
Submitted:
02 January 2024
Posted:
05 January 2024
You are already at the latest version
Abstract
Anti-inflammatory agents are widely used for the treatment of inflammatory diseases. Nevertheless, the associated side effects of available drugs make necessary the search for new anti-inflammatory conditions. Here, we investigated the anti-inflammatory activity of solidagenone. Initially, we observed that a single dose of 30, 60 or 90 mg/Kg of solidagenone did not result in mortality or elicit any discernible signs of toxicity in the animals. At the same doses, solidagenone promoted a significative reduction in the migration of neutrophils in an acute peritonitis model and decreased mortality in a lipopolysaccharide-induced endotoxic shock model. Interestingly, treatment with solidagenone conferred a protective effect against leukopenia and thrombocytopenia, hematological disorders commonly observed in sepsis conditions. In addition, treatment with all the doses of solidagenone promoted a significative reduction in nitric oxide, TNF-α, and IL-1β relative to the LPS-stimulated vehicle-treated cultures. Furthermore, gene expression and in silico analyses also supported the modulation of NF-κB pathway by solidagenone. Finally, in silico pharmacokinetics prediction indicated a favorable drugability profile for solidagenone. Taken together, the findings of the present investigation show that solidagenone exhibits significant anti-inflammatory properties in acute experimental models, potentially through the modulation of NF-κB signaling pathway.
Keywords:
Solidagenone
; Inflammation
; Endotoxic shock
; NF-κB
1. Introduction
Inflammation assumes a pivotal role in rectifying imbalances within the body's homeostasis and is crucial for the repair, remodeling, and renewal of various tissues under diverse and adverse conditions [1]. Functioning as the primary line of defense, inflammation safeguards the host from infections induced by a spectrum of pathogens, including bacteria, fungi, parasites, and virus [2]. Additionally, others stimuli, such as cellular damage, chemical agents, physical injuries, burns, radiation, freezing, ischemia, and reperfusion, can incite inflammatory responses [3].
Classically, the treatment of inflammation entails the administration of non-steroidal anti-inflammatory drugs or glucocorticoids [4,5]. Both exhibit varying degrees of effectiveness in alleviating inflammation and they use are accompanied by a spectrum of adverse effects such as, atherosclerosis, cardiac alterations, gastrointestinal disorders, hypertension, renal toxicity, type 2 diabetes, visceral obesity, and others [5,6].
Therefore, the development of new substances exhibiting anti-inflammatory properties holds significant importance in the clinical use, with the goal of securing alternative treatments characterized by efficacy and diminished adverse effects. An attractive strategy in this search for new drugs involves the exploration of molecules with anti-inflammatory potential derived from medicinal plants, given the vast and yet undiscovered diversity of bioactive compounds within the Plant Kingdom [7,8]. Natural products stand out as a noteworthy reservoir for therapeutic interventions against various diseases, with the anti-inflammatory effects of traditional herbal medicines, crude plant extracts, natural compounds, and their derivatives having already demonstrated significant promise [7,9].
In this context, Solidago chilensis Meyen (Asteraceae) emerges as a highly promising reservoir for the discovery of anti-inflammatory agents. This plant species, with a historical use spanning over 700 years in South American folk medicine, has been recognized for its diverse therapeutic attributes, including anticancer, antidepressant, diuretic, gastroprotective, burn treatment, and anti-inflammatory activities [10,11,12,13].
Phytochemical analyses of S. chilensis have revealed a spectrum of chemical compounds, including flavonoids such as quercitrin, quercetin, and rutin, as well as diterpenes such as deoxysolidagenone, solidagolactol, and solidagenone [14]. Notably, the relatively underexplored compound solidagenone, isolated from the rhizomes, leaves, and inflorescences of S. chilensis, exhibits promise biological activities [13,15,16,17]. Solidagenone has demonstrated gastroprotective activity in a murine model of gastric injury induced by hydrochloric acid [15], attenuated skin inflammation in experimental models [16], exhibited antidepressant-like effects in mice [13], and a protective effect in a model of airway inflammation induced by ovalbumin [17]. Consequently, solidagenone holds considerable potential for further investigations into its anti-inflammatory activity. The present study is designed to assess the anti-inflammatory properties of solidagenone in acute experimental models of inflammation and its mechanism of action.
2. Results and Discussion
Firstly, the toxicity effects of a single dose of solidagenone was investigated. Administration of 30, 60 or 90 mg/Kg of solidagenone did not result in mortality or elicit any discernible signs of toxicity in the animals (Table 1). Furthermore, no significant variance in body weight was noted among animals treated with different doses of solidagenone when compared to mice treated only with vehicle solution (Table 2).
Toxicological investigations of bioactive compounds derived from medicinal plants are essential to ensure the safety of using herbs in the treatment of a series of diseases [18]. Therefore, it is relevant to analyze the acute toxicity generated by treatment with solidagenone in mice. Here we verified that the compound did not present toxicity at any of the administered doses used (30, 60, and 90 mg/Kg). These findings are consistent with the literature, considering that Rodriguez and collaborators [19], used a similar methodology used in this study and verified that solidagenone, intraperitoneally injected, did not show any observable symptoms of toxicity or mortality in Swiss mice treated with doses ranging from 100 to 600 mg/Kg.
A crucial index for evaluating the physiological or pathological state of mice subjected to experimental tests is body weight. Alterations in body weight often correlate with significant physiological changes, warranting a thorough analysis of this parameter [20]. In our study, we found that treatment with solidagenone did not induce significant changes in the weight of the animals.
Next, we assessed the anti-inflammatory activity of solidagenone, firstly, in a mouse model of acute peritonitis induced by carrageenan. As revealed in Figure 1, mice subjected to carrageenan stimulation and treated with a vehicle solution exhibited a mean of 274.8 neutrophils in 300 counted cells, a significative increase (p < 0.05) compare to naïve group which presented a mean of 1.8 neutrophils in 300 counted cells. In comparison to the vehicle-treated group, pre-treatment with solidagenone at doses of 30, 60, and 90 mg/kg resulted in a statistically significant reduction (p < 0.05) in neutrophil migration by 58.9%, 47.6%, and 41.9%, respectively. Under the same conditions, the administration of dexamethasone, at a dose of 30 mg/kg, also promoted a significative reduction (p < 0.05) of 59.6% in neutrophil migration (Figure 1). In agreement with these data, Liz and colleagues [21] demonstrated that an aqueous rhizome extract of S. chilensis and its two derived fractions promoted inhibition of leukocytes migration, particularly affecting neutrophil migration, and exudation in a mouse model of air pouch using carrageenan. Moreover, these effects were accompanied by a reduction of IL-1β, TNFα, and nitric oxide production, as well as a decrease in myeloperoxidase and adenosine deaminase activity [21]. Similar results, using a model of pleurisy induced by carrageenan, were found with extracts from leaves or inflorescences of S. chilensis [22].
Next, the anti-inflammatory effect of solidagenone was further investigated in a mouse model of endotoxic shock. As revealed in Figure 2, in comparison with the vehicle group, the animals treated with solidagenone at 60 and 90 mg/Kg exhibited an increased survival rate of 45.5 and 63.3% respectively, which was significant (p < 0.05). Under the same conditions, dexamethasone, administered at a dose of 30 mg/kg, also promoted a significant increase (p < 0.05) in survival rate of 63.3%.
As expected, LPS administration induced a leukopenia and thrombocytopenia in the animals (Table 3). Interestingly, treatment with solidagenone also conferred a significant (p < 0,05) protective effect against these hematological disorders, commonly observed in sepsis conditions. Dexamethasone, while promoting a more pronounced leukocyte recovery, did not afford protection against thrombocytopenia (Table 3).
