
Submitted:
05 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Segmental bone defects caused by trauma, infection, tumor resection or osteoporotic fractures present significant surgical treatment challenges. Host bone autograft is considered the gold standard to restore function but comes with the cost of harvest site comorbidity. Allograft bone is a secondary option but has its own limitations in incorporation with host bone as well as cost. Therefore, developing new bone tissue engineering strategies to treat bone defects is critically needed. In the past three decades, the use of stem cells delivered with different scaffolds or growth factors for bone tissue engineering has made tremendous progress. Many varieties of stem cells have been isolated from different tissues for use in bone tissue engineering. This review summarizes the progress in using different postnatal stem cells including bone marrow mesenchymal stem cells, muscle-derived stem cells, adipose-derived stem cells, dental pulp stem cells/periodontal ligament stem cells, periosteum stem cells, umbilical cord-derived stem cells, peripheral blood stem cells and urine-derived stem cells for bone tissue engineering and repair. This review also summarizes the progress using exosomes or extracellular vesicles delivered with various scaffolds for bone repair. The advantages and disadvantages of each type of stem cell are also discussed and explained in detail. It is hopeful in the future that these preclinical results will translate into new therapies for bone defect repair.
Keywords:
Bone tissue engineering
; bone marrow mesenchymal stem cells
; muscle-derived stem cells
; adipose-derived stem cells
; dental pulp stem cells
; periodontal ligament stem cells
; periosteum stem cells
; umbi
1. Introduction
The repair of large bony defects caused by trauma, infection, tumor resection or fractures has traditionally relied on the use of bone autograft or allograft. Technological advances have allowed for the development of alternative approaches for the repair of bone defects and fracture non-unions using tissue engineering in combination with stem/progenitor cells, bone growth factors, and scaffolds. Many different types of stem cells derived from different tissue sources have been explored to promote stem cell-mediated bone regeneration with varying degrees of success.
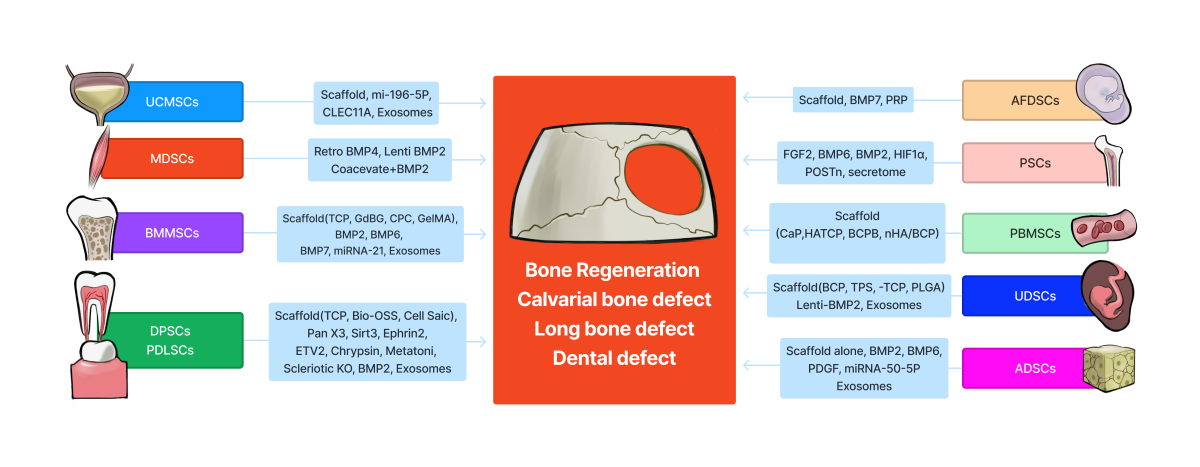
Adult or postnatal stem cells can be isolated from almost any tissue. The most commonly studied postnatal stem cells include bone marrow mesenchymal stem/stromal cells (BMMSCs), muscle-derived stem cells (MDSCs), adipose-derived stem cells (ADSCs), umbilical cord-derived mesenchymal stem cells (UC-MSCs), periosteal stem cells (PSCs), dental pulp-derived stem cells (DPSCs), periodontal ligament stem cells (PDLSCs), peripheral blood-derived mesenchymal stem cells (PB-MSCs) and urine derived system cells (UDSCs). Embryonic stem cells including induced pluripotent stem cells have also been investigated. This review focuses on the progress that has been made utilizing postnatal stem cells for bone tissue engineering and regeneration. Given the large number of literature available, in vivo preclinical studies are the focus of this review.
2. Bone marrow mesenchymal stem(stromal) cells (BMMSCs).
Bone marrow cells are both the earliest and mostly frequently investigated stem cells for use in bone tissue engineering. In 1991, Connolly et al. first explored the use of autologous bone marrow aspirate (BMA) for the treatment of fracture non-unions. They found that the BMA stimulated callus formation and eventual union in 8/10 patients who underwent cast fixation and 10/10 patients who underwent intramedullary nail fixation. The authors concluded that bone marrow injection was as effective as open autologous bone grafting but with considerably fewer disadvantages [1]. Furthermore, combining BMMSCs with demineralized bone matrix has been shown to be as effective as autologous iliac crest bone grafting for the treatment of non-unions [2]. In addition, loading BMMSCs onto a hydroxyapatite scaffold has also been shown to promote new bone formation within bone defects which may allow for the eventual recovery of skeletal function[2,3]. Over the past two decades, many in vitro and in vivo preclinical studies using animal models have demonstrated the effectiveness of using BMMSCs for bone tissue engineering. Combining BMMSCs with different scaffolds has been shown to be more efficient than applying a scaffold alone [4]. For the treatment of craniofacial defects, combining human BMMSCs with bone morphogenetic protein (BMP) 7 (an osteoinductive growth factor) improved the healing of the craniofacial bone defect [5]. In addition, Scotti C et al. proposed a novel strategy to enhance new bone formation by subcutaneously implanting a human BMMSC-derived cartilage intermediate. The group demonstrated that this cartilage intermediate formed a functional bone organ with an outer layer, mainly consisting of host cells which overlaid a pre-mineralized area, and an inner, trabecular-like, endochondral bone layer. The regenerated bone contained bone marrows with sinusoid-like structures of blood vessels, multiple lineages of hematopoietic stem cells and progenitor cells similar to that of native bone, which represents a strategy of a "developmental engineering" paradigm for functional bone regeneration[6,7]. Long T et al. has shown that BMMSC cell sheets closely mimic periosteum and allow osteogenic cells to promote new bone formation in critical-size bone defects as efficiently as autograft bone [8].
Lin et al. has already reviewed the progress of BMMSC aging and tissue engineering applications to enhance bone repair [9]. The clinical application of BMMSCs for skeletal tissues repair was also reviewed by Arthur A et al.[10,11]. Therefore, this section of the review will mainly focus on progress in the past 5 years of using human bone marrow stem cells for bone tissue engineering.
2.1. BMMSCs loaded with different scaffolds for bone tissue engineering.
Juan Francisco Blanco et al. reported that transplantation of human BMMSCs with a tri-calcium phosphate (TCP) scaffold into a rabbit femoral condyle critical size bone defect achieved improved bone regeneration and better osteointegration without eliciting inflammation, compared with the TCP scaffold alone [12]. Yang C et al. reported that rat BMMSCs transduced with lentiviral-miRNA-21 loaded within a TCP scaffold enhanced bone regeneration in a critical-size, calvarial bone defect model in rats, compared to the scaffold alone or the BMMSCs alone. Canine BMMSCs transduced with lentiviral-miRNA-21 were also found to enhance alveolar bone regeneration. The enhanced osteogenesis and bone regeneration was attributed to the upregulation of P-AKT/HIF1α and endogenous BMP2 expression [13]. Human BMMSCs seeded with gadolinium-doped bioglass (Gd-BG) scaffolds fabricated by combining hollow mesoporous Gd-BG microspheres with chitosan demonstrated enhanced osteogenesis in vitro and enhanced critical-size defect healing in a rat cranial bone defect model [14]. Du FF et al. compared fresh, autologous bone marrow mononuclear cell concentrate (BMMNC) with BMMSCs for repairing a segmental femur bone defect (1.5cm in length) in beagle dogs using β-TCP as a scaffold. The results showed that the BMMNCs group promoted greater bone regeneration than the cultured BMMSC group. The grafts in the BMMNC group also demonstrated improved mineralization, with a collagen arrangement and micro-biomechanical properties more similar to the contralateral native tibia bone [15].
BMP-6-loaded nano-hydroxyapatite (nHA)/gelatin (Gel)/gelatin microsphere (GMS) scaffolding, pre-seeded with BMMSCs was shown to promote BMMSC osteogenic differentiation in vitro and significantly accelerated bone regeneration in a rat critical-size calvarial defect model [16] .
Human BMMSCs loaded into calcium deficient hydroxyapatite (CDHA) scaffolds showed complete bone healing of a calvarial bone defect in nude mice with bone formation demonstrated in the pores of the CDHA scaffold. In contrast, when delivered with sintered β-TCP, the new bone formed along the periphery of β-TCP. CDHA scaffolds can be prepared at ambient temperature, yielding closer-to-native bone mineralization than β-TCP scaffolding. Also, BMMSCs showed better engraftment when loaded onto CDHA scaffolding, compared with the β-TCP [17]. Others reported that autologous BMMSCs and platelet-rich plasma (PRP) combined with a calcium phosphate cement (CPC) scaffold for femoral, critical-size bone defects resulted in healing in mini-pigs. The newly formed bone area was higher in the group with CPC scaffold-loaded MSCs and PRP than in the CPC scaffold-only group, at each time point (all p < 0.05). Thus, the strategy of CPC combined with BMMSCs and PRP may also be an effective method to repair bone defects [18].
Lin H et al. reported using a new nanocomposite that incorporated the GO-based nanosheets or silica-coated graphene oxide (SiGO) into methacrylated gelatin (GelMA)-based scaffolds to evaluate bone formation by human BMMSCs. Incorporation of GO markedly increased mineralization within human BMMSC-laden constructs, which was further increased by replacing GO with SiGO. Mechanistic analysis revealed that the nanosheet enhanced the production, retention, and biological activity of endogenous BMPs, resulting in robust osteogenesis in the absence of exogenous osteoinductive growth factors. The functional involvement of BMP was verified when the investigators inhibited the BMP signaling pathway in some of the GO-nanosheets and SiGO/GelMA scaffolds using the BMP-antagonist, LDN-193189, demonstrating significantly reduced mineralization via reduction of BMP-induced SMAD 1/5 phosphorylation [19]. The bone formation potential of this technology was further tested in vivo using subcutaneous implantation in a mouse model, which revealed that human BMMSC-laden GO/GelMA and SiGO/GelMA samples resulted in bone volumes 108 and 385 times larger, respectively, than the GelMA control group. These results demonstrated the biological activity and mechanism of action of GO-based nanosheets in augmenting the osteogenic capability of human BMMSCs and highlighted the potential of leveraging nanomaterials such as GO and SiGO for bone tissue engineering applications [19]. The same group also constructed a pre-vascularized bone-like tissue using human BMMSCs embedded in their own self-generated extracellular matrix. Then, a 3D culture of human umbilical vein endothelial cells (HUVECs)/HBMSCs was introduced to cover a bone-like construct’s surface for vascularization. The authors showed that this endochondral ossification-inspired procedure resulted in a robust osteogenic differentiation of human BMMSCs and markedly promoted the HUVEC/HBMSCs network formation in vitro. When the pre-vascularized bone-like tissues were subcutaneously implanted into mice, they exhibited significantly more functional blood vessel formation than the control group that contained HUVEC and HBMSC single cells, along with increased bone formation and remodeling [20].
Several recent studies also highlight the potential for specialized scaffold engineering using a variety of materials and production methods. BMMSCs differentiated on silk scaffolds into either hypertrophic chondrogenic pellets or osteogenic pellets were shown to form immature and mature bone when implanted subcutaneously, though the hypertrophic chondrogenic pellets form more vascularized bone, compared with osteogenic pellets, better mimicking the endochondral ossification process [21]. Rat MSCs loaded with a photo crosslinked biomimetic GelMA hydrogel scaffold resulted in robust new bone formation in a rat femoral bone defect model [22]. Machado-Paula et al. combined polycaprolactone (PCL) fibers, carbon nanotubes (CNT), and hydroxyapatite nanoparticles (nHap) and demonstrated that the fibers formed by rotary jet spinning (RJS) instead of traditional electrospinning (ES) with embedded rat BMMSCs showed the best outcomes to repair rat calvarial bone defects after 6 weeks, as demonstrated by a 10-fold increase in new bone formation compared to the RJS scaffold or BMMSCs only in a rat 5 mm critical-size bone defect [23].
Liu X et al. also engineered scaffold-free, functional spheroids with rat MSCs and two-dimensional hetero-nano-layers (2DHNL), consisting of black phosphorus (BP) and graphene oxide (GO), to create a three-dimensional (3D), cell-instructive microenvironment for large bone defect repair. After transplantation of the spheroids into the critical-size calvarial defects of rats, the authors demonstrated enhanced bone regeneration and neovascularization, as well as improved support for the osteogenic differentiation of the rat MSCs. Furthermore, adding the osteogenic factor, dexamethasone (DEX), on the 2DHNL showed high in vivo osteogenic induction and bone regrowth without prior in vitro culture in osteogenic medium. These functional, 2DHNL-impregnated spheroids that enable rat MSCs and osteogenic factor co-delivery could be a promising strategy for effective in vivo bone repair [24]. Hang Lin’s group developed an innovative idea by seeding human BMMSCs on their own secreted extracellular matrix (mECM) without an exogenous scaffold to allow human BMMSCs to undergo the N-cadherin-mediated developmental condensation process and subsequent chondrogenesis. Furthermore, the BMMSC-mECM constructs significantly enhanced bone formation in vivo via endochondral ossification [25]. It has also been shown that locally delivered, fluorescent nanoparticle (fNP)-labeled MSCs enhanced tibial defect repair, increased the number of stem cells, and supported vascular maturation in mice. fNP-MSCs also survived in the defect throughout repair. While only a small portion of transplanted cells underwent osteogenic differentiation (OSX+), a significant portion maintained their expression of MSC and skeletal stem cell markers (SCA-1 and PRRX1) [26]. Pitacco P et al. designed a new bone tissue engineering strategy of incorporating human BMMSCs into fibrin-based bioinks, then bioprinting these into PCL frameworks to produce mechanically reinforced constructs. Different in vitro culture regimens were used to produce chondrogenic and early-hypertrophic engineered grafts. Following their implantation into femoral bone defects of transiently immunosuppressed rats, the bioprinted constructs rapidly remodeled into bone in vivo, with early hypertrophic constructs supporting higher levels of vascularization and bone formation compared to the chondrogenic constructs. The early-hypertrophic, bioprinted constructs also supported higher levels of vascularization and spatially distinct patterns of new bone formation, compared to BMP-2 loaded collagen scaffolds (a positive control) [27].
2.2. BMMSCs delivered with scaffold and bone growth factors for bone tissue engineering.
Herberg S et al. designed MSC tube condensates to mimic the femoral diaphysis, then applied transforming growth factor (TGF)-β1 and BMP-2, attempting to recapitulate endochondral ossification. First, localized TGF-β1 + BMP-2-morphogen delivery stimulated chondrogenic priming/endochondral differentiation of the tubular condensates in vitro. When implanted subcutaneously, the human BMMSC tubes formed cartilage templates that underwent bony remodeling. Application of the MSC tubes were found to stimulate more robust endochondral defect healing than human BMMSC sheets, without significant ectopic bone formation, in a segmental femoral defect model [28].
Human BMMSC spheroids loaded with BMP-2-HA in viscoelastic gels demonstrated greater calcium deposition than human BMMSCs spheroids entrapped in elastic alginate gels. After implantation into critical-size calvarial bone defects, the viscoelastic hydrogels encased with human BMMSCs spheroids appeared to be a more potent stimulator of osteogenesis than hydrogels with BMP2-HA alone. However, increases in bone formation were evident in the viscoelastic gels, regardless of the BMP-2 integration method (i.e., soluble delivery versus HA nanoparticles) [29].
2.3. BMMSCs derived exosomes or extracellular vesicles in bone tissue engineering.
Fan J et al. developed an approach to amass exosome mimetics (EMs) from human BMMSCs. The human BMMSCs-EMs had a significantly increased proportion of vesicles positive for the exosome-specific CD63 marker, compared with BMMSC-derived exosomes using the traditional method, and showed enhanced bone regeneration in vivo using a chitosan hydrogel scaffold. Further, EMs from noggin knockdown BMMSCs enhanced bone regeneration via inhibition of miR-29a [30]. Another study showed that human BMMSCs osteogenically pre-differentiated for 10 and 15 days led to the production of osteogenic exosomes. When the purified exosomes were loaded into the 3D-printed titanium alloy scaffolds, the cell-free, exosome-coated scaffolds regenerated bone tissue as efficiently as human BMMSC-seeded exosome-free scaffolds within 12 weeks. Furthermore, RNA-sequencing indicated that the osteogenic exosomes induced the osteogenic differentiation by using their cargo, including upregulated osteogenic miRNAs (Hsa-miR-146a-5p, Hsa-miR-503-5p, Hsa-miR-483-3p, and Hsa-miR-129-5p) or downregulated anti-osteogenic miRNAs (Hsa-miR-32-5p, Hsa-miR-133a-3p, and Hsa-miR-204-5p), and by activating the PI3K/Akt and MAPK signaling pathways [31]. The rat BMMSC-derived exosome (BMSC-OI-exosome) contains the miRNA’s let-7a-5p, let-7c-5p, miR-328a-5p and miR-31a-5p which target Acvr2b/Acvr1 and regulate the competitive balance of Bmpr2/Acvr2b toward Bmpr-elicited Smad1/5/9 phosphorylation . Delivery of the BMSC-OI-exosome using mesoporous bioactive glass (MBG) rapidly induced bone regeneration in a rat skull defect model [32]. In a different study, BMMSC-derived functionally engineered EVs (FEEs) could bind to mimetic peptides from collagen (DGEA, GFPGER) and fibronectin (RGD). Using photo cross-linkable alginate hydrogels containing RGD to encapsulate, tether, and retain the FEEs over a period of 7 days, helped maintain the structural integrity and osteoinductive functionality of the EVs. When implanted in a calvarial defect model in vivo, alginate-RGD hydrogels containing the FEEs enhanced bone regeneration 4-fold when compared to controls lacking FEEs, and 2-fold when compared to controls without the tethering peptide [33].
