
Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Endophytic fungi colonize the inner tissues of plants and provide direct and indirect benefits to the plant. Although Mongolia is rich in medicinal plants, due to climatic and anthropogenic reasons the resources are being depleted and many species are under the threat of gradual extinction, while the endophytic fungi of Mongolian plants are largely unknown. Understanding the diversity and metabolic potential of endophytic fungi is of great significance in terms of the utilization of beneficial fungal strains in plant propagation and the discovery of bioactive compounds. In this study, a total of 24 culturable endophytic fungal strains were isolated from Cynomorium songaricum (Rupr.), a medicinal and vulnerable plant species of Mongolia. Based on the morphological characteristics and the sequences of the rDNA internal transcribed spacer (ITS) region, the isolates were identified to 6 genera: Fusarium (8), Clonostachys (7), Penicillium (6), Alternaria (1), Aspergillus (1), and Madurella (1). The antimicrobial activity was assessed by agar-diffusion method, revealing that 15 strains were able to inhibit the growth of at least one of the test organisms. Among them, 1 strain showed inhibitory activity against Escherichia coli, 12 against Bacillus subtilis, 13 against Staphylococcus aureus, 8 against Aspergillus niger, respectively. All fungal strains were screened for their ability to solubilize complex phosphorus and zinc minerals and for their production of indole-3-acetic acid (IAA). The strains were preserved in the national culture collection for further sustainable exploitation of their biotechnological potential.
Keywords:
fungal diversity
; ITS region
; Cynomorium songaricum
; Ulaan Goyo
; indole acetic acid
; zinc solubilization
; phosphate solubilization
; culture collection
1. Introduction
To implement the Convention on Biological Diversity [1] and the Law on Genetic resources of Mongolia, which entered into force on the 30th December 2021 [2], the establishment and maintenance of facilities for ex-situ conservation of biological diversity, research and management of the collection of biological resources, especially microorganisms, are in demand in Mongolia. As the nation's growing and largest culture collection, we are committed to exploring, preserving, and supplying microbial resources and associated data for research, education and industrial applications. Mongolia is one of the countries most affected by climate change, with the temperature increase of 2.14°C confirmed between 1940 and 2008 [3], which can lead to the loss of biodiversity. In addition to climatic reason, overexploitation makes medicinal plants more vulnerable. Therefore, we focus on endophytic microorganisms, isolate the key culturable representatives residing the medicinal, endangered and vulnerable plants in Mongolia, preserve them in the culture collection, and explore their biotechnological potential for further sustainable use.
Endophytic fungi are microorganisms that live in internal plant tissue for at least a certain period of their life cycle without causing harm to the host plant under any circumstance [4]. They are well known to provide direct (nutrient acquisition and phytohormone production) and indirect (activation of systemic resistance, production of secondary metabolites and protection for abiotic and biotic stresses) benefits to the host plant [5]. The direct benefits of endophytic fungi result in enhanced root development, increased plant height, biomass production and overall yield, hence they can be referred to as biofertilizers [6]. Endophytes can improve the uptake of both macronutrients such as phosphorus, nitrogen and potassium and micronutrients such as zinc, iron and copper from soil and organic matter [7]. Phosphorus is the second important nutrient for overall plant development and productivity. It required for biological growth and development of plants and also performs a significant part in metabolic processes including photosynthesis, transfer of energy, signal transduction and respiration [8]. However, its structural and chemical characteristics make it a limiting nutrient for plant growth since they reduce its free availability. Zinc is one of the essential micronutrients for plants, and a deficiency of this element causes a slowdown in a number of processes, such as photosynthesis and the synthesis of biomolecules (carbohydrates, RNA and proteins), which leads to a decrease in the quality of the crop [9]. Zinc-solubilizing microorganisms can solubilize the inaccessible form of zinc by secreting organic acids, siderophores and other chelating compounds, but this property is well known to bacteria and, to a lesser extent, fungi [7,9]. Endophytic fungi produce phytohormones such as auxins, gibberellins (GAs) and cytokinins. A kind of auxin known as indole-3-acetic acid (IAA) generally affects plant cell division, nodule formation, seed initiation and tuber germination, development of xylem and root, photosynthesis, and provides resistance to plants against various stressful conditions and is the main auxin produced by endophytes [5,7].
Over the past years, endophytic fungi have attracted attention due to their ability to produce novel bioactive secondary metabolites and have become known as a treasure house of bioactive compounds of medicinal importance [10]. Their metabolites are progressively being studied and metabolites are categorized into various functional groups, alkaloids, benzopyranones, chinones, flavonoids, phenolic acids, quinones, steroids, saponins, tannins, terpenoids, tetralones, xanthones, and many others that serve as a potential candidate for antimicrobial, anti-insect, anticancer and many more properties [11,12,13,14]. However, it is estimated that only 1–2% of approximately 300,000 plant species have been studied, meaning that the vast majority of endophytic fungal symbiotic relationships remain unexplored [15].
Cynomorium songaricum Rupr., called Ulaan Goyo or Zuungariin Goyo in Mongolia, is a medicinal, parasitic and rare plant species distributed in southern Mongolia and northwest Inner Mongolia in China [16]. This plant usually parasitizes on the roots of Nitraria tangutorum Bobr. and Nitraria sibirica Pall located in dry sandy regions, and it is widely used as a functional food and medicine in traditional Mongolian medicine and in traditional Chinese medicine [17]. In the traditional Mongolian medicine, it has been used to treat kidney diseases, high blood pressure, liver and bile dysfunction, diabetes, weakness, dropsy, nervousness and constipation [18]. Numerous studies on the bioactive compounds and functions of C. songaricum have been conducted worldwide and Cui et al. summarized them and reported that at least 76 biologically active compounds have been isolated and identified from this amazing plant species, including flavonoids, terpenoids, steroids, organic acids, saccharides, glycosides and phloroglucinol adducts. These compounds have pharmacological functions of this plant such as anti-aging, anti-oxidation, anti-fatigue and anti-HIV effects, as well as effects on the immune system, nervous system, reproductive system and other biological activities [17]. Despite extensive research of the bioactive compounds and pharmacological actions of the plant, a study on the distribution and dynamics of endophytic fungi in C. songaricum and its host N. tangutorum was first reported in 2018, suggesting a possible exchange of endophytic fungi between C. songaricum and its host N. tangutorum [19]. Further investigations revealed significant correlations between differential secondary metabolites and endophytic fungi in C. songaricum distributed across different locations [20], and moreover, plant species and lifestyle, as well as local environment strongly influenced the abundance and diversity of the endophytic fungal species in C. songaricum and its host N. tangutorum [21].