For better understanding the anti-inflammatory effect of solidagenone, we measured the amount of nitric oxide and cytokines in the supernatant of resident macrophages from animals previously treated with solidagenone and stimulated in vitro with LPS plus IFNγ. As revealed in Figure 3, treatment with all the doses of solidagenone promoted a significative reduction in nitric oxide (Figure 3A), TNF-α (Figure 3B), and IL-1β (Figure 3C) relative to the LPS-stimulated vehicle-treated cultures. In a similar way, dexamethasone, administered at a dose of 30 mg/kg, also promoted a significative (p < 0.05) reduction of all the pro-inflammatory molecules evaluated (Figure 3). In agreement with these data, solidagenone also reduced NO, IL-1β, and TNFα production and gene expression of several inflammatory mediators (NOS2, IL1β, TNFα, and Cox2) in peritoneal macrophages stimulated in vitro with LPS plus IFNγ [17]. Moreover, the anti-inflammatory activity of solidagenone was also associated with nitric oxide, IL-6, and TNFα reduction in croton-oil-, arachidonic acid-, and phenol-induced ear oedema mouse models [16].
It is well-established that LPS induces Toll-like receptor-4 (TLR4) activation, triggering intracellular signaling pathways, including NF-ƙB activation, a pivotal transcription factor that modulates diverse pro-inflammatory genes such as IL-1β, NOS2, and TNF-α [23,24]. Notably, as revealed in Figure 4, treatment of activated macrophages in vitro with different concentrations of solidagenone resulted in a significant (p < 0.05) reduction in the gene expression of NF-ƙB. Similar results were obtained with the treatment with dexamethasone.
To understand the interaction between solidagenone and NF-κB, we investigate how solidagenone may interact with p65 subunit of NF-κB transcription factor and IκB kinase enzyme (IKK), through docking simulations. Both molecules are critical to preventing the displacement of NF-κB into the nucleus and consequently its activation [25,26]. Figure 5 shows solidagenone docked into the binding site of p65 subunit of NF-κB. As can be seen, solidagenone docks to NF-Κb system due seven van der Waals interactions (Lys-218, Arg-30, Phe-184, Asn-155, Pro189, Lys-79, and Thr-191), one alkyl interaction (Ala-192), one Pi-alkyl interaction (Ala-188), and one hydrogen bond interaction (Asn-190). The calculated docking energy was equal to -6.4 kcal/mol. Some other images regarding solidagenone-p65 subunit of NF-κB are shown in Supplementary material (Figure S1).
Regarding the IKK system, it is important to mention that it is organized in two chains (A and B), comprising four domains each: C-terminal dimerization domain (SDD), C-terminal kinase domain (KDC), N-terminal kinase domain (KDN), and ubiquitin-like domain (ULD) (Suplementary material, Figure S2). Since, according to Liu and coworkers [27], chain B is the one in the active conformation, this was the chosen chain used as the protein target for docking solidagenone. Figure 6 shows detains of simulated IKK-solidagenone complex. Solidagenone docks to IKK system due six van der Waals interactions (Lys-44, Asp-166, Lys-147, Thr-23, Gly-22, and Asn-150), one Pi-Sigma interaction (Leu-2), four Pi-alkyl interactions (Ala-42, Va-l29, Ile-165, and Val-152), and two hydrogen bond interactions (Asn-28 and Glu-149). The calculated docking energy was equal to -7.8 kcal/mol. Consequently, the solidagenone-IKK interaction is more stable than the solidagenone-NF-κB interaction. Figure S3 shows some images regarding solidagenone-IKK interaction.
In silico ADME profile of solidagenone is shown summarized in Figure 7 and Table S1. The Bioavailability Radar (Figure 7) bears six axes for six relevant properties for oral bioavailability. Each property is determine by a descriptor and a range of optimal values is depicted as a pink area. For saturation, the ratio of sp3 hybridized carbons over the total carbon count of the molecule (Fraction Csp3) should be at least 0.25. For size, the molecular weight should be between 150 and 500 g/mol. For polarity, the TPSA should be between 20 and 130 Å. For solubility, log S should not exceed 6. For lipophilicity, XLOGP3 should be in the range from − 0.7 to + 6.0. For flexibility, the molecule should not have more than 9 rotatable bonds. In this context, solidagenone ca be estimated as drug-like, since the red line is fully included in the pink area. Table S1 shows computed parameters for solidagenone, grouped in different sections (physicalchemical properties, lipofilicity, water solubility, pharmacokinetics, druglikeness, and medicinal chemistry).
3. Materials and Methods
3.1. Drugs
Solidagenone was obtained from Solidago chilensis inflorescences at Pharmaceutical Technology Institute (FarManguinhos, Oswaldo Cruz Foundation, Rio de Janeiro, Brazil) as previously described [16,28]. Dexamethasone (Sigma-Aldrich, St. Louis, MO) was used as positive control in anti-inflammatory experiments. All compounds were solubilized in Dimethyl sulfoxide (DMSO; PanReac, Barcelona, Spain) and diluted in culture medium for use in in vitro assays or saline for in vivo assays. The final concentration of DMSO wdid not extrapolate 0.1% in all in vitro assays or 10% in all in vivo analysis.
3.2. Animals
BALB/c mice (4 to 8 weeks old) were bred and housed at the Gonçalo Moniz Institute (Oswaldo Cruz Foundation, Bahia, Brazil) in sterilized cages, under controlled environmental conditions, and provided with a balanced rodent diet and water ad libitum. All animal experiments and procedures were conducted in accordance with the institution’s committee on the ethical handling of laboratory animals and were approved under the number L-IGM-29/2009.
3.3. Acute toxicity in mice
BALB/c mice (male; 6-8 weeks of age; n=6/ group) were divided into four experimental groups and treated orally with a single dose of with different doses of solidagenone (30, 60, and 90 mg/kg) or vehicle (saline solution with 10% DMSO). Following the completion of the treatment, the mice were monitored for general toxicity signs over a 15-day period. This involved the observation of morphological and behavioral changes. Additionally, the body mass of the animals was measured on days 0, 7, and 14.
3.4. Induction of acute peritonitis in mice
BALB/c mice (males; 6-8 weeks old) were divided into six experimental groups (n = 6) and treated orally with different doses of solidagenone (30, 60, and 90 mg/kg), dexamethasone (30 mg/kg) or vehicle (saline solution with 10% DMSO) 24 and 1 hour before the challenge. Subsequently, animals were challenged through an intraperitoneal injection of 250 μL of carrageenan (Sigma-Aldrich; 1 mg/mL; 250 μL). After 4 h, mice were euthanized and peritoneal exudates were harvested by peritoneal lavage using saline solution. Cells were centrifuged at x400 g for 10 min, at 4º C. Cytospin preparations were stained with rapid panoptic and a differential count of 300 cells was performed by a blinded investigator.
3.5. LPS-induced endotoxin shock
BALB/c mice (male;4-5 weeks of age;) were randomized into five experimental groups (n=11/ group) and treated orally with different doses of solidagenone (30, 60, and 90 mg/kg), dexamethasone (30 mg/kg) or vehicle (saline solution with 10% DMSO) 24 and 3 hour before the challenge. Subsequently, animals received a lethl dose of LPS (600 µg; from serotype 0111:B4 Escherichia coli, Sigma-Aldrich) in saline, by intraperitoneally route. The animals were monitored for 4 days to observe survival. In addition, a second set of experiments was performed and heparinized blood samples were collected after 6 h after challenge with LPS (600 µg) and to analyses leukocytes and thrombocytes using the Hematology Analyzer PE 7010VET (Shenzhen Prokan Eletronics Inc.).
3.6. Cytokine and nitric oxide production by resident macrophages
To evaluate nitric oxide and cytokines production by resident macrophages, groups of male BALB/c mice were orally treated with solidagenone, dexamethasone or vehicle in the doses described above. After 90 min, the mice were subjected to euthanasia for macrophage collection by means of peritoneal wash using cold Dulbecco's modified Eagle's medium (DMEM; Life Technologies, GIBCO-BRL, Gaithersburg, MD). Cells were washed twice with DMEM, resuspended in DMEM supplemented with 10% fetal bovine serum (FBS; GIBCO) and 50 μg/mL of gentamycin (Life Technologies, Carlsbad, CA), and plated in 96-well plates at a cell density of 2 × 105 cells/well. After 2 hours of incubation at 37°C, the plate was washed with saline solution and new medium was added to remove non-adherent cells. Cells were then activated or not with LPS (500 ng/mL) ) and IFNγ (5 ng/mL), and further incubated at 37° C and 5% CO2. Cell-free supernatants were collected at two different times after incubation for quantification of TNFα (4 hours) and for quantification of nitric oxide and IL-1β (24 hours) and kept at -80 °C until use.
