Submitted:
02 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
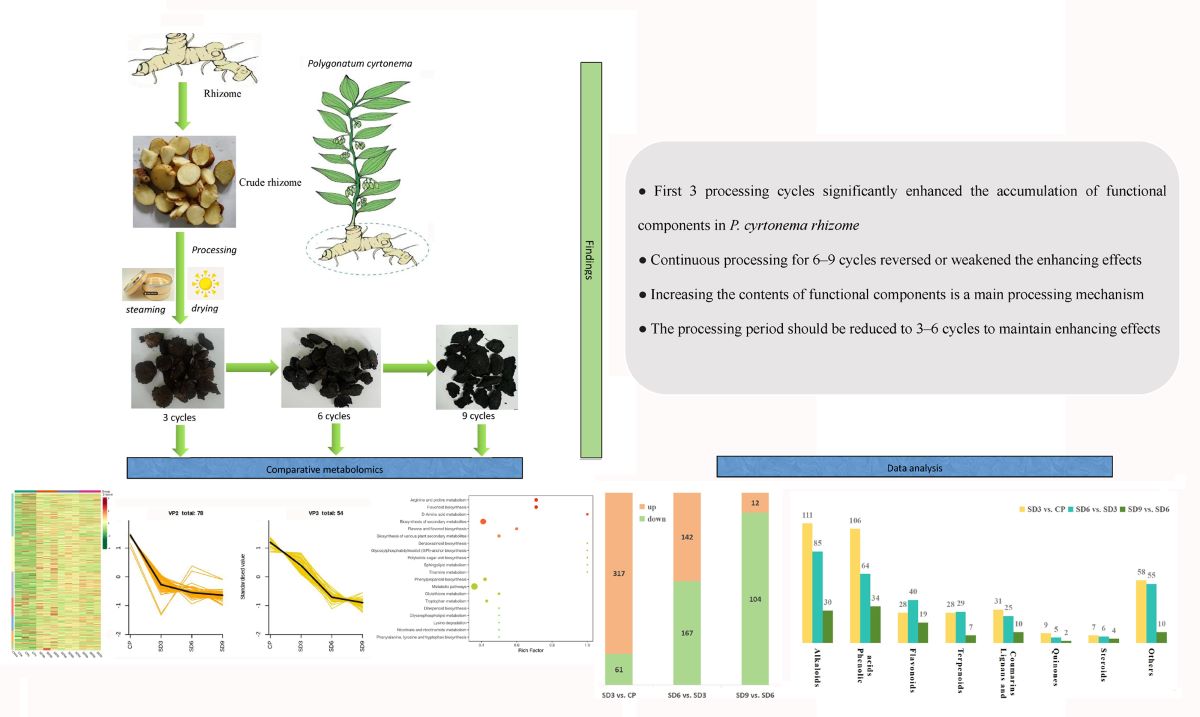
Polygonatum cyrtonema Hua. is an important medicinal and edible food in China and the traditional processing method of nine-steaming and nine-drying has been used to process its rhizome. However, the global dynamic changes in secondary metabolites during the processing and whether this method could enhance the accumulation of functional components remain unclear. This study aimed to explore the processing-induced mechanisms involved in the accumulation of secondary metabolites and provide theoretical support for the pharmacological study on P. cyrtonema. Results showed the first 6 processing cycles induced drastic changes in the accumulation of secondary metabolites, while the last 3 did not induce further changes. The accumulation of most secondary metabolites significantly increased after the first 3 cycles and stabilized after 6 cycles. The first 3 cycles led to numerous newly added metabolites while the last 6–9 cycles led to a small number of disappeared metabolites. Processing enhanced the biosynthesis of various secondary metabolites and significantly affected the metabolisms of amino acids. Together, a short period of processing was helpful for enhancing the biosynthesis of secondary metabolites. However, the enhancing effects will be reversed or weakened under continuous processing. The processing cycles should be reduced to 3–6 to maintain the enhancing effects.
Keywords:
Polygonatum cyrtonema
; medicinal plant
; processing
; secondary metabolites
; functional components
; metabolomics
1. Introduction
Medicinal plants are a rich source of traditional and modern medicine and play an important role in the development of human life. Species in the genus Polygonatum are perennial herbs of Liliaceae. The species P. cyrtonema Hua (“Duohua Huangjin” in Chinese), a known medicinal plant, is recorded in China Pharmacopoeia and has a long history of application in Chinese folk medicine [1,2]. Modern pharmacological studies have shown that P. cyrtonema rhizome plays an important role in anti-aging, immune function regulation, control of blood glucose and blood lipid, memory improvement, and antitumor and antibacterial effects [3,4,5]. This function is attributed to the abundant functional medicinal components derived from the rhizome, such as polysaccharides and secondary metabolites (e.g., saponins, flavonoids, and alkaloids) [6,7]. Therefore, P. cyrtonema rhizome is regarded as an important medicinal and edible food resource and has received increasing attention in recent years in China.
Processing, Paozhi in Chinese, is an ancient Chinese pharmaceutic technique to facilitate the clinical use of Chinese herbal medicines according to traditional Chinese medicine theory [8]. Processing can reduce toxicity, reinforce efficacy, alter the energetic nature and therapeutic direction, and improve the flavor of herbal medicines; these transformations increase the therapeutic effectiveness and applicability of herbal medicines in individualized treatment [9]. For the purpose of detoxification and enhancing efficacy, P. cyrtonema rhizome (crude medicine) is usually processed using a traditional method known as “nine-steaming and nine-drying,” which involves nine cycles of steaming and sun-drying [10,11]. In recent years, several studies have investigated Polygonatum rhizome treated with nine processing cycles. These works include examining the structural changes of polysaccharides, analyzing polysaccharide and extract contents, and determining the relationship between processing degree and the internal and external quality of rhizoma based on color change [11,12,13]. Some findings suggest that nine processing cycles may lead to the loss and change of polysaccharide in P. cyrtonema rhizome, which affect factors such as molecular weight, monosaccharide composition, and particle size distribution [14]. Secondary metabolites (e.g., alkaloids, flavonoids, amines, glycosides, and steroids), a large group of active compounds with low molecular weight are produced at specific stages of the life cycle or morphological differentiation [15]. These secondary metabolites play an important role in the interaction of plants with their environment and have been used in drug and pharmaceutical industries to treat various disorders [16,17,18,19]. Therefore, investigating the changes in secondary metabolites of P. cyrtonema rhizome in abundance and structural transformation is crucial to understand its nutritional value and processing efficacy. However, only a limited number of studies have focused on secondary metabolites in P. cyrtonema rhizome to date. Thus, the global dynamic changes in this species during processing remain unclear. In addition, whether nine processing cycles can retain the functional components to the greatest extent is uncertain. Accordingly, a comprehensive investigation into the processing-induced dynamic changes in secondary metabolites of P. cyrtonema rhizome is necessary.
Plant metabolomics is based on the nonbiased and high-throughput analyses of complex metabolites contained in plant extracts by HPLC-MS (for nonvolatile compounds), GC-MS (for volatile oil), or NMR [20]. Metabolomics is considered an important functional genomics tool because metabolites link genotypes and phenotypes [21,22,23,24,25]. In recent years, metabolomics has been widely employed in various fields, such as medicinal plants, biological interaction, fruit nutritional quality, and biological activity [26]. Our present study focuses on the secondary metabolites derived from P. cyrtonema rhizome, and metabolomics analysis was performed to investigate the dynamic changes in secondary metabolites during traditional processing. Our objectives were to obtain insights into the processing-induced mechanisms involved in the accumulation of secondary metabolites and provide valuable theoretical and metabolic data to support the pharmacological study on P. cyrtonema.
2. Materials and Methods
2.1. Plant Material Selection and Pre-Treatment
Nine fresh, healthy, five-year-old Polygonatum cyrtonema Hua. plants were collected from plantations in Quzhou (28°37′N, 118°49′E; altitude 446 m), Zhejiang Province (Southeast China), in April 2023. The P. cyrtonema rhizomes were harvested from the plants and cleaned with water, and then, the rhizomes of 3 P. cyrtonema plants were mixed as a biological replicate. The whole rhizomes were divided uniformly into two groups: one as crude rhizome (CP group, non-processed) with three replicates, and the other stored at 4 °C for a short period for follow-up “nine-steaming and nine-drying” as processed rhizome (SD groups) with three replicates. The samples in the CP group were further cut into slices and quickly frozen in liquid nitrogen, which were then stored at −80 °C until further analysis.
2.2. Polygonatum Rhizome Processing
The processing of P. cyrtonema rhizome followed a previously described method [13]. In brief, the fresh P. cyrtonema rhizome was placed in a steamer to steam for 6 h, subsequently moisturized overnight, and then dried in an oven at 50 °C for 8 h with 1–9 cycles. Next, rhizomes subjected to 3, 6, and 9 cycles of processing were collected as SD3, SD6, and SD9 groups, respectively. Similarly, the samples in the SD groups were further cut into slices and quickly frozen in liquid nitrogen, which were then stored at −80 °C until further analysis (Figure 1).
2.3. Sample Preparation and Extraction for Widely Targeted Metabolomic Analysis
Sample preparation and extraction followed the methods provided by Metware Biotechnology Co., Ltd. (Wuhan, China). Briefly, the P. cyrtonema rhizome samples were freeze-dried in a lyophilizer (Scientz-100F) using vacuum freeze-drying technology, followed by grinding for 1.5 min at 30 Hz with a grinder (MM 400, Retsch, Germany). Then, 50 mg of powder from each sample was weighted and then dissolved in 1.2 mL of 70% methanolic aqueous pre-cooled at −20 °C. After extraction, the mixtures were centrifuged at 12000 rpm for 3 min, and the supernatant was collected and filtered through a microporous membrane (SCAA-104, 0.22 μm pore size; ANPEL, Shanghai, China). Finally, the filtered samples were stored in injection vials for UPLC-MS/MS analysis.
