
Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
The ZIPs (ZRT/IRT-like proteins) play critical roles in the absorption, transport, and intracellular balance of metal ions essential for various physiological processes in plants. However, little is known about the pan-genomic characteristics and properties of ZIP genes in cucumber (Cucumis sativus L.). In this study, we identified 10 CsZIP genes from the pan-genome of 13 C. sativus accessions. Among them, only CsZIP10 showed no variation in protein sequence length. We analyzed the gene structure, conserved domains, promoter cis-elements, and phylogenetic relationships of these 10 CsZIP genes derived from "9930". Based on phylogenetic analysis, the CsZIP genes were classified into three branches. Amino acid sequence comparison revealed the presence of conserved histidine residues in the ZIP proteins. Analysis of promoter cis-elements showed that most promoters contained elements responsive to plant hormones. Expression profiling in different tissues showed that most CsZIP genes were expressed at relatively low levels in C. sativus leave, stem, and tendril, and CsZIP7 and CsZIP10 were specifically expressed in root, indicating their potential involvement in the absorption and transport of metal ions. Transcriptomic data indicated that these 10 ZIP genes displayed responses to both downy mildew and powdery mildew, and CsZIP1 were significantly downregulated after both salt and heat treatments. In conclusion, this study deepens our understanding of the ZIP gene family and enhances our knowledge of the biological functions of CsZIP genes in C. sativus.
Keywords:
pan-genome
; Cucumis sativus
; CsZIP
1. Introduction
The homeostasis of metal ions plays a crucial role in the growth and development of plants. Appropriate concentrations of metal ions are required by plants to sustain biochemical reactions and physiological processes within cells. However, an excess or deficiency of metal ions may lead to severe growth issues and physiological disruptions. Therefore, maintaining suitable concentrations of metal ions is vital for the survival of plants. The ZIP gene family performs a significant role in the metal ion homeostasis of plants. This family encodes zinc ion transport proteins responsible for regulating the absorption and distribution of zinc as well as other metal ions. Through the activity of these proteins, plants can respond to changes in environmental metal ion levels and maintain a balanced state of metal ions within cells [1,3]. Most ZIP proteins consist of 309–476 amino acid residues and possess eight putative transmembrane (TM) domains. They exhibit a common membrane topology, with the N- and C-terminal ends located extracytoplasmically [2]. The cytoplasmic loop between TM3 and TM4 contains a histidine-rich domain (HRD), which functions as the metal binding domain involved in metal transport. Additionally, the amphipathic nature of TM4 and TM5 creates cavities that allow for the passage of metal ions [2,3].
Currently, more than 300 members of the ZIP protein family have been identified from plants, and some ZIP members have been functionally characterized. The ZIP gene, AtIRT1, was first identified in Arabidopsis thaliana. It encodes an iron transporter protein and is mainly expressed in the roots [4]. The previous study has shown that AtIRT1 also exhibits specific expression in the companion cells of the cortex and plays a role in iron transport in aerial organs [5]. AtIRT2 is another IRT gene which functions similarly to AtIRT1 in A. thaliana. AtIRT2 is localized to the vacuolar membrane and compartmentalizes Fe into vacuoles to prevent its toxicity in the cytoplasmic compartment [6]. AtZIP1 and AtZIP3 are expressed in the roots. AtZIP1 is a vacuolar transporter protein responsible for the reactivation of Mn/Zn from vacuoles to cytoplasm in root cells [7]. AtZIP3 plays a role in the uptake of Zn and Fe from the soil to the plant roots [3].
AtZIP2 mediates Mn/Zn uptake into parenchyma cells in the xylem, thereby facilitating the transport of Mn/Zn towards the shoots [7]. In contrast, AtZIP4 transports zinc ions into cells or between plant tissues [3]. In addition, some ZIP transporter proteins directly participate in the accumulation of Zn in edible plant parts [8,9]. The expression levels of ZIP family genes in plants show dynamic changes. For example, under zinc-deficient conditions, some ZIP transporter proteins are expressed at higher levels, but when zinc levels return to normal or high, their expression decreases [10].
In rice (Oryza sativa L.), OsIRT1 and OsIRT2 are iron transport proteins that directly absorb Fe2+ and Zn2+ into root cells [11,12,13,14]. When O. sativa is exposed to iron deficiency conditions, the expression of OsIRT1 and OsIRT2 in the root system is upregulated [15]. OsZIP1 is localized in the endoplasmic reticulum and plasma membrane, and it functions as a metal ion transporter in O. sativa under conditions of metal ion excess [16]. Furthermore, studies have shown that overexpression of OsZIP1 enhances the accumulation of Zn and Fe in roots, shoots, and seeds [13,14]. OsZIP4 and OsZIP5, as plasma membrane-localized Zn transporters, participate in Zn2+ uptake in O. sativa [17,18]. Overexpression of OsZIP4 and OsZIP5 can increase the Zn2+ concentration in plant roots [17,18]. OsZIP8 is also a Zn transporter protein in O. sativa, participating in the uptake and distribution of Zn2+ [19]. Generally speaking, ZIP transport proteins are primarily involved in the absorption and transport of Zn and Fe. However, these proteins are also involved in the transport and uptake of other metal ions, such as Mn and Cu [7].
Additionally, they could also participate in the absorption, transport, and accumulation of various toxic heavy metals, such as cadmium (Cd), in plants [20]. For example, overexpression of OsIRT1 in O. sativa has been found to enhance its resistance to Cd [21]. ZIP2 and ZIP3 have been demonstrated the involvement in the uptake and transport of Cd in cabbage, with an increase in their expression levels [22]. In tomato (Solanum lycopersicum), the expression of SlZIP4 is positively correlated with Cd concentration within a certain range [23]. Additionally, ZIP genes also participate in the absorption and transport of Cd in mulberry and tobacco [24,25].
Previous studies have reported the genome-wide identification of the ZIP gene family in various plant species including A. thaliana, peanut, and Populus trichocarpa . Arabidopsis had 15 ZIP members, peanut 30, and Populus.trichocarpa 16, respectively [45,46,47]. However, there is no report on the CsZIP gene family yet in C. sativus . Therefore, it is necessary to identify the CsZIP gene family.
C. sativus is a popular cash crop due to its unique flavor and refreshing crisp texture [26]. Due to the crucial role of the CsZIP gene family in the absorption and transportation of metal ions, it is imperative to identify and characterize the CsZIP gene family in C. sativus. The updated genome database of C. sativus provides a better resource for identifying and characterizing CsZIP genes. In this study, we used bioinformatics methods to investigate the sequence similarity, gene structure, and expression patterns of the CsZIP gene family, with the aim of clarifying the phylogenetic relationship of the ZIP gene family, deepening our understanding of CsZIP genes, and improving our knowledge of their biological functions.
2. Materials and Methods
2.1. Identification of ZIP genes in C. sativus
The identification and analysis of C. sativus ZIP genes were performed using genome sequence data of C. sativus deposited at Cucurbit Genomics Data website (http://cucurbitgenomics.org/organism/20). The hidden Markov model (HMM) profile files of the ZIP conserved domain (PF02535) were downloaded from the Pfam database (http://pfam.xfam.org/). The ZIP genes of C. sativus were identified from the genome database using HMMER 3.0 with the default parameters and a cutoff value of 0.01. The Pfam (http://pfam.janelia.org)and SMART (http://smart.embl-heidelberg.de/) tools were used to confirm the ZIP conserved domain. ZIP candidates were predicted using the ExPASy Proteomics Server (http://expasy.org/)(Artimo et al. 2012), and their subcellular localization was investigated using Cell-PLoc (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/) [27].
2.2. Gene structure and motif analysis
The CDS sequences and genomic of CsZIP genes were downloaded from the C. sativus genome database (http://cucurbitgenomics.org/organism/20), and the gene structures were visualized using the Gene Structure Display Server online tool (http://gsds.cbi.pku.edu.cn/) [28]. The conserved motifs of CsZIP proteins were then identified with MEME 4.9.1 (http://meme-suite. org/) [29] and visualized with WebLogo (http://weblogo.berkeley.edu/logo.cgi) [30]. The total number of motifs ( nmotifs ) is 10, the minimum length of motifs ( minw ) is 6 amino acids, and the maximum length of motifs ( maxw ) is 10 amino acids.
2.3. Phylogenetic analysis and multiple sequence alignment
The MEGA software (v7.0) was utilized to align the protein sequences of ZIP from A. thaliana, C. sativus, melon, and O. sativa using the ClustalW algorithm. Subsequently, an analysis of the evolutionary relationships among the ZIP proteins was conducted employing the neighbor-joining method with 1000 bootstrap replicates. The resulting phylogenetic trees underwent graphical representation and enhancement through Evolview (https://evolgenius.info//evolview-v2/#login).
2.4. Gene duplication analysis and genome distribution
CsZIP loci were extracted from the C. sativus genome database (http://cucurbitgenomics.org/organism/20) and their locations on chromosomes were visualized using MapChart software [31]. From Orthomcl [32], the homology of ZIP gene between C. sativus and A. thaliana, Cucumis melo and O. sativa were obtained. Circos shows the chromosomal location and collinearity of the ZIP gene [33]. All CDS between Arabidopsis and C. sativus genomes are compared by Clustal W. Then, repeated pairs and homologous pairs are identified based on the alignment result. The Computational Biology Unit (CBU) Ka/Ks calculation tool (http://services.cbu.uib.no/tools/kaks) was used to calculate the non-synonymous (Ka) and synonymous (Ks) nucleotide substitution rates. Then, the formula T =Ks/2r Mya (millions of years) was used to calculate the divergence times of the duplicated genes, where r is equal to 1.5 x10−8 [34].
2.5. Analysis of promoter regions of CsZIP genes
The 1.5-kb sequences upstream of the initiation codons (ATG) of CsZIP genes were downloaded from the cucurbit genomics data website (http://cucurbitgenomics. org/organism/20), and analyzed for cis-elements in the promoter region using the online tool PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) [35].
2.6. Analysis of transcriptome data
The data from different C. sativus RNA seq libraries were downloaded from the Cucurbit Genomics Data website (http://cucurbitgenomics.org/organime\/20) and NCBI SAR databases (accession Nos.:PRJNA388584,GSE151055,GSE81234 and GSE11265). The expression profile of CsZIP gene was visualized using TBtools software [36].
2.7. Plant materials and growth conditions
In this study, the temporal and spatial expression patterns of ZIP genes were analyzed using the C. sativus inbred line 'China long' 9930 (North China type). C. sativus seedlings were grown in greenhouses at Shandong Agricultural University in China, following standard water management and pest control practices.
2.8. RNA isolation and real-time PCR analysis
To collect samples for expression analysis, the roots, stems, leaves, female and male flowers, ovaries, and tendrils of C. sativus plants were collected. These samples were immediately frozen in liquid nitrogen and stored at -80 ℃ for RNA isolation. Total RNAwas extracted with TRIzol (Thermo Fisher Scientific, USA) by following the manufacturer’sinstructions and then treated with DNase I (Thermo Fisher Scientific, USA) to remove the possible DNA contamination. cDNA synthesiswas carried out with a RevertAid First Strand Synthesis Kit (Thermo Fisher Scientific, USA). Primers were designed for qRT-PCR verification of the select genes with Primer Premier 5.0 (Table S4) and C. sativus actin gene was used as an internal control. qRT-PCR for the select genes was performed with an Ultra SYBR Green Mixture qPCRkit (CWBIO, China) on an iCycler iQ™ real-time PCR detection system (Bio-Rad, USA) by following the PCR program of 95°C for 30 s, 40 cycles of 95°C for 15 s and 60°C for 15 s, and finally 72°C for 30 s. Relative expression levels of the select genes were measured using the 2−∆∆Ct method [37].
