
Submitted:
28 December 2023
Posted:
04 January 2024
You are already at the latest version
Abstract
Mountain ecosystems are crucial for global biodiversity conservation. However, their landscape features are constantly changing owing to urban expansion. Understanding the relationships between biotic communities and landscape features is essential for biodiversity conservation. This study aimed to examine the effect of land-cover type on avian communities in Lishui, a mountainous urban area in eastern China. Avian surveys were conducted using a total of 168 line transects across different land-cover types once per season from December 2019 to January 2021. We assessed the diversity of bird communities by calculating various metrics at both taxonomic and functional levels. Among the land-cover types measured, woodland, built-up land, cultivated land, and water bodies significantly influenced bird community diversity and composition. Species richness, species abundance, and functional richness were negatively correlated with the proportion of woodland but positively correlated with the proportion of non-natural land cover, such as built-up and cultivated land. In contrast, functional evenness was positively correlated with the proportion of woodland and grassland but negatively correlated with the proportion of non-natural land cover. Land-cover type also exhibited significant correlations with avian functional characteristics such as diet, foraging strata, and body mass, thereby influencing the overall community structure. Our results indicated that mountainous landscape patterns substantially affect avian communities. Different land-cover types possess varying resource endowments that affect the distribution of avian species. Therefore, urban landscape planning in mountainous areas should carefully consider the various functions provided to organisms by different types of land cover to promote biodiversity.
Keywords:
Bird community
; Functional diversity
; Taxonomic diversity
; Landscape pattern
; Mountain city
1. Introduction
Mountain ecosystems are an important component of terrestrial ecosystems worldwide and are considered pivotal areas for biodiversity conservation [1]. Although mountains comprise only 25% of Earth’s land area, they are home to approximately 87% of the world’s wild animals, including amphibians, reptiles, birds, and mammals [2,3]. However, human population growth and economic development have accelerated deforestation to clear land for agricultural or residential needs [4]. These activities have intensified the modification of natural landscapes in mountainous regions, imposing considerable pressure on the species inhabiting these areas [1].
In recent years, extensive research has been conducted investigating the impact of natural landscape modifications on biodiversity. In particular, birds, which occupy a diverse range of habitats and are highly susceptible to environmental fluctuations [5], have become the most frequently studied species for examining the effects of anthropogenic habitat alterations on animals [6,7,8].
Anthropic landscape changes encompassing alterations in land-cover type, quantity, and composition often result in the loss or increase of avian species and changes in community species composition [9,10,11]. On the one hand, landscapes that encompass a substantial proportion of natural or semi-natural land cover tend to have positive effects on species richness and abundance. This may be attributed to the provision of ample natural shelter and high habitat connectivity in such landscapes [12,13,14]. However, anthropogenic landscape modification often reduces natural habitats, leading to fragmentation of the remaining native land cover into isolated patches within a matrix of non-natural land-cover types [10,14]. According to the habitat heterogeneity hypothesis, heterogeneous habitats generally contain more species because they provide more diverse ways of exploiting the available environmental resources (niches) [15]. However, access to resources may be impeded or entirely prevented when isolated native patches are encompassed by non-natural land cover that is unsuitable for survival [16]. On the other hand, anthropogenic landscapes can also provide benefits to certain species in some cases [17]. For instance, some habitat generalists, such as house sparrows and barn swallows, are found in diverse arrays of anthropogenic habitats [18], whereas other species are endemic to a particular type of natural habitat [19]. Therefore, understanding the potential responses of different species to various forms of anthropogenic land-cover change is crucial to effectively guide urban landscape planning in mountainous regions.
The ecological traits of a species influence its ability to thrive in specific environments [20]. Environmental factors act as selective forces that eliminate species that cannot tolerate the conditions at a particular site [21]. Hence, in addition to taxonomic modifications (such as variations in species richness, abundance, and composition), biomes also exhibit functional changes owing to alterations in landscape patterns [5,22]. However, numerous studies have demonstrated that the pattern of changes in functional diversity may differ from that of taxonomic diversity [6,23]. For instance, the study conducted by Coetzee and Chown (2016) found that land-use change resulted in an increase in the diversity of avian species in the local area. However, land-use change also caused a decrease in the abundance of species that possess unique functional traits. The simultaneous consideration of taxonomic and functional diversity has the potential to enhance our understanding of the impact of anthropogenic habitat alteration on biological communities.
China’s terrain is predominantly mountainous, with approximately two-thirds of its land area covered by mountains. Furthermore, approximately one-third of cities in China are located in mountainous areas [24]. Expansion of urban areas is an unavoidable consequence of the exponential development of China’s economy. In this study, we examined the impact of landscape patterns on bird communities in a mountainous city in eastern China. Specifically, we conducted a comprehensive analysis of bird community diversity by calculating multiple metrics at both the taxonomic and functional levels, and assessed the impact of landscape patterns on these metrics. Furthermore, we conducted an analysis to investigate the relationship between species composition and functional traits of the assemblage and landscape patterns. We hypothesized that: (1) the diversity of bird species, taxonomically and functionally, would increase to some extent owing to the emergence of non-natural land cover and expansion of bioavailable niches; and (2) different land-cover types may have varying associations with the functional characteristics of birds, thereby influencing the composition of bird communities
2. Materials and Methods
2.1. Study region
Lishui city, located in the Zhejiang province of eastern China (27°25′–28°57′ N; 118°41′–120°26′ E), is a typical mountainous urban area. It spans a total area of 17275 km2 and is home to a population of 2.70 million residents (Figure 1). Mountains account for more than 90% of the total land area. Within this region, more than 3500 peaks exceed an elevation of 1000 m, with 240 peaks surpassing 1500 m. The region has a subtropical monsoon climate and is characterized by a wide range of vegetation types and substantial vegetation coverage, with forest cover exceeding 80%. Intricate and diverse terrains and landforms, along with a wide array of vegetation types, have resulted in the formation of numerous high-quality habitats that support a diverse range of invertebrate and vertebrate species [25].
2.2. Bird survey
In total, 168 line transects, each measuring 1.5 km in length, were evenly distributed throughout the study region (Figure 1). Each transect was surveyed once per season from December 2019 to November 2021. To mitigate the potential impact of light and temperature, surveys were conducted within the specific timeframe of three hours after sunrise and three hours before sunset. Furthermore, the surveys were conducted only when weather conditions were optimal, specifically, when there was no rain or wind. During the surveys, the observers conducted transect walks at a consistent speed of approximately 2.0 km/h. They utilized binoculars to document all visible bird species and listened for bird vocalizations to record the presence of non-visible species in the area. To prevent the duplication of records, birds that flew over the transect were not recorded. Additionally, as our objective was to investigate the impact of land cover on bird communities, rather than to measure the exact bird population densities, direct bird count data were used in the subsequent analyses without adjusting for detectability. The bird taxonomy and nomenclature used in this study adhered to the guidelines outlined in A Checklist on the Classification and Distribution of the Birds of China [26].