In this study, culturable endophytic fungal strains were isolated and identified from Cynomorium songaricum Rupr. parasitizing on the roots of Nitraria sibirica Pall, and their antimicrobial activity as well as their plant growth-promoting properties, such as phosphate solubilization, zinc solubilization and IAA production were determined.
2. Materials and Methods
2.1. Collection of Plant Samples
The medicinal and vulnerable plant samples of Cynomorium songaricum Rupr. were collected in July 2021 in Govi-Altai province (45°34’43” N, 98°13’12” E, h1730m), Mongolia. The aboveground and underground parts, fresh and old rhizomes on the root of host plant Nitraria sibirica Pall. were collected (Figure 1).
2.2. Isolation of Endophytic Fungi
Plant samples were subjected to a three-step surface sterilization procedure according to the method described by Thi Minh Le et al. [22] with minor modification. Portions of healthy parts and rhizomes were washed thoroughly under running tap water to remove adhered debris, and then each sample was sterilized sequentially by washing with 70% ethanol for 1 min, 3% sodium hypochlorite for 2-4 min, and 70% ethanol for 1 min, rinsed three times in sterile distilled water. After drying on sterile filter paper, each sample was cut into small pieces of less than 1 cm in size with a sterile scalpel and placed on potato dextrose agar (PDA, Biolab Diagnostics Laboratory Inc., Hungary) supplemented with 50 mg/L chloramphenicol to suppress bacterial growth. All plates were incubated at 28°C and the growth of endophytic fungal hyphae emerging from the segments was monitored daily for up to 3 weeks. Emerging fungi were transferred to fresh PDA plates, incubated for 1-2 weeks, and periodically checked for purity. The effectiveness of the surface sterilization procedure was ascertained by spreading 200 mL of the last wash water on the agar plates and incubating at 28°C for 1 week to check for microbial growth. The pure cultures were preserved in glycerol suspensions (20%, V/V) at -80°C.
All isolated fungi were deposited into the Mongolian National Culture Collection of Microorganisms (MNCCM), Institute of Biology, Mongolian Academy of Sciences.
2.3. Identification of the Isolates
Fungal isolates were cultured on Low carbon agar (LCA) medium composed of glucose 1 g/L, potassium dihydrogen phosphate 1 g/L, magnesium sulphate 0.2 g/L, potassium chloride 0.2 g/L, sodium nitrate 2 g/L, yeast extract 0.2 g/L, and agar 15 g/L at 28°C for 7 days. The purity and monosporic cultures were confirmed by observing the isolates under an Olympus CX41 microscope (Olympus, Japan) at 40-500x magnifications, and spore-forming fungi were preliminary identified by morphological features, such as conidia, conidiophores, and hyphae.
Molecular identification was carried out using fungal isolates grown on PDA at 28°C for 7 days. Total genomic DNA was extracted using the PrepMan™ Ultra Sample Preparation Reagent (Thermo Fisher Scientific Inc., USA) according to the manufacturer’s instructions.
The internal transcribed spacer (ITS) region, 5.8S gene and the D1/D2 domain of the large subunit (LSU) ribosomal RNA (rRNA) gene was amplified using the primers ITS5 (5'-GGAAGTAAAAGTCGTAACAAGG-3') and NL4 (5'-GGTCCGTGTTTCAAGACGG-3') as described previously [23]. The amplification was performed with a FastStart Taq DNA polymerase (Roche, China) in the GeneAmp® PCR System 9700 (Thermo Fisher Scientific Inc., USA) according to the following conditions: initial denaturation at 94⁰C for 2 min, followed by 30 cycles of denaturation at 94⁰C for 1 min, annealing at 56⁰C for 30 sec, and extension at 68⁰C for 90sec; and a final extension step at 68⁰C for 5 min. The PCR products were visualized by electrophoresis on 1% agarose gel and subsequently purified using an AccuPrep® PCR/Gel Purification Kit (Bioneer, Korea), and sent to Macrogen, Korea for commercial sequencing. The sequences were analyzed by BLAST similarity search at the website of the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/BLAST) based on their identity values.
The obtained sequences were submitted to GenBank and the accession numbers are LC663161- LC663164, LC769415- LC769442.
2.4. Antimicrobial Activity Test
The endophytic fungal isolates were screened using the agar plug diffusion method for antimicrobial activity against potentially pathogenic bacteria [Escherichia coli (NBRC 102203T), Bacillus subtilis (NBRC 13719T), and Staphylococcus aureus (NBRC 100910T)] and two fungi [Candida albicans (NBRC 1385T) and Aspergillus niger (NBRC 33023T)]. Inoculums of the test bacteria (approximately 1 x 108 CFU/mL) and yeast (approximately 1 x 106 CFU/mL) were prepared by comparison with 0.5 McFarland standards, whereas inoculums of test fungi were prepared as 1 x 106 spores/mL. The test bacterial inoculums were seeded on Nutrient agar (Biolab Diagnostics Laboratory Inc., Hungary), yeast and fungal inoculums were seeded on PDA, respectively. For the preparation of the agar plug, the fungal isolates were precultured on PDA plates at 28℃ for 7 days and then the cultures were cut into plugs 6 mm in diameter, and the plugs were placed on the agar medium seeded with test microorganisms. The plates were incubated at either 37°C (bacteria and yeast) or 30°C (fungi). Ampicillin sodium (50 µg/disk) and cycloheximide (25 µg/disk) were taken as positive controls for bacteria and fungi, respectively. Antimicrobial activity was assessed by the size (diameter in mm) of sterile zones formed around the fungal agar plugs [24,25,26,27].