2.4. UPLC Conditions
The P. cyrtonema rhizome sample extracts were analyzed using the ultra-performance liquid chromatography–electrospray ionization–tandem mass spectrometry system (UPLC-ESI-MS/MS, UPLC, ExionLC™ AD, https://sciex.com.cn). The UPLC analytical conditions were as follows: column, Agilent SB-C18 (1.8 µm, 2.1 mm×100 mm); the mobile phase consisted of solvent A, pure water with 0.1% formic acid, and solvent B, acetonitrile with 0.1% formic acid. Sample measurements were performed with a gradient program that employed the starting conditions of 95% A, 5% B. Within 9 min, a linear gradient to 5% A, 95% B was programmed, and a composition of 5% A, 95% B was kept for 1 min. Subsequently, a composition of 95% A, 5.0% B was adjusted within 1.1 min and kept for 2.9 min. The flow velocity was set at 0.35 mL per minute, the column oven was set to 40 °C, and the injection volume was 2 μL. The effluent was alternatively connected to an ESI-triple quadrupole-linear ion trap (QTRAP)-MS.
2.5. ESI-Q TRAP-MS/MS
The ESI source operation parameters were as follows: source temperature of 500 °C; ion spray voltage (IS) of 5500 V (positive ion mode)/ −4500 V (negative ion mode); ion source gas I (GSI), gas II (GSII), and curtain gas were set at 50, 60, and 25 psi, respectively; the collision-activated dissociation was set to high. Triple quadrupole (QQQ) scans were acquired as multiple reaction monitoring (MRM) experiments, with collision gas (nitrogen) set to medium. Declustering potential (DP) and collision energy (CE) for individual MRM transitions were obtained with further DP and CE optimization. A specific set of MRM transitions was monitored for each period according to the metabolites eluted within that period.
2.6. Qualitative and Quantitative Analysis of Metabolites
Based on the reference library Metware database MWDB v2.0 (Metware Biotechnology Co., Ltd. Wuhan, China) and publicly available metabolite databases, primary and secondary mass spectrometry data obtained using targeted MRM were subjected to qualitative and quantitative analysis. In brief, high-resolution mass spectrometry AB Sciex TripleTOF 6600 was used for qualitative detection of mixed samples in widely target metabolomics, followed by accurate metabolite quantification using AB Sciex 6500 QTRAP for accurate metabolite quantification. This approach combines the advantages of non-targeted and targeted metabolomics by using high-resolution QQQ mass spectrometry with high sensitivity, high specificity, and excellent quantitation capabilities.
During the qualitative analysis of metabolites, repeat signals of K+, Na+, NH4+, and other substances with large molecular weights were eliminated based on the information of secondary spectrum [27,28]. Quantitative analysis was conducted in the MRM mode, where characteristic ions of each metabolite were screened through the QQQ mass spectrometry to obtain signal strengths. After the metabolite profile data of different samples were obtained, integration and correction of chromatographic peaks were performed using MultiQuant v3.0.2 (AB Sciex, Concord, Ontario, Canada). All chromatographic peak area integral data were derived, and the relative content of the corresponding metabolites was calculated based on the peak area integrals [29,30].
2.7. Multivariate Statistical Analysis
Metabolite data were log2-transformed and underwent autoscaling before any statistical analysis. Metabolite data from 12 P. cyrtonema rhizome samples were used for unsupervised principal component analysis (PCA), hierarchical clustering analysis (HCA), and orthogonal partial least squares discriminant analysis (OPLA-DA) using the Metware Cloud, which is a free online platform for data analysis (https://cloud.metware.cn). Briefly, unsupervised PCA was performed using the “prcomp” function within R v3.5.1 (base package, https://www.r-project.org). The HCA results of samples and metabolites were presented as heatmaps with dendrograms. Pearson’s correlation coefficients (PCC) between samples were calculated using the “cor” function in R v3.5.1 and v4.4.0 (base package; Hmisc) and presented as only heatmaps. HCA and PCC were obtained using R v2.8.0 (Complex Heatmap). For HCA, normalized signal intensities of metabolites (unit variance scaling, UV scaling) were visualized as a color spectrum. OPLS-DA was performed using R v1.0.1 (MetaboAnalystR). Before OPLS-DA, the data were log2-transformed and underwent mean centering. A permutation test (200 permutations) was performed to avoid overfitting. The OPLS-DA results also include score and permutation plots [31].
For the two-group analysis, differentially accumulated metabolites were determined by variable importance in project scores (VIP ≥ 1) and fold change ≥ 2 or fold change ≤ 0.5, with VIP values extracted from the OPLS-DA results. The identified differential metabolites were annotated using the Kyoto Encyclopedia of Genes and Genomes (KEGG) compound database (http://www.kegg.jp/kegg/compound), and annotated metabolites were then mapped to the KEGG pathway database (http://www.kegg.jp/kegg/pathway.html). Pathways with significantly regulated metabolites were subjected to metabolite set enrichment analysis, and their significance was determined by hypergeometric test’s p-values.
3. Results
3.1. Global Metabolomic Profile of P. cyrtonema Rhizome
Based on UPLC-MS/MS and the Metware database and publicly available metabolite databases, secondary metabolites in crude P. cyrtonema rhizome (CP group) and processed P. cyrtonema rhizome (SD3, SD6, and SD9 groups) were determined. A total of 755 metabolites were identified from crude and processed rhizome, including 206 alkaloids, 171 phenolic acids, 86 flavonoids, 72 terpenoids, 65 lignans and coumarins, 18 quinones, 13 steroids, and 124 others (Figure 2A). This result indicates that the major secondary metabolites in P. cyrtonema rhizome were polyphenols (phenolic compounds), including phenolic acids and flavonoids, which accounted for 34.04% of the total. Alkaloids, a huge group of low-molecular-weight nitrogen-containing compounds, accounted for 27.28% of the total. The hierarchical clustering heatmap based on UV scaling showed that most secondary metabolites in processed rhizome (SD3, SD6, and SD9 groups) were abundant than those in crude rhizome (CP group), which indicates that the 3–9 processing cycles of steaming and drying led to a significant global accumulation of secondary metabolites (Figure 2B).
UV scaling and PCA were performed on 12 samples to gain insights into the differences in overall metabolites among the four groups and the degree of variability between samples within the same group. The PCA plot showed that the explained variance ratios of PC1 and PC2 were 46.98% and 22.02%, respectively. Three mixed samples as QC were grouped together near the center of the PCA plot, which confirms QC samples with similar metabolic profiles and ensures the stability and repeatability of the entire analysis (Figure 3). In addition, the values of PCC in the correlation analysis show that 12 samples in the same group were close to 1.0 (Figure S1), which indicates that the data from this metabolome analysis were credible and could be used for further analysis. In the PCA plot, we noticed that three replicates within the SD6 group and three replicates within the SD9 group were not clearly separated and nearly clustered into one group, which implies that the secondary metabolites between the SD6 and SD9 groups had no remarkable differences. The initial findings from PCA suggest that, in the course of the traditional processing of P. cyrtonema rhizome, the last 3 cycles of steaming and drying had not induced further significant changes in the abundance of secondary metabolites.
3.2. Differential Accumulated Secondary Metabolite Profiling between Crude and Processed Rhizomes
OPLS-DA is a multivariate statistical analysis method with supervised pattern recognition and can solve the problem that PCA is not sensitive to variables with little correlation. The score plot of each group was generated based on the differential variables to further illustrate the differences between groups. OPLS-DA was performed to reveal the separation among P. cyrtonema rhizome groups and identify significantly differential secondary metabolites between these groups. The OPLS-DA score plots for the pairwise comparisons of SD3 vs. CP, SD6 vs. SD3, and SD9 vs. SD6 are shown in Figure 4. In the three OPLS-DA models, the values of R2Y and Q2 for the comparison groups were closer to 1.0, except for the comparison of SD9 vs. SD6 (Q2=0.83). This result indicates that these models were reliable and suitable for the following screening of differential metabolites.
The significantly differential secondary metabolites were identified as differential accumulated secondary metabolites (DASMs) within each pairwise comparison based on fold change≥2.0 [log2 (2) ≥1.0] or ≤0.5 [log2 (0.5) ≤ −1] and VIP≥1.0 as the screening standards. These DASMs identified from the three comparison groups are displayed in a volcano plot in Figure 5, wherein detailed information on the top 20 DASMs with maximum values of log2 (fold change) in each group is presented in Table 1. A total of 378 DASMs (317 up- and 61 down-regulated) were identified from SD3 vs. CP, 309 DASMs (142 up- and 167 down-regulated) from SD6 vs. SD3, and 116 DASMs (12 up- and 104 down-regulated) from SD9 vs. SD6 (Figure 6). Among the top 20 DASMs from each comparison group, all from SD3 vs. CP were up-regulated, only 9 from SD6 vs. SD3 were up-regulated, and none from SD9 vs. SD6 were up-regulated. In addition, some unique DASMs including newly added and disappeared DASMs were found in the three comparison groups. In detail, in the SD3 vs. CP group, 164 newly added DASMs and 3 disappeared DASMs were observed in SD3 and CP, respectively; in the SD6 vs. SD3 group, 16 newly added DASMs and 33 disappeared DASMs were found in SD6 and SD3, respectively; and in the SD9 vs. SD6 group, 4 newly added DASMs and 30 disappeared DASMs were detected in SD9 and SD6, respectively (Table 2 and Table S1).
A large quantity of DASMs were identified from SD3 vs. CP and SD6 vs. SD3, while only a small quantity was observed from SD9 vs. SD6. This finding indicates that drastic changes occurred in the abundance of secondary metabolites from crude P. cyrtonema rhizome to processed rhizome subjected to the first 6 processing cycles, and the last 3 processing cycles did not induce further significant changes. Moreover, these newly added and disappeared secondary metabolites suggest complex chemical changes in the processed rhizome, including the formation of numerous novel secondary metabolites mainly occurring in the first 3 processing cycles and a small number of original secondary metabolites decomposing mainly occurring in the last 6–9 processing cycles. This result from DASM analysis is consistent with that from PCA and further suggests that the traditional processing method produces a series of chemical changes in P. cyrtonema rhizome. This phenomenon could make the processed rhizome significantly different from the crude rhizome in nutritional and healthy values.
