastocyst stage, the maximum telomere length is observed, which exceeds that observed both at earlier stages of preimplantation development and during subsequent embryogenesis [191]. Telomerase activity emerged as a crucial factor driving telomere elongation during the transition from morula to blastocyst. Analysis of telomere length in 8-cell, morula, and blastocyst-stage embryos obtained from mTERC−/− or mTERC+/+ mouse models revealed that blastocysts from mTERC+/+ mice displayed significantly longer telomeres compared to those at the 8-cell and morula stages. Conversely, blastocysts from mTERC−/− mice did not exhibit this telomere elongation [163,192]. The blastocyst comprises two main cell types: the trophectoderm (TE) covering the outer surface of the embryo and the inner cell mass (ICM) situated inside of the blastocyst. While the ICM forms embryonic structures, the TE has more limited potential and contributes to the development of extraembryonic membranes. Interestingly, despite these distinctions in cell fate, telomere length exhibits only minor variations between the cells of the inner cell mass and the trophoblast [193]. Telomere elongation during the blastocyst stage significantly influences telomere length during subsequent stages of development. In the comparison of telomerase activity in bovine embryos undergoing development in vivo, in vitro, as well as after in vitro fertilization (IVF), and parthenogenetic activation, researchers noted only minor quantitative differences [186]. However, a recent study involving children born through assisted reproductive technology (ART) revealed that those born following a blastocyst transplant procedure exhibited shorter telomeres in their white blood cells by the age of one year [194]. Furthermore, research conducted in mice has shown that in vitro culture of mouse embryos suppresses telomerase activity during the early blastocyst stage, which subsequently leads to telomere shortening [194]. The intricate and dynamic regulation of telomere length during early development renders this mechanism susceptible to various negative influences, both external and internal. For instance, studies have demonstrated that a high-fat diet and obesity in female mice can lead to reduced telomerase activity and telomere shortening in oocytes and early embryos [195].
Embryonic stem cells (ESC) obtained from the inner cell mass (ICM) of a blastocyst have longer telomeres than mouse embryonic fibroblasts (MEF) of the same genetic background, which are typically obtained at embryonic day 13.5. It confirms that telomeres shorten after the blastocyst stage during cell differentiation to promote embryo development. Generation of induced pluripotent cells (iPSCs) from MEFs is accompanied by the telomere elongation up to length similar to the telomeres in ESCs. Moreover, telomeres are elongated in MEFs during cultivation and at early passages (P5) their length is comparable with the telomeres in ICM of blastocyst, however, telomeres length increased up to 24 passage and maintained constant for a long time. It was demonstrated the decreased localization of heterochromatin markers H3K9me3 and H4K20me3 at telomeric regions of ESCs and in iPSCs that could activate the mechanisms of telomere lengthening [191].
Interestingly, correlation between metabolism program, telomere length and ability to differentiate was demonstrated for human embryonic stem cells (hESC) recently. It was determined that the expression level of genes involved in OxPhos metabolic pathway is increased in hESCs with short telomeres, however, the expression level of genes related to glycolysis metabolism is up-regulated in cells with long telomeres [196]. Moreover, pluripotent ESCs was characterized with high glycolysis activity, but primed and differentiated cells demonstrate the increased OxPhos activity [197].
The switching between different metabolic programs in T cells during their development and maintaining is well studied. Quiescent T cells use catabolic pathways such as OxPhos, which provide efficient robust energy output, while activation of T cells is supported with anabolic glycolysis pathway, which provide nutrients necessary for protein production and cell division. Activation of T cells requires the increased proliferation which is accompanied by telomerase activation and telomere lengthening [140,198]. Naïve and memory T cells maintain their energy levels by relying on OxPhos, as their energy demand is relatively low during homeostasis. Upon activation, T cells proliferate at an incredibly rate and differentiate into effector cells. Early upregulation of glycolysis during T cells activation is supported with activation of PDH kinase 1, which phosphorylates and inactivates PDH. The inhibition of PDH drives engagement of aerobic glycolysis [199].
Switching between metabolism programs is regulated by different aspects. Activation of OxPhos requires an increased level of healthy mitochondria, which biogenesis is stimulated by transcription factors (Nrf1 and PPARg coactivator-1a) which drive expression of nuclear encoded mitochondrial genes [200].
Anabolic growth of cells is regulated by genes which expression is activated by mammalian target of rapamycin (mTOR), a serine/threonine kinase that integrates a multiple of extra- and intracellular signals and promotes glycolysis, growth and proliferation [201,202]. mTOR is regulated by a sensor of AMP/ATP ratio AMPK which drives catabolic metabolism when energy stores are depleted. It stimulates mitochondria biogenesis and inhibits mTOR activity. Both mTOR and AMPK form an axis of reciprocal regulation of catabolic and anabolic pathways [203,204,205]. Interestingly, that hTERP protein encoded in precursor of human telomerase RNA component is involved in regulation of autophagy and cell proliferation through AMPK pathway and stimulates autophagy and proliferation [206,207]. Autophagy is activated when glycolysis is inhibited, so we could hypothesize that the switching between biogenesis pathway of primary transcript of telomerase RNA gene [208] may be regulated by intracellular signals metabolism reprogramming in order to modulate the telomerase activity in accordance with cell requirement in proliferation rate.
5. Conclusions
Cell proliferation is important for organism development, regeneration, immune system function, cancer progression, and more. Cell division relies on energy, nutrition, metabolism regulation, genome integrity, and the maintenance of telomere length and structure. In this review we performed the analysis of mechanisms of metabolism and telomere maintenance in processes of activation of proliferation. We focused on different models that have experimental data on homogeneous cellular populations. These models have well-studied metabolism reprogramming and information on telomere elongation. This review provides the evidence that it is possible to reprogram metabolism in order to regulate the telomere length, proliferative activity of cells that may be important for the development of approaches to regeneration, immune response modulation and cancer therapy and further investigations in this area are necessary to improve the understanding and manipulating of molecular mechanisms used for regulation of proliferation.
Author Contributions
Conceptualization, M.R.; writing—original draft preparation, M.R., D.N., M.K.; writing—review and editing, O.D., A.V.; supervision, O.D., A.V.; project administration, M.R..; funding acquisition, M.R., D.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant #23-SH04-20 of the Development Program of the MSU Interdisciplinary Scientific and Educational School “Molecular technologies of living systems and synthetic biology” at the Lomonosov Moscow State University to M.R. and D.N.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Olovnikov, A.M. A Theory of Marginotomy. Journal of Theoretical Biology 1973, 41, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Chow, T.T.; Zhao, Y.; Mak, S.S.; Shay, J.W.; Wright, W.E. Early and Late Steps in Telomere Overhang Processing in Normal Human Cells: The Position of the Final RNA Primer Drives Telomere Shortening. Genes & Development 2012, 26, 1167–1178. [Google Scholar] [CrossRef]
- Levy, M.Z.; Allsopp, R.C.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere End-Replication Problem and Cell Aging. Journal of Molecular Biology 1992, 225, 951–960. [Google Scholar] [CrossRef]
- Fumagalli, M.; Rossiello, F.; Clerici, M.; Barozzi, S.; Cittaro, D.; Kaplunov, J.M.; Bucci, G.; Dobreva, M.; Matti, V.; Beausejour, C.M.; et al. Telomeric DNA Damage Is Irreparable and Causes Persistent DNA-Damage-Response Activation. Nat Cell Biol 2012, 14, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.E.; Shay, J.W. Role of Telomerase in Cellular Proliferation and Cancer. J. Cell. Physiol. 1999, 180, 10–18. [Google Scholar] [CrossRef]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Weng, N. Telomere and Adaptive Immunity. Mechanisms of Ageing and Development 2008, 129, 60–66. [Google Scholar] [CrossRef]
- Weng, N.P.; Levine, B.L.; June, C.H.; Hodes, R.J. Regulated Expression of Telomerase Activity in Human T Lymphocyte Development and Activation. J. Exp. Med. 1996, 183, 2471–2479. [Google Scholar] [CrossRef]