3.7. Real time reverse transcription polymerase chain reaction (qRT-PCR)
Peritoneal exudate macrophages were obtained as previously described [17]. Then, cells were plated into 24-well plate at 1×106 cells/well in DMEM medium supplemented with FBS and gentamicin for 24 h at 37°C and 5% CO2. Cells were then pretreated or not with solidagenone (50, 25, and 12.5 µM) or dexamethasone (12.5 µM) for 1 h and then stimulated or not with LPS (500 ng/mL) and IFNγ (5 ng/mL) and incubated at 37oC for 3 h. After treatment, gene expression of NF-ƙb was performed as previously described [17]. The following primer sequences were used in real-time PCR assays: NFkb:5´-ATGGCAGACGATGATCCCTAC-3 and 3´-TGTTGACAGTGGTATTTCTGGTG-5.
3.8. Solidagenone structure
For molecular modeling purpose, the solidagenone structure was obtained from PubChem, a public chemical database at the National Center for Biotechnology Information (NCBI) of the National Library of Medicine (NLM), an institute within the U.S. National Institutes of Health (NIH) [29]. That structure was energy-minimized by using quantum mechanical modeling method DFT B3LYP/def2-TZVP, as implemented in Orca 5.0.3 software [30].
3.9. Crystallographic protein structures
The crystallographic structures of human proteins p65 subunit of NF-κB transcriptor factor, with resolution of 2.70 Å (PDB ID: 1NFI) [31], and IκB kinase enzyme (IKK), with resolution 2.83 Å (PDB ID: 4KIK) [27], were both obtained from the Protein Data Bank [32], and used in the docking simulations with solidagenone. The proteins preparation steps were carried out as follows: (1) non-essential water molecules were removed; (2) polar hydrogens were added to the protein; (3) partial charges were calculated using both Kollman [33], and Gasteiger’s approaches [34].
3.10. Molecular docking simulations
3.11. ADME in silico
SwissADME platform [38] was used for computing physicochemical and pharmacokinetics parameters related to adsorption, distribution, metabolism, and excretion (ADME) properties of solidagenone, as well as its drug-likeness.
3.12. Statistical analyses
Statistical analyses were performed using GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA). One-way analysis of variance (ANOVA) followed by Newman-Keuls multiple comparison tests were used for the comparison of groups. Statistical significance was considered when p-values were less than 0.05. The presented data are representative of at least two or three independent experiments.
4. Conclusions
Taken together, the findings of the present investigation show that solidagenone exhibits significant anti-inflammatory properties in acute experimental models, potentially through the modulation of NF-κB signaling pathway. Furthermore, solidagenone shows no acute toxicity and its in silico ADME profile suggests favorable drug-like characteristics. The presented data fortify the potential of solidagenone to leading to the development of alternative treatments with reduced adverse effects for inflammatory conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Solidagenone docked into the p65 subunit of NF-κB; Figure S2: IKK system; Figure S3: IKK system, and detail of solidagenone-IKK interaction. Table S1: ADME parameters, druglikeness, and medicinal chemistry of solidagenone.
Author Contributions
I.P.S.: Investigation, conceptualization, formal analysis, writing-original draft, L.P.S.: formal analysis, writing-original draft, D.K.C.S.: methodology, formal analysis, B.P.Z.C.d.R.: methodology, formal analysis, T.B.d.O.: methodology, formal analysis, A.M.K.: methodology, formal analysis, E.d.S.R.: methodology, formal analysis, C.V.C.d.S.: methodology, conceptualization, writing-review and editing, J.F.O.C.: conceptualization, investigation, formal analysis, S.S.V.: conceptualization, writing-review and editing, O.A.S.-F.: conceptualization, resources, writing-review, supervision, and editing, M.B.P.S.: conceptualization, writing—review and editing, supervision, funding acquisition, C.S.M.: conceptualization, formal analysis, writing—review and editing, supervision.
Funding
This work was supported by grants from PRONEX (grant number 0002/2014). M.B.P.S is a recipient of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) fellowship. E.d.S.R. is grateful for the PhD scholarship from Coordenação de Aperfeiçoamento de Pessoal de Nível superior—Brasil (CAPES)—Finance Code 001.
Institutional Review Board Statement
The study was approved by the Ethics Committee Institutional Animal Care and Use Committee FIOCRUZ (IGM 009/2009).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rathinam, V.A.K.; Chan, F.K. Inflammasome, Inflammation, and Tissue Homeostasis. Trends Mol. Med. 2018, 24, 304–318. [Google Scholar] [CrossRef]
- Hawiger, J.; Zienkiewicz, J. Decoding inflammation, its causes, genomic responses, and emerging countermeasures. Scand. J. Immunol. 2019, 90, e12812. [Google Scholar] [CrossRef] [PubMed]
- Roe, K. An inflammation classification system using cytokine parameters. Scand. J. Immunol. 2021, 93, e12970. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, S.; Migliorati, G.; Bruscoli, S.; Riccardi, C. Defining the role of glucocorticoids in inflammation. Clin. Sci. 2018, 132, 1529–1543. [Google Scholar] [CrossRef] [PubMed]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114–147. [Google Scholar] [CrossRef]
- Oray, M.; Samra, K.A.; Ebrahimiadib, N.; Messe, H.; Foster, C.S. Long-term side effects of glucocorticoids. Expert Opin. Drug Saf. 2016, 15, 457–465. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M. Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-Inflammatory Activity of Natural Products. Molecules. 2016, 21, 13–21. [Google Scholar] [CrossRef]
- Mors, W.; Rizzini, C.; Pereira, N. Medicinal plants of Brazil. J. Nat. Prod. 2001, 64, 1258–1259. [Google Scholar] [CrossRef]
- Catarino, H.R.; de Godoy, N.P.; Scharlack, N.K.; Neves, L.M.; de Gaspi, F.O.; Esquisatto, M.A.; do Amaral, M.E.; Mendonça, F.A.; dos Santos, G.M. InGaP 670-nm laser therapy combined with a hydroalcoholic extract of Solidago chilensis Meyen in burn injuries. Lasers Med. Sci. 2015, 30, 1069–79. [Google Scholar] [CrossRef]
