
Submitted:
04 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Urinary tract infections (UTIs) caused by Klebsiella pneumoniae can lead to severe clinical complications and even death. An alternative treatment option for infected patients is using bacteriophages (phages). In the present study, we isolated phage KP-1 from sewage water using K. pneumoniae as a host. Whole genome analysis indicated that the genome was a double-stranded linear 176,096-bp long DNA molecule with 41.8% GC content and did not contain virulence or antibiotic resistance genes. The inactivation potential of phage KP-1 was assessed in broth at an MOI of 1 and 10, and a maximum inactivation of 4.9 and 5.4 log CFU/mL was observed after 9 h, respectively. The MOI of 10 efficacy was also assessed in urine to evaluate the phage’s performance in an acidic environment. A maximum inactivation of 3.8 log CFU/mL was observed after 9 h. The results indicate that phage KP-1 could be used to control UTIs caused by K. pneumoniae; however, further studies using phage cocktails to broader the phage’s spectrum of activity and phages in micro- or nanocarriers to avoid the negative effect of the low pH of the urine on phage viability, are necessary.
Keywords:
Bacteriophage
; Phage therapy
; Klebsiella pneumoniae
; Urinary tract infections
1. Introduction
The World Health Organization has classified ESKAPEE, a group of seven highly virulent bacteria (Enterococcus faecium, Staphylococcus aureus, K. pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterobacter spp. and Escherichia coli), as number one priority for the development of new antimicrobial drugs [1]. Among these, the Gram-negative bacterium K. pneumoniae poses a significant global health threat due to the widespread dissemination of carbapenemase genes [2]. In clinical settings, K. pneumoniae is a common cause of bloodstream, respiratory and urinary tract infections (UTIs) [3]. Notably, in intensive care units, most healthcare-associated infections are associated with endotracheal tubes (cases of pneumonia), vascular catheters (bloodstream infections) or urinary catheters (UTIs) [4]. According to the 2019 ECDC report, 94% of UTIs were associated with the presence of urinary catheters [4]. The adhesive fimbriae of K. pneumoniae enables the formation of robust biofilms on medical devices [5]. These biofilms confer additional resistance to the patient’s immune response and antibiotics, complicating clinical outcomes and limiting treatment options [6]. In the most recent antimicrobial resistance surveillance report, K. pneumoniae isolates presented widespread resistance to third-generation cephalosporins and increasing resistance to carbapenems in a worsening scenario across Europe [7]. The resistance to these antibiotics can also be transferred to other bacteria during the biofilm matrix formation, affecting the treatment options available for other bacteria [6]. Complications arise in patients with UTIs where antimicrobial-resistant bacteria are detected, leading to prolonged hospitalizations, higher rates of treatment failure and even death [8]. As the antimicrobial treatment landscape worsens, there is an urgent need for novel treatment options in clinical settings.
Phages, viruses specific to bacteria, offer several advantages over antibiotics, including low environmental impact, preservation of the microbiota, ease and cost-effectiveness of isolation, and resistance to cross-resistance [9]. Furthermore, phages can infect antibiotic-resistant bacteria and, in some cases, enhance bacterial susceptibility to antibiotics where resistance has been observed [10]. Therefore, researchers worldwide are exploring their potential applications in various fields [11]. Given that patients in intensive care units are prone to UTIs [12,13], the use of phages to treat these infections has received considerable attention [14]. Commercially available phage preparations from the ELIAVA Institute have been used against isolated E. coli and K. pneumoniae strains obtained from UTI patients [15]. These phages were even directly administered to treat UTIs in patients who underwent transurethral resection of the prostate [16]. In a different approach, Le and his coworkers (2023) successfully treated a recurrent UTI caused by K. pneumoniae through intravenous administration of phages without antibiotics. Other researchers have isolated and characterized phages specific to K. pneumoniae, testing their efficacy against biofilm formation and in animal models [18,19]. However, some isolated and characterized phages exhibit a very narrow host range and cannot infect different bacterial strains of the same species [9]. Implementing phage libraries may address this limitation, allowing researchers to select the most effective phage preparations for a specific strain [20]. As the levels of K. pneumoniae resistance to carbapenems continue to increase in Europe and specifically in Portugal [7], the isolation and characterization of new phages capable of infecting this bacterium become crucial for providing new treatment options for patients. Therefore, this study focuses on isolating and characterizing a new phage capable of infecting an environmentally isolated carbapenemase strain-producing K. pneumoniae. In addition, in vitro tests in liquid medium and human urine were conducted to assess the phage's potential for use in urinary tract infections, providing insight into its efficacy in the acidic environment of the urinary tract.
2. Results
2.1. Phage isolation and virion morphology
Phage KP-1 formed very small (diameter <0.1 mm) clear plaques on the host strain (Figure 1. A). High titre suspensions [109 plaque-forming units (PFU/mL)] were produced. Transmission electron microscopy analysis revealed virions with an icosahedral head of approximately 76 nm and a contractile tail of 110 ± 7.9 nm. Phage KP-1 presents a myovirus morphotype and has been classified as a Caudoviricetes.
2.2. Genome analysis
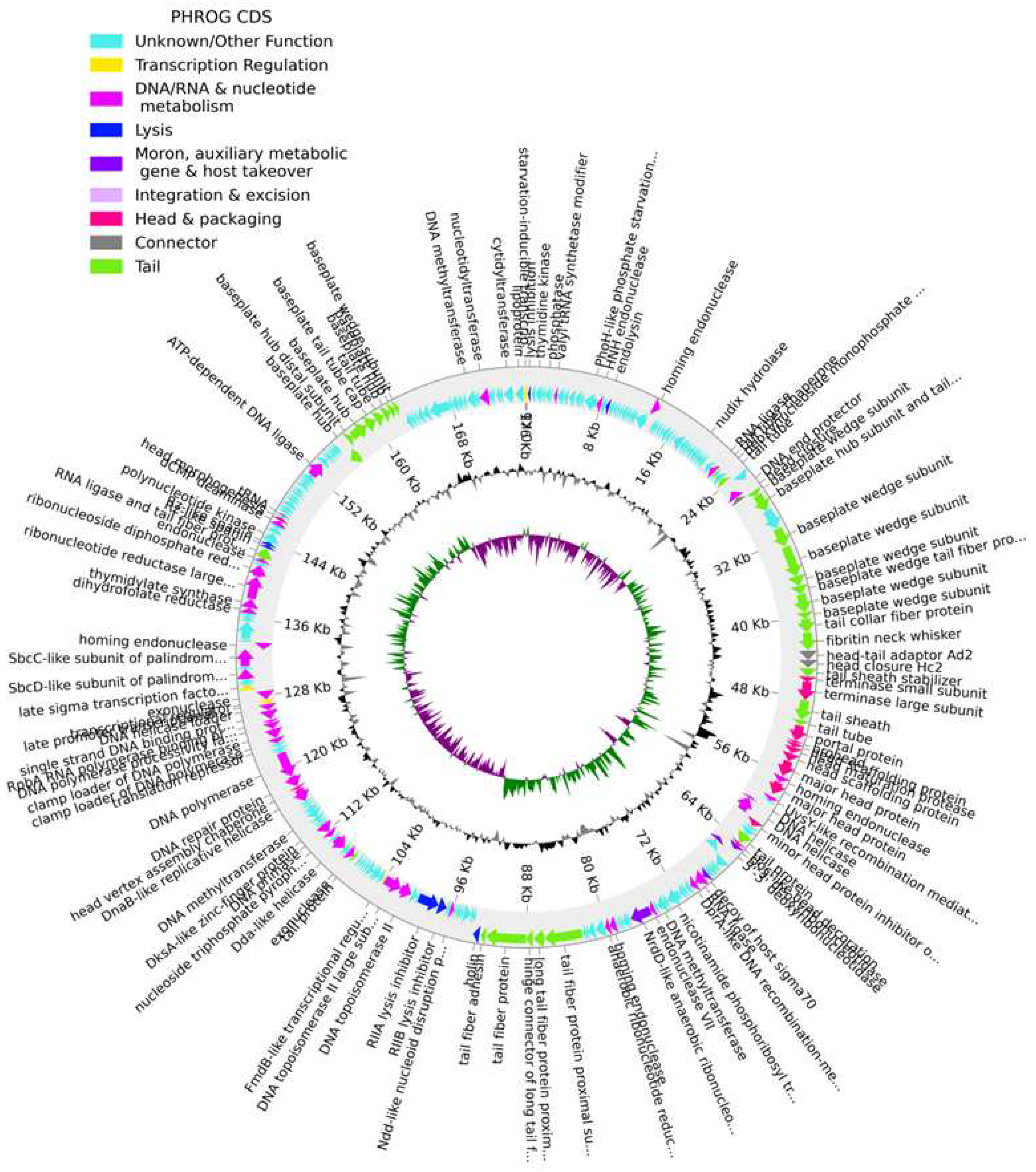
The genome of phage KP-1 (GenBank: PP002985) is a double-stranded DNA with a size of 176,096 bp, and the GC content is 41.8 %. The CheckV results showed 98.8 % of completeness and no host contamination. A total of 268 CDSs were predicted in the phage KP-1 genome, with 148 CDSs predicted as hypothetical proteins and 120 CDSs as functional proteins (Figure 2, Table S1). Our analysis could not detect any genes encoding antibiotic resistance or virulence determinants in the phage KP-1 genome. The absence of the phage integrase gene indicates that phage KP-1 could potentially be exploited for biocontrol applications. We further detected the presence of two tRNA genes in the KP-1 genome suggesting that the phage may rely on its own tRNA for translation after infecting the host cell (Figure 2, Table S1). Overall we detected putative genes encoding for phage proteins involved in DNA replication and modification, phage structural and packaging and host lysis (e.g., endolysin and holin) (Figure 2, Table S1). The genome sequence of phage KP-1 was classified under the Slopekvirus genus and had a similarity of 98.87 % with Klebsiella phage phiKp_26 (GenBank: LC768496) as well with a set of Klebsiella phages isolated from sewage.