- Lu, W.-Y.; Forbes, S.J. Telomerase Activity Links to Regenerative Capacity of Hepatocytes. Transplantation 2018, 102, 1587–1588. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. A Telomeric Sequence in the RNA of Tetrahymena Telomerase Required for Telomere Repeat Synthesis. Nature 1989, 337, 331–337. [Google Scholar] [CrossRef]
- Lingner, J.; Hughes, T.R.; Shevchenko, A.; Mann, M.; Lundblad, V.; Cech, T.R. Reverse Transcriptase Motifs in the Catalytic Subunit of Telomerase. Science 1997, 276, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; Yadav, T.; Ouyang, J.; Lan, L.; Zou, L. Alternative Lengthening of Telomeres through Two Distinct Break-Induced Replication Pathways. Cell Reports 2019, 26, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A Highly Conserved Repetitive DNA Sequence, (TTAGGG)n, Present at the Telomeres of Human Chromosomes. Proc. Natl. Acad. Sci. U.S.A. 1988, 85, 6622–6626. [Google Scholar] [CrossRef] [PubMed]
- Meyne, J.; Ratliff, R.L.; Moyzis, R.K. Conservation of the Human Telomere Sequence (TTAGGG)n among Vertebrates. Proc. Natl. Acad. Sci. U.S.A. 1989, 86, 7049–7053. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Hirose, Y.; Langmore, J.P. Long G Tails at Both Ends of Human Chromosomes Suggest a C Strand Degradation Mechanism for Telomere Shortening. Cell 1997, 88, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres Shorten during Ageing of Human Fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, R.C.; Vaziri, H.; Patterson, C.; Goldstein, S.; Younglai, E.V.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere Length Predicts Replicative Capacity of Human Fibroblasts. Proceedings of the National Academy of Sciences 1992, 89, 10114–10118. [Google Scholar] [CrossRef]
- Morin, G.B. The Human Telomere Terminal Transferase Enzyme Is a Ribonucleoprotein That Synthesizes TTAGGG Repeats. Cell 1989, 59, 521–529. [Google Scholar] [CrossRef]
- Bryan, T.M.; Englezou, A.; Dalla-Pozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an Alternative Mechanism for Maintaining Telomere Length in Human Tumors and Tumor-Derived Cell Lines. Nat Med 1997, 3, 1271–1274. [Google Scholar] [CrossRef]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase Activity in Human Germline and Embryonic Tissues and Cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; O’Connor, M.S.; Qin, J.; Songyang, Z. Telosome, a Mammalian Telomere-Associated Complex Formed by Multiple Telomeric Proteins. Journal of Biological Chemistry 2004, 279, 51338–51342. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P.; Cech, T.R. Pot1, the Putative Telomere End-Binding Protein in Fission Yeast and Humans. Science 2001, 292, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Cech, T.R. Shaping Human Telomeres: From Shelterin and CST Complexes to Telomeric Chromatin Organization. Nat Rev Mol Cell Biol 2021, 22, 283–298. [Google Scholar] [CrossRef]
- Bianchi, A. TRF1 Is a Dimer and Bends Telomeric DNA. The EMBO Journal 1997, 16, 1785–1794. [Google Scholar] [CrossRef]
- Broccoli, D.; Smogorzewska, A.; Chong, L.; de Lange, T. Human Telomeres Contain Two Distinct Myb–Related Proteins, TRF1 and TRF2. Nat Genet 1997, 17, 231–235. [Google Scholar] [CrossRef]
- van Steensel, B.; de Lange, T. Control of Telomere Length by the Human Telomeric Protein TRF1. Nature 1997, 385, 740–743. [Google Scholar] [CrossRef]
- Bilaud, T.; Brun, C.; Ancelin, K.; Koering, C.E.; Laroche, T.; Gilson, E. Telomeric Localization of TRF2, a Novel Human Telobox Protein. Nat Genet 1997, 17, 236–239. [Google Scholar] [CrossRef]
- Kim, S.; Kaminker, P.; Campisi, J. TIN2, a New Regulator of Telomere Length in Human Cells. Nat Genet 1999, 23, 405–412. [Google Scholar] [CrossRef]
- Seimiya, H.; Muramatsu, Y.; Smith, S.; Tsuruo, T. Functional Subdomain in the Ankyrin Domain of Tankyrase 1 Required for Poly(ADP-Ribosyl)Ation of TRF1 and Telomere Elongation. Molecular and Cellular Biology 2004, 24, 1944–1955. [Google Scholar] [CrossRef]
- M. Stansel, R. T-Loop Assembly in Vitro Involves Binding of TRF2 near the 3’ Telomeric Overhang. The EMBO Journal 2001, 20, 5532–5540. [Google Scholar] [CrossRef] [PubMed]
- Fouché, N.; Cesare, A.J.; Willcox, S.; Özgür, S.; Compton, S.A.; Griffith, J.D. The Basic Domain of TRF2 Directs Binding to DNA Junctions Irrespective of the Presence of TTAGGG Repeats. Journal of Biological Chemistry 2006, 281, 37486–37495. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.R.; Raghuraman, M.K.; Cech, T.R. Monovalent Cation-Induced Structure of Telomeric DNA: The G-Quartet Model. Cell 1989, 59, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Sundquist, W.I.; Klug, A. Telomeric DNA Dimerizes by Formation of Guanine Tetrads between Hairpin Loops. Nature 1989, 342, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Konishi, A.; Izumi, T.; Shimizu, S. TRF2 Protein Interacts with Core Histones to Stabilize Chromosome Ends. Journal of Biological Chemistry 2016, 291, 20798–20810. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; Kabir, S.; van Overbeek, M.; Celli, G.B.; de Lange, T. Loss of Rap1 Induces Telomere Recombination in the Absence of NHEJ or a DNA Damage Signal. Science 2010, 327, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Chen, Y.; Lei, M.; Chang, S. TRF2-RAP1 Is Required to Protect Telomeres from Engaging in Homologous Recombination-Mediated Deletions and Fusions. Nat Commun 2016, 7, 10881. [Google Scholar] [CrossRef]
- Lototska, L.; Yue, J.; Li, J.; Giraud-Panis, M.; Songyang, Z.; Royle, N.J.; Liti, G.; Ye, J.; Gilson, E.; Mendez-Bermudez, A. Human RAP 1 Specifically Protects Telomeres of Senescent Cells from DNA Damage. EMBO Reports 2020, 21, e49076. [Google Scholar] [CrossRef]
- Ye, J.Z.-S.; Hockemeyer, D.; Krutchinsky, A.N.; Loayza, D.; Hooper, S.M.; Chait, B.T.; de Lange, T. POT1-Interacting Protein PIP1: A Telomere Length Regulator That Recruits POT1 to the TIN2/TRF1 Complex. Genes Dev. 2004, 18, 1649–1654. [Google Scholar] [CrossRef]
- O’Connor, M.S.; Safari, A.; Xin, H.; Liu, D.; Songyang, Z. A Critical Role for TPP1 and TIN2 Interaction in High-Order Telomeric Complex Assembly. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 11874–11879. [Google Scholar] [CrossRef]
- Chen, C.; Gu, P.; Wu, J.; Chen, X.; Niu, S.; Sun, H.; Wu, L.; Li, N.; Peng, J.; Shi, S.; et al. Structural Insights into POT1-TPP1 Interaction and POT1 C-Terminal Mutations in Human Cancer. Nat Commun 2017, 8, 14929. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Podell, E.R.; Cech, T.R. Structure of Human POT1 Bound to Telomeric Single-Stranded DNA Provides a Model for Chromosome End-Protection. Nat Struct Mol Biol 2004, 11, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Rai, R.; Huang, C.; Broton, C.; Long, J.; Xu, Y.; Xue, J.; Lei, M.; Chang, S.; Chen, Y. Structural and Functional Analyses of the Mammalian TIN2-TPP1-TRF2 Telomeric Complex. Cell Res 2017, 27, 1485–1502. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Y.; van Overbeek, M.; Donigian, J.R.; Baciu, P.; de Lange, T.; Lei, M. A Shared Docking Motif in TRF1 and TRF2 Used for Differential Recruitment of Telomeric Proteins. Science 2008, 319, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Janoušková, E.; Nečasová, I.; Pavloušková, J.; Zimmermann, M.; Hluchý, M.; Marini, V.; Nováková, M.; Hofr, C. Human Rap1 Modulates TRF2 Attraction to Telomeric DNA. Nucleic Acids Research 2015, 43, 2691–2700. [Google Scholar] [CrossRef]
- Choi, K.H.; Farrell, A.S.; Lakamp, A.S.; Ouellette, M.M. Characterization of the DNA Binding Specificity of Shelterin Complexes. Nucleic Acids Research 2011, 39, 9206–9223. [Google Scholar] [CrossRef]
- Takai, K.K.; Hooper, S.; Blackwood, S.; Gandhi, R.; de Lange, T. In Vivo Stoichiometry of Shelterin Components. Journal of Biological Chemistry 2010, 285, 1457–1467. [Google Scholar] [CrossRef]
- Pisano, S.; Marchioni, E.; Galati, A.; Mechelli, R.; Savino, M.; Cacchione, S. Telomeric Nucleosomes Are Intrinsically Mobile. Journal of Molecular Biology 2007, 369, 1153–1162. [Google Scholar] [CrossRef]
- Fajkus, J.; Trifonov, E.N. Columnar Packing of Telomeric Nucleosomes. Biochemical and Biophysical Research Communications 2001, 280, 961–963. [Google Scholar] [CrossRef]
- Soman, A.; Liew, C.W.; Teo, H.L.; Berezhnoy, N.V.; Olieric, V.; Korolev, N.; Rhodes, D.; Nordenskiöld, L. The Human Telomeric Nucleosome Displays Distinct Structural and Dynamic Properties. Nucleic Acids Research 2020, 48, 5383–5396. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric Repeat Containing RNA and RNA Surveillance Factors at Mammalian Chromosome Ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally Regulated Transcription of Mammalian Telomeres by DNA-Dependent RNA Polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; White, R.L.; Davis, R.W. Hybridization of RNA to Double-Stranded DNA: Formation of R-Loops. Proc. Natl. Acad. Sci. U.S.A. 1976, 73, 2294–2298. [Google Scholar] [CrossRef] [PubMed]
- White, R.L.; Hogness, D.S. R Loop Mapping of the 18S and 28S Sequences in the Long and Short Repeating Units of Drosophila Melanogaster rDNA. Cell 1977, 10, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Graf, M.; Bonetti, D.; Lockhart, A.; Serhal, K.; Kellner, V.; Maicher, A.; Jolivet, P.; Teixeira, M.T.; Luke, B. Telomere Length Determines TERRA and R-Loop Regulation through the Cell Cycle. Cell 2017, 170, 72–85. [Google Scholar] [CrossRef]
