
Submitted:
02 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
Human adenovirus 36 (HAdV-D36) is the only currently known infectious agent capable of promoting obesity in humans and animals, but the effects on the embryonated chicken egg (ECE) have not been described yet. In our study, we used ECEs as a model of a dynamically developing organism. The allantoic cavity and the yolk sack of ECEs was inoculated with HAdV-D36 to simulate the different routes of infection. Each day, a part of the embryos was weighed and histologically examined and compared to the controls to analyze pathological changes induced by the virus. Our study confirmed a significant increase in the ECEs body weight; however, this process was not caused by adipose tissue development, but increased cell proliferation of all tissues. We suggest that HAdV-D36 must affect the developing organisms via another mechanism inducing enhanced the ECEs growth.
Keywords:
embryonated chicken eggs
; human adenovirus 36
; organogenesis
1. Introduction
The virus-associated obesity has gain interest in scientific world in recent years. The viral infection and obesity, two seemingly unrelated events, come together in case of infectious obesity. The term includes an increase in body weight associated with the presence of an infectious agent, e.g., virus or bacteria [1]. Pasarica et al. indicate four types of viruses whose presence in the host organism may cause obesity: canine distemper virus (CDV), Rous-associated virus number 7 (RAV-7), Borna disease virus (BDV), and adenoviruses, mainly avian adenovirus (SMAM-1) and human adenovirus 36 (HAdV-D36) [2]. However, only human adenovirus 36 has been confirmed to infect both human and animals [3].
Human adenovirus 36 (HAdV-D36) belongs to the Adenoviridae family in group D. It is a non-enveloped virus with stable, highly conservative genome consisting of double stranded DNA (dsDNA), with only one mutation reported in a 14-year period [4]. This virus causes mainly the respiratory and the digestive system diseases [5]. Research on adenoviruses associated with obesity has been conducted since 1990, when SMAM-1 pro-obesity effects were first discovered in birds [6]. Further research of the Adenoviridae family, with particular emphasis on human adenovirus 36, showed that this virus significantly reduces the amount of triglycerides and cholesterol in the blood and has simultaneous adipogenic effect. The virus affects the differentiation of 3T3-L1 preadipocytes into adipocytes [7]. Interestingly, insulin plays important, inductive role in adipocytogenesis [8]. Rats infected with HAdV-D36 present weight gain, greater insulin sensitivity, and increased glucose uptake. The studies on rats, adult chickens, and monkeys showed a 15-30% increase in body weight after HAdV-D36 infection [9]. Moreover, HAdV-D36 was easily transmitted between infected and uninfected animals via droplets. Adenovirus 36 causes obesity by simultaneously activating several physiological processes: activation of pre-adipocyte differentiation into adipocytes, increase in glucose uptake, decrease in leptin and norepinephrine levels, and increase in MCP-1 level [3]. The worldwide studies on various populations show the presence of adenovirus 36 in the majority of the studied subjects. Almgren et al., researching the Swedish population, noted the presence of HAdV-D36 in more than half of the patients and the increase in virus prevalence from7% in 1992-1998 to 15-20% in 2002-2006 [10]. Zhou et al., studying 824 patients in China, estimated the overall seroprevalence of human adenovirus 36 at 49.8% [11]. Similar studies conducted worldwide present similar results indicating the increasing role of HAdV-D36 in the incidence of infectious obesity.
So far, the mechanisms of action have been studied and described only in adult patients or adult model organisms. Since the metabolism of embryos is very distinct from adult organisms, more focused towards growth and differentiation, it may respond differently to virus presence. Viruses, being similir to insulin, may possibly act as glucose uptake inductors or growth factors [8,9]. Viruses are also associated with teratogenic effects on the developing embryo or fetus. Viruses like rubellavirus, cytomegalovirus, herpesviruses, parvoviruses, and viruses causing hepatitis are well known for their harmful effects on embryonic development, hence screening the pregnant women and animals for them became a standard [12]. Since, in case of HAdV-D36 no data on its influence on embryos exist and the virus affects the adults via complex mechanism, we believe that studies on developing organisms such as embryonated chicken eggs (ECEs) can deliver a lot of new knowledge applicable in both veterinary and human medicine
2. Results
2.1. ECEs weight changes from the 6th do 20th day of development
Every day from the 6th to the 20th day of the incubation period, 28 ECEs from a total of 420 were examined. All ECEs were opened and embryos were weighed. Embryos inoculated with HAdV-D36 had significantly different weight compared to control embryos (inoculated with PBS). The greatest differences between the HAdV-D36-infected and control embryos were found for these that were inoculated into the yolk. In this group, weight differences between HAdV-D36-infected and control embryos ranged between 0.14 g and 1.09 g up to day 12 of development. In embryos older than 12 days, the greatest weight differences were noted for day 15 (3.51 g) and day 18 (5.41 g). In the case of embryos inoculated into the allantoic cavity, the differences were not as significant, ranging between 0.1 g (6th day of development) and 3.33 g (day 20 of development). On days 6, 10, 17, and 19 the weight of control embryos was even higher than of the infected ones.
Detailed data about the weight of embryos infected in the yolk sack and in the allantoic cavity are presented in the Table 1.
The analysis of weight changes of the studied embryos showed that the polynomial relationship of second degree describes the situation the most accurately (Figure 1). We observed a significant change in weight gain occurring between day 12 and 13 (the area marked a grey dashed line on Figure 1), followed by greater weight gains past day 12 in all study groups.
Further analysis of data, including the observed shift in weight gains in older embryos, showed that the embryo development is better described by separate linear correlations, one for day 6 to day 12 period, and second for day 13 to day 20 (Figure 1).
For the earlier development (only 6th to 12th day), all the obtained linear correlations were statistically significant (p < 0.001), regression coefficients showed a very strong positive correlation. We noticed that β coefficients for HAdV-D36- infected ECEs were almost equal when comparing the ECEs inoculated in the allantoic cavity and into the yolk sac (0.6437 and 0.6483, respectively). In the case of control ECEs inoculated in the yolk sack, we observed a much lower value of β coefficient when compared to control ECEs inoculated into allantoic cavity, (0.4936 vs. 0.7429). For later ECEs development (13th to 20th day), we obtained much higher slope coefficients (β) than for the early development for all ECEs groups, indicating greater mean weight gains in the later embryo’s development. We observed the same tendency as for the earlier development – the ECEs infected with HAdV-D36 into the allantoic cavity and into the yolk sack had similar values of β coefficient (3.7137 and 3.3257, respectively), however, surprisingly the ECEs inoculated into the allantoic cavity presented slightly stronger increase in weight. This tendency occurred also in the control ECEs; we observed greater weight gain in control ECEs inoculated into the allantoic cavity compared to these inoculated into the yolk sack.
2.2. Histopathological examinations
We observed very clear changes in embryonic development as early as 48 hafter HAdV-D36 infection. In general, we noted accelerated proliferation of whole-body cells. In the 7-day-old embryos, mesenchyme of the dermis in the HAdV-D36-infected ECEs was at a significantly more advanced stage of development compared to control ECEs. This early connective tissue contained more collagen fibres and was more optically dense than in the control. Both the liver and the kidney presented accelerated growth and increased angiogenesis.
On day 8, HAdV-D36-infected embryos had cartilage, intervertebral muscles, the liver, and kidneys much more advanced in development, than those observed in the control embryos.
After observing the embryos for the first 4 days, it was possible to conclude that the HAdV-D36-infected embryos development was about 2 days more advanced than the development of the control embryos.
On the 9th (Figure 2) day of development, it was possible to notice the initial stages of cartilage tissue remodelling, the formation of joints in the limbs or the initial phases of eye tissue organisation. in HAdV-D36-infected ECEs. The liver and kidneys were still enlarged and more advanced than in the control group. No increased number of adipocytes in the chicken body was observed.
On the 10th day of HAd-VD36 infection we observed, the initial phases of respiratory tract development, including clear formation of lungs, which is unique for this stage of development of birds, as normally they begin to develop later.
On the 13th day of HAdV-D36-infected embryos development, we observed a glandular period in the developing lungs and the first drops of fat appearing in the liver. We observed a constantly increasing growth of all organs, with particular emphasis on the liver and kidneys, which were growing significantly every day until day 20 of embryo development. However, on day 15 (Figure 3), we observed first inflammatory changes in the liver, which followed by the extensive limphocyte-macrophage infiltration to the liver parenchyma and the vicinity of blood vessels in the next days.