Figure 2.
The genome map of phage KP-1. The outer circle with arrow-headed bands represents the coding DNA sequences (CDS) color coded according to the functional category of the predicted gene in the direction of the transcription. The innermost ring represents the genome GC skew (green/pink) followed by GC content (black/grey). The labels show the predicted functions of the functional CDSs, color-coded by the PHROGs category.
Figure 2.
The genome map of phage KP-1. The outer circle with arrow-headed bands represents the coding DNA sequences (CDS) color coded according to the functional category of the predicted gene in the direction of the transcription. The innermost ring represents the genome GC skew (green/pink) followed by GC content (black/grey). The labels show the predicted functions of the functional CDSs, color-coded by the PHROGs category.
2.3. Phage host range and efficiency of plating
The spot test analysis revealed that phage KP-1 presented a very narrow host range infecting only E. coli ATCC 13706 of the 52 strains tested, with an efficiency of plating of 32.98 ± 2.85% (Table 1).
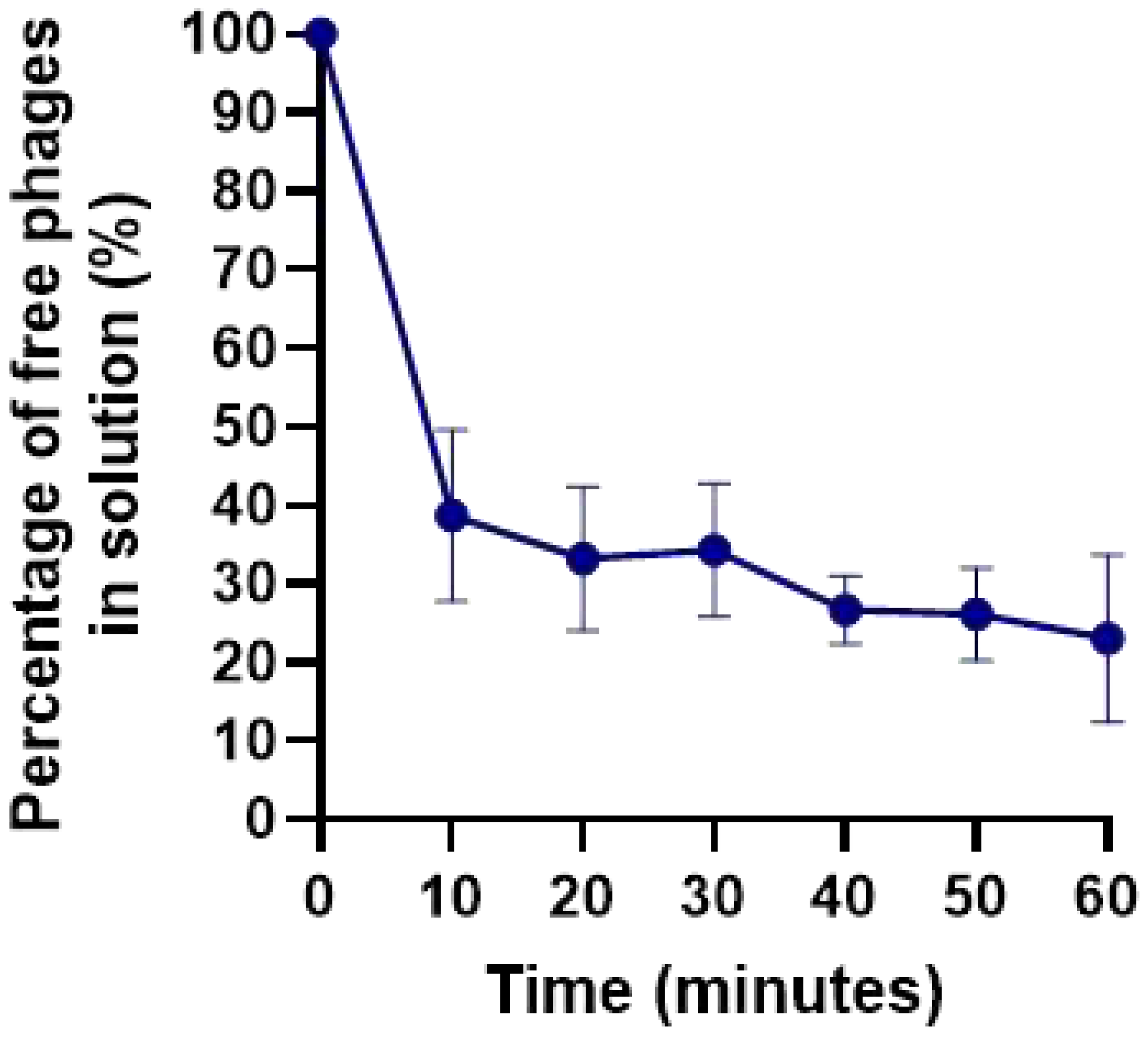
2.4. Adsorption curve
Phages KP-1 adsorption assays showed that approximately 60% and 75% of the phage particles adsorb to K. pneumoniae Scc 24 cells after 10 and 60 min, respectively (Figure 7).
Figure 2.
Adsorption curves of phages KP-1 in the presence of K. pneumoniae as the host. Values represent the mean of three independent experiments, and error bars represent the standard deviation.
Figure 2.
Adsorption curves of phages KP-1 in the presence of K. pneumoniae as the host. Values represent the mean of three independent experiments, and error bars represent the standard deviation.
2.5. One-step growth curve
The growth curve for phage KP-1 was determined in Tryptic Soy Broth (TSB, Liofilchem, Roseto degli Abruzzi, Italy) at a temperature of 37 °C (Figure 3). From the observed results, phage KP-1 presented an eclipse time of 80 min, a latent period of 100 min and a burst size of 7.9 ± 0.3 PFU/host cell.
2.6. Bacterial kill curve
Bacterial kill curves were performed in TSB at 37°C using two different MOIs to determine whether increasing the phage dose would promote higher bacterial inactivation. In both assays, the bacterial control increased from 4.7 ± 0.16 x 105 CFU/mL to 8.9 ± 0.14 x 108 CFU/mL (ANOVA, p < 0.05, Figure 4A). When the bacterium was challenged with the phage at MOIs of 1 and 10, a maximum bacterial decrease of 4.9 and 5.4 log CFU/mL was observed after 9 h of incubation, respectively.
When higher doses of phage were used, it was observed that bacterial densities started to decrease earlier (after 6 h) than when a MOI of 1 was used (ANOVA, p < 0.05, Figure 4A). However, the maximum inactivation, observed after 9 hours, did not present statistically significant differences (ANOVA, p > 0.05, figure 4A). Nonetheless, when a MOI of 10 was used, the bacterial numbers remained constant for a more prolonged period (12-24 h), with statistically significant differences when compared with the MOI of 1 (ANOVA, p < 0.05, Figure 3A). Therefore, an MOI of 10 was selected for further studies.
Phage KP-1 remained stable during this assay with no variation in phage titre during the 24 h in the control group of both MOIs studied (Figure 3B). When phages were incubated in the presence of the host bacteria, a significant increase in phage titre of 2.9 and 1.7 log PFU/mL was observed for both test groups (ANOVA, p < 0.05, Figure 3B).
2.7. Bacterial kill curve in urine
Fresh, early morning urine was filtered using a 0.22 µm syringe filter and inoculated with phage KP-1 and phage to a final MOI of 10. During the 24-hour bacterial challenge, a maximum bacterial inactivation of 3.8 log CFU/mL was observed after 9 h (ANOVA, p < 0.05, Figure 5A). Despite bacterial regrowth, the differences in bacterial density between the test and control groups remained statistically significant until the end of the experiment (ANOVA, p < 0.05, Figure 5A). Phage remained stable with no significant differences in the control group (ANOVA, p > 0.05, Figure 5B). In the presence of the host, the phage titer increased significantly by 2.7 log PFU/mL(ANOVA, p < 0.05, Figure 5B).
2.8. Frequency of emergence of phage-resistant mutants
When K. pneumoniae Scc 24 was challenged with phage KP-1, a frequency of emergence of phage-resistant mutants of 5.55x10-3 ± 0.002 was observed. After incubation for 24 h at 37 °C, a bacterial density of 4.51x108 ± 0.61 CFU/mL was observed for the control group. However, only 2.52x104 ± 0.93 CFU/mL was observed after 24 h when the bacteria were treated with phage.
3. Discussion
The rise of carbapenem-resistant K. pneumoniae poses a global public health threat [21,22]. Phages are increasingly considered an alternative or adjuvant treatment option, especially when antibiotics prove ineffective in treating bacterial infections [23,24,25,26]. Phage therapy is explored when patients do not respond to conventional antibiotic treatments [17]. However, it is still considered an experimental approach [27] and requires emergency regulatory approval [27], meaning that more data and research are needed to advance its integration into modern healthcare strategies. In the present study, phage KP-1 was isolated using a strain of carbapenemase-producing K. pneumoniae, and its biological properties and in vitro efficacy were investigated.
The genome of the K. pneumoniae phage KP-1 (size range 19,260–346,602bp) [28] is a lytic phage classified as Slopekvirus (with a 176,096 bp linear double-stranded DNA). It does not appear to encode any known endolysin, toxin, virulence or antibiotic resistance genes and can be considered safe for use in phage therapy.