2.4. Targeting cell senescence to improve BMMSC-mediated bone tissue engineering.
Cell senescence due to serial in vitro expansion can negatively affect MSCs’ functionality when implanted in vivo for bone defect healing. Therefore, eliminating senescent cells before cells transplantation represents a new strategy to enhance BMMSC-mediated bone defect healing. For example, Ming Xu’s group reported that treatment of aged murine BMMSCs with dasatinib +quercetin improved bone regeneration of aged murine BMMSCs [34]. Xing X et al. screened senolytic drugs using induced-senescent BMMSCs and found that quercetin, among others, can reliably eliminate senescent MSCs during culture, restore MSC self-renewal potential, and promote osteogenic differentiation in vitro. In vivo quercetin, when delivered with a hydrogel locally, enhanced long bone and calvarial defect healing in aged rats by decreasing MMP secretion from the local environment [35].
3. Muscle-derived stem cells (MDSCs)
David Yaffe first isolated myoblasts from rat skeletal muscle tissues using pre-plate techniques [36]. These cells could be cultured in vitro for many months, maintaining their capacity of continuous proliferation, fusion, and differentiation into postnatal multinucleated myofibers. Virtually all the cells in this cell line have the potential to differentiate into myofibers [36]. Later, Dr. Yaffe established a C2 myoblast cells line from the injured thigh muscle of two-month-old C3H mice[37] . Blau et al. further re-cloned C2 cells and expanded the cells to the C2C12 myoblast cell line [38]. Furthermore, Blau’s group isolated primary myoblasts use the pre-plating method and found that these cells are primarily myogenic, both in vitro and in vivo [39]. Subsequently, the Huard group isolated a subpopulation of muscle stem cells, termed muscle-derived stem cells (MDSCs, pre-plate 6), using the modified pre-plate technique, demonstrating that MDSCs are not only myogenic, but also multipotent in vitro and in vivo [40-43].
Earlier study from Huard group demonstrated that after transduction with BMP2 using an adenoviral vector, the MDSCs were found to be very efficient at regenerating new bone within a critical-size calvarial bone defect created in mice [41]. Additionally, it has been demonstrated in vivo that transplantation of allogenic retroviral BMP-4-transduced murine MDSCs (mMDSCs) regenerated bone efficiently in both immunocompetent mice, using a heterotopic bone formation model, and in a critical-size calvarial bone defect model in immunocompromised mice [44]. Further studies found that the transplantation of mMDSCs with retroviral transduced BMP-4 or BMP-2 in combination with retroviral-transduced VEGFα further enhance retro-BMP-2 mediated bone regeneration by promoting angiogenesis. Vice versa, when retroviral BMP-2 transduced mMDSCs were co-transplanted with retroviral soluble fms-like tyrosine kinase-1 (sFlt-1), a soluble receptor of VEGFA, bone regeneration was inhibited [45,46]. The bone regeneration capacity of MDSCs can also be fine-tuned by co-transplanting MDSCs transduced to express BMP-4 and noggin (BMP antagonist) with the latter prevents bone overgrowth [47]. In addition, retro-BMP-4/green fluorescent protein (GFP) MDSCs not only healed critical-size bone defects via their direct differentiation into osteoblasts and osteocytes, they also influenced the host cells response through the secretion various paracrine factors (MCP1, VEGFα, FGF2, IGF2, PDGF, and TGFβ1) [48]. The capacity of MDSCs to promote new bone formation is also affected by the sex of the donor and recipient. Male MDSCs were found to be more efficient than female MDSCs at promoting osteogenic differentiation in vitro and new bone formation in vivo [49]. This gender difference in bone formation was not related to variations in the concentration of circulating hormonal factors between male and female recipients [50].
Human MDSCs isolated using the pre-plate technique and transduced with BMP-2 using retroviral and adenoviral vectors, were able to promote healing of critical-size calvarial defects in SCID mice as demonstrated by gross observation and histology and a small fraction of the cells were found to contribute to bone formation [51]. Mastrogiacomo M et al. isolated human skeletal muscle-derived cells (MDCs) using a similar technique and found that the human MDCs underwent osteogenic, chondrogenic and adipogenic differentiation in vitro and could form new bone and cartilage in vivo when implanted subcutaneously [52]. Gao X et al. reported successful isolation of hMDSCs using pre-plate techniques and found 95% of hMDSCs expressing CD73, CD90, CD105, and CD44. Furthermore, 95% of the hMDSCs also are positive for CD56 and CD146 while also being negative for UEA and CD45[53]. These cells do not display small, round and shiny morphology as reported for mMDSCs [54], but rather showed a spindle-like MSC-like morphology [53]. The hMDSCs also underwent adipogenesis, chondrogenesis, osteogenesis, and myogenesis [53]. When the hMDSCs were transduced with lentiviral BMP-2, they regenerated a substantial amount of new bone and healed more than 75% of critical-size bone defects in immunocompromised mice [53]. It was further demonstrated that hMDSCs transduced with lentiviral-BMP-2 were just as efficient as human BMMSCs in terms of regenerating new, functional bone in a critical-size calvarial bone defect model [55]. Interestingly, bone regeneration with lentiviral BMP-2 transduced hMDSCs is not affected by the donor cells’ age but is impaired by host age, which indicated host micromilieu is also important for stem cell mediated bone regeneration [56]. Also, the quality of the bone formed from retro-BMP4 transduced mMDSCs is dependent on the scaffold used for delivery. Usas A et al compared Gelfoam, collagen gel (CG) and fibrin sealant (FS) as scaffold and found that FS and CG based scaffolds healed calvarial bone defects with closer resemblance to native bone compared with the bone overgrowth observed in the Gelfoam group. The FS scaffolds induced less ectopic ossification, further demonstrating fine-tuned MDSC delivery via absorbable bioengineered scaffold delivery systems. However, none of these scaffold induced new bone formation when loaded with mMDSCs transduced with LacZ (reported gene) which indicated the importance of BMP to induce mMDSC mediated bone formation [57].
Human muscle-derived cells isolated using cell sorting techniques have also shown bone regeneration capacity. For example, Zheng et al. found a population of myogenic endothelial stem cells (sorted for CD56+CD34+CD144+CD45-) could promote new bone formation in a muscle heterotopic bone formation model when transduced with BMP2 [58,59]. Another group has shown that the MSCs isolated from traumatically injured human muscles were also able to differentiate into osteoblasts, adipocytes and chondrocytes [60]. Because orthopaedic trauma often involves disruption of both muscle and bone within localized areas, it is possible that skeletal muscle tissue could be an important source of MSCs for future applications in orthopaedic trauma surgery.
4. Adipose derived stem cells (ASCs).
The use of adipose-derived stem cells (ADSCs) has garnered significant research interest in the field of tissue engineering because they are easily harvested and accessible in human patients. Since the techniques used for the isolation, characterization, and cryopreservation of ADSCs for applications in other areas of tissue engineering have been thoroughly reviewed by Levi B, Longaker MT [61]and De Francesco F [62], this section will only focus on the use of ADSCs for bone regeneration.
4.1. ADSCs alone with scaffold for bone tissue engineering
The first in vivo study that investigated the bone regenerative capacity of murine ADSCs seeded on apatite-coated, PLGA scaffolds was performed by Longaker's group [63]. The authors found that, in both juvenile and adult mice, transplantation of non-transduced ADSCs produced large, yet comparable, amounts of new bone in calvarial bone defects when compared to bone marrow cells and osteoblasts [63]. It was also shown that the transplanted ADSCs contributed to 84-99% of the new bone formation in the defect area [63]. In the same year, a case report demonstrated that autologous fibrin glue loaded with ADSCs significantly promoted healing of a traumatic calvarial defect [64]. Furthermore, it has been shown that the capacity of ADSCs to induce new bone formation is compromised by freezing and subsequent thawing the cells [65]. In vitro studies revealed a significant negative impact of the freeze-thaw process on cell proliferation as well as osteogenic and adipogenic differentiation (*P < 0.01)[65]. In vivo experiments showed near-complete healing in calvarial defects treated with fresh human adipose stem cells (hASCs), in contrast to minimal healing observed with freeze-thaw hASCs (*P < 0.01)[65]. However, adding either recombinant insulin-like growth factor 1 (rIGF-1) or recombinant BMP-4 (rBMP-4) significantly offset the impaired osteogenic differentiation in frozen hASCs (*P < 0.01)[65]. Kim Y et al investigated the bone regeneration potential of canine ADSCs and an ADSC osteogenic cell sheet (OCS), on a critical size radial 15 mm long segmental defect in a canine model using composite PCL/β-tricalcium phosphate (β-TCP) scaffolds. The authors found that the alkaline phosphatase (ALP) activity of osteogenic ADSCs and OCS was significantly higher when compared to undifferentiated ADSCs . ALP, runt-related transcription factor 2(RUX2), osteopontin, and bone morphogenetic protein 7 (BMP7) mRNA levels were upregulated in osteogenic ADSCs and OCS when compared to undifferentiated ADSCs [66]. In vivo, the study revealed that the combination of OCS with aPCL/β-TCP composite scaffold maximized new bone mass volume (28.11± 5.5 cm³) in canine radial defects, outperforming PCL/β-TCP alone or with undifferentiated ADSCs or osteogenic ADSCs. Though the defect was not completely healed in any of these groups, this study highlights the superior efficacy of OCS in conjunction with the composite scaffold for enhanced bone regeneration in critical-sized defects [66]. Orbay H et al. conducted a study using ADSCs harvested from the inguinal fat pads of male Lewis rats, differentiating them toward endothelial and osteoblastic lineages before transplantation into critical-size calvarial defects. The rats (n=30) were randomized into four groups, utilizing hydroxyapatite/poly(lactide-co-glycolide) [HA-PLG] scaffolds alone or seeded with non-differentiated ADSCs, ADSCs derived endothelial cells, or ADSCs derived osteoblasts. Micro-CT analysis eight weeks post-operation revealed the highest bone mineral density in the ADSC group derived osteoblast group (1.46 ± 0.01 g/cm3), followed by the ADSC derived endothelial cell group (1.43 ± 0.05 g/cm3), scaffold only group (1.42 ± 0.05 g/cm3), and non-differentiated ADSC group (1.3 ± 0.1 g/cm3) [67]. Although the osteogenic differentiated ADSC group exhibited the highest vascular density, the differences among the groups did not achieve statistical significance (P > 0.05), indicating that ADSC-derived endothelial cells and osteoblasts provided a limited increase in calvarial bone healing when combined with HA-PLG scaffolds [67] Bernhard J et al. created tissue-engineered grafts using human ADSCs by differentiating them into hypertrophic chondrocytes in decellularized bone scaffolds and compared these to acellular scaffolds and grafts engineered using ADSC-derived osteoblasts. After implanting these grafts into critical size femoral defects in athymic rats for 12 weeks, the grafts engineered using hypertrophic chondrocytes recapitulated endochondral ossification [68]. Highly enhanced bone deposition was associated with extensive bone remodeling and the formation of bone marrow, and with the presence of pro-regenerative M2 macrophages within the hypertrophic grafts[68]. As a result, hypertrophic chondrocyte grafts bridged 7/8 defects, as compared to only 1/8 for osteoblast grafts and 3/8 acellular scaffolds. These results suggested that the ADSC-derived hypertrophic chondrocytes in osteogenic scaffolds can markedly improve long bone repair [68]. Liu J et al. investigated rat allogeneic ADSCs combined with heterogeneous deproteinized bone (HDB) to repair segmental 4 mm radial defects in rats. The authors found that ADSCs-HDB with in vitro pre-osteogenic differentiation group regenerated the radius defects in 8 weeks completely, the ADSCs-HDB group without pre-differentiation also promoted bone defect healing compared to the HDB scaffold and blank control groups, with the blank control group resulting in a non-union. These results indicate in vitro pre-osteogenic differentiation of ADSCs is more effective to promote bone defect healing than undifferentiated ADSCs and represents an effective way for bone tissue engineering [69]. In another study, hADSCs were seeded in 3D culture systems, using spheroids and polystyrene scaffolds to mimic the native stem cell niche. The spheroids, in particular, exhibited enhanced osteogenic differentiation, as evidenced by ALP activity and the upregulated expression of key osteogenic markers such as RUNX2, osterix, integrin-binding sialoprotein (IBSP) and osteocalcin, compared to both polystyrene scaffolds and traditional 2D culture [70]. Zhang H et al. using a rabbit ADSC double cell sheet (DCS) and a composite scaffold made with polylysine (PLL)-modified coralline hydroxyapatite (CHA) with the aim of engineering vascularized bone to repair large radius bone defects in rabbits. The authors found that the DCS complex provided a very large cell reserve and formed abundant osteoblasts and vascular endothelial cells in vitro [68]. In vivo, at 12 weeks after surgery, the defect surface of the DCS-PLL-CHA group was completely wrapped by bone tissue and osteoids, the cortical bone was continuous, and the medullary cavity was perforated. A large amount of well-organized lamellar bone was formed, a small amount of undegraded CHA exhibited a linear pattern, and a significant amount of bone filling could be seen in the pores. Further, although the expression levels of BGLAP, SPP1 and VEGF were similar in each group, but PECAM1 expression was higher in the DCS-PLL-CHA group than in the autogenous bone group and the CHA group [71]. ADSCs seeded with tricalcium phosphates (TCP) and a PLGA scaffold enhanced mandibular bone defects healing in mini-pig with a significantly high bone volume regenerated (34.8% ± 4.80%) than scaffolds implanted without cells (n = 6, 22.4% ± 9.85%) as revealed by micro-CT (p < 0.05). Moreover, an increased amount of osteocalcin deposition was found in the experimental group in comparison to the control group (27.98 ± 2.81% vs 17.10 ± 3.57%, p < 0.001) [72].
4.2. ADSCs modified with different growth factors for bone tissue engineering.
Peterson B et al. found that human processed lipoaspirate (HPLA) cells genetically modified to overexpress BMP2 induced complete healing of femoral defects within a period of 8 weeks [73]. In contrast, implantation of HPLA cells that were not transduced to overexpress BMP2 resulted in no significant new bone formation. BMP2 alone also healed the femoral defect [73]. Hsu MN et al. designed a hybrid baculovirus (BV) system for the delivery of the BMP-2 gene and the CRISPRi system targeting Noggin of rat ADSCs. After BV-mediated co-delivery into ADSCs, the system induced prolonged BMP2 expression and simultaneously stimulated Nog expression while the CRISPRi system effectively repressed Nog upregulation for at least 14 days. The CRISPRi-mediated Nog knockdown, along with BMP2 overexpression, further stimulated the osteogenic differentiation of ADSCs. Implantation of the CRISPRi-engineered ADSCs into critical-size defects at the calvaria significantly enhanced the calvarial bone healing and matrix mineralization [74]. Chou YF et al. incorporated different doses of recombinant human BMP-2 (rhBMP-2) onto apatite-coated porous poly(l-lactide-co-dl-lactide) (70:30) (PLDLA) scaffolds, seeded them with ADSCs, then implanted them into athymic rats to observe critical-size femoral defect healing. Interestingly, the results showed that the combination of ADSCs and rhBMP-2 onto a scaffold did not enhance healing of calvarial defects when compared to rhBMP-2 alone. This result may indicate that at certain dosages, rhBMP-2 plays a dominant role in bone regeneration, potentially masking the therapeutic effects of implanted stem cells [75].