On days 19 and 20 (Figure 4), we observed substantial liver inflammation with the initial stages of a very pronounced liver lipidosis.
2.3. Real-time PCR
The analysis of real-time PCR results showed that the highest CQ (quantification cycle) concentration of HAdV-D36 genetic material in the embryos liver can be found on day 20 and the lowest on day 17 for both tested groups (Figure 5 and Figure 6). Despite the low Cq value, the viral DNA was present in each of the tested samples from HaDV-D36-infected embryos. The negative control showed no rise in CQ levels.
2.4. Antioxidant and lipid peroxidation markers

Table 2.
Antioxidant and lipid peroxidation markers levels in the heart tissue of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36. .
Table 2.
Antioxidant and lipid peroxidation markers levels in the heart tissue of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36. .
| HEART | control n = 16 |
HAdV-D36-infected n = 18 |
t/z* | p |
|---|---|---|---|---|
| CAT (IU/g protein) | 1584±205 | 2585±798 | 5.13 | <0.001 |
| GR (IU/g protein) | 2.6 (2.3-3.0) | 3.0 (2.2-3.9) | 0.57* | 0.569 |
| TOS (μmol/g protein) | 2.6±0.5 | 2.5±0.8 | 0.22 | 0.826 |
| SOD (NU/mg protein) | 58.0 (52.5-59.0) | 74.6 (70.9-76.2) | 4.47* | <0.001 |
| MnSOD (NU/mg protein) | 33.8±4.1 | 40.8±8.0 | 2.41 | <0.05 |
| CuZnSOD (NU/mg protein) | 22.4±5.1 | 31.6±12.9 | 2.81 | <0.05 |
| GPx (IU/g protein) | 4.0±0.9 | 3.8±0.9 | 0.62 | 0.542 |
| GST(IU/g protein) | 1.24±0.23 | 1.62±0.49 | 2.19 | <0.05 |
| MDA (μmol/g protein) | 1.6 (1.2-1.8) | 3.3(1.8-5.1) | 3.05* | <0.01 |
Legend: CAT (IUiu/g protein) – catalase, CuZnSOD (NU/mg protein)- Copper-zinc-superoxide dismutase, GPx (IU/g protein)-glutathione peroxidase, GR (IU/g protein)- glutathione reductase, GST (IU/g protein)- glutathione S transferase, MDA (μmol/g protein)- malondialdehyde, MnSOD (NU/mg protein)- manganase superoxide dismutase, SOD (NU/mg protein)- superoxide dismutase, TOS (μmol/g protein)- total oxidative status, t – test statistics in t-Student test for independent sample, z – test statistics in Mann-Whitney test, p – statistical significance, M±SD – mean ± standard deviation, Me(Q1-Q3) – median(lower-upper quartile). .
We found statistically significant differences between the CAT activity in the heart tissue of HAdV-D36-infected and control embryos with higher CAT activity in the heart tissue of HAdV-D36-infected embryos (average 1000 IU/g protein, 95% CI: 593-1408 IU/g protein). 3Differences were found in SOD levels between groups (p<0.001). Higher results were observed in the study group. Statistically significant differences in MnSOD levels were shown between the study and control groups (p<0.05). The study group had higher results with an average of 7.0 IU/g protein. 95% CI: 0.8-13.3 IU/g protein. There were statistically significant differences in CuZnSOD concentrations between the study and control groups (p<0.05). The study group had higher results with an average of 9.3 IU/g protein, 95% CI: 2.4-16.1 IU/g protein. Statistically significant differences in GST levels were found between the study and control groups (p<0.05). The study group had higher results with an average of 0.28 IU/g protein, 95% CI: 0.02-0.55 IU/g protein. There were statistically significant differences in MDA levels between groups (p<0.01). Higher results were observed in the study group.
There were no differences in the GPX parameter between the analyzed groups (p=0.542). There were no differences between groups in GR concentration (p=0.569) or in the TOS parameter between the groups analyzed (p=0.826).
Table 3.
Antioxidant and lipid peroxidation markers levels in the liver tissue of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36. .
Table 3.
Antioxidant and lipid peroxidation markers levels in the liver tissue of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36. .
| control n = 16 |
HAdV-D36-infected n = 18 |
t/z* | p | |
|---|---|---|---|---|
| CAT (IU/g protein) | 4596±1255 | 4834±1487 | 0.50 | 0.617 |
| GR (IU/g protein) | 0.84±0.15 | 1,31±0,37 | 5.04 | <0.001 |
| TOS (μmol/g protein) | 2.1±0.5 | 2.9±0.9 | 2.93 | <0.01 |
| SOD (NU/mg protein) | 63.3±7.9 | 90.4±30.6 | 3.30 | <0.01 |
| MnSOD (NU/mg protein) | 40.1±7.1 | 38.0±16.8 | 0.50 | 0.623 |
| CuZnSOD (NU/mg protein) | 28.0 (18.8-31.2) | 38.0 (31.9-66.7) | 3.36* | <0.001 |
| GPx (IU/g protein) | 7.0±1.3 | 5.5±1.3 | 3.58 | <0.01 |
| GST (IU/g protein) | 5.14±0.99 | 4.72±1.14 | 1.15 | 0.258 |
| MDA (μmol/g protein) | 0.77 (0.60-1.08) | 1.35(1.16-1.92) | 2.43* | <0.05 |
Legend: CAT (IUiu/g protein) – catalase, CuZnSOD (NU/mg protein)- Copper-zinc-superoxide dismutase, GPx (IU/g protein)-glutathione peroxidase, GR (IU/g protein)- glutathione reductase, GST (IU/g protein)- glutathione S transferase, MDA (μmol/g protein)- malondialdehyde, MnSOD (NU/mg protein)- manganase superoxide dismutase, SOD (NU/mg protein)- superoxide dismutase, TOS (μmol/g protein)- total oxidative status, t – test statistics in t-Student test for independent sample, z – test statistics in Mann-Whitney test, p – statistical significance, M±SD – mean ± standard deviation, Me(Q1-Q3) – median(lower-upper quartile.
There were differences in GR concentrations between groups (p<0.001). Higher concentrations were observed in the study group with an average of 0.47 IU/g protein, 95%PU: 0.28-0.66 IU/g protein. TOS also showed differences in concentrations between groups (p<0.01). In the study group, higher concentrations were observed with an average of 0.8 mol/g protein, 95%PU: 0.3-1.3 mol /g protein. There were statistically significant differences in SOD concentrations between the study and control groups (p<0.01). The study group had higher results with an average of 24.1 NU/mg protein, 95% CI: 8.8-39.2 NU/mg protein.
Differences were found between groups in CuZnSOD concentrations (p<0.001). Higher CuZnSOD values were observed in the study group compared to the control. GPX shown differences in concentrations between groups (p<0.01). Higher GPX values were observed in the study group with an average of 1.53 IU/g protein, 95%CI: 0.7-2.4 IU/g protein. There were differences in MDA concentrations between groups (p<0.05). Higher results were observed in the study group. There were no differences in concentration between groups in GST (p=0.258) and MnSOD (p=0.623). There were no differences in liver CAT concentration between the study groups (p=0.617).
3. Discussion
The influence of the virus on the host organism are comprehensively described. However, the studies conducted on animal models and humas so far include only adult individuals. Due to ethical reasons, no human studies have been carried out on an embryo or fetus. Our data suggest that HADV-D36 effect on embryonated chicken eggs (ECEs) results from completely different mechanism than the one described by Poterio et al. [3]. Our measurements of the chicken embryos weight changes agree with the observations made by Dhurandhar et al., Pasarica et al., and Shirani et al., who also showed a significant increase in the weight of infected animals compared to control animals [2,6,22]. Also, statistical analysis of our results confirms significant difference in the weight between the virus-infected and control embryos. However, our histopathological examinations showed no increase in fat cells growth, no tissue hypertrophy but accelerated cell proliferation in the infected embryos. The amount of fat in their cells and tissues remained at a normal level, but in some cases, steatosis of some organs was also observed. However, due to the absence of any accompanying cytopathic effects on the tissues allowing to determine their individual characteristics, the changes did not result from viral infection. It suggests that the mechanism behind it is significantly different from the known effects of HADv-D36. It seems that the virus finds a susceptible environment in the embryo. Histopathological analyses are consistent with the weight measurements of the embryos. However, the presented data indicate that the increased mass of the examined embryos does not result from an increase in the number of fat cells in the tissues of infected embryos, but from increased cell division in all tissues of the body, which accelerated development and increased organs size. The observed changes might result from an unknown mechanism induced by HAdV-D36 infection. Whether it is due to the high number of differentiating stem cells or what is the mechanism behind it still remains unknown. Perhaps it is related to the expression of developmental genes, whose activation suppresses the genes necessary for adipocyte transformation, or the genes responsible for adipocyte transformation are not yet active.