3.3. Variation Patterns in the Relative Abundance of Secondary Metabolites
The secondary metabolites in 12 P. cyrtonema rhizome samples were subjected to K-means clustering analysis to understand the variation patterns in their relative abundance. After standardization by UV scaling, 557 secondary metabolites were clustered into 7 variation patterns according to their changes in relative abundance during processing (Figure 7 and Table 3). The clustering analysis showed that the relative abundance of most secondary metabolites (420, 75.4%) clustering in variation patterns 4–7 increased in SD3 samples unlike in CP samples. Only 137 DASMs (24.6%) clustering in variation patterns 1–3 decreased in SD3 samples unlike in CP samples. Therefore, for most secondary metabolites in crude P. cyrtonema rhizome, the first 3 processing cycles of steaming and drying significantly enhanced their accumulation in relative abundance. However, the first 3 processing cycles also induced a certain level of inhibition in a small number of secondary metabolites, which led to a significant decrease in their relative abundance.
In all the 557 secondary metabolites with 7 different variation patterns, most exhibited 4 change trends in relative abundance during processing. Among them, 34.11% presented as continuously increasing from CP to SD9, which mainly consisted of alkaloids, phenolic acids, and terpenoids (VP6); 23.69% showed a continuous decrease from CP to SD9, which primarily comprised alkaloids, phenolic acids, and flavonoids (VP2 and VP3); 24.96% demonstrated a trend of first increasing from CP to SD3 and then decreasing from SD3 to SD9, which mainly included alkaloids, phenolic acids, and flavonoids (VP4 and VP7); and 16.34% exhibited a trend of first significantly increasing from CP to SD3, then slightly rising from SD3 to SD6, and last slightly decreasing from SD6 to SD9, which primarily consisted of alkaloids and phenolic acids (VP5). Therefore, alkaloids, phenolic acids, and flavonoids were the main secondary metabolites with diversified variation patterns during the 9 processing cycles. In addition, nearly half of the terpenoid compounds, which were mainly ditepenoids, presented a continuously increasing trend from CP to SD9, which indicates that continuous processing from 1–9 cycles led to continuous accumulation in their relative abundance.
3.4. Annotation and Functional Classification of Secondary Metabolites
Enrichment analysis of KEGG pathways was performed based on all the identified secondary metabolites to clarify which secondary metabolic pathways in P. cyrtonema rhizome were significantly affected by the 9 processing cycles of steaming and drying. A total of 289 secondary metabolites were mapped to the KEGG database categories and annotated in metabolic pathways, with 94 from SD3 vs. CP, 99 from SD6 vs. SD3, and 96 from SD9 vs. SD6 being annotated to 38, 29, and 22 significantly enriched pathways, respectively. Among all the 289 annotated secondary metabolites, 125 were DASMs, including 63 from SD3 vs. CP, 34 from SD6 vs. SD3, and 28 from SD9 vs. SD6 (Table S2). The top 20 significantly enriched KEGG pathways in the 3 comparison groups are shown in Figure 8, most of which were related to the biosynthesis of various secondary metabolites, including alkaloids, flavonoids, and terpenoids, as well as the metabolisms of many kinds of amino acids, such as lysine degradation and biosynthesis of amino acids (Table 4). Meanwhile, most DASMs involved in these pathways were up-regulated after the first 3 processing cycles. However, after the 6 processing cycles, nearly half of the DASMs involved in the biosynthesis of various secondary metabolites and most of the DASMs involved in the metabolisms of amino acids were down-regulated. After the last 9 processing cycles, nearly all of DASMs enriched in these pathways were down-regulated. The KEGG enrichment analysis supports the results obtained from DASMs and variation pattern analysis, which indicates that the first 3 processing cycles significantly enhanced the biosynthesis of various secondary metabolites. However, these enhancing effects were gradually reversed by the continuous 6–9 processing cycles. Meanwhile, the processing significantly affected the metabolisms of many kinds of amino acids, which suggests significant changes in the biosynthesis or degradation of amino acids.
3.5. Processing-Induced Changes in the Functional Components of P. cyrtonema Rhizome
Alkaloids, phenolic acids, flavonoids, terpenoids, and steroids were selected to determine processing-induced changes in the key functional components of P. cyrtonema rhizome, and changes in their relative abundance during processing were further analyzed. The results showed that continuous processing of steaming and drying induced global changes (up- and down-regulated) in the relative abundance of these functional components. As for alkaloids and phenolic acids (Figure 9), some of them were up- or down-regulated in relative abundance after the 3 processing cycles. However, with increased processing cycles from 6–9, the number of up-regulated metabolites decreased gradually, and down-regulated metabolites increased gradually. With regard to flavonoids, terpenoids, and steroids (Figure 9), some of them were up-regulated in relative abundance after the 3 processing cycles. Similarly, with increased processing cycles from 6–9, the number of up-regulated metabolites decreased gradually, and down-regulated metabolites began to occur. Among these regulated functional components, the most abundant were alkaloid, phenolamine and plumerane belonging to the alkaloid class; flavones, flavonols and flavonoids belonging to the flavonoid class; monoterpenoids belonging to the terpenoid class; and steroidal saponins belonging to the steroid class. This result suggests that processing for a short period can significantly induce enhancing effect, which will promote the biosynthesis of functional components in P. cyrtonema rhizome. However, these enhancing effects will unavoidably be reversed or weakened under continuous processing for a long period. Accordingly, the processing period of steaming and drying should be reduced to 3–6 cycles to maintain enhancing effects on the accumulation of functional components. In addition, different functional components, even if belonging to the same class, have different response mechanisms toward processing, which are presented as up-regulated, down-regulated, or stable in their relative abundance.
4. Discussion
Processing often leads to the structural transformation of herbal components. During processing, herbal components may undergo oxidation, decomposition, isomerization, hydrolysis, and/or reaction with other constituents to eventually form novel compounds [32]. Some herbal medicines, such as Aconitum root, Ginseng Radix et Rhizome, and Rhei Radix et Rhizoma, have been demonstrated to possess distinct chemical profiles after processing and showed reduced toxicity or altered therapeutic activities [9]. Understanding these complicated processing mechanisms provides a theoretical and metabolic data basis for improving processing techniques and for the pharmacological study of P. cyrtonema.
In the processed rhizome, we found numerous newly added secondary metabolites and a small number of disappeared secondary metabolites, which might result from the formation of novel secondary metabolites and the decomposition of the original secondary metabolites induced by processing. Liang et al. found that, with an increase in processing time, drastic changes occurred in the chemical components of P. cyrtonema rhizome, such as component isomerization, decreased type and content of primary glycosides, and increased content of aglycones [33]. Therefore, besides the increased contents of functional components, the structural transformation of secondary metabolites in crude P. cyrtonema rhizome might also be an important processing mechanism related to the enhancing effect. In this mechanism, the synthesis of novel compounds and decomposition of original secondary metabolites did not occur synergistically: one mainly in the first 3 processing cycles, and the other mainly in the last 6–9 processing cycles.
Apart from the structural transformation of components, directly reducing the contents of toxic components is also a main processing mechanism in Chinese herbal medicines [9]. Alkaloids are one of the largest groups of plant secondary metabolites, present in several economically relevant plant families [34]. Extracts from alkaloid-containing plants have been used to treat several ailments such as fever, snakebite, and insanity. However, despite their significant benefits to health and pharmaceutical industries, the toxicity of some types of plant alkaloids, such as pyrrolizidine, tropane, piperidine, indolizidine, and steroidal alkaloids, has been observed in animals and humans [35,36]. Alkaloid compounds were found to be the most abundant secondary metabolites in P. cyrtonema rhizome. Among the 206 alkaloids identified, 20 of them were regarded as potential toxic alkaloids according to the above mentioned reported literature, including 7 piperidine alkaloids, 7 pyridine alkaloids, 2 isoquinoline alkaloids, 2 tropan alkaloids, and 2 steroidal alkaloids. Continuous processing did not lead to the complete decomposition of these alkaloids, and only the relative abundance of piperidine and 6-Deoxyfagomine (piperidine alkaloids) in rhizomes processed for 6 cycles were significantly decreased unlike in crude rhizomes (vp 3 in Figure 7). Although the toxicity of crude P. cyrtonema rhizome and related toxic components in it were unclear, our present study suggested that reducing the contents of potential toxic alkaloids and decomposing toxic alkaloids may not be the main mechanisms for the detoxification of P. cyrtonema rhizome. Therefore, some chemical changes or physically structural alterations may have occurred during processing, which were related to the detoxification of P. cyrtonema rhizome.
Flavonoids comprise a large and diverse group of polyphenolic compounds with antioxidant, hypoglycemic, hypolipidemic, and anticancer properties [37]. Flavonoids and their glycosides are important bioactive constituents of the rhizomes of Polygonatum plants. Currently, 61 flavonoids have been isolated from Polygonatum plants, with 13 of them identified in P. cyrtonema [4]. The stability of different flavonoids, such as flavanones, flavonols, and flavones, is influenced not only by the processing but also by the flavonoid structure [38]. A total of 86 flavonoids were identified in this study, which were mainly flavones, flavonols, isoflavones, chalcones, and other types of flavonoids. Continuous processing induced diversified variation patterns in the relative abundance of different flavonoids, as well as the synthesis of novel secondary metabolites and the decomposition of original secondary metabolites. The change profiles observed in most flavonoids suggest that 3–6 processing cycles can enhance efficacy by increasing their contents and inducing the synthesis of 85% newly added secondary metabolites. Continuous processing until 9 cycles will unavoidably weaken the enhancing effect by decreasing the increased contents during 3–6 processing cycles and further decomposing more flavonoids.
Phenolic acids, which are a subclass of plant phenolics, exhibit tremendous antioxidant activity and protective effects, including antimicrobial, anticancer, anti-inflammatory, and anti-mutagenic [39]. As the main secondary metabolites rich in P. cyrtonema rhizome, the variation patterns exhibited by most of them suggest that 3 processing cycles induced their significant accumulation and the addition of 91% newly formed secondary metabolites. Saponins in medicinal plants exhibit essential medicinal properties, such as anti-inflammatory, antiviral, insecticidal, and anticancer actions [40]. Saponins are primarily classified into steroid and triterpenoid saponins based on the structure of the hydrophobic aglycone unit [41]. As the primary active ingredient, Polygonatum plants are rich in steroidal saponins, and 10 steroidal saponins have been isolated and identified from P. cyrtonema to date [42]. A total of 13 steroidal saponins and 3 triterpene saponins were identified in this study. The variation patterns suggest that 3 processing cycles were sufficient to increase the contents of saponins and induce 100% newly added secondary metabolites. Conversely, continuous processing for more than 3 cycles will unavoidably weaken the enhancing effects induced by the first 3 cycles. In addition, saponins might have higher stability than other secondary metabolites in P. cyrtonema rhizome because the decomposition of only 1 steroidal saponin in crude rhizome was found during the entire processing. A total of 17 kinds of amino acids were detected in crude Polygonatum rhizome, which were important flavor and nutritional components [43]. However, the effects of traditional processing on amino acid composition, contents, and nutritional value of Polygonatum plants have not been fully understood to date. KEGG enrichment analysis suggests that processing has significant effects on amino acid metabolisms. Therefore, in future work, changes in amino acid metabolisms during processing will be explored to further provide metabolic data support for the nutritional and healthy values of processed P. cyrtonema rhizome from the aspect of amino acid nutrients.