Kim Y. et al. used lentivirus-mediated BMP-7 transduced ADSCs to osteogenically enhanced cell sheets loaded with a PCL/β-TCP scaffold or combined with demineralized bone matrix (DBM). When applied in a 15 mm-long, segmental radius defect in a canine model, the BMP-7 overexpressing cell sheets, particularly when used with DBM, significantly increased bone regeneration and vascularization, evidenced by micro-CT analysis, histological evaluation, and gene expression analyses [76]. Osinga R et al. developed a strategy to use hADSCs to regenerate bone through endochondral ossifications by inducing chondrogenesis in vitro before transplantation. ADSCs were cultured either as micromass pellets or into collagen sponges in chondrogenic medium containing TGF-β3 and BMP-6 for 4 weeks (early hypertrophic templates) or for two additional weeks in medium supplemented with β-glycerophosphate, l-thyroxin, and interleukin1-β to induce hypertrophic maturation[77]. The authors showed, in vitro, ADSC-deposited cartilaginous matrix positive for glycosaminoglycans, type II collagen, and Indian hedgehog. Hypertrophic maturation induced upregulation of type X collagen, bone sialoprotein, and matrix metalloproteinase 13 (MMP-13)[77]. When transplanted in vivo, both early and late hypertrophic templates underwent cartilage remodeling, as assessed by MMP13- and tartrate-resistant acid phosphatase-positive staining, and developed bone ossicles, including bone marrow elements, although to variable degrees of efficiency. In situ hybridization for human-specific sequences and staining with a human specific anti-CD146 antibody demonstrated the direct contribution of ADSCs to bone and stromal tissue formation through endochondral ossification [77] developed PDGF and bio-mineral coated fibers which were then assembled with hADSCs to form spheroids, aiming to enhance vascularization alongside osteogenesis. The authors found that PDGF incorporation within the spheroids increased hADSC proliferation, as revealed by Ki-67 staining and DNA contents, enhanced osteogenic differentiation by 2.46-fold (osteocalcin expression), and elevated endothelial differentiation of hADSCs by 12.85-fold as revealed by von Willebrand factor (vWF), compared to those without PDGF[78]. When transplanted in vivo into a mouse calvarial defect, the spheroids demonstrated enhanced bone regeneration area (42.48 ± 10.84%) and the greatest number of capillaries and arterioles derived from transplanted hADSCs. These findings indicate that stem cell spheroids incorporating PDGF and bio-minerals could be used as a module for successful vascularized bone regeneration [78].
Additionally, different surgical and transplantation techniques may impact ADSC-based therapeutic efficacy, though evidence is limited. For example, Wang Z et al. evaluated the effects of drilling through the growth plate and using ADSCs and BMP-2 to treat femoral head epiphyseal ischemic necrosis in rabbit model. The authors found that the combination of growth plate drilling with either ADSCs or both ADSCs and BMP-2 significantly enhanced the restoration of normal hip structures, improving femoral epiphyseal quotients and trabecular areas compared to controls and drilling-only groups (P<0.01)[79]. The drilling plus BMP-2 group also demonstrated improved femoral epiphyseal quotients and trabecular areas compared with those of untreated and drilling treatment only groups (P<0.05)[79].
Other studies used small molecules to enhance ADSCs mediated bone tissue engineering. Fan J et al used the small molecule phenamil alongside BMP-2 to promote in vitro osteogenic regeneration and enhanced calvarial defect regeneration in a mouse model. It was found treatment of ADSCs with BMP-2 in combination with phenamil significantly promoted the in vitro osteogenic differentiation of ADSCs. In vivo, the scaffolds treated with phenamil+BMP-2 significantly promoted mouse calvarial regeneration compared with the groups treated with phenamil or BMP-2 alone[80]. Moreover, the combination treatment reduced the BMP-2 dose without compromising calvarial healing efficacy [80]. Yao W et al. developed a bone-seeking molecular compound, LLP2A-Alendronate (LLP2A-Ale), to augment ADSCs homing to bone with the goal of accelerating bone healing in a mouse closed fracture model. Mice with mid-femur fractures were treated with placebo, LLP2A-Ale (500 μg/kg, IV), ADSCs derived from wild-type female Osx-mCherry adipose tissue (ADSC, 3 x 105, IV) or ADSC + LLP2A-Ale. LLP2A-Ale treatment increased the number of exogenous ADSCs homing to the fracture gaps, enhanced incorporation of these cells into bone callus, and stimulated endochondral bone formation [81]. Additionally, higher engraftment of exogenous ADSCs in fracture gaps seemed to contribute to overall fracture healing and improved bone strength [81] . Another study focused on the modulation of the Wnt signaling pathway, targeting inhibitory factors such as Dickkopf-1 (DKK1), a secreted Wnt pathway antagonist. Utilizing anti-DKK1 neutralizing antibodies increased osteogenic differentiation of hADSCs in vitro. In vivo, systemic anti-DKK1 treatment improved hADSC engraftment, survival, and vascular ingrowth when implanted into femoral segmental bone defects in NOD-SCID mice, compared with the isotype antibody control during the repair process [82]. The modulation of paracrine effects seen in ADSC-based therapies also plays a significant role bone regeneration. For example, Levi B et al. conducted in vitro and in vivo experiments combining hADSCs with mouse calvarial osteoblasts in a co-culture supplemented with Hedgehog modifiers and with an osteoconductive scaffold into calvarial defects in mice. Micro-CT, histological evaluation, in situ hybridization, and PCR revealed that hADSCs can promote bone defect healing via paracrine effects and activation of the Hedgehog signaling pathway [83]. Taken together, ADSCs combined with different growth factors or inhibiting negative bone regeneration factors delivered with scaffold further enhanced bone regenerative potential of ADSCs.
4.3. ADSC-derived exosomes for bone tissue engineering.
Human ADSC-derived exosomes immobilized on the polydopamine-coating poly (lactic-co-glycolic acid) (PLGA/pDA) scaffolds under mild chemical conditions can be slowly and consistent released in vitro and promoted osteogenic differentiation, proliferation and migration of human MSCs[84]. In vivo results showed that this cell-free system significantly enhanced bone regeneration in a critical-size bone defect model, attributing in part to its osteoinductive effects as well as its promotion of MSC migration and homing to the newly formed bone tissue [84]. Enrichment of hADSC-derived exosomes with miR-375, achieved through stable overexpression after lentiviral transfection, has been shown to improve osteogenic differentiation of human BMMSCs and promoted bone regeneration in rat calvarial bone defects [85]. Another study combined human ADSC-derived exosomes with Mg2+ and gallic acid (GA) and constructed a functionalized, cell-free PLGA/Exo-Mg-GA metal-organic framework (MOF). The composite demonstrated enhanced osteogenic, angiogenic and anti-inflammatory capabilities. In vivo experiments further corroborated the osteogenic effects of this unique composite, revealing promotion of new bone formation and satisfactory osseointegration by stabilizing the bone graft environment, increasing blood supply, promoting osteogenic differentiation of endogenous cells, and accelerating bone regeneration [86]. Additionally, small extracellular vesicles (sEVs) derived from ADSCs, functionalized with a bioactive pentapeptide (cysteine–arginine–glutamic acid–lysine–alanine) (CREKA), showed increased binding to fibrin-fibronectin in vitro [87]. The functionalized sEVs have also demonstrated enhanced bone repair capabilities in a 2.8 mm femur epiphyseal bone defect in vivo. These positive effects on bone repair was mediated by modulation of local inflammation and enhancement of angiogenesis, osteogenesis [87].
4.4. miRNA regulated ADSCs for bone tissue engineering
Human ADSCs overexpressing miR-450b have been shown not only to promote osteogenic differentiation in vitro, but also enhance ectopic bone formation in vivo. This was achieved through the downregulation of BMP-3, an abundant BMP member that inhibit bone formation [88]. Wang F et al. investigated the role of miR-150-5p in ADSC-mediated bone regeneration. ADSCs were transfected with miR-150-5p inhibitors, miR-150-5p ADV, or short hairpin RNA (shRNA) of Notch3, and the subsequent effects on osteogenesis were evaluated. A combination of hydroxyapatite/tricalcium phosphate (HA/TCP) ceramic powders and transfected ADSCs were implanted into BALB/C nude mice to study the in vivo effects. It was revealed that compared to the negative control (NC) and miR-150-5p overexpression groups, the inhibition of miR-150-5p (miR-150-5p ADV group) significantly increased ADSC osteogenesis by regulating Notch3. MiR-150-5p overexpression decreased the expression of pFAK, pERK1/2, and RhoA. Conversely, these expression levels were up-regulated when miR-150-5p was inhibited, or Notch3 was silenced [89]. Furthermore, miR-150-5p inhibition partially reversed the suppressive effect of notch3 knockdown on osteogenesis in vitro and in vivo. This study demonstrated that the combination of ADSCs with miR-150-5p inhibition and HA/TCP scaffold might be a promising strategy for bone defect repair [89].
5. Dental pulp stem cells and periodontal ligament stem cells
The study of human dental pulp-derived stem cells (hDPSCs) or periodontal ligament cells (PDLSCs) for tissue engineering has been a topic of increasing interest. Several reviews have been published that summarize the numerous characteristics of stem cells derived from human dental tissues that possess multipotency in vitro and may have significant implications in the field of regenerative medicine [90-93]. This section will mainly review the progress in the past 5 years.
5.1. Unmodified DPSCs loaded with different scaffold for bone tissue engineering.
Laino G pioneered the isolation of stem cells from human dental tissues using a c-kit+/CD34+/CD45- marker profile (via fluorescence activated cell sorting [FACS]) and demonstrate their self-renewal and multipotent differentiation capabilities, which facilitated the formation of living autologous fibrous bone tissue (LAB) in vitro. After implantation into immunocompromised mice, LAB contributed to the formation of lamellar bone composed of osteocytes [94]. Various studies have subsequently explored the osteogenic differentiation of DPSCs.
Zhang W et al. demonstrated that both rat and human DPSCs and BMMSCs can efficiently undergo osteogenic differentiation in vitro; however, only rat BMMSCs underwent in vivo bone formation when seeded on hydroxyapatite/TCP scaffolds [95]. Interestingly, another group found that the DPSCs underwent osteogenic differentiation much more efficiently than BMMSCs [96]. It has also been shown that un-transduced DPSCs seeded onto collagen gel constructs can improve the healing of calvarial bone defects in the absence of growth factor supplementation. These DPSCs generated new bone via direct differentiation into an osteogenic lineage as well as the stimulation of angiogenesis from the host [97]. Rabbit autologous DPSCs expressed vimentin and CD44 when used (1×108) with Bio-Oss scaffold in the bone defect of rabbit alveolar toothless jaw, regenerated bone with abundant osteoblasts compared to few osteoblasts in scaffold only group [98]. Jahanbin A et al. compared bone regeneration of human DPSCs with autologous bone grafting for maxillary alveolar defect repair in rats using collagen matrix as a scaffold. The authors found that at 1 month, there was no difference between the autologous bone graft group and the DPSCs group. But at 2 months, maximum bone formation was observed in the iliac crest bone graft group which regenerated significantly more new bone than the scaffold-only group [99]. Zhang W et al. combined human DPSCs with tyrosine-derived polycarbonate polymer scaffolds [E1001(1k)] containing beta-tricalcium phosphate (β-TCP) [E1001(1k)/β-TCP] and compared this group to BMP2 (4 µg) in a rat mandibular ramus critical bone defect bone repair model. Human DPSC-seeded and acellular E1001(1k)/β-TCP scaffolds were cultured in vitro in osteogenic media for 1 week before implantation. Live micro-CT imaging at 3 and 6 weeks post-implantation revealed robust bone regeneration in the BMP implant group. DPSCs seeded with high (5×105) and lower density (2.5×105) groups exhibited similar uniformly distributed mineralized tissue coverage throughout the defects but less than the BMP implants[100] . In addition, robust expression of dentin and bone differentiation marker expression was observed in human DPSC-seeded scaffolds, whereas, in contrast, BMP and scaffold-alone implants exhibited only bone and not dentin differentiation marker expression[100]. Human DPSCs were detected at 3 weeks but not at 6 weeks in the human DPSC-seeded scaffold groups, indicating that their survival was at least 3 weeks[100]. These results showed that hDPSC-seeded E1001(1k)/β-TCP scaffolds support the rapid regeneration of osteo-dentin-like mineralized jaw tissue, representing a promising new therapy for alveolar jaw bone repair and regeneration [100]. Human DPSCs/HUVECs seeded on acellular tyrosine-derived polycarbonate E1001(1K)/β-TCP constructs were implanted into rabbit craniomaxillofacial (CMF) bone defects at one and three months. The results showed that human DPSCs seeded E1001(1k)/β-TCP constructs support the formation of osteodentin-like mineralized jawbone tissue closely resembling that of natural rabbit jawbone. Although unseeded scaffolds supported limited alveolar bone regeneration, more robust and homogeneous bone formation was observed in the hDPSC/HUVEC-seeded constructs, suggesting that human DPSCs/HUVECs contributed to enhanced bone formation. Further, the bioengineered jaw bone recapitulated the typical morphology of natural rabbit jawbone, was highly vascularized, and exhibited active remodeling as evidenced by the presence of osteoblasts and osteoclasts on newly formed bone surfaces [101]. Li Y et al. isolated DPSCs from autologous inflammatory dental pulp from two patients and loaded these onto a scaffold of β-tricalcium phosphate and engrafted into the periodontal defect area in the root furcation. Clinical and radiographic evaluation showed that DPSCs from inflammatory dental pulp tissues were able to graft and regenerate new bone to repair periodontal defects 9 months after surgical reconstruction[102]. Hu J et al. isolated human DPSCs and used the DPSCs as cell sheets or single cell injections to treat miniature pig periodontitis with bone defects (5mm in width, 7 mm in length, and 3 mm in depth). After 12 weeks, both the human DPSC sheet treatment and human DPSC injection significantly improved periodontal tissue healing clinically in comparison with the control group[103]. The volume of regenerative bone in the human DPSC cell sheet group (52.7 ± 4.1 mm(3)) was significantly larger than in the human DPSC injection group (32.4 ± 5.1 mm(3)) (P < 0.05). The percentage of bone in the periodontium in the human DPSC injection group was 12.8 ± 4.4 %, while it was 17.4 ± 5.3 % in the human DPSC sheet group (P < 0.05). This indicated that transplantation of human DPSCs into this large animal model significantly improved periodontal bone regeneration and soft tissue healing [103]. Lyu J et al. compared the recombinant peptide Cellnest™ 3D stem cell matrix (CellSaic) containing rat DPSCs and BMMSCs for rat congenital cleft fracture repair. Cultured CellSaic in osteoinductive media generated more mineralized tissues than the control group without osteoinductive media. Overall, rat BMSC-Cell Saic and rat DPSC-Cell Saic made with Cellnest™ as a scaffold provided excellent support for promoting bone regeneration in rat mandibular congenital defects[104]. Both differentiated and undifferentiated rat DPSC-CellSaic, but only differentiated rat BMSC-CellSaic could induce the formation of new bone tissue. Rat DPSC-CellSaic represents a better source for craniofacial bone defect repair than rat BMSC-CellSaic [104](34502394, 2021). Another study evaluated the use of ceramic nanocomposites of hydroxyapatite/titania /calcium silicate (C1, C2, and C3) with hDPSCs, demonstrating enhancement in bone healing and osteointegration in a rabbit tibia defect model compared to the control group [105].
A single-center, double-blind, randomized, split-mouth, controlled clinical trial was conducted to evaluate the beneficial effects of uncultured DPSCs delivered in a collagen matrix on the post-extraction sockets of impacted mandibular third molars in 32 patients. The clinical, radiological, and surgical characteristics of the impacted third molars in both the control and experimental groups were homogeneous. No significant differences were observed in terms of bone repair when analyzing density (p=0.4203, neuroradiologist 1; p=0.2525, neuroradiologist 2) or interdental septum height (p=0.2280, neuroradiologist 1; p=0.4784, neuroradiologist 2). The study could not demonstrate that autologous dental pulp mesenchymal stem cells reduced socket bone resorption following the extraction of inferior third molar extraction [106].
5.2. DPSCs modified with different genes for bone tissue engineering.
Song F et al. discovered that Pannexin3 (Panx3) was upregulated during the osteogenic differentiation of DPSCs in a time-dependent manner. Overexpression of Panx3 promoted osteogenic differentiation of hDPSCs, as evidenced by the upregulated expression of mineralization-related markers, increased ALP activity, and enhanced ALP and Alizarin red staining. Conversely, depletion of Panx3 resulted in impaired osteogenic differentiation[107]. Panx3 was found to interact with the Wnt/β-catenin signaling pathway, forming a negative feedback loop. However, Wnt/β-catenin did not contribute to the enhancement of osteogenic differentiation as observed in Panx3 overexpression. Moreover, Panx3 promoted osteogenic differentiation of human DPSCs by increasing the ERK signaling pathway. In vivo, Micro-CT and histological staining results showed that Panx3-modified human DPSCs significantly improved ossification of critical-size bone defects [107].
Song D et al. utilized DPSCs transduced with Adeno-SIRT1 to enhance distraction osteogenesis in a rabbit tibia model. The authors demonstrated that the Ad-SIRT1 overexpressing DPSCs group exhibited improved bone formation, higher bone mineral density (BMD), and increased bone mineral content (BMC) compared to the AD-GFP-DPSCs and no cell group [108]. Wang W et al. revealed that ephrinB2 was upregulated following the osteogenic induction of human DPSCs. Overexpression of ephrinB2 enhanced the osteogenic differentiation capacity of human DPSCs in vitro. Additionally, p-ephrinB2, was upregulated by ephrinB2 overexpression [109]. In a canine bone defect model, Lenti-ephrinB2 transduced canine DPSCs significantly improved the quality of newly regenerated alveolar bone, as demonstrated by higher trabecular bone volume per tissue volume (BV/TV), trabecular thickness, and radiographic analysis[109]. Furthermore, Ets variant 2 (ETV2) transcription factor-transduced DPSCs exhibited enhanced osteogenesis in vitro compared to un-transduced human PDSCs. Transplantation of ETV2-transduced DPSCs using a β-TCP scaffold increased bone formation in a rat calvarial defect model and a nude mice ectopic bone formation model [110].