The only element that seems to work similarly in both adults and embryos is the inflammation caused by the virus. Ha-Na et al. proved that human adenovirus 36 is responsibl increased levels of MCP-1 (macrophage chemoattractant protein 1) because it activates the nuclear factor Kß (transcription factor), causing the inflammation in the adipose tissue [23].
The real-time PCR results has showed an unusual fluctuation in the CQ level in the livers of chickens’ embryos. The viral load between day 13 and 14 showed a slight increase, but on day 15 it started to decrease for the next two days. On day 18, levels of viral DNA increased again. The decrease in the amount of viral genetic material was most likely related to the enhanced cellular response and increased antibody titer [24]. The unusual increase in viral genetic material observed on day 18 was probably connected to the decline in antibody titer which could no longer stop the virus’ multiplication. However, this hypothesis requires further research, because the small amount of serum in studied embryos did not allow to measure the level of antibodies. The chicken embryos are characterised by the presence of an immature, but already functioning, immunological system, which comprises antibodies and inflammatory cells [25,26]. The observed fluctuation in CQ levels coincided with the histopathological picture showing high number of macrophages and lymphocytes between day 13 and day 20 of the embryos’ development.
The presented data indicate the accelerated tissue proliferation, however do not point out the exact mechanism. Therefore, the observed changes require further research that will allow to unveil more details and explain the phenomenon in question. Heart failure mechanism is associated with changes in fat tissue quantity, inflammatory processes, and altered cardiac physiology that is additionally complicated by co-morbidities [27]. In these conditions of heart dysfunctions, the impaired glucose metabolism and fatty acid ß-oxidation disrupt mitochondrial functions and promote the formation of reactive oxygen species (ROS). In the presented study, we observed significantly increased levels of oxidative stress markers: CAT, GPx, TOS, total SOD, CuZnSOD, and MDA, both in ECEs heart and liver tissue, indicative if increased oxidative stress and ROS levels. Reactive oxygen species negatively affect the function and structure of all cellular macromolecules, including nucleic acids, proteins, and lipids. They can alter Ca2+ regulation, activate pathways linked to electrical remodeling, stimulate cardiomyocytes hypertrophy, induce apoptosis, promote fibrosis, and activate or inhibit the inflammatory response. All of which are understood to be crucial factors in the onset of heart failure [28,29] Reactive oxygen species can modify numerous signaling pathways involved in the hypertrophy of cardiomyocytes. For instance, apoptotic signal-regulating kinase 1 (ASK1) in rat’s ventricular cardiomyocytes is activated in a redox-dependent manner by angiotensin II, endothelin-1, and phenylephrine causing their hypertrophy [30] Our findings indicate that CAT, SOD, GST and, to some extent, GR activity in the ECEs heart tissue is influenced by HAdV-D36 infection which lead to changes in the oxidative processes and hypertrophy of the heart tissue. In the liver, ROS overwhelm the enzymatic and non-enzymatic antioxidants, causing oxidative stress, hepatocellular dysfunction, and eventually hepatic fibrosis. Viral infections also increase levels of enzymatic and non-enzymatic oxidative stress markers in the liver [31,32]. According to Duygu et al., individuals with chronic HBV infections present higher levels of oxidative stress markers, such as hydroxyl, hydrogen peroxide, singlet oxygen, lipid hydroperoxide, and superoxide, and lower levels of antioxidant markers, such as total sulfhydryl, vitamin C, uric acid, vitamin E, and bilirubin in their blood [33] Increased glutathione reductase activity prevents the excessive quenching of intracellular ROS essential for insulin signalling [34]. Camini et al analyzed the impact of Caraparu virus (CARV) on the hepatic pathogenesis in the 6-week-old BALB/c mice and the role of oxidative stress and antioxidant defences in this pathology. CARV infection caused no change in the oxidative stress markers but caused an increase in glutathione content and altered SOD expression and activity [35]. Da Silva et al. studied impact of Oropouche virus (OROV) on the hepatic stress in male and female wild-type BALB/c mice. They reported that SOD and CAT activity in the liver and spleen decreased after OROV infection, indicating the antioxidant response as ineffective in stopping the oxidative damage [36]. In our study, we observed increased GR and total SOD activities and decreased MnSOD, CuZnSOD, GPx, and GST activities. Moreover, HAdV-D36 infection significantly increased MDA concentration and TOS leves. The presented data show that oxidative stress generated by HADV-D36 results from imbalance in redox homeostasis started by exccesive ROS production that negatively affects biomolecules or incapacitates the antioxidant system.
4. Materials and Methods
4.1. Ethical statement
The use of embryonated chicken eggs (ECE) was approved by the Local Ethics Committee in Wrocław (approval no. 021/2021; date of approval 17.03.2021).
4.2. Virus
Human adenovirus 36 (HAdV-D36) (ATCC VR1610TM, Manassas, Virginia, US) was used in the study.
4.3. Embryonated chicken eggs (ECEs)
Four hundred-twenty embryonated chicken eggs (ECEs) of the Rossa I hens (Gallus gallus domesticus) were used in the study. On the fifth day of incubation, ECEs were divided into 4 groups:
- Ad-AC: inoculated with HAdV-D36 into the allantoic cavity (n = 105),
- Ad-YS: inoculated with HAdV-D36 into the yolk sack (n = 105),
- C-AC: inoculated with PBS into allantoic cavity (n = 105) – control group,
- C-YS: inoculated with PBS into yolk sack (n = 105 eggs) – control group.
The inoculation via allantoic cavity and yolk sack was chosen to imitate oral and droplet route of infection.
4.4. Inoculation of ECEs
The edges of the air sac of ECEs were marked under the ovoscope. Then, the position of embryo, the location of the large blood vessels, and the site of viral inoculation were determined. Two holes in the shell were drilled: above the air sac (to reduce the pressure after the injection) and above the site of inoculation (into yolk sac or into allantoic cavity). 100 µl of 100 TCID50 = 10-6 of virus (or PBS in case of control embryos) were injected into the yolk sac or allantoic cavity. Then, ECEs were transferred to an incubator and kept at 37 °C and 80% of humidity.
4.5. ECEs weight measurements
Starting from day 6 to day 20 of the incubation period (15 days in total), 28 embryos (7 from each of 4 groups) were extracted and weighed each day on the laboratory scales (Radwag WLCX2, Poland).
4.6. Sampling ECEs for histopathological and molecular studies
Chicken embryos removed from the eggs were dissected with a surgical scalpel. Until the 10th day of incubation, the entire embryos were taken due to the small size of the internal organs. After the 10th day of incubation, only livers were sampled for future examinations.
4.7. Histopathological examinations
Whole embryos (up to 10 days) and livers taken from each embryos group were subjected to histopathological examination in order to observe the changes that caused HAdV-D36 infection in the tissues of the examined organism.
The sampled material was fixed in 4% solution of buffered formaldehyde (pH 7,2-7,4). Then, the material was rinsed in tap water, and dehydrated in alcohol series. Eventually, it was embedded in paraffin, sliced into 6 µm thick samples, and stained with haematoxylin and eosin. The material was observed under a light microscope (Nikon Eclipse 80i, Nikon, Japan).
4.8. Real Time PCR
Real time PCR tests were performed to confirm replication of the virus and to determine the viral load in liver cells. A commercial Bead-Beat Micro AX Gravity kit (A&A Biotechnology, Gdańsk) was used to isolate DNA from the samples. Quantitative polymerase chain reaction (qPCR) was done using an Amplifyme Probe no-rox mix (am04) (Blirt, Gdańsk). Genetic material of the virus was identified using the following primers and probe for the Eorf4 gene (GenBank accession number: GQ384080.1):
Forward primer: AAAGAGCAGCACAGAGAGATCA (Position 34417-34438),
Reverse primer: GAGTGAGCGTGCTGGTTC (Position 34533- 34550),
Probe: FAM-TTCAAGGCCATAAATCTGCCCTGATATCCA-BHQ1 (Position 34499 - 34528).