- Balk, B.; Dees, M.; Bender, K.; Luke, B. The Differential Processing of Telomeres in Response to Increased Telomeric Transcription and RNA–DNA Hybrid Accumulation. RNA Biology 2014, 11, 95–100. [Google Scholar] [CrossRef]
- Balk, B.; Maicher, A.; Dees, M.; Klermund, J.; Luke-Glaser, S.; Bender, K.; Luke, B. Telomeric RNA-DNA Hybrids Affect Telomere-Length Dynamics and Senescence. Nat. Struct. Mol. Biol. 2013, 20, 1199–1205. [Google Scholar] [CrossRef]
- Feretzaki, M.; Pospisilova, M.; Valador Fernandes, R.; Lunardi, T.; Krejci, L.; Lingner, J. RAD51-Dependent Recruitment of TERRA lncRNA to Telomeres through R-Loops. Nature 2020, 587, 303–308. [Google Scholar] [CrossRef]
- Yadav, T.; Zhang, J.-M.; Ouyang, J.; Leung, W.; Simoneau, A.; Zou, L. TERRA and RAD51AP1 Promote Alternative Lengthening of Telomeres through an R- to D-Loop Switch. Molecular Cell 2022, 82, 3985–4000. [Google Scholar] [CrossRef]
- Wang, H.; Nora, G.J.; Ghodke, H.; Opresko, P.L. Single Molecule Studies of Physiologically Relevant Telomeric Tails Reveal POT1 Mechanism for Promoting G-Quadruplex Unfolding. Journal of Biological Chemistry 2011, 286, 7479–7489. [Google Scholar] [CrossRef] [PubMed]
- Zaug, A.J.; Podell, E.R.; Cech, T.R. Human POT1 Disrupts Telomeric G-Quadruplexes Allowing Telomerase Extension in Vitro. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 10864–10869. [Google Scholar] [CrossRef] [PubMed]
- Patrick, E.M.; Slivka, J.D.; Payne, B.; Comstock, M.J.; Schmidt, J.C. Observation of Processive Telomerase Catalysis Using High-Resolution Optical Tweezers. Nat Chem Biol 2020, 16, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Latrick, C.M.; Cech, T.R. POT1–TPP1 Enhances Telomerase Processivity by Slowing Primer Dissociation and Aiding Translocation. EMBO J 2010, 29, 924–933. [Google Scholar] [CrossRef]
- Jansson, L.I.; Hentschel, J.; Parks, J.W.; Chang, T.R.; Lu, C.; Baral, R.; Bagshaw, C.R.; Stone, M.D. Telomere DNA G-Quadruplex Folding within Actively Extending Human Telomerase. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 9350–9359. [Google Scholar] [CrossRef]
- Paudel, B.P.; Moye, A.L.; Abou Assi, H.; El-Khoury, R.; Cohen, S.B.; Holien, J.K.; Birrento, M.L.; Samosorn, S.; Intharapichai, K.; Tomlinson, C.G.; et al. A Mechanism for the Extension and Unfolding of Parallel Telomeric G-Quadruplexes by Human Telomerase at Single-Molecule Resolution. eLife 2020, 9, e56428. [Google Scholar] [CrossRef]
- Wu, R.A.; Collins, K. Human Telomerase Specialization for Repeat Synthesis by Unique Handling of Primer-Template Duplex. The EMBO Journal 2014, 33, 921–935. [Google Scholar] [CrossRef]
- Wang, H. A Novel Specificity for the Primer-Template Pairing Requirement in Tetrahymena Telomerase. The EMBO Journal 1998, 17, 1152–1160. [Google Scholar] [CrossRef]
- Brown, A.F.; Podlevsky, J.D.; Qi, X.; Chen, Y.; Xie, M.; Chen, J.J.-L. A Self-Regulating Template in Human Telomerase. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 11311–11316. [Google Scholar] [CrossRef]
- Berman, A.J.; Akiyama, B.M.; Stone, M.D.; Cech, T.R. The RNA Accordion Model for Template Positioning by Telomerase RNA during Telomeric DNA Synthesis. Nat Struct Mol Biol 2011, 18, 1371–1375. [Google Scholar] [CrossRef]
- Qi, X.; Xie, M.; Brown, A.F.; Bley, C.J.; Podlevsky, J.D.; Chen, J.J.-L. RNA/DNA Hybrid Binding Affinity Determines Telomerase Template-Translocation Efficiency. EMBO J. 2012, 31, 150–161. [Google Scholar] [CrossRef]
- Nandakumar, J.; Bell, C.F.; Weidenfeld, I.; Zaug, A.J.; Leinwand, L.A.; Cech, T.R. The TEL Patch of Telomere Protein TPP1 Mediates Telomerase Recruitment and Processivity. Nature 2012, 492, 285–289. [Google Scholar] [CrossRef]
- Zhong, F.L.; Batista, L.F.Z.; Freund, A.; Pech, M.F.; Venteicher, A.S.; Artandi, S.E. TPP1 OB-Fold Domain Controls Telomere Maintenance by Recruiting Telomerase to Chromosome Ends. Cell 2012, 150, 481–494. [Google Scholar] [CrossRef]
- Sexton, A.N.; Youmans, D.T.; Collins, K. Specificity Requirements for Human Telomere Protein Interaction with Telomerase Holoenzyme. Journal of Biological Chemistry 2012, 287, 34455–34464. [Google Scholar] [CrossRef]
- Schmidt, J.C.; Zaug, A.J.; Kufer, R.; Cech, T.R. Dynamics of Human Telomerase Recruitment Depend on Template-Telomere Base Pairing. MBoC 2018, 29, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.C.; Zaug, A.J.; Cech, T.R. Live Cell Imaging Reveals the Dynamics of Telomerase Recruitment to Telomeres. Cell 2016, 166, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Safari, A.; O’Connor, M.S.; Chan, D.W.; Laegeler, A.; Qin, J.; Songyang, Z. PTOP Interacts with POT1 and Regulates Its Localization to Telomeres. Nat Cell Biol 2004, 6, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Zaug, A.J.; Podell, E.R.; Cech, T.R. Switching Human Telomerase On and Off with hPOT1 Protein in Vitro. Journal of Biological Chemistry 2005, 280, 20449–20456. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.L.; Centore, R.C.; O’Sullivan, R.J.; Rai, R.; Tse, A.; Songyang, Z.; Chang, S.; Karlseder, J.; Zou, L. TERRA and hnRNPA1 Orchestrate an RPA-to-POT1 Switch on Telomeric Single-Stranded DNA. Nature 2011, 471, 532–536. [Google Scholar] [CrossRef]
- Rubtsova, M.P.; Skvortsov, D.A.; Petruseva, I.O.; Lavrik, O.I.; Spirin, P.V.; Prasolov, V.S.; Kisseljov, F.L.; Dontsova, O.A. Replication Protein A Modulates the Activity of Human Telomerase in Vitro. Biochemistry Mosc. 2009, 74, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Hockemeyer, D.; Sfeir, A.J.; Shay, J.W.; Wright, W.E.; de Lange, T. POT1 Protects Telomeres from a Transient DNA Damage Response and Determines How Human Chromosomes End. EMBO J 2005, 24, 2667–2678. [Google Scholar] [CrossRef]
- Denchi, E.L.; de Lange, T. Protection of Telomeres through Independent Control of ATM and ATR by TRF2 and POT1. Nature 2007, 448, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Podell, E.R.; Zaug, A.J.; Yang, Y.; Baciu, P.; Cech, T.R.; Lei, M. The POT1-TPP1 Telomere Complex Is a Telomerase Processivity Factor. Nature 2007, 445, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Y.; Redon, S.; Lingner, J. The Human CST Complex Is a Terminator of Telomerase Activity. Nature 2012, 488, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Cervantes, R.B.; Mandell, E.K.; Otero, J.H.; Lundblad, V. RPA-like Proteins Mediate Yeast Telomere Function. Nat Struct Mol Biol 2007, 14, 208–214. [Google Scholar] [CrossRef]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like Mammalian Ctc1-Stn1-Ten1 Complex Binds to Single-Stranded DNA and Protects Telomeres Independently of the Pot1 Pathway. Molecular Cell 2009, 36, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Wang, Y.; Diao, J.; Price, C.M. Dynamic DNA Binding, Junction Recognition and G4 Melting Activity Underlie the Telomeric and Genome-Wide Roles of Human CST. Nucleic Acids Research 2017, 45, 12311–12324. [Google Scholar] [CrossRef]
- Hom, R.A.; Wuttke, D.S. Human CST Prefers G-Rich but Not Necessarily Telomeric Sequences. Biochemistry 2017, 56, 4210–4218. [Google Scholar] [CrossRef]
- Lim, C.J.; Barbour, A.T.; Zaug, A.J.; Goodrich, K.J.; McKay, A.E.; Wuttke, D.S.; Cech, T.R. The Structure of Human CST Reveals a Decameric Assembly Bound to Telomeric DNA. Science 2020, 368, 1081–1085. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, B.; Li, T.; Liu, R.; Xiao, Y.; Geng, X.; Li, G.; Liu, Q.; Price, C.M.; Liu, Y.; et al. Mammalian CST Averts Replication Failure by Preventing G-Quadruplex Accumulation. Nucleic Acids Research 2019, 47, 5243–5259. [Google Scholar] [CrossRef]
- Casteel, D.E.; Zhuang, S.; Zeng, Y.; Perrino, F.W.; Boss, G.R.; Goulian, M.; Pilz, R.B. A DNA Polymerase-α·Primase Cofactor with Homology to Replication Protein A-32 Regulates DNA Replication in Mammalian Cells. Journal of Biological Chemistry 2009, 284, 5807–5818. [Google Scholar] [CrossRef] [PubMed]
- Ganduri, S.; Lue, N.F. STN1–POLA2 Interaction Provides a Basis for Primase-Pol α Stimulation by Human STN1. Nucleic Acids Research 2017, 45, 9455–9466. [Google Scholar] [CrossRef]
- Nakaoka, H.; Nishiyama, A.; Saito, M.; Ishikawa, F. Xenopus Laevis Ctc1-Stn1-Ten1 (xCST) Protein Complex Is Involved in Priming DNA Synthesis on Single-Stranded DNA Template in Xenopus Egg Extract. Journal of Biological Chemistry 2012, 287, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Akıncılar, S.C.; Khattar, E.; Boon, P.L.S.; Unal, B.; Fullwood, M.J.; Tergaonkar, V. Long-Range Chromatin Interactions Drive Mutant TERT Promoter Activation. Cancer Discovery 2016, 6, 1276–1291. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Lorbeer, F.K.; Shain, A.H.; McSwiggen, D.T.; Schruf, E.; Oh, A.; Ryu, J.; Darzacq, X.; Bastian, B.C.; Hockemeyer, D. Mutations in the Promoter of the Telomerase Gene TERT Contribute to Tumorigenesis by a Two-Step Mechanism. Science 2017, 357, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Hertwig, F.; Roels, F.; Dreidax, D.; Gartlgruber, M.; Menon, R.; Krämer, A.; Roncaioli, J.L.; Sand, F.; Heuckmann, J.M.; et al. Telomerase Activation by Genomic Rearrangements in High-Risk Neuroblastoma. Nature 2015, 526, 700–704. [Google Scholar] [CrossRef]
