
Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Objectives of this study were to investigate effects of repeated superstimulation on the success of ovum-pick up and in vitro production of embryos (OPU-IVP) and donor health and evaluate the efficiency of a modified SOF sequential medium in producing viable embryos. Over 1 y, six cross-bred beef heifers and three beef cows were subjected to superstimulations and OPU-IVP. Oocytes recovered were used for in vitro embryo production and the resulting embryos were transferred as fresh or frozen-thawed. Number of follicles, oocytes recovered, cleavage and blastocyst rates substantially varied among individuals. However, repeated trials did not significantly affect the efficiency of OPU-IVP. The pregnancy rates (PR) 22 to 30 d after embryo transfer for in vitro-derived fresh embryos was similar to that of fresh in vivo embryos, whereas transfer of in vitro-derived frozen embryos reduced PR compared to frozen-thawed in vivo embryos. All recipient heifers diagnosed pregnant delivered a viable calf, suggesting developmental competence of the embryos. Repeated OPU-IVP had no apparent deleterious effects, as all donors had viable calves after ET. We concluded that multiple OPU-IVP cycles can be effectively performed for a prolonged interval to maximize embryo production from elite donors without compromising their reproductive health.
Keywords:
ovum pick up
; in vitro embryo production
; embryo transfer
; beef cattle
; cow
; heifer
1. Introduction
A burgeoning population requires increases in food production and cattle productivity. Existing and emerging reproductive technologies, including in vitro production (IVP) of embryos, are fundamental to propagating elite cows, improving productivity and global dissemination of superior genetics [1].
Traditional embryo transfer (ET) is commonly used to propagate offspring from superior beef and dairy cattle. However, reproductive efficiency can be increased several-fold by repeated ultrasound-guided ovum pick-up and in vitro production of embryos (OPU-IVP). Oocytes recovered by OPU can be fertilized with sperm from more than one bull or with sexed semen (with no reduction in fertility) [2,3]. Moreover, generation interval can be reduced, as OPU can be done before puberty or up to 3 mo of pregnancy. In addition, OPU-IVP is a viable option for cattle with reduced fertility due to reproductive tract pathology, impaired sperm transport, or terminal disease [4]. Clearly, OPU-IVP could substantially increase number of calves produced compared to conventional ET, boost livestock productivity and environmental sustainability, due to fewer cattle and less greenhouse gas emissions [5].
Although global in vivo bovine embryo production due to ET has largely stabilized, in vitro production of bovine embryos was 31.5% higher in 2021 versus 2020 (International Embryo Technology Society data) [6], with 34.5 and 37.0% increases in North and South America, respectively. Canada consistently ranks within the top 10 countries globally in exporting beef and other cattle products. Canada has ~ 11.5 million cattle (9.5 million beef and 2 million dairy), with > 40% of Canada’s beef herd in Alberta [7,8].
In vitro embryo production has enhanced genetics and production in dairy and beef, with OPU-IVP used mainly in dairy. Lactating dairy cows generally have lower oocyte quality, fertilization rate, and compromised development of early blastocysts, attributed to lactational metabolic challenges, and more metabolic and infectious diseases during the postpartum period [9,10]. In addition, high milk production hastens steroid metabolism, reducing progesterone and estradiol concentrations and decreasing estrus expression [11]. Thus, the OPU-IVP and superstimulation approach is growing, due to its greater efficiency in embryo production [12]. Also, sex-sorted sperm is used to produce female embryos from young heifers with high genetic value [13,14].
The goal of IVP is to replicate the maternal reproductive tract. Oocyte quality is critical, with breed differences [2]. The success of OPU-IVP can also be influenced by other factors, including donor age, nutritional status, milk yield, parity, lactational status, management, etc. [15,16,17]. Synchronizing follicular waves and stimulating them with follicle stimulating hormone (FSH) enhance oocyte quality and promote blastocyst formation [13], with follicle number and size positively affecting embryo production [18]. Additionally, sperm preparation is crucial for IVF, to eliminate seminal plasma and choose highly motile sperm.
OPU can be done in cattle with or without superstimulation, with the latter routinely done twice weekly [19,20] for long intervals without affecting donor cow fertility or well-being, preventing dominant follicles that reduce developmental competence of cumulus-oocyte complexes (COCs) [21,22]. Furthermore, hormonal stimulation increases number of COCs and rates of cleavage, morula and blastocyst formation [23].The usual regimen is FSH twice daily for 4 or 5 d [24]. In cattle, inducing ovarian stimulation and subsequently allowing a period of gonadotropin-free rest, commonly referred to as "coasting," can improve oocyte quality and blastocyst yields [25]. Coasting improves OPU/IVP results., e.g., an 80% blastocyst rate in females given six injections of FSH followed by 48-h coasting [23,26,27]. Limited studies have been conducted on OPU donors undergoing superstimulation, and the consequences of repeated superstimulation and egg retrieval on donor health and well-being, as well as the success of OPU-IVP, are still not fully known. There are concerns OPU may reduce animal welfare. Repeated OPU is mildly invasive [28,29]; although it elevated stress and decreased productive and reproductive herd performance, it did not appear to affect reproductive ability [30].
Embryos produced in vivo are exposed to variable fluid composition and gas atmosphere, with IVC attempting to replicate those conditions, typically using static or sequential culture. In the latter, the media and its components are changed based on embryo stage and stage-specific embryonic demands [31]. In contrast, static culture has a single formulation of culture media, allowing an embryo to select required components [32]. Both methods have advantages and disadvantages. Sequential culture mimics in vivo conditions but may cause more oxidative stress by changing media and increased handling. In contrast, static media allows for decreased disturbance of pH and temperature, reducing ROS production and allowing embryos to benefit from secreted growth factors, though waste and ROS can be deleterious [4].
Once embryos become blastocysts, they can be transferred fresh or cryopreserved. Unfortunately, IVP embryos have poor viability and cryotolerance compared to in vivo derived embryos [33], attributed to cytoplasmic accumulation of lipids, particularly if media contains fetal bovine serum (FBS). Serum withdrawal can reduce lipid accumulation and improve cryotolerance, but often compromises embryo development [34]. Freezing tolerance is highly dependent on these lipids, and embryos with altered lipid composition and content are more sensitive to cryoinjury. Cryodamage typically occurs during lipid phase transition and lipid peroxidation events [35]. Improving the OPU-IVP procedure and cryopreservation should increase embryo quality and live births.