The qPCR followed the protocol: 2 min at 50°C, 10 min at 95°C, 45 cycles of 20 s at 95°C, and 1 min at 60°C.
4.9. Antioxidant and lipid peroxidation markers
On the 18th day of ECEs development, the heart and liver of HAdV-D36-infected (Ad-AC n = 9, Ad-YS n = 9) and control (C-AC n = 8, C-YS n = 8) embryos were collected for antioxidant and lipid peroxidation markers analyses. The samples were homogenized, centrifuged at 12000 g, 4 °C for 10 min and then stored in -80 °C until the analyses. The analyses were done using colorimetric methods and the changes of absorbances were read on PERKIN ELMER Victor X3 reader (PerkinElmer, Inc., Waltham, Massachusetts, United States). The protein content was measured using the Lowry method [13].
4.9.1. Catalase (CAT) activity (EC 1.11.1.6)
CAT activity was assessed using the Aebi method [14]. CAT activity was expressed as units of activity per 1 g of protein (IU/g protein).
4.9.2. Superoxide Dismutase (SOD) activity (EC 1.15.1.1)
SOD activity was determined with the Oyanagui method [15]. SOD activity was expressed as nitrite units (NU) per 1 mg of protein, with1 NU equal to 50% inhibition of nitrite ion formation under the method’s condition. CuZnSOD activity was assessed by calculating the difference between total SOD and MnSOD activity in the presence of potassium cyanide (KCN) as CuZnSOD inhibitor.
4.9.3. Glutathione Peroxidase (GPx) activity (EC 1.11.1.9)
GPx activity was measured using the kinetic method [16], with t-butyl peroxide as a substrate and expressed as μmoles of NADPH oxidized in 1 min per 1 g of protein.
4.9.4. Glutathione Reductase (GR) activity (EC 1.8.1.7)
GR activity was assessed using the kinetic method and expressed as μmoles of nicotinamide adenine dinucleotide phosphate (NADPH) utilized in reaction with oxidized glutathione over 1 min per 1 g of protein [17].
4.9.5. Glutathione-S Transferase (GST) activity (EC 2.5.1.18)
GST activity was estimated using the Habig and Jakoby kinetic method [18] and expressed as μmoles of thioether formed over 1 min per 1 g of protein.
4.9.6. Total oxidant status (TOS)
4.9.7. Malondialdehyde (MDA) concentration
Malondialdehyde (MDA) concentration was measured using the method of Ohkawa et al. [21], using the reaction with thiobarbituric acid. MDA concentration in the plasma was calculated against the standard curve prepared from 1,1,3,3-tetraethoxypropane and was expressed in μmol/L.
4.10. Statistical analysis
The normality of distribution of ECEs weight and the antioxidant and lipid peroxidation markers levels were assessed using the Shapiro-Wilk test and a quantile-quantile plot. In the case of ECEs weight data, the relationship between quantitative variables was determined using polynomial regression or ordinary least square regression and data are presented as mean and the 95% confidence interval (95% CI). Data of antioxidant and lipid peroxidation markers with normal distribution were presented as mean value and standard deviation (M ± SD), while data with non-normal distribution as median and the lower and upper quartiles (Me(Q1-Q3)). The student’s t test for independent samples was used for group comparisons, and homogeneity of variance was assessed using the Fisher-Snedecor test. In the absence of homogeneity of variance, the Welch correction test was used. Differences between group were determined based on mean value and a a 95-confidence interval (CI). For variables deviating from normality, the Mann-Whitney U test was used. All tests were two-tailed and a significance level was < 0.05. Statistical analyses were performed using Statistica v. 13.3.0 (TIBCO Software Inc.).
Author Contributions
B.B. and A.P. convinced the project. Data analyses was performed by A.P., P.K., D.S., K.M., E.C. and J.Z.F. Manuscript was written by A.P, P.K and D.S., K.M. with input from A.K and P.J
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper
References
- Mitra, A. K., & Clarke, K. Viral obesity: Fact or fiction? In Obesity Reviews, 2010, 11, pp. 289–296,. [CrossRef]
- Pasarica, M., & Dhurandhar, N. V. Infectobesity: Obesity of Infectious Origin. Advances in Food and Nutrition Research, 2007, 52, 61–102. [CrossRef]
- Ponterio, E., & Gnessi, L. Adenovirus 36 and Obesity: An Overview. Viruses,1980.7, 3719–3740. [CrossRef]
- Nam, J. H., Na, H. N., Atkinson, R. L., & Dhurandhar, N. V. Genomic stability of adipogenic human adenovirus 36. International Journal of Obesity, 2014, 38(2), 321–324. [CrossRef]
- Adenoviridae ~ ViralZone. (n.d.). Retrieved October 17, 2023, from https://viralzone.expasy.org/4.
- Dhurandhar, N. v, Kulkarni A’ J, P., Ajinkya, S. M., & Sherikar, A. Effect of adenovirus infection on adiposity in chicken. In Veterinary Microbiology, 1992, Vol. 31.
- Rathod, M. A., Rogers, P. M., Vangipuram, S. D., McAllister, E. J., & Dhurandhar, N. V. Adipogenic cascade can be induced without adipogenic media by a human adenovirus. Obesity, 2009, 17(4), 657–664. [CrossRef]
- Laron, Z. Insulin - A growth hormone. In Archives of Physiology and Biochemistry,2008, 114(1), pp. 11–16). [CrossRef]
- Pasarica, M., Shin, A. C., Yu, M., Yang, H. M. O., Rathod, M., Catherine Jen, K. L., MohanKumar, S., MohanKumar, P. S., Markward, N., & Dhurandhar, N. v. Human adenovirus 36 induces adiposity, increases insulin sensitivity, and alters hypothalamic monoamines in rats. Obesity, 2006, 14(11), 1905–1913. [CrossRef]
- Almgren, M., Atkinson, R., He, J., Hilding, A., Hagman, E., Wolk, A., Thorell, A., Marcus, C., Näslund, E., Östenson, C. G., Schalling, M., & Lavebratt, C. Adenovirus-36 is associated with obesity in children and adults in Sweden as determined by rapid ELISA. PLoS ONE, 2012, 7(7). [CrossRef]
- Zhou, Y., Pan, Q., Wang, X., Zhang, L., Xiao, F., & Guo, L. The relationship between human adenovirus 36 and obesity in Chinese Han population. Bioscience Reports, 2018,38, 20180553. [CrossRef]
- Chudnovets, A., Liu, J., Narasimhan, H., Liu, Y., & Burd, I. Role of Inflammation in Virus Pathogenesis during Pregnancy. Journal of Virology, 2020,. [CrossRef]
- Lowry, O. H., Rosebrough, N. J., Farr, A. L., Randall, R. J. Protein measurement with the folin phenol reagent. Journal of Biological Chemistry, 1951,193(1), 265–275. [CrossRef]
- Aebi, H. [13] Catalase in vitro. Methods in Enzymology, 1984, 105(C), 121–126. [CrossRef]
- Oyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Analytical Biochemistry, 1984, 142(2), 290–296. [CrossRef]
- Mannervik, B. [60] Glutathione peroxidase. Methods in Enzymology, 1985, 113(C), 490–495. [CrossRef]
- Carlberg, I., & Mannervik, B. [59] Glutathione reductase. Methods in Enzymology, 1985,113(C), 484–490. [CrossRef]
- Habig, W. H., & Jakoby, W. B. [51] Assays for differentiation of glutathione S-Transferases. Methods in Enzymology,1981,77(C), 398–405. [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clinical Biochemistry, 2005 38(12), 1103–1111. [CrossRef]
- Erel, O. A novel automated method to measure total antioxidant response against potent free radical reactions. Clinical Biochemistry, 2004, 37(2), 112–119. [CrossRef]
- Ohkawa, H., Ohishi, N., & Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry, 1979, 95(2), 351–358. [CrossRef]
- Karandish, M., Shirani, F., Teimoori, A., Rashno, M., & Latifi, M. Using rats as a research model to investigate the effect of human adenovirus 36 on weight gain. In ARYA Atheroscler,2017, 13.