- Heaphy, C.M.; de Wilde, R.F.; Jiao, Y.; Klein, A.P.; Edil, B.H.; Shi, C.; Bettegowda, C.; Rodriguez, F.J.; Eberhart, C.G.; Hebbar, S.; et al. Altered Telomeres in Tumors with ATRX and DAXX Mutations. Science 2011, 333, 425–425. [Google Scholar] [CrossRef]
- Law, M.J.; Lower, K.M.; Voon, H.P.J.; Hughes, J.R.; Garrick, D.; Viprakasit, V.; Mitson, M.; De Gobbi, M.; Marra, M.; Morris, A.; et al. ATR-X Syndrome Protein Targets Tandem Repeats and Influences Allele-Specific Expression in a Size-Dependent Manner. Cell 2010, 143, 367–378. [Google Scholar] [CrossRef]
- Iwase, S.; Xiang, B.; Ghosh, S.; Ren, T.; Lewis, P.W.; Cochrane, J.C.; Allis, C.D.; Picketts, D.J.; Patel, D.J.; Li, H.; et al. ATRX ADD Domain Links an Atypical Histone Methylation Recognition Mechanism to Human Mental-Retardation Syndrome. Nat Struct Mol Biol 2011, 18, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.-Y.; Jones, D.T.W.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.-A.K.; Tönjes, M.; et al. Driver Mutations in Histone H3.3 and Chromatin Remodelling Genes in Paediatric Glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Mackay, A.; Burford, A.; Carvalho, D.; Izquierdo, E.; Fazal-Salom, J.; Taylor, K.R.; Bjerke, L.; Clarke, M.; Vinci, M.; Nandhabalan, M.; et al. Integrated Molecular Meta-Analysis of 1,000 Pediatric High-Grade and Diffuse Intrinsic Pontine Glioma. Cancer Cell 2017, 32, 520–537. [Google Scholar] [CrossRef]
- Mukherjee, J.; Johannessen, T.-C.; Ohba, S.; Chow, T.T.; Jones, L.; Pandita, A.; Pieper, R.O. Mutant IDH1 Cooperates with ATRX Loss to Drive the Alternative Lengthening of Telomere Phenotype in Glioma. Cancer Research 2018, 78, 2966–2977. [Google Scholar] [CrossRef]
- Sturm, D.; Witt, H.; Hovestadt, V.; Khuong-Quang, D.-A.; Jones, D.T.W.; Konermann, C.; Pfaff, E.; Tönjes, M.; Sill, M.; Bender, S.; et al. Hotspot Mutations in H3F3A and IDH1 Define Distinct Epigenetic and Biological Subgroups of Glioblastoma. Cancer Cell 2012, 22, 425–437. [Google Scholar] [CrossRef]
- Yeager, T.R.; Neumann, A.A.; Englezou, A.; Huschtscha, L.I.; Noble, J.R.; Reddel, R.R. Telomerase-Negative Immortalized Human Cells Contain a Novel Type of Promyelocytic Leukemia (PML) Body. Cancer Res 1999, 59, 4175–4179. [Google Scholar] [PubMed]
- Henson, J.D.; Cao, Y.; Huschtscha, L.I.; Chang, A.C.; Au, A.Y.M.; Pickett, H.A.; Reddel, R.R. DNA C-Circles Are Specific and Quantifiable Markers of Alternative-Lengthening-of-Telomeres Activity. Nat Biotechnol 2009, 27, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Grudic, A.; Jul-Larsen, Å.; Haring, S.J.; Wold, M.S.; Lønning, P.E.; Bjerkvig, R.; Bøe, S.O. Replication Protein A Prevents Accumulation of Single-Stranded Telomeric DNA in Cells That Use Alternative Lengthening of Telomeres. Nucleic Acids Research 2007, 35, 7267–7278. [Google Scholar] [CrossRef]
- Wu, G.; Jiang, X.; Lee, W.-H.; Chen, P.-L. Assembly of Functional ALT-Associated Promyelocytic Leukemia Bodies Requires Nijmegen Breakage Syndrome 1. Cancer Res 2003, 63, 2589–2595. [Google Scholar]
- Acharya, S.; Kaul, Z.; Gocha, A.S.; Martinez, A.R.; Harris, J.; Parvin, J.D.; Groden, J. Association of BLM and BRCA1 during Telomere Maintenance in ALT Cells. PLoS ONE 2014, 9, e103819. [Google Scholar] [CrossRef]
- Pan, X.; Drosopoulos, W.C.; Sethi, L.; Madireddy, A.; Schildkraut, C.L.; Zhang, D. FANCM, BRCA1, and BLM Cooperatively Resolve the Replication Stress at the ALT Telomeres. Proc. Natl. Acad. Sci. U.S.A. 2017, 114. [Google Scholar] [CrossRef]
- Barroso-González, J.; García-Expósito, L.; Hoang, S.M.; Lynskey, M.L.; Roncaioli, J.L.; Ghosh, A.; Wallace, C.T.; Modesti, M.; Bernstein, K.A.; Sarkar, S.N.; et al. RAD51AP1 Is an Essential Mediator of Alternative Lengthening of Telomeres. Molecular Cell 2019, 76, 217. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.K.; Cimprich, K.A. Causes and Consequences of Replication Stress. Nat Cell Biol 2014, 16, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Verma, P.; Cho, N.W.; Winters, H.D.; Wondisford, A.R.; Greenberg, R.A. Break-Induced Telomere Synthesis Underlies Alternative Telomere Maintenance. Nature 2016, 539, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.E.; Maréchal, A.; Flynn, R.L. SMARCAL1 Resolves Replication Stress at ALT Telomeres. Cell Reports 2016, 14, 1032–1040. [Google Scholar] [CrossRef]
- Silva, B.; Pentz, R.; Figueira, A.M.; Arora, R.; Lee, Y.W.; Hodson, C.; Wischnewski, H.; Deans, A.J.; Azzalin, C.M. FANCM Limits ALT Activity by Restricting Telomeric Replication Stress Induced by Deregulated BLM and R-Loops. Nat Commun 2019, 10, 2253. [Google Scholar] [CrossRef]
- Lu, R.; O’Rourke, J.J.; Sobinoff, A.P.; Allen, J.A.M.; Nelson, C.B.; Tomlinson, C.G.; Lee, M.; Reddel, R.R.; Deans, A.J.; Pickett, H.A. The FANCM-BLM-TOP3A-RMI Complex Suppresses Alternative Lengthening of Telomeres (ALT). Nat Commun 2019, 10, 2252. [Google Scholar] [CrossRef]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 Regulates TERRA-Telomeric DNA Hybrids and Telomere Maintenance in ALT Tumour Cells. Nat Commun 2014, 5, 5220. [Google Scholar] [CrossRef]
- Azzalin, C.M.; Lingner, J. Telomere Functions Grounding on TERRA Firma. Trends Cell Biol. 2015, 25, 29–36. [Google Scholar] [CrossRef]
- Root, H.; Larsen, A.; Komosa, M.; Al-Azri, F.; Li, R.; Bazett-Jones, D.P.; Stephen Meyn, M. FANCD2 Limits BLM-Dependent Telomere Instability in the Alternative Lengthening of Telomeres Pathway. Hum. Mol. Genet. 2016, 25, 3255–3268. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.W.; Dilley, R.L.; Lampson, M.A.; Greenberg, R.A. Interchromosomal Homology Searches Drive Directional ALT Telomere Movement and Synapsis. Cell 2014, 159, 108–121. [Google Scholar] [CrossRef]
- Potts, P.R.; Yu, H. The SMC5/6 Complex Maintains Telomere Length in ALT Cancer Cells through SUMOylation of Telomere-Binding Proteins. Nat Struct Mol Biol 2007, 14, 581–590. [Google Scholar] [CrossRef]
- Schrank, B.R.; Aparicio, T.; Li, Y.; Chang, W.; Chait, B.T.; Gundersen, G.G.; Gottesman, M.E.; Gautier, J. Nuclear ARP2/3 Drives DNA Break Clustering for Homology-Directed Repair. Nature 2018, 559, 61–66. [Google Scholar] [CrossRef]
- Garcia-Exposito, L.; Bournique, E.; Bergoglio, V.; Bose, A.; Barroso-Gonzalez, J.; Zhang, S.; Roncaioli, J.L.; Lee, M.; Wallace, C.T.; Watkins, S.C.; et al. Proteomic Profiling Reveals a Specific Role for Translesion DNA Polymerase η in the Alternative Lengthening of Telomeres. Cell Reports 2016, 17, 1858–1871. [Google Scholar] [CrossRef] [PubMed]
- Bizard, A.H.; Hickson, I.D. The Dissolution of Double Holliday Junctions. Cold Spring Harbor Perspectives in Biology 2014, 6, a016477–a016477. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Keirsey, J.; Russell, B.; Kavecansky, J.; Lillard-Wetherell, K.; Tahmaseb, K.; Turchi, J.J.; Groden, J. Telomerase-Associated Protein 1, HSP90, and Topoisomerase IIα Associate Directly with the BLM Helicase in Immortalized Cells Using ALT and Modulate Its Helicase Activity Using Telomeric DNA Substrates. Journal of Biological Chemistry 2009, 284, 14966–14977. [Google Scholar] [CrossRef] [PubMed]
- Sobinoff, A.P.; Allen, J.A.; Neumann, A.A.; Yang, S.F.; Walsh, M.E.; Henson, J.D.; Reddel, R.R.; Pickett, H.A. BLM and SLX4 Play Opposing Roles in Recombination-dependent Replication at Human Telomeres. The EMBO Journal 2017, 36, 2907–2919. [Google Scholar] [CrossRef] [PubMed]
- Hoang, S.M.; O’Sullivan, R.J. Alternative Lengthening of Telomeres: Building Bridges To Connect Chromosome Ends. Trends in Cancer 2020, 6, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Wright, W.E.; Shay, J.W. Clustered Telomeres in Phase-Separated Nuclear Condensates Engage Mitotic DNA Synthesis through BLM and RAD52. Genes Dev. 2019, 33, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Xue, C.; Kumar, S.; Crickard, J.B.; Yu, Y.; Wang, W.; Pham, N.; Li, Y.; Niu, H.; Sung, P.; et al. Rad52 Restrains Resection at DNA Double-Strand Break Ends in Yeast. Molecular Cell 2019, 76, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Malacaria, E.; Pugliese, G.M.; Honda, M.; Marabitti, V.; Aiello, F.A.; Spies, M.; Franchitto, A.; Pichierri, P. Rad52 Prevents Excessive Replication Fork Reversal and Protects from Nascent Strand Degradation. Nat Commun 2019, 10, 1412. [Google Scholar] [CrossRef]
- Verma, P.; Dilley, R.L.; Zhang, T.; Gyparaki, M.T.; Li, Y.; Greenberg, R.A. RAD52 and SLX4 Act Nonepistatically to Ensure Telomere Stability during Alternative Telomere Lengthening. Genes Dev. 2019, 33, 221–235. [Google Scholar] [CrossRef]
- West, S.C.; Blanco, M.G.; Chan, Y.W.; Matos, J.; Sarbajna, S.; Wyatt, H.D.M. Resolution of Recombination Intermediates: Mechanisms and Regulation. Cold Spring Harb Symp Quant Biol 2015, 80, 103–109. [Google Scholar] [CrossRef]
- Bhowmick, R.; Minocherhomji, S.; Hickson, I.D. RAD52 Facilitates Mitotic DNA Synthesis Following Replication Stress. Molecular Cell 2016, 64, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, S.K.; Kamileri, I.; Lugli, N.; Evangelou, K.; Da-Ré, C.; Huber, F.; Padayachy, L.; Tardy, S.; Nicati, N.L.; Barriot, S.; et al. Mammalian RAD52 Functions in Break-Induced Replication Repair of Collapsed DNA Replication Forks. Molecular Cell 2016, 64, 1127–1134. [Google Scholar] [CrossRef]