Objectives of this study were to: 1) investigate effects of repeated OPU-IVP sessions on ovarian follicular development, oocyte recovery, developmental competence of embryos, and donor health; and 2) evaluate the efficiency of a modified SOF sequential medium in producing viable embryos that can survive cryopreservation. We tested the hypothesis that repeated OPU-IVP efficiently produces viable offspring with no deleterious effects on donors.
2. Materials and Methods
2.1. Cattle and Ethics
This experiment was conducted as per the animal care protocol # AC19-0044 of the University of Calgary. Six cross-bred beef heifers (12-14 mo old) and three purebred cows (one Simmental and two Maine Anjou) were subjected to repeated superstimulations and OPU-IVP for 1 y.
2.2. Superstimulation
Intravaginal insert of a CIDR (CIDR 1380, Zoetis Canada, Kirkland, QC, Canada) was designated Day 0, with transvaginal aspiration of dominant follicles > 10 mm on Day 2. Injections of FSH (Folltropin-V, Vetoquinol, Lavaltrie, QC, Canada) (2 to 3 ml; IM) were given 12 h apart beginning on Day 4 (AM) and ending on Day 6 PM (six in total). Transvaginal ultrasound-guided oocyte aspiration was done after 38 to 40 h of coasting (interval between last FSH and oocyte collection).
2.3. Transvaginal Ultrasound-Guided Oocyte Aspiration
On Day 8, ovarian follicles were aspirated using an ultrasound-guided transvaginal approach. The COCs were immediately recovered, transferred to maturation medium (Hank’s medium with 10% FCS, 0.2 mM sodium pyruvate, 0.5 μg/mL FSH, 5 μg/mL LH, 1 μg/mL estradiol, and 50 μg/mL gentamicin) and maintained in 5-ml culture vials. Vials were gassed with CO2, sealed, and transported to the laboratory in a portable incubator maintained at 39 °C.
2.4. In Vitro Maturation, Fertilization, and Culture
At the laboratory, oocytes were transferred from vials and in vitro-matured in the IVM medium as described above and incubated at 39 °C and 5% CO2 for 22-24 h. In vitro-matured oocytes (oocytes with expanded cumulus cells and a homogenous cytoplasm) were transferred to pre-equilibrated fertilization drops (50 µl final volume; maximum 10 oocytes/drop).
Fertilization media contained 113.96 mM NaCl, 3.15 mM KCl, 25.04 mM NaHCO3, 0.34 mM NaH2PO4.H2O, 10 mM sodium lactate, 2.04 mM CaCl2.2H2O, 0.49 mM MgCl2.6H2O,1 mM sodium pyruvate, 6 mg/ml bovine serum albumin (BSA), 10 µg/ml heparin, PHE (18 µM D-penicillamine, 9 µM hypotaurine and 1.8 µM epinephrine), and 50 μg/mL gentamicin. Matured oocytes were incubated with frozen-thawed sperm (~ 1-2 x 106 sperm/mL of fertilization medium) from a bull with proven fertility subjected to swim-up to optimize sperm quality [36].
After 18 h of sperm-oocyte co-incubation, all presumptive zygotes were cumulus-dissociated by pipetting in H-SOF (107.7 mM NaCl, 7.16 mM KCl, 25.07 mM NaHCO3, 1.19 mM KH2PO4, 1.5 mM sodium lactate, 1.5 mM Glucose, 10 mM HEPES, 3 mg/ml BSA, 10 μM EDTA, 0.33 mM sodium pyruvate, 1 mM L-Glutamine, 1X MEM non-essential amino acid, and 50 μg/mL gentamicin). Cumulus-dissociated presumptive zygotes were washed and cultured in SOF-1 for 48 h. The SOF 1 media contained 107.7 mM NaCl, 7.16 mM KCl, 25.07 mM NaHCO3, 1.19 mM KH2PO4, 1.5 mM sodium lactate, 1.5 mM glucose, 1.71 CaCl2.2H2O, 0.49 mM MgCl2.6H2O, 8 mg/ml BSA, 10 μM EDTA, 0.33 mM sodium pyruvate, 1 mM L-Glutamine, 1X MEM non-essential amino acid, and 50 μg/mL gentamicin. On Day 3 (Day 0 = fertilization), cleavage was evaluated and cleaved embryos were transferred to SOF-2 (107.7 mM NaCl, 7.16 mM KCl, 25.07 mM NaHCO3, 1.19 mM KH2PO4, 1.5 mM sodium lactate, 1.5 mM glucose, 1.71 CaCl2.2H2O, 0.49 mM MgCl2.6H2O, 8 mg/ml BSA, 0.33 mM sodium pyruvate, 1 mM L-Glutamine, 1X MEM non-essential amino acids, 1X MEM essential amino acids, and 50 μg/mL gentamicin). On Day 5, embryos were transferred to SOF-3 [SOF2 media supplemented with 2.5% TCH serum replacement (Protide Pharmaceuticals, IL, USA) and 2.5% FCS].
Blastocyst production was evaluated on Day 7, based on International Embryo Transfer Society criteria [37]. Embryos from purebred cows were transferred fresh to estrus-synchronized recipient cows or cryopreserved. For cryopreservation, embryos were washed in holding media (BoviPro, MOFA Global, Verona, WI, USA) and transferred to the freezing medium (BoviPro, MOFA Global, Verona, WI, USA) [38]. Each blastocyst was loaded into a 0.25 ml straw, put in a programmable freezer (Beltron EFT 3002, Longmont, CO, USA) and after 5 min of exposure of embryos to freezing medium, maintained at – 6.5 oC for 1, min, then seeded by touching the top and bottom columns of the holding medium with a cotton swab dipped in liquid nitrogen, held for >10 min, then cooled to –35 °C at a controlled rate (0.5 °C/min), plunged, and stored in liquid nitrogen. Embryos were thawed for 1 min in a water bath (37 °C) and direct transferred.