3. Results
3.1. Identification of CsZIP genes from C. sativus pan-genome
With the release of a graph-based pan-genome [38], we could identify ZIP genes in C. sativus by analyzing different accession genomes. We obtained the HMM profile files and extracted CsZIP proteins from the C. sativus genome database. The candidate genes were further analyzed using NCBI CDD, SMART, and Pfam. As a result, we identified a total of 10 CsZIP genes in the 13 C. sativus accessions (Table 1). Of these accessions, only 'Cuc80' and 'PI183967' contain nine CsZIP genes (Table 2). To avoid confusion, we renamed these genes as CsZIP1-CsZIP10 based on their chromosomal order (Table 3).
To further investigate the variation of CsZIP genes among different C. sativus accessions, we counted the length of the identified CsZIP proteins (Table 2). Among the 13 C. sativus accessions, only one gene, CsZIP10, had the same protein length across all accessions. The lengths of CsZIP1, CsZIP4, CsZIP6 and CsZIP8 differed in only one accession. CsZIP2, CsZIP3 and CsZIP7 exhibited variations in protein length in two accessions. CsZIP9 showed protein variations in three accessions, while CsZIP5 exhibited protein length differences in multiple accessions. All CsZIP protein sequences can be found in Dataset S2.
3.2. Characterization of CsZIP genes from Chinese long 9930
Subsequent analyses primarily focused on the genes identified in the '9930' genome, as it encompassed all the 10 CsZIPs and is the first C. sativus genome sequenced and has been updated to the V3 version. The gene name, gene number, gene intron number, chromosome position, gene length, protein length, isoelectric point, relative molecular weight, and subcellular localization prediction of CsZIPs are presented in Table 3. Sequence analysis revealed that the length of CsZIP proteins ranged from 228 to 594 amino acids, with molecular weights ranging from 27.20 to 61.98 kDa. The longest and most complex gene structure was found in CsZIP6, while the shortest gene was CsZIP8. CsZIP4 had the highest relative molecular weight, whereas CsZIP3 the lowest. The isoelectric points of these proteins ranged from 5.58 to 8.53. Furthermore, subcellular localization prediction indicated that most CsZIP proteins were localized to the cell membrane, except CsZIP2 and CsZIP7, which were found in the chloroplasts (Table 3).

Table 1.
The origins of the different C. sativus accessions.
| Accession name | Accession group | Accession name | Accession group |
|---|---|---|---|
| 9930 | East Asian cultivated accession | Hx117 | Indian cultivated accession |
| XTMC | East Asian cultivated accession | Hx14 | Indian cultivated accession |
| Cu2 | East Asian cultivated accession | W4 | Indian wild accession |
| Cuc37 | Eurasian cultivated accession | W8 | Indian wild accession |
| Gy14 | Eurasian cultivated accession | Cuc64 | Indian wild accession |
| 9110gt | Eurasian cultivated accession | PI183967 | Indian wild accession |
| Cuc80 | Xishuangbanna cultivated accession |
Table 2.
The protein lengths of CsZIPs in different C. sativus accessions.
| Protein name | 9930 | XTMC | Cu2 | Cuc80 | Cuc37 | Gy14 | 9110gt | PI183967 | Cuc64 | W4 | W8 | Hx14 | Hx117 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CsZIP1 | 249 | 350 | 350 | - | 350 | 350 | 350 | 350 | 350 | 350 | 350 | 350 | 350 | |
| CsZIP2 | 417 | 417 | 417 | 417 | 417 | 441 | 417 | 417 | 417 | 417 | 417 | 408 | 417 | |
| CsZIP3 | 228 | 275 | 275 | 275 | 275 | 306 | 275 | 275 | 306 | 275 | 275 | 275 | 275 | |
| CsZIP4 | 594 | 594 | 639 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | 594 | |
| CsZIP5 | 367 | 367 | 367 | 365 | 367 | 367 | 367 | 367 | 369 | 367 | 365 | 369 | 815 | |
| CsZIP6 | 463 | 463 | 463 | 463 | 463 | 407 | 463 | 463 | 463 | 463 | 463 | 463 | 463 | |
| CsZIP7 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 335 | 302 | 335 | 335 | 346 | |
| CsZIP8 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 349 | 1096 | |
| CsZIP9 | 334 | 334 | 334 | 334 | 334 | 337 | 334 | - | 338 | 334 | 334 | 334 | 337 | |
| CsZIP10 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 | 354 |
Table 3.
CsZIP family in C. sativus.
| Gene | Introns | Chromosomal site | Gene length | protein length | Protein MW(kDa) | Isoelectric point | Subcellular location predicted |
|---|---|---|---|---|---|---|---|
| CsZIP1 | 2 | 1 | 2955 | 249 | 27.2 | 6.88 | Cell membrane |
| CsZIP2 | 3 | 2 | 3491 | 417 | 44.5 | 6.09 | Chloroplast |
| CsZIP3 | 9 | 4 | 9406 | 228 | 24.6 | 7.02 | Cell membrane |
| CsZIP4 | 4 | 4 | 5063 | 594 | 62.0 | 6.27 | Cell membrane |
| CsZIP5 | 2 | 4 | 2777 | 367 | 38.8 | 5.98 | Cell membrane |
| CsZIP6 | 10 | 5 | 21173 | 463 | 49.9 | 5.97 | Cell membrane |
| CsZIP7 | 1 | 6 | 2038 | 334 | 35.7 | 5.73 | Chloroplast |
| CsZIP8 | 0 | 6 | 1490 | 348 | 38.0 | 5.58 | Cell membrane |
| CsZIP9 | 2 | 7 | 7007 | 334 | 35.9 | 6.02 | Cell membrane |
| CsZIP10 | 2 | 7 | 4076 | 354 | 37.2 | 8.53 | Cell membrane |
3.3. Analysis of phylogenetic relationship and gene structure and protein motif
To analyze the phylogenetic relationship of CsZIP proteins, we constructed the phylogenetic tree of these 10 proteins (Figure 1). Based on the phylogenetic tree, the CsZIP family was divided into three distinct clades. CsZIP2, CsZIP5 and CsZIP10 belonged to the first clade, while CsZIP8 and CsZIP1 occupied the second clade, and the other CsZIPs formed the third clade (Figure 1). The number of introns varied among CsZIP genes, ranging from 0 to 10: CsZIP6 has the most introns, and CsZIP8 has none (Figure 1b). The structures of CsZIP6, CsZIP3 and CsZIP4 genes were similar, suggesting that they are more closely related (Figure 1).
We further analyzed the conserved motifs of CsZIP family and found that motif type and arrangement were very similar among the members of the first and second clades, in all of which only motif9 was present; but diverse among the members of the third clade, and motif number ranged from one (CsZIP3 and CsZIP4) to seven (CsZIP7) (Figure 2). Motif 2 was present in all the CsZIPs except CsZIP3 indicating that Motif 2 is highly conserved (Figure 2). CsZIP6, CsZIP3 and CsZIP4 are the CsZIP proteins that did not contain Motif 1 (Figure 2). The differences in motifs among CsZIP proteins may contribute to their functional differentiation. The amino acid sequence for each motif is shown in the Figure S1.
3.4. Multiple sequence alignment
To analyze the characteristics of ZIP protein homologous sequences in comparison to other species, we conducted a comparative study of ZIP proteins in Arabidopsis, Cucumis melo, O. sativa, and C. sativus. The transmembrane domains IV-VII exhibited the highest degree of conservation (Figure 3). Previous studies have reported that transmembrane domains IV and V are amphiphilic and play a crucial role in transport, a conclusion that has been further corroborated by subsequent research [42]. In the mutant irt1, mutations in the conserved histidine residues or adjacent polar charged residues within transmembrane domains IV and V were found to abolish their transport function [39]. Notably, the IV regions of the sequence alignment in Arabidopsis, Cucumis melo , O. sativa, and C. sativus showed the presence of conserved histidine residues (Figure 3).
3.5. Phylogenetic analysis among different species
We obtained a comprehensive understanding of the relative relationship of CsZIP proteins by constructing a phylogenetic tree using the amino acid sequences of ZIP proteins from Arabidopsis, C. sativus, Cucumis melo, and O. sativa (Figure 4). The phylogenetic analysis revealed three major clades: Group 1, 2 and 3. In Group 1, there were 7 proteins from Arabidopsis, 3 from C. sativus, 3 from Cucumis melo, and 8 from O. sativa. Group 2 consisted of 6 proteins from Arabidopsis, 3 from C. sativus, 3 from Cucumis melo, and 3 from O. sativa. Group 3 included 5 proteins from Arabidopsis, 4 from C. sativus, 4 from Cucumis melo, and 5 from O. sativa (Figure 4). It is worth noting that the number of ZIP proteins varied within each subgroup for Arabidopsis, C. sativus, Cucumis melo, and O. sativa. Specifically, half O. sativa ZIP proteins were grouped into Group 1, while the distribution among the three clades was relatively equal for the other species. Further investigation within the Group 3 revealed a close relationship between CsZIP6, CmZIP8, AtIAR1, and OsZIP14 (Figure 4). For reference, the amino acid sequences of the ZIP proteins can be found in the Dataset S1. This analysis provides valuable insights into the interrelationships of ZIP proteins in different plant species and serves as a foundation for further exploration of their biological functions.
3.6. Chromosome localization and collinearity analysis
The chromosomal distribution analysis of CsZIP genes in C. sativus revealed an uneven distribution pattern across seven chromosomes (Figure 5). Specifically, three ZIP genes (CsZIP3, 4 and 5) were located on chromosome 4, two on each of chromosome 6 and 7, respectively, and one on each of chromosome 1, 2 and 5, respectively, and none on chromosome 3 (Figure 5).
We further conducted a collinearity analysis of the ZIP gene family members between C. sativus and other plant species, including Arabidopsis, Cucumis melo , O. sativa, and Chinese cabbage (Figure 6). Our results revealed that a total of eight genes were identified to be present in both C. sativus and Arabidopsis, suggesting a conserved relationship between these two species (Figure 6). In the case of C. sativus and Cucumis melo, all members of the ZIP gene family exhibited homologous alignment, indicating a high level of homology during the long differentiation process between these two closely related species (Figure 6). However, only two genes showed homology between C. sativus and O. sativa. Similarly, we detected eight homologous genes in both C. sativus and Chinese cabbage, similar with the findings between C. sativus and Arabidopsis (Figure 6). These findings provide valuable insights into the evolutionary relationships and conservation of the ZIP gene family among different plant species.
3.7. Analysis of cis-acting elements in promoter region of CsZIP genes
The analysis of the 1500 bp sequence upstream of the ZIP gene in C. sativus revealed the presence of various cis-elements (Figure 7). Specifically, the promoter regions of all the genes in this family contained elements related to some of gibberellin, auxin, salicylic acid, jasmonic acid, low temperature, and drought stress. Most of the gene promoters contained hormone-related elements, indicating their potential involvement in hormone signaling pathways. Notably, gibberellin-related elements were found in the promoters of several family members, including CsZIP1, CsZIP2, CsZIP4, CsZIP6, CsZIP7, CsZIP8 and CsZIP10 (Figure 7). This suggests that these genes may play crucial roles in gibberellin-mediated processes. Auxin response elements were predominantly enriched in CsZIP6, CsZIP8 and CsZIP10 gene promoters (Figure 7). This finding suggests that these specific genes may be involved in the regulation of plant growth and development through auxin signaling pathways.