- Özer, Ö.; Bhowmick, R.; Liu, Y.; Hickson, I.D. Human Cancer Cells Utilize Mitotic DNA Synthesis to Resist Replication Stress at Telomeres Regardless of Their Telomere Maintenance Mechanism. Oncotarget 2018, 9, 15836–15846. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.T.; Cesare, A.J.; Rivera, T.; Karlseder, J. Cell Death during Crisis Is Mediated by Mitotic Telomere Deprotection. Nature 2015, 522, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Davoli, T.; Denchi, E.L.; de Lange, T. Persistent Telomere Damage Induces Bypass of Mitosis and Tetraploidy. Cell 2010, 141, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Maciejowski, J.; Li, Y.; Bosco, N.; Campbell, P.J.; de Lange, T. Chromothripsis and Kataegis Induced by Telomere Crisis. Cell 2015, 163, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Nassour, J.; Radford, R.; Correia, A.; Fusté, J.M.; Schoell, B.; Jauch, A.; Shaw, R.J.; Karlseder, J. Autophagic Cell Death Restricts Chromosomal Instability during Replicative Crisis. Nature 2019, 565, 659–663. [Google Scholar] [CrossRef]
- Bayne, S.; Liu, J.-P. Hormones and Growth Factors Regulate Telomerase Activity in Ageing and Cancer. Molecular and Cellular Endocrinology 2005, 240, 11–22. [Google Scholar] [CrossRef]
- Patrick, M.S.; Cheng, N.-L.; Kim, J.; An, J.; Dong, F.; Yang, Q.; Zou, I.; Weng, N. Human T Cell Differentiation Negatively Regulates Telomerase Expression Resulting in Reduced Activation-Induced Proliferation and Survival. Front. Immunol. 2019, 10, 1993. [Google Scholar] [CrossRef]
- Burgess, R.J.; Agathocleous, M.; Morrison, S.J. Metabolic Regulation of Stem Cell Function. J Intern Med 2014, 276, 12–24. [Google Scholar] [CrossRef]
- Almeida, L.; Lochner, M.; Berod, L.; Sparwasser, T. Metabolic Pathways in T Cell Activation and Lineage Differentiation. Seminars in Immunology 2016, 28, 514–524. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Miyazawa, H.; Aulehla, A. Revisiting the Role of Metabolism during Development. Development 2018, 145, dev131110. [Google Scholar] [CrossRef]
- Bao, Y.; Mukai, K.; Hishiki, T.; Kubo, A.; Ohmura, M.; Sugiura, Y.; Matsuura, T.; Nagahata, Y.; Hayakawa, N.; Yamamoto, T.; et al. Energy Management by Enhanced Glycolysis in G1-Phase in Human Colon Cancer Cells In Vitro and In Vivo. Molecular Cancer Research 2013, 11, 973–985. [Google Scholar] [CrossRef]
- Mitra, K.; Wunder, C.; Roysam, B.; Lin, G.; Lippincott-Schwartz, J. A Hyperfused Mitochondrial State Achieved at G 1 –S Regulates Cyclin E Buildup and Entry into S Phase. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 11960–11965. [Google Scholar] [CrossRef]
- Tudzarova, S.; Colombo, S.L.; Stoeber, K.; Carcamo, S.; Williams, G.H.; Moncada, S. Two Ubiquitin Ligases, APC/C-Cdh1 and SKP1-CUL1-F (SCF)-β-TrCP, Sequentially Regulate Glycolysis during the Cell Cycle. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 5278–5283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fan, M.; Candas, D.; Zhang, T.-Q.; Qin, L.; Eldridge, A.; Wachsmann-Hogiu, S.; Ahmed, K.M.; Chromy, B.A.; Nantajit, D.; et al. Cyclin B1/Cdk1 Coordinates Mitochondrial Respiration for Cell-Cycle G2/M Progression. Developmental Cell 2014, 29, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Sutter, B.M.; Li, B.; Tu, B.P. Acetyl-CoA Induces Cell Growth and Proliferation by Promoting the Acetylation of Histones at Growth Genes. Molecular Cell 2011, 42, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Intlekofer, A.M.; Dematteo, R.G.; Venneti, S.; Finley, L.W.S.; Lu, C.; Judkins, A.R.; Rustenburg, A.S.; Grinaway, P.B.; Chodera, J.D.; Cross, J.R.; et al. Hypoxia Induces Production of L-2-Hydroxyglutarate. Cell Metabolism 2015, 22, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, S.; Hau, M. Telomere Attrition: Metabolic Regulation and Signalling Function? Biol. Lett. 2019, 15, 20180885. [Google Scholar] [CrossRef]
- Zheng, J. Energy Metabolism of Cancer: Glycolysis versus Oxidative Phosphorylation (Review). Oncology Letters 2012, 4, 1151–1157. [Google Scholar] [CrossRef]
- Fice, H.; Robaire, B. Telomere Dynamics Throughout Spermatogenesis. Genes 2019, 10, 525. [Google Scholar] [CrossRef] [PubMed]
- Thilagavathi, J.; Mishra, S.S.; Kumar, M.; Vemprala, K.; Deka, D.; Dhadwal, V.; Dada, R. Analysis of Telomere Length in Couples Experiencing Idiopathic Recurrent Pregnancy Loss. J Assist Reprod Genet 2013, 30, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, F.; Dai, S.; Zhang, N.; Zhao, W.; Bai, R.; Sun, Y. Sperm Telomere Length Is Positively Associated with the Quality of Early Embryonic Development. Hum. Reprod. 2015, 30, 1876–1881. [Google Scholar] [CrossRef]
- Anifandis, G.; Samara, M.; Simopoulou, M.; Messini, C.I.; Chatzimeletiou, K.; Thodou, E.; Daponte, A.; Georgiou, I. Insights into the Role of Telomeres in Human Embryological Parameters. Opinions Regarding IVF. JDB 2021, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Berneau, S.C.; Shackleton, J.; Nevin, C.; Altakroni, B.; Papadopoulos, G.; Horne, G.; Brison, D.R.; Murgatroyd, C.; Povey, A.C.; Carroll, M. Associations of Sperm Telomere Length with Semen Parameters, Clinical Outcomes and Lifestyle Factors in Human Normozoospermic Samples. Andrology 2020, 8, 583–593. [Google Scholar] [CrossRef]
- Beygi, Z.; Forouhari, S.; Mahmoudi, E.; Hayat, S.M.G.; Nourimand, F. Role of Oxidative Stress and Antioxidant Supplementation in Male Fertility. CMM 2021, 21, 265–282. [Google Scholar] [CrossRef]
- Ahmed, W.; Lingner, J. PRDX1 and MTH1 Cooperate to Prevent ROS-Mediated Inhibition of Telomerase. Genes Dev. 2018, 32, 658–669. [Google Scholar] [CrossRef]
- Antunes, D.M.F.; Kalmbach, K.H.; Wang, F.; Dracxler, R.C.; Seth-Smith, M.L.; Kramer, Y.; Buldo-Licciardi, J.; Kohlrausch, F.B.; Keefe, D.L. A Single-Cell Assay for Telomere DNA Content Shows Increasing Telomere Length Heterogeneity, as Well as Increasing Mean Telomere Length in Human Spermatozoa with Advancing Age. J Assist Reprod Genet 2015, 32, 1685–1690. [Google Scholar] [CrossRef]
- De Frutos, C.; López-Cardona, A.P.; Fonseca Balvís, N.; Laguna-Barraza, R.; Rizos, D.; Gutierrez-Adán, A.; Bermejo-Álvarez, P. Spermatozoa Telomeres Determine Telomere Length in Early Embryos and Offspring. REPRODUCTION 2016, 151, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Tan, Y.; Qiu, X.; Luo, H.; Li, Y.; Li, R.; Yang, X. Sperm Telomere Length as a Novel Biomarker of Male Infertility and Embryonic Development: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2023, 13, 1079966. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Bailey, S.M.; Okuka, M.; Muñoz, P.; Li, C.; Zhou, L.; Wu, C.; Czerwiec, E.; Sandler, L.; Seyfang, A.; et al. Telomere Lengthening Early in Development. Nat Cell Biol 2007, 9, 1436–1441. [Google Scholar] [CrossRef]
- Bender, H.S.; Murchison, E.P.; Pickett, H.A.; Deakin, J.E.; Strong, M.A.; Conlan, C.; McMillan, D.A.; Neumann, A.A.; Greider, C.W.; Hannon, G.J.; et al. Extreme Telomere Length Dimorphism in the Tasmanian Devil and Related Marsupials Suggests Parental Control of Telomere Length. PLoS ONE 2012, 7, e46195. [Google Scholar] [CrossRef]
- Eisenhauer, K.M.; Gerstein, R.M.; Chiu, C.-P.; Conti, M.; Hsueh, A.J.W. Telomerase Activity in Female and Male Rat Germ Cells Undergoing Meiosis and in Early Embryos1. Biology of Reproduction 1997, 56, 1120–1125. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and Telomerase: Implications for Cancer and Aging. Radiation Research 2001, 155, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Blasco, M.A.; Trimarchi, J.R.; Keefe, D.L. An Essential Role for Functional Telomeres in Mouse Germ Cells during Fertilization and Early Development. Developmental Biology 2002, 249, 74–84. [Google Scholar] [CrossRef]
- Siderakis, M.; Tarsounas, M. Telomere Regulation and Function during Meiosis. Chromosome Res 2007, 15, 667–679. [Google Scholar] [CrossRef]
- Keefe, D.L.; Franco, S.; Liu, L.; Trimarchi, J.; Cao, B.; Weitzen, S.; Agarwal, S.; Blasco, M.A. Telomere Length Predicts Embryo Fragmentation after in Vitro Fertilization in Women—Toward a Telomere Theory of Reproductive Aging in Women. American Journal of Obstetrics and Gynecology 2005, 192, 1256–1260. [Google Scholar] [CrossRef]
- Liu, L.; Franco, S.; Spyropoulos, B.; Moens, P.B.; Blasco, M.A.; Keefe, D.L. Irregular Telomeres Impair Meiotic Synapsis and Recombination in Mice. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 6496–6501. [Google Scholar] [CrossRef]
- Treff, N.R.; Su, J.; Taylor, D.; Scott, R.T. Telomere DNA Deficiency Is Associated with Development of Human Embryonic Aneuploidy. PLoS Genet 2011, 7, e1002161. [Google Scholar] [CrossRef]
- Keefe, D.L. Telomeres and Genomic Instability during Early Development. European Journal of Medical Genetics 2020, 63, 103638. [Google Scholar] [CrossRef] [PubMed]
- Yamada-Fukunaga, T.; Yamada, M.; Hamatani, T.; Chikazawa, N.; Ogawa, S.; Akutsu, H.; Miura, T.; Miyado, K.; Tarín, J.J.; Kuji, N.; et al. Age-Associated Telomere Shortening in Mouse Oocytes. Reprod Biol Endocrinol 2013, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, M.; Ye, X.; Liu, K.; Huang, J.; Wang, L.; Ji, G.; Liu, N.; Tang, X.; Baltz, J.M.; et al. Delay in Oocyte Aging in Mice by the Antioxidant N-Acetyl-l-Cysteine (NAC). Human Reproduction 2012, 27, 1411–1420. [Google Scholar] [CrossRef]