2.5. Embryo Transfer
In vitro-produced embryos (fresh and frozen-thawed) were transferred to recipients (n=30 each) by a certified ET practitioner. In brief, all recipient cows were estrus-synchronized by intravaginal insertion of a CIDR (Day 0) and injection of GnRH (Fertiline, Vetoquinol). On Day 7, CIDR was removed a 750 µg cloprostenol (Estrumate, Merck Animal Health, Kirkland, QC, Canada) given im. On Day 9 morning (48 h after CIDR removal), recipients were given 100 µg GnRH (Fertiline) im. On Day 16, transrectal ultrasonography was used to evaluate the ovaries and those with a corpus luteum were deemed eligible to be an embryo recipient. Transrectal ultrasonography 30 d after embryo transfer was used to detect pregnancy. The ET practitioner provided pregnancy data from their practice from transfer of in vivo-produced fresh versus frozen embryos.
2.6. Statistical Analyses
To determine effects of cow or number of trials on OPU-IVP, one-way ANOVA with post-hoc Tukey HSD Test or a Kruskal-Wallis test was used, depending on the data sets. Chi square was used to compare pregnancy rates of embryos resulting from fresh in vivo versus fresh in vitro or frozen in vivo versus frozen in vitro. One-way ANOVA with post-hoc Tukey HSD was used to evaluate embryo development and post-thaw survival rate (arc sine transformed).
3. Results
3.1. Evaluation of the Donor Effect on OPU-IVP Outcomes in Crossbred Heifers
All crossbred heifers (n=6; 12 to 14 mo old at the outset) responded to superstimulatory treatments for a prolonged interval (~ 1 y). Superstimulatory responses, number of oocytes recovered, cleavage and blastocyst rates are in Figure 1.
3.2. Evaluation of Repeated Superstimulations on OPU-IVP Responses in Crossbred Heifers
Despite variations among heifers in number of follicles, number of OPU-IVP trials did not affect outcomes, with no change in number of follicles or embryo development over ~ 1 y of repeated superstimulatory treatments (Table 1). Furthermore, none had any health concerns and after the trial, all received an OPU-IVP embryo, became pregnant and calved.
3.3. Evaluation of OPU-IVP Outcomes among Purebred Cows
The OPU-IVP responses of three beef cows are provided in Table 2. Although cows differed in their OPU-IVP responses, range of cleavage rates (54 to 70%) and blastocyst production (20 to 78%) were higher than those from the crossbred heifers.
3.4. Developmental Competence of OPU-IVP Embryos
In this study, all recipients diagnosed as pregnant at 22 to 30 d after embryo transfer delivered a viable calf. A total of 29 fresh IVP embryos were transferred, yielding a pregnancy rate of 51.7% (15 pregnancies). However, only 33.3% of transferred frozen-thawed embryos resulted in a viable calf. These results were compared to the pregnancy rates resulted from the transfer of in vivo-derived fresh (607/964; 63.0%) and frozen (606/1124; 53.9%) embryos conducted during the same year by the same veterinarian. These data demonstrated that there were no significant differences in pregnancy rates between in vivo- and in vitro-derived fresh embryos (63 vs. 51%, respectively). However, pregnancy rates for in vivo frozen embryos were higher than that of IVP frozen embryos (54 vs. 33%, respectively; P<0.03)
4. Discussion
Our objectives were to evaluate the impacts of repeated superstimulation OPU-IVP on embryo production plus donor reproductive health. Over 1 y, both heifers and cows were subjected to nine rounds of ovarian superstimulation, followed by OPU-IVP. Despite differences among individual animals, it was clear that the number of OPU-IVP trials did not affect IVP outcomes, with production of viable embryos, some of which were transferred (fresh or frozen-thawed) and produced healthy calves. Also, after the end of the study, all heifers received an embryo, became pregnant and subsequently calved. These results supported our hypothesis that repeated OPU-IVP efficiently produces viable offspring with no deleterious effects on donors.
Despite variations among animals and trials, it was evident that these cattle responded to our superstimulatory treatments for a prolonged interval, consistent with previous reports [39,40]. In general, multiple injections of FSH enhanced superstimulatory responses and developmental competence of oocytes. Repeated superstimulatory treatments in dairy cows did not significantly affect number of follicles or embryo development [19], consistent with our study in beef cattle. Although variations among individuals in their responses to superstimulatory treatments is expected, further improvements to optimize follicular size and improve rates of oocyte recovery and blastocyst production are required to maximize outcomes of OPU-IVP.
Repeated ovarian superstimulations and OPU-IVP were done in purebred cows (n=3). Although cows differed in their OPU-IVP responses, in general, IVP outcomes were higher than those from the crossbred heifers. Perhaps this could be a reflection of our improved efficiency in the OPU-IVP and genetic backgrounds of the cows used in this study. It has been reported that cattle with higher genetic backgrounds for in vivo superovulation also typically have a high capacity to generate blastocysts through OPU-IVP [41]. Another study demonstrated the importance of the breed on oocyte quality and in vitro embryo development [42].
This study clearly demonstrated that repeated egg recovery over 1 y did not compromise reproductive health of heifers, as all became pregnant and calved following transfer of IVP embryos. Some producers have concerns that repeated egg recovery will affect reproductive health of their valuable cows. Potential consequences include ovarian scar tissue or adhesions, bleeding or infection. In our studies, both heifers and cows tolerated multiple sessions of OPU-IVP without effects on reproductive or general health throughout the study or thereafter.
When we transferred OPU-IVP embryos, both fresh and frozen, pregnancy rate of IVP fresh embryos was higher than their frozen counterparts. Also, we compared these results with the transfer of in vivo-derived fresh and frozen embryos data. There were no differences in pregnancy rates between in vivo and in vitro derived fresh embryos, although the wide difference in number of in vivo- and in vitro-derived embryos transferred compromised statistical analysis. In addition, pregnancy rates of in vivo-produced frozen embryos were significantly greater than that of IVP frozen embryos, indicating the need for further refinement of embryo culture and freezing procedures. In an older study [40], transfer of IVF-fresh embryos yielded 56% pregnancy rate compared to that of IVF-frozen embryos (42%). However in vivo-fresh and -frozen samples yielded pregnancy rates of 66 and 67%, respectively. Similarly, when pregnancy rates were compared between in vivo and in vitro embryos derived from the same Nelore donor cows [43], there was ~ a 10% decrease in pregnancy rates for OPU-IVP embryos. Several variables can affect embryo developmental competence and affect cryosurvival, maintenance of pregnancy [44], and birth of viable calves.