Our present study reveals complicated changes in secondary metabolites of P. cyrtonema rhizome during traditional processing with “nine-steaming and nine-drying.” Undoubtedly, processing produces beneficial effects on reinforcing efficacy by promoting the accumulation of functional components and on reducing toxicity to a certain extent by decreasing the contents of toxic components, which is meaningful for edible safety and increasing the medicinal values of P. cyrtonema rhizome. However, processing also brings adverse effects given that it can significantly decrease the contents of a small number of functional components in the rhizome, even after only 3 cycles. Apart from these functional components belonging to secondary metabolites, polysaccharides are also the main functional component in P. cyrtonema rhizome. Previous studies have found the highest content of polysaccharide in crude rhizome, with decreasing content in processed rhizome with increasing processing cycles, and 4 processing cycles may be the ideal processing method for Polygonatum to retain more active ingredients [14,44]. Based on the comprehensive quality evaluation index, 5 processing cycles were regarded as a turning point with the highest index [11]. Therefore, achieving the purpose of reinforcing efficacy, reducing toxicity by processing, and preserving functional components as much as possible in processed P. cyrtonema rhizome should be comprehensively considered. Thus, we propose that the traditional processing method should be improved in the processing period. Furthermore, the 9 processing cycles of steaming and drying used in P. cyrtonema rhizome should be reduced to 3–6 cycles.
Despite the extensive use of processed Chinese herbal medicines, the underlying scientific mechanism of processing remain unclear for most of them, including Polygonatum plants to date. During processing with heating and drying, complicated changes in functional components of P. cyrtonema rhizome may occur not only in regulated contents but also in formed novel compounds. In many cases, the contents and structures of components may be altered simultaneously. Future studies should be devoted to comprehensively elucidating the processing-induced scientific mechanisms of reinforcing efficacy and reducing toxicity. Thus, additional efforts should be made to investigate the association of chemical and pharmacological changes using advanced technologies and assess the contribution of processing-induced chemical alteration to the changed bioactivities of P. cyrtonema. Furthermore, a unified and scientific processing technology should be established in the future to improve the traditional processing method used in P. cyrtonema.
5. Conclusions
This study applied a comparative metabolomics strategy to reveal changes in secondary metabolites during the processing of Polygonatum cyrtonema rhizome. The rhizome samples underwent traditional processing involving “nine cycles of steaming and sun-drying.” After being subjected to 3, 6, and 9 processing cycles, the extracts from 12 processed rhizome samples were used for the qualitative and quantitative analysis of secondary metabolites. The results are as obtained as follows:
1) The major secondary metabolites in P. cyrtonema rhizome were polyphenols (phenolic compounds), including phenolic acids and flavonoids, followed by alkaloids, terpenoids, lignans, and coumarins, and finally quinones and steroids.
2) Processing significantly increased the global accumulation of secondary metabolites in processed rhizome unlike in unprocessed crude rhizome. In all 9 processing cycles, drastic changes in the relative abundance of secondary metabolites from crude rhizome to processed rhizome occurred after the first 6 cycles, and the last 3 cycles did not induce further significant changes. For most secondary metabolites, the first 3 processing cycles significantly enhanced their accumulation in relative abundance, while significantly decreasing their accumulation in a small number of secondary metabolites. Processing also led to numerous newly added secondary metabolites mainly occurring in the first 3 cycles, with a small number of disappeared secondary metabolites mainly occurring in the last 6–9 cycles.
3) Processing induced some significantly enriched KEGG pathways, in which the first 3 processing cycles enhanced the biosynthesis of various secondary metabolites and had significant effects on amino acid metabolisms.
4) Processing greatly influenced the relative abundance of functional components, and a short period of processing was helpful for enhancing their biosynthesis in P. cyrtonema rhizome. However, the enhancing effects will unavoidably be reversed or weakened under continuous processing for a long period. Therefore, the processing period of steaming and drying should be reduced to 3–6 cycles to maintain an enhancing effect on the accumulation of functional components.
5) Different functional components, even if belonging to the same class, had different response mechanisms toward processing, which were presented as up-regulated, down-regulated, or stable in relative abundance. 6) Future studies should be devoted to comprehensively elucidating the processing-induced scientific mechanisms of reinforcing efficacy and reducing toxicity. Furthermore, a unified and scientific processing technology should be established in the future to improve the traditional processing method used in P. cyrtonema.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Correlation analysis of secondary metabolites from 12 P. cyrtonema samples. The shades of color indicate higher or lower Pearson’s Correlation Coefficient, with redder representing higher and greener representing lower; Table S1: Newly added and disappeared secondary metabolites in three comparison groups; Table S2: KEGG pathways and annotated compounds in three comparison groups.
Author Contributions
Conceptualization, H.L. and B.Y.; methodology, W.P.; software, W.P.; validation, H.L., B.Y. and X.S.; formal analysis, W.P.; investigation, Q.S.; resources, Y.Y.; data curation, H.L.; writing—original draft preparation, J.S.; writing—review and editing, X.S.; visualization, J.S.; supervision, B.Y.; project administration, H.L.; funding acquisition, Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by The Science and Technology Planning Project of Suichang County (Grant No. 2022HZ05).
Data Availability Statement
Data is contained within the article or Supplementary Material.
Acknowledgments
The authors appreciate MetWare corporation (Wuhan, China) for providing the widely targeted metabolomic analysis.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Chen, X.; Tamura, M.N. Flora of China; Science Press: Beijing, China, 2000; Volume 24, pp. 223–232. [Google Scholar]
- National Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; Volume 4, pp. 387–388. [Google Scholar]
- Zhang, J.; Ma, B.; Yang, Y.; Sun, G. Development of steroidal saponins and pharmacologic activity from plants of Polygonatun genus (in Chinese). Chin. Pharm. J. 2006, 41, 330–332. [Google Scholar]
- Zhang, J.; Wang, Y.; Yang, W.; Yang, M.; Zhang, J. Research progress in chemical constituents in plants of Polygonatum and their pharmacological effects (in Chinese). China J. Chin. Mater. Med. 2019, 44, 1989–2008. [Google Scholar] [CrossRef]
- Bai, J.B.; Ge, J.C.; Zhang, W.J.; Liu, W.; Luo, J.P.; Xu, F.Q.; Wu, D.L.; Xie, S.Z. Physicochemical, morpho-structural, and biological characterization of polysaccharides from three Polygonatum spp. RSC. Adv. 2021, 11, 37952–37965. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhao, C.; Li, X.; Gao, Q.; Huang, L.; Xiao, P.; Gao, W. The genus Polygonatum: a review of ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 214, 274–291. [Google Scholar] [CrossRef]
- Shen, W.D.; Li, X.Y.; Deng, Y.Y.; Zha, X.Q.; Pan, L.H.; Li, Q.M.; Luo, J.P. Polygonatum cyrtonema Hua. polysaccharide exhibits anti-fatigue activity via regulating osteocalcin signaling. Int. J. Biol. Macromol. 2021, 175, 235–241. [Google Scholar] [CrossRef]
- Sheridan, H.; Kopp, B.; Krenn, L.; Guo, D.; Sendker, J. Traditional chinese herbal medicinal preparation: invoking the butterfly effect. Science. 2015, 350, S64–6. [Google Scholar]
