
Submitted:
04 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
This review focuses on the role of small extracellular vesicles in the pathophysiological mechanisms of retinal degenerative diseases. Many of these mechanisms are related or modulated by the oxidative burden of the retinal cells. It has been recently demonstrated that cellular communication in the retina involves extracellular vesicles and that their rate of release and cargo features might be affected by the cellular environment and in some instances also mediated by autophagy. The fate of these vesicles is diverse, and could end up in the circulation and be used as markers, or target neighbour cells modulating gene and protein expression and eventually also angiogenesis. Neovascularization in the retina promotes vision loss in diseases such as diabetic retinopathy and age-related macular degeneration. The importance of micro RNAs, either as small extracellular vesicles’ cargo or free circulating, on the regulation of retinal angiogenesis is also discussed.
Keywords:
retina
; exosomes
; oxidative stress
; autophagy
; microrna
1. Introduction
‘The retina is the brain’s window on the world’, as it appears in the magnificent neural sciences compendium of Kandel and others [1], and as such it is regarded as part of the central nervous system. The retina has several cell types, among them there are two types of photoreceptors: cones and rods, distributed asymmetrically: cones mostly in the center (macula lutea) and rods mostly in the periphery. Neurons that form the optic nerve, retinal ganglion cells, are also in the retina together with other nerve cell types: bipolar cells, horizontal cells, amacrine cells and Müller cells [2]. Since half of the cerebral cortex is used to process the signals arriving from the approximately one million fibers of the optic nerve, the visual information coming from the retina is a highly valued capacity, thus visual impairment and blindness are among the most feared disabilities of mankind. The retinal pigment epithelium (RPE) plays an important role in maintaining the function and the integrity of the retina and choroid [3], it absorbs light and protects against photooxidation [4]. RPE is part of the blood-retinal barrier, and it regulates the entrance of nutrients into the retina. In fact, RPE transports nutrients, ions, water, and metabolic-end products from one side of the retina to the other [5]. RPE is critical for phototransduction, is also responsible of phagocyting the tips of photoreceptors’ outer segments, and secretes a number of growth factors essential for the structural integrity of the retina and the choroid. Thus, it has been repeatedly involved in the pathophysiology of major retinal diseases such as age-related macular degeneration (AMD) [6] and diabetic retinopathy (DR) [7].
Photoreceptors and retinal ganglion cells are particularly vulnerable to oxidative burden due to a high oxygen environment, glucose oxidation and lipid polyunsaturated fat content, coupled with photo-transduction, leading to increased reactive oxygen species (ROS) production. It has been observed that ROS imbalance is involved in many retinal diseases [8,9,10,11]. Among the primary reactive species are superoxide anion, singlet oxygen, hydroxyl radical, hydrogen peroxide, peroxynitrite or nitric oxide. Furthermore, the oxidation of polyunsaturated fatty acids can lead to the formation of aldehydes such as malondialdehyde or 4-hydroxy-alkenals which have been described as fingerprints of oxidative stress, and the latter also having signaling properties [12].
Our interest on the retinal oxidative pathophysiology started in the early 90s, when two of us (M.D.-L. and F.J.R.) joined efforts to describe a very relevant part of the antioxidant system of the retina in human donors: the glutathione system [13]. Although diseases affecting the anterior segment of the eye are the primary causes of vision impairment and blindness globally (cataract and glaucoma), DR and AMD are also among those, especially in elderly population and developed countries. In these nations, retinitis pigmentosa (RP), a group of inherited forms of retinal degeneration, is the prevalent cause of blindness in the working age population [14]. Oxidative stress has been implicated in the pathogenesis of RP and antioxidants clearly delay RP progression, at least in experimental models [9].
The need to maintain retinal homeostasis for an adequate vision and the indisputable role that redox equilibrium plays in the pathophysiological mechanisms in so many retinal diseases, have showed a constantly growing interest in the last decades. Our aim in this manuscript is to review the pathophysiological mechanisms related or regulated by oxidative burden in retinal diseases, with specific emphasis on the role of angiogenesis and extracellular vesicles and their cargo.
2. Oxidative Pathophysiological Mechanisms
As mentioned above, this review focuses on pathophysiological mechanisms that have been clearly related or are regulated or affected by oxidation, where antioxidants, by different mechanisms, may exert beneficial effects. The angiogenesis mechanism as part of the pathophysiology of vision loss in retinal diseases has been already introduced above. The need of cellular communication for this process, and certainly others, led us to study in detail the phenomena involved in it. In multicellular organisms, cells communicate by extracellular signaling molecules such as nucleotides, lipids, and proteins. These molecules, after being released into the extracellular medium, bind to receptors of their neighboring cells, inducing intracellular signaling and modifying the physiological state of the recipient cell. In addition to the release of these signaling molecules, eukaryotic cells are able to release vesicles to the extracellular medium that contain different types of molecules, among which micro RNA (miRNA) sequences are also present and show regulatory actions in different retinal cells.
2.1. Extracellular Vesicles
Extracellular vesicles (EVs) contain numerous proteins, lipids, and nucleic acids that might affect the fate of the target cells [15]. It has been observed that EVs are involved in cell communication, transporting information (proteins, mRNA, miRNAs, and even DNA fragments) between cells [16]. EVs are thus involved in processes such as immune suppression of tumor metastasis, angiogenesis and autophagy [17,18,19,20,21]. The environment surrounding cells can promote the release of these EVs [22,23,24,25]. EVs can be sub-micro-sized, known as microparticles, or nanometric-sized, previously known as exosomes and actually named as small extracellular vesicles (sEVs) [26,27,28]. Normally, sEVs can include several molecules in their cargo, such as proteins, lipids, and genetic material, e.g. miRNA [29,30]. The fact that the composition of sEVs (proteins, lipids, mRNAs and miRNAs) reflects the type of cell and the physiological status of such cell, has led to propose sEVs as biomarkers for diagnosis of various human diseases [31,32], also those of the visual system [33], where their role in normal and diseased eye has been reviewed [34].
It is generally accepted that sEVs production is stimulated by oxidative stress: Malik and collaborators showed that either 2 h ischemia followed by 1 h reperfusion, or ethanol, were able to stimulate sEVs production in cardiac myocytes [35]. They showed that ethanol-induced sEVs appeared to have an increased protein content compared to hypoxia and reoxygenation-induced sEVs, based on similar acetylcholine-esterase activity, which reflects the amount of membrane present. Furthermore, after two hours of ethanol exposure, a dramatical increase of ROS in cardiac myocytes was also observed. Interestingly, after treatment with antioxidants to reduce ROS production, a decreased sEVs formation was obtained. A similar approach but with retinal pigment epithelial cells also confirmed that under oxidative conditions the amount of sEVs produced by these cells was increased and their cargo also varied [22,36]. A very innovative approach using human primary RPE cultures from human induced pluripotent stem cells (hiPSC)-derived retinal organoids has recently described that sEVs released from these cells under oxidative burden, contain drusen-associated proteins [37]; this represents a new insight in the pathophysiology of AMD that certainly confirms a crucial role for sEVs in this process.