Cryopreservation enables prolonged preservation of oocytes and embryos; the latter can be thawed and transferred, facilitating large-scale use [4]. Therefore, producing embryos that tolerate cryopreservation is an important challenge of OPU-IVP [33]. Cryopreservation causes thermal, mechanical, osmotic and toxic challenges, affecting gene expression [45]. Furthermore, culture conditions can alter metabolism and lipid profile of embryos and reduce viability of frozen-thawed IVP versus in vivo embryos. The majority of in vitro-produced (IVP) embryos display features linked to lower quality, including vacuoles in trophoblastic cells, a decreased number of mitochondria and microvilli, diminished intercellular junctions, variations in gene expression, and changes in lipid metabolism [46,47,48].
Cryotolerance is highly dependent on lipids; embryos with altered lipid composition and content are more sensitive to cryoinjury. Cryodamage typically occurs during lipid phase transition and lipid peroxidation events during freezing and thawing [35]. Lipid phase transition alters conformation states of lipid molecules from a fluid phase to a solid-like-ordered phase which can cause cryodamage [49]. Suboptimal culture conditions, particularly inclusion of fetal bovine serum (FBS), promote cytoplasmic accumulation of lipids. Withdrawal of serum can reduce lipid accumulation and improve cryotolerance of bovine embryos, but often compromises embryo development [34]. Strategies to increase cryotolerance of IVP embryos include culture under low O2 to minimize oxidative stress, use of antioxidants or apoptosis inhibitors [50,51] or substances to accelerate lipid metabolism or prevent lipid accumulation [52]. Further research is required for formulating a culture medium for efficient production of viable embryos that better withstand cryopreservation and thawing.
Quality control measures in the laboratory and the use of superior supplies are crucial for successful embryo production. This includes monitoring and maintaining laboratory air quality and temperature, ensuring the osmolarity and pH of media, oil overlay, and preventing from oxidative stress and toxic components exposure [53,54]. Optimizing pregnancy rates can be achieved through the careful selection of high-quality embryos at the appropriate developmental stage throughout the transfer process, by a skilled and experienced individual. Embryologists and those involved in gametes and embryo handling, assessment, and grading should undergo sufficient training and ongoing professional education development. Veterinarians supervising an OPU-IVP facility should havespecialized training, licensure, and certifications from the relevant regulatory authorities (i.e., provincial or state veterinary licensing, and be nationally accredited for the retrieval of oocytes, handling, and in vitro production of embryos) [55].
Improving efficiency and promoting widespread adoption of assisted reproductive technologies relies on optimizing the OPU-IVP procedure for the highest recovery of oocytes, in vitro production of viable embryos with higher cryotolerance, and prioritize the safety and well-being of donor. The development of oocytes can be affected by a range of factors, such as breed, age, nutritional conditions, environmental factors, as well as hormonal and metabolic status [56,57]. The success rate of IVEP is usually higher in oocytes derived from indicus cattle (Zebu, Bos indicus) when compared to oocytes from British or Continental taurus cattle (Bos taurus) both beef and dairy breeds [58,59]. Indicus cattle typically have more antral follicles, facilitating recovery of oocytes suitable for IVEP [60]. Overall, IVEP outcomes are more efficient in indicus than taurus due to greater number of recovered oocytes and blastocysts produced per OPU session [58,59,61].
5. Conclusions
This study advanced our knowledge to apply an evidence-based approach for OPU-IVP to formulate culture media that supported embryo development and post-thaw survival. Moreover, this study yielded new information that there were no specific negative consequences of repeated OPU sessions on general or reproductive health of donors. This information is highly relevant before applying this procedure for intense production of embryos from our superior animals to alleviate producer concerns regarding animal health and welfare.
Author Contributions
Conceptualization, J. Thundathil; Methodology, A. Guest, C. Johnson, J. Thundathil; Formal Analysis, A. Guest, C. Johnson; Investigation, A. Guest, C. Johnson, J. Thundathil; Resources, J. Thundathil, J.P. Kastelic; Writing – Original Draft Preparation, F. Salek; Writing – Review & Editing, F. Salek, J. Thundathil, J.P. Kastelic; Supervision, J. Thundathil; Project Administration, J. Thundathil; Funding Acquisition, J. Thundathil, J.P. Kastelic.
Funding
This work was supported by funding from the Natural Sciences and Engineering Research Council (NSERC) of Canada (Grant No. RGPIN-2020–04585).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We acknowledge the animal resource support received the DBK Genetics, Inc., AB, and the OPU service (fee for a service basis) received from the Bova-Tech Ltd., Rocky View, AB. An abstract of this study was presented at the ICAR 2022, Bologna, Italy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Daly, J.; Smith, H.; McGrice, H.A.; Kind, K.L.; Van Wettere, W.H.E.J. Towards Improving the Outcomes of Assisted Reproductive Technologies of Cattle and Sheep, with Particular Focus on Recipient Management. Animals 2020, 10, 293. [CrossRef]
- Humblot, P.; Le Bourhis, D.; Fritz, S.; Colleau, J.J.; Gonzalez, C.; Guyader Joly, C.; Malafosse, A.; Heyman, Y.; Amigues, Y.; Tissier, M.; et al. Reproductive Technologies and Genomic Selection in Cattle. Vet. Med. Int. 2010, 2010, 1–8. [CrossRef]
- Bó, G.A.; Cedeño, A.; Mapletoft, R.J. Strategies to Increment in Vivo and in Vitro Embryo Production and Transfer in Cattle. Anim. Reprod. 2019, 16, 411–422. [CrossRef]
- Ferré, L.B.; Kjelland, M.E.; Strøbech, L.B.; Hyttel, P.; Mermillod, P.; Ross, P.J. Review: Recent Advances in Bovine in Vitro Embryo Production: Reproductive Biotechnology History and Methods. Animal 2020, 14, 991–1004. [CrossRef]
- Davis, T.C.; White, R.R. Breeding Animals to Feed People: The Many Roles of Animal Reproduction in Ensuring Global Food Security. Theriogenology 2020, 150, 27–33. [CrossRef]
- Viana, J.H. 2021 Statistics of Embryo Production and Transfer in Domestic Farm Animals. N. Am. 2022.