2.5. Phosphate and Zinc oxide Solubilization Assay
Screening of fungal isolates for the ability to solubilize phosphate and zinc was carried out on Pikovskaya’s (PKV) agar medium and on mineral salts agar medium amended with 0.1% of insoluble ZnO, respectively, as described previously [28]. Agar plugs (6 mm), cut from 14 days old culture of endophytic fungi, were placed on the respective plates in triplicate and incubated at 28°C. The clear zones formed around the colonies were measured after the incubation period, 4 days and 14 days after inoculation, for the efficiency of solubilization of phosphorus and zinc, respectively.
2.6. IAA Production Assay
The production of indole acetic acid (IAA) of endophytic fungi was determined using the following colorimetric assay. Fungal isolates were incubated in 5mL PDB supplemented with 5mM L-tryptophan at 28℃ for 5 days. After cultivation, each culture was centrifuged at 5000 rpm for 10 min, and 1 mL of the clear supernatant was mixed with 2 mL of Salkowski reagent (1 mL 0.5 M FeCl3, 50 mL 35% perchloric acid) and the mixture was incubated in the dark at room temperature for 20 min. A mixture of PDB medium with 5mM L-tryptophan and Salkowski reagent was used as a control. Development of pink color indicated IAA production and the pink to red color produced by the isolates was categorized into low, medium, and high [8]. The absorbance of a positive reaction was determined with a UV-VIS spectrophotometer at 530 nm and the amount of IAA product was calculated from a standard graph prepared using known quantities of pure IAA [29].
3. Results
3.1. Isolation and Identification of Endophytic Fungi
In the present study, a total of 24 purified isolates of endophytic fungi were obtained from aboveground parts (4), underground parts (4), fresh rhizomes (8), and old rhizomes (8) of Cynomorium songaricum Rupr. parasitizing on the roots of Nitraria sibirica Pall, growing in sandy soil in the Govi-Altai province territory of Mongolia. They were morphologically highly diverse and mainly pigmented on PDA medium (Figure 2).
Molecular identification of fungal endophytes was performed using ITS rDNA sequences (ITS1-5.8S-ITS2) as a marker. ITS rDNA sequences from the 24 isolates were compared with sequences of organisms represented in the GenBank database. BLAST results showed that the 24 isolates belonged to 6 genera: Fusarium, Clonostachys, Penicillium, Alternaria, Aspergillus, and Madurella (Table 1). The genus Fusarium had the highest number of isolates (8), followed by Clonostachys with 7 isolates, Penicillium with 6 isolates, while the genera Alternaria, Aspergillus, and Madurella all had one isolate each.
3.2. Antimicrobial Activity
All isolates were evaluated in vitro for antimicrobial activity, and among the 24 isolates, 15 (62.5%) exhibited antimicrobial activity against at least one test microorganism whereas the rest yielded no activity. There was 1 strain having antimicrobial activity against Escherichia coli, 12 strains against Bacillus subtilis, 13 strains against Staphylococcus aureus, and 8 strains against Aspergillus niger (Table 2). Among the 15 antagonistic strains, 13 (86.6%) belonged to the genera Fusarium and Clonostachys. All 7 strains belonging to Clonostachys and 6 out 8 strains belonging to Fusarium had antimicrobial activity. Strain P26-ZN1-2 belonging to Madurella inhibited growth of E. coli along with B. subtilis and S. aureus with inhibitory zone of 7.5±0.7, 9.5±2.1 and 15.5±0.7, respectively. Moreover, 8 strains (P26-H1-3, P26-H2-2, P26-ZN1-2, P26-ZO1-3, P26-ZO1-4, P26-ZO1-5, P26-ZO1-6, P26-ZO2-3) showed a broader spectrum of antimicrobial activity (inhibition zone, or against test bacteria and fungi, both), and 2 strains (P26-H2-2, P26-R2-1) displayed strong inhibition to pathogenic fungi. None of the strains inhibited the Candida albicans test (Table 2).
3.3. In Vitro Test for Plant Growth Promoting Traits of Endophytes
The endophytic isolates were further studied for their plant growth-promoting traits, including IAA production, phosphate solubilization, and zinc oxide solubilization. All 28 strains exhibited positive results for one or more traits, IAA production was seen in 9 strains in PDB medium supplemented with 5mM L-Tryptophan (Figure 3), whereas phosphate solubilization, and zinc oxide solubilization activity was noticed in 3, and 21 strains respectively. Three Penicillium strains, P26-ZN2-3, P26-ZN2-5, and P26-ZO1-1, were positive for both phosphate and zinc oxide solubilization traits, however no production of IAA was detected in the presence and absence of L-Tryptophan. Penicillium strain P26-ZN2-5 showed the most significant phosphate solubilizing activity on a solid PVK medium with the solubilization index (SI) (2.16 ± 0.02 cm). This strain also showed the most significant zinc solubilizing activity on mineral salt agar with the solubilization index (SI) (3.4 ± 0.1 cm).
4. Discussion
It has been shown that the diversity of endophytic fungi is influenced by the plant genotype [30,31] or both the host genotype and geography combined [32]. A recent study by Miao et al. on speciation and genetic diversity of endophytic fungi from their host plants, C. songaricum, parasitic N. tangutorum and non-parasitic N. tangutorum at three geographic locations, found that only 0.41% to 4.48% of endophytic fungal species were shared between their host plants, consistent with previous studies indicating that the plant genotype strongly affects the endophytic fungal composition [21].