- De Barros, M.; Mota da Silva, L.; Boeing, T.; Somensi, L.B.; Cury, B.J.; de Moura Burci, L.; Santin, J.R.; de Andrade, S.F.; Monache, F.D.; Cechinel-Filho, V. Pharmacological reports about gastroprotective effects of methanolic extract from leaves of Solidago chilensis (Brazilian arnica) and its components quercitrin and afzelin in rodents. Naunyn-Schmiedebergs Arch. Pharmacol. 2016, 389, 403–17. [Google Scholar] [CrossRef] [PubMed]
- Locateli, G.; de Oliveira Alves, B.; Miorando, D.; Ernetti, J.; Alievi, K.; Zilli, G.A.L.; Serpa, P.Z.; Vecchia, C.A.D.; Mota da Silva, L.; Müller, L.G.; Roman Junior, W.A. Antidepressant-like effects of solidagenone on mice with bacterial lipopolysaccharide (LPS)-induced depression. Behav. Brain Res. 2020, 395, 112–863. [Google Scholar] [CrossRef]
- Valverde, S.S.; Oliveira, T.B. , Souza, S.P. Solidago chilensis Meyen (Asteraceae). Revista Fitos, 2012, 7, 131–136. [Google Scholar] [CrossRef]
- Schmeda-Hirschmann, G.; Rodriguez, J.; Astudillo, L. Gastroprotective activity of the diterpene solidagenone and its derivatives on experimentally induced gastric lesions in mice. J. Ethnopharmacol. 2002, 81, 111–5. [Google Scholar] [CrossRef]
- Valverde, S.S.; Santos, B.C.S.; de Oliveira, T.B.; Gonçalves, G.C.; de Sousa, O.V. Solidagenone from Solidago chilensis Meyen inhibits skin inflammation in experimental models. Basic Clin. Pharmacol. Toxicol. 2021, 128, 91–102. [Google Scholar] [CrossRef]
- Vasconcelos, J.F.; Santos, I.P.; de Oliveira, T.B.; Kelly, A.M.; do Reis, B.P.Z.C.; Orge, I.D.; Meira, C.S.; Valverde, S.S.; Soares, M.B.P. The protective effect of solidagenone from Solidago chilensis Meyen in a mouse model of airway inflammation. Basic Clin. Pharmacol. Toxicol. 2022, 130, 44–55. [Google Scholar] [CrossRef]
- Dashti, A.; Shokrzadeh, M.; Karami, M.; Habibi, E. Phytochemical identification, acute and subchronic oral toxicity assessments of hydroalcoholic extract of Acroptilon repens in BALB/c mice: A toxicological and mechanistic study. Heliyon. 2022, 8, e08940. [Google Scholar] [CrossRef]
- Rodríguez, J.A.; Bustamante, C.; Astudillo, L.; Schmeda-Hirschmann, G. Gastroprotective activity of solidagenone on experimentally-induced gastric lesions in rats. J. Pharm. Pharmacol. 2002, 54, 399–404. [Google Scholar] [CrossRef]
- Ugwah-Oguejiofor, C.J.; Okoli, C.O.; Ugwah, M.O.; Umaru, M.L.; Ogbulie, C.S.; Mshelia, H.E.; Umar, M.; Njan, A.A. Acute and sub-acute toxicity of aqueous extract of aerial parts of Caralluma dalzielii N. E. Brown in mice and rats. Heliyon. 2019, 5, e01179. [Google Scholar] [CrossRef]
- Liz, R.; Vigil, S.V.; Goulart, S.; Moritz, M.I.; Schenkel, E.P.; Fröde, T.S. The anti-inflammatory modulatory role of Solidago chilensis Meyen in the murine model of the air pouch. J. Pharm. Pharmacol. 2008, 60, 515–21. [Google Scholar] [CrossRef]
- Goulart, S.; Moritz, M.I.; Lang, K.L.; Liz, R.; Schenkel, E.P.; Fröde, T.S. Anti-inflammatory evaluation of Solidago chilensis Meyen in a murine model of pleurisy. J. Ethnopharmacol. 2007, 113, 346–53. [Google Scholar] [CrossRef]
- Bartuzi, P.; Hofker, M.H.; van de Sluis, B. Tuning NF-κB activity: a touch of COMMD proteins. Biochim. Biophys. Acta. 2013, 1832, 2315–21. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: mechanism and clinical study. Signal Transduct. Targ. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metabolism. 2011, 13, 11–22. [Google Scholar] [CrossRef]
- Wei, H.; Wang, B.; Miyagi, M.; She, Y.; Gopalan, B.; Huang, D.-B.; Ghosh, G.; Stark, G.R.; Lu, T. PRMT5 dimethylates R30 of the p65 subunit to activate NF-κB. PNAS. 2013, 110, 13516–13521. [Google Scholar] [CrossRef]
- Liu, S.; Misquitta, Y.R.; Olland, A.; Johnson, M.A.; Kelleher, K.S.; Kriz, R.; Lin, L.L.; Stahl, M.; Mosyak, L. Crystal structure of a human IκB kinase β asymmetric dimer. J Biol Chem. 2013, 288, 22758–22767. [Google Scholar] [CrossRef]
- Valverde, S.S.; Azevedo, S.S.; Tomassini, T.C.B. Utilização de CLAE, como paradigma na obtenção e controle do diterpeno solidagenona a partir de inflorescências de Solidago chilensis Meyen (arnica brasileira). Rev. Bras. Farm. 2009, 90, 196–99. [Google Scholar]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; Zaslavsky, L.; Zhang, J.; Bolton, E.E. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef]
- Neese, F. The ORCA Program System. Wiley Interdisciplinary Reviews: Computational Molecular Science, 2012, 2, 73–78. [Google Scholar] [CrossRef]
- Jacobs, M.D.; Harisson, S.C. Structure of an IkappaBalpha/NF-kappaB complex. Cell 1998, 95, 749–758. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gililand, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: the RESP model. J. Phys. Chem. 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Gasteiger, J.; Marsili, M. Iterative partial equalization of orbital electronegativity—a rapid access to atomic charges. Tetrahedron 1980, 36, 3129–3228. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–61. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–12. [Google Scholar] [CrossRef] [PubMed]
- BIOVIA, Dassault Systèmes; 21.1.0; Discovery Studio Visualizer; Dassault Systèmes: San Diego, CA, USA, 2021.
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
Figure 1.
Effect of solidagenone in a model of acute peritonitis. BALB/c mice (n=6/group) were submitted to a challenge with 1% carrageenan solution after treatment with solidagenone (30, 60, and 90 mg/Kg) or dexamethasone (Dexa; 30 mg/Kg) or vehicle (saline solution with 10% DMSO). Untreated and unchallenged animals consists of naïve group. Values represent the means ± S.D. of six mice/group. *p < 0.05 compared to the vehicle group; #p < 0.05 compared to naïve group.
Figure 1.
Effect of solidagenone in a model of acute peritonitis. BALB/c mice (n=6/group) were submitted to a challenge with 1% carrageenan solution after treatment with solidagenone (30, 60, and 90 mg/Kg) or dexamethasone (Dexa; 30 mg/Kg) or vehicle (saline solution with 10% DMSO). Untreated and unchallenged animals consists of naïve group. Values represent the means ± S.D. of six mice/group. *p < 0.05 compared to the vehicle group; #p < 0.05 compared to naïve group.
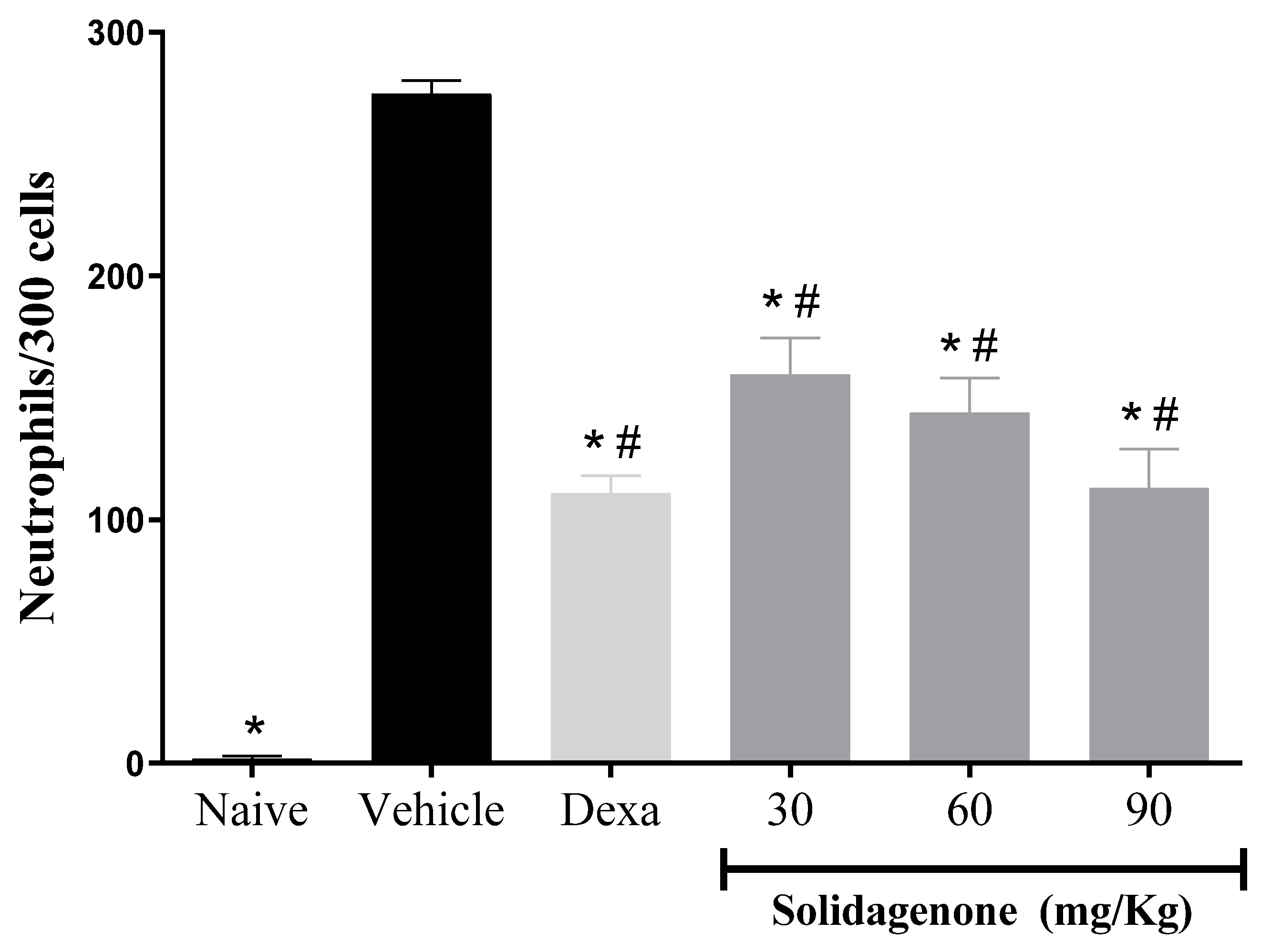
Figure 2.