Other authors have used rotenone to challenge ARPE-19 cells and compared the effects of the sEVs obtained from these cells with those of the ones from control cells. sEVs secreted by rotenone stimulated ARPE-19 cells could induce cell apoptosis, oxidative injury, and inflammation in control ARPE-19 cells; these same sEVs secreted under OS can damage retinal functions of rats and upregulate the expression of Apaf1. This overexpression of Apaf1 in sEVs secreted under OS can cause the inhibition of cell proliferation, the increase of cell apoptosis and elicitation of inflammatory response in ARPE-19 cells, through a caspase-9 apoptotic pathway [38]. Subretinal injection of sEVs derived from ARPE-19 cells, in a mouse model of retinal degeneration (accomplished by the prior i.p. injection of MNU) (Methyl-Nitroso-Urea), confirmed the amelioration of visual function impairments, and the structural damages in the retina of MNU-treated mice; furthermore, RPE-derived sEVs exert beneficial effects on the electrical response of the inner retinal circuits and suppressed the expression levels of inflammatory factors, and mitigated the oxidative damage, in MNU-treated mice [39]. Co-cultures of ARPE-19 (remember of human origin) and primary porcine RPE cells in a transwell model, stimulated with hydrogen peroxide, produced sEVs that contained HDAC6, which is known to reduce tight junction stability, action that was dependent on sEVs uptake; this led these authors to propose that sEVs are able to communicate stress messages to healthy RPE cells, eventually contributing to RPE dysfunction [40].
Kannan and collaborators have recently reviewed the role of alpha crystallins in the RPE and their implications for the pathogenesis and treatment of AMD [41]. This research group provided evidence that under severe oxidative conditions, RPE released alpha crystallin via the exosomal pathway and that it accumulated in the basolateral side of RPE, once the barrier function has broken down, suggesting sEVs as a source of alpha crystallin for the drusen [42]. α- Crystallins are prominent members of the small heat shock protein family, and apart from the well recognized chaperone effect, a wide variety of other properties have been described, including anti-inflammatory, antifibrillar, antiapoptotic properties, protection against ER stress and autophagy, and modulation of angiogenesis [43]. It is noteworthy, that most of the studies that address the antiapoptotic function and associated signaling mechanisms of α-crystallins use oxidative stimuli as a model for such studies, and in fact, RPE cells overexpressing either αA or αB crystallin contained increased cellular glutathione (GSH). Moreover, these same authors [43] have showed that a selective increase in mitochondrial GSH compartment of oxidatively stressed RPE in αA and αB overexpressing cells provided cellular protection, a formerly proposed role for the mitochondrial GSH pool in different cell types [44,45,46,47]. This is reinforced by the proposal that mitochondrial DNA damage is involved in the pathophysiology of AMD [48]. Interestingly, theses sEVs have been also related to other eye diseases such as glaucoma. It has been recently and elegantly confirmed that enriched sEVs (obtained under hypoxic conditions) derived from human amniotic membrane stem cells, containing large numbers of functional molecules, improved glaucoma by protecting retinal cells against oxidative and hypoxic injuries in vitro and by recovering IOP and retinal degeneration in vivo [49].
Using an experimental approach with primary polarized RPE cultures under subtoxic oxidative stress conditions, changes in sEVs proteins content involved in epithelial barrier integrity have been reported, that include basal-side specific desmosome and hemidesmosome shedding via sEVs [50], that would explain the RPE impaired outer blood-retinal-barrier function that occurrs in AMD. Cellular oxidative damage of the RPE cells induce by NaIO3 could be prevented by the administration of sEVs obtained from mesenchymal stem cells [51] that decreased the levels of reactive oxygen species and upregulated SOD activity by a mechanism that has been ascribed to the up regulation of the expression of the Nrf2 pathway, since Nrf2 inhibitors block this antioxidant effect [51]. Furthermore, it has been recently described that oxidative conditions induce in RPE cells the expression of CYLD-AS1, a long noncoding mRNA able to regulate the expression of Nrf2 (as just mentioned, related to oxidative stress), and NF-κB pathway members, which are related to inflammation. Remarkably, these two signaling pathways were mediated by the CYLD-AS1 interactor miR-134-5p. Moreover, sEVs secreted by CYLD-AS1 knockdown RPE cells had a lower proinflammatory effect than those secreted by control cells [52]. These authors proposed that targeting CYLD-AS1 would be an approach for the treatment of AMD.
It has been reported that the complement regulatory protein (CD59) was increased in regions of the RPE of early AMD patients, but decreased in advanced forms of AMD [53]. This same Dr. Handa’s group showed that this protein was released within sEVs to the subretinal space [54]. The capability of RPE-released sEVs to induce angiogenesis in an experimental model has also been demonstrated, proposing for the first time the hypotheses of sEVs as relevant players of retinal neovascularization [36].
2.2. Angiogenesis
Angiogenesis is the formation of new blood vessels from existing vasculature in response to different stimuli. Two types of angiogenesis are distinguished: physiological and pathological. The first one occurs during embryonic vasculature growth and continues after birth in early postnatal development. The formation of new blood vessels during development is necessary to provide oxygen and required nutrients to growing organs. On the other hand, pathological neovascularization mechanisms are characterized by an uncontrolled and disorderly growth of vasculature and are displayed when there is an imbalance between the stimulators and inhibitors of angiogenesis. The first description of this process was made by Judah Folkman in 1971 [55]. Two main cell types are involved in angiogenesis: the pericytes and endothelial cells, the formers by maintaining vascular stability and thus enhancing endothelial cells barrier function by direct contact and paracrine regulation (excellently reviewed in [56]). The pericytes activated by the hypoxia-inducible factor pathway in a hypoxia condition can secrete sEVs that regulate endothelial cell migration, sprouting, and angiogenesis in the wound healing model and spinal cord explant cultures [57]. Thus, pericytes themselves, sEVs from neurons, glia, and endothelial cells, as well as the circulation, can regulate EC integrity and intercellular crosstalk within NVU in both physiological and pathological conditions [58,59,60].
Abnormal angiogenesis allows the extravasation of plasma proteins, establishing a provisional matrix, necessary for the migration of activated endothelial cells. Basement membrane and extracellular matrix are degraded locally by metalloproteases, allowing the underlying endothelial cells to migrate following the trail of the angiogenic stimulus into the perivascular space [61]. Another study [62], not originally designed to look for retinal pathophysiological mechanisms but for the microvascular damage in diabetes, obtained results that fit again in demonstrating that sEVs from adipocytes obtained in a high glucose environment, were able to damage retina microvascular endothelial cells via a LINC00968/miR-361-5p/TRAF3 signaling pathway [62].