3.8. Expression of CsZIP genes in different tissues
This part aimed to investigate the roles of ZIP genes in C. sativus development by analyzing RNA-seq data from different tissues. The expression patterns of CsZIP gene family were examined using data from the Cucurbit Genomics Data website and NCBI SAR database, and heat maps were generated to visualize the expression levels across tissues. The results revealed that the expression of ZIP genes in C. sativus leaves, stems, and tendrils was relatively lower compared to other tissues (Figure 8a). In contrast, CsZIP7 and CsZIP10 showed the highest expression levels in roots, suggesting their possible involvement in regulating ion transport in the underground parts of C. sativus. CsZIP2, CsZIP5 and CsZIP9 were predominantly expressed in flowers, indicating their potential roles in flower differentiation and development (Figure 8a). On the other hand, CsZIP1, CsZIP3, CsZIP4, CsZIP6 and CsZIP8 exhibited major expression in the ovary, especially,CsZIP3, CsZIP4 and CsZIP8 displayed high expression levels in the unfertilized ovary (Figure 8a). These findings suggest that these five ZIP genes may have regulatory functions in fruit development.
To further validate the reliability of the RNA-seq results, qRT-PCR analysis of ZIP gene expression in different tissues was performed. Overall, the qRT-PCR results were consistent with the RNA-seq data, confirming the reliability of the datasets (Figure 8b). However, an inconsistency was observed in the expression of CsZIP4 between male flowers and tendrils, indicating some variations in gene expression in specific tissues (Figure 8b). Therefore, these genes are likely involved in ion transport, flower differentiation and development, and potentially regulate fruit development.
3.9. Expression profiles of CsZIP genes under abiotic and biotic stresses
Due to the lack of transcriptomic data on metal ion transport, and in order to enhance our understanding of the functional roles within the ZIP gene family and predict their potential applications in the future, We investigate the impact of various environmental stresses on the expression of CsZIP genes, we assessed their comprehensive expression patterns in response to different stress conditions, including salt, heat, downy mildew (DM, caused by Pseudoperonospora cubensis), and powdery mildew (PM, caused by Podosphaera fusca), based on publicly available transcriptome data. This analysis enabled us to gain further insights into the potential roles of CsZIP genes in mediating plant response to diverse stresses.
Under salt stress, specific expression patterns of CsZIP genes were revealed through transcriptome data analysis. CsZIP4, CsZIP5, CsZIP6 and CsZIP8 genes were up-regulated in response to NaCl treatment; In contrast, CsZIP1, CsZIP3 and CsZIP10 genes showed down-regulation (Figure 9a). It is worth noting that Si treatment has been reported to enhance stress resistance and stimulate plant growth [66]. Interestingly, when plants were solely subjected to Si treatment, most CsZIP genes exhibited up-regulation in their expression levels. However, the expression of the CsZIP1 gene was found to be down-regulated (Figure 9a). Additionally, we observed a similar down-regulation of CsZIP1 gene expression after exogenous salt and Si treatment. Therefore, it can be concluded that under salt stress conditions, most CsZIP genes tend to be up-regulated, while the CsZIP1 gene specifically demonstrates down-regulation in response to salt stress (Figure 9a).
We also examined the response of CsZIP genes to heat stress (Figure 9b). At 3 hours following exposure to high temperatures, the expression levels of CsZIP2, CsZIP3, CsZIP4, CsZIP5, CsZIP6, CsZIP7, CsZIP8 and CsZIP9 were up-regulated, whereas CsZIP1 was down-regulated, and similar patterns emerged at 6 hours after heat stress, suggesting that these genes may play a crucial role in heat tolerance (Figure 9b).
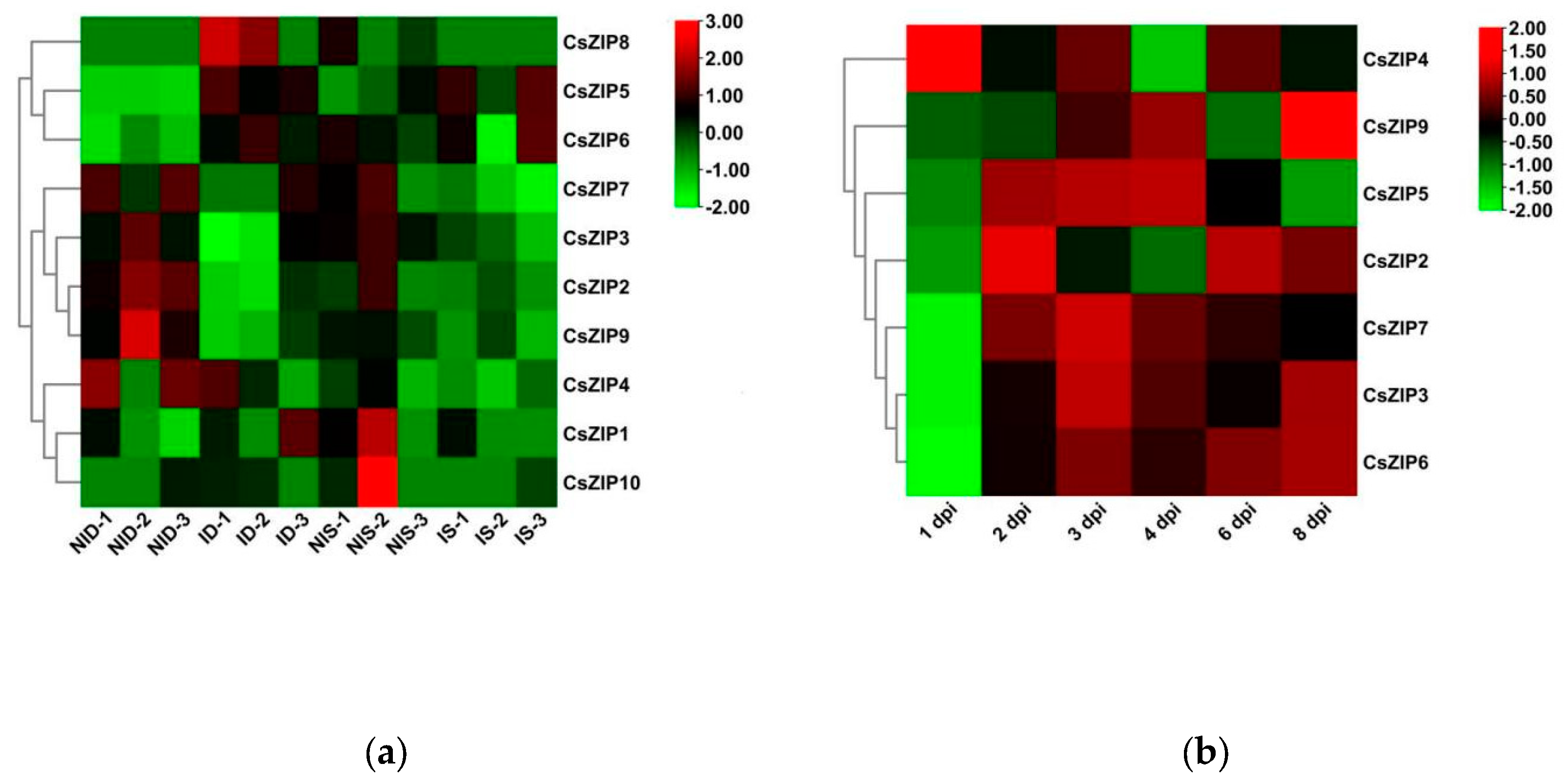
To explore the potential functions of CsZIPs in biotic stress resistance, we conducted an analysis of CsZIP expression using the RNA-Seq database. Following inoculation with powdery mildew (PM), we observed distinct gene expression patterns between the susceptible and resistant C. sativus lines (Figure 10a). After inoculation with PM, the expression of CsZIP3, CsZIP4, CsZIP7 and CsZIP9 was down-regulated in both the resistant and susceptible lines. In contrast, the expression of CsZIP5 was up-regulated (Figure 10a).
For DM inoculation, CsZIP2, CsZIP3, CsZIP5, CsZIP6, CsZIP7 and CsZIP9 genes were up-regulated at least at one time point after treatment, while CsZIP4 genes were down-regulated (Figure 10b).
These findings indicate that CsZIP genes are responsive to different environmental stresses and may play diverse roles in mediating plant responses to these stresses. Further research is needed to fully understand the specific functions of each CsZIP gene and their contributions to stress resistance in C. sativus.
4. Discussion
The ZIP (ZRT/IRT-like protein) gene family is involved in the absorption and transport of metal ions in plants, playing a crucial role in plant growth, development, and response to heavy metal stress [1]. In this study, a comprehensive genome-wide identification of CsZIPs was performed, and a total of 10 genes were identified in C. sativus. We further investigated their gene structure, phylogenetic relationship, composition of cis-regulatory elements in promoter, chromosome localization, collinearity analysis, and expression patterns under different elemental stresses. These findings enhance our understanding of the ZIP gene family and provide bases for better elucidating the function and evolutionary relationship of CsZIPs in C. sativus.
Previous studies have reported the identification of the ZIP gene family in various plant species including A. thaliana, peanut, and Populus.trichocarpa. Arabidopsis had 15 ZIP members, peanut 30, and P.trichocarpa 16, respectively [40,41,42]. However, in this study, only 10 CsZIP genes were identified (Figure 1). Of the 13 C. sativus germplasm resources analyzed, 'Cuc80' and 'PI183967' contained 9 CsZIP genes, whereas the remaining varieties harbored 10 ZIP genes (Table 2). Notably, only CsZIP10 exhibited a consistent amino acid sequence length, while the lengths of other CsZIPs varied among the different varieties (Table 2).
Previous studies have predicted and confirmed the subcellular localization of ZIP proteins. Most of ZIP proteins are predicted to be localized in membrane system. For instance, in O. sativa, it has been demonstrated that OsZIP1, OsZIP5, OsZIP7 and OsZIP8 are localized in the plasma membrane [17,21,22,51,52]. In peanuts, AhZIP1.2, AhZIP3.2, AhZIP3.5 and AhZIP7.8 are localized in the endomembrane system [42]. In Arabidopsis, AtIRT1, AtIRT3, AtZIP1 and AtZIP2 have been confirmed to be localized in the plasma membrane [6,7,53]. In this study, we predicted the subcellular localization of 10 ZIP proteins. The results showed that CsZIP1, CsZIP3, CsZIP4, CsZIP5, CsZIP6, CsZIP8, CsZIP9 and CsZIP10 are predicted to be localized in the cell membrane, while CsZIP2 and CsZIP7 in the chloroplast (Table 3).
In the evolutionary process of plant genomes, the emergence of gene family members often accompanies gene duplication, leading to new functions. For example, most of the AhZIP genes experienced gene duplication events except AhIRT1.1/1.3, AhZIP7.2/7.8 and AhZIP3.5/3.6 [42]. However, unfortunately, we did not detect any gene duplication events in the CsZIP genes in C. sativus.
The CsZIP gene promoters contain many cis-acting elements that respond to various hormones and stresses, including gibberellins, methyl jasmonate, abscisic acid, and low temperature. In S. lycopersicum, zinc deficiency tolerance is determined by auxin signaling [46]. Furthermore, the response elements of three ZIP genes in sunflower are regulated by salicylic acid and methyl jasmonate under iron deficiency [47]. These findings suggest that the expression of CsZIP genes can be induced by several plant hormones, such as salicylic acid, MeJA, gibberellins and auxins. Methyl jasmonate acts as a crucial plant hormone in defense against biotic and abiotic stresses by triggering defense mechanisms and regulating growth [48]. Auxin, on the other hand, controls plant development, with tryptophan being a key amino acid in the auxin synthesis pathway. As Zn participates in tryptophan synthesis, CsZIP genes might play a significant role in auxin biosynthesis [49]. Being similar to auxin, gibberellins also regulate various plant growth and development processes, including seed germination and fruit development [50]. Salicylic acid has been shown to greatly enhance plant stress resistance [51]. Additionally, several cis-acting elements associated with biotic and abiotic stress responses have been identified in the promoter region of CsZIP genes (Figure 7). This suggests that the CsZIP gene family may regulate plant growth in hormone signaling pathways to adapt to different environmental stresses.