A possible exchange of endophytic fungi between C. songaricum and its host N. tangutorum was previously suggested [19], so the endophytic fungal composition of C. songaricum parasitizing another host, N. sibirica Pall, growing in the Gobi Desert was of great interest. Although it is known that culture-based methods do not reflect the real diversity of fungi in a niche due to artificial selection pressure and some of these microorganisms cannot be cultivated under laboratory conditions [19,33,34] culturable endophytes are potential source for applications in biotechnology, medicine, agriculture and beyond.
In our study, representatives of the taxa Fusarium, Clonostachys, and Penicillium were predominated; representatives of Alternaria, Aspergillus, and Madurella were less prominent. Indeed, Fusarium spp. Penicillium spp., and Aspergillus spp. were the most abundant fungi isolated from C. songaricum parasitizing on the roots of N. tangutorum [19], and among the fungal genera from plants reported in general [11,35,36].
The second prevalent genus isolated from C. songaricum parasitizing the roots of N. sibirica was Clonostachys, with 7 isolates having 99.13–100% similarity to Clonostachys rosea based on ITS sequences. Clonostachys rosea is a well-recognized mycoparasite whose hyphae penetrate and destroy those of many host fungi, and there are several commercial products based on C. rosea available for biocontrol applications worldwide [37,38]. Moreover, Clonostachys fungi produce at least 229 secondary metabolites such as nitrogen-containing metabolites, polyketides and terpenoids, many of which exhibit biological activities such as cytotoxic, antimicrobial, antileishmanial, antimalarial activity [39]. Our isolates, assigned to the genera Fusarium and Clonostachys, exhibited high antibacterial and antifungal activity.
Representatives of the genus Penicillium were isolated exclusively from fresh rhizomes. They did not show antimicrobial activities, but solubilized both phosphate and zinc. Penicillium spp. are known to be excellent solubilizers of phosphate [41–43], and zinc solubilizing efficiency in less extent [44]. The Penicillium spp. solubilizing minerals found in fresh rhizomes suggest that they may play a role in promoting growth.
One isolate, P26-ZN1-2, was classified into the genus of Madurella. Species of Madurella are the most common agents of black-grain mycetoma [45]. However, in recent years, several Madurella strains have been isolated from different plant species as endophytes [46–48]. Madurella strain P26-ZN1-2 had antibacterial activity against Gram-negative and Gram-positive test bacteria, and produced the highest amount of IAA.
Our study provides the first insight into the cultivable endophytic fungal composition of C. songaricum, a rare medicinal plant parasitizing the roots of N. sibirica growing in the Gobi Desert of Mongolia. The resulting fungi, which have antimicrobial and plant growth-promoting properties, can be used to further exploit their biotechnological potential and be applied to propagate endangered and vulnerable medicinal plants.
Author Contributions
Conceptualization, E.J.; methodology, A.M., B.B., D.N., E.J.; investigation, A.M., B.B., D.N., E.J.; writing—original draft preparation, E.J., A.M., B.B.; writing—review and editing, E.J.; visualization, E.J., B.B.; formal analysis, A.M., B.B., D.N., E.J., data curation, B.B., D.N.; supervision, E.J.; project administration, E.J.; funding acquisition, E.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was commissioned by the Ministry of Environment and Tourism of Mongolia and funded by Mongolian Science and Technology Foundation, Project Agreement No. ShuUz_2020/01.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Dr. Gundegmaa Vanjil, lecturer at Mongolian State University of Education, for plant authentication, Galbadrakh Ayush, traditional medicine practitioner of Chandmani soum of Govi-Altai province, for his help in sampling of medicinal plants, Lkhagvadorj Lodonsharav, driver, for safe and comfortable driving.
Conflicts of Interest
The authors declare no conflict of interest.
References
- CBD Convention on Biological Diversity. United Nations, 1992. Available online: https://www.cbd.int/doc/legal/cbd-en.pdf (accessed on 11 March 2020).
- Law on Genetic Resources of Mongolia. 2021. Available online: https://legalinfo.mn/mn/detail?lawId=16390399395691.
- Climate Change in Mongolia. Outputs from GCM. M. Available online: https://www.env.go.jp/content/900448010.pdf.
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of endophytes for sustainable agricultural intensification. Mol Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: a tool for plant growth promotion and sustainable agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Wang, L. Fungal Endophytes: Beyond Herbivore Management. Front Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic microbes: biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Anton Leeuw Int J G. 2020, 113, 1075–1107. [Google Scholar] [CrossRef]
- Khan, M.S. , Zaidi, A., Ahemad, M., Oves, M., Wani, P.A. Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch Agron Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Upadhayay, V.K.; Singh, A.V.; Khan, A.; Sharma, A. Contemplating the role of zinc-solubilizing bacteria in crop biofortification: an approach for sustainable bioeconomy. Front Agron. 2022, 72, 1–13. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A Treasure House of Bioactive Compounds of Medicinal Importance. Front Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 2003. 67, 491–502. [Google Scholar] [CrossRef]
- Alam, B.; Lǐ, J.; Gě, Q.; Khan, M.A.; Gōng, J.; Mehmood, S.; Yuán, Y.; Gǒng, W. Endophytic Fungi: From Symbiosis to Secondary Metabolite Communications or Vice Versa? Front. Plant Sci. 2021, 12, 791033. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.J.; Palombo, E.A.; Moulton, S.E.; Zaferanloo, B. Exploring the Promise of Endophytic Fungi: A Review of Novel Antimicrobial Compounds. Microorganisms 2022, 10, 1990. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Kaur, T.; Chhabra, I.; Panja, A.; Paul, S.; Kumar, V.; Malik, T. Endophytic fungi: hidden treasure chest of antimicrobial metabolites interrelationship of endophytes and metabolites. Front Microbiol. 2023, 14, 1227830. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G. The emergence of endophytic microbes and their biological promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Tuvaanjav, S.; Shuqin, H.; Komata, M.; Ma, C.J.; Kanamoto, T.; Nakashima, H.; Yoshida, T. Isolation and antiviral activity of water-soluble Cynomorium songaricum Rupr. polysaccharides. J Asian Nat Prod Res. 2016, 18(2), 159–171. [Google Scholar] [CrossRef] [PubMed]
- 17. Cui, J-L.; Gong, Y.; Xue, X-Z.; Zhang, Y-Y; Wang, M-L.; Wang, J-H. A Phytochemical and Pharmacological Review on Cynomorium songaricum as Functional and Medicinal Food. Nat. Prod. Commun. 2018. 13. [CrossRef]
- Ligaa, U.; Davaasuren, B.; Ninjin, N. Medicinal plants of Mongolia used in Western and Eastern medicine (in Mongolian language), JKC printing: Ulaanbaatar, Mongolia, 2006; pp. 228–229.