2.3. Land-cover types and diversity
We quantified the composition of land-cover types around each sampling transect to determine the degree of human activity. First, Landsat 8 satellite images with a spatial resolution of 30 × 30 m were obtained from the geospatial data cloud (http://www.gscloud.cn/). We used a series of image preprocessing techniques, including image fusion, image mosaicing, radiometric calibration, and atmospheric correction, to mitigate the influence of different error sources. Second, supervised classification was performed on the Landsat 8 satellite images using the maximum likelihood classifier in ENVI 5.3. Subsequently, the percentage of various land-cover types within a buffer area (750 m radius) surrounding each sampling transect was computed using Geographic Information System processing tools. Ultimately, seven land-cover types (woodland, cultivated land, shrubland, bare land, grassland, water bodies, and built-up land) were identified and further verified during field surveys. Additionally, we calculated the Shannon–Wiener diversity index of land-cover types to characterize the landscape diversity within each sampling transect.
2.4. Functional traits
To assess functional diversity, we selected 12 avian functional traits associated with resource utilization (Table 1). These traits included body mass (a continuous trait), diet (five categorical traits), and foraging strata (six continuous traits). Functional traits were derived from a global dataset provided by Wilman et al.(2014) [27], in which each species is assigned to a specific dietary guild, and the foraging strata for each species are represented as percentages in multiple columns that sum up to 100.
2.5. Taxonomic and functional diversity
Taxonomic diversity was assessed by calculating species richness, species abundance, and the Shannon–Wiener diversity index for each line transect. Functional diversity was assessed by examining functional richness (FRic), functional evenness (FEve), and functional divergence (FDiv), each of which reveals distinct facets of functional diversity [28]. The FRic indicates the volume occupied by a species assemblage within a functional space, FEve reflects the regularity of the abundance distribution in the functional space, and FDiv is the degree to which the abundance of a community is distributed towards the extremities of the occupied trait space.
2.6. Statistical analyses
To assess the effects of land-cover type on the taxonomic and functional diversity of the avian community, we applied generalized linear models with Poisson distributions, and over-dispersion was adjusted using a quasi-Poisson procedure. We determined that the seven land-cover types (percentage of area) were correlated and exhibited high collinearity; thus, we first performed principal component analysis to summarize these land-cover types into three independent principal components (PC1, PC2, and PC3) that could explain most of the original variance. In the models, PC1, PC2, PC3, LD, and season were used as explanatory variables. The associations between the three extracted components and the original explanatory continuous variables are presented in Table 2. In addition, we used the Wilcoxon rank-sum test with Bonferroni correction to further examine the seasonal variability of avian diversity indices.
To assess the influence of land-cover type on avian community composition, land-cover data were integrated into the non-metric multidimensional scaling (NMDS) ordination space of avian community composition using the ‘envfit’ function in the ‘vegan’ R package. To improve the clarity of our findings, we visually represented the distribution of the avian dietary guilds in the ordination space. In this study, NMDS was calculated using a Bray-Curtis distance matrix based on species abundance data. Pearson’s correlation analysis was used to evaluate the correlations between NMDS axis scores and each land-cover type, and a permutation test (999 repetitions) was used to assess the significance of the correlation coefficients.
To further investigate the role of land-cover type in shaping bird communities, we related the functional traits of bird species to land-cover type using fourth-corner statistics [29]. By utilizing three datasets (i.e., species abundance, functional traits, and land-cover types), this methodology enabled the detection of positive or negative correlations between avian traits and the environmental characteristics of their respective habitats. Additionally, it provided a measure of the statistical significance of these associations.
All statistical analyses were performed using R version 4.0.1 [30], and P < 0.05 was considered significant.
3. Results
3.1. Principal component analysis results
The first three principal components (PC1, PC2, and PC3) explained 69% of the land-cover types (percentage of area) variation in the 168 samples (Table 2). In particular, PC1 (34.3%) was positively associated with woodland and negatively associated with built-up and cultivated land, PC2 (19.2%) was positively associated with both shrubland and water bodies, and PC3 (15.3%) was positively associated with grassland and negatively associated with shrubland.
3.2. Bird survey results
During the survey period, 45 027 birds from 316 species belonging to 18 orders and 67 families were recorded along the 168 line transects. Among the recorded species, 59 were listed as Class II Key Protected Wild Animal Species in China and 42 were listed as either near threatened, vulnerable, or endangered on the Red List of International Union for Conservation of Nature.
3.3. Influence of land-cover types on avian community diversity
The generalized linear models showed that land-cover types had significant impacts on the diversity of bird communities (Table 3). Specifically, PC1 (positively correlated with woodland but negatively correlated with built-up and cultivated land) was negatively correlated with species richness, species abundance, the Shannon–Wiener diversity index, and FRic and positively correlated with FEve. Additionally, PC3 (positively correlated with grassland and negatively correlated with shrubland) was positively correlated with FEve.
3.4. Influence of season on avian community diversity
The results from the generalized linear models also indicated that season was an important factor affecting the diversity indices, both at the taxonomic and functional levels (Table 3). The results of the Wilcoxon rank-sum test showed (Figure 2) that the species richness was significantly higher in autumn (16.63 ± 11.49), spring (16.46 ± 9.44), and summer (14.70 ± 8.12) than in winter (11.27 ± 6.74). Additionally, the species abundance was significantly higher in autumn (90.24 ± 104.57) and spring (64.40 ± 56.65) than in winter (57.40 ± 92.73) and summer (55.97 ± 53.69). The Shannon–Wiener diversity index was the highest in spring (2.35 ± 0.53) and the lowest in winter (1.95 ± 0.45). It was also significantly higher in summer (2.28 ± 0.48) and autumn (2.24 ± 0.52) than in winter.
In terms of functional diversity, the FRic was the highest in summer (10.34 ± 3.92) and spring (9.65 ± 3.77), followed by autumn (8.98 ± 3.51) and winter (7.18 ± 3.69). The FEve was the highest in summer (0.66 ± 0.09), followed by spring (0.63 ± 0.09) and autumn (0.62 ± 0.09), and it was the lowest in winter (0.61 ± 0.11). The FDiv in autumn (0.80 ± 0.09) and winter (0.79 ± 0.09) was significantly higher than that in spring (0.76 ± 0.09) and summer (0.74 ± 0.09), and the latter two values differed significantly from each other.