5.3. DPSCs treated with small molecule or its inhibitor enhanced bone repair.
DPSCs were harvested from six healthy patients aged 18-29 years and cultured in normal medium (NM), osteogenic medium (OM), or OM with a helioxanthin derivative, 4-(4-methoxyphenyl) pyrido[40,30:4,5]thieno[2,3-b]pyridine-2-carboxamide (TH). These cells were then fabricated into cell sheets and labeled with PKH26. After transplantation into mouse tibial fractures, it was demonstrated that the transplanted OM+TH-treated DPSC sheets localized to the fracture site and facilitated bone formation [111]. Human DPSCs treated with chrysin, a flavanol extracted from oroxylum seeds, exhibited increased osteogenic differentiation in vitro and enhanced β-TCP induced mineralization in a mouse heterotopic bone formation model and a rat calvaria defect model by upregulating SMAD3 [112]. DPSCs treated with melatonin showed enhanced osteogenic differentiation by increasing P38MAPK activity. In vivo, the transplantation of DPSCs in a calvarial defect model using an MBCP scaffold resulted in improved bone regeneration compared to just the scaffold or an empty control, but it was not significantly better than untreated DPSCs [113].
Combining sclerostin antibody with DPSCs increased bone regeneration in WT mice. Meanwhile, the implantation of SOSTKO DPSCs exhibited similar bone regeneration in WT mice compared to bone regeneration in SOSTKO mice, indicating that the regulation of bone formation factors of DPSCs is also important [114].
5.4. DPSCs or PDLSCs exosome for bone tissue engineering.
Healthy human periodontal ligament stem cells (PDLSCs) derived exosomes, loaded with hydrogel or β-TCP, accelerated bone formation in alveolar bone defects in rat models of periodontitis-induced bone loss. Mechanistically, human PDLSCs-exosomes suppressed the over-activation of canonical Wnt signaling, restored the osteogenic differentiation capacity of inflammatory PDLSCs [115]. Human PDLSC-derived sEVs (P-EVs) have been shown to enhance the proliferation and migration of BMMSCs through increased phosphorylation of AKT and extracellular signal-regulated kinase 1/2 (ERK1/2). The role of P-EV-induced adenosine receptor signaling in AKT and ERK1/2 phosphorylation was a key mediator contributing to enhanced BMMSC proliferation and migration. In vivo, P-EV/Matrigel accelerated bone tissue repair by increasing cell infiltration compared to the control group. Additionally, exosomes derived from PDLSCs enhanced alveolar bone defect healing in a rat model [116]. The above studies provide evidence that stem cells derived from dental tissues may be another source of stem cells for bone tissue engineering especially craniofacial bone regeneration.
6. Periosteal stem cells (PSCs).
Periosteum is indispensable in bone repair and is an important source of skeletal stem cells (SSCs) for endogenous bone regeneration [117]. However, relative fewer studies used isolated PSCs for bone regeneration and tissue engineering.
6.1. PSCs alone or combining with bone growth factors for bone tissue engineering.
Van Gastel N et al. demonstrated that murine PSCs, pre-treated with FGF2, resulted in complete healing of large bone defects in a mouse femoral via endochondral bone healing [118]. Ji W et al. compared human PSCs seeded with different doses of BMP6 or various clinical grades of calcium phosphate scaffold (ChronOs®, ReproBone™, & CopiOs®) for bone regeneration in an ectopic bone formation model. They reported that cells seeded on CaP scaffolds with an intermediate Ca2+ release rate, combined with low or medium dosages of BMP6 (810ng and 3240ng BMP6/scaffold), demonstrated robust new bone formation at 5 weeks, and the new bone was derived from both donor and host cells [119].
PSCs transplanted into 3cm fresh tibia defects in sheep showed similar effects as BMP6 or BMP2 protein treatment. However, in a 4.5cm biologically exhausted tibial defect, only PSCs in combination with the BMP group promoted bone defect healing in sheep. This result indicated that exogenous PSCs are very important when host cells are compromised [120]. To determine the role of HIF-1α in PSC-mediated bone regeneration, it was found that knockdown HIF-1α impaired the bone regeneration and osteogenesis of PSCs both in vivo and in vitro. Furthermore, the knockdown of HIF-1α also reduced periostin (POSTN) expression, and the addition of recombinant POSTN partially rescued the osteogenic inhibition. The phosphorylation levels of PI3K and AKT were enhanced with HIF-1α overexpression and inhibited when HIF-1α was knockdown, and the addition of a PI3K or AKT activator could partially rescue POSTN expression [121].
6.2. PSCs from different anatomic origins demonstrate variable bone regeneration capacities.
Human PSCs isolated from different anatomic origins (tibia, maxilla, and mandible) exhibited similar trilineage differentiation in vitro. However, for in vivo bone formation, 8 weeks after ectopic implantation in nude mice, it was observed that constructs seeded with tibial and mandibular human PSCs, but not maxilla PSCs, regenerated bone effectively [122]. Tang Y et al. investigated the differences in PSCs from the mandible and femur and their potential responses to YAP signaling. Mandibular PSCs were in cubic shape with better proliferation, while femoral PSCs were in slender with reduced cell viability compared to mandibular PSCs. Mandibular PSCs outperformed femoral PSCs in ALP activity, osteogenic related genes’ mRNA expression, and calcium deposition at a later stage. Interestingly, the downregulation of YAP enhanced the ALP activity, related gene’s mRNA expression, and calcium deposits of femoral PSCs, while inhibiting those characteristics in mandibular PSCs in vitro. Mandibular PSCs also demonstrated superior bone repair in both mandible and femoral defect models, likely due to their different embryonic origins and modes of bone formation[123].
6.3. PSC secretomes for bone tissue engineering
Pranskunas M et al. evaluated the functionality of the secretome isolated from PSCs in basal or osteogenic-induced conditions in healing critical-size calvarial bone defects in a rabbit model using a bioceramic xenograft scaffold. The osteogenic-induced PSC secretome showed increased diversity of proteins, especially those related to osteogenesis [124]. Micro-CT and histological morphometric analysis revealed that bioceramic xenografts implanted with secretomes enhanced the new bone formation process, with the osteogenic-induced secretome inducing the greatest bone tissue formation. Therefore, the application of the PSC secretome, particularly from osteogenic-induced PSCs, may be an effective therapeutic approach to enhance bone tissue healing and regeneration [124].
7. Amniotic fluid derived stem cells (AFDSCs).
Human amniotic fluid-derived stem cells (AFDSCs) represent a unique population of stem cells that are easily accessible and characterized as an intermediate stage between embryonic stem cells (ESCs) and adult stem cells. Human AFDSCs can be induced towards osteoblastic differentiation by rhBMP-7 and respond more strongly to rhBMP-7 than human BMMSCs). When seeded on nanofibrous scaffolds (NF scaffolds) with morphology similar to that of natural collagen fibers, showed significantly enhanced ALP activity, calcium content, von Kossa staining and the expression of osteogenic genes than those on the traditional scaffolds, i.e. solid walled scaffolds both in vitro and in bone formation in vivo [125]. Human AFDSCs have been shown to regenerate bone in critical-size calvarial bone defects via direct differentiation into osteogenic and endothelial cell lineages [97]. They also undergo in vitro osteogenic differentiation when seeded onto poly(lactide-co-glycolide) (PLGA)-bladder submucosa matrix (BSM) composite scaffolds[126]. Mohammed EEA et al. reported that human second-trimester AFDSCs, when seeded on a 30% Nano-hydroxyapatite chitosan scaffold, undergo osteogenic differentiation in vitro. When transplanted into a rabbit tibia defect, they enhanced bone formation, showing complete bone defect healing at 4 weeks after surgery [127]. Using human AFDSCs to fabricate osteogenic and vascular cell sheets and applying these sheets to a rat 4 mm calvarial bone defect enhanced bone regeneration more than osteogenic or vascular cell sheets alone and more than the control group [128].
Wang M reported that rat AFDSCs exhibit typical fibroblastoid morphology, stable proliferation activity, and multi-differentiation potential. Flow-cytometry analysis demonstrated that these cells were positive for CD29, CD44, and CD90, while negative for hematopoietic markers such as CD34 and CD45. These cells, when premixed with a platelet-rich plasma (PRP) gel, demonstrated superior regenerative capacity in restoring alveolar bone defects, as evidenced by micro-CT and histological analyses at 4 and 8 weeks post-surgery, compared to control groups. Moreover, the implanted AFDSCs survived in the defect site and directly participated in the bone tissue regeneration [129]. Ghaffarinovin Z et al. isolated rat AFDSCs and seeded them on random polycaprolactone (PCL) fibrous scaffolds combined with PRP, applying this combination to repair calvarial bone defect. The authors found that collagen type I was expressed by AFDSCs cultured on the scaffold. Adding PRP promoted the formation of blood vessels and collagen type I expression in the defect area [130].
8. Peripheral blood-derived mesenchymal stem cells (PBMSCs).
Recently, new methods have been investigated to isolate MSCs from peripheral blood due to its easy accessibility compared to other sources such as bone marrow. The Gang Li group first isolated PBMSCs from rabbit blood, comparing their bone regeneration capabilities with rabbit BMMSCs in a rabbit ulna 20 mm defect model. The authors demonstrated that rabbit PBMSCs possessed multi-differentiation potential comparable to BMMSCs [131]. Allogenic PBMSCs seeded onto a porous calcium phosphate resorbable scaffold enhanced bone regeneration in the rabbit ulna critical-sized bone defect model, suggesting that allogenic PBMSCs might be a new source of circulating osteogenic stem cells for bone regeneration and tissue engineering [131]. Zheng RC et al. also isolated PBMSCs from rabbits, showing that PBMSCs had a similar proliferation rate, as indicated by BrdU-positive cells, compared to BMMSCs. PBMSCs were positive for CD90 but negative for CD14. They exhibited osteogenic, adipogenic, and chondrogenic differentiation in vitro and bone formation in vivo in an immunocompromised mouse root canal model. Further histological results demonstrated that the PBMSC and BMMSC groups showed higher newly formed bone than the HA/TCP and defect groups in the upper and lower chambers at 6 weeks, as well as in the upper canal at 3 weeks; however, there was no difference in newly formed bone among all groups in the lower canal at 3 weeks. The PBMSCs exhibit characteristics and bone regenerative capacity similar to BMMSCs both in vitro and in vivo. Therefore, the authors concluded that PBMSCs are a promising cell source for bone regeneration clinical use [132]. Chen L et al. developed a three-dimensional (3D) co-culture system using a biphasic calcium phosphate bioceramic (BCPB) scaffold seeded with rabbit PBMSCs and endothelial progenitor cells (EPCs) to improve new bone formation and vascularization for long bone segmental defects. The results showed both osteogenic and vascular related genes were up-regulated when EPCs were co-cultured with PBMSCs. In addition, BCPB is biocompatible and the expression levels of osteogenic and vascular related markers were also up-regulated in the 3D co-culture system[133]. After implantation of seeded PBMSCs and EPCs within a modified BCPB resulted substantial new bone formation and promoted vascularization in a rabbit large bone defect model. This study demonstrate that PBMSCs represent a potentially alternative cell source in the treatment of large bone defects [133]. Wang H et al. used a similar concept to construct a novel vascularized tissue engineered bone (VTEB) by using rabbit PBMSCs and peripheral blood endothelial progenitor cells (PBEPC) seeded on a 3D-printed biphasic calcium phosphate (BCP) scaffold with highly bioactive nano hydroxyapatite (nHA) coating (nHA/BCP). They tested it in a rabbit femoral segmental bone defect (SBD) model. The 2D coculture of PBMSCs and PBEPCs showed that these constructs could promote the osteogenic or angiogenic differentiation of each cell type ratio, especially in the group with PBEPC/PBMSC = 75:25. The 3D coculture results demonstrated that the nHA coating could further promote PBEPC/PBMSC adhesion, proliferation, and osteogenic and angiogenic differentiation on the BCP scaffold[134]. In vivo, it was found that among the four groups (BCP, BCP-PBEPC/PBMSC, nHA/BCP, and nHA/BCP-PBEPC/PBMSC), the nHA/BCP-PBEPC/PBMSC group induced the best formation of blood vessels and newly formed bone and the best repair of the SBD. Therefore, taking advantage of the synergistic effect of nHA and PBEPC/PBMSC on angiogenesis and osteogenesis in the BCP scaffold is a new strategy for efficient bone repair [134]. Li S et al. isolated a subset of CD45- cells with fibroblast-like morphology from mouse peripheral blood. These cells were adherent to plastic, negative for CD34, CD19, CD11b, lineage and c-kit, and positive for Sca-1, CD73, CD44. CD90.1, CD29, CD105, CD106, CD140a. These cells exhibited osteogenic, chondrogenic, and adipogenic differentiation potentials and induced healing of critical-size calvarial bone defects in vivo [135].
Human PBMSCs were also isolated from peripheral blood. Purified populations of PBMSCs can be obtained within a short period of time using this protocol with a success rate of 60%. Human PBMSCs cultured under hypoxia possessed potent multi-lineage differentiation capacity. They also expressed Nanog and Lgr5, as well as a series of MSC surface antigens (including CD29, CD90, CD105, and CD73). Additionally, using an ectopic bone formation model, it was demonstrated that the transplantation of human PBMSCs regenerate bone in vivo [136].
9. Umbilical cord derived mesenchymal stem cells (UC-MSCs).
Human umbilical cords represent another source of stem cells that can be banked and used for both mother and infant.
9.1. UC-MSCs delivered with different scaffold for bone tissue engineering.
Diao Y et al demonstrated that human umbilical cord-derived mesenchymal stem cells (UC-MSCs) at the fourth passage were positive for CD29, CD44, CD71, CD73, CD90, and CD105, and negative for CD14, CD34, CD45, and CD117. Furthermore, these cells expressed HLA-A, B, C (MHC-I), but not HLA-DP, DQ, DR (MHC-II), or costimulatory molecules such as CD80 and CD86. Following incubation in specific inductive media for 3 weeks, cultured cells were shown to possess the potential to differentiate into adipogenic, osteogenic, or chondrogenic lineages in vitro [137]. When UC-MSCs loaded with a biomimetic artificial bone scaffold material were implanted subcutaneously into the back of Balb/c nude mice for four to twelve weeks, underwent osteogenesis in vivo [137]. Human UC-MSCs transfected with the pEGFP-OSX plasmid showed enhanced in vitro osteogenic differentiation, and co-delivery using PLGA scaffold promoted bone formation in nude mice 4 weeks after transplantation.
A retrospective analysis of the clinical effects (Randomized Clinical Trials) of transplanted human UC-MSCs for the treatment of osteonecrosis of the femoral head (ONFH) demonstrated that human UC-MSCs grafted by intra-arterial infusion organized effective perfusion for femur head as demonstrated by an increase in the oxygen delivery index (ODI) at three days post-operation. The MRI results revealed that at 12 and 24 months after treatment, the necrotic volume of the femoral heads was significantly reduced, and no obvious abnormalities were observed. These data indicate that intra-arterially infused human UC-MSCs migrate into the necrotic field of femoral heads and differentiate into osteoblasts, thus improving the avascular necrosis of femoral heads. This finding suggested that the intra-arterial infusion of human UC-MSCs is a feasible and relatively safe method for the treatment of femoral head necrosis [138] . Mesenchymal stem cells derived from the umbilical cord (Wharton’s jelly (WJ-MSC)) seeded onto Bio-Oss scaffold can differentiate into osteoblast-like cells, and when injected, the Bio-Oss scaffold significantly enhanced calvarial defect healing in a rat model [139]. Others have shown that miR-196a-5p could repress proliferation, stimulate osteogenic differentiation, and WJCMSC sheet-derived ECM deposition, thus promoting new bone formation and rat calvarial bone defect closure. Furthermore, SERPINB2 is a key downstream gene involved in the miR-196a-5p-promoted WJCMSC osteogenesis [140].
9.2. UC-MSC derived exosomes for bone tissue engineering.
Human UC-MSC-derived exosomes could effectively promote the proliferation, migration, and osteogenic differentiation of a murine calvariae pre-osteoblast cell line in vitro. When an injectable hydroxyapatite (HAP)-embedded in situ cross-linked hyaluronic acid-alginate (HA-ALG) hydrogel system was used with the exosome, the combination significantly enhanced bone regeneration in rats [141]. Human UM-MSC-derived extracellular vesicles (UC-MSC-EVs) have a size ranging from 60 nm to 150 nm and express CD9, CD63, CD81, and TSG101. Systemic administration of human UC-MSC-EVs prevented bone loss and maintained bone strength in osteoporotic mice by enhancing bone formation, reducing marrow fat accumulation, and decreasing bone resorption. The beneficial effect is thought to correlate with highly expressed levels of pro-osteogenic protein, C-type lectin domain family 11, member A (CLEC11A), in human UC-MSC-EVs. In addition, human UC-MSC-EVs enhanced the shift from adipogenic to osteogenic differentiation of BMMSCs via delivering CLEC11A in vitro [142].
Another study showed UC-MSC-derived exosomes (UC-MSC-EXOs) encapsulated in hyaluronic acid hydrogel (HA-Gel) and combined with customized nanohydroxyapatite/poly-ε-caprolactone (nHP) scaffolds markedly enhanced bone regeneration in vivo. Moreover, the in vitro results demonstrated that UC-MSC-EXOs promoted the proliferation, migration, and angiogenic differentiation of endothelial progenitor cells (EPCs) but did not significantly affect the osteogenic differentiation of BMMSCs[143]. Importantly, mechanistic studies revealed that exosomal miR-21 was the potential intercellular messenger that promoted angiogenesis by upregulating the NOTCH1/DLL4 pathway [143]. Human UC-MSC-derived exosome-loaded chitosan/hydroxyapatite (CS/HA) scaffold also regenerated significantly more bone than the scaffold only group or the control group in a rat calvarial bone defect model [144].