In general, the expression patterns of genes are correlated with their functions [52]. The expression data of CsZIPs in different tissues provide valuable insights into the possible functions of the ZIP gene family. Previous studies have confirmed that ZIP genes are predominantly expressed in roots, regardless of the plant species, such as Arabidopsis, O. sativa, or barley, with many genes being induced under deficiencies of environmental zinc or other metallic elements. [7,53,54,55]. Existing research has indicated that within tomatoes, the SlZIP gene family comprises 15 members, with 13 expressing in the roots, exhibiting significantly high expression levels. Among these, 11 SlZIP genes are expressed in fruits, albeit with varying expression across different developmental stages, indicating the involvement of SlZIP genes in the accumulation of zinc and iron in S lycopersicum fruits or their redistribution among different tissues [56]. Similarly, CsZIP7 and CsZIP10 exhibit high expression in C. sativus roots, while CsZIP10 has higher expression levels in stems compared to other tissues (Figure 8). This suggests that CsZIP7 and CsZIP10 may play functional roles in these two tissues. Furthermore, it was observed that CsZIP2, CsZIP5 and CsZIP9 show higher expression levels in flowers, while CsZIP3, CsZIP4 and CsZIP8 exhibit the highest expression levels in fertilized ovaries. CsZIP1 and CsZIP6 are also expressed in ovaries (Figure 8). Based on this information, it can be inferred that these CsZIP genes may participate in the regulation of fruit development by regulating the accumulation of Zn and Fe or their redistribution among different tissues.
Previous studies have primarily focused on the regulatory roles of ZIP genes in plant metal ion uptake and transport. For instance, AtZIP3 is known to play a crucial role in facilitating the absorption of Zn and Fe from the soil into plant roots [8]. On the other hand, AtZIP2 functions by mediating the uptake of Mn/Zn into the parenchyma cells of the xylem, thereby enabling the efficient transportation of Mn/Zn to the aboveground parts of plants [7]. However, in this study, we found that CsZIP genes could response to different environmental stresses. We investigated the comprehensive expression patterns of CsZIP genes under different environmental stresses, including powdery mildew, downy mildew, salt, and heat. The results showed differential expression of CsZIP genes in response to high temperature (Figure 9b), salt, silicon (Figure 9a), powdery mildew (Figure 10a), and downy mildew treatments (Figure 10a). These results provide valuable clues suggesting that CsZIP genes may have significant functions in multiple stress conditions, warranting further research and validation of their specific roles.
5. Conclusions
In this study, we performed pan-genome-wide identification of the ZIP gene family in C. sativus. A total of 10 members were identified. Eleven of the 13 accessions contained all the CsZIP genes. The ZIP gene family has three evolutionary branches and contains conserved histidine residues. The CsZIP gene promoters contained elements that responded to plant hormone signaling pathways, indicating that plant hormone signals may have an impact on CsZIP-mediated biotic and abiotic stresses. The expression patterns of CsZIPs in different tissues showed that the CsZIP gene family may be related to the growth and development of C. sativus. Finally, CsZIPs could response to different environmental stresses. In summary, this study helps to improve the current understanding of the CsZIP gene family and provides a basis for exploring their functions in C. sativus.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: The conserved motif LOGO of C. sativus ZIP proteins; Table S1: The FPKM values of CsZIP genes; Table S2: The FPKM value of CsZIP genes under salt and hot treatments; Table S3: The FPKM value of CsZIP genes under powdery mildew and downy mildew; Table S4: The primer sequences used for the qRT-PCR; Dataset S1: The amino acid sequences of the ZIP proteins. Dataset S2:All CsZIP protein sequences of 13 C. sativus accessions.
Author Contributions
Conceptualization, Z.R.; methodology, Z.W. and M.Y.; software, Z.W., X.W., J.C.; validation, Z.W.; formal analysis, Z.W., M.Y. and C.C.; investigation, Z.W. and C.C.; resources, Z.R. and L.W.; data curation, Z.W.; writing—original draft preparation, Z.W., and Z.R.; writing—review and editing, L.W. and Z.R.; visualization, L.W. and Z.R.; supervision, L.W. and Z.R.; project administration, L.W. and Z.R.; funding acquisition, Z.W. and Z.R.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China ( 31972419 and 32172605), the Agricultural Variety Improvement Project of Shandong Province (2022LZGCQY001) and the ‘Taishan Scholar’ Foundation of the People’s Government of Shandong Province (ts20130932).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and Supplementary Materials.
Acknowledgments
We extend our appreciation to the anonymous reviewers for their valuable suggestions to help improve this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Grotz N, Guerinot ML. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim Biophys Acta 2006 Jul;1763(7):595-608. [CrossRef]
- Gaither LA, Eide DJ. Eukaryotic zinc transporters and their regulation. Biometals 2001 Sep-Dec;14(3-4):251-70. [CrossRef]
- Guerinot, ML. The ZIP family of metal transporters. Biochim Biophys Acta 2000 May 1;1465(1-2):190-8. [CrossRef]
- Nishida S, Tsuzuki C, Kato A, Aisu A, Yoshida J, Mizuno T. AtIRT1, the primary iron uptake transporter in the root, mediates excess nickel accumulation in Arabidopsis thaliana. Plant Cell Physiol 2011 Aug;52(8):1433-42. [CrossRef]
- Krausko M, Labajová M, Peterková D, Jásik J. Specific expression of AtIRT1 in phloem companion cells suggests its role in iron translocation in aboveground plant organs. Plant Signal Behav 2021 Sep 2;16(9):1925020. [CrossRef]
- Vert G, Barberon M, Zelazny E, Séguéla M, Briat JF, Curie C. Arabidopsis IRT2 cooperates with the high-affinity iron uptake system to maintain iron homeostasis in root epidermal cells. Planta 2009 May;229(6):1171-9. [CrossRef]
- Milner MJ, Seamon J, Craft E, Kochian LV. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J Exp Bot 2013 Jan;64(1):369-81. [CrossRef]
- Gaitán-Solís E, Taylor NJ, Siritunga D, Stevens W, Schachtman DP. Overexpression of the transporters AtZIP1 and AtMTP1 in cassava changes zinc accumulation and partitioning. Front Plant Sci 2015 Jul 9;6:492. [CrossRef]
- Ramegowda, Y. , Venkategowda, R., Jagadish, P. et al. Expression of a rice Zn transporter, OsZIP1, increases Zn concentration in tobacco and finger millet transgenic plants. Plant Biotechnol Rep 2013, 7, 309–319. [Google Scholar] [CrossRef]
- van de Mortel JE, Almar Villanueva L, Schat H, Kwekkeboom J, Coughlan S, Moerland PD, Ver Loren van Themaat E, Koornneef M, Aarts MG. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2006 Nov;142(3):1127-47. [CrossRef]
- Itai RN, Ogo Y, Kobayashi T, Nakanishi H, Nishizawa NK. rice genes involved in phytosiderophore biosynthesis are synchronously regulated during the early stages of iron deficiency in roots. rice (N Y) 2013 Jun 25;6(1):16. [CrossRef]
- Ishimaru Y, Suzuki M, Tsukamoto T, Suzuki K, Nakazono M, Kobayashi T, Wada Y, Watanabe S, Matsuhashi S, Takahashi M, Nakanishi H, Mori S, Nishizawa NK. rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J 2006 Feb;45(3):335-46. [CrossRef]
- Lee S, An G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ 2009 Apr;32(4):408-16. [CrossRef]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Ishimaru Y, Kim S, Tsukamoto T, Oki H, Kobayashi T, Watanabe S, Matsuhashi S, Takahashi M, Nakanishi H, Mori S, Nishizawa NK. Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil. Proc Natl Acad Sci U S A 2007 May 1;104(18):7373-8. [CrossRef]
- Liu XS, Feng SJ, Zhang BQ, Wang MQ, Cao HW, Rono JK, Chen X, Yang ZM. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol 2019 Jun 27;19(1):283. [CrossRef]
- shimaru Y, Masuda H, Suzuki M, Bashir K, Takahashi M, Nakanishi H, Mori S, Nishizawa NK. Overexpression of the OsZIP4 zinc transporter confers disarrangement of zinc distribution in rice plants. J Exp Bot 2007;58(11):2909-15, 2909. [CrossRef]
- Lee S, Jeong HJ, Kim SA, Lee J, Guerinot ML, An G. OsZIP5 is a plasma membrane zinc transporter in rice. Plant Mol Biol 2010 Jul;73(4-5):507-17. [CrossRef]