3.5. Influence of land-cover types on avian community composition
The bird community composition was significantly influenced by five of the eight environmental variables (seven land-cover types and landscape diversity). Among the land-cover types, woodland, built-up land, cultivated land, and water bodies showed the strongest correlations with variations in species composition across all seasons (Figure 3). Of these, woodland provided the highest explanatory rate (spring: 0.20; summer: 0.20; autumn: 0.24; winter: 0.12), followed by built-up land (spring: 0.15; summer: 0.16; autumn: 0.17; winter: 0.08) and cultivated land (spring: 0.11; summer: 0.11; autumn: 0.13; winter: 0.05). Additionally, the winter bird community was affected by bare land. In the ordination diagrams (Figure 3), we observed that the distribution of avian diet guilds was influenced by land-cover type. For example, the PlantSeed guild (which feeds on plants and seeds) appeared more frequently in habitats with large areas of non-natural land cover, such as built-up and cultivated land, particularly in spring and autumn. During winter, the VertFishScav guild (which feeds on vertebrates, fish, and carrion) appeared more frequently in habitats with expansive visibility and a high proportion of water bodies. Insectivorous (which feed on invertebrates) and omnivorous birds were more evenly distributed in the ordination diagrams.
3.6. Associations between land-cover types and bird functional characteristics
The fourth-corner analysis revealed numerous stable associations between land-cover type and bird functional characteristics, particularly in the foraging strata (Figure 4). In most seasons, foraging in water was positively correlated with the proportion of water bodies and built-up land, whereas it was negatively correlated with the proportion of woodland. The opposite was true when foraging in the vegetation layer. We found that the functional characteristics of foraging on the vegetation layer were negatively correlated with the proportion of water bodies and non-natural land covers (i.e., built-up and cultivated land) but positively correlated with the proportion of woodland. Additionally, a positive relationship was observed between foraging on the ground and the proportion of cultivated land during spring and winter. In terms of diet, the relationships between most dietary characteristics and land-cover type exhibited large seasonal variations, except for the characteristic of VertFishScav guild, which was significantly and positively associated with the proportion of water bodies. Body mass had a strong positive correlation with the proportion of built-up land and water bodies but showed a negative correlation with the proportion of woodland. This suggests that large-bodied birds occur more frequently in open habitats, whereas small-bodied birds are more common in dense habitats.
4. Discussion
The preservation of biodiversity has emerged as a paramount concern in the context of the rapid decline of species worldwide. Montane environments are important biodiversity hotspots that have been subjected to habitat changes due to human activity. This study examined the impact of land-cover types on avian communities in the mountainous region of Lishui, a typical urban area in eastern China. Our results revealed that the diversity and composition of avian communities in mountainous urban areas exhibit seasonal variations and are influenced by land-cover type. The results of this study provide a valuable resource for the future planning of urban areas in mountainous regions and the conservation of biodiversity.
Land-cover distribution is closely associated with the survival of birds, making it a key factor influencing bird diversity [6,23]. Moreover, diverse land-cover types offer varying resources and habitats for birds, potentially resulting in distinct impacts on avian diversity [8,31]. Forests or woodlands are typically the primary habitats for birds when selecting nesting and roosting locations. This study identified woodland, cultivated land, and built-up land as the most dominant land-cover types affecting avian diversity. The proportion of woodland had a negative effect on species richness and abundance, while that of non-natural land covers, such as built-up and cultivated land, had a positive effect. Although this finding is contrary to those of previous studies [8,31,32], these studies were primarily conducted in landscapes dominated by human activities, such as urban areas and farmland, where non-natural land cover predominated and woodland habitats were relatively scarce. In contrast, our research was conducted in a mountainous landscape characterized by a forest cover of over 80%, providing ample and abundant habitats for birds. Under these circumstances, as the proportion of woodland decreases and that of non-natural landscapes increases, the variety of available habitats for birds also increases. In particular, generalist birds capable of inhabiting a variety of habitats with diverse resources can clearly derive benefits from the supplementary resources offered by non-natural land cover [9]. Similar results have been obtained in earlier studies conducted in mountainous regions [7,31]. This finding also reveals that the expansion of non-natural land cover could be strategically increased while preserving abundant natural forest or woodland remnants during mountain landscape development. Furthermore, we found that functional richness was negatively correlated with the proportion of woodland and positively correlated with non-natural land cover. This phenomenon can also be explained by the habitat diversity hypothesis, which states that niche diversity increases with habitat type [15]. Mosaic forest landscapes, characterized by anthropogenic land cover interspersed with woodland areas, offer a greater number of ecological niches than more homogenous landscapes. Consequently, they support higher functional richness than areas that are entirely forested.
In this study, bird community diversity also exhibited seasonal variation, with higher richness and abundance in spring and autumn and lower richness and abundance in winter (Figure 2). The availability of food resources, including variety, distribution, and quantity, is considered one of the most influential factors [14,33]. In contrast to the scarcity of food resources during the colder winter months, there is a notable increase in both the variety and quantity of food available during spring and autumn. For example, flowering plants can offer ample pollen and nectar resources, whereas cultivated land provides a substantial quantity of invertebrates and seed grains following spring plowing. Autumn is the period during which various types of grains and fruits mature. In addition, the reasons for the low number of birds in winter may include bird migration, hibernation, and decreased activity aimed at minimizing heat expenditure. Species richness and functional richness cannot be described by simple linear relationships [34]. Theoretically, an increase in species richness typically leads to an increase in functional richness. However, functional redundancy may occur when additional species fill the same ecological role [35,36]. In this study, the number of species observed in autumn exceeded that in spring and summer, whereas functional diversity exhibited a notable decrease (Figure 2). Functional evenness describes the regularity of the distribution of functional traits within a functional space and is related to resource utilization [37]. The greater the value, the more comprehensive the utilization of resources. We found that the bird community exhibited the highest utilization of environmental resources during summer, followed by that in spring and autumn, with the lowest utilization during winter (Figure 2). Functional divergence describes the differences in functional characteristics within a community and is related to niche differentiation and resource competition [37]. We found that resource competition within the bird community was less pronounced during autumn and winter but more intense during spring and summer (Figure 2).