Although host specificity is considered one of the most advantageous properties of phages, distinguishing them from antibiotics, the narrow host range can be an obstacle to efficient phage therapy [9]. Phage KP-1 showed a narrow host spectrum but can also infect E. coli ATCC 13706 besides its host. These results suggest that KP-1 phage can control K. pneumoniae in vitro and diseases caused by E. coli, two closely related species of the Enterobacteriaceae family and the most predominant in UTIs [4]. The efficacy of phage KP-1 was also tested in this study against other strains of K. pneumoniae and other bacterial genera of the Enterobacteriaceae family. However, the phage infected none of these bacteria. Generally, phages are highly specific, often infecting only one bacterial genus or even specific strains [9,10]. Shah et al. (2023) showed that phage RAM-1 infected only 3 of 16 K. pneumoniae strains tested [29]. Phage vB_kpnM_17-11 also showed a narrow host range, able to lyse only 4 out of 96 strains of K. pneumoniae tested [30]. In another study, phages LASTA and SJM3 infected only 5 K. pneumoniae of the 140 tested [24]. The narrow host range of phage KP-1 may be due to bacterial resistance to phage adsorption, exopolysaccharide production, and/or capsule formation, which have been described for a range of bacteria [31]. For effective phage KP-1 applications, it will probably be necessary to design highly multi-component cocktails and thus extend their action range.
The success of phage therapy is generally attributed to parameters like adsorption rate and burst size [32]. The adsorption profile of phage KP-1 showed that after 10 and 60 min, approximately 60 and 75%, respectively, of the phage particles were adsorbed to the host cells. Similar results have been obtained with phage K2a [33]. However, in general, the adsorption rate of phage KP-1 is lower than in other studies for Klebsiella phages [24,33,34]. Phage LASTA and SJM3 adsorption assays showed that approximately 97 and 94%, respectively, of the phage particles were adsorbed to K. pneumoniae after 20 min [24]. In another study, the adsorption rate of phage HS106 was approximately 84.2% after 6 min [34].
Phage KP-1 has a long latent period (100 min) and a small burst size (7.9 ± 0.3 PFU/host cell). It has been observed that K. pneumoniae phages have a wide range of latent periods and burst sizes, ranging from high burst sizes (410 PFU/infected cell) and short latent periods (21 min) [29] to low burst sizes (31.7 PFU/infected cell) and long latent periods (30 min) [30]. Baqer et al. (2022) demonstrated that K. pneumoniae infecting phages K2a, K2b, K2w5, K2w6, Kp99, K9w5, K9w6, K9coc had burst sizes of 116, 41, 354, 106, 214, 66, 130 and 210 PFU/host cell, respectively, with latency periods of approximately 5, 20, 20, 25, 20, 30, 10 and 10 min, respectively [33]. Phages LASTA and SJM3 presented a higher burst size of 187 ± 37 and 155 ± 34 PFU/host cell, respectively, and a long latent period of 80 min [24]. In another study, phage HS106 presented higher burst size (183 PFU/host cell) and low latent period (10 min) [34]. Although phage KP-1 presents a small burst size (7.9 ± 0.3 PFU/host cell) and a long latency period (100 min) (Figure 3), phages replicate efficiently in the host, causing a high reduction in K. pneumoniae growth, suggesting that other factors regulate the phage–bacteria interaction. A phage’s burst size and latent period depend on the phage type, the host’s physiological state, the growth medium’s composition, the pH and the temperature of the incubation [33]. The smaller burst size can be caused by the phage’s large size and the host‘s small size. The host cell’s size is critical as it modulates the availability of receptors and its protein synthesis machinery for phage binding and growth [35]. Larger burst sizes and longer latent periods increase the likelihood of successful dispersal in the environment [36], indicating that the phage is widely available for isolation.
Klebsiella pneumoniae was effectively inactivated by KP-1 phage in TSB medium (maximum inactivation of 5.4 log CFU/mL after 9 h). The increase in the MOI from 1 to 10 increased treatment efficiency. The maximum inactivation of phage KP-1 at MOI 1 (maximum inactivation of 4.9 log CFU/mL) and 10 (5.4 log CFU/mL) was statistically similar (Figure 4A). However, when an MOI of 10 was used, bacterial regrowth was delayed, and the bacterial concentration remained constant until the end of the experiment. Chen et al. (2023) also observed that lower phage concentrations can induce bacterial regrowth more rapidly than those treated with higher concentrations [34]. The authors observed that bacterial density (OD600) increased more rapidly when incubated at a lower MOI of 1 compared to higher MOIs (10 and 100) [34]. The number of phage particles of KP-1 when incubated with K. pneumoniae for 12 h at an MOI of 1 and 10 increased by 2.7 and 1.9 log PFU/mL, respectively. These results demonstrate that high initial phage doses may not be essential due to the self-perpetuating nature revealed by increasing phage titre and bacteria.
The efficiency of phage KP-1 was tested in human urine samples to evaluate the potential application of this phage for the inactivation of UTI caused by K. pneumoniae. It was observed that phage KP-1 could successfully inactivate K. pneumoniae in urine (maximum inactivation of 3.8 log CFU/mL). The concentration of phage KP-1 remained constant in the absence of the host. Its titer increased significantly in the presence of the bacterium during the experiment. However, its effectiveness in inactivating K. pneumoniae in urine (maximum concentration of 3.8 log CFU/mL after 9 h of incubation) was significantly lower compared to experiments in TSB medium (maximum inactivation of 5.4 log CFU/mL after 9 h of incubation). Although the phage concentration remained constant throughout the experiment, and phage KP-1 replication in the presence of K. pneumoniae was similar in TSB medium and urine (2.9 and 2.7 log PFU/mL, respectively), the lower bacterial inactivation in urine can be due to lower phage viability. Silva et al. (2014) showed that although phages survive at different pH values, their efficiency in inactivating bacteria is affected by low pH values [37]. In general, phage lytic activity decreases at pH values 10 < pH < 5, with optimum pH conditions around neutrality (pH of 6-8) [38,39]. Similar results were obtained by Pereira et al. (2016) [40]. These authors showed that single phage suspensions of phages E-2 and E-4 and the phage cocktail E-2/E-4 reduced approximately 2.0 log CFU/mL of Enterobacter cloacae in urine. The efficacy of single phage suspensions and the phage cocktail was lower than that of phosphate-buffered saline (PBS), which reduced 3.4 log CFU/mL [40]. Further studies using the phage KP-1 in micro- and/or nanocarriers should be done to avoid the negative effect of the urine’s low pH on the phage viability.
A major concern of bacterial inactivation by phages is the emergence of phage-resistant bacteria [41,42,43,44]. The development of resistant mutants, which only occurred in bacteria exposed to the KP-1 phage, was limited (5.55 x 10-3). These results are in close agreement with results obtained by other researchers [44,45,46,47]. Phage cocktails in phage therapy can help overcome the problem of bacterial phage resistance [39]. However, their success requires phages that do not have overlapping cross-resistance, i.e. bacterial mutants resistant to one phage but sensitive to another and vice versa [48], for example, using phages from different families.
4. Materials and Methods
4.1. Bacterial strains and growth conditions
The bacterial strains used in this study are listed in Table 1. The bacterial strain K. pneumoniae Scc 24 was previously isolated [49] and used as a phage host. Escherichia coli (ATCC 13706 and ATCC 25922), Salmonella Typhimurium (ATCC 13311 and ATCC 14028), S. flexneri DSM 4782 were purchased from the ATCC and DSM collections, respectively. Enterobacter cloacae was previously isolated by our group [50]. Escherichia coli (Scc: 9, 33, 34, 35, 36, 37, 40, 41, 43, 45, 47, 48, 49, 51, 52, 53, 55, 56, 58, 69, 77, 78 and 91) and K. pneumoniae (Scc: 1, 5, 11, 15, 17 and 98) were previously isolated [49]. Escherichia coli (BC30, AE11, AD6, AF15, AN19, AC5, AJ23, BN65 and BM62) were isolated in previous works from water samples collected in Ria de Aveiro [51]. Salmonella Enteritidis (CVA, CVB, CVC, CVD and CVE) were provided by the Controlvet Laboratory in previous work[50]. Fresh bacterial cultures were kept at 4 °C in Tryptic Soy Agar (TSA, Liofilchem, Roseto degli Abruzzi, Italy). Before each assay, one isolated colony was transferred to 30 mL of TSB and grown overnight at 37 °C with orbital shaking (120 rpm) until an optical density (O.D. 600) of 0.8, corresponding to about 109 cells/mL.