10. Urine derived stem cells (UDSCs).
10.1. UDSCs loaded with different scaffold materials for bone tissue engineering.
Guan et al. isolated human UDSCs via a simple centrifuge method to obtain cell pellets that were adherent to plastic. The human UDSCs demonstrated multipotent differentiation potentials and expressed CD29, CD44, CD73, CD90, and were negative for CD34, CD45, CD133, and HLA-DR. When these human UDSCs were loaded with β-TCP, they induced substantial new bone formation and subsequent healing of segmental bone defects in rat femora[145]. Human UDSCs transduced with Lenti-BMP2 showed enhanced osteogenic differentiation capacity and enhanced ectopic bone formation in nude mice, and the direct contribution of human cells was observed in the newly regenerated bone[146] . Human UDSCs seeded on calcium silicate (CS) particles incorporated into poly (lactic-co-glycolic acid) (PLGA) composite scaffolds showed enhanced cell proliferation, ALP activity, calcium deposition, and expression of certain osteoblast-related genes and proteins via inducing the Wnt/β-catenin signaling pathway. Furthermore, human UDSCs seeded on the CS/PLGA scaffold promoted bone formation in vivo in immune-compromised mice [147].
Others also showed UDSCs loaded with surface mineralized biphasic calcium phosphate ceramics (BCPs) significantly promoted bone defect healing in a New Zealand white rabbit segmental bone defect model [148]. Another study demonstrated that a graphene oxide-modified silk fibroin/nanohydroxyapatite scaffold loaded with human UDSCs significantly promoted rat bone defect healing via modulation of the polarization of macrophages to the M2 macrophage phenotype that promotes bone regeneration [149]. Liu M et al. combined human UDSCs with a biphasic calcium phosphate (BCP) bioceramic ornamented with chitosan sponges (CS) (CS/BCP) hybrid scaffold to construct tissue-engineered bone and evaluated whether the combination promotes bone regeneration in large segmental bone defects in rabbits. The results demonstrated that human UDSCs can differentiate into osteoblasts, and the human UDSCs adhered, proliferated, and differentiated on CS/BCP hybrid scaffolds. Micro-CT, biomechanical detection, and histological analyses revealed that the combination of human UDSCs and the CS/BCP hybrid scaffold enhanced bone regeneration more effectively compared with conventional pure BCP scaffolds, indicating that human UDSCs can be used as a cell source for bone tissue engineering [150]. Wu S et al. reported an injectable BMP2-releasing chitosan microspheres/type I collagen hydrogel (BMP2-CSM/Col I hydrogel) loaded with human UDSCs for bone regeneration. The results showed human UDSCs proliferated in a time-dependent fashion, spread with good extension, and interconnected with each other in different hydrogels both for 2D and 3D models. Sustained-released BMP2 increased the ALP activities and mineral depositions of UDSCs in 2D culture, and enhanced the expression of osteogenic genes and proteins in 3D culture [151]. In vivo, the mixture of human UDSCs and BMP2-CSM/Col I hydrogels effectively enhanced bone regeneration, and the ratio of new bone volume to total bone volume was 38% after 8 weeks of implantation, although the defect was not completely healed. Further, human UDSCs differentiated into osteoblasts in the newly regenerated bone as demonstrated by human nuclear positive staining [151].
Xing F et al. fabricated a 3D-printed poly(ε-caprolactone) (PCL) scaffold with a nano-topographical surface and loaded it with human UDSCs for bone regeneration. The topological 3D-printed PCL scaffolds (TPS), fabricated by surface epiphytic crystallization, possessed uniformly patterned nanoridges, with an element composition and functional grouping of nanoridges the same as the native PCL. Compared with bare 3D-printed PCL scaffolds (BPS), TPS have a higher ability for protein adsorption and mineralization in vitro[152]. The proliferation, cell length, and osteogenic gene expression of UDSCs on the surface of TPS were significantly higher than that of BPS. TPS loaded with UDSCs exhibited enhanced bone regeneration in cranial bone defects when compared to the BPS/UDSCs group and the scaffold-only group, but the new regenerated bone appeared as mesh [152].
Zhang X et al. fabricated a 3D-printed polylactic acid and hydroxyapatite (PLA/HA) composite scaffold with human UDSCs to study its therapeutic effect in a rat model of skull defects (5mm). Human UDSCs were inoculated onto PLA/HA and PLA scaffolds using 3D printing and implanted into a 5 mm calvarial bone defect of Sprague-Dawley rats. The results demonstrated that the PLA/HA scaffold loaded with human UDSCs effectively promoted new bone regeneration in the defect area[153]. Micro-CT images showed that in the PLA/HA group, the defect area was almost entirely covered by newly formed bone (coverage of 96.7 ± 1.6%), and the coverage was greater than that in the PLA group (coverage of 74.6 ± 1.9%) at 12 weeks [153].
10.2. UDSC exosomes for bone tissue engineering.
Li H et al. reported that human UDSC-derived extracellular vesicles (UDSC-EVs) exhibited a cup-like morphology with a double-layered membrane structure, positive for CD63 and TSG101 and negative for calnexin. In vitro, UDSC-EVs promoted the osteogenic differentiation of BMSCs and reduced proinflammatory factor production and osteoclastic activity in RAW264.7 macrophage cells [154]. In vivo, local injection of UDSC-EVs around the central sites of the calvaria of ultra-high-molecular-weight polyethylene (UHMWPE) particle-induced osteolysis decreased inflammatory cytokine generation and osteolysis compared with the control groups, and significantly increased bone formation [154].
Human UDSC-derived exosomes delivered with Gelatin methacrylate (GelMA) and hyaluronic acid methacrylate (HAMA)/nano-hydroxyapatite (nHAP) hydrogels could promote the osteogenesis of bone marrow mesenchymal stem cells (BMMSCs) and the angiogenesis of endothelial progenitor cells (EPCs), respectively, in vitro[155]. Additionally, the in vivo results demonstrated that this composite hydrogel could significantly promote the defect repair of cranial bone in the rat model. It was also found that UDSC-EXOs/GelMA-HAMA/nHAP composite hydrogel can promote the formation of H-type vessels in the bone regeneration area, enhancing the therapeutic effect of bone regeneration [155].
11. Comparison of bone regenerative potential of different stem cells
Given stem cells can be isolated from almost every tissue. What cells are more effective remains a critical, unanswered question. Several studies have compared different source of stem cells for bone regeneration.
Hayashi et al. compared the osteogenic differentiation of rat MSCs from bone marrow, periosteum, and adipose tissues in vitro and in vivo and found that MSCs from bone marrow and periosteum were more osteogenic and formed significantly more bone in vivo than ADSCs [156]. Stockmann P et al. demonstrated that culture expanded PSCs were as efficient as ADSCs and BMMSCs in terms of treatment of unicortical calvarial defects [157]. Another group found that, in critical-size sheep tibiae defects, ADSCs were not as efficient as BMMSCs for defect healing when seeded within a collagen scaffold, but the ADSCs was much more efficient in bone healing when combined with platelet-rich plasma [158]. Xu L et al. compared the human BMMSCs and ADSCs isolated from bone marrow and adipose tissue obtained after total hip arthroplasty patients for epigenetic differences and in vivo bone regeneration. They found that BMMSCs possessed stronger osteogenic and lower adipogenic differentiation potentials compared to ADSCs. There was no significant difference in the chondrogenic differentiation potential[159]. The CpG sites of the RUNX2 promoter in BMMSCs were hypomethylated, while in ADSCs they were hypermethylated. The CpG sites of the PPARγ promoter in ADSCs were hypomethylated, while in BMMSCs they were hypermethylated. The methylation status of the Sox9 promoter in BMMSCs was only slightly lower than that of the ADSCs. In vivo, BMMSCs regenerated more bone in a critical-size bone defect model in mice than ADSCs [159]. Mohamed-Ahmed S et al. compared osteogenesis of human ADSCs and BMMSCs from the same donors using poly(L-lactide-co-ε-caprolactone) scaffolds both in vitro and in vivo. They found both ADSCs and BMMSCs demonstrated mineralization in vitro. However, BMMSCs showed higher ALP activity than ADSCs. In vivo, human osteogenesis-related genes RUNX2 and collagen type I were expressed in the defects with scaffold/cells. Defects with scaffold/BMMSCs had higher cellular activity than defects with scaffold/ADSCs. Moreover, bone formation in defects with scaffold/BMMSCs was greater than in defects with scaffold/ADSCs, especially at the early timepoints. These results suggest that although ADSCs have the potential to regenerate bone, the rate of bone regeneration with ADSCs may be slower than with BMMSCs[160]. They conclude that BMMSCs are more suitable for bone regenerative applications [160]. Rodrigues MT et al. compared osteogenic differentiation of BMMSCs and AFDSCs using both two and three dimensions using a tissue culture plate and by seeding the cells onto microfibrous starch and poly(ɛ-caprolactone) scaffolds (a blend of starch and polycaprolactone), respectively. BMMSCs and AFDSCs were successfully differentiated into the osteogenic cells, as revealed by production of a mineralized extracellular matrix[161]. However,, the two types of cells presented different expression patterns of bone-related markers as well as different timing of differentiation, indicating that both cell origin and the culture environment have a significant impact on the differentiation into the osteogenic phenotype in AFDSCs and BMMSCs [161]. Mohammed EEA also compared human AFDSCs and BMMSCs for their repair capacities for rat lumbar spine defects using a gel-foam scaffold. The results showed that human AFDSCs are more effective than the human BMMSCs for spine fusion repair [162]. Maraldi T et al. compared DPSCs and AFDSCs for bone regeneration in critical-sized calvarial bone defect using collagen as scaffold. The authors found that both DPSCs and AFDSCs promoted bone regeneration by direct differentiation into osteoblasts and increased blood vessel formation in the regenerated bone area using human mitochondria as a tracing marker [97].
Nakajima K et al. compared stem cells from human exfoliated deciduous teeth (SHED) to that of human DPSCs and BMMSCs for bone regeneration using a polylactic-coglycolic acid barrier membrane as a scaffold in 4 mm calvaria defects of immunodeficient mice. Micro-CT results showed the degree of bone regeneration with SHED relative to the bone defect was almost equivalent to that with human DPSCs and BMMSCs 12 weeks after transplantation. The ratio of new bone formation relative to the pre-created bone defect was not significantly different among groups with SHED, hDPSCs and hBMMSCs. In addition, histology demonstrated SHED produced the greatest amount of osteoid and widely distributed collagen fibers compared to human DPSC and BMMSC groups. Thus, SHED transplantation exerted bone regeneration ability sufficient for the repair of bone defects [163]. Lee Y et al. compared BMMSC and DPSC cell morphology, cell proliferation, trilineage differentiation, mineral synthesis, and osteogenic gene expression in vitro and bone regeneration in vivo using Bio-Oss as scaffold. It was shown the BMMSCs and DPSCs exhibited similar morphology, proliferative ability, surface marker profile, and trilineage-differentiation potential in vitro. However, the BMMSCs exhibited a higher mineral deposition and expression levels of osteogenic marker genes, including ALP, RUNX2, and osteocalcin (OCN)[164]. In the in vivo studies, the new bone volume density in both cells groups was significantly greater than that in the empty control or Bio-Oss only group. Moreover, the new bone formation and Collagen I/osteoprotegerin protein expressions of the Bio-Oss BMMSCs or Bio-Oss-DPSCs groups were higher than those of the Bio-Oss only group[164]. Finally, the Bio-Oss+BMMSCs and Bio-Oss+DPSCs groups had a similar bone mineral density, new bone formation, and osteogenesis-related protein expression[164].
Vater C et al. also compared DPSC and BMMSC for proliferation and bone regeneration in a critical-size calvarial bone defect. The authors found that DPSCs showed a 2-fold lower population doubling time and a 9-fold higher increase in proliferation when seeded onto mineralized collagen matrix (MCM) scaffolds as compared to BMMSCs, but DPSCs showed a significantly lower osteogenic capability than BMMSCs. However, pre-seeding of MCM scaffolds with DPSCs and BMMSCs did not enhance bone defect healing in vivo as the healing of the critical-size bone defect in NMRI nude mice was comparable among all groups[165] . Another study compared bone regeneration of the DPSCs and BMMSCs using MBCP and Bio-Oss scaffolds in a rabbit calvarial bone defect model. Despite the inferior bone regenerative capacity of DPSCs and BMMSCs at early timepoints after bone injury compared to autologous bone grafting; at 8 weeks, the efficiency of the BMMSCs combined with MBCP and Bio-Oss was comparable to that of the autogenous bone after 8 weeks of healing. DPSCs in combination with both scaffolds showed slightly inferior bone formation compared to autologous bone grafting[166].
Furthermore, Zhu Y et al. compared ADSCs and DPSCs for bone regeneration using bovine-derived xenografts with 10% porcine collagen as scaffold. The study found that although DPSCs had higher proliferative abilities, ADSCs exhibited greater mineral depositions and higher osteogenic-related gene expression, indicating better osteogenic differentiation potential of ADSCs [167]. After applying cryopreserved ADSCs and DPSCs in a critical-sized calvarial defect model, both cryopreserved mesenchymal stem cells significantly improved bone volume density and new bone area at 2, 4, and 8 weeks. Furthermore, the combined treatment with ADSCs and xenografts was more efficient in enhancing bone repair compared to combined treatment with DPCSs at all timepoints[167]. The authors further evaluated the sequential early bone healing process both histologically and radiographically, confirming a high agreement between these two methods which support the conclusion [167].
Nyberg E et al. compared the stromal vascular fraction (SVF) cells obtained from human lipoaspirate (point of care) and ADSCs purified from the more heterogeneous population of SVF cells. Isogenic SVF and ADSCs were suspended in fibrin hydrogels and seeded in three-dimensional-printed osteoinductive scaffolds of decellularized bone matrix and polycaprolactone. The results showed that both cell populations successfully mineralized the scaffold in vitro, demonstrating the robust bone formation properties of SVF[168]. In murine critical-sized cranial defects, ADSC-loaded scaffolds had greater (but not statistically significant) bone volume and bone coverage area than SVF-loaded scaffolds. However, both cell-laden interventions resulted in significantly greater bone healing than the contralateral acellular controls [168]. Zhang Y et al. also compared SVFs from human adipose tissue and hADSCs derived from the same donor and their interaction with human monocytes to mimic the clinical setting of intraoperative preparation of bone constructs to repair bone. The results demonstrated that SVF displayed equal in vitro osteogenic differentiation compared to donor-matched expanded ADSCs, which for both was significantly enhanced upon co-culture with monocytes. Moreover, SVFs and ADSCs displayed different immunoregulatory effects on monocytes/macrophages [169]. Upon implantation in rat femoral bone defects, SVF constructs demonstrated superior bone formation compared to ADSC constructs and cell-free controls and there were no effects observed with monocyte addition[169].
Gao X et al. compared bone regeneration of Lenti-BMP2 transduced human BMMSCs and MDSCs in a critical-size mouse calvarial bone defect model using fibrin sealant as scaffold. The authors found both Lenti-BMP2 transduced BMMSCs and MDSCs regenerated functional bone in 6 weeks, with near complete defect healing. No significant differences were found in term of new bone volume and defect healing percentage between Lenti-BMP2 transduced human BMMSCs and MDSCs[55]. However, non-transduced human BMMSCs and MDSCs all formed negligible new bone which indicated BMP2 signaling are required [55]. Lough D et al. compared MDSCs with ADSCs and BMMSCs isolated from same mice for bone regeneration. The authors found that all populations exhibited mesenchymal stem cell multilineage capacity, but ADSC- and BMMSC-enriched constructs were capable of forming small bone aggregates . In contrast, MDSCs self-assembled a form of organized cortico-cancellous bone structures within two- and three-dimensional in vitro systems. MDSCs also augmented defect healing, angiogenesis, and diploic space formation in a cranial defect mice model in vivo [170].
Agata H et al. compared the bone regeneration capacities of BMMSCs and PSCs and found that PSCs were capable of osteogenic differentiation in vitro, although less efficiently than BMMSCs; however, when PSCs were pretreated with FGF2 and BMP2, they induced greater bone formation in vivo when compared to the BMMSCs [171]. González-Gil AB et al. compared the therapeutic potential of PSCs and BMMSCs in combination with biomaterials in a bone nonunion model. PSCs, BMMSCs and bone graft were isolated from green fluorescent protein (GFP)-transgenic rats. Animals were divided into six groups, depending on the treatment to be applied: bone defect left empty (CTL); treated with live bone allograft (LBA); hrBMP-2 in collagen scaffold (CSBMP2); acellular polycaprolactone scaffold (PCL group); PCL scaffold containing PSCs (PCL-PSCs) and PCL containing BMMSCs (PCL-BMMSCs). In the LBA group, all the animals showed bone bridging (n = 6) whereas in the CSBMP2 group, four out of six animals demonstrated healing. In the PCL and PCLPSC groups, a reduced number of animals showed radiological healing, whereas no healing was detected in the PCL-BMMSC group. MicroCT results showed significant new bone formation in the LBA, CSBMP2, and PCL-PSC groups when compared with the CTL group. At 10 weeks, GFP-positive cells were detected only in the LBA group and were restricted to the outer cortical bone in close contact with the periosteum. Tracking of cellular implants demonstrated significant higher survival of the PSCs when compared with BMMSCs [172].