- Cui, J.L.; Vijayakumar, V.; Zhang, G. Partitioning of Fungal Endophyte Assemblages in Root-Parasitic Plant Cynomorium songaricum and Its Host Nitraria tangutorum. Front Microbiol. 2018, 9, 666. [Google Scholar] [CrossRef]
- Cui, J.L.; Gong, Y.; Vijayakumar, V.; Zhang, G.; Wang, M.L.; Wang, J.H.; Xue, X.Z. Correlation in chemical metabolome and endophytic mycobiome in Cynomorium songaricum from different desert locations in China. J Agr Food Chem. 2019, 67, 3554–3564. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.M.; Zhang, Y.Y.; Cui, J.L.; Zhang, G. Species and geographic specificity between endophytic fungi and host supported by parasitic Cynomorium songaricum and its host Nitraria tangutorum distributed in desert. Arch Microbiol. 2021, 203, 2511–2519. [Google Scholar] [CrossRef]
- Thi Minh Le, T.; Thi Hong Hoang, A.; Thi Bich Le, T.; Thi Bich Vo, T.; Van Quyen, D.; Hoang Chu, H. Isolation of endophytic fungi and screening of Huperzine A–producing fungus from Huperzia serrata in Vietnam. Sci Rep. 2019, 9, 16152. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Lv, Y.L.; Zhang, F.S.; Chen, J.; Cui, J.L.; Xing, Y.M.; Li, X.D.; Guo, S.X. Diversity and antimicrobial activity of endophytic fungi associated with the alpine plant Saussurea involucrata. Biol Pharm Bull. 2010, 33, 1300–1306. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J Pharm Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Tan, Xm.; Zhou, Yq.; Zhou, Xl. Diversity and bioactive potential of culturable fungal endophytes of Dysosma versipellis; a rare medicinal plant endemic to China. Sci Rep. 2018, 8, 5929. [Google Scholar] [CrossRef]
- Ibrahim, D.; Lee, C.C.; Sheh-Hong, L. Antimicrobial Activity of Endophytic Fungi Isolated from Swietenia macrophylla Leaves. Nat. Prod. Commun. 2014, 9, 1934578X1400900. [Google Scholar] [CrossRef]
- Senthil Kumar, C.M.; Jacob, T.K.; Devasahayam, S.; Thomas, S.; Geethu, C. Multifarious plant growth promotion by an entomopathogenic fungus Lecanicillium psalliotae. Microbiol. Res. 2018, 207, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Rajini, S.B.; Nandhini, M.; Udayashankar, A.C.; Niranjana, S.R.; Lund, O.S.; Prakash, H.S. Diversity, plant growth-promoting traits, and biocontrol potential of fungal endophytes of Sorghum bicolor. Plant Pathol. [CrossRef]
- Saikkonen, K. Forest structure and fungal endophytes. Fungal Biol. Rev. 2007, 21, pp–67. [Google Scholar] [CrossRef]
- Mohammad Golam Dastogeer, K.; Oshita, Y.; Yasuda, M.; Kanasugi, M.; Matsuura, E.; Xu, Q.; Okazaki, S. Host Specificity of Endophytic Fungi from Stem Tissue of Nature Farming Tomato (Solanum lycopersicum Mill.) in Japan. Agronomy 2020, 10, 1019. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Li, H.; Sivasithamparam, K.; Jones, M.G.K.; Wylie, S.J. Host Specificity of Endophytic Mycobiota of Wild Nicotiana Plants from Arid Regions of Northern Australia. Microbial ecology 2018, 75, 74–87. [Google Scholar] [CrossRef]
- Fan, S., Miao, L., Li, H., Lin, A., Song, F., Zhang, P. Illumina-based analysis yields new insights into the diversity and composition of endophytic fungi in cultivated Huperzia serrata. PLoS ONE 2020, 15 (11), e0242258. [CrossRef]
- Lucking, R.; Aime, M.C. , Robbertse, B. et al. Unambiguous identification of fungi: where do we stand and how accurate and precise is fungal DNA barcoding? IMA fungus 2020, 11, 14. [Google Scholar] [CrossRef]
- Paul, N.C.; Yu, S.H. Endophytic fungi from medicinal plants in Korea. Lap Lambert Academic Publishing; Saarbrücken, Germany, 2011.