- Wu, X.; Wang, S.; Lu, J.; Jing, Y.; Li, M.; Cao, J.; Bian, B.; Hu, C. Seeing the unseen of Chinese herbal medicine processing (Paozhi): advances in new perspectives. Chin. Med. 2018, 13, 4. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, L.; Zhao, Q.; Bao, K.; Jiang, C. Research progress on Polygonatum cyrtonema processed by nine times steaming and nine times shining (in Chinese). Chin. Tradit. Herb. Drugs. 2020, 51, 5631–5637. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, C.; Jin, C.; Liu, J.; Liu, C.; Li., L. Research on relationship between processing degree and internal and external quality of Polygonatum cyrtonema processed by “nine-steaming and nine-suncuring” based on color change (in Chinese). Chin. Tradit. Herb. Drugs. 2022, 53, 1719–1729. [Google Scholar] [CrossRef]
- Xu, R.; Liang, J.; Yu, N.; Wu, H.; Wu, Z.; Zhou, A. Structural change of polysaccharide constituents from Polygonatum cyrtonema in Jiuhua Mountain after processing (in Chinese). J. Anhui. Univ. Chin. Med. 2021, 40, 91–96. [Google Scholar]
- Guo, T.; Wang, R.; Song, Y.; Ye, J.; Jiao, H.; Yu, D.; Chang, H. Chemical components dynamic variation of root of Polygonati rhizoma during a nine-time repeat of the steaming and sundrying process (in Chinese). J. Pharm. Res. 2022, 41, 220–229. [Google Scholar] [CrossRef]
- Yao, X.; Deng, Z.; Li, H.; Zhang, B. Effect of processing cycles on the composition of Polygonatum cyrtonema Hua during nine-steam-nine-bask processing. Food. Biosci. 2022, 102081, 1–11. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism-from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A. Natural products and plant disease resistance. Nature. 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K.M.; Inze, D. Plant cell factories in the post-genomic era: new ways to produce designer secondary metabolites. Trends. Plant. Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Kutchan, T.; Dixon, R.A. Physiology and metabolism-Secondary metabolism: Nature’s chemical reservoir under deconvolution. Curr. Opin. Plant Biol. 2005, 8, 227–229. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Pinto, D.C.G.A. Biological potential and medical use of secondary. Metabolites. 2019, 6, 66. [Google Scholar] [CrossRef]
- Yang, D.; Du, X.; Yang, Z.; Liang, Z.; Guo, Z.; Liu, Y. Transcriptomics, proteomics, and metabolomics to reveal mechanisms underlying plant secondary metabolism. Eng. Life Sci. 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics-the link between genotypes and phenotypes. Plant. Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Hall, R.; Beale, M.; Fiehn, O.; Hardy, N.; Sumner, L.; Bino, R. Plant metabolomics: The missing link in functional genomics strategies. Plant Cell. 2002, 14, 1437–1440. [Google Scholar] [CrossRef]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; Trethewey, R.N.; Lange, B.M.; Wurtele, E.S.; Sumner, L.W. Potential of metabolomics as a functional genomics tool. Trends. Plant. Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef]
- Saito, K. Phytochemical genomics-a new trend. Curr. Opin. Plant. Biol 2013, 16, 370–380. [Google Scholar] [CrossRef] [PubMed]
- La, G.; Hao, X.; Li, X.; Ou, M.; Yang, T. Application of metabolomics in plant research (in Chinese). Bot. Res. 2016, 5, 26–33. [Google Scholar]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: application in the study of rice metabolomics. Mol. Plant. 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Cao, X. Rewiring of the fruit metabolome in tomato breeding. Cell 2018, 172, 249–261.e12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qiu, X.; Tan, Q.; Xiao, Q.; Mei, S. A comparative metabolomics study of flavonoids in Radish with different skin and flesh colors (Raphanus sativus L.). J. Agric. Food. Chem. 2020, 68, 14463–14470. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, Y.; Liu, C.; Shi, L.; Li, J.; Zeng, Y.; Guo, L.; Wang, S. Identification of medicinal compounds of Fagopyri dibotryis rhizome from different origins and its varieties using UPLC-MS/MS based metabolomics. Metabolites. 2022, 12, 790. [Google Scholar] [CrossRef]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome. Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Qin, K.; Hao, W.; Hao, C.; Lu, T.; Zhang, X. Chemical mechanism during Chinese medicine processing. Prog Chem. 2012, 77, 637–649. [Google Scholar] [CrossRef]
- Liang, Z.; Pan, Y.; Qiu, L.; Wu, X.; Xu, X.; Shu, Y.; Yuan, Q. Analysis on chemical components changes of Polygonati Rhizoma in processing of nine times steaming and nine times sunning by UPLC-Q-TOF-MS/MS (in Chinese). Chin. Tradit. Herb. Drugs. 2022, 53, 4948–4957. [Google Scholar] [CrossRef]
- Matsuura, H.N.; Fett-Neto, A.G. Plant alkaloids: main features, toxicity, and mechanisms of action. In Plant Toxins; Gopalakrishnakone, P., Carlini, C., Ligabue-Braun, R., Eds.; Toxinology; Springer press: Dordrecht, The Netherland, 2015; pp. 1–15. [Google Scholar] [CrossRef]
- Adibah, K.Z.M.; Azzreena, M.A. Plant toxins: alkaloids and their toxicities. GSC Biol. Pharm. Sci 2019, 6, 21–29. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Panter, K.E. Alkaloids toxic to livestock. The Alkaloids. 2009, 67, 143–216. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, M.; Silakari, O. Flavones, an important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef] [PubMed]
- Chaabana, H.; Ioannoua, I.; Parisb, C.; Charbonnela, C.; Ghoula, M. The photostability of flavanones, flavonols and flavones and evolution of their antioxidant activity. J. Photochem. Photobiol. A: Chemistry. 2017, 336, 131–139. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- El Aziz, M.M.A.; Ashour, A.S.; Melad, A.S.G. A review on saponins from medicinal plants: chemistry, isolation, and determination. J. Nanomed. Res. 2019, 7, 282–288. [Google Scholar] [CrossRef]
- Kavya, N.M.; Adil, L.; Senthilkumar, P. A review on saponin biosynthesis and its transcriptomic resources in medicinal plants. Plant. Mol. Biol. Rep. 2021, 39, 833–840. [Google Scholar] [CrossRef]
- Shi, Y.; Si, D.; Chen, D.; Zhang, X.; Han, Z.; Yu, Q.; Liu, J.; Si, J. Bioactive compounds from Polygonatum genus as anti-diabetic agents with future perspectives. Food Chem. 2023, 408, 135183. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Huang, Z.; Zhou, D.; Lu, Y.; Tang, Q.; Zou, H.; Shi, X.; Xie, H.; Zeng, J.; Zheng, Y. Effect of processing on amino acid composition and nutritional value of protein from Polygonatum (in Chinese). Food. Ferment. Ind. 2023, 7. [Google Scholar] [CrossRef]
- Wu, W.; Huang, N.; Huang, J.; Wang, L.; Wu, L.; Wang, Q.; Zhao, H. Effects of the steaming process on the structural properties and immunological activities of polysaccharides from Polygonatum cyrtonema. J. Func. Foods. 2022, 88, 104866. [Google Scholar] [CrossRef]
Figure 1.
The P. cyrtonema rhizome samples during 9 processing cycles of steaming and drying. CP represents crude rhizome samples and SD3, SD6 and SD9 represent processed samples subjected to 3, 6 and 9 processing cycles, respectively.
Figure 1.
The P. cyrtonema rhizome samples during 9 processing cycles of steaming and drying. CP represents crude rhizome samples and SD3, SD6 and SD9 represent processed samples subjected to 3, 6 and 9 processing cycles, respectively.

Figure 2.
Basic information of secondary metabolites detected in P. cyrtonema rhizome. (A) quantity and classification of secondary metabolites. (B) clustered heatmap of secondary metabolites in crude rhizome samples (CP group) and in processed rhizome samples (SD3, SD6 and SD9 groups). The shades of color indicate more or less metabolites, with redder representing more and greener representing less.
Figure 2.
Basic information of secondary metabolites detected in P. cyrtonema rhizome. (A) quantity and classification of secondary metabolites. (B) clustered heatmap of secondary metabolites in crude rhizome samples (CP group) and in processed rhizome samples (SD3, SD6 and SD9 groups). The shades of color indicate more or less metabolites, with redder representing more and greener representing less.
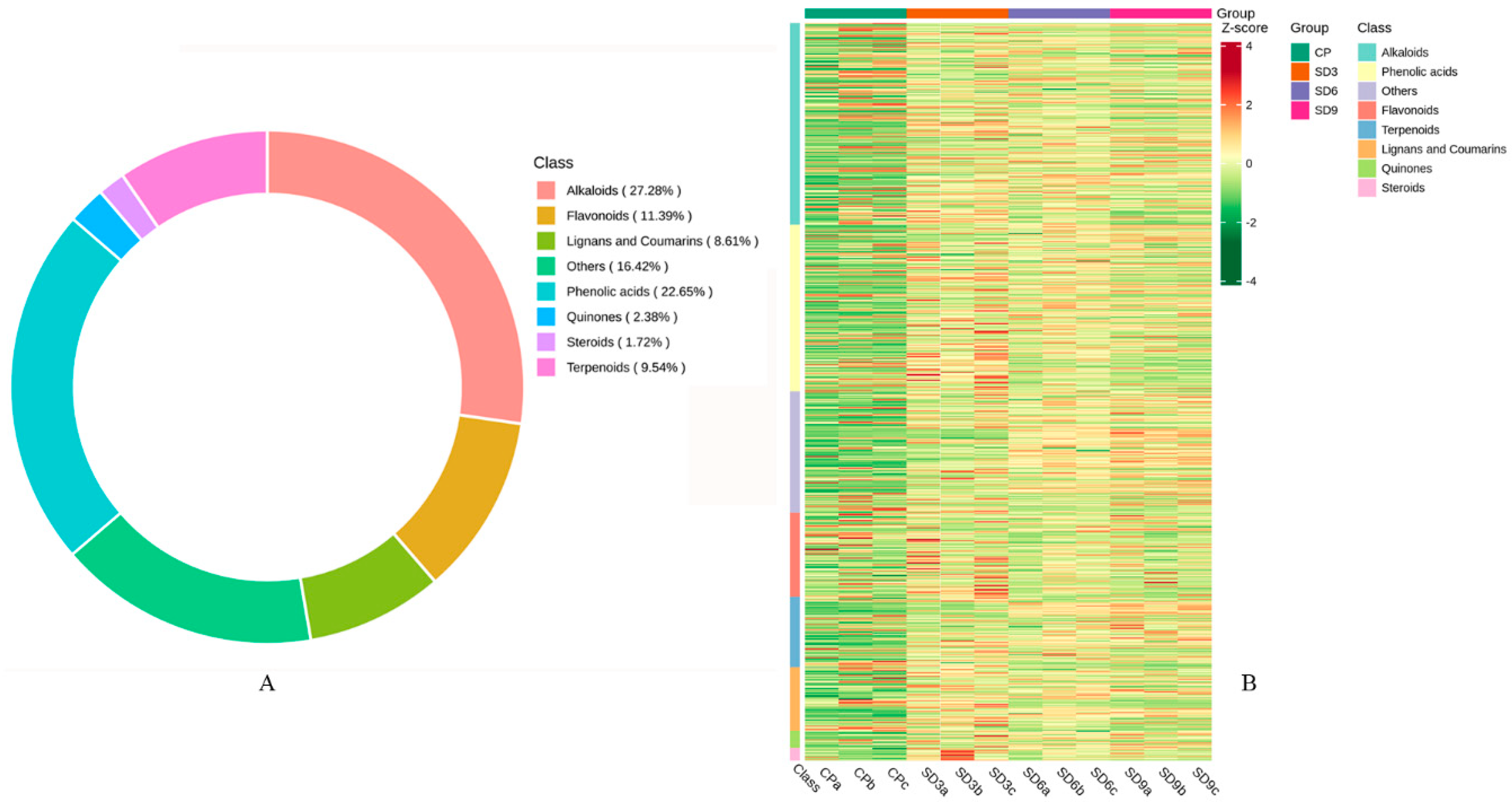
Figure 3.
Principal component analysis (PCA) plot of secondary metabolites from 12 P. cyrtonema samples. The figure in brackets refer to the explained variance ratio.
Figure 3.