Hajrasouliha’s group has tested the hypothesis that cells within the neurosensory retina may release sEVs that regulate angiogenesis. They observed that these sEVs, derived from retinal astrocytes of normal mice, contain multiple antiangiogenic factors that inhibit the development of choroidal neovascularization in a laser-induced model, whereas those derived from RPE do not. The results of these studies indicate that in the retina, sEVs derived from different cells may influence the balance of anti- and proangiogenesis. The mechanisms by which retinal astrocytes sEVs suppress retinal vascular leakage and inhibit choroidal neovascularization in this laser-induced mouse model might be that retinal astrocytes sEVs target both macrophages and vascular endothelial cells. Infiltrating macrophages are a major source of inflammatory cytokines, complement, and VEGF, all of which are important in the pathogenesis of inflammation and new vessel formation. Retinal astrocytes derived sEVs suppressed the infiltration and migration of macrophages in a chemotactic chamber. These sEVs also inhibited endothelial cells migration and vascular tubule formation. The antiangiogenic effect of retinal astrocytes sEVs on macrophages and endothelial cells might be possible by their cargo which includes proteins, lipids, mRNA, and miRNAs. The analysis with an antibody array of retinal astrocytes-derived sEVs revealed the presence of molecules with anti-angiogenic properties such as endostatin, PEDF, and TIMP-1. However, several chemokines and metalloproteases were detected in these sEVs that can either promote or inhibit angiogenesis, depending on their environmental context [63].
The role of RPE cell-derived sEVs in the pathogenesis of dry (specially after the findings by Flores-Bellver and coworkers [37] mentioned above), and wet AMD cannot be ignored. RPE cell-derived sEVs may directly promote the occurrence and development of choroidal neovascularization in wet AMD. It has been demonstrated that oxidative burden induces RPE cells to secrete more sEVs, and that these sEVs effectively transmit signals that were able to induce tube formation in endothelial cells [22]. Furthermore, it was demonstrated that stressed RPE cells released a higher amount of sEVs than controls, with a higher expression of VEGF receptor (VEGFR) in the membrane, and enclosed an extra cargo of VEGFR mRNA. Angiogenesis assays confirmed that endothelial cells increased their tube formation capacity when exposed to stressed RPE sEVs [22]. In following experiments, we established a link between two phenomena: autophagy and sEVs release in RPE cells under oxidative conditions and the contribution of these two cellular mechanisms to the regulation of angiogenesis [36]. In a physiological environment, there is equilibrium between angiogenic activators and inhibitors, resulting in very limited new blood vessel formation. Nevertheless, angiogenic stimulators can break the balance in a number of visual conditions, such as AMD and DR [64,65], or, the other way around, as it has been recently proposed, the potential efficacy of bone marrow-derived mesenchymal stem cell-derived sEVs in diabetes-induced retinal injury via suppressing Wnt/ b-catenin signaling and subsequent reduction of oxidative stress, inflammation and angiogenesis [66].
2.3. Autophagy and sEVs
Perturbing RPE homeostasis by impairing autophagy promotes inflammasome activation and activates macrophage-mediated angiogenesis, which influences key features of AMD development [67]. Some authors have demonstrated the link between autophagy and exosome biogenesis in disease. Bhattacharya and collaborators described how GAIP interacting protein C terminus (GIPC) regulates both mechanisms in pancreatic tumor cells [68]. Wang described how autophagy and RPE-derived sEVs work together for the generation of drusen in AMD [69,70]. An increased autophagy enhances tube formation and endothelial cells migration. Moreover, if autophagy is inhibited endothelial cells migration and tube formation are reduced [71]. Oxidative stress-induced retinal astrocytes modulate proliferation and migration of endothelial cells by increasing retinal astrocytes’ autophagy and the release of sEVs [72].
Increasing evidences indicate that impaired autophagy is associated with angiogenesis, both in the development of the chick embryo [73], where Atg7 plays an important role, as in RF/6A cells, from choroid, where hypoxia-induced autophagy stimulated endothelial growth [74]. Results from our group indicate that inhibiting autophagy in ARPE-19 cells by Atg7 siRNA, decreases the expression of VEGFR2 in low-stressed RPE cells. This effect was accompanied by a decrease in the total number of sEVs and the VEGFR2-positive sEVs released from ARPE-19 cells [36]. Interestingly, high doses of the antioxidant curcumin have shown activation of autophagy [75]; these authors did not evaluate sEVs release though their results allow the speculation that, under these conditions, an increase in sEVs could be certainly expected.
Similarly to what other authors have done, photo-oxidative stimulation with blue-light induced ARPE-19 cells to release sEVs with higher levels of IL-1β, IL-18, and caspase-1 than those unstimulated, whereas the levels of these same factors in these cells were significantly enhanced when treated with sEVs from stimulated cells; furthermore, the NLRP3 mRNA and protein levels were found to be markedly higher in the treated group than in the untreated one [76]. An interesting correlation has been proposed between NLRP3 inflammasome activity and autophagy [77].
Paraquat addition to ARPE-19 cells somehow mimics the enhanced oxidative burden of the cellular environment in neovascular-AMD. A proteomic analysis of the sEVs released under this condition was performed and the outcome showed upregulation of proteins related to autophagy pathway, such as cathepsin D, similar results to what was observed in the aqueous humor of AMD patients [78].
sEVs released from the RPE seem to cross the Bruch’s membrane and target choroidal endothelial cells, influencing their fate. Besides, sEVs from retinal astrocytes seem to reduce vessel leakage in a model of AMD, whereas RPE-originating sEVs do not stop the new vessels from leaking [63]. Moreover, it has been proposed in another AMD model that sEVs released by ageing RPE cells are able to increase autophagy in neighboring cells, and that this can contribute to drusen formation [69]. In parallel, another group demonstrated that sEVs derived from mesenchymal stem cells inhibited neovascularization by down-regulating VEGF expression [79]. It is therefore well established already that both processes, autophagy and exosome biogenesis are in some ways related. It was also observed that oxidative challenge in ARPE-19 cells does not kill them, but rather it enhances autophagy [80], VEGF release [81,82], and sEVs liberation [22,36]. RPE might not be the only cellular type of the retina that might release different types of extracellular vesicles, as a modulation response, as has been recently described, studying the EVs activity within the retina during poly ADP ribosylation in degenerating rd10 [83], and rd1 [84] mice retinae.
2.4. Micro RNAs
As mentioned above, sEVs might contain different types of molecules, among which are micro RNA (miRNA) sequences [25], short (around 22 nucleotides), single-stranded RNA molecules that have been proposed as potential therapeutic tools and diagnostic markers in DR [85,86] and also AMD [87,88]. They are found inside the cell as well as the extracellular medium, including plasma. MiRNAs are non coding RNAs involved in transcription and post-transcriptional messenger RNA (mRNA) sequences normally by degrading mRNA transcription or even repressing translational process. As an example, in RPE samples from elderly humans it has been described that miR-21 from EVs that were then taken up by microglia, led to a change in expression of genes in the p53 signaling pathway, implicating microglia in the regulation of aging and AMD [89]. It has been also demonstrated that injection of up-regulated exosomal miR-17-3p contained in sEVs from human umbilical cord mesenchymal stem cells, reduced the blood glucose and HbAlc, increased body weight, Hb content and glutamine synthetase level, decreased contents of inflammatory factors and VEGF, alleviated oxidative injury, and inhibited retinal cell apoptosis in an experimental mouse diabetes model, through inhibiting signal transducer and activator of transcription 1 (STAT1) [90].