- Lee S, Kim SA, Lee J, Guerinot ML, An G. Zinc deficiency-inducible OsZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol Cells 2010 Jun;29(6):551-8. [CrossRef]
- Wu X, Zhu ZB, Chen JH, Huang YF, Liu ZL, Zou JW, Chen YH, Su NN, Cui J. Transcriptome analysis revealed pivotal transporters involved in the reduction of cadmium accumulation in pak choi (Brassica chinensis L.) by exogenous hydrogen-rich water. Chemosphere 2019 Feb;216:684-697. [CrossRef]
- Guo J, Li K, Zhang X, Huang H, Huang F, Zhang L, Wang Y, Li T, Yu H. Genetic properties of cadmium translocation from straw to brown rice in low-grain cadmium rice line. Ecotoxicol Environ Saf 2019 Oct 30;182:109422. [CrossRef]
- Yu R, Li D, Du X, Xia S, Liu C, Shi G. Comparative transcriptome analysis reveals key cadmium transport-related genes in roots of two pak choi (Brassica rapa L. ssp. chinensis) cultivars. BMC Genomics 2017 Aug 8;18(1):587. [CrossRef]
- Bienert S, Waterhouse A, de Beer TA, Tauriello G, Studer G, Bordoli L, Schwede T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res 2017 Jan 4;45(D1):D313-D319. [CrossRef]
- Fan W, Liu C, Cao B, Qin M, Long D, Xiang Z, Zhao A. Genome-Wide Identification and Characterization of Four Gene Families Putatively Involved in Cadmium Uptake, Translocation and Sequestration in Mulberry. Front Plant Sci 2018 Jun 29;9:879. [CrossRef]
- Palusińska M, Barabasz A, Kozak K, Papierniak A, Maślińska K, Antosiewicz DM. Zn/Cd status-dependent accumulation of Zn and Cd in root parts in tobacco is accompanied by specific expression of ZIP genes. BMC Plant Biol 2020 Jan 22;20(1):37. [CrossRef]
- Liu M, Zhang C, Duan L, Luan Q, Li J, Yang A, Qi X, Ren Z. CsMYB60 is a key regulator of flavonols and proanthocyanidans that determine the colour of fruit spines in cucumber. J Exp Bot 2019 Jan 1;70(1):69-84. [CrossRef]
- Chou KC, Shen HB. Cell-PLoc: a package of Web servers for predicting subcellular localization of proteins in various organisms. Nat Protoc 2008;3(2):153-62. [CrossRef]
- Hu B, Jin J, Guo AY, Zhang H, Luo J, Gao G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015 Apr 15;31(8):1296-7. [CrossRef]
- Bailey TL, Williams N, Misleh C, Li WW. MEME: discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006 Jul 1;34(Web Server issue):W369-73. [CrossRef]
- Crooks GE, Hon G, Chandonia JM, Brenner SE. WebLogo: a sequence logo generator. Genome Res 2004 Jun;14(6):1188-90, 1188. [CrossRef]
- Voorrips, RE. MapChart: software for the graphical presentation of linkage maps and QTLs. J Hered 2002 Jan-Feb;93(1):77-8. [CrossRef]
- Li L, Stoeckert CJ Jr, Roos DS. OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res 2003 Sep;13(9):2178-89, 2178. [CrossRef]
- Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA. Circos: an information aesthetic for comparative genomics. Genome Res 2009 Sep;19(9):1639-45. [CrossRef]
- Koch MA, Haubold B, Mitchell-Olds T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol Biol Evol 2000 Oct;17(10):1483-98. [CrossRef]
- Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 2002 Jan 1;30(1):325-7. [CrossRef]
- Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, Xia R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020 Aug 3;13(8):1194-1202. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001 Dec;25(4):402-8. [CrossRef]
- Li H, Wang S, Chai S, Yang Z, Zhang Q, Xin H, Xu Y, Lin S, Chen X, Yao Z, Yang Q, Fei Z, Huang S, Zhang Z. Graph-based pan-genome reveals structural and sequence variations related to agronomic traits and domestication in cucumber. Nat Commun 2022 Feb 3;13(1):682. [CrossRef]
- Eng BH, Guerinot ML, Eide D, Saier MH Jr. Sequence analyses and phylogenetic characterization of the ZIP family of metal ion transport proteins. J Membr Biol 1998 Nov 1;166(1):1-7. [CrossRef]
- Fu XZ, Zhou X, Xing F, Ling LL, Chun CP, Cao L, Aarts MGM, Peng LZ. Genome-Wide Identification, Cloning and Functional Analysis of the Zinc/Iron-Regulated Transporter-Like Protein (ZIP) Gene Family in Trifoliate Orange (Poncirus trifoliata L. Raf.). Front Plant Sci 2017 Apr 19;8:588. [CrossRef]
- Zhang, H. , Zhao, S., Li, D. et al. Genome-Wide Analysis of the ZRT, IRT-Like Protein (ZIP) Family and Their Responses to Metal Stress in Populus trichocarpa. Plant Mol Biol Rep 35, 534–549 (2017). [CrossRef]
- Zhang Z, Chen N, Zhang Z, Shi G. Genome-Wide Identification and Expression Profile Reveal Potential Roles of Peanut ZIP Family Genes in Zinc/Iron-Deficiency. Tolerance. Plants (Basel) 2022 Mar 16;11(6):786. [CrossRef]
- Gindri RG, Navarro BB, da Cruz Dias PV, Tarouco CP, Nicoloso FT, Brunetto G, Berghetti ÁLP, da Silva LOS, Fett JP, Menguer PK, Ricachenevsky FK. Physiological responses of rice oszip7 loss-of-function plants exposed to varying Zn concentrations. Physiol Mol Biol Plants 2020 Jul;26(7):1349-1359. [CrossRef]
- Tan L, Zhu Y, Fan T, Peng C, Wang J, Sun L, Chen C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem Biophys Res Commun 2019 Apr 23;512(1):112-118. [CrossRef]
- Vert G, Grotz N, Dédaldéchamp F, Gaymard F, Guerinot ML, Briat JF, Curie C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002 Jun;14(6):1223-33. [CrossRef]
- Akther MS, Das U, Tahura S, Prity SA, Islam M, Kabir AH. Regulation of Zn uptake and redox status confers Zn deficiency tolerance in tomato. Scientia Horticulturae 2020; 273. [CrossRef]
- Kabir AH, Tahura S, Elseehy MM, El-Shehawi AM. Molecular characterization of Fe-acquisition genes causing decreased Fe uptake and photosynthetic inefficiency in Fe-deficient sunflower. Sci Rep 2021 Mar 10;11(1):5537. [CrossRef]
- Cheong JJ, Choi YD. Methyl jasmonate as a vital substance in plants. Trends Genet 2003 Jul;19(7):409-13. [CrossRef]
- Castillo-Gonzalez J, Ojeda-Barrios D, Hernandez-Rodriguez A, Gonzalez-Franco AC, Robles-Hernandez L, Lopez-Ochoa GR. ZINC METALLOENZYMES IN PLANTS. Interciencia 2018, 43, 242–248.
- Eriksson S, Böhlenius H, Moritz T, Nilsson O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 2006 Sep;18(9):2172-81. [CrossRef]
- Preciado-Rangel P, Reyes-Pérez JJ, Ramírez-Rodríguez SC, Salas-Pérez L, Fortis-Hernández M, Murillo-Amador B, Troyo-Diéguez E. Foliar Aspersion of Salicylic Acid Improves Phenolic and Flavonoid Compounds, and Also the Fruit Yield in Cucumber (Cucumis sativus L.). Plants (Basel) 2019 Feb 16;8(2):44. [CrossRef]
- Li S, Liu X, Zhou X, Li Y, Yang W, Chen R. Improving Zinc and Iron Accumulation in Maize Grains Using the Zinc and Iron Transporter ZmZIP5. Plant Cell Physiol 2019 Sep 1;60(9):2077-2085. [CrossRef]
- Lee S, Kim SA, Lee J, Guerinot ML, An G. Zinc deficiency-inducible OsZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol Cells. 2010 Jun;29(6):551-8. [CrossRef]
- Pedas P, Schjoerring JK, Husted S. Identification and characterization of zinc-starvation-induced ZIP transporters from barley roots. Plant Physiol Biochem. 2009 May;47(5):377-83. [CrossRef]
- Wintz H, Fox T, Wu YY, Feng V, Chen W, Chang HS, Zhu T, Vulpe C. Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J Biol Chem. 2003 Nov 28;278(48):47644-53. [CrossRef]
- GONG Yuanyong, ZHAO Lihua, YAN Fei. Genome-Wide Identification and Expression Pattern Analysis of the Tomato ZIP Gene Family[J].Northeast Agricultural Science, 48(02), 42-48+109.
Figure 1.
Phylogenetic tree and gene structure of ZIP family members in C. sativus. The phylogenetic tree was constructed using the neighbor-joining (NJ) method with 1000 bootstrap replicates, based on the alignment of the identified 10 ZIP proteins in C. sativus. The gene structures of the identified 10 ZIP genes in C. sativus were generated using the Gene Structure Display Server v2.0. In the structures, the green box represents the UTR, the yellow box represents the exon, and the black line represents the intron.
Figure 1.
Phylogenetic tree and gene structure of ZIP family members in C. sativus. The phylogenetic tree was constructed using the neighbor-joining (NJ) method with 1000 bootstrap replicates, based on the alignment of the identified 10 ZIP proteins in C. sativus. The gene structures of the identified 10 ZIP genes in C. sativus were generated using the Gene Structure Display Server v2.0. In the structures, the green box represents the UTR, the yellow box represents the exon, and the black line represents the intron.
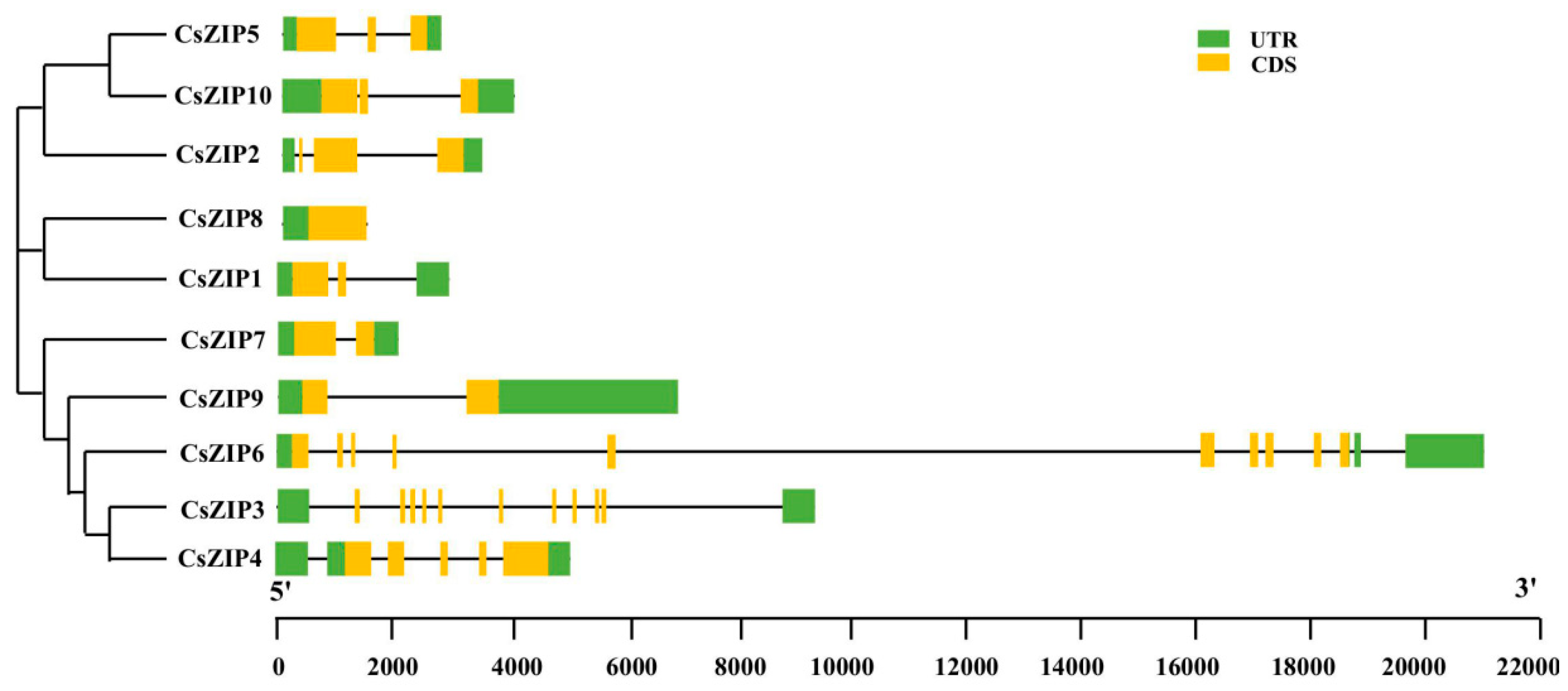
Figure 2.
The conserved motifs for C. sativus ZIP proteins. The conserved motifs identified by MEME are highlighted with colored boxes.
Figure 2.
The conserved motifs for C. sativus ZIP proteins. The conserved motifs identified by MEME are highlighted with colored boxes.