Solidagenone promoted an increase in the survival rate of animals challenged with LPS.. Mice were orally treated with solidagenone at doses of 30 mg/kg (■), 60 mg/Kg (▲) and 90 mg/kg (▼) or dexamethasone at a dose of 30 mg/kg (♦), or vehicle (●). The animals were monitored for 4 days to observe survival. Data from two independent experiments are represented in the graph.. *p < 0.05 compared to the vehicle group. **p < 0.01 compared to the vehicle group. Statistical analysis were performed using Logrank (Mantel Cox).
Figure 2.
Solidagenone promoted an increase in the survival rate of animals challenged with LPS.. Mice were orally treated with solidagenone at doses of 30 mg/kg (■), 60 mg/Kg (▲) and 90 mg/kg (▼) or dexamethasone at a dose of 30 mg/kg (♦), or vehicle (●). The animals were monitored for 4 days to observe survival. Data from two independent experiments are represented in the graph.. *p < 0.05 compared to the vehicle group. **p < 0.01 compared to the vehicle group. Statistical analysis were performed using Logrank (Mantel Cox).
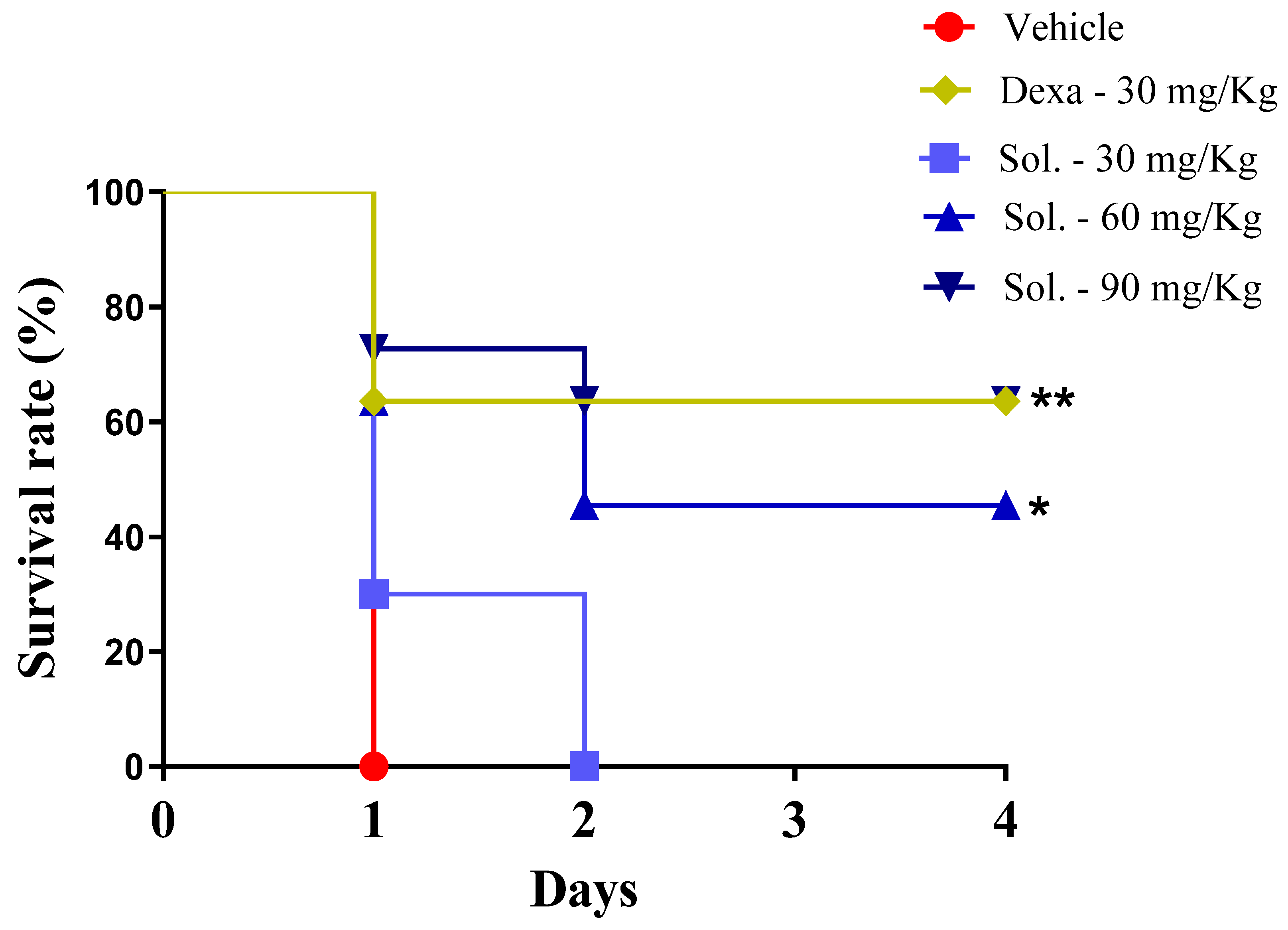
Figure 3.
In vivo treatment with solidagenone reduces nitric oxide and cytokines production in macrophages stimulated with LPS. Resident peritoneal macrophages were collected from animals treated with different doses of solidagenone (30, 60, and 90 mg/Kg) or dexamethasone (Dexa; 30 mg/Kg) or vehicle (saline solution with 10% DMSO). Macrophages were plated in 96-well plates in the absence or presence of LPS plus IFNγ and, cell-free supernatants were collected after 4 or 24 h,. Levels of nitrite (A), TNF-α (B), and IL-1β (C) were obtained throw Griess method and ELISA respectively. Values represent the means ± S.D. of six mice/group. *p < 0.05 compared to the vehicle group; #p < 0.05 compared to naïve group. $p < 0.05 compared to dexamethasone group.
Figure 3.