These miRNAs contained in sEVs in fact have been proposed as mediators of homeostasis [91], which in fact we believe they might be. In this report photo-oxidative conditions induced a decrease in the number, but not in their size, of sEVs production (contrary to what has been reported by others under photo-oxidative stimuli [76]), obtained from whole mouse retina homogenates. Other publications showed, as mentioned above, that other oxidative stimuli in fact increase the amount of sEVs production [22,35,36], it might well be that this discrepancy resides in the sEVs collecting procedure. Nevertheless, these authors proposed that retinal health requires optimal levels of sEVs, and their cargo; thus, replenishing sEVs loads in the retina may prove to be an efficacious therapy, as mentioned above [39].
Furthermore, some miRNAs have been involved in eye-related vascular proliferation. Initially proposed as an hypothesis that RPE derived sEVs and most probably their miRNA content could be related to choroidal neovascularization in AMD [92], it was relatively soon confirmed in different experimental models [22,25,36,82,93,94]. Collectively identified as ¨angiomiRNAs¨, they are expressed in endothelial cells, mir-126, mir-210, mir-221/222, among others, and are able to promote or repress angiogenesis (reviewed in [95]). In this sense, bone marrow mesenchymal stem cells derived sEVs are able to reduce inflammation and angiogenesis, concomitantly with the up regulation of antioxidant enzymes, in diabetes-induced retinal injury [96].
As already mentioned, oxidative stimuli promoted sEVS release from RPE. These sEVS contain different miRNA cargoes with pro-angiogenic properties [25,36]. Recent data from our lab indicate that mir-205 negatively modulates HIF-1a and VEGFA mRNA expression [93]. Under oxidative conditions (high glucose), mir-205 levels significantly decreased while HIF-1a, and VEGFA mRNA expression levels were increased finally, leading to angiogenesis in human umbilical vein endothelial cells (HUVEC) cultures. After adding the antioxidant NAC, mir205 levels were normalized as well as HIF-1a and VEGFA mRNA expression levels also did it. Under these conditions, HUVEC cells were unable to produce tubes and branches [93]. In a similar approach, we transfected ARPE-19 cells with a miR-302a-3p mimic and compared the effect of adding sEVs obtained from these cells to HUVEC, on tube and vessel formation, with the effect of sEVs from ARPE-19 cells incubated with hydrogen peroxide, and confirmed that miR-302a-3p contained in sEVs can modify VEGFA mRNA expression levels as part of its antiangiogenic features [94]. These results strongly indicate the pivotal role of miRNAs as angiogenic modulators specially under oxidative conditions.
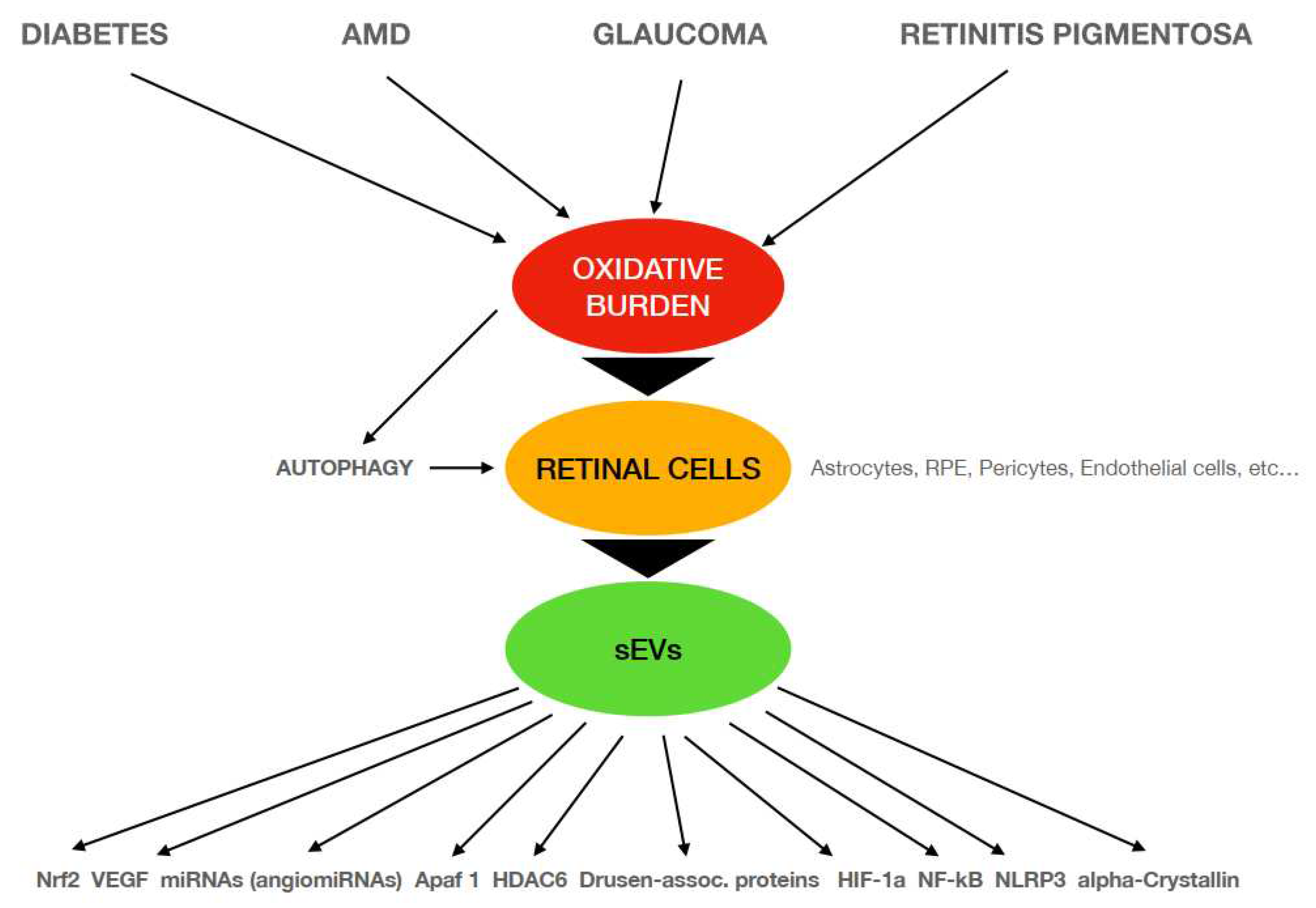
Figure 1 represents an schematic diagram of the main relationships described in this review that confirm sEVs release and their cargo as mediators in the retinal diseases studied and the different pathways, directly or indirectly involved in these oxidative-related pathophysiological mechanisms.
Funding
Authors’ research receives support from: ‘Consolidación de Indicadores’ CEU-UCH 2022-2023 and ‘Proyectos Puente y en Consolidación’ CEU FUSP 2022-2023 (MMi), Agencia Estatal de Investigación (AEI), grant number PID2020-117875GB-I00/AEI/ 10.13039/501100011033, and Instituto de Salud Carlos III (ISCIII), grant number PI21/00083 (MMu, JS-P and JMB).