- Guevara-Araya, M.J.; Vilo, C.; Urzúa, A. Differences in community composition of endophytic fungi between above- and below-ground tissues of Aristolochia chilensis in an arid ecosystem. Rev. Chil. de Hist. Nat. 2020, 93, 3. [Google Scholar] [CrossRef]
- Gams, W.; Diederich, P.; Poldmaa, K. Fungicolous fungi. In: Biodiversity of Fungi: Inventory and Monitoring Methods, eds Mueller, G.M.; Bills, G.F.; Foster, M.S. Elsevier Academic Press: Burlington, MA, pp 343–392. [CrossRef]
- Dan, F.J.; Dubey, Mukesh, D.; Birgit, J., Magnus, K. Clonostachys rosea to control plant diseases. Burleigh Dodds Science Publishing: UK. [CrossRef]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys Fungi and Their Biological Activities. Journal of fungi (Basel, Switzerland) 2020, 6, 229. [Google Scholar] [CrossRef]
- Qiao, H.; Sun, X. R.; Wu, X.Q.; Li, G.E.; Wang, Z.; Li, D.W. The phosphate-solubilizing ability of Penicillium guanacastense and its effects on the growth of Pinus massoniana in phosphate-limiting conditions. Biology open 2019, 8, bio046797. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Warren, R.A.; Harvey, P.R. Phosphate solubilization by Penicillium spp. closely associated with wheat roots. Biol Fertil Soils 2004, 40, 36–43. [Google Scholar] [CrossRef]
- Qiao, H.; Sun, X.R.; Wu, X. Q.; Li, G.E.; Wang, Z.; Li, D.W. The phosphate-solubilizing ability of Penicillium guanacastense and its effects on the growth of Pinus massoniana in phosphate-limiting conditions. Biology open 2019, 8, bio046797. [Google Scholar] [CrossRef]
- Doilom, M.; Guo, J.-W.; Phookamsak, R.; Mortimer, P.E.; Karunarathna, S.C.; Dong, W.; Liao, C.-F.; Yan, K.; Pem, D.; Suwannarach, N.; et al. Screening of Phosphate-Solubilizing Fungi from Air and Soil in Yunnan, China: Four Novel Species in Aspergillus, Gongronella, Penicillium, and Talaromyces. Front. Microbiol 2020, 11(585215). [Google Scholar] [CrossRef] [PubMed]