4.2. Phage isolation and purification
Phage KP-1 was isolated from the sewage network of Aveiro (station EEIS9 of SIMRIA Multi Sanitation System of Ria de Aveiro). About 50 millilitres of water were filtered through 0.45 µm pore size polycarbonate membranes (Millipore, Bedford, MA, USA) and added to 50 mL of a twice-concentrated TSB medium. The mixture was inoculated with 1 mL of K. pneumoniae Scc 24 in exponential phase and incubated for 24 h at 25 °C and 120 rpm. After incubating, the solution was centrifuged at 10.000 × g for 10 min at 4 °C and filtered through a polyethersulfone layer with a 0.22 µm pore size (Merck-Millipore, Darmstadt, Germany). The filtrate was stored at 4 °C, and the titter was determined according to [52]. Serial dilutions of the filtrate stock were performed in phosphate buffer saline (PBS) [137 mmol−1 NaCl (Sigma, St. Louis, MO, USA), 8.1 mmol−1 Na2HPO4·2H2O (Sigma, St. Louis MO, USA), 2.7 mmol−1 KCl (Sigma, St. Louis, MO, USA), and 1.76 mmol−1 KH2PO4 (Sigma, St. Louis, MO, USA), pH 7.4). Five hundred microlitres of each dilution, along with 200 µL of fresh bacterial culture, were added to 5 mL of molten TSB 0.6% top agar layer [30 g/L TSB (Liofilchem, Roseto degli Abruzzi, Italy), 6 g/L agar (Liofilchem, Roseto degli Abruzzi, Italy), 0.12 g/L MgSO4 (Sigma, St. Louis, MO, USA), and 0.05 g/L CaCl2 (Sigma, St. Louis, MO, USA), pH 7.4] and poured over a TSA plate. Plates were incubated at 37 °C and observed for the presence of lytic plaques after 16 to 18 h. One single plaque was selected from the solid medium and added to the TSB medium with a fresh culture of the host. The sample was centrifuged, and the supernatant was used as a phage source for a second isolation procedure. Three successive single-plaque isolation cycles were performed to acquire pure phage stock. All lysates were centrifuged at 10.000× g for 10 min at 4 °C to remove bacteria or bacterial debris. The phage suspensions were kept at 4 °C.
4.3. Electron Microscope Examination
Phage particles of a highly concentrated suspension (109 PFU/mL) were negatively stained with uranyl acetate (Electron Microscopy Sciences). Briefly, 15 µL of each sample were adsorbed to carbon-coated collodion 400 mesh nickel grids (Gilder) for 2 min and stained with 2% aqueous uranyl acetate (Electron Microscopy Sciences) for 1 min. Grids were visualized in a JEOL JEM-1011 transmission electron microscope (operating at 100 kV). Micrographs were taken with a MEGA VIEW III (SIS) digital camera at various magnifications.
4.4. Phage DNA Extraction
The phage suspension (>109 PFU/mL) was centrifuged 3 times at 12.000 x g for 10 min. DNA The extraction of the virion's nucleic acid was performed in duplicate according to Jakočiūnė & Moodley (2018) [53], with bacterial DNA and RNA removal and digestion of the bacteriophage capsid [54] and DNA purification performed with the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany). During DNA purification, twice the volume of phage lysate and equivalent volumes were used to increase the yield. The quantity and quality of the DNA were measured in a NanoDrop One UV-VIS scanning spectral microdroplet spectrophotometer (ThermoScientific, USA), according to the manufacturer's instructions.
4.5. Phage Genome Assembly and Annotation
Phage genome sequencing was performed by Stabvida (Lisboa, Portugal) using Illumina MiSeq using 300bp paired-end. The library construction was done using the Kit Kapa HyperPlus according to manufacturers’ protocol.
The quality control of raw sequence reads were performed using FASTQC v0.11.9 (FastQC source: Bioinformatics Group at the Babraham Institute, UK.) before and after trimming low-quality reads with Trimmomatic v.0.39 with the parameters: ILLUMINACLIP:adaptors.fasta:2:30:1, Leading:8, Trailing:8, Slidingwindow:4:15 and Minlen:100 [55]. The trimmed reads were subjected to de novo genome assembly, using SPAdes v.3.13.1 with the –careful parameter [56]. The assembly graph was inspected using Bandage v0.8.1 [57]. The reads were then mapped back against the resulting assembly using BBMap v.38.18 to determine the average coverage of each contig [58]. Manual filtering was performed to remove contigs with dubious coverage. The reads were map back to the filtering contig file using Bowtie2 v.2.5.1 [59] and assembly with SPAdes v.3.13.1. Further assembly error correction and polishing were performed using Pilon v1.24 [60]. Termini could not be predicted using PhageTerm [61], however, the genome was found to be circularly permuted using apc.pl (https://github.com/jfass/apc), and the repeated sequence artifacts were removed. The phage genome was manually reordered to match the most closely related phage based on the method reported by Shen and Millard [62]. The full-genome sequence of the KP-1 phage was compared to the phage genome sequences in GenBank using BLASTN (somewhat similar sequences) in the NCBI database, and the most closely related phages were identified. The completeness and contamination of the phage genome sequence was tested using CheckV v1.0.1 [63]. The phage genome was annotated for coding DNA sequences (CDS), tRNA, tmRNA, CRISPRs, virulence factors (VFs), toxins and antimicrobial resistance genes (ARGs) using Pharokka v1.3.2 [64], and the CDS were assigned to functional categories using PHROGs [65].
4.6. Phage host range and efficiency of plating
The phage host range was assessed through spot testing according to [52] for the bacterial strains in Table 1. Briefly, molten top agar 0.6% agar was inoculated with 300 µL of fresh exponential bacteria culture, poured over a TSA plate and allowed to dry. Subsequently, 50 to 100 µL of phage stock was spotted over the bacterial lawn and plates were incubated at 37 °C for 16 to 18 h before observation. A clear lysis zone on the area where the phage had been spotted would indicate sensitivity to the phage. Bacterial sensitivity to phage infection was recorded according to a clear lysis zone (x) or absence of lysis zone (-) (Table 1).
4.7. Adsorption curve
The phage adsorption curve was determined according to [52,66]. Briefly, phages (103 PFU/mL) were added to a mid-exponential bacterial culture (cell density of about 106 CFU/mL) at 37 °C in TSB. Every 10 min, an aliquot was collected, a few drops of chloroform were added, and the sample was centrifuged at 10.000 × g for 5 min at 4 °C. The number of unadsorbed phage particles was determined by plating the sample using the double agar layer technique [52]. The percentage of adsorbed phages was calculated by comparing the free phage titre at a given point with the initial phage titre. Three independent assays were performed.
4.8. One-step growth assay
Phage suspension (final concentration of 106 PFU/mL) was added to 10 mL of fresh bacterial culture of K. pneumoniae (with a final concentration of 109 PFU/mL) to obtain a final MOI of 0.001 and incubated for 5 min at 37 °C without shaking. The suspension was then centrifuged at 10.000× g for 5 min, the supernatant was discarded, and the pellet was resuspended in 10 mL of TSB and incubated at 37 °C. A 1 mL sample was collected and immediately tittered by the double-layer agar technique. Plates were incubated at 25 °C and observed after 18 h. Samples were collected every 20 min for 140 min and three independent assays were performed.
4.9. Bacterial kill curve in TSB culture media
To characterize the bacterial kill curves, 30 mL of sterile TSB was inoculated with the strain K. pneumoniae Scc 24, which was then challenged with phage Kp-1 at MOIs of 1 and 10 (final concentration 105 and 106 PFU/mL, respectively). Two controls were included in this assay: the bacterial control (BC) and the phage control (PC). The bacterial control was inoculated with K. pneumoniae but not with phage, and the phage control was inoculated only with the phage. Controls and test samples were kept at the same conditions (37 °C and no agitation). Aliquots were collected from all controls and samples at 0, 3, 6, 9, 12 and 24 h of incubation. Phage titre was determined in duplicate by the double agar layer [52] after an incubation period of 16-18 h at 37 °C. Bacterial concentration was determined in duplicate in the TSA medium after an incubation period of 24 h at 37 °C. Three independent assays were performed for each MOI.
4.10. Bacterial kill curve in urine samples.
4.10.1. Urine sample collection and handling.
According to Pereira and coworkers [40], early urine samples were collected using a post-hygiene, midstream clean-catch technique. After performing daily hygiene, the micturition middle stream was collected directly into sterile containers after discarding the initial portion. The end portions were also discarded. The samples were immediately transported to the laboratory, centrifuged at 10.000 x g for 10 min and filtered through a 0.22 µm pore size (Merck-Millipore, Darmstadt, Germany) into sterile containers to remove bacteria eventually present in the urine samples. The urine samples were always collected from the same person at the same hours.
4.10.2. Bacterial kill curves in urine
The efficacy of the phage KP-1 to control K. pneumoniae was assessed in the sterile urine for an MOI of 10 to evaluate the possible application of this phage during UTI. An Erlenmeyer flask containing 30 mL of sterile urine was inoculated with bacteria (final concentration of 105 CFU/mL) and phage (final concentration of 106 PFU/mL). Two other flasks were included as control groups. The phage control group was only inoculated with phage (PC), and the bacteria control group was only inoculated with bacteria without the phage (BC). The flasks were incubated at 37 °C with no agitation, and aliquots were collected after 0, 3, 6, 9, 12 and 24 h. Bacterial concentration was determined, in duplicate, in the TSA plate after an incubation period of 24 h. In duplicate, the double agar layer method [52] determined phage titre after an incubation period of 24 h at 37 °C. Three independent assays were performed.
4.11. Rate of emergence of phage-resistant bacterial mutant
The emergence rate of phage-resistant bacterial mutants was determined according to Filippov et al., 2011. Ten phage-sensitive colonies were isolated from a TSA plate and incubated at 37 °C in test tubes containing 5 mL of TSB medium for 24 h (final concentration of about 109 CFU/mL). Each bacterial culture was serially diluted, and aliquots of 100 µL from the 100 to 10-2 dilutions were transferred to molten top TSB 0.6% agar, previously inoculated with 100 µL of phage (from a phage stock with a final titre of about 109 PFU/mL) and poured over a TSA plate. The plates were incubated at 37 °C for 3-5 days since some mutants grow slowly. In parallel, an aliquot of 100 µL of the 10-5 to 10-7 dilutions was incorporated on TSA plates without the addition of phage and incubated for 24 h at 37 °C. The frequency of spontaneous phage-resistant mutation was calculated by dividing the number of resistant bacteria (colonies counted in the presence of phage) by the total number of sensitive bacteria (CFU counted in the plates without phage) and multiplied by 100. For this assay, three independent experiments were performed.