The species distribution in avian communities is influenced by a combination of environmental conditions and species traits [6,8,17]. Birds with narrow dietary niches are restricted to a limited number of locations where their specific niche requirements are met. For example, as demonstrated in this study, avian species that feed on fish tended to inhabit areas in close proximity to water bodies, whereas those that feed on plants and seeds were more commonly observed in habitats characterized by a high proportion of built-up and cultivated land, typically in the vicinity of home gardens and farmland (Figure 3) Moreover, environmental conditions within a given habitat fluctuate with the changing seasons, and the reliance of birds on specific habitats fluctuates throughout the year (Figure 3 and Figure 4). Birds with wide niches thrive in various habitats [13]. For example, the omnivorous birds in our study were evenly distributed in the ordination diagrams (Figure 3). In addition, consistent with previous findings [38,39], our study revealed a close association between land-cover type and the foraging strata of avian species. For instance, the functional characteristics of foraging in water were positively correlated with the proportion of water bodies and built-up land (typically accompanied by water sources such as ponds or irrigation canals), whereas they were negatively correlated with the proportion of woodland (Figure 4). Foraging on the vegetation layer exhibited an inverse relationship, showing a positive correlation with woodland but a negative correlation with water bodies and non-natural land cover (i.e., built-up and cultivated land) (Figure 4). These findings have important implications for the construction of cities in mountainous regions. In the urban planning and construction process, it is imperative for planners to prioritize landscape configurations, particularly the type of land cover, to create a conducive environment for diverse bird species that occupy different ecological niches. Moreover, our findings indicate a significant positive correlation between body mass and the proportion of built-up land and water bodies and a negative correlation with the proportion of woodland (Figure 4). This suggests that larger-bodied birds are more likely to occur in open habitats, whereas smaller-bodied birds are more prevalent in dense habitats. Smaller body sizes result in more rapid heat consumption [40]. To offset the energy expenditure, smaller birds typically need to forage more frequently, which may attract greater attention from predators and humans. Consequently, they inhabit enclosed environments with dense vegetation to minimize exposure risks.
5. Conclusions
Our results indicated that mountainous landscape patterns have considerable effects on avian diversity and community structure. Non-natural land cover can increase the taxonomic diversity and functional richness to a certain extent. However, this increase was observed when an adequate natural environment was maintained. Hence, it is imperative that we continue to protect the natural environment and enact measures to prevent habitat destruction. Furthermore, we found that various land-cover types possess distinct resource endowments that influence bird distribution. Therefore, we recommend that in the planning of urban mountain landscapes, attention should be given not only to the extent of natural landscape development but also to the functions provided to organisms by different land-cover types. Maintaining diverse landscapes enables different species to occupy different niches and survive in mountainous urban areas, thereby preserving biodiversity.
Author Contributions
Conceptualization, C.L.; methodology, W.Z. and S.Z.; software, W.Z. and S.Z.; formal analysis; W.Z. and S.Z.; investigation, Y.X., J.T., D.C., Y.Y., and J.S.; writing—original draft preparation; X.W.; writing—review and editing, P.C. All authors read and approved the final manuscript.
Funding
This research was funded by the Open Research Fund of the State Environmental Protection Key Laboratory on Biodiversity and Biosafety.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to data protection restrictions.
Acknowledgments
We would like to thank the Lishui Municipal Bureau of Ecological Environment for their support of this study and the people who participated in the bird field survey.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Z.; Li, X.; Song, W.; Li, Q.; Onditi, K.; Khanal, L.; Jiang, X. Small mammal species richness and turnover along elevational gradient in Yulong Mountain, Yunnan, Southwest China. Ecol. Evol. 2020, 10, 2545–2558. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, A.; Meng, F.; Zhao, W.; Yang, Y.; Soininen, J.; Shen, J.; Zhou, J. Embracing mountain microbiome and ecosystem functions under global change. New Phytol. 2022, 234, 1987–2002. [Google Scholar] [CrossRef]
- Rahbek, C.; Borregaard, M.K.; Colwell, R.K.; Dalsgaard, B.; Holt, B.G.; Morueta-Holme, N.; Nogues-Bravo, D.; Whittaker, R.J.; Fjeldså, J. Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science 2019, 365, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ziegler, A.D.; Elsen, P.R.; Feng, Y.; Baker, J.C.A.; Liang, S.; Holden, J.; Spracklen, D.V.; Zeng, Z. Accelerating global mountain forest loss threatens biodiversity hotspots. One Earth 2023, 6, 303–315. [Google Scholar] [CrossRef]
- Matuoka, M.A.; Benchimol, M.; de Almeida-Rocha, J.M.; Morante-Filho, J.C. Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis. Ecol. Indic. 2020, 116, 106471. [Google Scholar] [CrossRef]
- Coetzee, B.W.T.; Chown, S.L. Land-use change promotes avian diversity at the expense of species with unique traits. Ecol. Evol. 2016, 6, 7610–7622. [Google Scholar] [CrossRef] [PubMed]
- Desrochers, R.E.; Kerr, J.T.; Currie, D.J. How, and how much, natural cover loss increases species richness. Global Ecol. Biogeogr. 2011, 20, 857–867. [Google Scholar] [CrossRef]
- Hayes, W.M.; Fisher, J.C.; Pierre, M.A.; Bicknell, J.E.; Davies, Z.G. Bird communities across varying landcover types in a Neotropical city. Biotropica. 2020, 52, 151–164. [Google Scholar] [CrossRef]