In vivo treatment with solidagenone reduces nitric oxide and cytokines production in macrophages stimulated with LPS. Resident peritoneal macrophages were collected from animals treated with different doses of solidagenone (30, 60, and 90 mg/Kg) or dexamethasone (Dexa; 30 mg/Kg) or vehicle (saline solution with 10% DMSO). Macrophages were plated in 96-well plates in the absence or presence of LPS plus IFNγ and, cell-free supernatants were collected after 4 or 24 h,. Levels of nitrite (A), TNF-α (B), and IL-1β (C) were obtained throw Griess method and ELISA respectively. Values represent the means ± S.D. of six mice/group. *p < 0.05 compared to the vehicle group; #p < 0.05 compared to naïve group. $p < 0.05 compared to dexamethasone group.
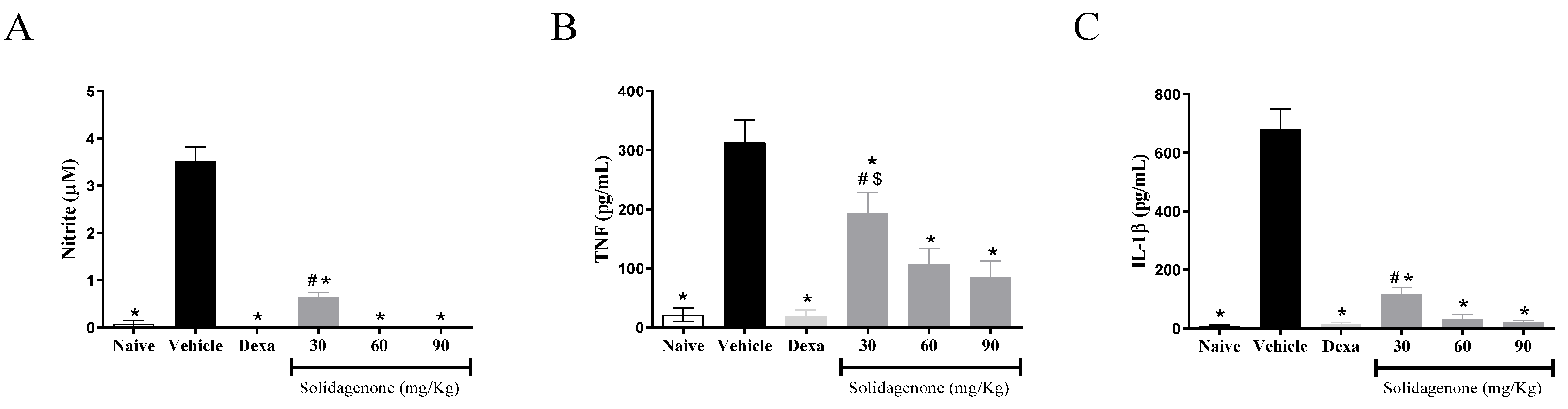
Figure 4.
Solidagenone treatment reduces NF-ƙβ expression levels in peritoneal macrophages. Cells were stimulated or not with LPS + IFN-γ and treated or not with solidagenone or dexamethasone. Analysis of gene expression was made by real-time qRT-PCR. Values represent the means ± S.D. of four determinations obtained in one of two assays performed. *p < 0.05 compared to stimulated and untreated cells; #p < 0.05 compared to unstimulated and untreated cells; $p < 0.05 compared to cells treated with dexamethasone.
Figure 4.
Solidagenone treatment reduces NF-ƙβ expression levels in peritoneal macrophages. Cells were stimulated or not with LPS + IFN-γ and treated or not with solidagenone or dexamethasone. Analysis of gene expression was made by real-time qRT-PCR. Values represent the means ± S.D. of four determinations obtained in one of two assays performed. *p < 0.05 compared to stimulated and untreated cells; #p < 0.05 compared to unstimulated and untreated cells; $p < 0.05 compared to cells treated with dexamethasone.
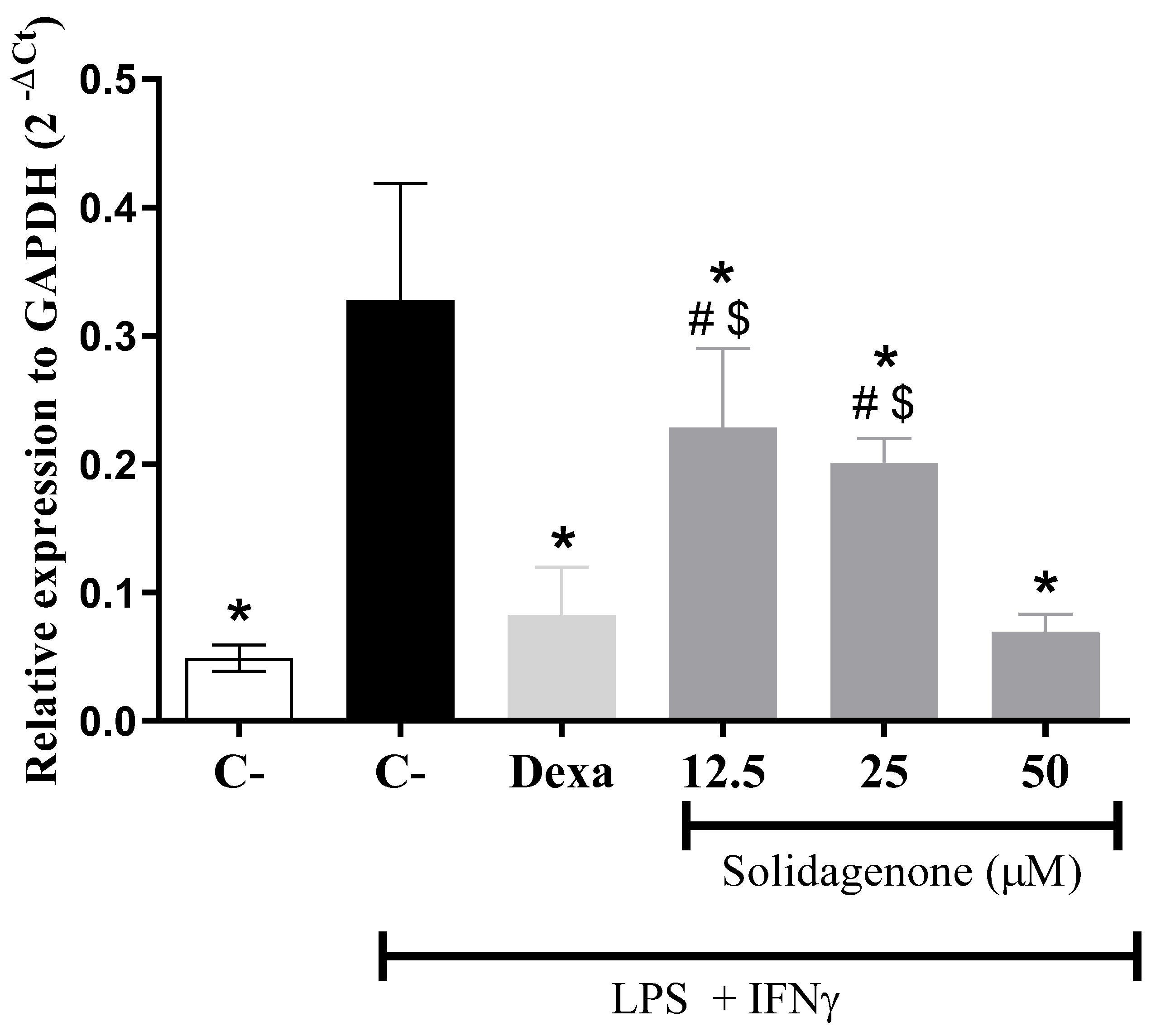
Figure 5.
Docking analysis of solidagenone into the bind site of p65 subunit of NF-κB transcription factor. (a) x of selected protein (PDB ID: 1NFI) and its interactions with solidagenone. (b) 2D interactions of solidagenone with amino acid residues in the p65 subunit of NF-κB.
Figure 5.
Docking analysis of solidagenone into the bind site of p65 subunit of NF-κB transcription factor. (a) x of selected protein (PDB ID: 1NFI) and its interactions with solidagenone. (b) 2D interactions of solidagenone with amino acid residues in the p65 subunit of NF-κB.