In summary, BMMSCs are still the most commonly used postnatal stem cells. BMMSCs are more efficient than ADSCs for bone regeneration and are equivalent to PSCs and MDSCs, but slightly better than DPSCs. AFDSCs are more effective than BMMSCs and comparable to DPSCs for bone regeneration.
12. Advantages and disadvantages of different stem cells for potential clinical applications.
In summary, many sources of stem cells are available for bone tissue engineering. The source of the stem cells to be utilized in clinical practice will depend on the patient’s needs and availability of donor tissues. BMMSCs are the most commonly used and overall are most effective in term of promoting bone regeneration. PSCs are also as effective as BMMSCs, but its isolation procedures are invasive[157]. MDSCs are often available during treatment of orthopaedic trauma and can be isolated with easy muscle access and regenerate functional bone efficiently but require BMP4 or BMP2 stimulation[55]. ADSCs are easily available as point of care despite less effective than BMMSCs. DPSCs are also attractive due to their nearly similar efficacy as BMMSCs for bone regeneration[167]. They represent the great cells source for dental bone loss or craniofacial bone reconstruction. UC-MSCs are also promising because their harvest is not invasive[137]. They can be used for both mother and infant (children) treatment and allow for stem cell banking for later use. Most recent studies on PBMSCs for bone regeneration are also encouraging due to their effectiveness and relatively easy availability for isolation. Recent investigations on UDSCs are especially encouraging in that they are effective, easily available at point of care, non-invasive, cheap cost to isolate, and suitable for stem cell banking.
13. Prospective applications of stem cells for bone tissue engineering for human bone tissue repair.
Despite extensive preclinical studies using many different stem cells from different tissue resources, clinical applications of stem cells are still limited due to most of the stem cells’ need culture and expansion. Therefore, development of new methods or devices allowing for point of care isolation of stem cells for bone defect repair or non-union fracture repair is critical. For example, Zhang Y et al. reported a point of care device for isolating and processing BMMSCs and forming a composite with a scaffold in 5 minutes and achieved clinical satisfactory bone repair for 42 patients [173]. Furthermore, stem cells banking of different stem cells for future application is also an important strategy. UC-MSCs, PBMSCs, UDSCs, DPSCs, MDSCs, ADSCs and BMMSCs are all excellent stem cells resources and suitable for stem cell banking. The choice of scaffold to deliver cells is also important. Fibrin sealant scaffolds are FDA approved (such as Tisseel Fibrin Sealant) and when used for delivery of cells or growth factors, it is easily absorbable while the newly formed bone is similar to native bone with normal blood vessel anatomy and bone marrow [55,57,174]. BCP or TCP or hydroxyapatite scaffolds are not easily absorbable and often remain in the new regenerated bone, these residues do not integrate with host bone, and likely offer inferior bone biomechanical properties. Ideally bone tissue engineering using stem cells will need the stem cells from both the donor and the host to differentiate into osteoblasts and secrete collagen I and other organic bone matrix which then mineralizes to form functional bone. Orchestration of osteogenesis and angiogenesis are also important. The combination of stem cells with bone growth factors and scaffolds will be most effective. Finally, exosome or extracellular vesicles derived from stem cells offers a cell free strategy of delivering osteogenic cargos to enhance bone formation. This approach is also promising due to its nature of not eliciting an immune response when used allogenically.
Author Contributions
Conceptualization, X.G. and J.H.; Software, X.X and X.G.; Validation, X.G. and J.H.; Writing, Original Draft Preparation, X.G.; Writing, Review and Editing, J.J,R, J.E.L,J.H.; Supervision, J.H.; Project Administration, J.H.; Funding Acquisition, J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by Philanthropic gifts from Mary Sue and Mike Shannon Network.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This project was funded by a Philanthropic gift from Mary Sue and Mike Shannon Network.
Conflicts of Interest
J.H. received royalties from Cook Myocyte. All other authors declare no conflicts of interest.
References
- Connolly, J.F.; Guse, R.; Tiedeman, J.; Dehne, R. Autologous marrow injection as a substitute for operative grafting of tibial nonunions. Clin Orthop Relat Res 1991, 259–270. [Google Scholar] [CrossRef]
- Tiedeman, J.J.; Garvin, K.L.; Kile, T.A.; Connolly, J.F. The role of a composite, demineralized bone matrix and bone marrow in the treatment of osseous defects. Orthopedics 1995, 18, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Quarto, R.; Mastrogiacomo, M.; Cancedda, R.; Kutepov, S.M.; Mukhachev, V.; Lavroukov, A.; Kon, E.; Marcacci, M. Repair of large bone defects with the use of autologous bone marrow stromal cells. N Engl J Med 2001, 344, 385–386. [Google Scholar] [CrossRef]
- Yang, Y.; Hallgrimsson, B.; Putnins, E.E. Craniofacial defect regeneration using engineered bone marrow mesenchymal stromal cells. J Biomed Mater Res A 2011, 99, 74–85. [Google Scholar] [CrossRef]
- Burastero, G.; Scarfi, S.; Ferraris, C.; Fresia, C.; Sessarego, N.; Fruscione, F.; Monetti, F.; Scarfo, F.; Schupbach, P.; Podesta, M. , et al. The association of human mesenchymal stem cells with BMP-7 improves bone regeneration of critical-size segmental bone defects in athymic rats. Bone 2010, 47, 117–126. [Google Scholar] [CrossRef]
- Scotti, C.; Tonnarelli, B.; Papadimitropoulos, A.; Scherberich, A.; Schaeren, S.; Schauerte, A.; Lopez-Rios, J.; Zeller, R.; Barbero, A.; Martin, I. Recapitulation of endochondral bone formation using human adult mesenchymal stem cells as a paradigm for developmental engineering. Proc Natl Acad Sci U S A 2010, 107, 7251–7256. [Google Scholar] [CrossRef] [PubMed]
- Scotti, C.; Piccinini, E.; Takizawa, H.; Todorov, A.; Bourgine, P.; Papadimitropoulos, A.; Barbero, A.; Manz, M.G.; Martin, I. Engineering of a functional bone organ through endochondral ossification. Proc Natl Acad Sci U S A 2013, 110, 3997–4002. [Google Scholar] [CrossRef]
- Long, T.; Zhu, Z.; Awad, H.A.; Schwarz, E.M.; Hilton, M.J.; Dong, Y. The effect of mesenchymal stem cell sheets on structural allograft healing of critical sized femoral defects in mice. Biomaterials 2014, 35, 2752–2759. [Google Scholar] [CrossRef]
- Lin, H.; Sohn, J.; Shen, H.; Langhans, M.T.; Tuan, R.S. Bone marrow mesenchymal stem cells: Aging and tissue engineering applications to enhance bone healing. Biomaterials 2019, 203, 96–110. [Google Scholar] [CrossRef]
- Arthur, A.; Gronthos, S. Clinical Application of Bone Marrow Mesenchymal Stem/Stromal Cells to Repair Skeletal Tissue. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Stamnitz, S.; Klimczak, A. Mesenchymal Stem Cells, Bioactive Factors, and Scaffolds in Bone Repair: From Research Perspectives to Clinical Practice. Cells 2021, 10. [Google Scholar] [CrossRef]
- Blanco, J.F.; Garcia-Brinon, J.; Benito-Garzon, L.; Pescador, D.; Muntion, S.; Sanchez-Guijo, F. Human Bone Marrow Mesenchymal Stromal Cells Promote Bone Regeneration in a Xenogeneic Rabbit Model: A Preclinical Study. Stem Cells Int 2018, 2018, 7089484. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liu, X.; Zhao, K.; Zhu, Y.; Hu, B.; Zhou, Y.; Wang, M.; Wu, Y.; Zhang, C.; Xu, J. , et al. miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1alpha pathway and enhances bone regeneration in critical size defects. Stem Cell Res Ther 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.Y.; Lu, B.; Yin, J.H.; Ke, Q.F.; Xu, H.; Zhang, C.Q.; Guo, Y.P.; Gao, Y.S. Gadolinium-doped bioglass scaffolds promote osteogenic differentiation of hBMSC via the Akt/GSK3beta pathway and facilitate bone repair in vivo. Int J Nanomedicine 2019, 14, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Wang, Q.; Ouyang, L.; Wu, H.; Yang, Z.; Fu, X.; Liu, X.; Yan, L.; Cao, Y.; Xiao, R. Comparison of concentrated fresh mononuclear cells and cultured mesenchymal stem cells from bone marrow for bone regeneration. Stem Cells Transl Med 2021, 10, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, R.; Tan, X.; Li, B.; Liu, Y.; Wang, X. Synthesis and Evaluation of BMMSC-seeded BMP-6/nHAG/GMS Scaffolds for Bone Regeneration. Int J Med Sci 2019, 16, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Monahan, D.S.; Brulin, B.; Gallinetti, S.; Humbert, P.; Tringides, C.; Canal, C.; Ginebra, M.P.; Layrolle, P. Biomimetic versus sintered macroporous calcium phosphate scaffolds enhanced bone regeneration and human mesenchymal stromal cell engraftment in calvarial defects. Acta Biomater 2021, 135, 689–704. [Google Scholar] [CrossRef]
- Li, G.; Shen, W.; Tang, X.; Mo, G.; Yao, L.; Wang, J. Combined use of calcium phosphate cement, mesenchymal stem cells and platelet-rich plasma for bone regeneration in critical-size defect of the femoral condyle in mini-pigs. Regen Med 2021, 16, 451–464. [Google Scholar] [CrossRef]
- Li, Z.; Xiang, S.; Lin, Z.; Li, E.N.; Yagi, H.; Cao, G.; Yocum, L.; Li, L.; Hao, T.; Bruce, K.K. , et al. Graphene oxide-functionalized nanocomposites promote osteogenesis of human mesenchymal stem cells via enhancement of BMP-SMAD1/5 signaling pathway. Biomaterials 2021, 277, 121082. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, X.; Fritch, M.R.; Li, Z.; Kuang, B.; Alexander, P.G.; Hao, T.; Cao, G.; Tan, S.; Bruce, K.K. , et al. Engineering pre-vascularized bone-like tissue from human mesenchymal stem cells through simulating endochondral ossification. Biomaterials 2022, 283, 121451. [Google Scholar] [CrossRef]
- Bernhard, J.C.; Marolt Presen, D.; Li, M.; Monforte, X.; Ferguson, J.; Leinfellner, G.; Heimel, P.; Betti, S.L.; Shu, S.; Teuschl-Woller, A.H. , et al. Effects of Endochondral and Intramembranous Ossification Pathways on Bone Tissue Formation and Vascularization in Human Tissue-Engineered Grafts. Cells 2022, 11. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Li, M.; Song, P.; Lei, H.; Gui, X.; Zhou, C.; Liu, L. Biomimetic Methacrylated Gelatin Hydrogel Loaded With Bone Marrow Mesenchymal Stem Cells for Bone Tissue Regeneration. Front Bioeng Biotechnol 2021, 9, 770049. [Google Scholar] [CrossRef]
- Machado-Paula, M.M.; Corat, M.A.F.; de Vasconcellos, L.M.R.; Araujo, J.C.R.; Mi, G.; Ghannadian, P.; Toniato, T.V.; Marciano, F.R.; Webster, T.J.; Lobo, A.O. Rotary Jet-Spun Polycaprolactone/Hydroxyapatite and Carbon Nanotube Scaffolds Seeded with Bone Marrow Mesenchymal Stem Cells Increase Bone Neoformation. ACS Appl Bio Mater 2022, 5, 1013–1024. [Google Scholar] [CrossRef]
- Liu, X.; Li, L.; Gaihre, B.; Park, S.; Li, Y.; Terzic, A.; Elder, B.D.; Lu, L. Scaffold-Free Spheroids with Two-Dimensional Heteronano-Layers (2DHNL) Enabling Stem Cell and Osteogenic Factor Codelivery for Bone Repair. ACS Nano 2022, 16, 2741–2755. [Google Scholar] [CrossRef]
- Liu, Y.; Kuang, B.; Rothrauff, B.B.; Tuan, R.S.; Lin, H. Robust bone regeneration through endochondral ossification of human mesenchymal stem cells within their own extracellular matrix. Biomaterials 2019, 218, 119336. [Google Scholar] [CrossRef]
- Yang, C.; Li, Z.; Liu, Y.; Hou, R.; Lin, M.; Fu, L.; Wu, D.; Liu, Q.; Li, K.; Liu, C. Single-cell spatiotemporal analysis reveals cell fates and functions of transplanted mesenchymal stromal cells during bone repair. Stem Cell Reports 2022, 17, 2318–2333. [Google Scholar] [CrossRef] [PubMed]
- Pitacco, P.; Sadowska, J.M.; O'Brien, F.J.; Kelly, D.J. 3D bioprinting of cartilaginous templates for large bone defect healing. Acta Biomater 2023, 156, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Herberg, S.; Varghai, D.; Alt, D.S.; Dang, P.N.; Park, H.; Cheng, Y.; Shin, J.Y.; Dikina, A.D.; Boerckel, J.D.; Rolle, M.W. , et al. Scaffold-free human mesenchymal stem cell construct geometry regulates long bone regeneration. Commun Biol 2021, 4, 89. [Google Scholar] [CrossRef]
- Whitehead, J.; Griffin, K.H.; Gionet-Gonzales, M.; Vorwald, C.E.; Cinque, S.E.; Leach, J.K. Hydrogel mechanics are a key driver of bone formation by mesenchymal stromal cell spheroids. Biomaterials 2021, 269, 120607. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Lee, C.S.; Kim, S.; Chen, C.; Aghaloo, T.; Lee, M. Generation of Small RNA-Modulated Exosome Mimetics for Bone Regeneration. ACS Nano 2020, 14, 11973–11984. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.; Zhu, Y.; Yang, M.; Mao, C. Human Mesenchymal Stem Cell Derived Exosomes Enhance Cell-Free Bone Regeneration by Altering Their miRNAs Profiles. Adv Sci (Weinh) 2020, 7, 2001334. [Google Scholar] [CrossRef]
- Liu, A.; Lin, D.; Zhao, H.; Chen, L.; Cai, B.; Lin, K.; Shen, S.G. Optimized BMSC-derived osteoinductive exosomes immobilized in hierarchical scaffold via lyophilization for bone repair through Bmpr2/Acvr2b competitive receptor-activated Smad pathway. Biomaterials 2021, 272, 120718. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Kang, M.; Shirazi, S.; Lu, Y.; Cooper, L.F.; Gajendrareddy, P.; Ravindran, S. 3D Encapsulation and tethering of functionally engineered extracellular vesicles to hydrogels. Acta Biomater 2021, 126, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xin, X.; Wang, L.; Wang, B.; Chen, L.; Liu, O.; Rowe, D.W.; Xu, M. Senolytics improve bone forming potential of bone marrow mesenchymal stem cells from aged mice. NPJ Regen Med 2021, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Huang, H.; Gao, X.; Yang, J.; Tang, Q.; Xu, X.; Wu, Y.; Li, M.; Liang, C.; Tan, L. , et al. Local Elimination of Senescent Cells Promotes Bone Defect Repair during Aging. ACS Appl Mater Interfaces 2022, 14, 3885–3899. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D. Retention of differentiation potentialities during prolonged cultivation of myogenic cells. Proc Natl Acad Sci U S A 1968, 61, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D.; Saxel, O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef] [PubMed]
- Blau, H.M.; Chiu, C.P.; Webster, C. Cytoplasmic activation of human nuclear genes in stable heterocaryons. Cell 1983, 32, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Rando, T.A.; Blau, H.M. Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J Cell Biol 1994, 125, 1275–1287. [Google Scholar] [CrossRef]
- Torrente, Y.; Tremblay, J.P.; Pisati, F.; Belicchi, M.; Rossi, B.; Sironi, M.; Fortunato, F.; El Fahime, M.; D'Angelo, M.G.; Caron, N.J. , et al. Intraarterial injection of muscle-derived CD34(+)Sca-1(+) stem cells restores dystrophin in mdx mice. J Cell Biol 2001, 152, 335–348. [Google Scholar] [CrossRef]