- Na, H. N., & Nam, J. H. Adenovirus 36 as an obesity agent maintains the obesity state by increasing MCP-1 and inducing inflammation. Journal of Infectious Diseases, 2012, 205(6), 914–922. [CrossRef]
- [24] Ribatti, D., Tamma, R., & Elieh Ali Komi, D. The morphological basis of the development of the chick embryo immune system. In Experimental Cell Research, 2019, 381(2) , pp. 323–329). Elsevier Inc. [CrossRef]
- Alexis, L., Romanoff, M., The Avian Embry Structural and functional development. Science, 1960,16,1305,.
- Garcia, P., Wang, Y., Viallet, J., & Macek Jilkova, Z. The Chicken Embryo Model: A Novel and Relevant Model for Immune-Based Studies. In Frontiers in Immunology ,2021, 12,. [CrossRef]
- Kindel, T. L., & Strande, J. L. Bariatric surgery as a treatment for heart failure: review of the literature and potential mechanisms. Surgery for Obesity and Related Diseases, 2018, 14(1), 117–122. [CrossRef]
- Burgoyne, J. R., Mongue-Din, H., Eaton, P., & Shah, A. M. Redox signaling in cardiac physiology and pathology. In Circulation Research, 2018, 111(8), pp. 1091–1106,. [CrossRef]
- Aimo, A., Castiglione, V., Borrelli, C., Saccaro, L. F., Franzini, M., Masi, S., Emdin, M., & Giannoni, A. Oxidative stress and inflammation in the evolution of heart failure: From pathophysiology to therapeutic strategies. In European Journal of Preventive Cardiology, 2020, 27(5), pp. 494–510),. [CrossRef]
- Higuchi, Y., Otsu, K., Nishida, K., Hirotani, S., Nakayama, H., Yamaguchi, O., Matsumura, Y., Ueno, H., Tada, M., & Hori, M. Involvement of Reactive Oxygen Species-mediated NF- κ B Activation in TNF- α -induced Cardiomyocyte Hypertrophy. Journal of Molecular and Cellular Cardiology, 2002, 34(2), 233–240. [CrossRef]
- Pal, S., Polyak, S. J., Bano, N., Qiu, W. C., Carithers, R. L., Shuhart, M., Gretch, D. R., & Das, A. Hepatitis C virus induces oxidative stress, DNA damage and modulates the DNA repair enzyme NEIL1, 2010. [CrossRef]
- Smirnova, O. A., Ivanova, O. N., Bartosch, B., Valuev-Elliston, V. T., Mukhtarov, F., Kochetkov, S. N., & Ivanov, A. v. Hepatitis C Virus NS5A Protein Triggers Oxidative Stress by Inducing NADPH Oxidases 1 and 4 and Cytochrome P450 2E1.2016,. [CrossRef]
- Duygu, F., Karsen, H., Aksoy, N., & Taskin, A. (2012). Relationship of oxidative stress in hepatitis B infection activity with HBV DNA and fibrosis. Annals of Laboratory Medicine, 2012, 32(2), 113–118. [CrossRef]
- Raza, H., John, A., & Howarth, F. C. Alterations in Glutathione Redox Metabolism, Oxidative Stress, and Mitochondrial Function in the Left Ventricle of Elderly Zucker Diabetic Fatty Rat Heart. Int. J. Mol. Sci, 2012, 13, 16241–16254. [CrossRef]
- Caetano, F., Letícia, C. , Almeida, T., Carolina, Bernardes, S., Silva, M., Lú Cia Pedrosa, M., Caldeira, D., Wanderson, C., de Lima, G., Carla, Pinto, A., César, P., Ferreira, P., & Carlos De Magalhães, J. (n.d.). Caraparu virus induces damage and alterations in antioxidant defenses in the liver of BALB/c mice after subcutaneous infection. [CrossRef]
- Menegatto, M. B. da S., Ferraz, A. C., Lima, R. L. S., Almeida, L. T., Brito, R. C. F. de, Reis, A. B., Carneiro, C. M., Lima, W. G. de, Silva, B. de M., Magalhães, J. C. de, & Magalhães, C. L. de B. Oropouche virus infection induces ROS production and oxidative stress in liver and spleen of mice. Journal of General Virology, 2023,104(5), 001857. [CrossRef]
Figure 1.
Mean embryo weight in relation to the development day of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into the allantoic cavity (AC) or yolk sac (YC) over 15 days of incubation. In the equations notation EW means embryo weight [g] and DD means development day Gray vertical dashed line marks the moment of change in weight gain for all study groups.
Figure 1.
Mean embryo weight in relation to the development day of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into the allantoic cavity (AC) or yolk sac (YC) over 15 days of incubation. In the equations notation EW means embryo weight [g] and DD means development day Gray vertical dashed line marks the moment of change in weight gain for all study groups.
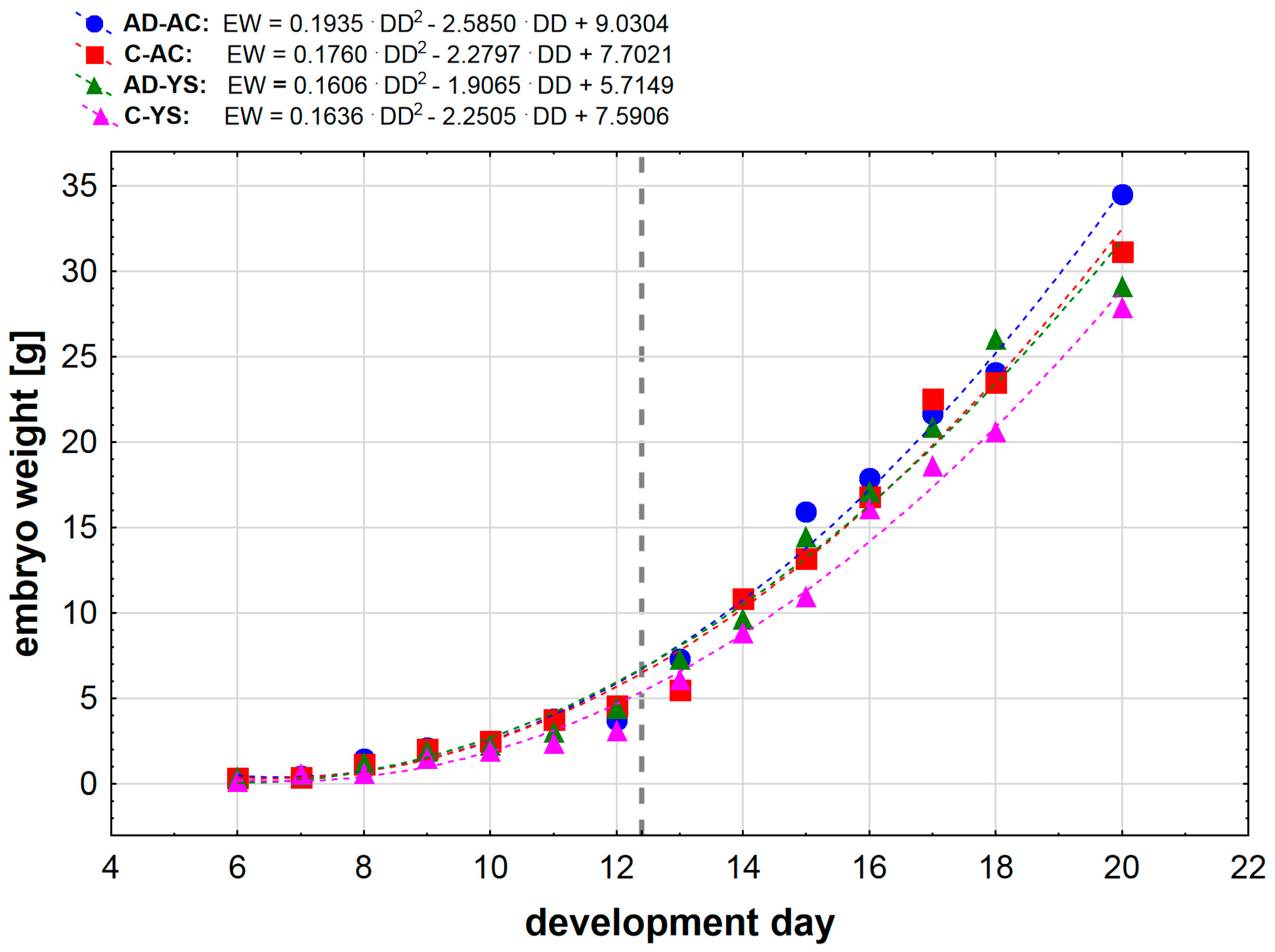
Figure 2.