References
- Meister, M.; Tessies-Lavigne, M. Low-level visual processing: the retina. In Principles of Neural Sciences; Kandel, E.R., Schwartz, J.M., Jessell, T.M., Siegelbaum, S.A., Hudspeth, A.J., Eds.; McGraw-Hill: New York, 2013; pp. 577-601.
- Wässle, H.; Boycott, B.B. Functional architecture of the mammalian retina. Physiol. Rev. 1991, 71, 447-480. [CrossRef]
- Panda-Jonas, S.; Jonas, J.B.; Jakobczyk-Zmija, M. (1996). Retinal pigment epithelial cell count, distribution, and correlations in normal human eyes. Am. J. Ophthalmol. 1996, 121, 181-189. [CrossRef]
- Boulton, M.; Dayhaw-Barker, P. The role of the retinal pigment epithelium: topographical variation and ageing changes. Eye 2001, 15, 384-389. [CrossRef]
- Marmor, M.F. Structure and function of the retinal pigment epithelium. Int. Ophthalmol. Clin. 1975, 15, 115-130. [CrossRef]
- Somasundaran, S.; Constable, I.J.; Mellough, C.B.; Carvalho, L.S. Retinal pigment epithelium and age-related macular degeneration: A review of major disease mechanisms. Clin. Exp. Ophthalmol. 2020, 48, 1043-1056. [CrossRef]
- Tonade, D.; Kern, T.S. Photoreceptor cells and RPE contribute to the development of diabetic retinopathy. Prog. Retin. Eye Res. 2021, 83, 100919. [CrossRef]
- Beatty, S.; Koh, H.-H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [CrossRef]
- Miranda, M.; Johnson, L.E.; Ahuja, S.; Ekstrom, P.A.; Romero, F.J.; van Veen, T. Significant photoreceptor rescue by treatment with a combination of antioxidants in an animal model for retinal degeneration. Neuroscience 2007, 145, 1120–1129. [CrossRef]
- Cantó, A.; Martínez-González, J.; Almansa, I.; López-Pedrajas, R.; Hernández-Rabaza, V.; Olivar, T.; Miranda, M. Time-course changes in oxidative stress and inflammation in the retinas of rds mice: a retinitis pigmentosa model. Antioxidants (Basel) 2022, 11, 1950. [CrossRef]
- van Reyk, D.M.; Gillies, M.C.; Davies, M.J. The retina: oxidative stress and diabetes. Redox Rep. 2003, 8, 187-192. [CrossRef]
- Romero F.J.; Bosch-Morell, F.; Romero M.J.; Jareño, E.J.; Romero, B.; Marín, N.; Romá J. Lipid peroxidation products and antioxidants in human disease. Environ. Health Perspect. 1998, 106 (suppl. 5), 1229-1234.
- Puertas, F.J.; Díaz-Llopis, M.; Chipont, E.; Romá, J.; Raya, A.; Romero, F.J. Glutathione system of human retina. Enzymatic conjugation of lipid peroxidation products. Free Radical Biol. Med. 1993, 14, 549-551. [CrossRef]
- Farrar, G.J. On the genetics of retinitis pigmentosa and on mutation-independent approaches to therapeutic intervention. EMBO J. 2002, 21, 857–864.
- Stoorvogel, W.; Kleijmeer, M.J.; Geuze, H.J.; Raposo, G. The biogenesis and functions of exosomes. Traffic 2002, 3, 321-330. [CrossRef]
- Balaj, L.; Lessard, R.; Dai, L.; Cho, Y.J.; Pomeroy, S.L.; Breakefield, X.O.; Skog, J. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nature Comm. 2011, 2, 180. [CrossRef]
- Aliotta, J. M., Pereira, M., Johnson, K. W., de Paz, N., Dooner, M. S., Puente, N., Ayala, C., Brilliant, K.; Berz, D.; Lee, D.; Ramratnam, B.; McMillan, P. N.; Hixson, D. C.; Josic, D.; Quesenberry, P. J. Microvesicle entry into marrow cells mediates tissue-specific changes in mRNA by direct delivery of mRNA and induction of transcription. Exp. Hematol. 2010, 38, 233-245. [CrossRef]
- Baixauli, F.; López-Otín, C.; Mittelbrunn, M. Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness. Front. Immunol. 2014, 5, 403. [CrossRef]
- Castellana, D.; Zobairi, F.; Martinez, M.C.; Panaro, M.A.; Mitolo, V.; Freyssinet, J.M.; Kunzelmann, C. Membrane microvesicles as actors in the establishment of a favorable prostatic tumoral niche: a role for activated fibroblasts and CX3CL1-CX3CR1 axis. Cancer Res. 2009, 69, 785-793. [CrossRef]
- Sadallah, S.; Eken, C.; Schifferli, J.A. Ectosomes as modulators of inflammation and immunity. Clin. Exp. Immunol. 2011, 163, 26-32. [CrossRef]
- Tetta, C.; Bruno, S.; Fonsato, V.; Deregibus, M.C.; Camussi, G. The role of microvesicles in tissue repair. Organogenesis 2011, 7, 105-115. [CrossRef]
- Atienzar-Aroca, S.; Flores-Bellver, M.; Serrano-Heras, G.; Martinez-Gil, N.; Barcia, J. M.; Aparicio, S.; Perez-Cremades, D.; Garcia-Verdugo, J.M.; Diaz-Llopis, M.; Romero, F.J.; Sancho-Pelluz, J. Oxidative stress in retinal pigment epithelium cells increases exosome secretion and promotes angiogenesis in endothelial cells. J. Cell. Mol. Med. 2016, 20, 1457–1466. [CrossRef]
- Biasutto, L.; Chiechi, A.; Couch, R.; Liotta, L.A.; Espina, V. Retinal pigment epithelium (RPE) exosomes contain signaling phosphoproteins affected by oxidative stress. Exp. Cell Res. 2013, 319, 2113–2123. [CrossRef]
- Freeman, D.W.; Noren Hooten, N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered extracellular vesicle concentration, cargo, and function in diabetes. Diabetes 2018, 67, 2377–2388. [CrossRef]
- Oltra, M.; Vidal-Gil, L.; Maisto, R.; Oltra, S.S.; Romero, F.J.; Sancho-Pelluz, J.; Barcia, J. M. MiR302a and 122 are deregulated in small extracellular vesicles from ARPE-19 cells cultured with H2O2. Sci. Rep. 2019, 9, 17954. [CrossRef]
- Greening, D.W.; Simpson, R.J. Understanding extracellular vesicle diversity–current status. Expert Rev. Proteomics 2018, 15, 887–910.