- Greenwood, P.L. Review: An Overview of Beef Production from Pasture and Feedlot Globally, as Demand for Beef and the Need for Sustainable Practices Increase. Animal 2021, 15, 100295. [CrossRef]
- Cordeiro, M.R.C.; Mengistu, G.F.; Pogue, S.J.; Legesse, G.; Gunte, K.E.; Taylor, A.M.; Ominski, K.H.; Beauchemin, K.A.; McGeough, E.J.; Faramarzi, M.; et al. Assessing Feed Security for Beef Production within Livestock-Intensive Regions. Agric. Syst. 2022, 196, 103348. [CrossRef]
- Leroy, J.L.M.R.; Rizos, D.; Sturmey, R.; Bossaert, P.; Gutierrez-Adan, A.; Van Hoeck, V.; Valckx, S.; Bols, P.E.J. Intrafollicular Conditions as a Major Link between Maternal Metabolism and Oocyte Quality: A Focus on Dairy Cow Fertility. Reprod. Fertil. Dev. 2012, 24, 1. [CrossRef]
- Ribeiro, E.S.; Gomes, G.; Greco, L.F.; Cerri, R.L.A.; Vieira-Neto, A.; Monteiro, P.L.J.; Lima, F.S.; Bisinotto, R.S.; Thatcher, W.W.; Santos, J.E.P. Carryover Effect of Postpartum Inflammatory Diseases on Developmental Biology and Fertility in Lactating Dairy Cows. J. Dairy Sci. 2016, 99, 2201–2220. [CrossRef]
- Lopez, H.; Caraviello, D.Z.; Satter, L.D.; Fricke, P.M.; Wiltbank, M.C. Relationship between Level of Milk Production and Multiple Ovulations in Lactating Dairy Cows. J. Dairy Sci. 2005, 88, 2783–2793. [CrossRef]
- Baruselli, P.S.; Souza, A.H.D.; Sá, M.F.D.; Marques, M.O.; Sales, J.N.D.S. Genetic Market in Cattle (Bull, Ai, Ftai, Moet and Ivp): Financial Payback Based on Reproductive Efficiency in Beef and Dairy Herds in Brazil. Anim. Reprod. 2018, 15, 247–255. [CrossRef]
- Demetrio, D.G.B.; Benedetti, E.; Demetrio, C.G.B.; Fonseca, J.; Oliveira, M.; Magalhaes, A.; Santos, R.M.D. How Can We Improve Embryo Production and Pregnancy Outcomes of Holstein Embryos Produced in Vitro? (12 Years of Practical Results at a California Dairy Farm). Anim. Reprod. 2020, 17, e20200053. [CrossRef]
- Soares, J.G.; Martins, C.M.; Carvalho, N.A.T.; Nicacio, A.C.; Abreu-Silva, A.L.; Campos Filho, E.P.; Torres Júnior, J.R.S.; Sá Filho, M.F.; Baruselli, P.S. Timing of Insemination Using Sex-Sorted Sperm in Embryo Production with Bos Indicus and Bos Taurus Superovulated Donors. Anim. Reprod. Sci. 2011, 127, 148–153. [CrossRef]
- Macmillan, K.; Lean, I.; Westwood, C. The Effects of Lactation on the Fertility of Dairy Cows. Aust. Vet. J. 1996, 73, 141–147. [CrossRef]
- Snijders, S.E.M.; Dillon, P.; O’Callaghan, D.; Boland, M.P. Effect of Genetic Merit, Milk Yield, Body Condition and Lactation Number on in Vitro Oocyte Development in Dairy Cows. Theriogenology 2000, 53, 981–989. [CrossRef]
- Ferré, L.B.; Alvarez-Gallardo, H.; Romo, S.; Fresno, C.; Stroud, T.; Stroud, B.; Lindsey, B.; Kjelland, M.E. Transvaginal Ultrasound-guided Oocyte Retrieval in Cattle: State-of-the-art and Its Impact on the in Vitro Fertilization Embryo Production Outcome. Reprod. Domest. Anim. 2023, 58, 363–378. [CrossRef]
- Zangirolamo, A.F.; Morotti, F.; Silva, N.C.D.; Sanches, T.K.; Seneda, M.M. Ovarian Antral Follicle Populations and Embryo Production in Cattle. Anim. Reprod. 2018, 15, 310–315. [CrossRef]
- Chaubal, S.A.; Molina, J.A.; Ohlrichs, C.L.; Ferre, L.B.; Faber, D.C.; Bols, P.E.J.; Riesen, J.W.; Tian, X.; Yang, X. Comparison of Different Transvaginal Ovum Pick-up Protocols to Optimise Oocyte Retrieval and Embryo Production over a 10-Week Period in Cows. Theriogenology 2006, 65, 1631–1648. [CrossRef]
- Roover, R.D.; Genicot, G.; Leonard, S.; Bols, P.; Dessy, F. Ovum Pick up and in Vitro Embryo Production in Cows Superstimulated with an Individually Adapted Superstimulation Protocol. Anim. Reprod. Sci. 2005, 86, 13–25. [CrossRef]
- Chastant-Maillard, S.; Quinton, H.; Lauffenburger, J.; Cordonnier-Lefort, N.; Richard, C.; Marchal, J.; Mormede, P.; Renard, J. Consequences of Transvaginal Follicular Puncture on Well-Being in Cows. Reproduction 2003, 555–563. [CrossRef]
- Van Wagtendonk-de Leeuw, A.M. Ovum Pick Up and In Vitro Production in the Bovine after Use in Several Generations: A 2005 Status. Theriogenology 2006, 65, 914–925. [CrossRef]
- Blondin, P.; Bousquet, D.; Twagiramungu, H.; Barnes, F.; Sirard, M.-A. Manipulation of Follicular Development to Produce Developmentally Competent Bovine Oocytes1. Biol. Reprod. 2002, 66, 38–43. [CrossRef]
- Bó, G.A.; Mapletoft, R.J. Historical Perspectives and Recent Research on Superovulation in Cattle. Theriogenology 2014, 81, 38–48. [CrossRef]