- Kinugawa, C.; Murakami, T.; Okamura, K.; Yajima, A. Telomerase Activity in Normal Ovaries and Premature Ovarian Failure. Tohoku J. Exp. Med. 2000, 190, 231–238. [Google Scholar] [CrossRef]
- Lavranos, T.C.; Mathis, J.M.; Latham, S.E.; Kalionis, B.; Shay, J.W.; Rodgers, R.J. Evidence for Ovarian Granulosa Stem Cells: Telomerase Activity and Localization of the Telomerase Ribonucleic Acid Component in Bovine Ovarian Follicles1. Biology of Reproduction 1999, 61, 358–366. [Google Scholar] [CrossRef]
- Russo, V.; Berardinelli, P.; Martelli, A.; Giacinto, O.D.; Nardinocchi, D.; Fantasia, D.; Barboni, B. Expression of Telomerase Reverse Transcriptase Subunit (TERT) and Telomere Sizing in Pig Ovarian Follicles. J Histochem Cytochem. 2006, 54, 443–455. [Google Scholar] [CrossRef]
- Yamagata, Y.; Nakamura, Y.; Umayahara, K.; Harada, A.; Takayama, H.; Sugino, N.; Kato, H. Changes in Telomerase Activity in Experimentally Induced Atretic Follicles of Immature Rats. Endocr J 2002, 49, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Bayne, S.; Li, H.; Jones, M.E.E.; Pinto, A.R.; Van Sinderen, M.; Drummond, A.; Simpson, E.R.; Liu, J.-P. Estrogen Deficiency Reversibly Induces Telomere Shortening in Mouse Granulosa Cells and Ovarian Aging in Vivo. Protein Cell 2011, 2, 333–346. [Google Scholar] [CrossRef]
- Goto, H.; Iwata, H.; Takeo, S.; Nisinosono, K.; Murakami, S.; Monji, Y.; Kuwayama, T. Effect of Bovine Age on the Proliferative Activity, Global DNA Methylation, Relative Telomere Length and Telomerase Activity of Granulosa Cells. Zygote 2013, 21, 256–264. [Google Scholar] [CrossRef]
- Endo, M.; Kimura, K.; Kuwayama, T.; Monji, Y.; Iwata, H. Effect of Estradiol during Culture of Bovine Oocyte–Granulosa Cell Complexes on the Mitochondrial DNA Copies of Oocytes and Telomere Length of Granulosa Cells. Zygote 2014, 22, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-P.; Li, H. Telomerase in the Ovary. REPRODUCTION 2010, 140, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Butts, S.; Riethman, H.; Ratcliffe, S.; Shaunik, A.; Coutifaris, C.; Barnhart, K. Correlation of Telomere Length and Telomerase Activity with Occult Ovarian Insufficiency. The Journal of Clinical Endocrinology & Metabolism 2009, 94, 4835–4843. [Google Scholar] [CrossRef]
- Yung, Y.; Maydan, S.A.; Bart, Y.; Orvieto, R.; Aizer, A. Human Granulosa Cells of Poor Ovarian Responder Patients Display Telomeres Shortening. J Assist Reprod Genet 2023, 40, 1943–1947. [Google Scholar] [CrossRef]
- Cheng, G.; Kong, F.; Luan, Y.; Sun, C.; Wang, J.; Zhang, L.; Jiang, B.; Qi, T.; Zhao, J.; Zheng, C.; et al. Differential Shortening Rate of Telomere Length in the Development of Human Fetus. Biochemical and Biophysical Research Communications 2013, 442, 112–115. [Google Scholar] [CrossRef]
- Ozturk, S.; Sozen, B.; Demir, N. Telomere Length and Telomerase Activity during Oocyte Maturation and Early Embryo Development in Mammalian Species. Molecular Human Reproduction 2014, 20, 15–30. [Google Scholar] [CrossRef]
- Turner, S.; Wong, H.P.; Rai, J.; Hartshorne, G.M. Telomere Lengths in Human Oocytes, Cleavage Stage Embryos and Blastocysts. Molecular Human Reproduction 2010, 16, 685–694. [Google Scholar] [CrossRef]
- Le, R.; Huang, Y.; Zhang, Y.; Wang, H.; Lin, J.; Dong, Y.; Li, Z.; Guo, M.; Kou, X.; Zhao, Y.; et al. Dcaf11 Activates Zscan4-Mediated Alternative Telomere Lengthening in Early Embryos and Embryonic Stem Cells. Cell Stem Cell 2021, 28, 732–747. [Google Scholar] [CrossRef]
- Kordowitzki, P.; López De Silanes, I.; Guío-Carrión, A.; Blasco, M.A. Dynamics of Telomeric Repeat-Containing RNA Expression in Early Embryonic Cleavage Stages with Regards to Maternal Age. Aging 2020, 12, 15906–15917. [Google Scholar] [CrossRef]
- Nagaraj, R.; Sharpley, M.S.; Chi, F.; Braas, D.; Zhou, Y.; Kim, R.; Clark, A.T.; Banerjee, U. Nuclear Localization of Mitochondrial TCA Cycle Enzymes as a Critical Step in Mammalian Zygotic Genome Activation. Cell 2017, 168, 210–223. [Google Scholar] [CrossRef]
- Varela, E.; Schneider, R.P.; Ortega, S.; Blasco, M.A. Different Telomere-Length Dynamics at the Inner Cell Mass versus Established Embryonic Stem (ES) Cells. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 15207–15212. [Google Scholar] [CrossRef]
- Schaetzlein, S.; Lucas-Hahn, A.; Lemme, E.; Kues, W.A.; Dorsch, M.; Manns, M.P.; Niemann, H.; Rudolph, K.L. Telomere Length Is Reset during Early Mammalian Embryogenesis. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 8034–8038. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Kues, W.A.; Baulain, U.; Garrels, W.; Herrmann, D.; Niemann, H. Species-Specific Telomere Length Differences Between Blastocyst Cell Compartments and Ectopic Telomere Extension in Early Bovine Embryos by Human Telomerase Reverse Transcriptase. Biology of Reproduction 2011, 84, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gu, Y.; Zhou, J.; Zang, J.; Ling, X.; Li, H.; Hu, L.; Xu, B.; Zhang, B.; Qin, N.; et al. Leukocyte Telomere Length in Children Born Following Blastocyst-Stage Embryo Transfer. Nat Med 2022, 28, 2646–2653. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Li, C.; Sun, H.; Xin, Y.; Zhu, S.; Liu, Y.; Tang, S.; Han, L.; Huang, Z.; Wang, Q. Telomere Dysfunction in Oocytes and Embryos From Obese Mice. Front. Cell Dev. Biol. 2021, 9, 617225. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, K.; Liu, Y.; Fu, Y.; Gao, S.; Gong, P.; Wang, H.; Zhou, Z.; Zeng, M.; Wu, Z.; et al. Telomere Heterogeneity Linked to Metabolism and Pluripotency State Revealed by Simultaneous Analysis of Telomere Length and RNA-Seq in the Same Human Embryonic Stem Cell. BMC Biol 2017, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Tsogtbaatar, E.; Landin, C.; Minter-Dykhouse, K.; Folmes, C.D.L. Energy Metabolism Regulates Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Weng, N.; Palmer, L.D.; Levine, B.L.; Lane, H.C.; June, C.H.; Hodes, R.J. Tales of Tails: Regulation of Telomere Length and Telomerase Activity during Lymphocyte Development, Differentiation, Activation, and Aging. Immunol Rev 1997, 160, 43–54. [Google Scholar] [CrossRef]
- Quinn, K.M.; Palchaudhuri, R.; Palmer, C.S.; La Gruta, N.L. The Clock Is Ticking: The Impact of Ageing on T Cell Metabolism. Clin & Trans Imm 2019, 8, e01091. [Google Scholar] [CrossRef]
- Buck, M.D.; O’Sullivan, D.; Klein Geltink, R.I.; Curtis, J.D.; Chang, C.-H.; Sanin, D.E.; Qiu, J.; Kretz, O.; Braas, D.; van der Windt, G.J.W.; et al. Mitochondrial Dynamics Controls T Cell Fate through Metabolic Programming. Cell 2016, 166, 63–76. [Google Scholar] [CrossRef]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted Role of mTOR (Mammalian Target of Rapamycin) Signaling Pathway in Human Health and Disease. Sig Transduct Target Ther 2023, 8, 375. [Google Scholar] [CrossRef]
- Shliapina, V.L.; Yurtaeva, S.V.; Rubtsova, M.P.; Dontsova, O.A. At the Crossroads: Mechanisms of Apoptosis and Autophagy in Cell Life and Death. Acta Naturae 2021, 13, 106–115. [Google Scholar] [CrossRef]
- González, A.; Hall, M.N.; Lin, S.-C.; Hardie, D.G. AMPK and TOR: The Yin and Yang of Cellular Nutrient Sensing and Growth Control. Cell Metabolism 2020, 31, 472–492. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A Nutrient and Energy Sensor That Maintains Energy Homeostasis. Nat Rev Mol Cell Biol 2012, 13, 251–262. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat Rev Mol Cell Biol 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, M.; Naraykina, Y.; Vasilkova, D.; Meerson, M.; Zvereva, M.; Prassolov, V.; Lazarev, V.; Manuvera, V.; Kovalchuk, S.; Anikanov, N.; et al. Protein Encoded in Human Telomerase RNA Is Involved in Cell Protective Pathways. Nucleic Acids Research 2018. [Google Scholar] [CrossRef] [PubMed]
- Shliapina, V.; Koriagina, M.; Vasilkova, D.; Govorun, V.; Dontsova, O.; Rubtsova, M. Human Telomerase RNA Protein Encoded by Telomerase RNA Is Involved in Metabolic Responses. Front. Cell Dev. Biol. 2021, 9, 754611. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, M.; Dontsova, O. How Structural Features Define Biogenesis and Function of Human Telomerase RNA Primary Transcript. Biomedicines 2022, 10, 1650. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Human telomeric chromatin structure. Telomeric chromatin is organized by shelterin complex composed by TRF1, TRF2, TIN2, TPP1, POT1 and RAP1. To prevent the degradation and reparation of chromosome ends T-loop and D-loop protect telomeres. G-rich 3’-overhangs form G-quadruplex structures which stimulate the telomerase to synthesize the telomeric repeats and prevent the synthesis of C-rich strand by CST complex. Transcription of subtelomeric regions produces TERRA RNA which promotes R-loop formation stabilized by G-quadruplex structures at the opposite strand.
Figure 1.
Human telomeric chromatin structure. Telomeric chromatin is organized by shelterin complex composed by TRF1, TRF2, TIN2, TPP1, POT1 and RAP1. To prevent the degradation and reparation of chromosome ends T-loop and D-loop protect telomeres. G-rich 3’-overhangs form G-quadruplex structures which stimulate the telomerase to synthesize the telomeric repeats and prevent the synthesis of C-rich strand by CST complex. Transcription of subtelomeric regions produces TERRA RNA which promotes R-loop formation stabilized by G-quadruplex structures at the opposite strand.