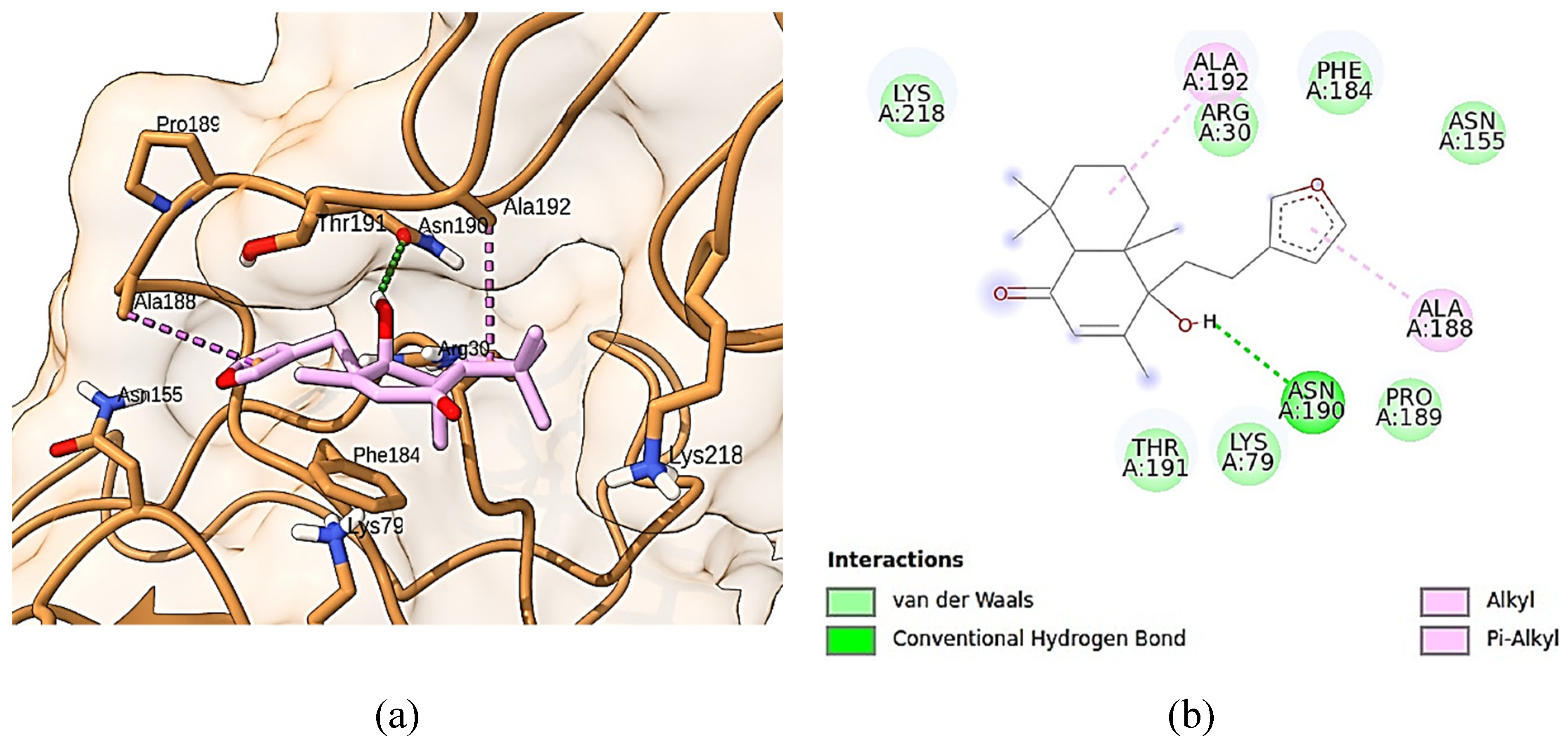
Figure 6.
Docking analysis of solidagenone into IKK domain. (a) Three-dimensional structure of selected protein (PDB ID: 4KIK) and its interaction with solidagenone. (b) 2D interactions of solidagenone with amino acid residues in the IKK domain.
Figure 6.
Docking analysis of solidagenone into IKK domain. (a) Three-dimensional structure of selected protein (PDB ID: 4KIK) and its interaction with solidagenone. (b) 2D interactions of solidagenone with amino acid residues in the IKK domain.
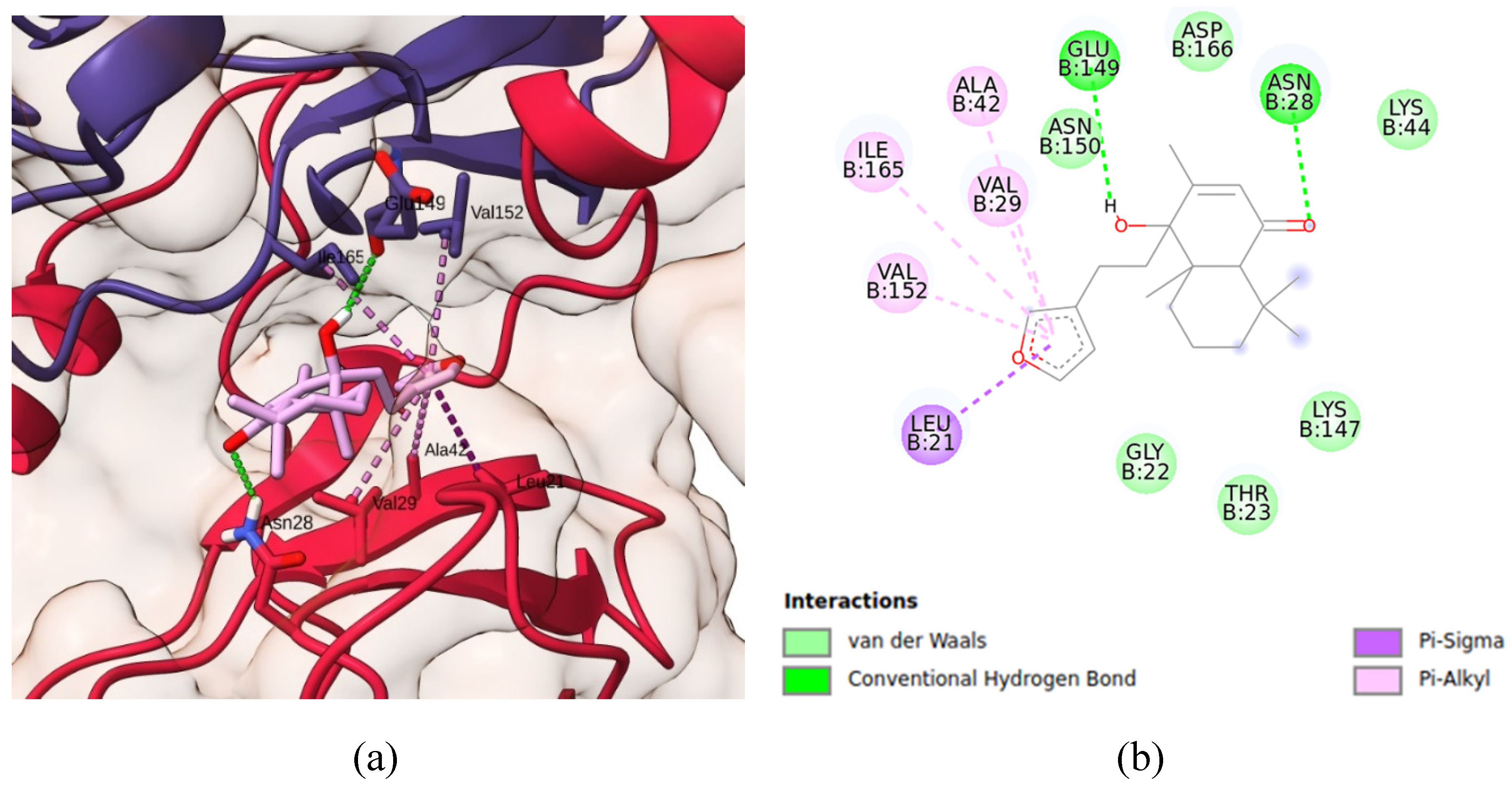
Figure 7.