- Shoffner, A.; Wilson, A.M.; Tang, W.; Gagné, S.A. The relative effects of forest amount, forest configuration, and urban matrix quality on forest breeding birds. Sci. Rep. 2018, 8, 17140. [Google Scholar] [CrossRef]
- Maseko, M.S.T.; Zungu, M.M.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Downs, C.T. Effects of habitat-patch size and patch isolation on the diversity of forest birds in the urban-forest mosaic of Durban, South Africa. Urban Ecosyst. 2020, 23, 533–542. [Google Scholar] [CrossRef]
- Soares, F.C.; Panisi, M.; Sampaio, H.; Soares, E.; Santana, A.; Buchanan, G.M.; Leal, A.I.; Palmeirim, J.M.; Lima, R.F. Land-use intensification promotes non-native species in a tropical island bird assemblage. Anim. Conserv. 2020, 23, 573–584. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Basnet, T.B.; Rokaya, M.B.; Bhattarai, B.P.; Münzbergová, Z. Heterogeneous Landscapes on Steep Slopes at Low Altitudes as Hotspots of Bird Diversity in a Hilly Region of Nepal in the Central Himalayas. PLoS One 2016, 11, e0150498. [Google Scholar] [CrossRef]
- Yabuhara, Y.; Yamaura, Y.; Akasaka, T.; Yamanaka, S.; Nakamura, F. Seasonal variation in patch and landscape effects on forest bird communities in a lowland fragmented landscape. Forest Ecol. Manag. 2019, 454, 117140. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Miller-Rushing, A.J.; Primack, R.B.; Devictor, V.; Corlett, R.T.; Cumming, G.S.; Loyola, R.; Maas, B.; Pejchar, L. How does habitat fragmentation affect biodiversity? A controversial question at the core of conservation biology. Biol. Conserv. 2019, 232, 271–273. [Google Scholar] [CrossRef]
- Newbold, T.; Scharlemann, J.P.W.; Butchart, S.H.M.; Sekercioğlu, C.H.; Alkemade, R.; Booth, H.; Purves, D.W. Ecological traits affect the response of tropical forest bird species to land-use intensity. P. Roy. Soc. B-Biol. Sci. 2013, 280, 20122131. [Google Scholar] [CrossRef]
- Parra-Torres, Y.; Ramírez, F.; Afán, I.; Aguzzi, J.; Bouten, W.; Forero, M.G.; Navarro, J. Behavioral rhythms of an opportunistic predator living in anthropogenic landscapes. Mov. Ecol. 2020, 8, 17. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, G.; Ma, H.; Wu, Y.; Zhang, W.; Zhang, Y.; Li, C.; de Boer, W.F. Bird communities’ responses to human-modified landscapes in the southern Anhui Mountainous Area. Avian Res. 2022, 13, 100006. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.; Zha, D.; Yang, S.; Huang, Z.Y.X.; Boer, W.F. Assembly processes of waterbird communities across subsidence wetlands in China: A functional and phylogenetic approach. Divers. Distrib. 2019, 25, 1118–1129. [Google Scholar] [CrossRef]
- Schaaf, A.A.; Gomez, D.; Tallei, E.; Vivanco, C.G.; Ruggera, R.A. Responses of functional traits in cavity-nesting birds to logging in subtropical and temperate forests of the Americas. Sci. Rep. 2021, 11, 24309. [Google Scholar] [CrossRef] [PubMed]
- Weideman, E.A.; Slingsby, J.A.; Thomson, R.L.; Coetzee, B.T.W. Land cover change homogenizes functional and phylogenetic diversity within and among African savanna bird assemblages. Landscape Ecol. 2020, 35, 145–157. [Google Scholar] [CrossRef]
- Li, W.; Li, X.; Tan, M.; Wang, Y. Influences of population pressure change on vegetation greenness in China’s mountainous areas. Ecol. Evol. 2017, 7, 9041–9053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, Y.; Fang, X.; Zhao, S.; Wu, Y.; Zhang, H.; Cui, L.; Cui, P. Effects of Environmental Factors on Bird Communities in Different Urbanization Grades: An Empirical Study in Lishui, a Mountainous Area of Eastern China. Animals (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of China, 3rd ed.; Science Press: Beijing, 2017. [Google Scholar]
- Wilman, H.; Belmaker, J.; Simpson, J.; de La Rosa, C.; Rivadeneira, M.M.; Jetz, W. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef]
- R Core Team. R: a Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, 2020. [Google Scholar]
- Sasaki, K.; Hotes, S.; Kadoya, T.; Yoshioka, A.; Wolters, V. Landscape associations of farmland bird diversity in Germany and Japan. Glob. Ecol. Conserv. 2020, 21, e00891. [Google Scholar] [CrossRef]
- Yang, X.; Tan, X.; Chen, C.; Wang, Y. The influence of urban park characteristics on bird diversity in Nanjing, China. Avian Res. 2020, 11. [Google Scholar] [CrossRef]
- Katuwal, H.B.; Pradhan, N.M.B.; Thakuri, J.J.; Bhusal, K.P.; Aryal, P.C.; Thapa, I. Effect of Urbanization and Seasonality in Bird Communities of Kathmandu Valley. Nepal. Proc. Zool. Soc. 2018, 71, 103–113. [Google Scholar] [CrossRef]
- Neves, K.; Moura, M.R.; Maravalhas, J.; Pacheco, R.; Pie, M.R.; Schultz, T.R.; Vasconcelos, H.L. Functional richness shows spatial scale dependency in Pheidole ant assemblages from Neotropical savannas. Ecol. Evol. 2019, 9, 11734–11741. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, J.T.; Martin, K. Biotic homogenization: Loss of avian functional richness and habitat specialists in disturbed Andean temperate forests. Biol. Conserv. 2015, 192, 418–427. [Google Scholar] [CrossRef]
- Altamirano, T.A.; de Zwaan, D.R.; Ibarra, J.T.; Wilson, S.; Martin, K. Treeline ecotones shape the distribution of avian species richness and functional diversity in south temperate mountains. Sci. Rep. 2020, 10, 18428. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Jacoboski, L.I.; Hartz, S.M. Using functional diversity and taxonomic diversity to assess effects of afforestation of grassland on bird communities. Perspect. Ecol. Conser. 2020, 18, 103–108. [Google Scholar] [CrossRef]
- Ding, Z.; Liang, J.; Hu, Y.; Zhou, Z.; Sun, H.; Liu, L.; Liu, H.; Hu, H.; Si, X. Different responses of avian feeding guilds to spatial and environmental factors across an elevation gradient in the central Himalaya. Ecol. Evol. 2019, 9, 4116–4128. [Google Scholar] [CrossRef]
- Aschoff, J. Thermal conductance in mammals and birds: Its dependence on body size and crcadian phase. Comparative Biochemistry and Physiology Part A: Physiology 1981, 69, 611–619. [Google Scholar] [CrossRef]
Figure 1.
Location of study area and distribution of avian sampling line transects.