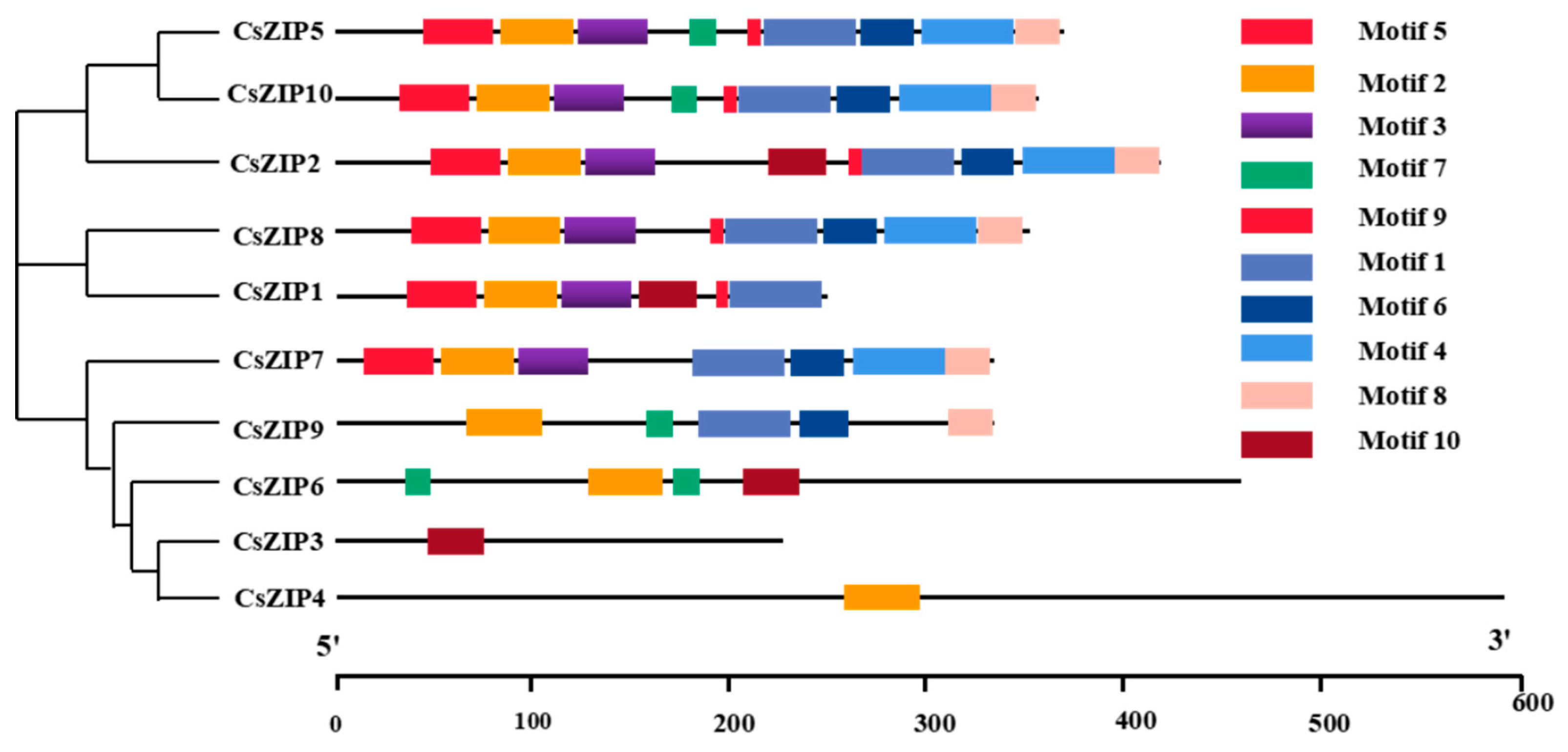
Figure 3.
Amino acid sequence alignment for ZIP proteins from Arabidopsis, C. sativus, Cucumis melo and O. sativa. The ZIP proteins were aligned using MAFFT v5.3 with the default settings. Completely conserved residues are shaded in black boxes, while highly conserved residues are shaded in gray boxes.
Figure 3.
Amino acid sequence alignment for ZIP proteins from Arabidopsis, C. sativus, Cucumis melo and O. sativa. The ZIP proteins were aligned using MAFFT v5.3 with the default settings. Completely conserved residues are shaded in black boxes, while highly conserved residues are shaded in gray boxes.
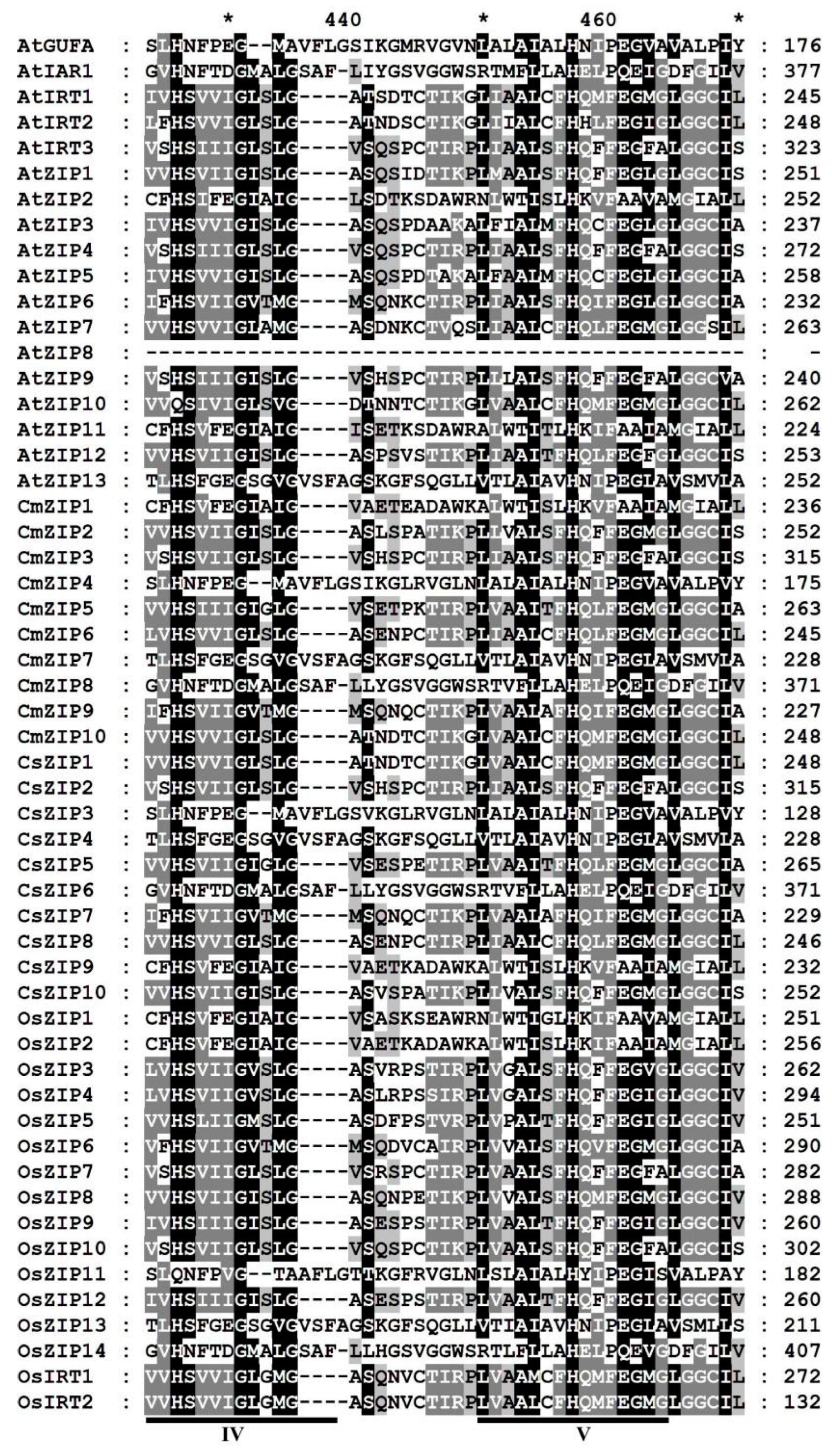
Figure 4.
Phylogenetic tree for the ZIP proteins from Arabidopsis, C. sativus, Cucumis melo and O. sativa. The unrooted amino acid sequence similarity tree was generated using MEGA 7.0 by neighbor-joining (NJ) method with 1000 bootstrap replicates.
Figure 4.
Phylogenetic tree for the ZIP proteins from Arabidopsis, C. sativus, Cucumis melo and O. sativa. The unrooted amino acid sequence similarity tree was generated using MEGA 7.0 by neighbor-joining (NJ) method with 1000 bootstrap replicates.
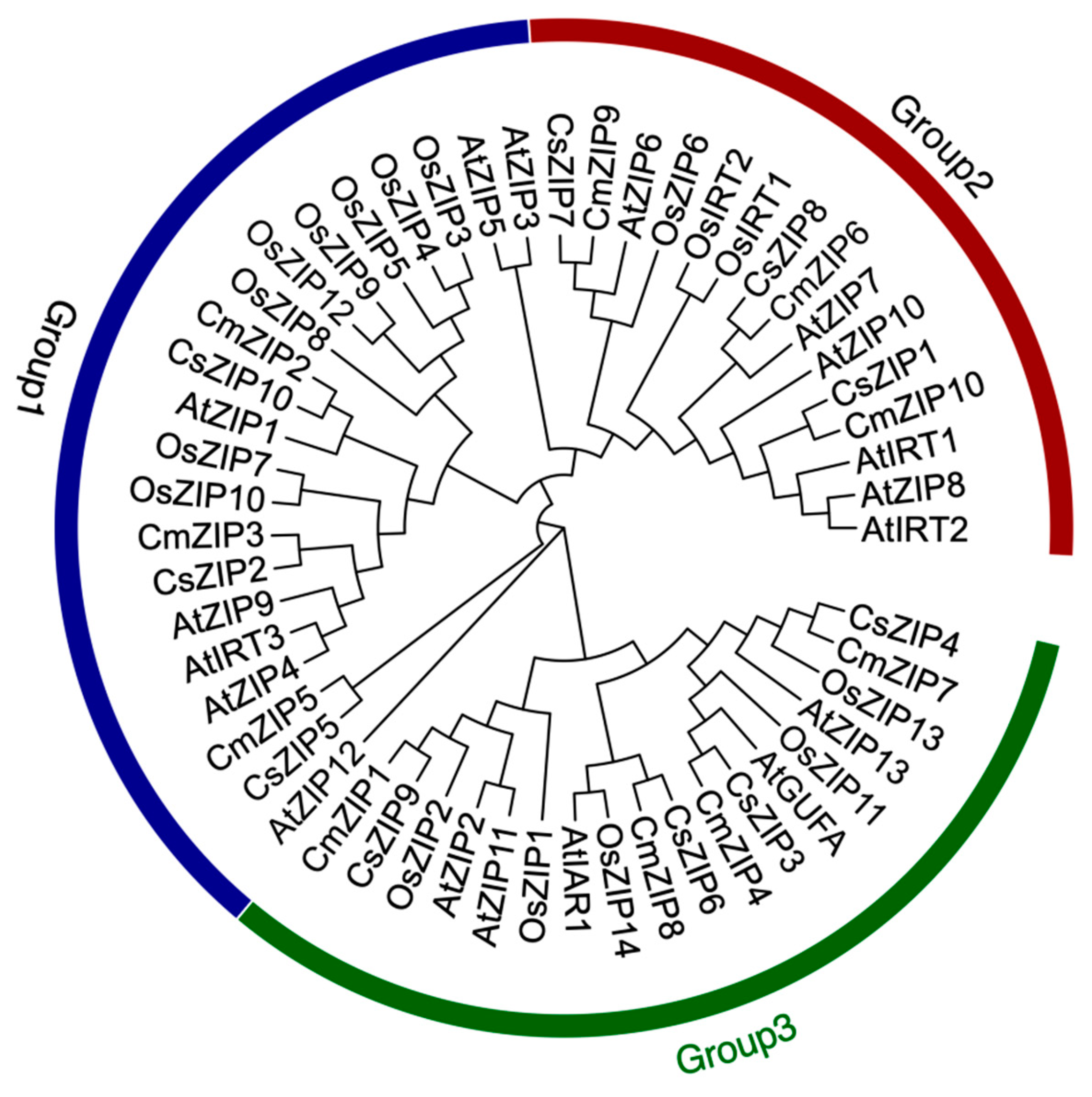
Figure 5.
Chromosomal location of C. sativus ZIP genes.