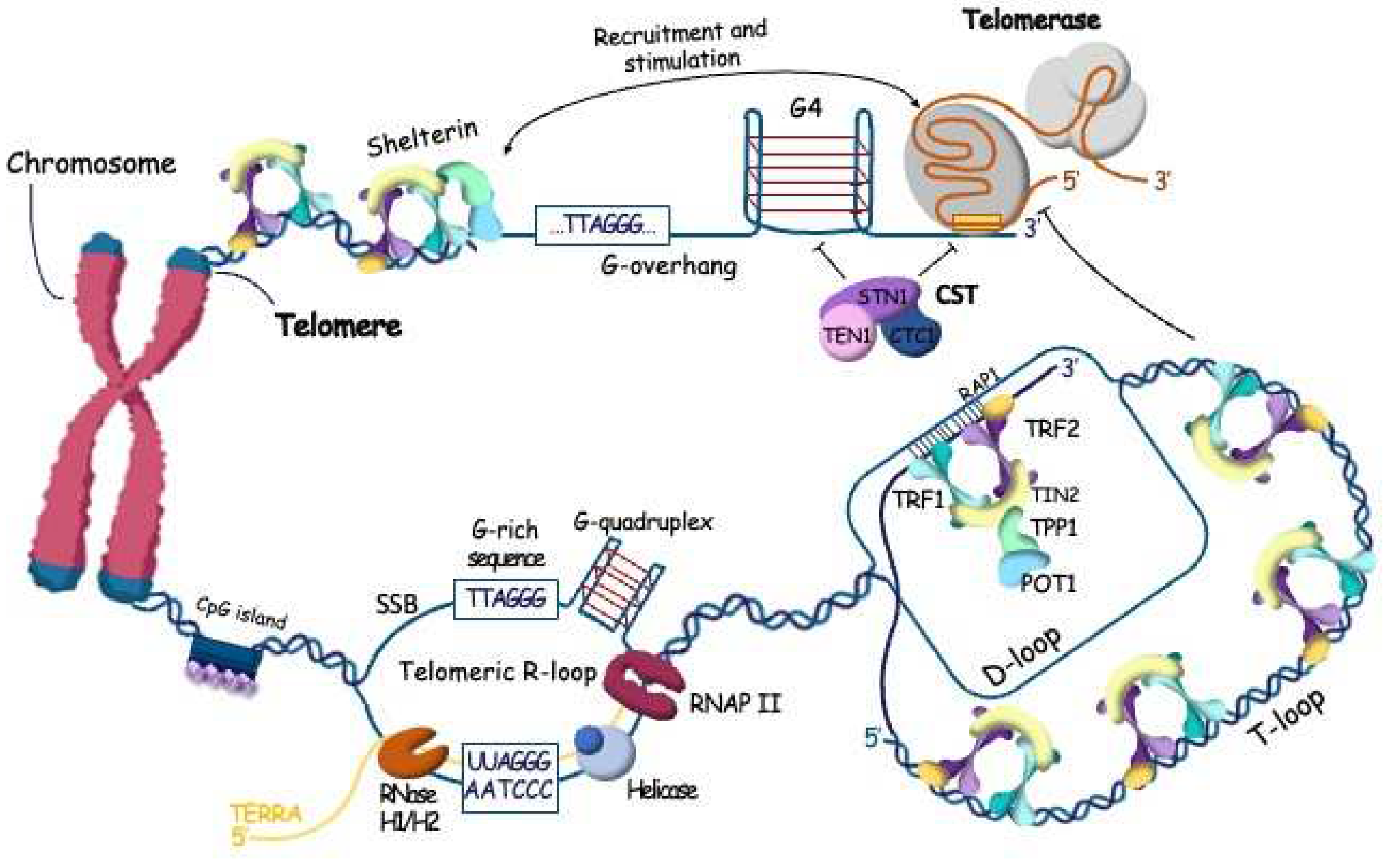
Figure 2.
Scheme of telomerase action at telomeres. Telomeric chromatin is organized in closed state. Subtelomere DNA is methylated at CpG dinucleotides. Histones are characterized with heterochromatin modifications (H3K9me3 and H4K20me3). ATRX/DAXX complex stimulates the accumulation of H3.3 nucleosomes and prevents the homological recombination of telomeric regions. Shelterin complex through TPP1 loads telomerase to the very end of telomere and stimulates telomeric repeats synthesis at processive manner when enzyme translocates along the telomere in order to add more telomeric repeats after single repeat synthesis.
Figure 2.
Scheme of telomerase action at telomeres. Telomeric chromatin is organized in closed state. Subtelomere DNA is methylated at CpG dinucleotides. Histones are characterized with heterochromatin modifications (H3K9me3 and H4K20me3). ATRX/DAXX complex stimulates the accumulation of H3.3 nucleosomes and prevents the homological recombination of telomeric regions. Shelterin complex through TPP1 loads telomerase to the very end of telomere and stimulates telomeric repeats synthesis at processive manner when enzyme translocates along the telomere in order to add more telomeric repeats after single repeat synthesis.
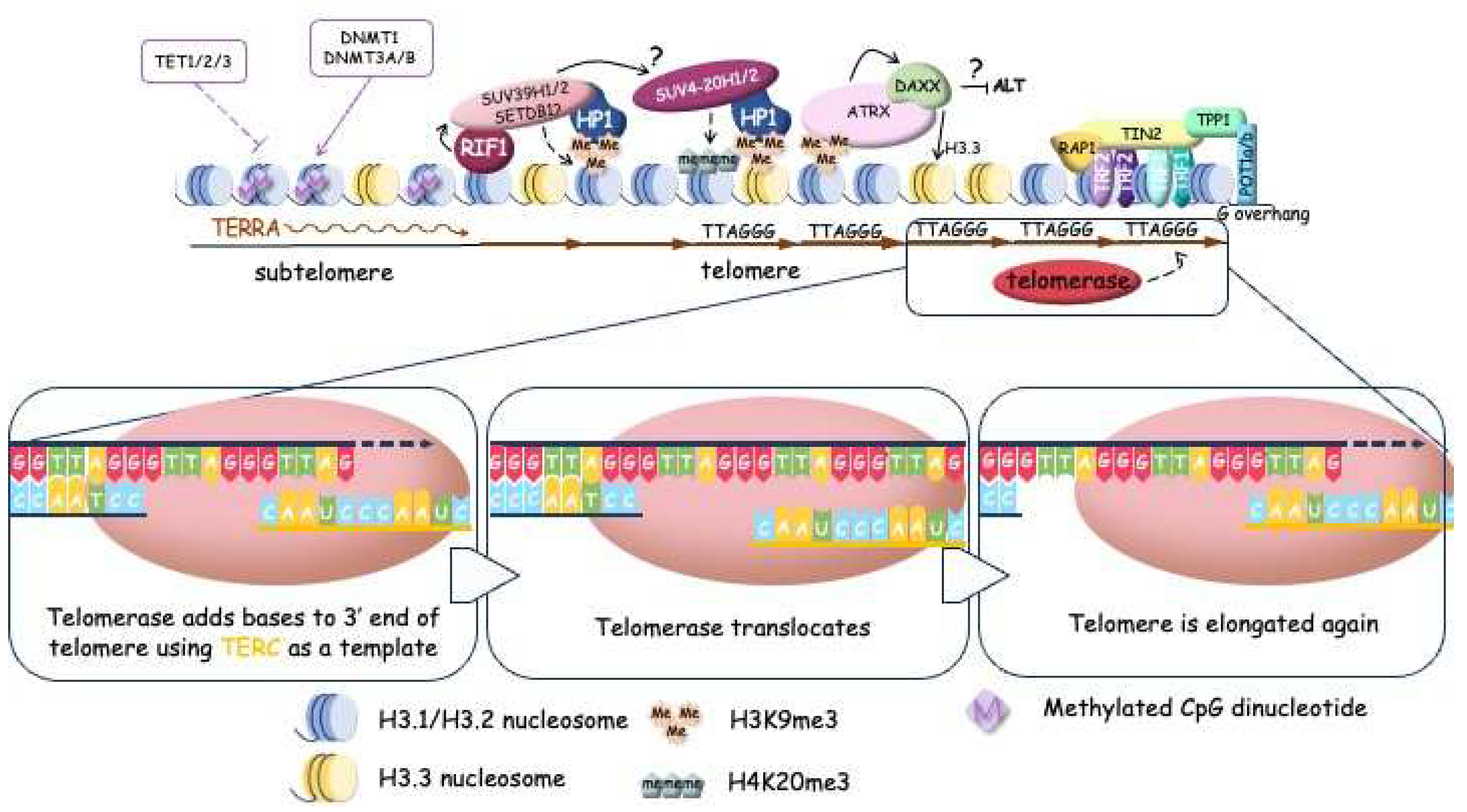
Figure 3.
ALTernative mechanism of telomere maintenance. Absence of ATRX at telomeres promotes recombination at telomeric chromatin. Replication fork stalling, DNA-damage and alternative structures of telomercic DNA (G-quadruplexes and R-loops) at telomeres also involved in activation of ALT.
Figure 3.
ALTernative mechanism of telomere maintenance. Absence of ATRX at telomeres promotes recombination at telomeric chromatin. Replication fork stalling, DNA-damage and alternative structures of telomercic DNA (G-quadruplexes and R-loops) at telomeres also involved in activation of ALT.
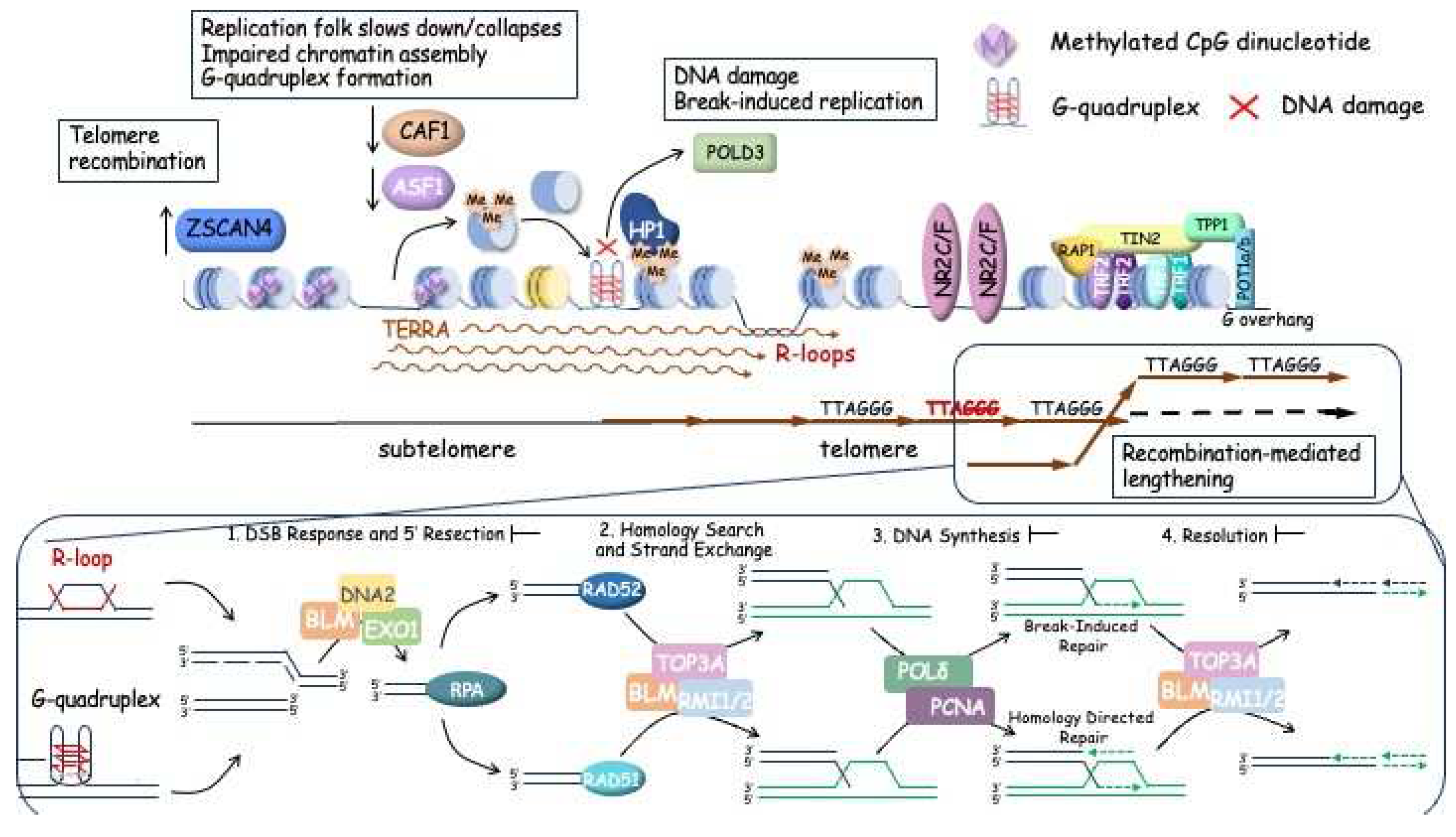
Figure 4.