Principal component analysis (PCA) plot of secondary metabolites from 12 P. cyrtonema samples. The figure in brackets refer to the explained variance ratio.
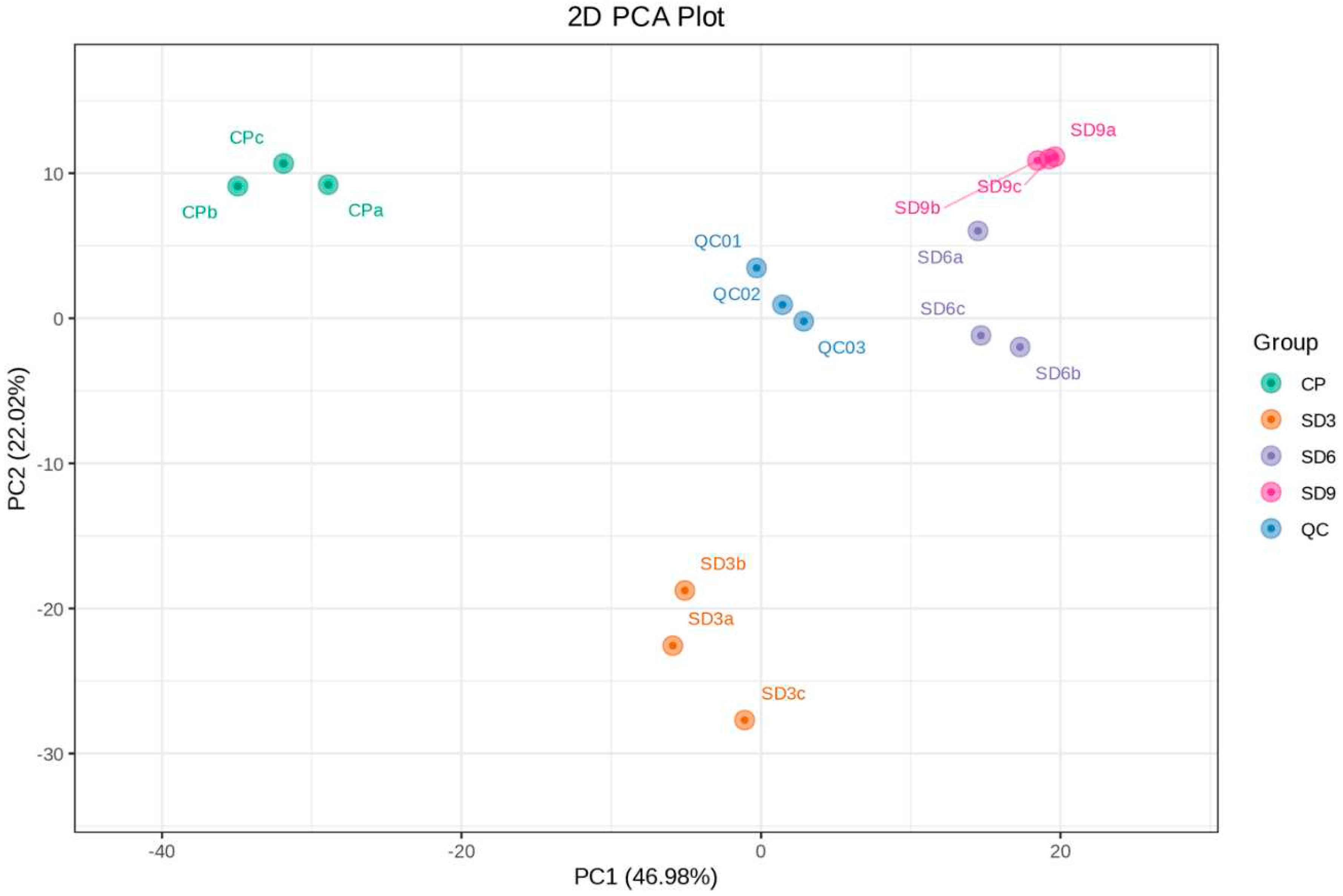
Figure 4.
The OPLS-DA score plots of the pairwise comparison on SD3 vs. CP, SD6 vs. SD3 and SD9 vs. SD6.
Figure 4.
The OPLS-DA score plots of the pairwise comparison on SD3 vs. CP, SD6 vs. SD3 and SD9 vs. SD6.
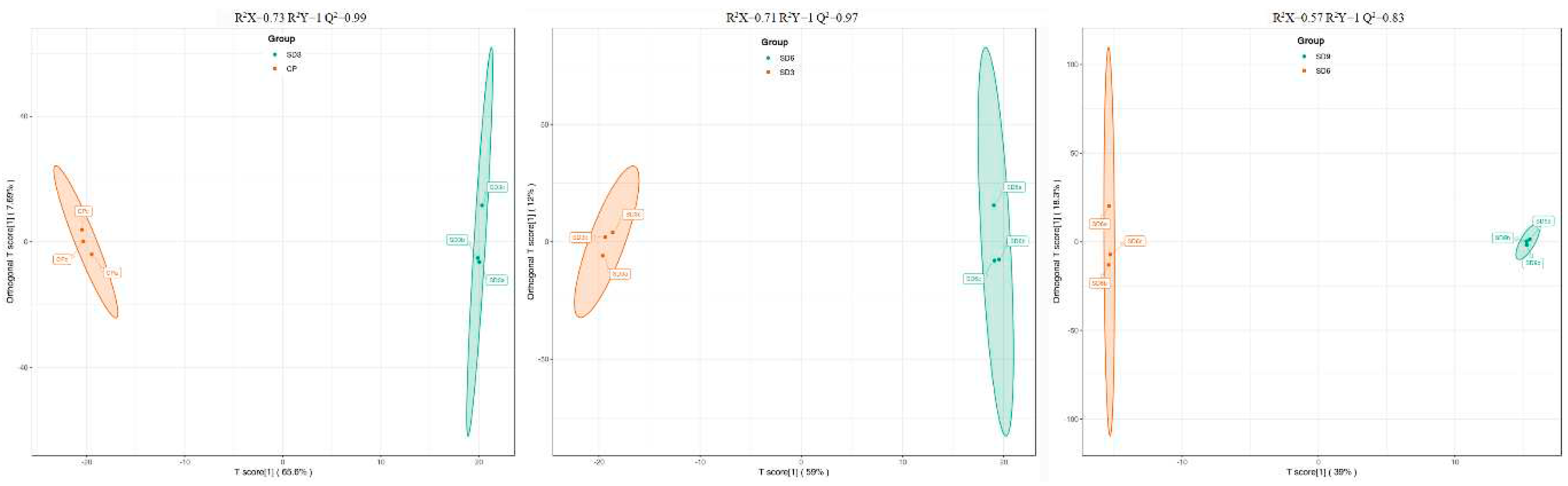
Figure 5.
Volcano plots of differential accumulated secondary metabolites. The dots in plots represent differential accumulated secondary metabolites, with red representing up-regulated, green representing down-regulated, and gray indicating insignificant.
Figure 5.
Volcano plots of differential accumulated secondary metabolites. The dots in plots represent differential accumulated secondary metabolites, with red representing up-regulated, green representing down-regulated, and gray indicating insignificant.
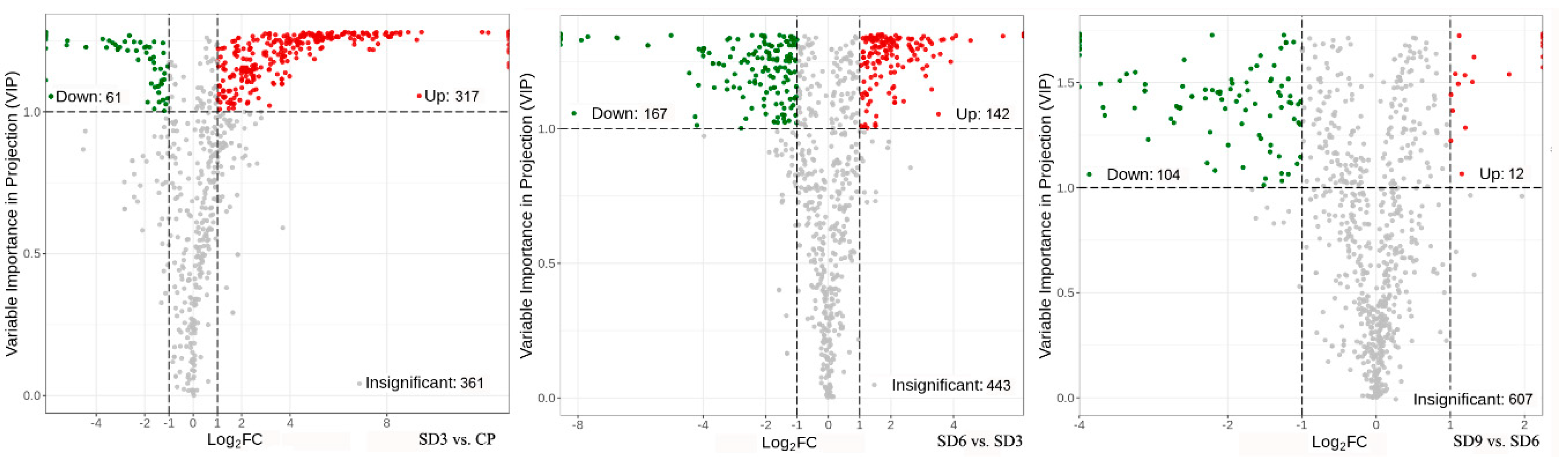
Figure 6.
Quantity and class of differential accumulated secondary metabolites from three comparison groups. (A) total quantity of differential accumulated secondary metabolites. (B) specific quantity of differential accumulated secondary metabolites within each class.
Figure 6.
Quantity and class of differential accumulated secondary metabolites from three comparison groups. (A) total quantity of differential accumulated secondary metabolites. (B) specific quantity of differential accumulated secondary metabolites within each class.