- Devi, D.; Gupta, S.B.; Mishra, B.K.; Verma, N.P. Isolation and identification of zinc solubilizing fungal isolates from cumin of semi-arid region of Rajasthan. The Pharma Innovation 2022, 11, 1036–1040. [Google Scholar]
- de Hoog, G.S.; van Diepeningen, A.D; Mahgoub, E-S.; van de Sande, W.W.J. New Species of Madurella, Causative Agents of Black-Grain Mycetoma. J Clin Microbiol. 2012, 50, 988 – 994. [CrossRef]
- Pushpa, H.; Kavya, S.; Pooja, K.; Sneha, L.; Arer, V.O. An isolation, identification and diversity of endophytic fungi from Catharanthus roseus and screening for their L-asparaginase activity. Int J Env Ecol Fam Urban Stud 2018, 8(7). Available online: https://issuu.com/tjprc/docs/2.ijeefusdec20182.
- Hapida, Y. ; Elfita.; Widjajanti, H.; Salni. Biodiversity and antibacterial activity of endophytic fungi isolated from jambu bol (Syzygium malaccense). Biodiversitas J Biol Diversity, 5677. [Google Scholar] [CrossRef]
- Orole, O.O.; Adejumo, T.O.; Link, T.; Voegele, R.T. Molecular identification of endophytes from maize roots and their biocontrol potential against toxigenic fungi of Nigerian maize. Science progress 2023, 106, 368504231186514. [Google Scholar] [CrossRef]
Figure 1.
Habitat of the collected plant samples. Cynomorium songaricum (a) growing in sandy soil, and rhizome of Cynomorium songaricum on the root of host plant Nitraria sibirica Pall. (b).
Figure 1.
Habitat of the collected plant samples. Cynomorium songaricum (a) growing in sandy soil, and rhizome of Cynomorium songaricum on the root of host plant Nitraria sibirica Pall. (b).

Figure 2.
Morphological diversity of endophytic fungi isolated from Cynomorium songaricum.

Figure 3.
Production of IAA by endophytic fungal strains.
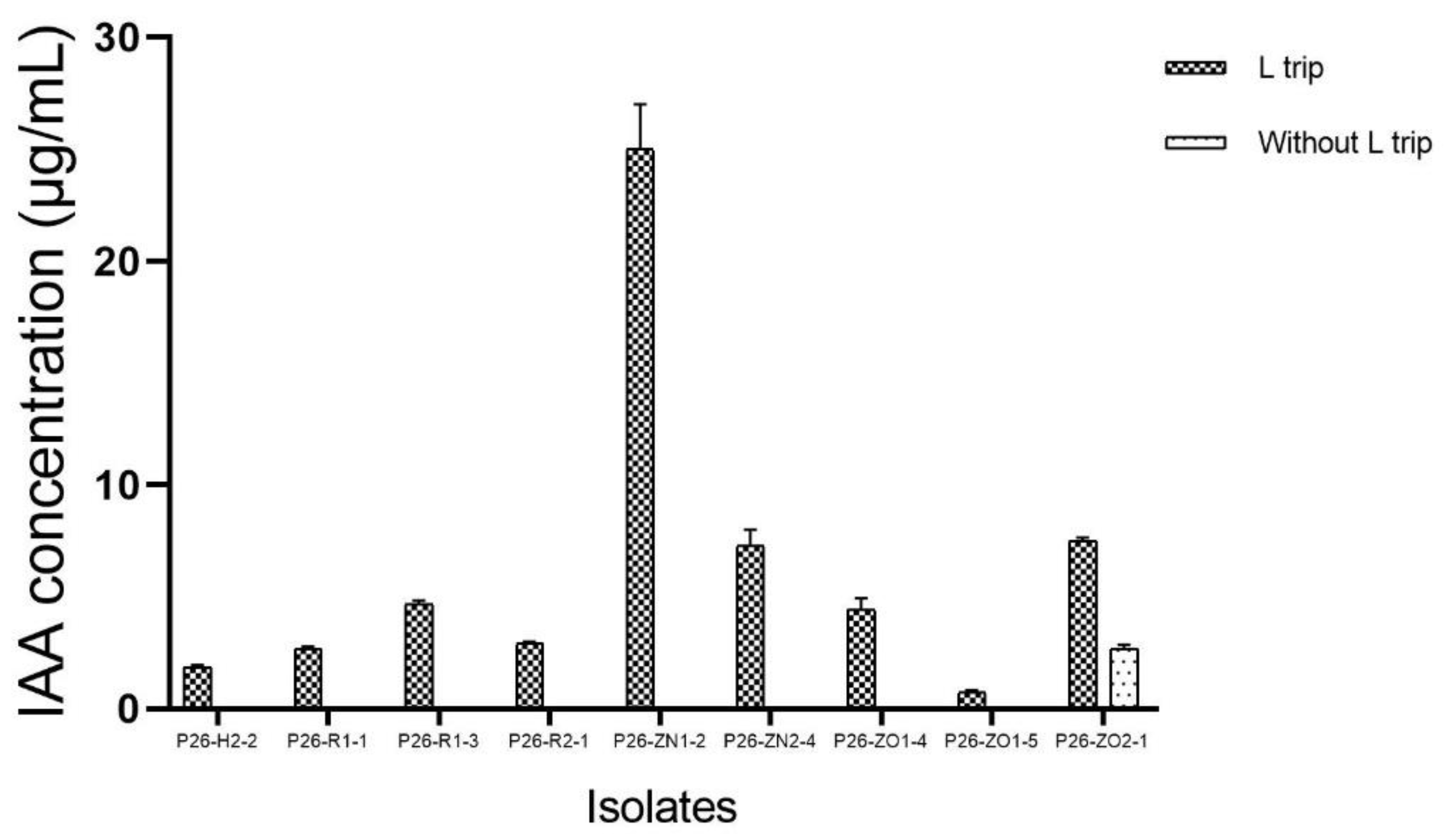
Figure 4.
Phosphate and zinc solubilization by endophytic fungal strains.
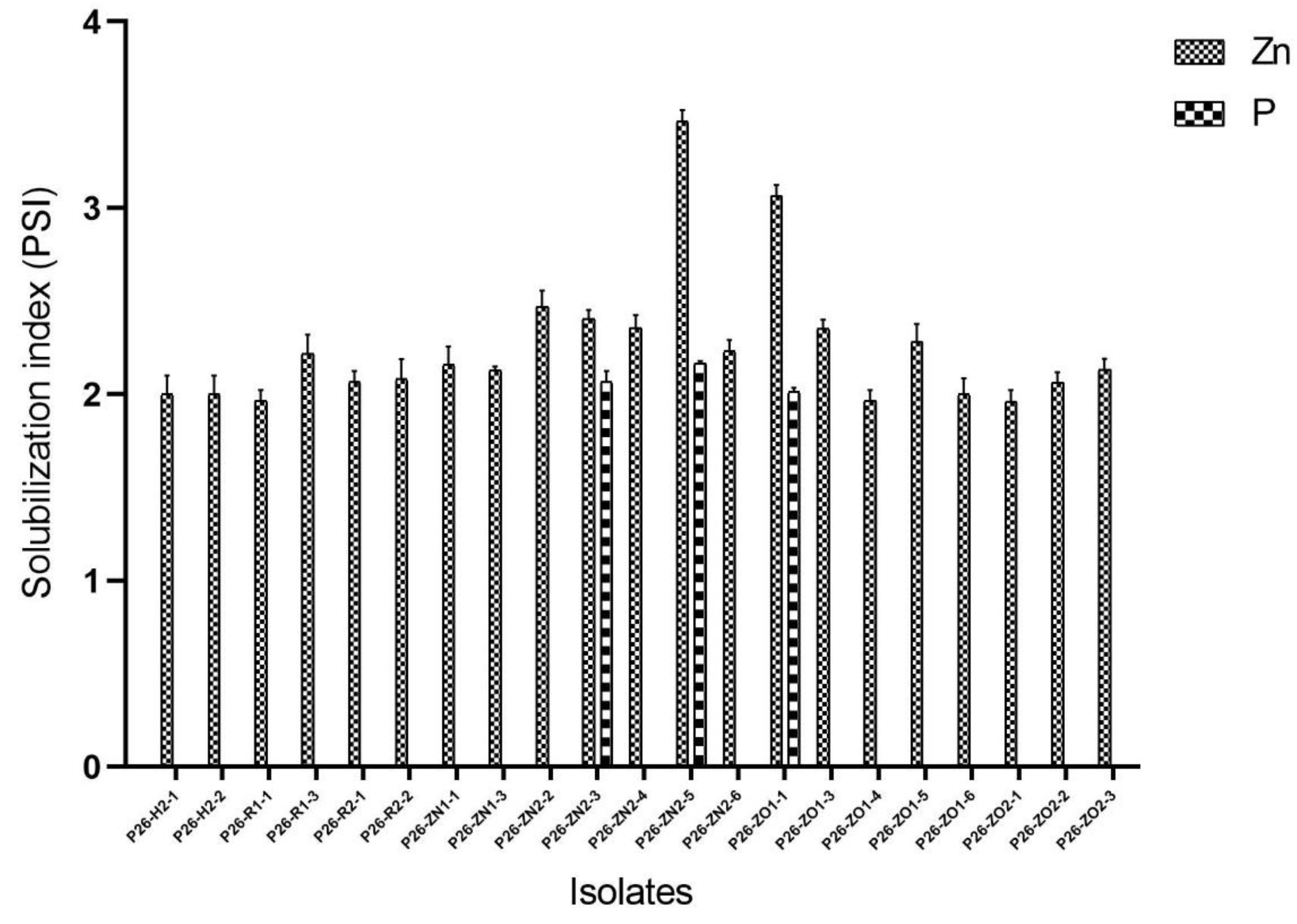

Table 1.