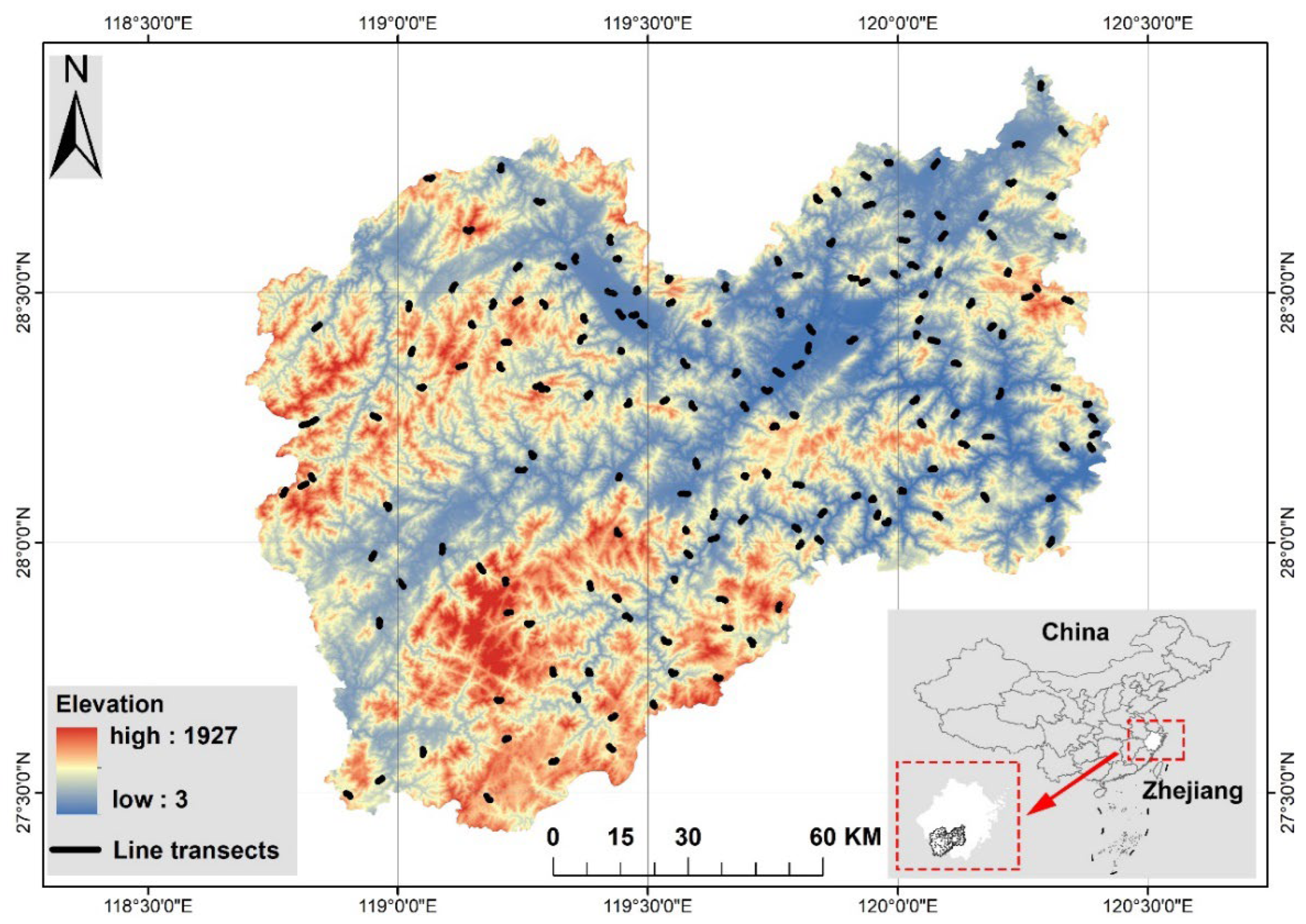
Figure 2.
Metrics of the avian communities in different seasons. Same letters indicate no significant differences between seasons and significance was determined by Wilcoxon rank-sum test. Significance is P ≤ 0.05.
Figure 2.
Metrics of the avian communities in different seasons. Same letters indicate no significant differences between seasons and significance was determined by Wilcoxon rank-sum test. Significance is P ≤ 0.05.
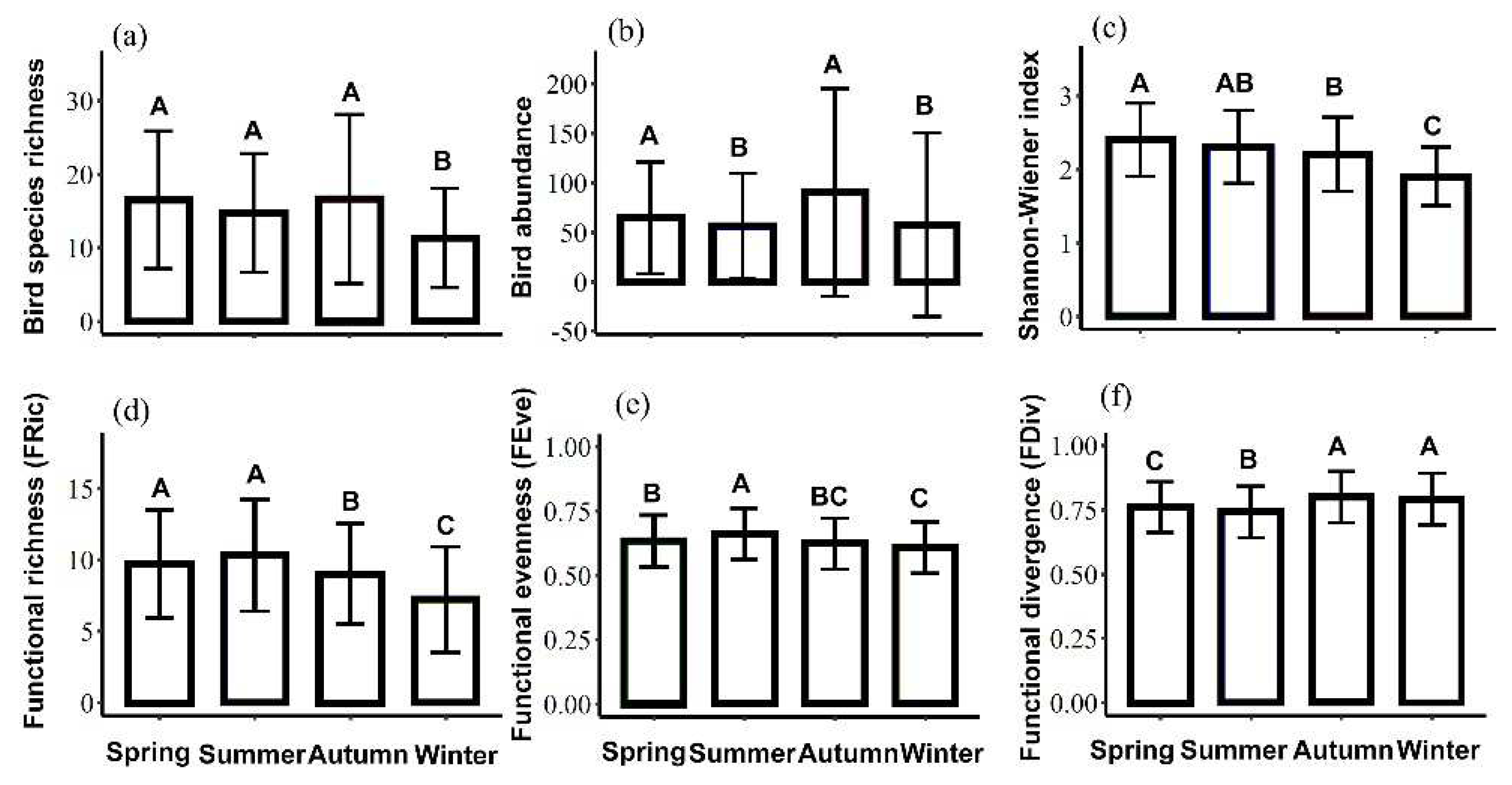
Figure 3.