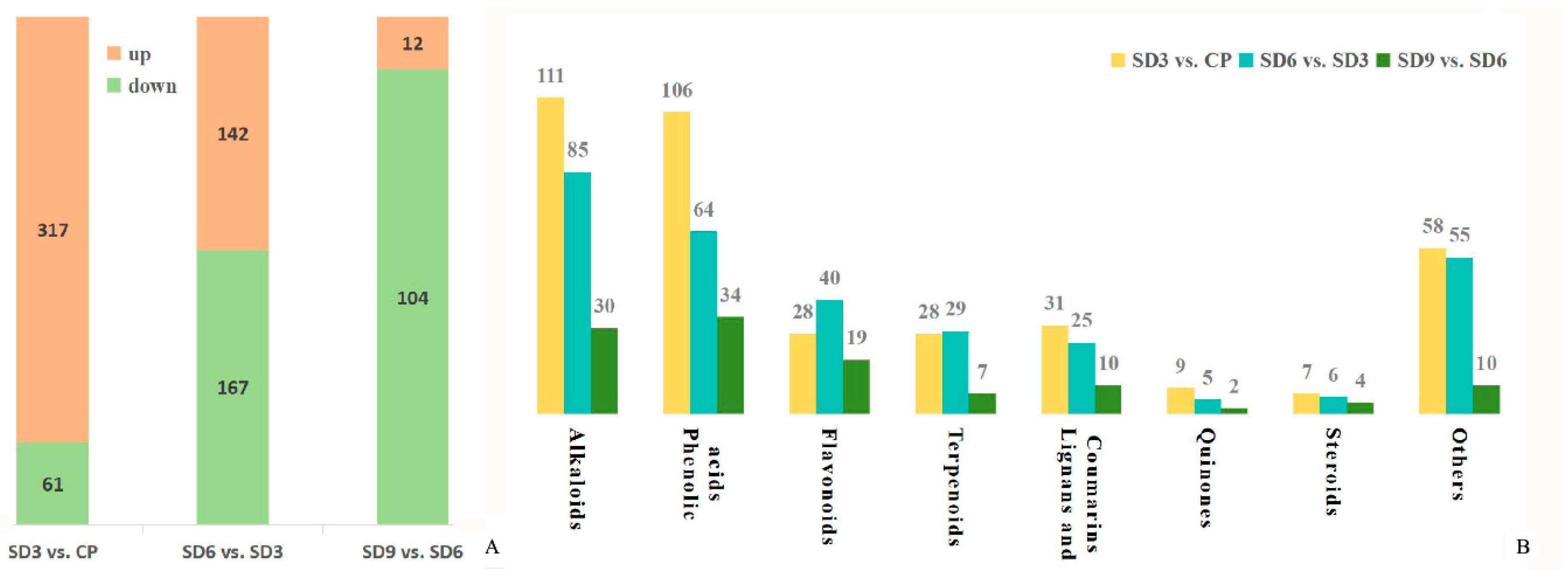
Figure 7.
K-Means clustering analysis of secondary metabolites. The number represent the total quantity of secondary metabolites with same variation pattern.
Figure 7.
K-Means clustering analysis of secondary metabolites. The number represent the total quantity of secondary metabolites with same variation pattern.
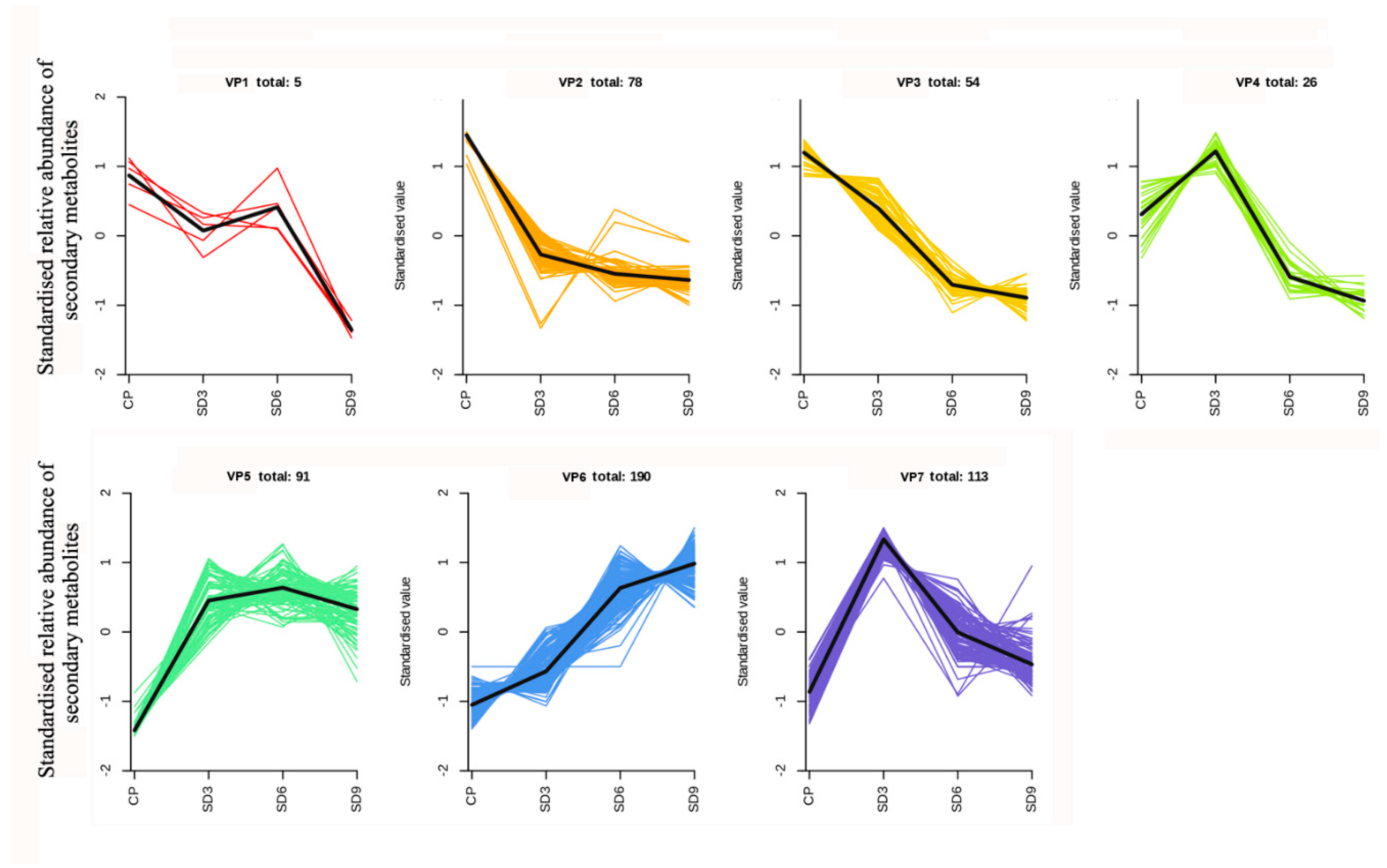
Figure 8.
The top 20 significantly enriched KEGG pathways. The colour of dots in plots refer to higher or lower P-value, with bluer representing higher, redder representing lower. The size of a dot represents more or less secondary metabolites, with bigger representing more and smaller representing less.
Figure 8.
The top 20 significantly enriched KEGG pathways. The colour of dots in plots refer to higher or lower P-value, with bluer representing higher, redder representing lower. The size of a dot represents more or less secondary metabolites, with bigger representing more and smaller representing less.
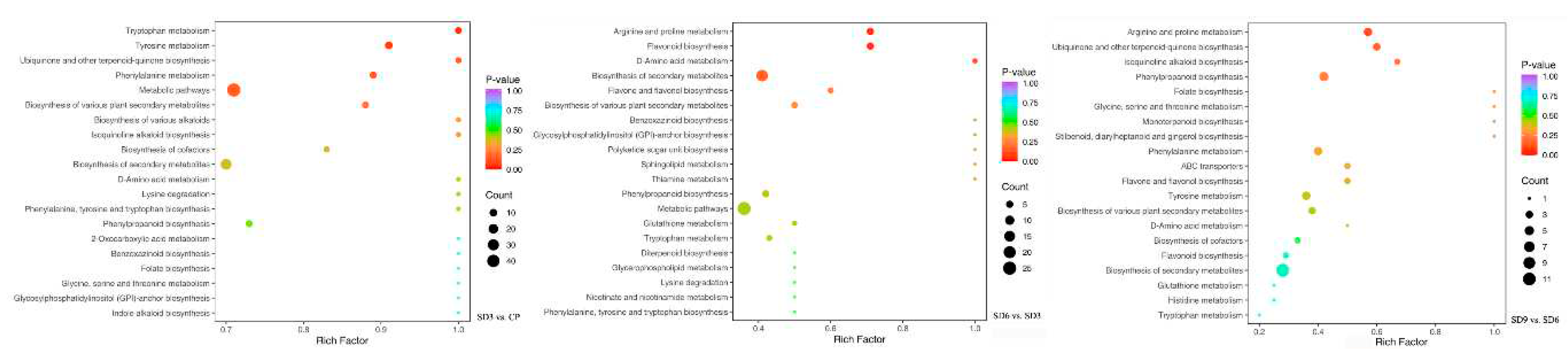
Figure 9.
Changes in relative abundance of functional components in P. cyrtonema rhizome during 9 processing cycles. The shades of color and arrows represent up- or down-regulated components, with orange and ↑ representing up-regulated, green and ↓ representing down-regulated, and gray representing unchanged. The number represent the quantity of up- or down-regulated and unchanged components.
Figure 9.
Changes in relative abundance of functional components in P. cyrtonema rhizome during 9 processing cycles. The shades of color and arrows represent up- or down-regulated components, with orange and ↑ representing up-regulated, green and ↓ representing down-regulated, and gray representing unchanged. The number represent the quantity of up- or down-regulated and unchanged components.


Table 1.
The detail information of top 20 differential accumulated secondary metabolites with maximum values of Log2FC.
Table 1.