- Landry, D.A.; Bellefleur, A.-M.; Labrecque, R.; Grand, F.-X.; Vigneault, C.; Blondin, P.; Sirard, M.-A. Effect of Cow Age on the in Vitro Developmental Competence of Oocytes Obtained after FSH Stimulation and Coasting Treatments. Theriogenology 2016, 86, 1240–1246. [CrossRef]
- Vieira, L.M.; Rodrigues, C.A.; Castro Netto, A.; Guerreiro, B.M.; Silveira, C.R.A.; Moreira, R.J.C.; Sá Filho, M.F.; Bó, G.A.; Mapletoft, R.J.; Baruselli, P.S. Superstimulation Prior to the Ovum Pick-up to Improve in Vitro Embryo Production in Lactating and Non-Lactating Holstein Cows. Theriogenology 2014, 82, 318–324. [CrossRef]
- Sirard, M.-A. 40 Years of Bovine IVF in the New Genomic Selection Context. Reproduction 2018, 156, R1–R7. [CrossRef]
- Petyim, S.; Båge, R.; Madej, A.; Larsson, B. Ovum Pick-up in Dairy Heifers: Does It Affect Animal Well-being? Reprod. Domest. Anim. 2007, 42, 623–632. [CrossRef]
- Pieterse, M.C.; Kappen, K.A.; Kruip, Th.A.M.; Taverne, M.A.M. Aspiration of Bovine Oocytes during Transvaginal Ultrasound Scanning of the Ovaries. Theriogenology 1988, 30, 751–762. [CrossRef]
- Kruip, Th.A.M.; Den Daas, J.H.G. In Vitro Produced and Cloned Embryos: Effects on Pregnancy, Parturition and Offspring. Theriogenology 1997, 47, 43–52. [CrossRef]
- Thompson, J.G.; Peterson, A.J. Bovine Embryo Culture in Vitro: New Developments and Post-Transfer Consequences. Hum. Reprod. 2000, 15, 59–67. [CrossRef]
- Gardner, D.K.; Lane, M. Development of Viable Mammalian Embryos in Vitro. In Principles of Cloning; Elsevier, 2002; pp. 187–213 ISBN 978-0-12-174597-4.
- Marsico, T.V.; Camargo, J.D.; Valente, R.S.; Sudano, M.J. Embryo Competence and Cryosurvival: Molecular and Cellular Features. Anim. Reprod. 2019, 16, 423–439. [CrossRef]
- Lonergan, P.; Rizos, D.; Gutierrez-Adan, A.; Fair, T.; Boland, M. Oocyte and Embryo Quality: Effect of Origin, Culture Conditions and Gene Expression Patterns. Reprod. Domest. Anim. 2003, 38, 259–267. [CrossRef]
- Tharasanit, T.; Thuwanut, P. Oocyte Cryopreservation in Domestic Animals and Humans: Principles, Techniques and Updated Outcomes. Animals 2021, 11, 2949. [CrossRef]
- Magdanz, V.; Boryshpolets, S.; Ridzewski, C.; Eckel, B.; Reinhardt, K. The Motility-Based Swim-up Technique Separates Bull Sperm Based on Differences in Metabolic Rates and Tail Length. PLOS ONE 2019, 14, e0223576. [CrossRef]
- Stringfellow, D.A.; Givens, M. Manual of the International Embryo Transfer Society (IETS): A Procedural Guide and General Information for the Use of Embryo Transfer Technology Emphasizing Sanitary Procedures; 4th ed.; International Embryo Transfer Society, 2010; Vol. Savory, Ill Champaign, IL: IETS;
- Youngs, C.R. Cryopreservation of Preimplantation Embryos of Cattle, Sheep, and Goats. J. Vis. Exp. 2011, 2764. [CrossRef]
- De Roover, R.; Bols, P.E.J.; Genicot, G.; Hanzen, Ch. Characterisation of Low, Medium and High Responders Following FSH Stimulation Prior to Ultrasound-Guided Transvaginal Oocyte Retrieval in Cows. Theriogenology 2005, 63, 1902–1913. [CrossRef]
- Hasler, J.F.; Henderson, W.B.; Hurtgen, P.J.; Jin, Z.Q.; McCauley, A.D.; Mower, S.A.; Neely, B.; Shuey, L.S.; Stokes, J.E.; Trimmer, S.A. Production, Freezing and Transfer of Bovine IVF Embryos and Subsequent Calving Results. Theriogenology 1995, 43, 141–152. [CrossRef]
- Jaton, C.; Koeck, A.; Sargolzaei, M.; Price, C.A.; Baes, C.; Schenkel, F.S.; Miglior, F. Short Communication: Genetic Correlations between Number of Embryos Produced Using in Vivo and in Vitro Techniques in Heifer and Cow Donors. J. Dairy Sci. 2016, 99, 8222–8226. [CrossRef]
- Abraham, M.C.; Ruete, A.; Brandt, Y.C.B. 260 Breed Influences Outcome of in Vitro Production of Embryos in Cattle. Reprod. Fertil. Dev. 2010, 22, 287. [CrossRef]
- Pontes, J.H.F.; Melo Sterza, F.A.; Basso, A.C.; Ferreira, C.R.; Sanches, B.V.; Rubin, K.C.P.; Seneda, M.M. Ovum Pick up, in Vitro Embryo Production, and Pregnancy Rates from a Large-Scale Commercial Program Using Nelore Cattle (Bos Indicus) Donors. Theriogenology 2011, 75, 1640–1646. [CrossRef]
- Valente, R.S.; Marsico, T.V.; Sudano, M.J. Basic and Applied Features in the Cryopreservation Progress of Bovine Embryos. Anim. Reprod. Sci. 2022, 239, 106970. [CrossRef]
- Valente, R.S.; Almeida, T.G.D.; Alves, M.F.; Paschoal, D.M.; Basso, A.C.; Sudano, M.J. Cellular and Apoptotic Status Monitoring According to the Ability and Speed to Resume Post-Cryopreservation Embryonic Development. Theriogenology 2020, 158, 290–296. [CrossRef]