- Lee, J.Y.; Musgrave, D.; Pelinkovic, D.; Fukushima, K.; Cummins, J.; Usas, A.; Robbins, P.; Fu, F.H.; Huard, J. Effect of bone morphogenetic protein-2-expressing muscle-derived cells on healing of critical-sized bone defects in mice. J Bone Joint Surg Am 2001, 83, 1032–1039. [Google Scholar] [CrossRef]
- Qu-Petersen, Z.; Deasy, B.; Jankowski, R.; Ikezawa, M.; Cummins, J.; Pruchnic, R.; Mytinger, J.; Cao, B.; Gates, C.; Wernig, A. , et al. Identification of a novel population of muscle stem cells in mice: potential for muscle regeneration. J Cell Biol 2002, 157, 851–864. [Google Scholar] [CrossRef]
- Qu, Z.; Balkir, L.; van Deutekom, J.C.; Robbins, P.D.; Pruchnic, R.; Huard, J. Development of approaches to improve cell survival in myoblast transfer therapy. J Cell Biol 1998, 142, 1257–1267. [Google Scholar] [CrossRef]
- Wright, V.; Peng, H.; Usas, A.; Young, B.; Gearhart, B.; Cummins, J.; Huard, J. BMP4-expressing muscle-derived stem cells differentiate into osteogenic lineage and improve bone healing in immunocompetent mice. Mol Ther 2002, 6, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Wright, V.; Usas, A.; Gearhart, B.; Shen, H.C.; Cummins, J.; Huard, J. Synergistic enhancement of bone formation and healing by stem cell-expressed VEGF and bone morphogenetic protein-4. J Clin Invest 2002, 110, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Usas, A.; Olshanski, A.; Ho, A.M.; Gearhart, B.; Cooper, G.M.; Huard, J. VEGF improves, whereas sFlt1 inhibits, BMP2-induced bone formation and bone healing through modulation of angiogenesis. J Bone Miner Res 2005, 20, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Usas, A.; Hannallah, D.; Olshanski, A.; Cooper, G.M.; Huard, J. Noggin improves bone healing elicited by muscle stem cells expressing inducible BMP4. Mol Ther 2005, 12, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Usas, A.; Proto, J.D.; Lu, A.; Cummins, J.H.; Proctor, A.; Chen, C.W.; Huard, J. Role of donor and host cells in muscle-derived stem cell-mediated bone repair: differentiation vs. paracrine effects. FASEB J 2014, 28, 3792–3809. [Google Scholar] [CrossRef] [PubMed]
- Corsi, K.A.; Pollett, J.B.; Phillippi, J.A.; Usas, A.; Li, G.; Huard, J. Osteogenic potential of postnatal skeletal muscle-derived stem cells is influenced by donor sex. J Bone Miner Res 2007, 22, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Meszaros, L.B.; Usas, A.; Cooper, G.M.; Huard, J. Effect of host sex and sex hormones on muscle-derived stem cell-mediated bone formation and defect healing. Tissue Eng Part A 2012, 18, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Peng, H.; Usas, A.; Musgrave, D.; Cummins, J.; Pelinkovic, D.; Jankowski, R.; Ziran, B.; Robbins, P.; Huard, J. Enhancement of bone healing based on ex vivo gene therapy using human muscle-derived cells expressing bone morphogenetic protein 2. Hum Gene Ther 2002, 13, 1201–1211. [Google Scholar] [CrossRef]
- Mastrogiacomo, M.; Derubeis, A.R.; Cancedda, R. Bone and cartilage formation by skeletal muscle derived cells. J Cell Physiol 2005, 204, 594–603. [Google Scholar] [CrossRef]
- Gao, X.; Usas, A.; Lu, A.; Tang, Y.; Wang, B.; Chen, C.W.; Li, H.; Tebbets, J.C.; Cummins, J.H.; Huard, J. BMP2 is superior to BMP4 for promoting human muscle-derived stem cell-mediated bone regeneration in a critical-sized calvarial defect model. Cell Transplant 2013, 22, 2393–2408. [Google Scholar] [CrossRef]
- Gharaibeh, B.; Lu, A.; Tebbets, J.; Zheng, B.; Feduska, J.; Crisan, M.; Peault, B.; Cummins, J.; Huard, J. Isolation of a slowly adhering cell fraction containing stem cells from murine skeletal muscle by the preplate technique. Nat Protoc 2008, 3, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Usas, A.; Tang, Y.; Lu, A.; Tan, J.; Schneppendahl, J.; Kozemchak, A.M.; Wang, B.; Cummins, J.H.; Tuan, R.S. , et al. A comparison of bone regeneration with human mesenchymal stem cells and muscle-derived stem cells and the critical role of BMP. Biomaterials 2014, 35, 6859–6870. [Google Scholar] [CrossRef]
- Gao, X.; Lu, A.; Tang, Y.; Schneppendahl, J.; Liebowitz, A.B.; Scibetta, A.C.; Morris, E.R.; Cheng, H.; Huard, C.; Amra, S. , et al. Influences of donor and host age on human muscle-derived stem cell-mediated bone regeneration. Stem Cell Res Ther 2018, 9, 316. [Google Scholar] [CrossRef]
- Usas, A.; Ho, A.M.; Cooper, G.M.; Olshanski, A.; Peng, H.; Huard, J. Bone regeneration mediated by BMP4-expressing muscle-derived stem cells is affected by delivery system. Tissue Eng Part A 2009, 15, 285–293. [Google Scholar] [CrossRef]
- Zheng, B.; Cao, B.; Crisan, M.; Sun, B.; Li, G.; Logar, A.; Yap, S.; Pollett, J.B.; Drowley, L.; Cassino, T. , et al. Prospective identification of myogenic endothelial cells in human skeletal muscle. Nat Biotechnol 2007, 25, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Li, G.; Chen, W.C.; Deasy, B.M.; Pollett, J.B.; Sun, B.; Drowley, L.; Gharaibeh, B.; Usas, A.; Peault, B. , et al. Human myogenic endothelial cells exhibit chondrogenic and osteogenic potentials at the clonal level. J Orthop Res 2013, 31, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Jackson, W.M.; Lozito, T.P.; Djouad, F.; Kuhn, N.Z.; Nesti, L.J.; Tuan, R.S. Differentiation and regeneration potential of mesenchymal progenitor cells derived from traumatized muscle tissue. J Cell Mol Med 2011, 15, 2377–2388. [Google Scholar] [CrossRef]
- Levi, B.; Longaker, M.T. Concise review: adipose-derived stromal cells for skeletal regenerative medicine. Stem Cells 2011, 29, 576–582. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, F.; Ricci, G.; D'Andrea, F.; Nicoletti, G.F.; Ferraro, G.A. Human Adipose Stem Cells: From Bench to Bedside. Tissue Eng Part B Rev 2015, 21, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.M.; Shi, Y.Y.; Aalami, O.O.; Chou, Y.F.; Mari, C.; Thomas, R.; Quarto, N.; Contag, C.H.; Wu, B.; Longaker, M.T. Adipose-derived adult stromal cells heal critical-size mouse calvarial defects. Nat Biotechnol 2004, 22, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Lendeckel, S.; Jodicke, A.; Christophis, P.; Heidinger, K.; Wolff, J.; Fraser, J.K.; Hedrick, M.H.; Berthold, L.; Howaldt, H.P. Autologous stem cells (adipose) and fibrin glue used to treat widespread traumatic calvarial defects: case report. J Craniomaxillofac Surg 2004, 32, 370–373. [Google Scholar] [CrossRef] [PubMed]
- James, A.W.; Levi, B.; Nelson, E.R.; Peng, M.; Commons, G.W.; Lee, M.; Wu, B.; Longaker, M.T. Deleterious effects of freezing on osteogenic differentiation of human adipose-derived stromal cells in vitro and in vivo. Stem Cells Dev 2011, 20, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, S.H.; Kang, B.J.; Kim, W.H.; Yun, H.S.; Kweon, O.K. Comparison of Osteogenesis between Adipose-Derived Mesenchymal Stem Cells and Their Sheets on Poly-epsilon-Caprolactone/beta-Tricalcium Phosphate Composite Scaffolds in Canine Bone Defects. Stem Cells Int 2016, 2016, 8414715. [Google Scholar] [CrossRef]
- Orbay, H.; Busse, B.; Leach, J.K.; Sahar, D.E. The Effects of Adipose-Derived Stem Cells Differentiated Into Endothelial Cells and Osteoblasts on Healing of Critical Size Calvarial Defects. J Craniofac Surg 2017, 28, 1874–1879. [Google Scholar] [CrossRef]
- Bernhard, J.; Ferguson, J.; Rieder, B.; Heimel, P.; Nau, T.; Tangl, S.; Redl, H.; Vunjak-Novakovic, G. Tissue-engineered hypertrophic chondrocyte grafts enhanced long bone repair. Biomaterials 2017, 139, 202–212. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, P.; Long, Y.; Huang, C.; Chen, D. Repair of bone defects in rat radii with a composite of allogeneic adipose-derived stem cells and heterogeneous deproteinized bone. Stem Cell Res Ther 2018, 9, 79. [Google Scholar] [CrossRef]
- Ruminski, S.; Kalaszczynska, I.; Dlugosz, A.; Lewandowska-Szumiel, M. Osteogenic differentiation of human adipose-derived stem cells in 3D conditions - comparison of spheroids and polystyrene scaffolds. Eur Cell Mater 2019, 37, 382–401. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Y.; Yu, N.; Ma, H.; Wang, K.; Liu, J.; Zhang, W.; Cai, Z.; He, Y. Construction of vascularized tissue-engineered bone with polylysine-modified coral hydroxyapatite and a double cell-sheet complex to repair a large radius bone defect in rabbits. Acta Biomater 2019, 91, 82–98. [Google Scholar] [CrossRef]
- Probst, F.A.; Fliefel, R.; Burian, E.; Probst, M.; Eddicks, M.; Cornelsen, M.; Riedl, C.; Seitz, H.; Aszódi, A.; Schieker, M. , et al. Bone regeneration of minipig mandibular defect by adipose derived mesenchymal stem cells seeded tri-calcium phosphate- poly(D,L-lactide-co-glycolide) scaffolds. Sci Rep 2020, 10, 2062. [Google Scholar] [CrossRef]
- Peterson, B.; Zhang, J.; Iglesias, R.; Kabo, M.; Hedrick, M.; Benhaim, P.; Lieberman, J.R. Healing of critically sized femoral defects, using genetically modified mesenchymal stem cells from human adipose tissue. Tissue Eng 2005, 11, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.N.; Yu, F.J.; Chang, Y.H.; Huang, K.L.; Pham, N.N.; Truong, V.A.; Lin, M.W.; Kieu Nguyen, N.T.; Hwang, S.M.; Hu, Y.C. CRISPR interference-mediated noggin knockdown promotes BMP2-induced osteogenesis and calvarial bone healing. Biomaterials 2020, 252, 120094. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.F.; Zuk, P.A.; Chang, T.L.; Benhaim, P.; Wu, B.M. Adipose-derived stem cells and BMP2: part 1. BMP2-treated adipose-derived stem cells do not improve repair of segmental femoral defects. Connect Tissue Res 2011, 52, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kang, B.J.; Kim, W.H.; Yun, H.S.; Kweon, O.K. Evaluation of Mesenchymal Stem Cell Sheets Overexpressing BMP-7 in Canine Critical-Sized Bone Defects. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Osinga, R.; Di Maggio, N.; Todorov, A.; Allafi, N.; Barbero, A.; Laurent, F.; Schaefer, D.J.; Martin, I.; Scherberich, A. Generation of a Bone Organ by Human Adipose-Derived Stromal Cells Through Endochondral Ossification. Stem Cells Transl Med 2016, 5, 1090–1097. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Ahmad, T.; Madhurakkat Perikamana, S.K.; Lee, J.; Kim, E.M.; Shin, H. Human adipose-derived stem cell spheroids incorporating platelet-derived growth factor (PDGF) and bio-minerals for vascularized bone tissue engineering. Biomaterials 2020, 255, 120192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.L.; He, R.Z.; Tu, B.; He, J.S.; Cao, X.; Xia, H.S.; Ba, H.L.; Wu, S.; Peng, C.; Xiong, K. Drilling Combined with Adipose-derived Stem Cells and Bone Morphogenetic Protein-2 to Treat Femoral Head Epiphyseal Necrosis in Juvenile Rabbits. Curr Med Sci 2018, 38, 277–288. [Google Scholar] [CrossRef]
- Fan, J.; Im, C.S.; Cui, Z.K.; Guo, M.; Bezouglaia, O.; Fartash, A.; Lee, J.Y.; Nguyen, J.; Wu, B.M.; Aghaloo, T. , et al. Delivery of Phenamil Enhances BMP-2-Induced Osteogenic Differentiation of Adipose-Derived Stem Cells and Bone Formation in Calvarial Defects. Tissue Eng Part A 2015, 21, 2053–2065. [Google Scholar] [CrossRef]
- Yao, W.; Lay, Y.E.; Kot, A.; Liu, R.; Zhang, H.; Chen, H.; Lam, K.; Lane, N.E. Improved Mobilization of Exogenous Mesenchymal Stem Cells to Bone for Fracture Healing and Sex Difference. Stem Cells 2016, 34, 2587–2600. [Google Scholar] [CrossRef] [PubMed]
- Negri, S.; Wang, Y.; Sono, T.; Qin, Q.; Hsu, G.C.; Cherief, M.; Xu, J.; Lee, S.; Tower, R.J.; Yu, V. , et al. Systemic DKK1 neutralization enhances human adipose-derived stem cell mediated bone repair. Stem Cells Transl Med 2021, 10, 610–622. [Google Scholar] [CrossRef]
- Levi, B.; James, A.W.; Nelson, E.R.; Li, S.; Peng, M.; Commons, G.W.; Lee, M.; Wu, B.; Longaker, M.T. Human adipose-derived stromal cells stimulate autogenous skeletal repair via paracrine Hedgehog signaling with calvarial osteoblasts. Stem Cells Dev 2011, 20, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, Y.; Zhang, P.; Tang, Y.; Zhou, M.; Jiang, W.; Zhang, X.; Wu, G.; Zhou, Y. Tissue-Engineered Bone Immobilized with Human Adipose Stem Cells-Derived Exosomes Promotes Bone Regeneration. ACS Appl Mater Interfaces 2018, 10, 5240–5254. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tang, Y.; Liu, Y.; Zhang, P.; Lv, L.; Zhang, X.; Jia, L.; Zhou, Y. Exosomes derived from miR-375-overexpressing human adipose mesenchymal stem cells promote bone regeneration. Cell Prolif 2019, 52, e12669. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Xu, C.; Meng, L.; Dong, X.; Qi, M.; Jiang, D. Exosome-functionalized magnesium-organic framework-based scaffolds with osteogenic, angiogenic and anti-inflammatory properties for accelerated bone regeneration. Bioact Mater 2022, 18, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Fu, X.; Li, X.; Li, J.; Han, W.; Wang, Y. Modification of adipose mesenchymal stem cells-derived small extracellular vesicles with fibrin-targeting peptide CREKA for enhanced bone repair. Bioact Mater 2023, 20, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Fan, J.; Liu, Y.; Li, T.; Xu, H.; Yang, Y.; Deng, L.; Li, H.; Zhao, R.C. miR-450b Promotes Osteogenic Differentiation In Vitro and Enhances Bone Formation In Vivo by Targeting BMP3. Stem Cells Dev 2018, 27, 600–611. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Q.; Zhao, Y.; Tian, Z.; Chang, S.; Tong, H.; Liu, N.; Bai, S.; Li, X.; Fan, J. Adipose-derived stem cells with miR-150-5p inhibition laden in hydroxyapatite/tricalcium phosphate ceramic powders promote osteogenesis via regulating Notch3 and activating FAK/ERK and RhoA. Acta Biomater 2023, 155, 644–653. [Google Scholar] [CrossRef]
- Liu, J.; Yu, F.; Sun, Y.; Jiang, B.; Zhang, W.; Yang, J.; Xu, G.T.; Liang, A.; Liu, S. Concise reviews: Characteristics and potential applications of human dental tissue-derived mesenchymal stem cells. Stem Cells 2015, 33, 627–638. [Google Scholar] [CrossRef]
- Tatullo, M.; Marrelli, M.; Shakesheff, K.M.; White, L.J. Dental pulp stem cells: function, isolation and applications in regenerative medicine. J Tissue Eng Regen Med 2015, 9, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Mayo, V.; Sawatari, Y.; Huang, C.Y.; Garcia-Godoy, F. Neural crest-derived dental stem cells--where we are and where we are going. J Dent 2014, 42, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- La Noce, M.; Paino, F.; Spina, A.; Naddeo, P.; Montella, R.; Desiderio, V.; De Rosa, A.; Papaccio, G.; Tirino, V.; Laino, L. Dental pulp stem cells: state of the art and suggestions for a true translation of research into therapy. J Dent 2014, 42, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Laino, G.; d'Aquino, R.; Graziano, A.; Lanza, V.; Carinci, F.; Naro, F.; Pirozzi, G.; Papaccio, G. A new population of human adult dental pulp stem cells: a useful source of living autologous fibrous bone tissue (LAB). J Bone Miner Res 2005, 20, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Walboomers, X.F.; van Osch, G.J.; van den Dolder, J.; Jansen, J.A. Hard tissue formation in a porous HA/TCP ceramic scaffold loaded with stromal cells derived from dental pulp and bone marrow. Tissue Eng Part A 2008, 14, 285–294. [Google Scholar] [CrossRef]
- Alge, D.L.; Zhou, D.; Adams, L.L.; Wyss, B.K.; Shadday, M.D.; Woods, E.J.; Gabriel Chu, T.M.; Goebel, W.S. Donor-matched comparison of dental pulp stem cells and bone marrow-derived mesenchymal stem cells in a rat model. J Tissue Eng Regen Med 2010, 4, 73–81. [Google Scholar] [CrossRef]