Non-metric multidimensional scaling (NMDS) showing the relationships between avian communities and land-cover types in the different seasons. Each symbol represents a species and the shape of the symbol represents the feeding guild. Land-cover types with significant effects are represented by arrows with labels. WOL = woodland; LD = landscape diversity; BUA = Built-up areas; CUL = Cultivated land; WAB = Water bodies; BAL = Bare land.
Figure 3.
Non-metric multidimensional scaling (NMDS) showing the relationships between avian communities and land-cover types in the different seasons. Each symbol represents a species and the shape of the symbol represents the feeding guild. Land-cover types with significant effects are represented by arrows with labels. WOL = woodland; LD = landscape diversity; BUA = Built-up areas; CUL = Cultivated land; WAB = Water bodies; BAL = Bare land.
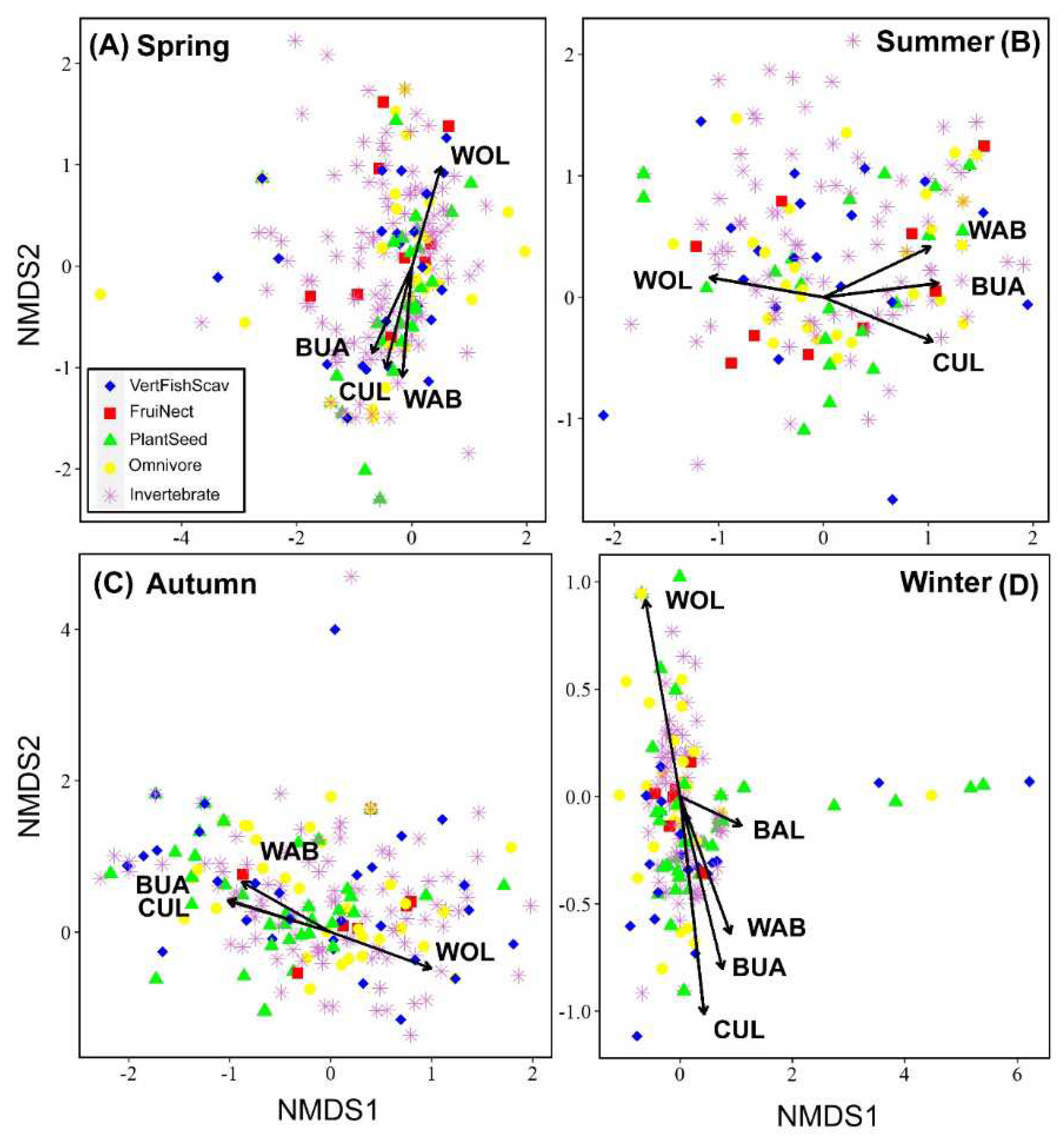
Figure 4.
Interaction coefficient from fourth corner analysis testing the relationship between bird functional characteristics and land cover in the different seasons:(A) in spring; (B) in summer; (C) in autumn; (D) in winter. Brighter squares show stronger associations than paler ones, positive associations are red, and negative associations are blue. F = foraging characteristic; D = diet characteristic.
Figure 4.
Interaction coefficient from fourth corner analysis testing the relationship between bird functional characteristics and land cover in the different seasons:(A) in spring; (B) in summer; (C) in autumn; (D) in winter. Brighter squares show stronger associations than paler ones, positive associations are red, and negative associations are blue. F = foraging characteristic; D = diet characteristic.