4.12. Statistical analysis.
The statistical analysis was performed using GraphPad Prism software 8.4.3, San Diego, California, USA. Normal distribution of the data was assessed by the Kolmogorov–Smirnov test, and the homogeneity of variance was confirmed by Levene’s test. Two-way ANOVA with repeated measures and Tukey’s multiple comparison post-hoc test was used to study if the variations between bacterial and viral concentrations presented any significant differences (sections 2.9 and 2.10.2). Two-way ANOVA was used to infer if the differences between the growth curve of the test groups and the control groups were significant (sections 2.9 and 2.10.2). A p-value below 0.05 was considered statistically significant.
5. Conclusions
The efficacy of phage KP-1 and its safety paves the way for in-depth studies, particularly in vivo, to combat urinary tract infections caused by K. pneumoniae. However, due to the urine’s low pH, which affects phage viability, phages should be applied in micro- and/or nanocarriers. In the future, new phages need to be isolated and tested with phage KP-1 to produce a cocktail with a broader spectrum of activity against K. pneumoniae and even to control other UTI-causing bacteria.
Supplementary Materials
Table S1. Coding sequences identified in KP-1 phage.
Author Contributions
J.D, C.M and P.C performed the experiments. J.D, C.M, P.C, J.R, C.P and A.A participated in the conception and design of the experiments, J.D, C.P, V.O, N.C.M.G., J.L.R., C.P and A.A participated in analyzing the results, J.D and C.P wrote the paper, and P.C, V.O and N.C.M.G. also contributed to the writing. J.L.R. and A.A. supervised the work, revised the paper, and contributed with reagents and analysis tools. All authors have read and agreed to the published version of the manuscript.
Funding
Through national funds, we acknowledge financial support to CESAM by FCT/MCTES (UIDP/50017/2020+UIDB/50017/2020+ LA/P/0094/2020).
Data Availability Statement
Not applicable.
Acknowledgements
Thanks to the Department of Biology and the University of Aveiro, where this research was carried out. The authors are also grateful to CESAM and its funding sources. João Duarte and Pedro Costa thank the Portuguese Foundation for Science and Technology (FCT) for their doctoral grant (2021.05519.BD and PD/BD/150360/2019, respectively). Carla Pereira acknowledges the FCT for Junior Research contract (CEEC Individual/03974/2017). Vanessa Oliveira was funded by National funds (OE), through FCT, IP., in the scope of the framework contract foreseen in the numbers 4, 5 and 6 of the article 23, of the Decree-Law 57/2016, of August 29, changed by Law 57/2017, of July 19.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization (WHO) Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis; World Heal.; World Health Organization, 2017; Vol. 13;
- Lee, Y.-L.; Ko, W.-C.; Hsueh, P.-R. Geographic patterns of global isolates of carbapenem-resistant Klebsiella pneumoniae and the activity of ceftazidime/avibactam, meropenem/vaborbactam, and comparators against these isolates: Results from the Antimicrobial Testing Leadership and Surveillan. Int. J. Antimicrob. Agents 2022, 60, 106679. [Google Scholar] [CrossRef]
- Gorrie, C.L.; Mirčeta, M.; Wick, R.R.; Judd, L.M.; Lam, M.M.C.; Gomi, R.; Abbott, I.J.; Thomson, N.R.; Strugnell, R.A.; Pratt, N.F.; et al. Genomic dissection of Klebsiella pneumoniae infections in hospital patients reveals insights into an opportunistic pathogen. Nat. Commun. 2022, 13, 3017. [Google Scholar] [CrossRef]
- European Center for Disease Prevention and Control and World Health Organization (WHO) Annual epidemiological report for 2019: Healthcare - associated infections acquired in intensive care units; Stockholm, 2023.
- Karampatakis, T.; Tsergouli, K.; Behzadi, P. Carbapenem-Resistant Klebsiella pneumoniae: Virulence Factors, Molecular Epidemiology and Latest Updates in Treatment Options. Antibiotics 2023, 12, 234. [Google Scholar] [CrossRef]
- Guerra, M.E.S.; Destro, G.; Vieira, B.; Lima, A.S.; Ferraz, L.F.C.; Hakansson, A.P.; Darrieux, M.; Converso, T.R. Klebsiella pneumoniae Biofilms and Their Role in Disease Pathogenesis. Front. Cell. Infect. Microbiol. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- European Center for Disease Prevention and Control and World Health Organization (WHO) Antimicrobial resistance surveillance in Europe; 2023.
- Abbo, L.; Hooton, T. Antimicrobial Stewardship and Urinary Tract Infections. Antibiotics 2014, 3, 174–192. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hong, Q.; Chang, R.Y.K.; Kwok, P.C.L.; Chan, H.-K. Phage–Antibiotic Therapy as a Promising Strategy to Combat Multidrug-Resistant Infections and to Enhance Antimicrobial Efficiency. Antibiotics 2022, 11, 570. [Google Scholar] [CrossRef]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med. Virol. 2019, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tandogdu, Z.; Cai, T.; Koves, B.; Wagenlehner, F.; Bjerklund-Johansen, T.E. Urinary Tract Infections in Immunocompromised Patients with Diabetes, Chronic Kidney Disease, and Kidney Transplant. Eur. Urol. Focus 2016, 2, 394–399. [Google Scholar] [CrossRef]
- European Center for Disease Prevention and Control Annual Epidemiological Report for 2018: Healthcare- associated infections acquired in intensive care unitsHealthcare-associated infections acquired in intensive care units; 2018.
- Al-Anany, A.M.; Hooey, P.B.; Cook, J.D.; Burrows, L.L.; Martyniuk, J.; Hynes, A.P.; German, G.J. Phage Therapy in the Management of Urinary Tract Infections: A Comprehensive Systematic Review. PHAGE 2023, 4, 112–127. [Google Scholar] [CrossRef]
- Sybesma, W.; Zbinden, R.; Chanishvili, N.; Kutateladze, M.; Chkhotua, A.; Ujmajuridze, A.; Mehnert, U.; Kessler, T.M. Bacteriophages as Potential Treatment for Urinary Tract Infections. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Leitner, L.; Mehnert, U.; Chkhotua, A.; Kessler, T.M.; Sybesma, W. Adapted Bacteriophages for Treating Urinary Tract Infections. Front. Microbiol. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Le, T.; Nang, S.C.; Zhao, J.; Yu, H.H.; Li, J.; Gill, J.J.; Liu, M.; Aslam, S. Therapeutic Potential of Intravenous Phage as Standalone Therapy for Recurrent Drug-Resistant Urinary Tract Infections. Antimicrob. Agents Chemother. 2023, 67. [Google Scholar] [CrossRef] [PubMed]
- Laforêt, F.; Antoine, C.; Blasdel Reuter, B.; Detilleux, J.; Pirnay, J.-P.; Brisse, S.; Fall, A.; Duprez, J.-N.; Delcenserie, V.; Thiry, D. In Vitro and In Vivo Assessments of Two Newly Isolated Bacteriophages against an ST13 Urinary Tract Infection Klebsiella pneumoniae. Viruses 2022, 14, 1079. [Google Scholar] [CrossRef] [PubMed]
- Zurabov, F.; Glazunov, E.; Kochetova, T.; Uskevich, V.; Popova, V. Bacteriophages with depolymerase activity in the control of antibiotic resistant Klebsiella pneumoniae biofilms. Sci. Rep. 2023, 13, 15188. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Du, F.; Long, M.; Li, P. Limitations of Phage Therapy and Corresponding Optimization Strategies: A Review. Molecules 2022, 27, 1857. [Google Scholar] [CrossRef] [PubMed]
- Patient, S.; Zhang, B. Comparison of two distinct subpopulations of Klebsiella pneumoniae ST16 co-occurring in a single patient. Microbiol. Spectr. 2022, 10, e0262421. [Google Scholar]
- Cai, R.; Wang, Z.; Wang, G.; Zhang, H.; Cheng, M.; Guo, Z.; Ji, Y.; Xi, H.; Wang, X.; Xue, Y.; et al. Biological properties and genomics analysis of vB_KpnS_GH-K3, a Klebsiella phage with a putative depolymerase-like protein. Virus Genes 2019, 55, 696–706. [Google Scholar] [CrossRef]