Liver development (L) on the 9th day of incubation of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into yolk sac. The liver of control ECEs is dominated by wide blood vessels filled with blood (black arrow). In the HAdV-D36-infected ECEs, hepatocytes comprise a significant part of the liver parenchyma, the blood vessels are much less filled with blood, and lymphatic vessels are clearly visible. H&E, Mag. 100 (upper line) and 200x (lower line).
Figure 2.
Liver development (L) on the 9th day of incubation of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into yolk sac. The liver of control ECEs is dominated by wide blood vessels filled with blood (black arrow). In the HAdV-D36-infected ECEs, hepatocytes comprise a significant part of the liver parenchyma, the blood vessels are much less filled with blood, and lymphatic vessels are clearly visible. H&E, Mag. 100 (upper line) and 200x (lower line).
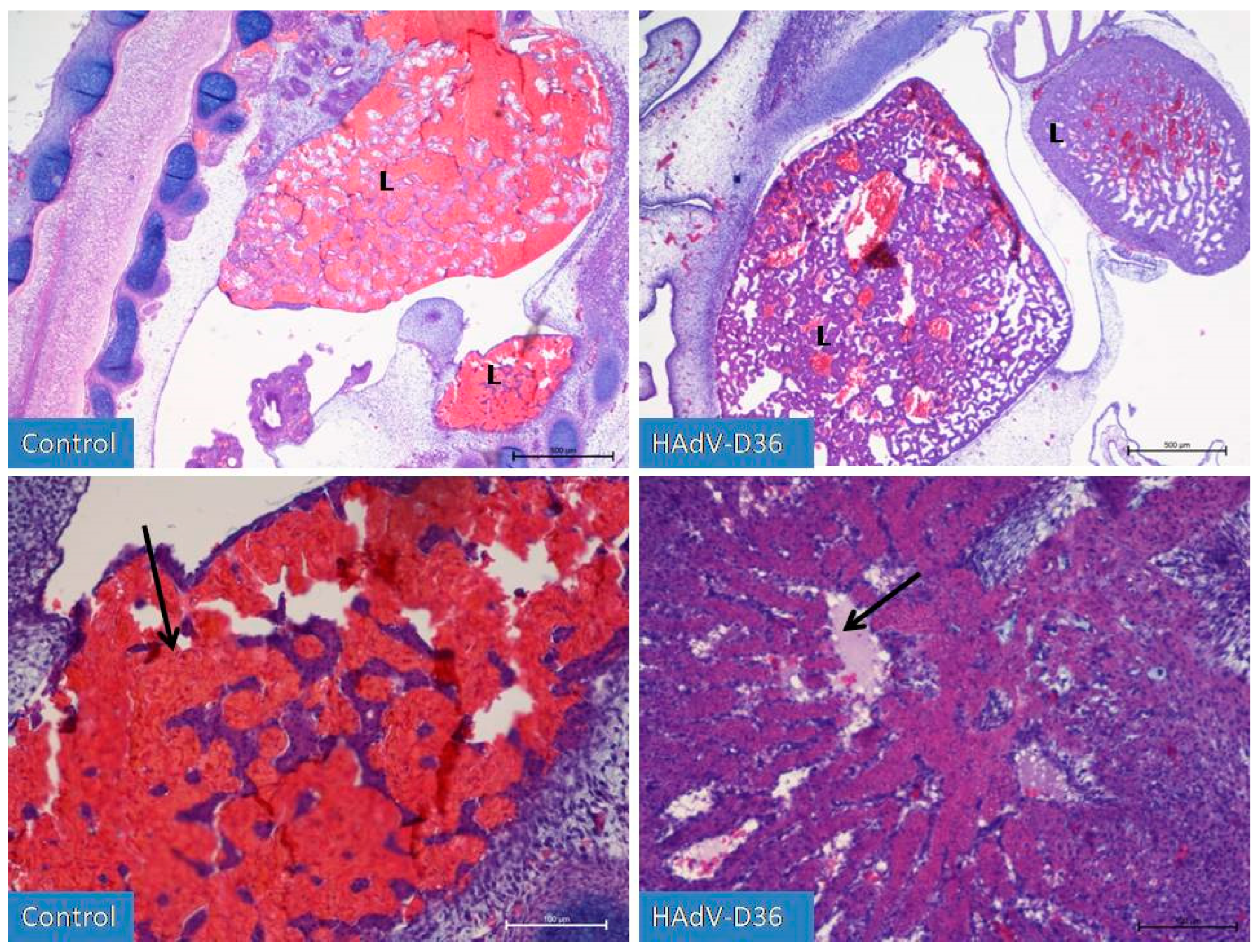
Figure 3.
Comparison of the degree of liver development on the 15th day of incubation. The hepatic trabeculae and blood vessels filled with blood to varying degrees are clearly visible in both in control and HAdV-D36-infected to the yolk sack ECEs. In the HAdV-D36-infected ECEs, clusters of lymphoid tissue appear in the liver parenchyma causing local inflammation (red arrow) followed by hepatocyte dystrophy. Fat droplets (black arrow) appear in the hepatocytes. H&E, Mag. 100 and 200x (right bottom picture).
Figure 3.
Comparison of the degree of liver development on the 15th day of incubation. The hepatic trabeculae and blood vessels filled with blood to varying degrees are clearly visible in both in control and HAdV-D36-infected to the yolk sack ECEs. In the HAdV-D36-infected ECEs, clusters of lymphoid tissue appear in the liver parenchyma causing local inflammation (red arrow) followed by hepatocyte dystrophy. Fat droplets (black arrow) appear in the hepatocytes. H&E, Mag. 100 and 200x (right bottom picture).
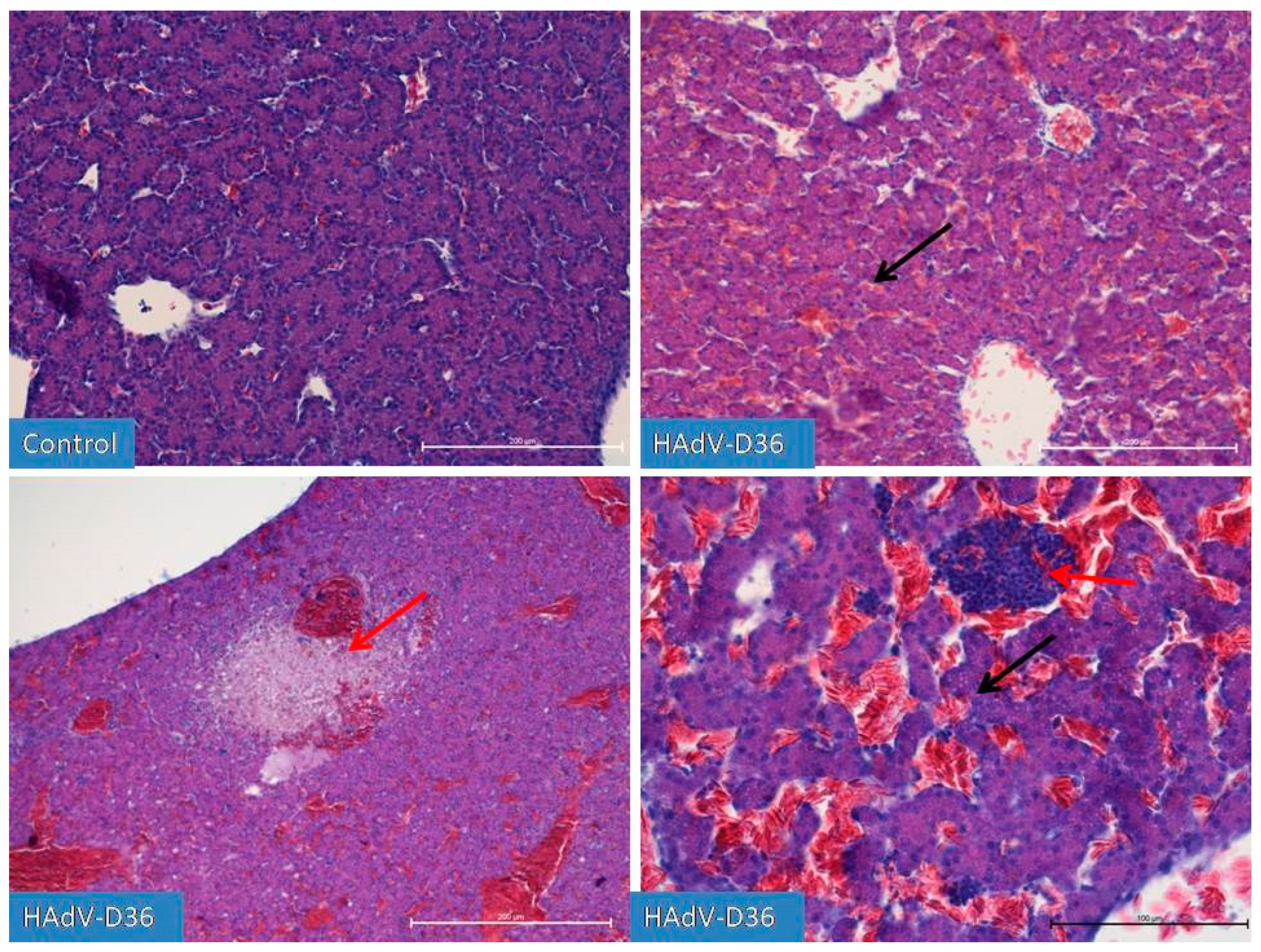
Figure 4.