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell. Biol. 2018, 19, 213–228. [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J. D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G. K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [CrossRef]
- Rashed, M.H.; Bayraktar, E.; Helal, G.K.; Abd-Ellah, M.F.; Amero, P.; Chavez-Reyes, A.; Rodriguez-Aguayo, C. Exosomes: from garbage bins to promising therapeutic targets. Int. J. Mol. Sci. 2017, 18, 538. [CrossRef]
- Kao, C.Y.; Papoutsakis, E.T. Extracellular vesicles: exosomes, microparticles, their parts, and their targets to enable their biomanufacturing and clinical applications. Curr. Opin. Biotechnol. 2019, 60, 89–98. [CrossRef]
- Fries, G.R.; Quevedo, J. Exosomal microRNAs as potential biomarkers in neuropsychiatric disorders. Methods Mol. Biol. 2018, 1733, 79-85.
- Roy, S.; Hochberg, F. H.; Jones, P. S. Extracellular vesicles: the growth as diagnostics and therapeutics; a survey. J. Extracell. Vesicles 2018, 7, 1438720. [CrossRef]
- Van der Merwe, Y.; Steketee, M.B. Extracellular vesicles: biomarkers, therapeutics, and vehicles in the visual system. Curr. Ophthalmol. Rep. 2017, 5, 276–28.
- Klingeborn, M.; Dismuke, W.M.; Bowes Rickman, C.; Stamer, W.D. Roles of exosomes in the normal and diseased eye. Prog. Retin. Eye Res. 2017, 59, 158-177. [CrossRef]
- Malik, Z.A.; Kott, K.S.; Poe, A.J.; Kuo, T.; Chen, L.; Ferrara, K.W.; Knowlton, A.A. Cardiac myocyte exosomes: stability, HSP60, and proteomics. Am. J. Physiol.-Heart Circ. Physiol. 2013, 304, H954-H965. [CrossRef]
- Atienzar-Aroca, S.; Serrano-Heras, G.; Freire Valls, A.; Ruiz de Almodovar, C.; Muriach, M.; Barcia, J.M.; Garcia-Verdugo, J.M.; Romero, F.J.; Sancho-Pelluz, J. Role of retinal pigment epithelium-derived exosomes and autophagy in new blood vessel formation. J. Cell. Mol. Med. 2018, 22, 5244-5256. [CrossRef]
- Flores-Bellver, M.; Mighty, J.; Aparicio-Domingo, S.; Li, K.; Shi, C.; Zhou, J.; Cobb, H.; McGrath, P.; Michelis, G.; Lenhart, P.; et al. Extracellular vesicles released by human retinal pigment epithelium mediate increased polarised secretion of drusen proteins in response to AMD stressors. J. Extracell. Vesicles 2021, 10, e12165. [CrossRef]
- Ke, Y.; Fan, X.; Rui, H.; Xinjun, R.; Dejia, W.; Chuanzhen, Z.; Li, X. Exosomes derived from RPE cells under oxidative stress mediate inflammation and apoptosis of normal RPE cells through Apaf1/caspase-9 axis. J. Cell. Biochem. 2020, 121, 4849-4861.
- Wang, Y.; Zhang, Q.; Yang, G.; Wei, Y.; Li, M.; Du, E.; Li, H.; Song, Z.; Tao, Y. RPE-derived exosomes rescue the photoreceptors during retina degeneration: an intraocular approach to deliver exosomes into the subretinal space. Drug Deliv. 2021, 28, 218-228.
- 40. Shah N, Ishii M, Brandon C, Ablonczy Z, Cai J, Liu Y, Chou CJ, Rohrer B. Extracellular vesicle-mediated long-range communication in stressed retinal pigment epithelial cell monolayers. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2610-2622. [CrossRef]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. Novel roles for α-crystallins in retinal function and disease. Prog. Retin. Eye Res. 2012, 31, 576-604. [CrossRef]
- Sreekumar, P.G.; Kannan, R.; Kitamura, M.; Spee, C.; Barron, E.; Ryan, S.J.; Hinton, D.R. αB crystallin is apically secreted within exosomes by polarized human retinal pigment epithelium and provides neuroprotection to adjacent cells. PLoS One 2010, 5, e12578. [CrossRef]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. Alpha crystallins in the retinal pigment epithelium and implications for the pathogenesis and treatment of age-related macular degeneration. Biochim. Biophys. Acta 2016, 1860, 258-268. [CrossRef]
- Almansa, I.; Fernández, A.; García-Ruiz, C.; Muriach, M.; Barcia, J. M.; Miranda, M.; Fernández-Checa, J.C.; Romero, F.J. Brain mitochondrial alterations after chronic alcohol consumption. J. Physiol. Biochem. 2009, 65, 305-312. [CrossRef]
- Romero, F.J.; Romá, J. Careful consideration of the effects induced by glutathione depletion in rat liver and heart. The involvement of cytosolic and mitochondrial glutathione pools. Chem. Biol. Interact. 1989, 70, 29-37. [CrossRef]
- Romero, F.J.; Sies, H. Subcellular glutathione contents in isolated hepatocytes treated with L-buthionine sulfoximine. Biochem. Biophys. Res. Commun. 1984, 123, 1116-1121. [CrossRef]
- Romero, F.J.; Soboll, S.; Sies, H. Mitochondrial and cytosolic glutathione after depletion by phorone in isolated hepatocytes. Experientia 1984, 40, 365-367. [CrossRef]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of mitochondrial DNA damage in ROS-mediated pathogenesis of age-related macular degeneration (AMD). Int. J. Mol. Sci. 2019, 20, 2374. [CrossRef]
- Seong, H.R.; Noh, C.H.; Park, S.; Cho, S.; Hong, S.J.; Lee, A.Y.; Geum, D.; Hong, S.C.; Park, D.; Kim, T.M.; et al. Intraocular pressure-lowering and retina-protective effects of exosome-rich conditioned media from human amniotic membrane stem cells in a rat model of glaucoma. Int. J. Mol. Sci. 2023, 24, 8073. [CrossRef]
- Hernandez, B.J.; Skiba, N.P.; Plössl, K.; Strain, M.; Liu, Y.; Grigsby, D.; Kelly, U.; Cady, M.A.; Manocha, V.; Maminishkis, A.; et al. Polarized desmosome and hemidesmosome shedding via small extracellular vesicles is an early indicator of outer blood-retina barrier dysfunction. J. Extracell. Biol. 2023, 2, e116. [CrossRef]
- Tang, Y.; Kang, Y.; Zhang, X.; Cheng, C. Mesenchymal stem cell exosomes as nanotherapeutics for dry age-related macular degeneration. J. Control Release 2023, 357, 356-370. [CrossRef]
- Du, H.; Huang, Z.; Zhou, X.; Kuang, X.; Long, C.; Tang, H.; Zeng, J.; Huang, H.; Liu, H.; Zhu B.; et al. Oxidative stress-induced lncRNA CYLD-AS1 promotes RPE inflammation via Nrf2/miR-134-5p/NF-κB signaling pathway. FASEB J. 2022, 36, e22577.