The detail information of top 20 differential accumulated secondary metabolites with maximum values of Log2FC.
| Class | DASM//Log2FC | ||
|---|---|---|---|
| SD3 vs. CP | SD6 vs. SD3 | SD9 vs.SD6 | |
| Alkaloids | Cephalanthrin A//12.57 1-Acetyl-β-carboline//8.98 (R)-1,2,3,4-Tetrahydro-3-carboxy-2-carboline//8.52 N-benzoyl-2-aminoethyl-β-D-glucopyranoside//8.18 Valerine//7.27 (1R,3S)-1-Methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid//6.6 |
4′-O-Methylnorbelladine//4.14 Casuarine Analogue//-3.2 Folicanthine//-3.66 |
|
| Phenolic acids | Chlorogenic acid methyl ester//9.32 4-O-Caffeoylquinic acid methyl ester//7.92 3-hydroxyphenylacetic acid//7.38 methyl 5-caffeoylquinate//6.91 4-Hydroxybenzoic acid//6.87 1-O-Feruloylquinic acid//6.8 2,3-Dihydroxybenzoic Acid//6.54 |
Ethyl malto//4.14 Antiarol; 3,4,5-Trimethoxyphenol//3.64 methyl 5-caffeoylquinate//-3.33 1-O-Feruloylquinic acid//-3.41 3-O-Feruloylquinic acid//-3.48 4-O-Caffeoylquinic acid methyl ester//-3.64 p-Hydroxyphenyl 6-O-(E)-caffeoyl-β-D-allopyranoside//-4.7 |
1-O-p-Coumaroylquinic acid//-2.64 4-O-Caffeoylquinic acid methyl ester//-2.8 Chlorogenic acid methyl ester//-2.85 1-O-Feruloylquinic acid//-2.9 2-Hydroxycinnamic acid//-3.04 2-(Formylamino)benzoic acid//-3.13 α-Hydroxycinnamic Acid//-3.28 3-Hydroxycinnamic Acid//-3.33 Methyl 5-caffeoylquinate//-3.43 3,4,5-Trimethoxycinnamic acid//-3.73 |
| Flavonoids | Sesuvioside A//7.41 |
3-Hydroxy-4′,5,7-Trimethoxyflavanone//4.03 Butin; ,3′,4′-Trihydroxyflavanone//3.41 Sesuvioside A//-3.25 Isorhamnetin-3-O-neohesperidoside//-3.73 3,5,7-Trihydroxy-6,8-dimethyl-3-(4′-hydroxybenzyl)-chroman-4-one (Polygonatone C)//-4.23 |
3-[(3,4-dihydroxyphenyl)methylidene]-5,7-dihydroxy-6-methoxy-2h-1-benzopyran-4-one glucosyl rhamnoside//-2.72 Apigenin-6-C-(2″-glucosyl)arabinoside//-2.78 Tricin (5,7,4′-Trihydroxy-3′,5′-dimethoxyflavone)//-2.95 |
| Steroids | Spirost-5-en-12-one-3-O-glucosyl(1→2)glucosyl(1→4)galactoside (Pratioside D1)//-4.0 |
Spirost-5-ene-3,27-diol-27-O-glucoside-3-O-[rhamnosyl(1→4)]glucoside (Polygonatoside D)//-2.86 27-Hydroxyspirost-5-en-3-yl-O-rhamnosyl-(1→2)-O-[glucosyl-(1→6)]-glucoside//-3.17 Spirost-5-en-3-ol-3-O-glucosyl(1→2)glucosyl(l→4)galactoside (Neosibiricoside D)//-3.32 |
|
| Lignans and Coumarins | Phellodenol E//8.25 Guaiacylglycerol-β-Guaiacyl Ether//6.69 |
7,8-Dihydroxy-4-methylcoumarin//5.33 7-Hydroxycoumarin;Umbelliferone//3.38 5,7-Dihydroxy-4-Phenylcoumarin//3.22 |
Phellodenol E//-3.12 |
| Others | 2,5-Dihydroxybenzaldehyde//7.4 Protocatechualdehyde//6.96 4-Methyl-5-thiazoleethanol//6.75 4-Hydroxybenzaldehyde//6.46 |
Squamocin K//3.56 |
4-hydroxyphenyl acrylaldehyde//-3.67 4-Methylbenzaldehyde//-3.92 3-Methylbenzaldehyde//-4.11 |
| Total | Up: 20; down: 0 | Up: 9; down: 11 | Up: 0; down: 20 |
Table 2.
The number of newly added and disappeared secondary metabolites identified from three comparison groups.
Table 2.
The number of newly added and disappeared secondary metabolites identified from three comparison groups.
| Secondary metabolites | Comparison group | Total number | Class | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Alkaloids | Phenolic acids | Flavonoids | Terpenoids | Lignans and Coumarins | Quinones | Steroids | Others | |||
| Newly added | SD3 vs. CP | 164 | 45 | 42 | 23 | 13 | 15 | 8 | 5 | 13 |
| SD6 vs. SD3 | 16 | 6 | 3 | 4 | 1 | 2 | ||||
| SD9 vs. SD6 | 4 | 1 | 1 | 2 | ||||||
| Disappeared | SD3 vs. CP | 3 | 1 | 1 | 1 | |||||
| SD6 vs. SD3 | 33 | 10 | 6 | 11 | 1 | 1 | 1 | 3 | ||
| SD9 vs. SD6 | 30 | 9 | 10 | 4 | 2 | 3 | 1 | 1 | ||
Table 3.
Quantity and class of secondary metabolites clustered into seven variation patterns.
| Variation pattern | Number (%) | Class | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Alkaloids | Phenolic acids | Flavonoids | Terpenoids | Lignans and Coumarins | Quinones | Steroids | Others | ||
| 1 | 5 (0.90) | 3 | 1 | 1 | |||||
| 2 | 78 (14.00) | 26 | 17 | 13 | 8 | 6 | 1 | 7 | |
| 3 | 54 (9.69) | 19 | 10 | 6 | 6 | 6 | 7 | ||
| 4 | 26 (4.67) | 7 | 5 | 4 | 1 | 1 | 2 | 2 | 4 |
| 5 | 91 (16.34) | 36 | 22 | 5 | 7 | 4 | 2 | 1 | 14 |
| 6 | 190 (34.11) | 39 | 47 | 14 | 26 | 14 | 4 | 2 | 44 |
| 7 | 113 (20.29) | 24 | 33 | 13 | 6 | 14 | 4 | 5 | 14 |
| Total | 557 | 154 | 135 | 56 | 54 | 45 | 13 | 10 | 90 |
Table 4.
Significantly enriched KEGG pathways related to biosynthesis of secondary metabolites and amino acid metabolisms.
Table 4.
Significantly enriched KEGG pathways related to biosynthesis of secondary metabolites and amino acid metabolisms.
| Comparison group | Biosynthesis of secondary metabolites | Number of DASMs | Amino acid metabolism | Number of DASMs |
|---|---|---|---|---|
| SD3 vs. CP | ko00130: Ubiquinone and other terpenoid-quinone biosynthesis | 4↑; 1↓ | ko00380: Tryptophan metabolism | 4↑; 3↓ |
| ko00999: Biosynthesis of various plant secondary metabolites | 5↑; 2↓ | ko00350: Tyrosine metabolism | 7↑; 3↓ | |
| ko00996: Biosynthesis of various alkaloids | 3↑ | ko00360: Phenylalanine metabolism | 8↑ | |
| ko00950: Isoquinoline alkaloid biosynthesis | 3↑ | ko00470: D-Amino acid metabolism | 2↓ | |
| ko01110: Biosynthesis of secondary metabolites | 23↑; 5↓ | ko00310: Lysine degradation | 1↑; 1↓ | |
| ko00901: Indole alkaloid biosynthesis | 1↑ | ko00400: Phenylalanine, tyrosine and tryptophan biosynthesis | 1↑; 1↓ | |
| ko00902: Monoterpenoid biosynthesis | 1↑ | ko00260: Glycine, serine and threonine metabolism | 1↑ | |
| ko00904: Diterpenoid biosynthesis | 1↓ | ko00300: Lysine biosynthesis | 1↑ | |
| ko00943: Isoflavonoid biosynthesis | 1↑ | ko01230: Biosynthesis of amino acids | 1↑ | |
| ko00941: Flavonoid biosynthesis | 2↑ | ko00340: Histidine metabolism | 2↑ | |
| ko00960: Tropane, piperidine and pyridine alkaloid biosynthesis | 1↑; 2↓ | ko00330: Arginine and proline metabolism | 2↓ | |
| SD6 vs. SD3 | ko00941: Flavonoid biosynthesis | 5↑ | ko00470: D-Amino acid metabolism | 2↓ |
| ko01110: Biosynthesis of secondary metabolites | 9↑; 9↓ | ko00380: Tryptophan metabolism | 3↓ | |
| ko00944: Flavone and flavonol biosynthesis | 3↓ | ko00400: Phenylalanine, tyrosine and tryptophan biosynthesis | 1↓ | |
| ko00999: Biosynthesis of various plant secondary metabolites | 1↑; 3↓ | ko00340: Histidine metabolism | 1↑ | |
| ko00904: Diterpenoid biosynthesis | 1↑ | ko00350: Tyrosine metabolism | 1↑; 1↓ | |
| ko00960: Tropane, piperidine and pyridine alkaloid biosynthesis | 2↓ | ko00360: Phenylalanine metabolism | 1↑ | |
| ko00943: Isoflavonoid biosynthesis | 1↑ | |||
| ko00950: Isoquinoline alkaloid biosynthesis | 1↓ | |||
| ko00130: Ubiquinone and other terpenoid-quinone biosynthesis | 1↓ | |||
| SD9 vs. SD6 | ko00130: Ubiquinone and other terpenoid-quinone biosynthesis | 3↓ | ko00330: Arginine and proline metabolism | 4↓ |
| ko00950: Isoquinoline alkaloid biosynthesis | 2↓ | ko00260: Glycine, serine and threonine metabolism | 1↓ | |
| ko00902: Monoterpenoid biosynthesis | 1↓ | ko00360: Phenylalanine metabolism | 4↓ | |
| ko00944: Flavone and flavonol biosynthesis | 2↓ | ko00350: Tyrosine metabolism | 4↓ | |
| ko00999: Biosynthesis of various plant secondary metabolites | 1↑; 2↓ | ko00470: D-Amino acid metabolism | 1↓ | |
| ko00941: Flavonoid biosynthesis | 1↑; 1↓ | ko00340: Histidine metabolism | 1↓ | |
| ko01110: Biosynthesis of secondary metabolites | 2↑; 10↓ | ko00380: Tryptophan metabolism | 1↓ | |
| ko00960: Tropane, piperidine and pyridine alkaloid biosynthesis | 1↓ |
↑: up-regulated DASMs; ↓: down-regulated DASMs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