Closest Relatives of Endophytic Fungal Strains Based on ITS Sequence BLAST Analyses.
| Strain | Accession No | Plant part | Species with most homologous sequence (Accession No) | Similarity % |
|---|---|---|---|---|
| P26-H1-1 | LC769420 | Aboveground |
Fusarium equiseti CB33-4 (MT558601) |
99.81 |
| P26-H1-3 | LC769421 | Aboveground |
Clonostachys rosea MR44 (KY320599) |
99.65 |
| P26-H2-1 | LC769422 | Aboveground |
Fusarium solani GBC-Fungus 27 (MN077430) |
100 |
| P26-H2-2 | LC769423 | Aboveground |
Fusarium solani N-49-1 (MT560378) |
100 |
| P26-R1-1 | LC769424 | Underground |
Fusarium equiseti NL-374-D (OQ561206) |
99.63 |
| P26-R1-3 | LC769425 | Underground |
Alternaria sp. INM5 (KY781740) |
99.30 |
| P26-R2-1 | LC769426 | Underground |
Fusarium solani N-13-2 (MT560338) |
100 |
| P26-R2-2 | LC769427 | Underground |
Clonostachys rosea MR44 (KY320599) |
99.13 |
| P26-ZN1-1 | LC769428 | Fresh rhizome |
Penicillium chrysogenum MZC-0 (MN069559) |
99.66 |
| P26-ZN1-2 | LC769429 | Fresh rhizome |
Madurella fahalii 332- pus (OQ421454) |
98.96 |
| P26-ZN1-3 | LC769430 | Fresh rhizome |
Aspergillus tabacinus fung8 (MT635280) |
100 |
| P26-ZN2-2 | LC769431 | Fresh rhizome | Penicillium chrysogenum MZC-0 (MN069559) | 99.83 |
| P26-ZN2-3 | LC769432 | Fresh rhizome | Penicillium roseopurpureum IHEM:28005 (OU989457) | 99.83 |
| P26-ZN2-4 | LC769433 | Fresh rhizome |
Fusarium sp. GFR18 (MT447523) |
100 |
| P26-ZN2-5 | LC769434 | Fresh rhizome |
Penicillium vinaceum 533 (DQ681340) |
100 |
| P26-ZN2-6 | LC769435 | Fresh rhizome | Penicillium roseopurpureum G5-2 (MN206951) | 100 |
| P26-ZO1-1 | LC769436 | Old rhizome |
Penicillium sp. FP-027-A7 (MH102087) |
99.49 |
| P26-ZO1-3 | LC769437 | Old rhizome |
Clonostachys rosea daef27 (MH550497) |
99.12 |
| P26-ZO1-4 | LC663164 | Old rhizome |
Clonostachys rosea Potato root (MT448899) |
100 |
| P26-ZO1-5 | LC769438 | Old rhizome |
Clonostachys sp. 1R1D (OR365747) |
99.82 |
| P26-ZO1-6 | LC769439 | Old rhizome |
Fusarium sp. GFR18 (MT447523) |
99.82 |
| P26-ZO2-1 | LC769440 | Old rhizome |
Fusarium proliferatum CBB-6 (MT560216) |
100 |
| P26-ZO2-2 | LC769441 | Old rhizome |
Clonostachys rosea MR44 (KY320599) |
99.83 |
| P26-ZO2-3 | LC769442 | Old rhizome |
Clonostachys rosea N25 (MH259861) |
100 |
Table 2.
Antimicrobial Activity of Endophytic Fungi.
| Strain | Taxa | Escherichia coli | Bacillus subtilis | Staphylococcus aureus | Candida albicans | Aspergillus niger |
|---|---|---|---|---|---|---|
| P26-H1-1 | Fusarium equiseti | - | 15.5±0.7 | 12±0 | - | - |
| P26-H1-3 | Clonostachys rosea | - | 10.5±0.7 | - | - | 15.5±0.7 |
| P26-H2-1 | Fusarium proliferatum | - | 11.5±1.4 | 13±0.7 | - | - |
| P26-H2-2 | Fusarium solani | - | - | 10.5±0.7 | - | 23±2.8 |
| P26-R1-1 | Fusarium equiseti | - | 15.5±0.7 | 13.5±0.7 | - | - |
| P26-R2-1 | Fusarium solani | - | - | - | - | 22.5±0.7 |
| P26-R2-2 | Clonostachys rosea | - | - | 7.5±2.1 | - | - |
| P26-ZN1-2 | Madurella fahalii | 7.5±0.7 | 9.5±2.1 | 15.5±0.7 | - | - |
| P26-ZN1-3 | Aspergillus amoenus | - | 8±1.4 | 7±0 | - | - |
| P26-ZO1-3 | Clonostachys rosea | - | 15±0 | 18.5±0.7 | - | 13.5±2.1 |
| P26-ZO1-4 | Clonostachys rosea | - | 9±4.2 | 9.5±0.7 | - | 12.5±2.1 |
| P26-ZO1-5 | Clonostachys rosea | - | 13.5±0.7 | 16.5±0.7 | - | 14±1.4 |
| P26-ZO1-6 | Fusarium tonkinense | - | 11.5±0.7 | 17.5±0.7 | - | 12±0 |
| P26-ZO2-2 | Clonostachys rosea | - | 12±0 | 15.5±2.1 | - | - |
| P26-ZO2-3 | Clonostachys rosea | - | 13.5±0.7 | 16±0 | - | 13±1.4 |
Note: Diameter of the inhibitory zone: mm, -: no inhibitory activity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