- Ebrahimi, K.B.; Fijalkowski, N.; Cano, M.; Handa, J.T. Decreased membrane complement regulators in the retinal pigmented epithelium contributes to age-related macular degeneration. J. Pathol. 2013, 229, 729-742. [CrossRef]
- Ebrahimi, K.B.; Fijalkowski, N.; Cano, M.; Handa, J.T. Oxidized low-density-lipoprotein-induced injury in retinal pigment epithelium alters expression of the membrane complement regulatory factors CD46 and CD59 through exosomal and apoptotic bleb release. Adv. Exp. Med. Biol. 2014, 801, 259-65.
- Folkman J. Tumor angiogenesis: therapeutic implications. N. Engl. J. Med. 1971, 285, 1182-1186. [CrossRef]
- Huang, H. Pericyte-endothelial interactions in the retinal microvasculature. Int. J. Mol. Sci. 2020, 21, 7413. [CrossRef]
- Mayo, J.N.; Bearden, S.E. Driving the hypoxia-inducible pathway in human pericytes promotes vascular density in an exosome-dependent manner. Microcirculation 2015, 22, 711–723. [CrossRef]
- Xu, B.; Zhang, Y.; Du, X.F.; Li, J.; Zi, H.X.; Bu, J.W.; Yan, Y.; Han, H.; Du, J.L. Neurons secrete miR-132-containing exosomes to regulate brain vascular integrity. Cell Res. 2017, 27, 882–897. [CrossRef]
- Huang, C.; Fisher, K.P.; Hammer, S.S.; Navitskaya, S.; Blanchard, G.J.; Busik, J.V. Plasma exosomes contribute to microvascular damage in diabetic retinopathy by activating the classical complement pathway. Diabetes 2018, 67, 1639–1649. [CrossRef]
- Zagrean, A.M.; Hermann, D.M.; Opris, I.; Zagrean, L.; Popa-Wagner, A. Multicellular crosstalk between exosomes and the neurovascular unit after cerebral ischemia. Therapeutic implications. Front. Neurosci. 2018, 12, 811. [CrossRef]
- Bergers, G., Benjamin, L.E. Tumorigenesis and the angiogenic switch. Nature Rev. Cancer 2003, 3, 401-410. [CrossRef]
- He, W.; Lin, A.; Wang, C. Adipocyte-derived exosomal LINC00968 promotes mouse retina microvascular endothelial cell dysfunction in a high-glucose environment by modulating the miR-361-5p/TRAF3 axis. Horm. Metab. Res. 2023, 55, 124-135. [CrossRef]
- Hajrasouliha, A.R.; Jiang, G.; Lu, Q.; Lu, H.; Kaplan, H. J.; Zhang, H. G.; Shao, H. Exosomes from retinal astrocytes contain antiangiogenic components that inhibit laser-induced choroidal neovascularization. J. Biol. Chem. 2013, 288, 28058-28067. [CrossRef]
- D’Amore P.A. Mechanisms of retinal and choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 1994, 35, 3974-3979.
- Campochiaro, P.A. Molecular pathogenesis of retinal and choroidal vascular diseases. Prog. Retin. Eye Res. 2015, 49, 67-81. [CrossRef]
- Ebrahim, N.; El-Halim, H.E.A.; Helal, O.K.; El-Azab, N.E.; Badr, O.A.M.; Hassouna, A.; Saihati, H.A.A.; Aborayah, N.H.; Emam, H.T.; El-Wakeel, H.S.; et al. Effect of bone marrow mesenchymal stem cells-derived exosomes on diabetes-induced retinal injury: Implication of Wnt/ b-catenin signaling pathway. Biomed. Pharmacother. 2022, 154, 113554. [CrossRef]
- Liu, J.; Copland, D.A.; Theodoropoulou, S.; Chiu, H. A.A.; Barba, M.D.; Mak, K.W.; Dick, A.D. Impairing autophagy in retinal pigment epithelium leads to inflammasome activation and enhanced macrophage-mediated angiogenesis. Sci. Rep. 2016, 6, 20639. [CrossRef]
- Bhattacharya, S.; Pal, K.; Sharma, A.K.; Dutta, S.K.; Lau, J.S.; Yan, I.K.; Patel, T.C. GAIP interacting protein C-terminus regulates autophagy and exosome biogenesis of pancreatic cancer through metabolic pathways. PloS One 2014, 9, e114409. [CrossRef]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Du, N.; Tso, M.O.; Neufeld, A.H. Autophagy and exosomes in the aged retinal pigment epithelium: possible relevance to drusen formation and age-related macular degeneration. PLoS One 2009, 4, e4160. [CrossRef]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Du, N.; Tso, M.O.; Neufeld, A.H. Autophagy, exosomes and drusen formation in age-related macular degeneration. Autophagy 2009, 5, 563–564. [CrossRef]
- Du, J.; Teng, R.J.; Guan, T.; Eis, A.; Kaul, S.; Konduri, G.G.; Shi, Y. Role of autophagy in angiogenesis in aortic endothelial cells. Am. J. Physiol.-Cell Physiol. 2012, 302, C383-C391. [CrossRef]
- Zhu, L.; Zang, J.; Liu, B.; Yu, G.; Hao, L.; Liu, L.; Zhong, J. Oxidative stress-induced RAC autophagy can improve the HUVEC functions by releasing exosomes. J. Cell Physiol. 2020, 235, 7392-7409. [CrossRef]
- Lu, W.H.; Shi, Y.X.; Ma, Z.L.; Wang, G.; Liu, L.; Chuai, M.; Yang, X. Proper autophagy is indispensable for angiogenesis during chick embryo development. Cell Cycle 2016, 15, 1742-1754. [CrossRef]
- Li, R.; Du, J.; Chang, Y. Role of autophagy in hypoxia-induced angiogenesis of RF/6A cells in vitro. Curr. Eye Res. 2016, 41, 1566-1570. [CrossRef]
- Carozza, G.; Tisi, A.; Capozzo, A.; Cinque, B.; Giovanelli, A.; Feligioni, M.; Floti, V.; Maccarone, R. New insights into dose-dependent effects of curcumin on ARPE-19 cells. Int. J. Mol. Sci. 2022, 23, 14771. [CrossRef]
- Zhang, W.; Ma, Y.; Zhang, Y.; Yang, J.; He, G.; Chen, S. Photo-oxidative blue-light stimulation in retinal pigment epithelium cells promotes exosome secretion and increases the activity of the NLRP3 inflammasome. Curr. Eye Res. 2019, 44, 67-75. [CrossRef]
- Xue, Z.; Zhang, Z.; Liu, H.; Li, W.; Guo, X.; Zhang, Z.; Liu, Y.; Jia, L.; Li, Y.; Ren, Y.; et al. lincRNA-Cox2 regulates NLRP3 inflammasome and autophagy mediated neuroinflammation. Cell Death Differ. 2019, 26, 130-145.