An overview of glucose metabolism. Glucose is catabolized in a series of enzymatic reactions yielding pyruvate. Pyruvate is converted in lactate or transported in mitochondria and metabolized into acetyl-CoA, fueling TCA cycle. FBP-fructose biphosphate, 1,3-BPG-1,3-biphosphoglycerate, 3PG-3-phosphoglycerate, PEP-phosphoenolpyruvate. Glycolysis also fuels pentose phosphate pathway, which produces nucleotides, aminoacids and fatty acids.
Figure 4.
An overview of glucose metabolism. Glucose is catabolized in a series of enzymatic reactions yielding pyruvate. Pyruvate is converted in lactate or transported in mitochondria and metabolized into acetyl-CoA, fueling TCA cycle. FBP-fructose biphosphate, 1,3-BPG-1,3-biphosphoglycerate, 3PG-3-phosphoglycerate, PEP-phosphoenolpyruvate. Glycolysis also fuels pentose phosphate pathway, which produces nucleotides, aminoacids and fatty acids.
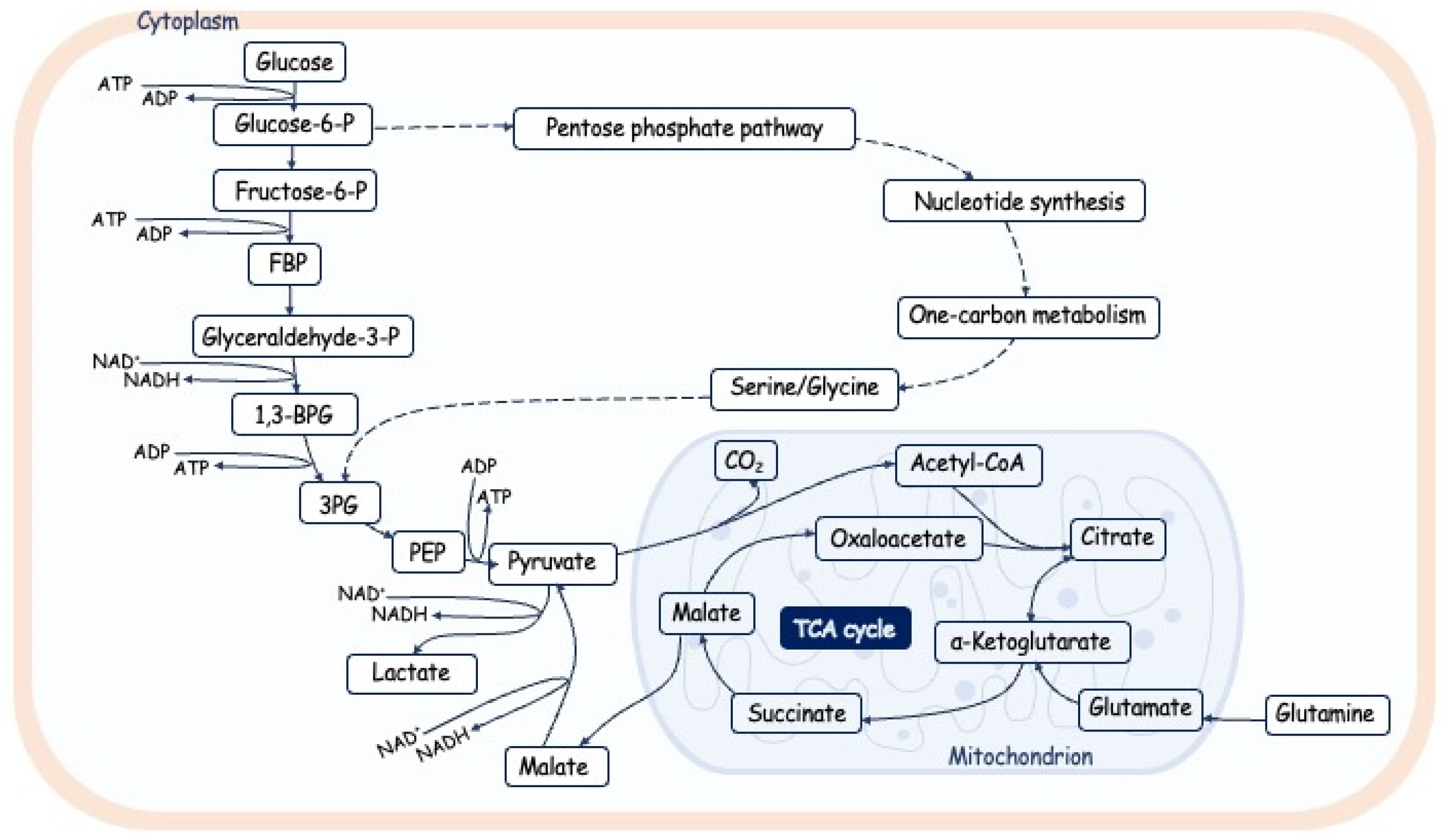
Figure 5.
Telomere dynamics and metabolic program preferencies during mouse embryo preimplantation development. At first stage after fertilization extensive chromatin remodeling occurs with a histone-to-protamine replacement, formation of the two parental pronuclei and major zygotic genome activation (ZGA) at the two-cell stage, accompanied with elongation of telomeres by a recombination-mediated mechanism that is telomerase-independent and with activation of OxPhos metabolism program. Then, at the stage from the morula-blastocyst transition telomerase is activated. This process is characterized by switching metabolism program from OxPhos to glycolysis. In mouse zygotes, at the morula stage, ATRX is clearly targeted to telomeres. The expression of H3K9 lysine demethylases from the Kdm4 family and Zscan4 favors telomere elongation by recombination in mouse and human zygotes.
Figure 5.
Telomere dynamics and metabolic program preferencies during mouse embryo preimplantation development. At first stage after fertilization extensive chromatin remodeling occurs with a histone-to-protamine replacement, formation of the two parental pronuclei and major zygotic genome activation (ZGA) at the two-cell stage, accompanied with elongation of telomeres by a recombination-mediated mechanism that is telomerase-independent and with activation of OxPhos metabolism program. Then, at the stage from the morula-blastocyst transition telomerase is activated. This process is characterized by switching metabolism program from OxPhos to glycolysis. In mouse zygotes, at the morula stage, ATRX is clearly targeted to telomeres. The expression of H3K9 lysine demethylases from the Kdm4 family and Zscan4 favors telomere elongation by recombination in mouse and human zygotes.
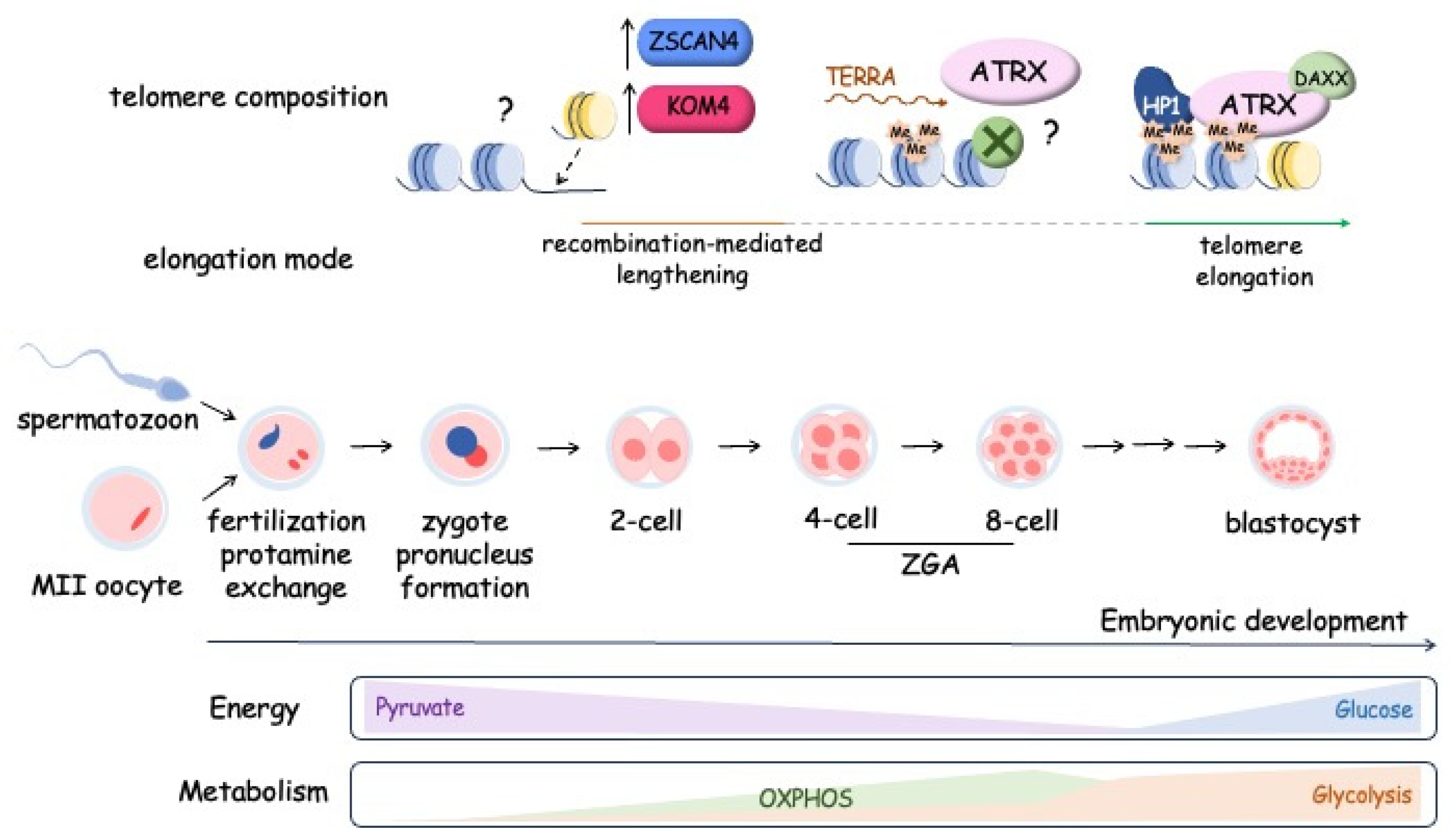
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