- Lonergan, P.; Rizos, D.; Ward, F.; Boland, M.P. Factors Influencing Oocyte and Embryo Quality in Cattle. Reprod. Nutr. Dev. 2001, 41, 427–437. [CrossRef]
- Wrenzycki, C. Gene Expression Analysis and in Vitro Production Procedures for Bovine Preimplantation Embryos: Past Highlights, Present Concepts and Future Prospects. Reprod. Domest. Anim. 2018, 53, 14–19. [CrossRef]
- De Andrade Melo-Sterza, F.; Poehland, R. Lipid Metabolism in Bovine Oocytes and Early Embryos under in Vivo, in Vitro, and Stress Conditions. Int. J. Mol. Sci. 2021, 22, 3421. [CrossRef]
- Edidin, M. Lipids on the Frontier: A Century of Cell-Membrane Bilayers. Nat. Rev. Mol. Cell Biol. 2003, 4, 414–418. [CrossRef]
- Lin, T.; Lee, J.E.; Kang, J.W.; Oqani, R.K.; Cho, E.S.; Kim, S.B.; Il Jin, D. Melatonin Supplementation during Prolonged in Vitro Maturation Improves the Quality and Development of Poor-quality Porcine Oocytes via Anti-oxidative and Anti-apoptotic Effects. Mol. Reprod. Dev. 2018, 85, 665–681. [CrossRef]
- Pero, M.E.; Zullo, G.; Esposito, L.; Iannuzzi, A.; Lombardi, P.; De Canditiis, C.; Neglia, G.; Gasparrini, B. Inhibition of Apoptosis by Caspase Inhibitor Z-VAD-FMK Improves Cryotolerance of in Vitro Derived Bovine Embryos. Theriogenology 2018, 108, 127–135. [CrossRef]
- Dias, L.R.O.; Leme, L.O.; Sprícigo, J.F.W.; Pivato, I.; Dode, M.A.N. Effect of Delipidant Agents during in Vitro Culture on the Development, Lipid Content, Gene Expression and Cryotolerance of Bovine Embryos. Reprod. Domest. Anim. 2020, 55, 11–20. [CrossRef]
- Mortimer, D.; Cohen, J.; Mortimer, S.T.; Fawzy, M.; McCulloh, D.H.; Morbeck, D.E.; Pollet-Villard, X.; Mansour, R.T.; Brison, D.R.; Doshi, A.; et al. Cairo Consensus on the IVF Laboratory Environment and Air Quality: Report of an Expert Meeting. Reprod. Biomed. Online 2018, 36, 658–674. [CrossRef]
- Gardner, D.K.; Kelley, R.L. Impact of the IVF Laboratory Environment on Human Preimplantation Embryo Phenotype. J. Dev. Orig. Health Dis. 2017, 8, 418–435. [CrossRef]
- Consensus Group, C. ‘There Is Only One Thing That Is Truly Important in an IVF Laboratory: Everything’ Cairo Consensus Guidelines on IVF Culture Conditions. Reprod. Biomed. Online 2020, 40, 33–60. [CrossRef]
- Sartori, R.; Spies, C.; Wiltbank, M.C. Effects of Dry Matter and Energy Intake on Quality of Oocytes and Embryos in Ruminants. Reprod. Fertil. Dev. 2017, 29, 58. [CrossRef]
- Baruselli, P.S.; Ferreira, R.M.; Vieira, L.M.; Souza, A.H.; Bó, G.A.; Rodrigues, C.A. Use of Embryo Transfer to Alleviate Infertility Caused by Heat Stress. Theriogenology 2020, 155, 1–11. [CrossRef]
- Pontes, J.H.F.; Silva, K.C.F.; Basso, A.C.; Rigo, A.G.; Ferreira, C.R.; Santos, G.M.G.; Sanches, B.V.; Porcionato, J.P.F.; Vieira, P.H.S.; Faifer, F.S.; et al. Large-Scale in Vitro Embryo Production and Pregnancy Rates from Bos Taurus, Bos Indicus, and Indicus-Taurus Dairy Cows Using Sexed Sperm. Theriogenology 2010, 74, 1349–1355. [CrossRef]
- Watanabe, Y.F.; Souza, H.A.; Mingoti, R.D.; Ferreira, R.M.; Batista, E.O.S.; Dayan, A.; Watanabe, O.Y.; Meirelles, F.V.; Nogueira, M.F.G.; Ferraz, J.B.S.; et al. Number of Oocytes Retrieved per Donor during OPU and Its Relationship with in Vitro Embryo Production and Field Fertility Following Embryo Transfer. Anim. Reprod. 2017, 14, 635–644. [CrossRef]
- Batista, E.; Macedo, G.; Sala, R.; Ortolan, M.; Sá Filho, M.; Del Valle, T.; Jesus, E.; Lopes, R.; Rennó, F.; Baruselli, P. Plasma Antimullerian Hormone as a Predictor of Ovarian Antral Follicular Population in Bos Indicus (Nelore) and Bos Taurus (Holstein) Heifers. Reprod. Domest. Anim. 2014, 49, 448–452. [CrossRef]
- Guerreiro, B.M.; Batista, E.O.S.; Vieira, L.M.; Sá Filho, M.F.; Rodrigues, C.A.; Castro Netto, A.; Silveira, C.R.A.; Bayeux, B.M.; Dias, E.A.R.; Monteiro, F.M.; et al. Plasma Anti-Mullerian Hormone: An Endocrine Marker for in Vitro Embryo Production from Bos Taurus and Bos Indicus Donors. Domest. Anim. Endocrinol. 2014, 49, 96–104. [CrossRef]
Figure 1.
Donor effects on mean (± SEM) number of follicles, oocytes recovered, percentage cleavage, and percentage blastocyst production (BC) in six crossbred heifers subjected to repeated OPU-IVP cycles over 1 y (data compiled from nine trials). a,bWithin an end point, heifers without a common superscript differed (P < 0.05).
Figure 1.