SwissADME plot of drug-likeness of solidagenone. The pink area represents the optimal range for each property. For saturation (INSATU), the ratio of sp3 hybridized carbons over the total carbon count of the molecule (Fraction Csp3) should be at least 0.25. For size, the molecular weight should be between 150 and 500 g/mol. For polarity (POLAR), the TPSA should be between 20 and 130 Å. For solubility (INSOLU), log S should not exceed 6. For lipophilicity (LIPO), XLOGP3 should be in the range from − 0.7 to + 6.0. For flexibility (FLEX), the molecule should not have more than 9 rotatable bonds.
Figure 7.
SwissADME plot of drug-likeness of solidagenone. The pink area represents the optimal range for each property. For saturation (INSATU), the ratio of sp3 hybridized carbons over the total carbon count of the molecule (Fraction Csp3) should be at least 0.25. For size, the molecular weight should be between 150 and 500 g/mol. For polarity (POLAR), the TPSA should be between 20 and 130 Å. For solubility (INSOLU), log S should not exceed 6. For lipophilicity (LIPO), XLOGP3 should be in the range from − 0.7 to + 6.0. For flexibility (FLEX), the molecule should not have more than 9 rotatable bonds.
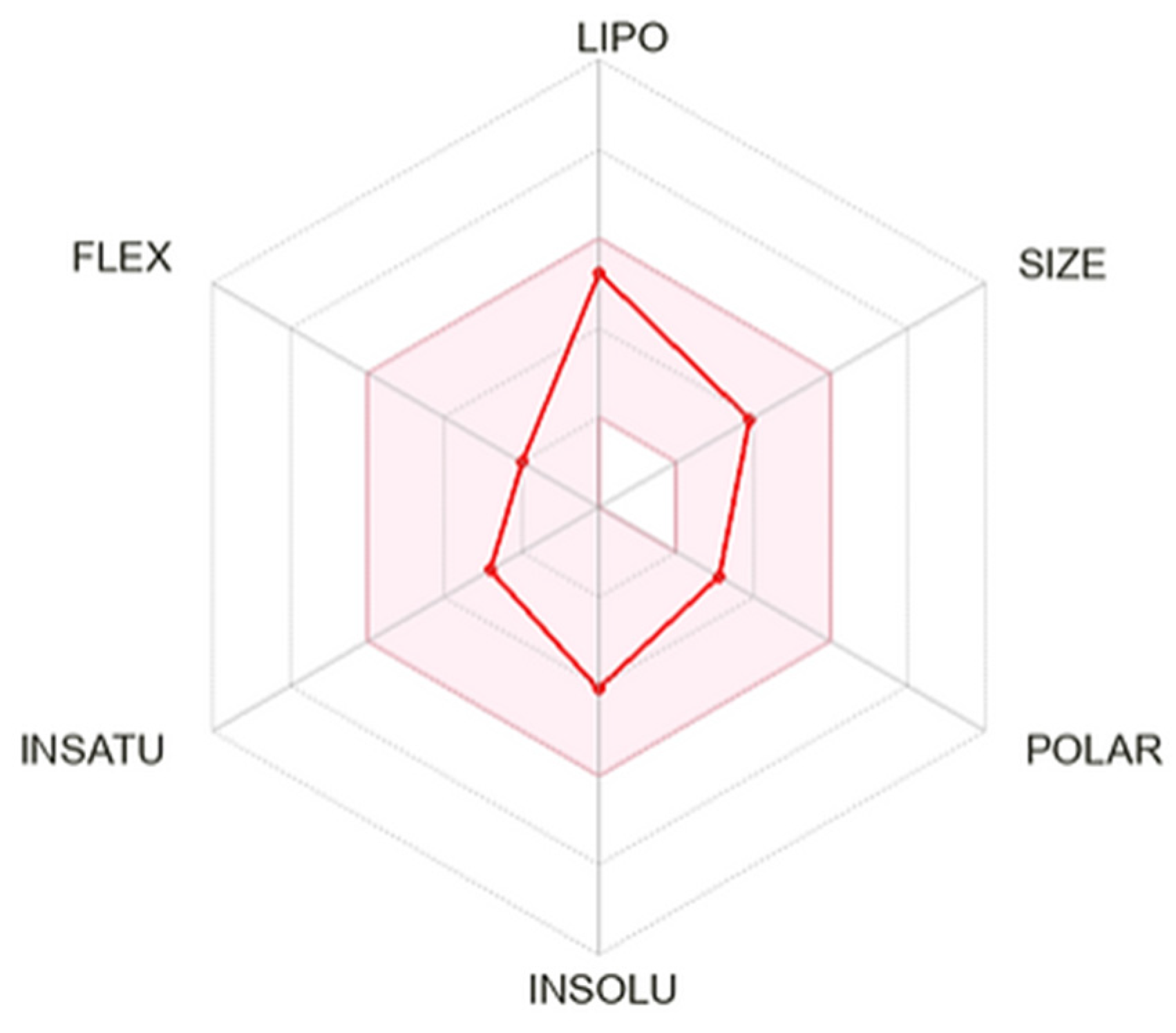

Table 1.
Effect of Solidagenone on behavioral and general appearance of male BALB/c mice.
| Behavior and general appearance | Observations | |||
|---|---|---|---|---|
| Vehicle | Solidagenone (30 mg/Kg) |
Solidagenone (60 mg/Kg) |
Solidagenone (90 mg/Kg) |
|
| Changes in the eyes | No changes | No changes | No changes | No changes |
| Changes in the fur | No changes | No changes | No changes | No changes |
| Changes in the skin | No changes | No changes | No changes | No changes |
| Coma | Absent | Absent | Absent | Absent |
| Convulsions | Absent | Absent | Absent | Absent |
| Diarrhea | Absent | Absent | Absent | Absent |
| Lethargy | Absent | Absent | Absent | Absent |
| Salivation | Absent | Absent | Absent | Absent |
| Sleep | Usual | Usual | Usual | Usual |
| Tremors | Absent | Absent | Absent | Absent |
The animals were subjected to daily observations for a duration of 14 days.
Table 2.
Body weight of BALB/c mice treated with the compound solidagenone.
| Days | Vehicle | Solidagenone (30 mg/Kg) |
Solidagenone (60 mg/Kg) |
Solidagenone (90 mg/Kg) |
|---|---|---|---|---|
| 0 | 24.3 (± 0.6) | 24.1 (± 0.3) | 23.8 (± 1.6) | 22.8 (± 0.9) |
| 7 | 24.2 (± 0.3) | 24.7 (± 0.5) | 24.0 (± 1.8) | 22.8 (± 1.2) |
| 14 | 24.4 (± 0.5) | 24.8 (± 0.6) | 24.5 (± 1.5) | 23.0 (± 0.7) |
Values represent the mean ± standard deviation of six animals per group.
Table 3.
Solidagenone attenuates leukopenia and thrombocytopenia in mice challenged with a LPS.
| Group | Dose (mg/Kg) | Leukocytes (103 cells/µL) |
Thrombocytes (103 mm3) |
|---|---|---|---|
| Naive | - | 3.8 ± 0.9* | 449.8 ± 49.2* |
| Vehicle | - | 1.7 ± 0.2 | 334.5 ± 36.7 |
| Dexamethasone | 30 | 3.8 ± 0.4* | 337.0 ± 26.8 |
| Solidagenone | 30 | 1.9 ± 0.3 | 365.8 ± 15.2 |
| Solidagenone | 60 | 2.2 ± 0.4 | 399.4 ± 3.4* |
| Solidagenone | 90 | 2.7 ± 0.5* | 403.5 ± 40.4* |
Values represent the mean ± standard deviation of six animals per group. *p < 0.05 compared to the vehicle group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