Liver after infection with HAdV-D36 into the yolk sack on day 19 of incubation. Increasing lipogenesis in hepatocytes (black arrow), accompanied by macrophage and lymphocytic infiltration (red arrow) in the vicinity of blood vessels and in the liver parenchyma. The increased presence of congestive blood is strongly diverse and region dependent. Changes in the liver parenchyma are accompanied by the degradation of hepatocytes. H&E, Mag. 100 and 200x (right bottom line).
Figure 4.
Liver after infection with HAdV-D36 into the yolk sack on day 19 of incubation. Increasing lipogenesis in hepatocytes (black arrow), accompanied by macrophage and lymphocytic infiltration (red arrow) in the vicinity of blood vessels and in the liver parenchyma. The increased presence of congestive blood is strongly diverse and region dependent. Changes in the liver parenchyma are accompanied by the degradation of hepatocytes. H&E, Mag. 100 and 200x (right bottom line).
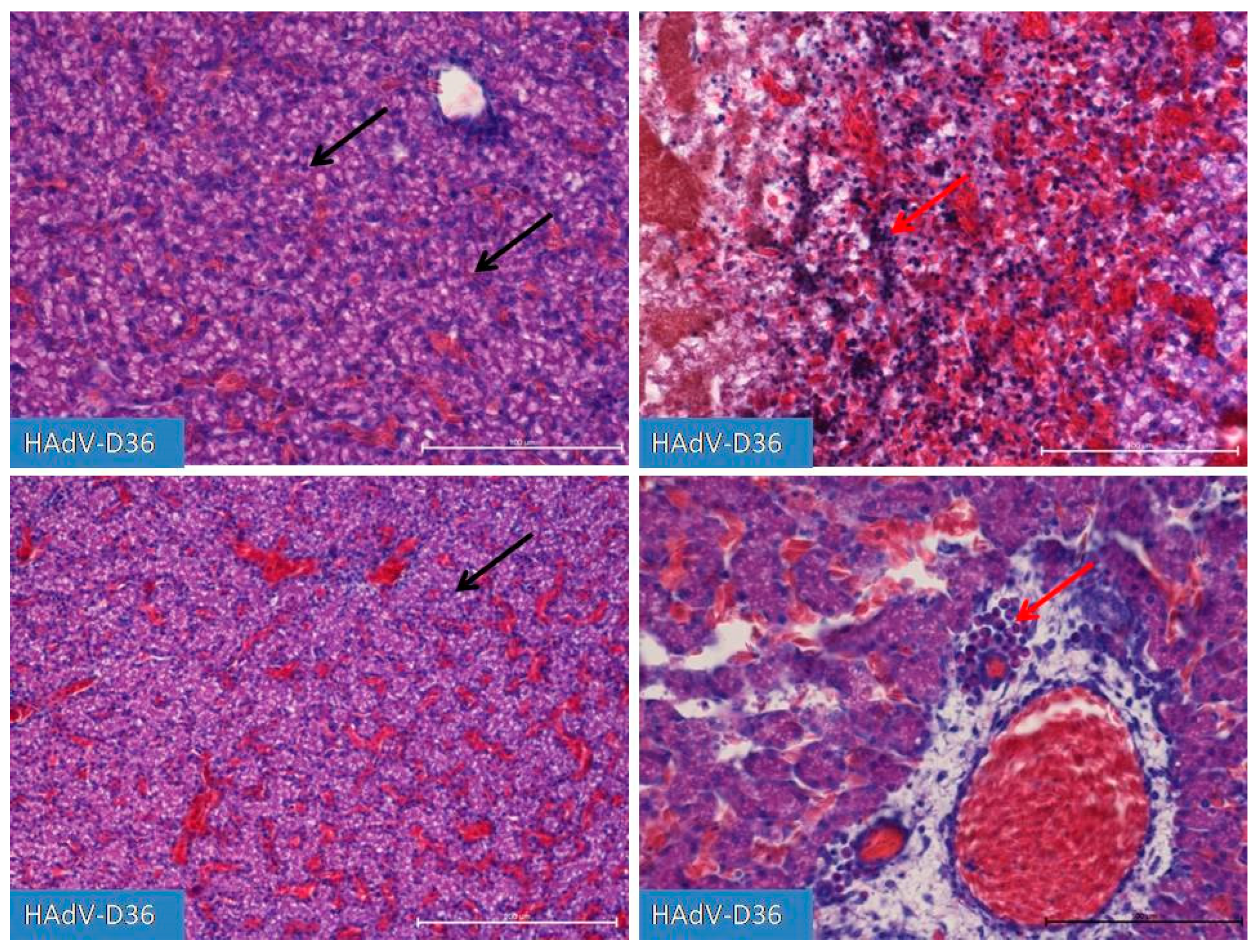
Figure 5.
Quantification cycle (CQ) level of HAdV-D36 genetic material isolated from the livers of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 or PBS into the yolk sack.
Figure 5.
Quantification cycle (CQ) level of HAdV-D36 genetic material isolated from the livers of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 or PBS into the yolk sack.
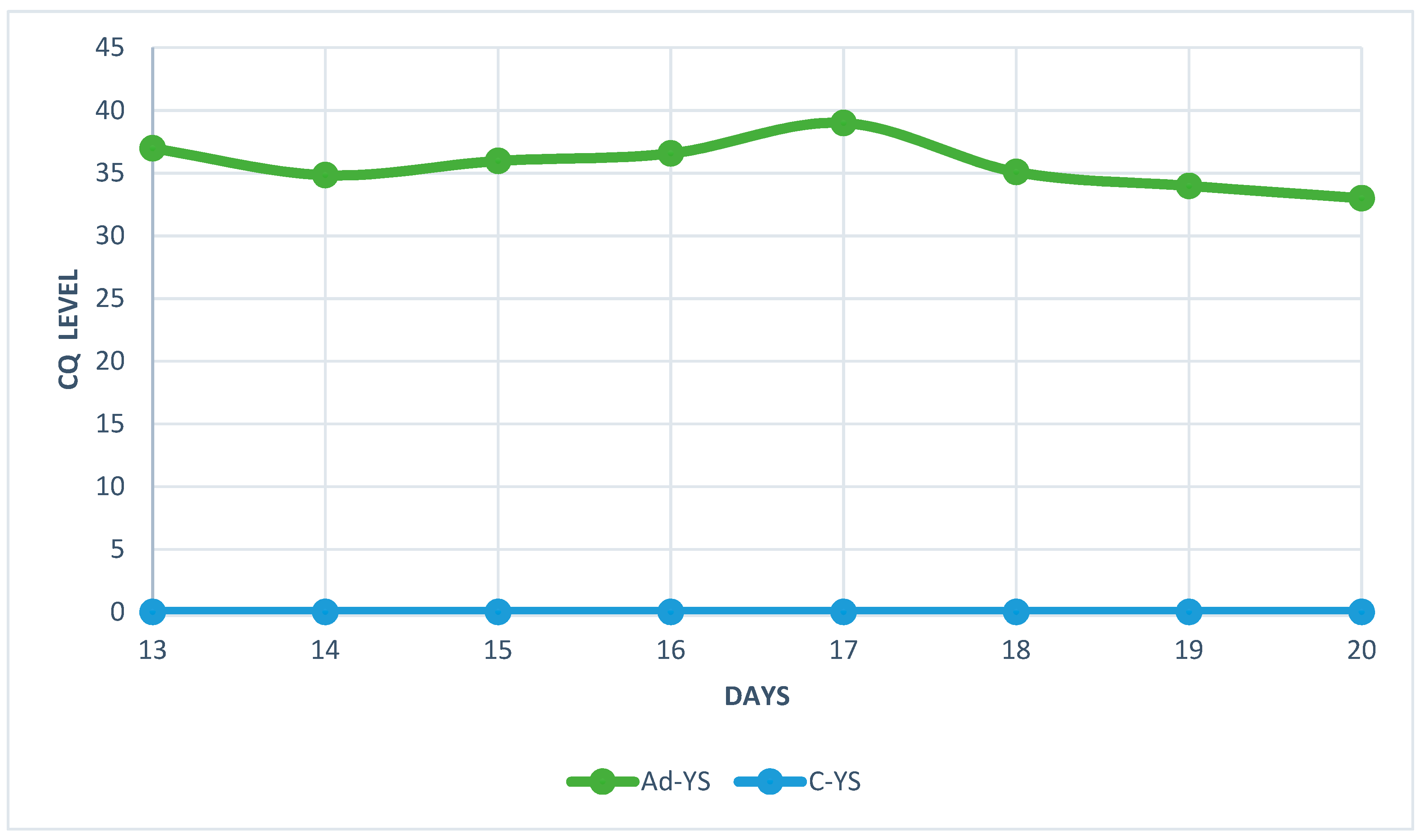
Figure 6.