- Majkowska-Skrobek, G.; Latka, A.; Berisio, R.; Squeglia, F.; Maciejewska, B.; Briers, Y.; Drulis-Kawa, Z. Phage-borne depolymerases decrease Klebsiella pneumoniae resistance to innate defense mechanisms. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Obradović, M.; Malešević, M.; Di Luca, M.; Kekić, D.; Gajić, I.; McAuliffe, O.; Neve, H.; Stanisavljević, N.; Vukotić, G.; Kojić, M. Isolation, characterization, genome analysis and host resistance development of two novel lastavirus phages active against pandrug-resistant Klebsiella pneumoniae. Viruses 2023, 15, 628. [Google Scholar] [CrossRef]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Alves, D.R. Phage therapy in the 21st century: Is there modern, clinical evidence of phage-mediated efficacy? Pharmaceuticals 2021, 14, 1–25. [Google Scholar] [CrossRef]
- Suh, G.A.; Lodise, T.P.; Tamma, P.D.; Knisely, J.M.; Alexander, J.; Aslam, S.; Barton, K.D.; Bizzell, E.; Totten, K.M.C.; Campbell, J.L.; et al. Considerations for the Use of Phage Therapy in Clinical Practice. Antimicrob. Agents Chemother. 2022, 66, e0207121. [Google Scholar] [CrossRef]
- Peng, Q.; Fang, M.; Liu, X.; Zhang, C.; Liu, Y.; Yuan, Y. Isolation and Characterization of a Novel Phage for Controlling Multidrug-Resistant Klebsiella pneumoniae. Microorganisms 2020, 8, 542. [Google Scholar] [CrossRef]
- Shah, M.; Bukhari, S.; Redaina; Adnan, M.; Imran, M.; Jamal, M. Isolation and characterization of lytic bacteriophage from waste water to control clinical multidrug resistant Klebsiella pneumoniae. Kuwait J. Sci. 2023, 50, 681–689. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, F.; Liang, S.; Chen, Q.; Wang, W.; Wang, Y.; Mart, A.J. Isolation and characterization of vB _ kpnM _ 17-11, a novel pnhage ffficcient against carbapenem- resistant Klebsiella pneumoniae. 2022, 12, 1–13. [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, H.; McKean, K.; Wang, I. Phage fitness may help predict phage therapy efficacy. Bacteriophage 2014, 4, e964081. [Google Scholar] [CrossRef] [PubMed]
- Baqer, A.; Fang, K.; Mohd-Assaad, N.; Adnan, S.; Nor, N. In Vitro Activity, Stability and Molecular Characterization of Eight Potent Bacteriophages Infecting Carbapenem-Resistant Klebsiella pneumoniae. Viruse 2023, 15, 117. [Google Scholar] [CrossRef]
- Chen, C.; Li, T.; Chen, H. Isolation and characterization of novel bacteriophage vB _ KpP _ HS106 for Klebsiella pneumonia K2 and applications in foods. Front. Microbiol. 2023, 14, 1227147. [Google Scholar] [CrossRef]
- Choi, C.; Kuatsjah, E.; Wu, E.; Yuan, S. The Effect of Cell Size on the Burst Size of T4 Bacteriophage Infections of Escherichia coli B23. J. Exp. Microbiol. Immunol. (JEMI 2010, 14, 85–91. [Google Scholar]
- Droubogiannis, S.; Katharios, P. Genomic and Biological Profile of a Novel Bacteriophage, Vibrio phage Virtus, Which Improves Survival of Sparus aurata Larvae Challenged with Vibrio harveyi. Pathogens 2022, 11, 630. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.J.; Costa, L.; Pereira, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Almeida, A. Influence of environmental variables in the efficiency of phage therapy in aquaculture. Microb. Biotechnol. 2014, 7, 401–413. [Google Scholar] [CrossRef]
- Nakai, T.; Sugimoto, R.; Park, K.H.; Matsuoka, S.; Mori, K.; Nishioka, T.; Maruyama, K. Protective effects of bacteriophage on experimental Lactococcus garvieae infection in yellowtail. Dis. Aquat. Organ. 1999, 37, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Duarte, J.; Costa, P.; Braz, M.; Almeida, A. Bacteriophages in the control of Aeromonas sp. in aquaculture systems: An integrative view. Antibiotics 2022, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Pereira, C.; Santos, L.; Klumpp, J.; Almeida, A. Potential of phage cocktails in the inactivation of Enterobacter cloacae — An in vitro study in a buffer solution and in urine samples. Virus Res. 2016, 211, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.; Moutinho, C.; Pinto, F.; Del Fiol, F.; Jozala, A.; Chaud, M.; Vila, M.; Teixeira, J.; Balcão, V. Alternatives to overcoming bacterial resistances: State-of-the-art. Microbiol. Res. 2016, 191, 51–80. [Google Scholar] [CrossRef]
- Lima, R.; Del Fiol, F.S.; Balcão, V.M. Prospects for the use of new technologies to combat multidrug-resistant bacteria. Front. Pharmacol. 2019, 10, 692. [Google Scholar] [CrossRef] [PubMed]
- Filippov, A.; Sergueev, K. V.; He, Y.; Huang, X.Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS One 2011, 6, 1–11. [Google Scholar] [CrossRef]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeidab, P.; Clemente, C.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Characterization and in vitro evaluation of new bacteriophages for the biocontrol of Escherichia coli. Virus Res. 2016, 227, 171–182. [Google Scholar] [CrossRef]
- Wright, R.C. .; Friman, V.-P.; Smith, M.C.M.; Brockhurst, M.A.; Cross-resistance is modular in bacteria–phage interactions. PLoS Biol. 2018, 16, e2006057. [Google Scholar] [CrossRef]
- Pinheiro, L.; Pereira, C.; Frazão, C.; Balcão, V.; Almeida, A. Efficiency of phage φ6 for biocontrol of Pseudomonas syringae pv. syringae: An in vitro preliminary study. Microorganisms 2019, 7, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Pereira, C.; Costa, P.; Almeida, A. Bacteriophages with potential to inactivate Aeromonas hydrophila in cockles : In vitro and in vivo preliminary studies. Antibiotics 2021, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Culot, A.; Grosset, N.; Gautier, M. Overcoming the challenges of phage therapy for industrial aquaculture: A review. Aquaculture 2019, 513, 734423. [Google Scholar] [CrossRef]
- Silva, I.; Tacão, M.; Tavares, R.D.S.; Miranda, R.; Araújo, S.; Manaia, C.M.; Henriques, I. Fate of cefotaxime-resistant Enterobacteriaceae and ESBL-producers over a full-scale wastewater treatment process with UV disinfection. Sci. Total Environ. 2018, 639, 1028–1037. [Google Scholar] [CrossRef]
- Pereira, S.R. do S. Viruses as therapeutic and infectious agents.Thesis MsC, University of Aveiro, Portugal, 2014.
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Cunha, Â.; Delgadillo, I.; Romalde, J.; Nunes, M.L.; Almeida, A. Bacteriophages with potential to inactivate Salmonella Typhimurium: Use of single phage suspensions and phage cocktails. Virus Res. 2016, 220, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophages; John Wiley and Sons Inc: New York, 1959. [Google Scholar]
- Jakočiūnė, D.; Moodley, A. A Rapid Bacteriophage DNA Extraction Method. Methods Protoc. 2018, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; ISBN 9780879695774. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge – Accurate paired shotgun read merging via overlap. PLoS One 2017, 12, 1–15. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Millard, A. Phage genome annotation: Where to begin and end. PHAGE Ther. Appl. Res. 2021, 2, 183–193. [Google Scholar] [CrossRef]
- Nayfach, S.; Camargo, A.P.; Schulz, F.; Eloe-Fadrosh, E.; Roux, S.; Kyrpides, N.C. CheckV assesses the quality and completeness of metagenome-assembled viral genomes. Nat. Biotechnol. 2021, 39, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Bouras, G.; Nepal, R.; Houtak, G.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Pharokka: a fast scalable bacteriophage annotation tool. Bioinformatics 2023, 39, 1–4. [Google Scholar] [CrossRef]
- Terzian, P.; Ndela, E.O.; Galiez, C.; Lossouarn, J.; Bucio, R.E.P.; Mom, R.; Toussaint, A.; Petit, M.-A.; Enault, F. PHROG: families of prokaryotic virus proteins clustered using remote homology. NAR Genomics Bioinforma. 2021, 3, lqab067. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. In Bacteriophages: methods and protocols, volume 1: isolation, characterization, and interactions; 2009; Vol. 501 ISBN 9781603271646. [CrossRef]
Figure 1.
Phage plaque morphology (A) and electron micrograph photography (B).
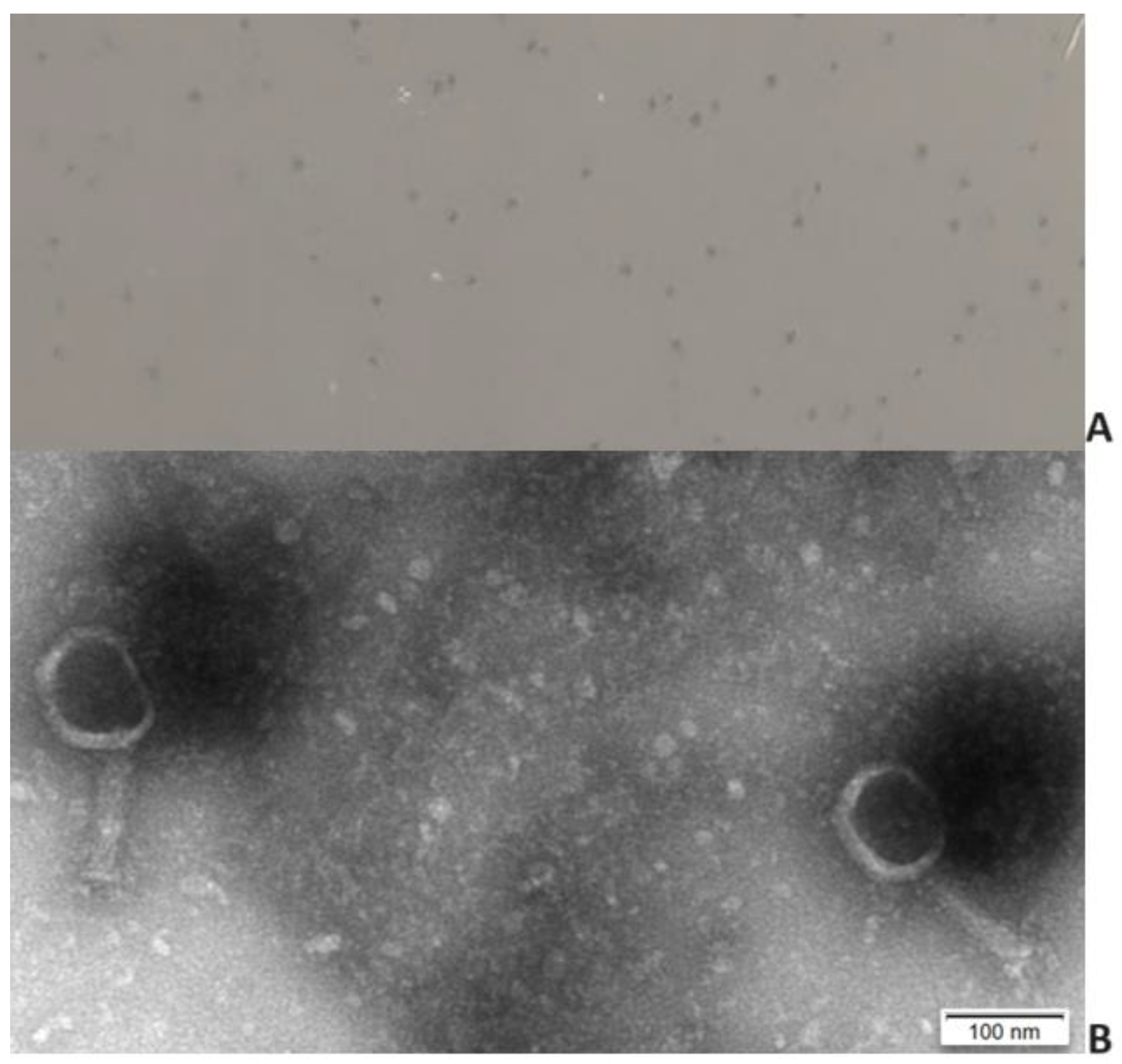
Figure 3.