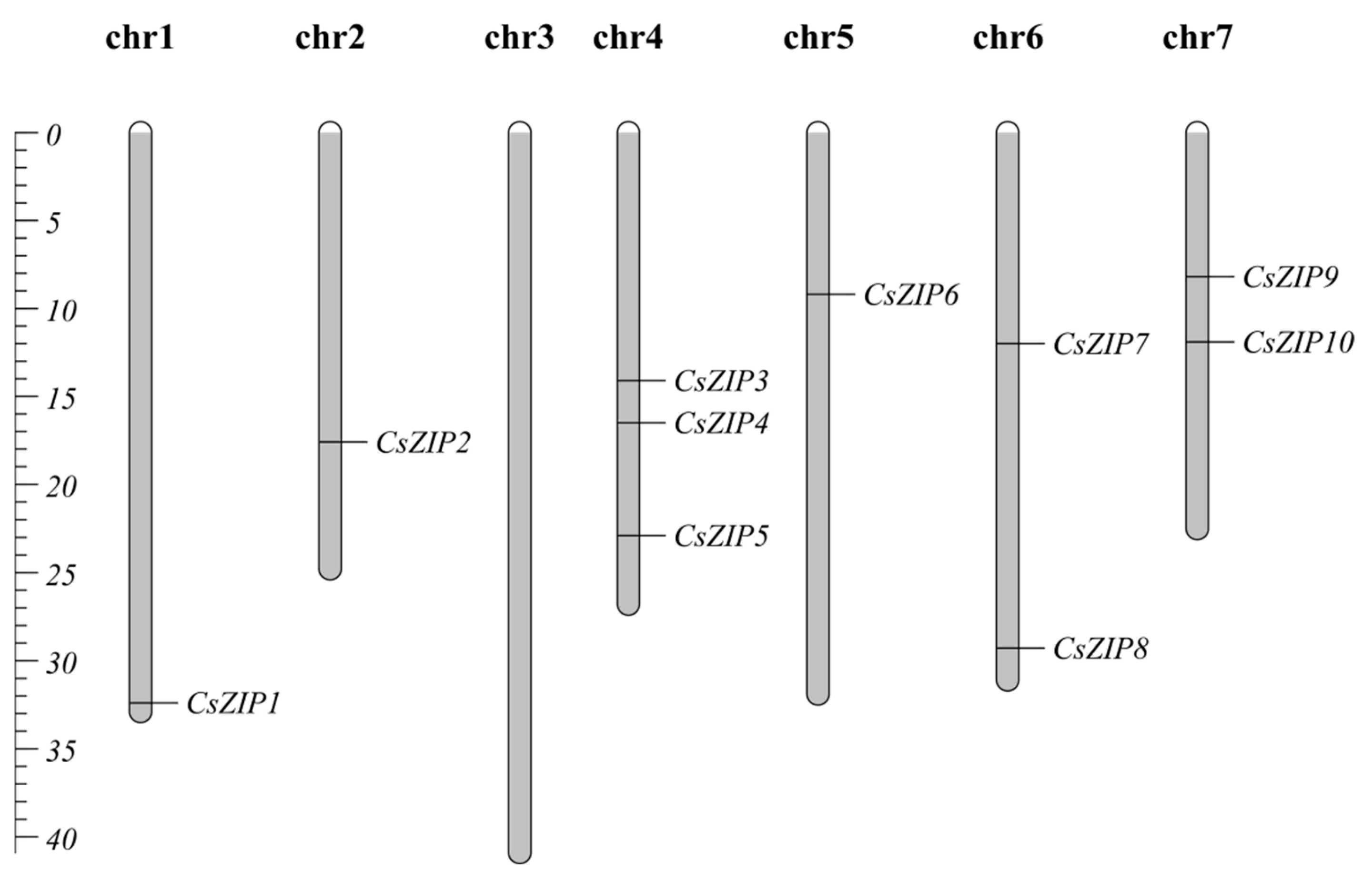
Figure 6.
Synteny analysis of ZIPs between C. sativus and other plant species (A. thaliana, Oryza sativa and Cucumis melo): The collinear blocks are marked by gray lines, while the collinear gene pairs with ZIP genes are highlighted by red lines.
Figure 6.
Synteny analysis of ZIPs between C. sativus and other plant species (A. thaliana, Oryza sativa and Cucumis melo): The collinear blocks are marked by gray lines, while the collinear gene pairs with ZIP genes are highlighted by red lines.
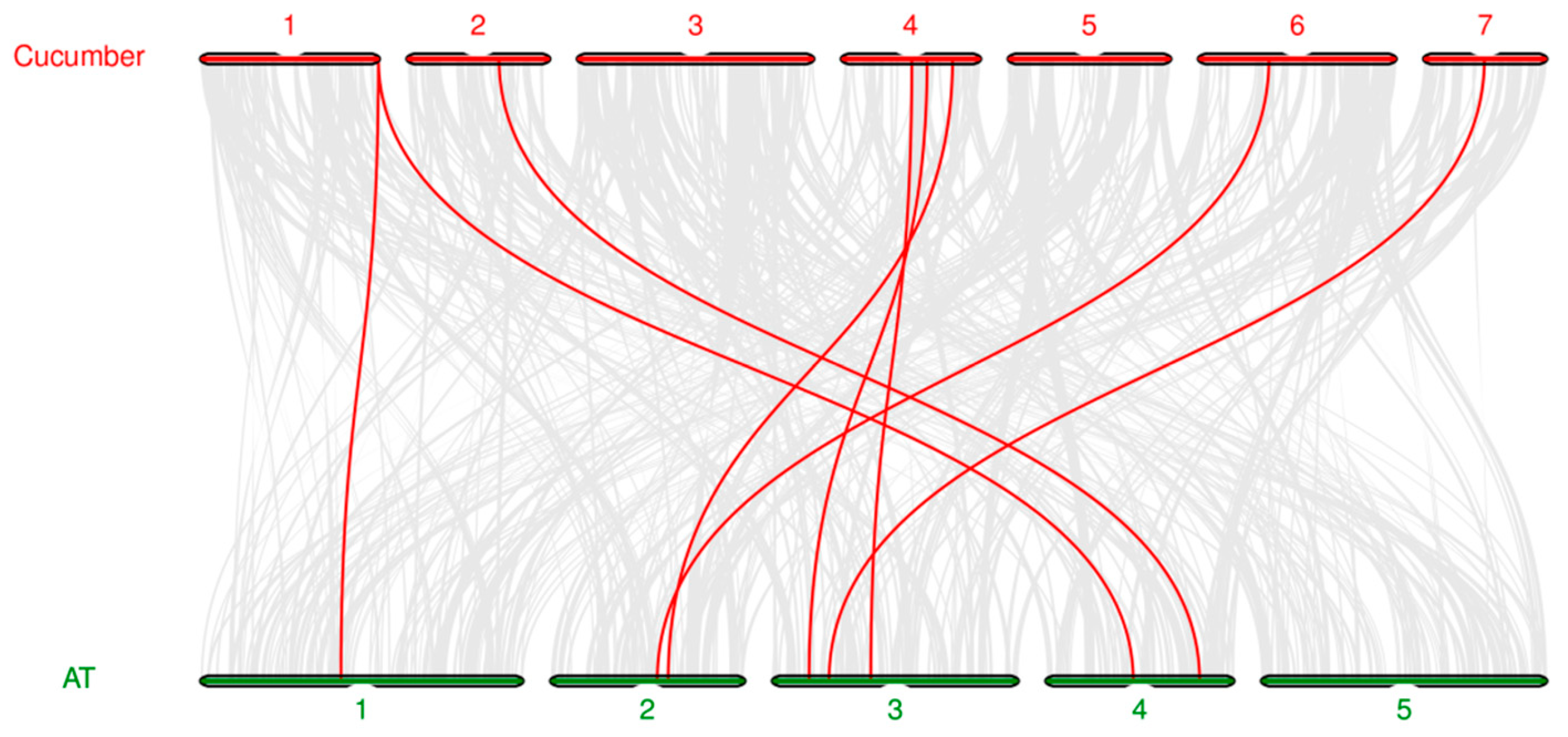

Figure 7.
Predicted cis-elements in promoter regions of C. sativus ZIP genes. The promoter region was defined as a 1.5 kb sequence upstream of the translation initiation codon of the CsZIP gene. Identification of cis-acting elements using the online tool Plant CARE. Different types of cis-acting elements are represented by closed boxes of different colors.
Figure 7.
Predicted cis-elements in promoter regions of C. sativus ZIP genes. The promoter region was defined as a 1.5 kb sequence upstream of the translation initiation codon of the CsZIP gene. Identification of cis-acting elements using the online tool Plant CARE. Different types of cis-acting elements are represented by closed boxes of different colors.
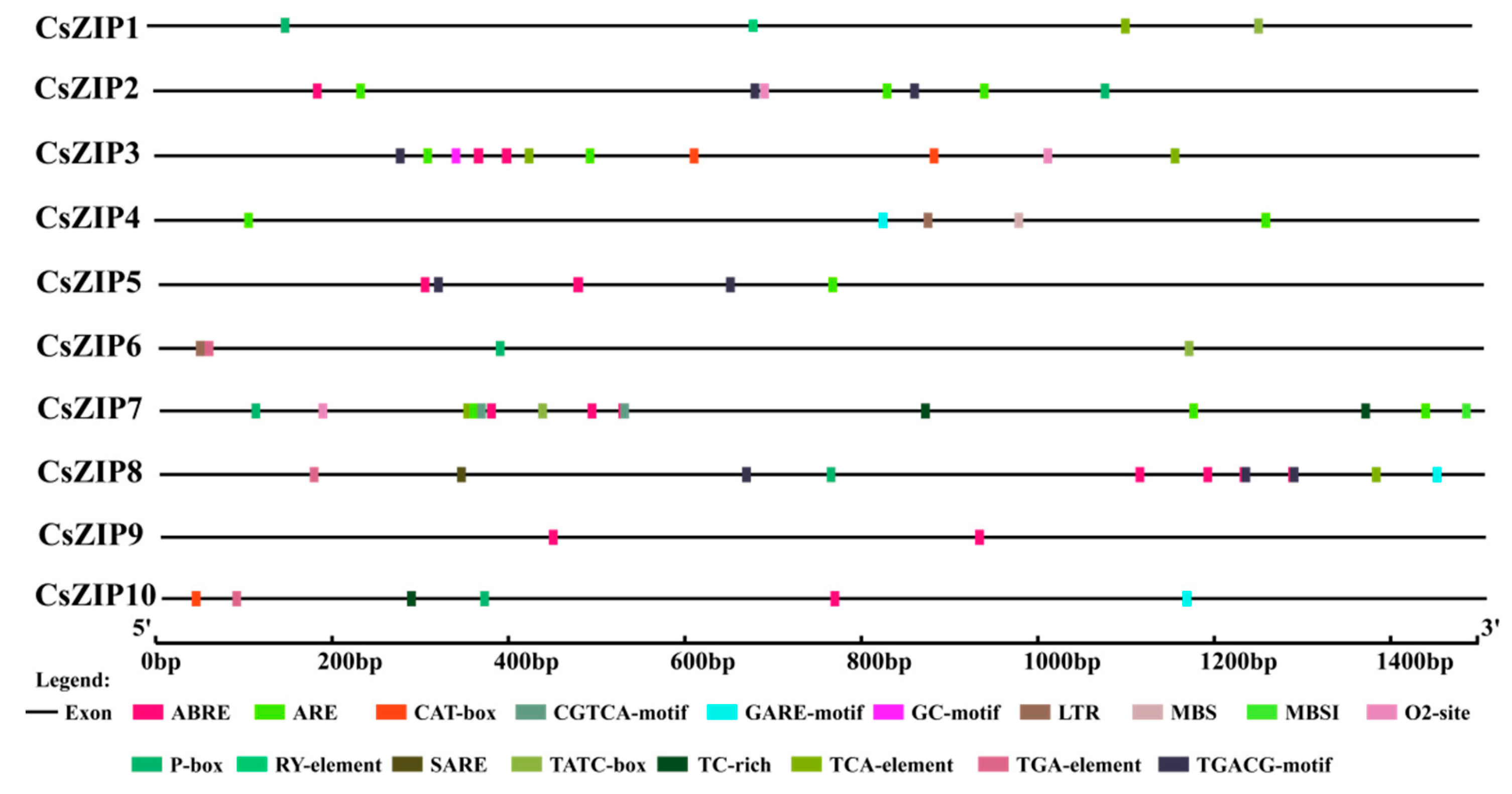
Figure 8.