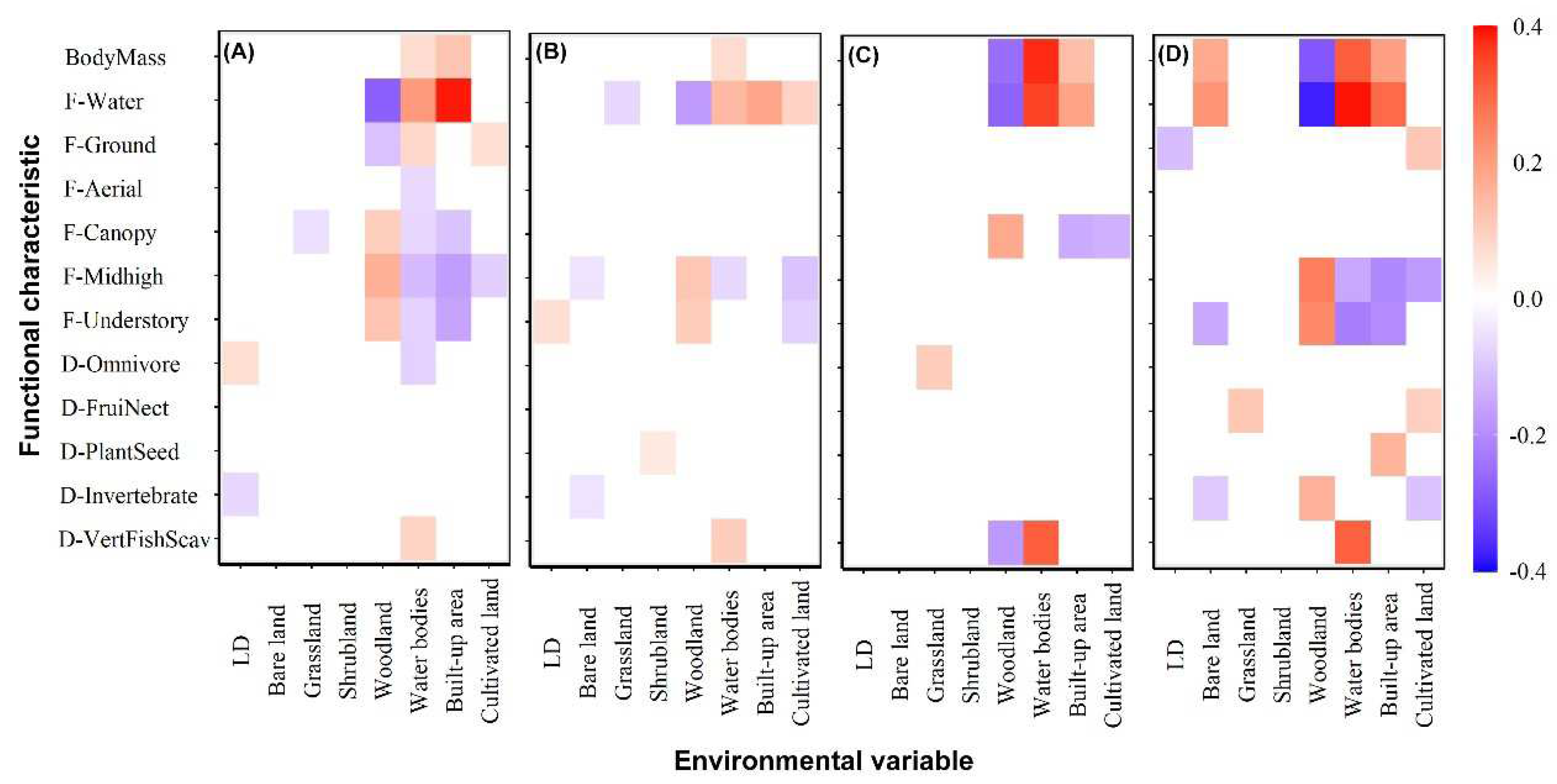

Table 1.
Traits used for the estimation of functional diversity.
| Trait type | Traits | Categories |
|---|---|---|
| Resource quantity | Body mass | Continuous |
| Diet guild | PlantSeed (feeding on plant and seeds); FruiNect (feeding on fruits and nectar); Invertebrate (feeding on invertebrates); VertFishScav (feeding on vertebrates, fish and carrion); Omnivore |
categorical |
| Foraging stratum | Ground; Understory; Midstorey; Canopy; Air; Water |
Continuous |
Table 2.
Results of the principal components analysis of land-cover types (percentage of area), PC = principal components.
Table 2.
Results of the principal components analysis of land-cover types (percentage of area), PC = principal components.
| Land-cover types | PC1 | PC2 | PC3 |
|---|---|---|---|
| Woodland | 0.638 | 0.059 | 0.007 |
| Shrubland | 0.028 | 0.298 | -0.300 |
| Built-up area | -0.486 | 0.051 | -0.259 |
| Grassland | -0.127 | -0.099 | 0.848 |
| Water bodies | -0.281 | 0.573 | -0.069 |
| Cultivated land | -0.491 | -0.437 | -0.032 |
| Bare land | -0.143 | 0.614 | 0.345 |
| Proportion of Variance (%) | 0.343 | 0.192 | 0.153 |
| Cumulative Proportion (%) | 0.343 | 0.534 | 0.687 |
Table 3.
The effect of land-cover types on bird diversity indices. Generalized linear model estimates of slopes of functions and their standard errors (in brackets) are presented. ***means P < 0.001; ** means P < 0.01, and *means P < 0.05. See Table 1 for an explanation of the principal components used in these analyses.
Table 3.
The effect of land-cover types on bird diversity indices. Generalized linear model estimates of slopes of functions and their standard errors (in brackets) are presented. ***means P < 0.001; ** means P < 0.01, and *means P < 0.05. See Table 1 for an explanation of the principal components used in these analyses.
| Explanatory variables | Models for | |||||
|---|---|---|---|---|---|---|
| Richness | Abundance | Shannon | FRic | FEve | FDiv | |
| PC1 | -0.08 (0.08) *** |
-0.16 (0.02) *** |
-0.05 (0.01) *** |
-0.05 (0.10) *** |
0.01 (0.00) * |
0.00 (0.00) |
| PC2 | 0.03 (0.02) |
0.05 (0.04) |
0.01 (0.01) |
0.02 (0.01) |
0.00 (0.01) |
0.00 (0.01) |
| PC3 | 0.04 (0.02) |
0.03 (0.03) |
0.01 (0.02) |
0.01 (0.01) |
-0.02 (0.01) * |
0.00 (0.02) |
| LD | 0.01 (0.01) |
0.05 (0.05) |
-0.00 (0.01) |
0.01 (0.01) |
-0.00 (0.00) |
-0.00 (0.01) |
| Spring | -0.01 (0.06) |
-0.34 (0.11) ** |
0.05 (0.02) * |
0.07 (0.04) |
0.02 (0.02) |
-0.05 (0.01) *** |
| Summer | -0.12 (0.06) * |
-0.48 (0.11) *** |
0.02 (0.02) |
0.14 (0.04) ** |
0.06 (0.02) *** |
-0.09 (0.01) *** |
| Winter | -0.38 (0.07) *** |
-0.45 (0.11) *** |
-0.14 (0.02) *** |
-0.22 (0.05) *** |
-0.03 (0.02) |
-0.02 (0.01) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