One-step growth curve of phage KP-1 in the presence of the host, K. pneumoniae. Values represent the mean of three independent experiments, and error bars represent the standard deviation.
Figure 3.
One-step growth curve of phage KP-1 in the presence of the host, K. pneumoniae. Values represent the mean of three independent experiments, and error bars represent the standard deviation.
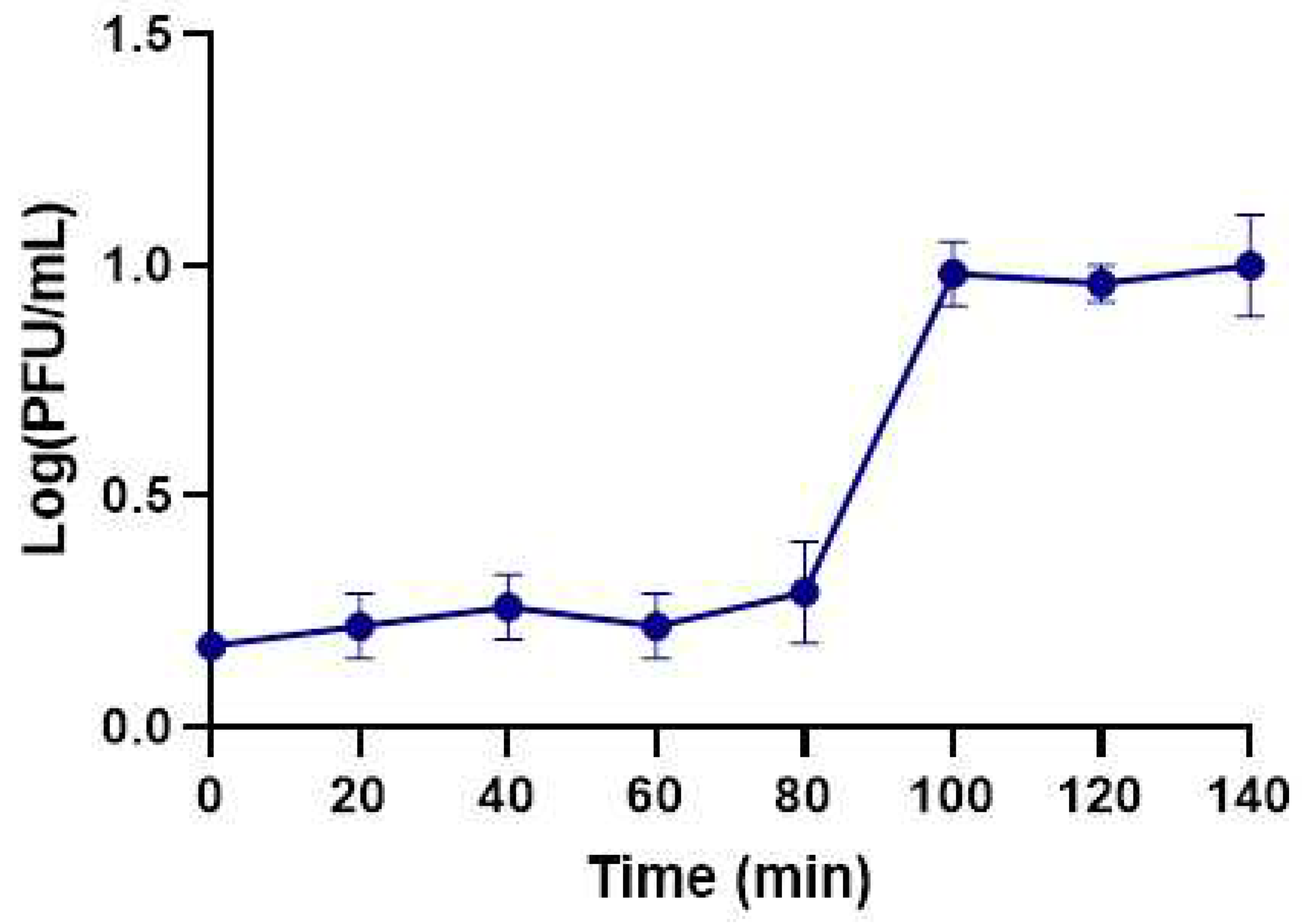
Figure 4.
Inactivation of K. pneumoniae using phage KP-1 at two different MOIs. (A) Bacterial concentrations: BC – Bacterial control; BP MOI 1 – Bacteria plus phage at a MOI of 1; BP MOI 10 – Bacteria plus phage at a MOI of 10. (B) Phage concentration: PC MOI 1 – phage control at a MOI of 1; PC MOI 10: phage control at a MOI of 10; PB MOI 1 - Phage and bacteria at a MOI of 1; PB MOI 10 – Phage and bacteria at a MOI of 10. The experiments were performed in TSB at 37 °C, and values represent the mean of three independent experiments. Error bars represent the standard deviation.
Figure 4.
Inactivation of K. pneumoniae using phage KP-1 at two different MOIs. (A) Bacterial concentrations: BC – Bacterial control; BP MOI 1 – Bacteria plus phage at a MOI of 1; BP MOI 10 – Bacteria plus phage at a MOI of 10. (B) Phage concentration: PC MOI 1 – phage control at a MOI of 1; PC MOI 10: phage control at a MOI of 10; PB MOI 1 - Phage and bacteria at a MOI of 1; PB MOI 10 – Phage and bacteria at a MOI of 10. The experiments were performed in TSB at 37 °C, and values represent the mean of three independent experiments. Error bars represent the standard deviation.
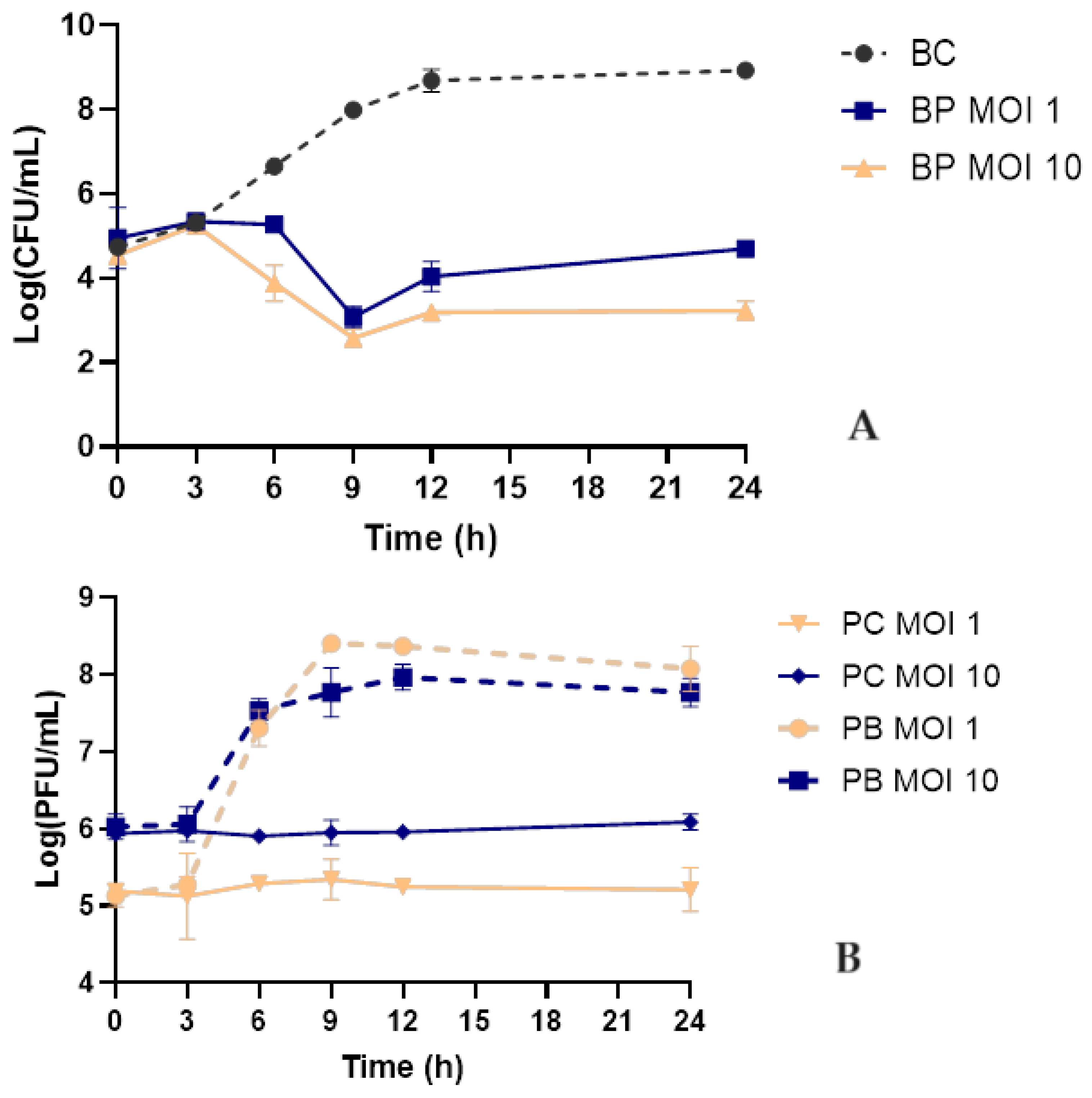
Figure 5.