Temporal-spatial expression of C. sativus ZIP genes. (a) Heatmap displaying the expression profile of CsZIP genes in nine different C. sativus tissues. The RNA-seq datasets with accession number PRJNA80169 were obtained from the Cucurbit Genomics Data website. The color scale represents Log2(FPKM) values, where blue and red indicate low and high expression levels, respectively. The FPKM values of CsZIP genes can be found in Table S1. (b) Validation of RNA-seq results for four CsZIP genes using qRT-PCR. The error bars represent the standard error of the mean (n = 3). R: root; S: stem; L: leaf; FF: female flower; MF: male flower; O: unexpanded ovary; O-fer: expanded fertilized ovary; O-unfer: expanded unfertilized ovary; T: tendril.
Figure 8.
Temporal-spatial expression of C. sativus ZIP genes. (a) Heatmap displaying the expression profile of CsZIP genes in nine different C. sativus tissues. The RNA-seq datasets with accession number PRJNA80169 were obtained from the Cucurbit Genomics Data website. The color scale represents Log2(FPKM) values, where blue and red indicate low and high expression levels, respectively. The FPKM values of CsZIP genes can be found in Table S1. (b) Validation of RNA-seq results for four CsZIP genes using qRT-PCR. The error bars represent the standard error of the mean (n = 3). R: root; S: stem; L: leaf; FF: female flower; MF: male flower; O: unexpanded ovary; O-fer: expanded fertilized ovary; O-unfer: expanded unfertilized ovary; T: tendril.
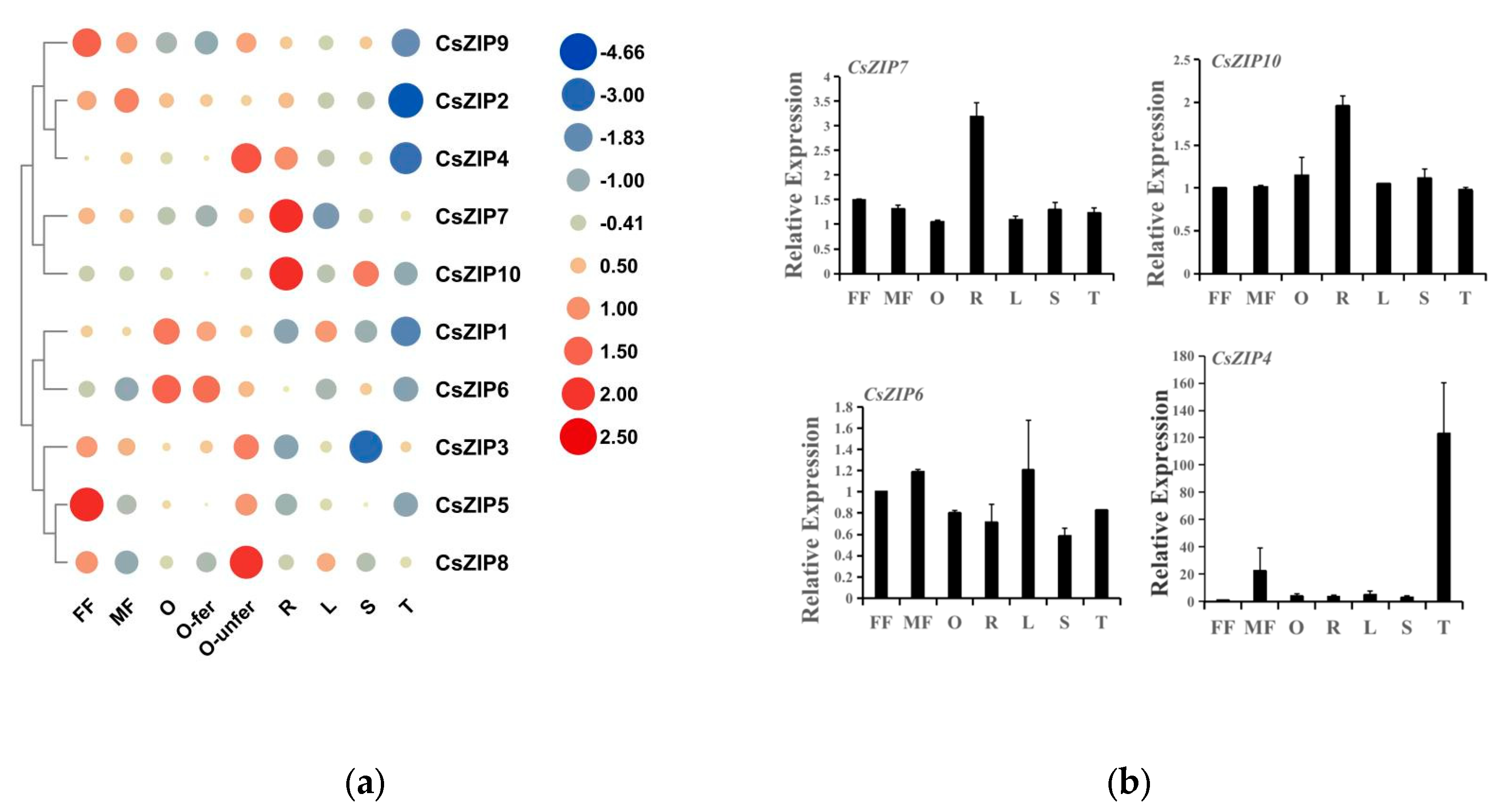
Figure 9.
Expression of CsZIP genes in response to salt and hot stimuli. (a) Heatmap for displaying the expression profile of CsZIP genes in response to salt stresses. (b) Heatmap for displaying the expression profile of CsZIP genes in response to hot (42°C). RNA-seq datasets with accession numbers of GSE151055 and GSE116265 were downloaded from NCBI SAR database. Color scale represents Log2(FPKM) values. Green, red, and black color indicates low, high, and no expression, respectively. The FPKM value of CsZIP genes under salt and hot treatments are listed in Table S2.
Figure 9.
Expression of CsZIP genes in response to salt and hot stimuli. (a) Heatmap for displaying the expression profile of CsZIP genes in response to salt stresses. (b) Heatmap for displaying the expression profile of CsZIP genes in response to hot (42°C). RNA-seq datasets with accession numbers of GSE151055 and GSE116265 were downloaded from NCBI SAR database. Color scale represents Log2(FPKM) values. Green, red, and black color indicates low, high, and no expression, respectively. The FPKM value of CsZIP genes under salt and hot treatments are listed in Table S2.
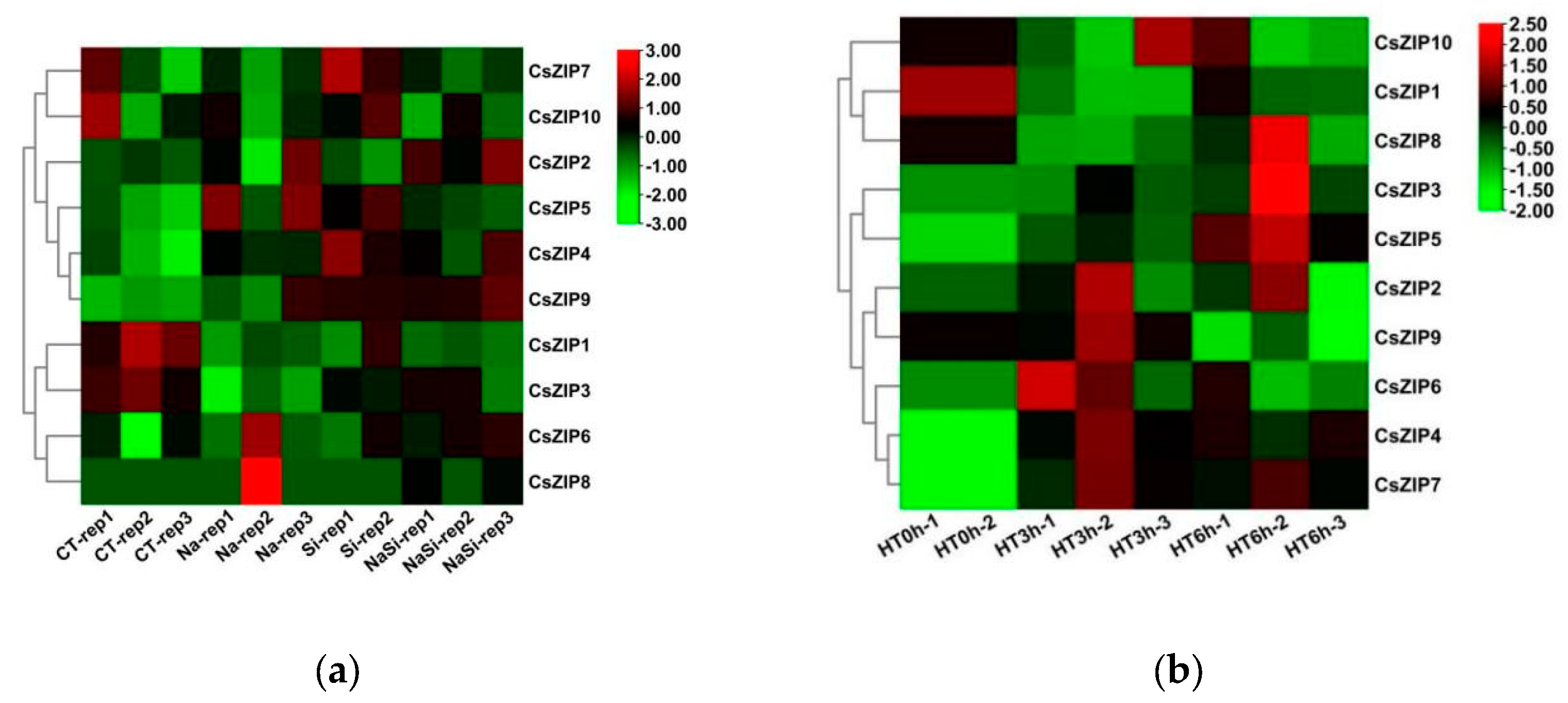
Figure 10.
Expression analysis of CsZIPs under biotic stresses: The transcriptional levels of CsZIP genes after infection with powdery mildew (PM) for 48 h (a) and with downy mildew (DM) for 1–8 days post-inoculation (b) are shown on the heatmaps. The color scale shows increasing expression levels from green to red. ID, PM-inoculated susceptible C. sativus line D8 leaves; NID, non-inoculated D8 leaves; IS, PM-inoculated resistant C. sativus line SSL508-28 leaves; NIS, non-inoculated SSL508-28 leaves; CT, without inoculation; DPI, days post inoculation. The FPKM value of CsZIP genes under powdery mildew and downy mildew are listed in Table S3.
Figure 10.
Expression analysis of CsZIPs under biotic stresses: The transcriptional levels of CsZIP genes after infection with powdery mildew (PM) for 48 h (a) and with downy mildew (DM) for 1–8 days post-inoculation (b) are shown on the heatmaps. The color scale shows increasing expression levels from green to red. ID, PM-inoculated susceptible C. sativus line D8 leaves; NID, non-inoculated D8 leaves; IS, PM-inoculated resistant C. sativus line SSL508-28 leaves; NIS, non-inoculated SSL508-28 leaves; CT, without inoculation; DPI, days post inoculation. The FPKM value of CsZIP genes under powdery mildew and downy mildew are listed in Table S3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