Donor effects on mean (± SEM) number of follicles, oocytes recovered, percentage cleavage, and percentage blastocyst production (BC) in six crossbred heifers subjected to repeated OPU-IVP cycles over 1 y (data compiled from nine trials). a,bWithin an end point, heifers without a common superscript differed (P < 0.05).
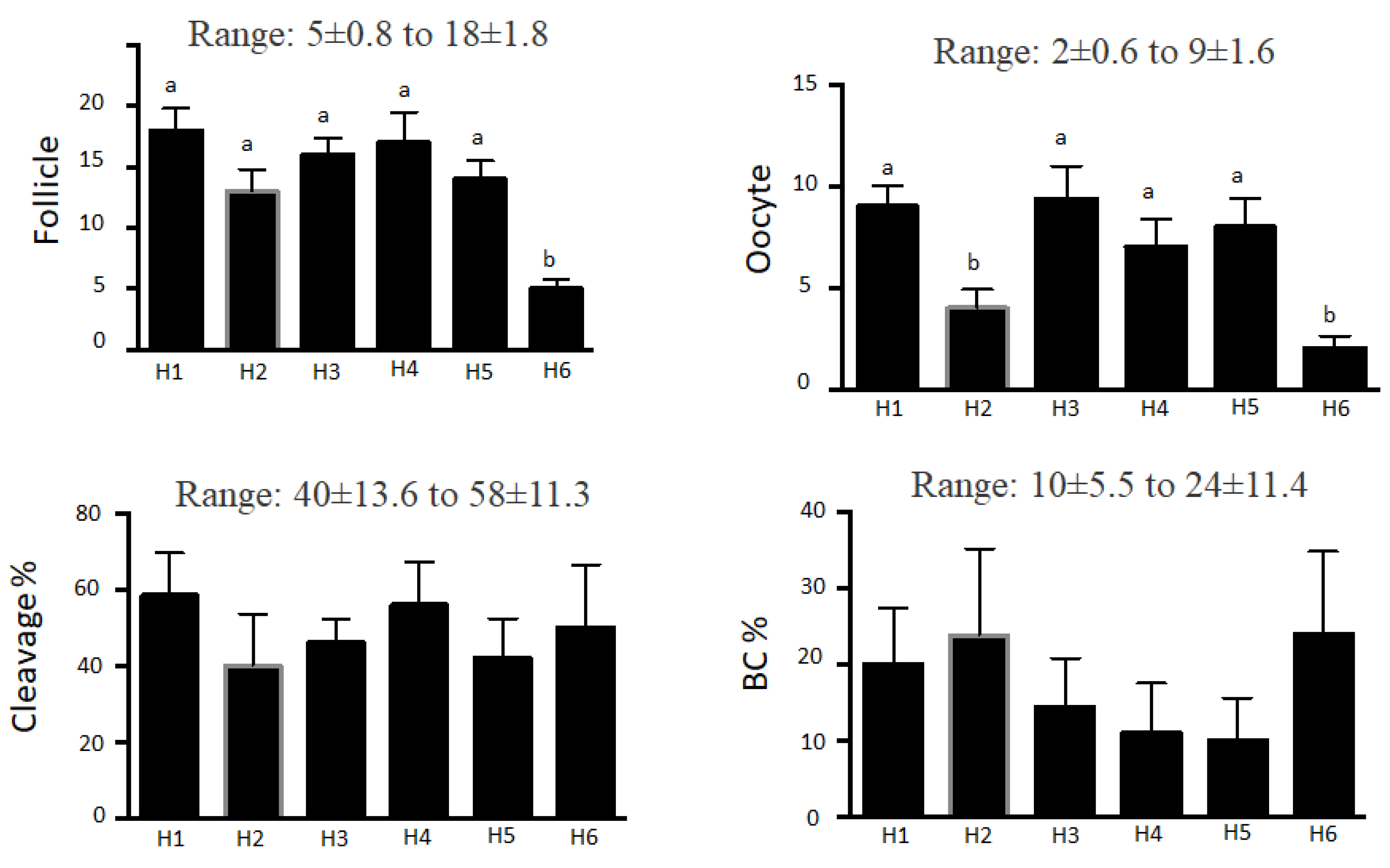

Table 1.
Mean (± SEM) effects of repeated OPU-IVP trials on follicles, oocytes recovered, and embryonic development in crossbred beef heifers (n=6).
Table 1.
Mean (± SEM) effects of repeated OPU-IVP trials on follicles, oocytes recovered, and embryonic development in crossbred beef heifers (n=6).
| Trial | No. follicles | No. oocytes | Cleavage (%) | Blastocyst rate (%) |
|---|---|---|---|---|
| 1 | 14±2.8 | 6±1.7 | 66±15 | 31±14.2 |
| 2 | 14±3.5 | 6±1.1 | 67±5.5 | 20±13.3 |
| 3 | 19±2.4 | 11±2.1 | 68±7.9 | 20±7.1 |
| 4 | 16±2.0 | 9±1.1 | 56±8.2 | 23±11.5 |
| 5 | 14±2.9 | 5±2.1 | 42±17 | 39±13.1 |
| 6 | 11±2.1 | 5±1.1 | 34±16.2 | 18±10.2 |
| 7 | 13±2.1 | 7±1.8 | 27±13 | 5.5±3.7 |
| 8 | 12±2.4 | 6±2.5 | 44±12.1 | 15±7.1 |
| 9 | 12±2.4 | 6±1.2 | 33±17.1 | 0 |
Table 2.
Mean (± SEM) effects of repeated OPU-IVP trials on follicles, oocytes recovered, and embryonic development in purebred beef cows (n=3).
Table 2.
Mean (± SEM) effects of repeated OPU-IVP trials on follicles, oocytes recovered, and embryonic development in purebred beef cows (n=3).
| Cow (No. trials) | No. follicles | No. oocytes | Cleavage (%) | Blastocyst rate (%) |
|---|---|---|---|---|
| A (2) | 56.0±1.0a | 17.5±3.5a | 54.5±16.5 | 20.0±1a |
| B (4) | 24.7±5.6b | 9.5±2.3ab | 70±15.9 | 52.5±18.3ab |
| C (4) | 9.0±1.2c | 4±0.8b | 89±6.2 | 78±7.7b |
a-c Within a column, means without a common superscript differed (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