Inactivation of K. pneumoniae in sterile urine by phage KP-1 using an MOI of 10. (A): bacterial concentrations; Bacteria control - bacteria in the absence of phage; BP MOI 10 - bacteria plus phage at an MOI of 10. (B): Phage concentration; PC MOI 10 – phage control; PB MOI 10 – phage in the presence of bacteria at a final MOI of 10. Values represent the mean of three independent experiments. Error bars represent the standard deviation.
Figure 5.
Inactivation of K. pneumoniae in sterile urine by phage KP-1 using an MOI of 10. (A): bacterial concentrations; Bacteria control - bacteria in the absence of phage; BP MOI 10 - bacteria plus phage at an MOI of 10. (B): Phage concentration; PC MOI 10 – phage control; PB MOI 10 – phage in the presence of bacteria at a final MOI of 10. Values represent the mean of three independent experiments. Error bars represent the standard deviation.
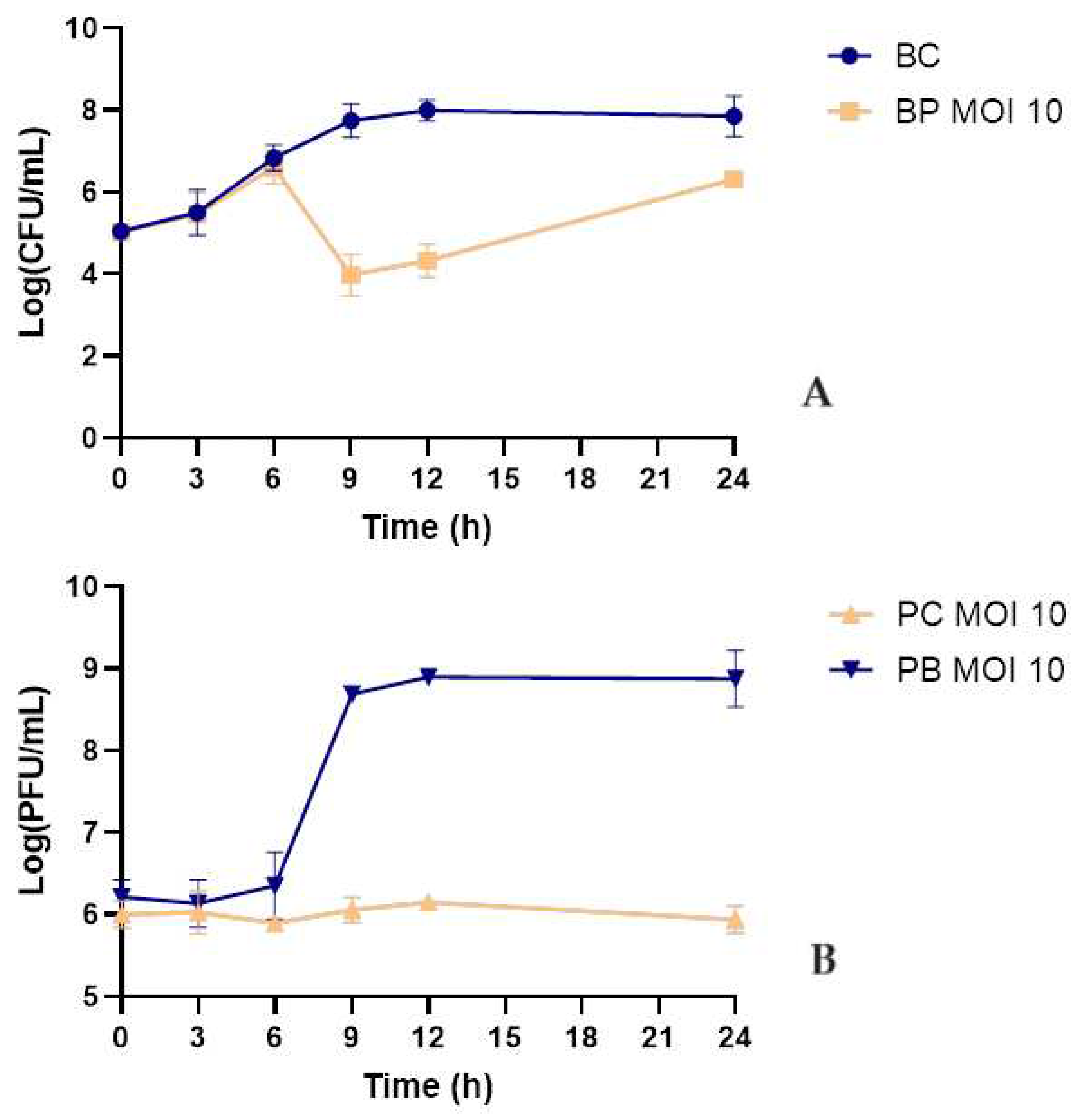

Table 1.
Phage host range and efficiency of plating on 52 different bacterial strains.
| Bacterial strains | Phage KP-1 | |
|---|---|---|
| Spot test | EOP (%) | |
| Klebsiella pneumoniae Scc 24 | + | 100 |
| Klebsiella pneumoniae Scc 1 | - | 0 |
| Klebsiella pneumoniae Scc 5 | - | 0 |
| Klebsiella pneumoniae Scc 11 | - | 0 |
| Klebsiella pneumoniae Scc 15 | - | 0 |
| Klebsiella pneumoniae Scc 17 | - | 0 |
| Escherichia coli ATCC 13706 | + | 32.98 ± 2.85 |
| Escherichia coli ATCC 25922 | - | 0 |
| Escherichia coli Scc 9 | - | 0 |
| Escherichia coli Scc 33 | - | 0 |
| Escherichia coli Scc 34 | - | 0 |
| Escherichia coli Scc 35 | - | 0 |
| Escherichia coli Scc 36 | - | 0 |
| Escherichia coli Scc 37 | - | 0 |
| Escherichia coli Scc 39 | - | 0 |
| Escherichia coli Scc 40 | - | 0 |
| Escherichia coli Scc 41 | - | 0 |
| Escherichia coli Scc 43 | - | 0 |
| Escherichia coli Scc 45 | - | 0 |
| Escherichia coli Scc 47 | - | 0 |
| Escherichia coli Scc 48 | - | 0 |
| Escherichia coli Scc 49 | - | 0 |
| Escherichia coli Scc 51 | - | 0 |
| Escherichia coli Scc 52 | - | 0 |
| Escherichia coli Scc 53 | - | 0 |
| Escherichia coli Scc 55 | - | 0 |
| Escherichia coli Scc 56 | - | 0 |
| Escherichia coli Scc 58 | - | 0 |
| Escherichia coli Scc 69 | - | 0 |
| Escherichia coli Scc 77 | - | 0 |
| Escherichia coli Scc 78 | - | 0 |
| Escherichia coli Scc 91 | - | 0 |
| Escherichia coli Scc 98 | - | 0 |
| Escherichia coli BC30 | - | 0 |
| Escherichia coli AE11 | - | 0 |
| Escherichia coli AD6 | - | 0 |
| Escherichia coli AF15 | - | 0 |
| Escherichia coli AN19 | - | 0 |
| Escherichia coli AC5 | - | 0 |
| Escherichia coli AJ23 | - | 0 |
| Escherichia coli BN65 | - | 0 |
| Escherichia coli BM62 | - | 0 |
| Citrobacter freundii 6F | - | 0 |
| Enterobacter cloacae | - | 0 |
| Providencia sp. | - | 0 |
| Salmonella enterica serovar Enteriditis CVA | - | 0 |
| Salmonella enterica serovar Enteriditis CVB | - | 0 |
| Salmonella enterica serovar Enteriditis CVC | - | 0 |
| Salmonella enterica serovar Enteriditis CVD | - | 0 |
| Salmonella enterica serovar Enteriditis CVE | - | 0 |
| Salmonella enterica serovar Typhimurium ATCC 14028 | - | 0 |
| Salmonella enterica serovar Typhimurium ATCC 13311 | - | 0 |
| Shigella flexneri DSM 4782 | - | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