Quantification cycle (CQ) level of HAdV-D36 genetic material isolated from the livers of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 or PBS into the yolk sack.
Figure 6.
Quantification cycle (CQ) level of HAdV-D36 genetic material isolated from the livers of embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 or PBS into the yolk sack.
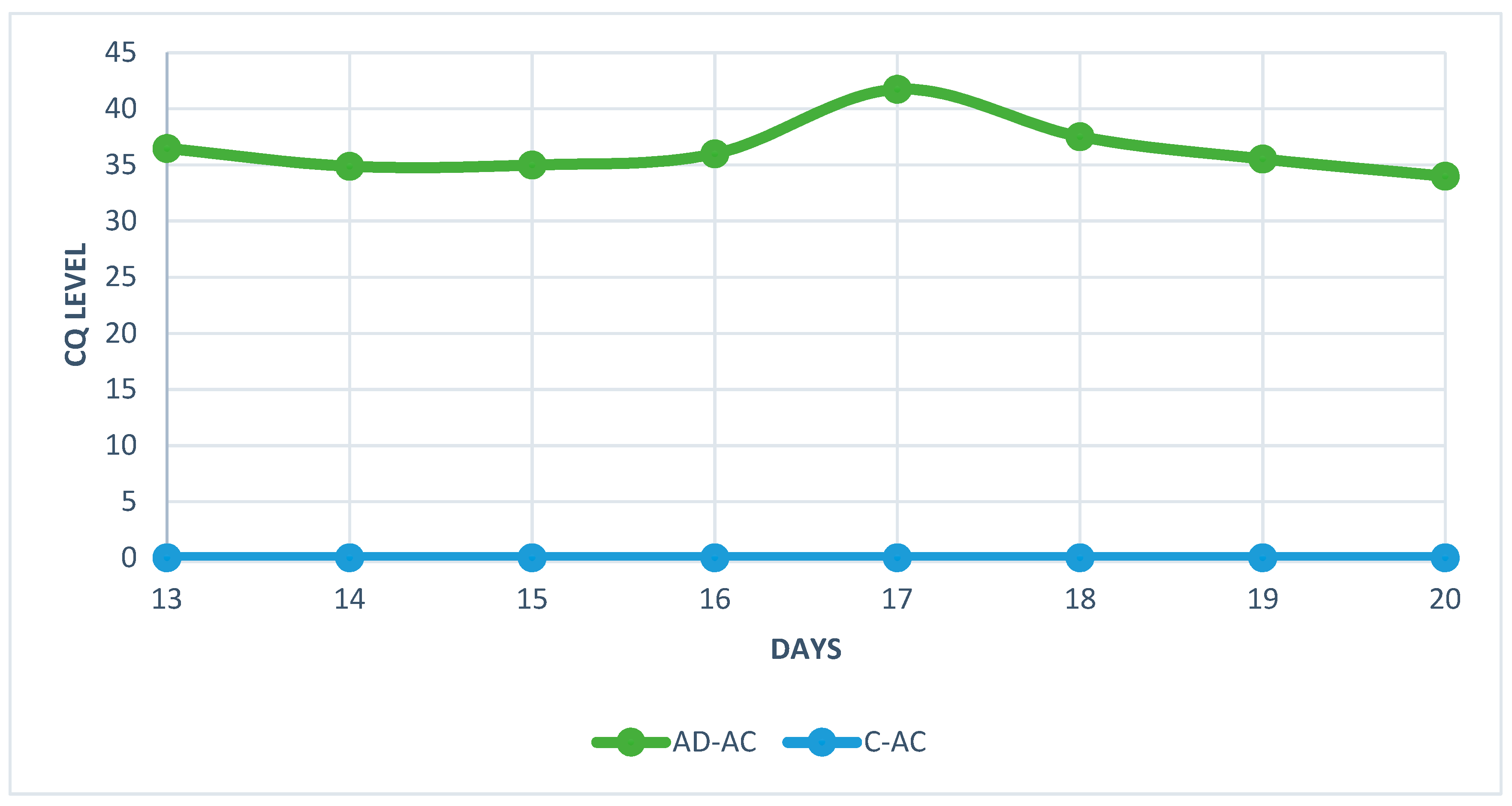
Table 1.
Weight of embryos from embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into the allantoic cavity (AC) or yolk sac (YC) over 15 days of incubation. Results are presented as mean ± standard deviation (Me ± SD).
Table 1.
Weight of embryos from embryonated chicken eggs (ECEs) inoculated with human adenovirus 36 (HAdV-D36) or PBS (control) into the allantoic cavity (AC) or yolk sac (YC) over 15 days of incubation. Results are presented as mean ± standard deviation (Me ± SD).
|
Day of incubation |
ECEs weight [g] | |||
| Ad-YS | C-YS | Ad-AC | C-AC | |
| 6 | 0.38 ± 0.12 | 0.15 ± 0.04 | 0.24 ± 0.10 | 0.34 ± 0.02 |
| 7 | 0.55 ± 0.17 | 0.57 ± 0.01 | 0.47 ± 0.14 | 0.40 ± 0.11 |
| 8 | 1.23 ± 0.38 | 0.63 ± 0.08 | 1.47 ± 0.51 | 1.16 ± 0.26 |
| 9 | 1.93 ± 0.39 | 1.50 ± 0.38 | 2.15 ± 0.21 | 2.03 ± 0.43 |
| 10 | 2.28 ± 0.43 | 1.93 ± 0.13 | 2.26 ± 0.45 | 2.51 ± 0.47 |
| 11 | 3.06 ± 0.65 | 2.39 ± 0.23 | 3.82 ± 0.90 | 3.78 ± 0.33 |
| 12 | 4.41 ± 1.32 | 3.12 ± 0.54 | 3.78 ± 1.00 | 4.58 ± 0.15 |
| 13 | 7.33 ± 2.04 | 6.13 ± 0.83 | 7.30 ± 1.44 | 5.52 ± 0.12 |
| 14 | 9.64 ± 2.37 | 8.84 ± 2.78 | 10.86 ± 0.46 | 10.86 ± 2.00 |
| 15 | 14.48 ± 2.25 | 10.97 ± 2.12 | 15.93 ± 0.53 | 13.20 ± 1.60 |
| 16 | 17.12 ± 4.29 | 16.12 ± 1.51 | 17.87 ± 0.33 | 16.82 ± 0.62 |
| 17 | 20.91 ± 0.65 | 18.62 ± 2.91 | 21.66 ± 0.50 | 22.63 ± 3.62 |
| 18 | 26.04 ± 1.13 | 20.63 ± 2.65 | 24.08 ± 0.39 | 23.52 ± 3.62 |
| 19 | 27.59 ± 2.01 | 26.45 ± 2.04 | 26.19 ± 0.19 | 27.66 ± 0.98 |
| 20 | 29.13 ± 2.32 | 27.91 ± 1.83 | 34.51 ± 0.30 | 31.18 ± 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