- Kang, G.Y.; Bang, J.Y.; Choi, A.J.; Yoon, J.; Lee, W.C.; Choi, S.; Yoon, S.; Kim, H. C.; Baek, J. H.; Park, H. S.; Lim, H. J.; Chung, H. Exosomal proteins in the aqueous humor as novel biomarkers in patients with neovascular age related macular degeneration. J. Proteome Res. 2014, 13, 581–595. [CrossRef]
- Lee, J.K.; Park, S.R.; Jung, B.K.; Jeon, Y.K.; Lee, Y.S.; Kim, M.K.; Kim, C.W. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PloS One 2013, 8, e84256. [CrossRef]
- Flores-Bellver, M.; Bonet-Ponce, L.; Barcia, J.M.; Garcia-Verdugo, J.M.; Martinez-Gil, N.; Saez-Atienzar, S.; Romero, F.J. Autophagy and mitochondrial alterations in human retinal pigment epithelial cells induced by ethanol: implications of 4-hydroxy-nonenal. Cell Death Dis. 2014, 5, e1328. [CrossRef]
- Kannan, R.; Zhang, N.; Sreekumar, P.G.; Spee, C.K.; Rodriguez, A.; Barron, E.; Hinton, D.R. Stimulation of apical and basolateral VEGF-A and VEGF-C secretion by oxidative stress in polarized retinal pigment epithelial cells. Mol. Vis. 2006, 12, 1649-1659.
- Maisto, R.; Oltra, M.; Vidal-Gil, L.; Martínez-Gil, N.; Sancho-Pellúz, J.; Filippo, C.D.; Rossi, S.; D’Amico, M.; Barcia, J.M.; Romero, F.J. ARPE-19-derived VEGF-containing exosomes promote neovascularization in HUVEC: the role of the melanocortin receptor 5. Cell Cycle 2019, 18, 413-424. [CrossRef]
- Vidal-Gil, L.; Sancho-Pelluz, J.; Zrenner, E.; Oltra, M.; Sahaboglu, A. Poly ADP ribosylation and extracellular vesicle activity in rod photoreceptor degeneration. Sci. Rep. 2019, 9, 3758. [CrossRef]
- Sahaboglu, A.; Vidal-Gil, L.; Sancho-Pelluz, J. Release of retinal extracellular vesicles in a model of retinitis pigmentosa. Adv. Exp. Med. Biol. 2019, 1185, 431-436.
- Greco, M.; Chiefari, E.; Accattato, F.; Corigliano, D. M.; Arcidiacono, B.; Mirabelli, M.; Liguori, R.; Brunetti, F.S.; Pullano, S.A.; Scorcia, V.; et al. Microrna-1281 as a novel circulating biomarker in patients with diabetic retinopathy. Front. Endocrinol. 2020, 11, 1–12. [CrossRef]
- Li, X.; Yu, Z. W.; Wang, Y.; Fu, Y.H.; Gao, X.Y. MicroRNAs: potential targets in diabetic retinopathy. Horm. Metab. Res. 2020, 52, 142–148. [CrossRef]
- Ren, C.; Liu, Q.;Wei, Q.; Cai, W.; He, M.; Du, Y.; Xu, D.;Wu, Y.; Yu, J. Circulating miRNAs as potential biomarkers of age-related macular degeneration. Cell Physiol. Biochem. 2017, 41, 1413–1423. [CrossRef]
- ElShelmani, H.; Wride, M.A.; Saad, T.; Rani, S.; Kelly, D.J.; Keegan, D. The role of deregulated microRNAs in age-related macular degeneration pathology. Transl. Vis. Sci. Technol. 2021, 10, 12. [CrossRef]
- Morris, D.R.; Bounds, S.E.; Liu, H.; Ding, W.Q.; Chen, Y.; Liu, Y.; Cai, J. Exosomal miRNA transfer between retinal microglia and RPE. Int. J. Mol. Sci. 2020, 21, 3541. [CrossRef]
- Li, W.; Jin, L.Y.; Cui, Y.B.; Xie, N. Human umbilical cord mesenchymal stem cells-derived exosomal microRNA-17-3p ameliorates inflammatory reaction and antioxidant injury of mice with diabetic retinopathy via targeting STAT1. Int. Immunopharmacol. 2021, 90, 107010. [CrossRef]
- Wooff, Y.; Cioanca, A.V.; Chu-Tan, J.A.; Aggio-Bruce, R.; Schumann, U.; Natoli, R. Small-medium extracellular vesicles and their miRNA cargo in retinal health and degeneration: mediators of homeostasis, and vehicles for targeted gene therapy. Front. Cell. Neurosci. 2020, 14, 160. [CrossRef]
- Tong, Y.; Zhou, Y.L.; Wang, Y.X.; Zhao, P.Q.; Wang, Z.Y. Retinal pigment epithelium cell-derived exosomes: Possible relevance to CNV in wet-age related macular degeneration. Med. Hypotheses 2016, 97, 98-101. [CrossRef]
- Oltra, M.; Vidal-Gil, L.; Maisto, R.; Sancho-Pelluz, J.; Barcia J. M. Oxidative stress-induced angiogenesis is mediated by miR-205-5p. J. Cell. Mol. Med. 2020, 24, 1428-1436.
- Oltra, M.; Martínez-Santos, M.; Ybarra, M.; Rowland, H.; Muriach, M.; Romero F.J.; Sancho-Pelluz, J.; Barcia, J.M. Oxidative-induced angiogenesis is modulated by small extracellular vesicle miR-302a-3p cargo in retinal pigment epithelium cells. Antioxidants (Basel) 2022, 11, 818. [CrossRef]
- Liu, C.-H.; Huang, S.; Britton, W. R.; Chen, J. MicroRNAs in vascular eye diseases. Int. J. Mol. Sci. 2020, 21, 649. [CrossRef]
- Ebrahim, N.; El-Halim, H.E.A.; Helal, O.K.; El-Azab, N.E.; Badr, O.A.M.; Hassouna, A.; Saihati, H.A.A.; Aborayah, N.H.; Emam, H.T.; El-Wakeel, H.S.; et al. Effect of bone marrow mesenchymal stem cells-derived exosomes on diabetes-induced retinal injury: Implication of Wnt/ b-catenin signaling pathway. Biomed. Pharmacother. 2022, 154, 113554. [CrossRef]
Figure 1.
Schematic diagram of the pathophysiological mechanisms involved in different retinal diseases that are related to oxidative conditions. AMD: Age-related Macular Degeneration; sEVs: small Extracellular Vesicles; RPE: Retinal Pigment Epithelium; VEGF: Vascular Endothelial Growth Factor; Apaf-1: Apoptosis protease-activating factor-1; HDAC6: Histone DeACetylase 6; HIF-1a: Hypoxia-Inducible Factor 1-alpha; NLRP3: NOD-, LRR- and pyrin domain-containing Protein 3.
Figure 1.
Schematic diagram of the pathophysiological mechanisms involved in different retinal diseases that are related to oxidative conditions. AMD: Age-related Macular Degeneration; sEVs: small Extracellular Vesicles; RPE: Retinal Pigment Epithelium; VEGF: Vascular Endothelial Growth Factor; Apaf-1: Apoptosis protease-activating factor-1; HDAC6: Histone DeACetylase 6; HIF-1a: Hypoxia-Inducible Factor 1-alpha; NLRP3: NOD-, LRR- and pyrin domain-containing Protein 3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