- Maraldi, T.; Riccio, M.; Pisciotta, A.; Zavatti, M.; Carnevale, G.; Beretti, F.; La Sala, G.B.; Motta, A.; De Pol, A. Human amniotic fluid-derived and dental pulp-derived stem cells seeded into collagen scaffold repair critical-size bone defects promoting vascularization. Stem Cell Res Ther 2013, 4, 53. [Google Scholar] [CrossRef]
- Huojia, M.; Wu, Z.; Zhang, X.; Maimaitiyiming, M.; Rong, M. Effect of Dental Pulp Stem Cells (DPSCs) in Repairing Rabbit Alveolar Bone Defect. Clin Lab 2015, 61, 1703–1708. [Google Scholar] [CrossRef]
- Jahanbin, A.; Rashed, R.; Alamdari, D.H.; Koohestanian, N.; Ezzati, A.; Kazemian, M.; Saghafi, S.; Raisolsadat, M.A. Success of Maxillary Alveolar Defect Repair in Rats Using Osteoblast-Differentiated Human Deciduous Dental Pulp Stem Cells. J Oral Maxillofac Surg 2016, 74, 829–e821. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Z.; Chen, S.; Macri, L.; Kohn, J.; Yelick, P.C. Mandibular Jaw Bone Regeneration Using Human Dental Cell-Seeded Tyrosine-Derived Polycarbonate Scaffolds. Tissue Eng Part A 2016, 22, 985–993. [Google Scholar] [CrossRef]
- Zhang, W.; Saxena, S.; Fakhrzadeh, A.; Rudolph, S.; Young, S.; Kohn, J.; Yelick, P.C. Use of Human Dental Pulp and Endothelial Cell Seeded Tyrosine-Derived Polycarbonate Scaffolds for Robust in vivo Alveolar Jaw Bone Regeneration. Front Bioeng Biotechnol 2020, 8, 796. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, S.; Nan, X.; Wei, H.; Shi, J.; Li, A.; Gou, J. Repair of human periodontal bone defects by autologous grafting stem cells derived from inflammatory dental pulp tissues. Stem Cell Res Ther 2016, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cao, Y.; Xie, Y.; Wang, H.; Fan, Z.; Wang, J.; Zhang, C.; Wang, J.; Wu, C.T.; Wang, S. Periodontal regeneration in swine after cell injection and cell sheet transplantation of human dental pulp stem cells following good manufacturing practice. Stem Cell Res Ther 2016, 7, 130. [Google Scholar] [CrossRef]
- Lyu, J.; Hashimoto, Y.; Honda, Y.; Matsumoto, N. Comparison of Osteogenic Potentials of Dental Pulp and Bone Marrow Mesenchymal Stem Cells Using the New Cell Transplantation Platform, CellSaic, in a Rat Congenital Cleft-Jaw Model. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, E.E.A.; Beherei, H.H.; El-Zawahry, M.; Farrag, A.R.H.; Kholoussi, N.; Helwa, I.; Mabrouk, M.; Abdel Aleem, A.K. Osteogenic enhancement of modular ceramic nanocomposites impregnated with human dental pulp stem cells: an approach for bone repair and regenerative medicine. J Genet Eng Biotechnol 2022, 20, 123. [Google Scholar] [CrossRef] [PubMed]
- Barbier, L.; Ramos, E.; Mendiola, J.; Rodriguez, O.; Santamaria, G.; Santamaria, J.; Arteagoitia, I. Autologous dental pulp mesenchymal stem cells for inferior third molar post-extraction socket healing: A split-mouth randomised clinical trial. Med Oral Patol Oral Cir Bucal 2018, 23, e469–e477. [Google Scholar] [CrossRef]
- Song, F.; Sun, H.; Huang, L.; Fu, D.; Huang, C. The Role of Pannexin3-Modified Human Dental Pulp-Derived Mesenchymal Stromal Cells in Repairing Rat Cranial Critical-Sized Bone Defects. Cell Physiol Biochem 2017, 44, 2174–2188. [Google Scholar] [CrossRef]
- Song, D.; Xu, P.; Liu, S.; Wu, S. Dental pulp stem cells expressing SIRT1 improve new bone formation during distraction osteogenesis. Am J Transl Res 2019, 11, 832–843. [Google Scholar]
- Wang, W.; Yuan, C.; Geng, T.; Liu, Y.; Zhu, S.; Zhang, C.; Liu, Z.; Wang, P. EphrinB2 overexpression enhances osteogenic differentiation of dental pulp stem cells partially through ephrinB2-mediated reverse signaling. Stem Cell Res Ther 2020, 11, 40. [Google Scholar] [CrossRef]
- Li, J.; Du, H.; Ji, X.; Chen, Y.; Li, Y.; Heng, B.C.; Xu, J. ETV2 promotes osteogenic differentiation of human dental pulp stem cells through the ERK/MAPK and PI3K-Akt signaling pathways. Stem Cell Res Ther 2022, 13, 495. [Google Scholar] [CrossRef]
- Yamakawa, D.; Kawase-Koga, Y.; Fujii, Y.; Kanno, Y.; Sato, M.; Ohba, S.; Kitaura, Y.; Kashiwagi, M.; Chikazu, D. Effects of Helioxanthin Derivative-Treated Human Dental Pulp Stem Cells on Fracture Healing. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Huo, J.F.; Zhang, M.L.; Wang, X.X.; Zou, D.H. Chrysin induces osteogenic differentiation of human dental pulp stem cells. Exp Cell Res 2021, 400, 112466. [Google Scholar] [CrossRef]
- Chan, Y.H.; Ho, K.N.; Lee, Y.C.; Chou, M.J.; Lew, W.Z.; Huang, H.M.; Lai, P.C.; Feng, S.W. Melatonin enhances osteogenic differentiation of dental pulp mesenchymal stem cells by regulating MAPK pathways and promotes the efficiency of bone regeneration in calvarial bone defects. Stem Cell Res Ther 2022, 13, 73. [Google Scholar] [CrossRef] [PubMed]
- Maillard, S.; Sicard, L.; Andrique, C.; Torrens, C.; Lesieur, J.; Baroukh, B.; Coradin, T.; Poliard, A.; Slimani, L.; Chaussain, C. Combining sclerostin neutralization with tissue engineering: An improved strategy for craniofacial bone repair. Acta Biomater 2022, 140, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Lei, F.; Li, M.; Lin, T.; Zhou, H.; Wang, F.; Su, X. Treatment of inflammatory bone loss in periodontitis by stem cell-derived exosomes. Acta Biomater 2022, 141, 333–343. [Google Scholar] [CrossRef]
- Zhao, Y.; Gong, Y.; Liu, X.; He, J.; Zheng, B.; Liu, Y. The Experimental Study of Periodontal Ligament Stem Cells Derived Exosomes with Hydrogel Accelerating Bone Regeneration on Alveolar Bone Defect. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Perrin, S.; Colnot, C. Periosteal Skeletal Stem and Progenitor Cells in Bone Regeneration. Curr Osteoporos Rep 2022, 20, 334–343. [Google Scholar] [CrossRef]
- van Gastel, N.; Stegen, S.; Stockmans, I.; Moermans, K.; Schrooten, J.; Graf, D.; Luyten, F.P.; Carmeliet, G. Expansion of murine periosteal progenitor cells with fibroblast growth factor 2 reveals an intrinsic endochondral ossification program mediated by bone morphogenetic protein 2. Stem Cells 2014, 32, 2407–2418. [Google Scholar] [CrossRef]
- Ji, W.; Kerckhofs, G.; Geeroms, C.; Marechal, M.; Geris, L.; Luyten, F.P. Deciphering the combined effect of bone morphogenetic protein 6 and calcium phosphate on bone formation capacity of periosteum derived cells-based tissue engineering constructs. Acta Biomater 2018, 80, 97–107. [Google Scholar] [CrossRef]
- Lammens, J.; Marechal, M.; Delport, H.; Geris, L.; Oppermann, H.; Vukicevic, S.; Luyten, F.P. A cell-based combination product for the repair of large bone defects. Bone 2020, 138, 115511. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhao, Z.; Cheng, M.; Li, M.; Si, J.; Lin, K.; Yu, H. HIF-1alpha Regulates Osteogenesis of Periosteum-Derived Stem Cells Under Hypoxia Conditions via Modulating POSTN Expression. Front Cell Dev Biol 2022, 10, 836285. [Google Scholar] [CrossRef] [PubMed]
- Groeneveldt, L.C.; Herpelinck, T.; Marechal, M.; Politis, C.; van, I.W.F.J.; Huylebroeck, D.; Geris, L.; Mulugeta, E.; Luyten, F.P. The Bone-Forming Properties of Periosteum-Derived Cells Differ Between Harvest Sites. Front Cell Dev Biol 2020, 8, 554984. [Google Scholar] [CrossRef]
- Yufei, T.; Bingfeng, W.; Jiayi, L.; Hu, L.; Wenli, L.; Lin, X. Distinct osteogenic effect of different periosteum derived cells via Hippo-YAP cascade signaling. Cell Cycle 2023, 22, 183–199. [Google Scholar] [CrossRef]
- Pranskunas, M.; Simoliunas, E.; Alksne, M.; Martin, V.; Gomes, P.S.; Puisys, A.; Kaupinis, A.; Juodzbalys, G. Assessment of the Bone Healing Process Mediated by Periosteum-Derived Mesenchymal Stem Cells' Secretome and a Xenogenic Bioceramic-An In Vivo Study in the Rabbit Critical Size Calvarial Defect Model. Materials (Basel) 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Feng, K.; Hu, J.; Soker, S.; Atala, A.; Ma, P.X. Osteogenic differentiation of human amniotic fluid-derived stem cells induced by bone morphogenetic protein-7 and enhanced by nanofibrous scaffolds. Biomaterials 2010, 31, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jeong, S.Y.; Ju, Y.M.; Yoo, J.J.; Smith, T.L.; Khang, G.; Lee, S.J.; Atala, A. In vitro osteogenic differentiation of human amniotic fluid-derived stem cells on a poly(lactide-co-glycolide) (PLGA)-bladder submucosa matrix (BSM) composite scaffold for bone tissue engineering. Biomed Mater 2013, 8, 014107. [Google Scholar] [CrossRef]
- Mohammed, E.E.A.; Beherei, H.H.; El-Zawahry, M.; Farrag, A.R.H.; Kholoussi, N.; Helwa, I.; Gaber, K.; Allam, M.A.; Mabrouk, M.; Aleem, A.K.A. Combination of Human Amniotic Fluid Derived-Mesenchymal Stem Cells and Nano-hydroxyapatite Scaffold Enhances Bone Regeneration. Open Access Maced J Med Sci 2019, 7, 2739–2750. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Wang, Y.; Xu, J.; Huang, T.; Luo, X. Construction of biomimetic cell-sheet-engineered periosteum with a double cell sheet to repair calvarial defects of rats. J Orthop Translat 2023, 38, 1–11. [Google Scholar] [CrossRef]
- Wang, M.; Li, H.; Si, J.; Dai, J.; Shi, J.; Wang, X.; Guo, L.; Shen, G. Amniotic fluid-derived stem cells mixed with platelet rich plasma for restoration of rat alveolar bone defect. Acta Biochim Biophys Sin (Shanghai) 2017, 49, 197–207. [Google Scholar] [CrossRef]
- Ghaffarinovin, Z.; Soltaninia, O.; Mortazavi, Y.; Esmaeilzadeh, A.; Nadri, S. Repair of rat cranial bone defect by using amniotic fluid-derived mesenchymal stem cells in polycaprolactone fibrous scaffolds and platelet-rich plasma. Bioimpacts 2021, 11, 209–217. [Google Scholar] [CrossRef]
- Wan, C.; He, Q.; Li, G. Allogenic peripheral blood derived mesenchymal stem cells (MSCs) enhance bone regeneration in rabbit ulna critical-sized bone defect model. J Orthop Res 2006, 24, 610–618. [Google Scholar] [CrossRef]
- Zheng, R.C.; Park, Y.K.; Kim, S.K.; Cho, J.; Heo, S.J.; Koak, J.Y.; Lee, S.J.; Park, J.M.; Lee, J.H.; Kim, J.H. Bone Regeneration of Blood-derived Stem Cells within Dental Implants. J Dent Res 2015, 94, 1318–1325. [Google Scholar] [CrossRef]
- Chen, L.; Wu, J.; Wu, C.; Xing, F.; Li, L.; He, Z.; Peng, K.; Xiang, Z. Three-Dimensional Co-Culture of Peripheral Blood-Derived Mesenchymal Stem Cells and Endothelial Progenitor Cells for Bone Regeneration. J Biomed Nanotechnol 2019, 15, 248–260. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Lai, S.; Cao, Q.; Liu, Y.; Li, J.; Zhu, X.; Fu, W.; Zhang, X. Construction of Vascularized Tissue Engineered Bone with nHA-Coated BCP Bioceramics Loaded with Peripheral Blood-Derived MSC and EPC to Repair Large Segmental Femoral Bone Defect. ACS Appl Mater Interfaces 2023, 15, 249–264. [Google Scholar] [CrossRef]
- Li, S.; Huang, K.J.; Wu, J.C.; Hu, M.S.; Sanyal, M.; Hu, M.; Longaker, M.T.; Lorenz, H.P. Peripheral blood-derived mesenchymal stem cells: candidate cells responsible for healing critical-sized calvarial bone defects. Stem Cells Transl Med 2015, 4, 359–368. [Google Scholar] [CrossRef]
- Lin, W.; Xu, L.; Lin, S.; Shi, L.; Wang, B.; Pan, Q.; Lee, W.Y.W.; Li, G. Characterisation of multipotent stem cells from human peripheral blood using an improved protocol. J Orthop Translat 2019, 19, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Diao, Y.; Ma, Q.; Cui, F.; Zhong, Y. Human umbilical cord mesenchymal stem cells: osteogenesis in vivo as seed cells for bone tissue engineering. J Biomed Mater Res A 2009, 91, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Qu, Z.; Yin, X.; Shang, C.; Ao, Q.; Gu, Y.; Liu, Y. Efficacy of umbilical cord-derived mesenchymal stem cell-based therapy for osteonecrosis of the femoral head: A three-year follow-up study. Mol Med Rep 2016, 14, 4209–4215. [Google Scholar] [CrossRef] [PubMed]
- Kosinski, M.; Figiel-Dabrowska, A.; Lech, W.; Wieprzowski, L.; Strzalkowski, R.; Strzemecki, D.; Cheda, L.; Lenart, J.; Domanska-Janik, K.; Sarnowska, A. Bone Defect Repair Using a Bone Substitute Supported by Mesenchymal Stem Cells Derived from the Umbilical Cord. Stem Cells Int 2020, 2020, 1321283. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Yang, H.; Cao, Y.; Yu, D.; Zhao, Y.; Cao, Y. MicroRNA-196a-5p overexpression in Wharton's jelly umbilical cord stem cells promotes their osteogenic differentiation and new bone formation in bone defects in the rat calvarium. Cell Tissue Res 2022, 390, 245–260. [Google Scholar] [CrossRef]
- Yang, S.; Zhu, B.; Yin, P.; Zhao, L.; Wang, Y.; Fu, Z.; Dang, R.; Xu, J.; Zhang, J.; Wen, N. Integration of Human Umbilical Cord Mesenchymal Stem Cells-Derived Exosomes with Hydroxyapatite-Embedded Hyaluronic Acid-Alginate Hydrogel for Bone Regeneration. ACS Biomater Sci Eng 2020, 6, 1590–1602. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Ni, C.Y.; Chen, C.Y.; Rao, S.S.; Yin, H.; Huang, J.; Tan, Y.J.; Wang, Z.X.; Cao, J. , et al. Human umbilical cord mesenchymal stromal cells-derived extracellular vesicles exert potent bone protective effects by CLEC11A-mediated regulation of bone metabolism. Theranostics 2020, 10, 2293–2308. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, Y.; Hao, Z.; Zhou, P.; Wang, P.; Fang, S.; Li, L.; Xu, S.; Xia, Y. Umbilical Mesenchymal Stem Cell-Derived Exosome-Encapsulated Hydrogels Accelerate Bone Repair by Enhancing Angiogenesis. ACS Appl Mater Interfaces 2021, 13, 18472–18487. [Google Scholar] [CrossRef]
- Bahar, D.; Gonen, Z.B.; Gumusderelioglu, M.; Onger, M.E.; Tokak, E.K.; Ozturk-Kup, F.; Ozkan, B.B.; Gokdemir, N.S.; Cetin, M. Repair of Rat Calvarial Bone Defect by Using Exosomes of Umbilical Cord-Derived Mesenchymal Stromal Cells Embedded in Chitosan/Hydroxyapatite Scaffolds. Int J Oral Maxillofac Implants 2022, 37, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Zhang, J.; Li, H.; Zhu, Z.; Guo, S.; Niu, X.; Wang, Y.; Zhang, C. Human Urine Derived Stem Cells in Combination with beta-TCP Can Be Applied for Bone Regeneration. PLoS One 2015, 10, e0125253. [Google Scholar] [CrossRef]
- Guan, J.; Zhang, J.; Zhu, Z.; Niu, X.; Guo, S.; Wang, Y.; Zhang, C. Bone morphogenetic protein 2 gene transduction enhances the osteogenic potential of human urine-derived stem cells. Stem Cell Res Ther 2015, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Zhang, J.; Guo, S.; Zhu, H.; Zhu, Z.; Li, H.; Wang, Y.; Zhang, C.; Chang, J. Human urine-derived stem cells can be induced into osteogenic lineage by silicate bioceramics via activation of the Wnt/beta-catenin signaling pathway. Biomaterials 2015, 55, 1–11. [Google Scholar] [CrossRef]
- Xing, F.; Li, L.; Sun, J.; Liu, G.; Duan, X.; Chen, J.; Liu, M.; Long, Y.; Xiang, Z. Surface mineralized biphasic calcium phosphate ceramics loaded with urine-derived stem cells are effective in bone regeneration. J Orthop Surg Res 2019, 14, 419. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, L.; Xing, F.; Yang, Y.; Gong, M.; Liu, G.; Wu, S.; Luo, R.; Duan, X.; Liu, M. , et al. Graphene oxide-modified silk fibroin/nanohydroxyapatite scaffold loaded with urine-derived stem cells for immunomodulation and bone regeneration. Stem Cell Res Ther 2021, 12, 591. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Gong, M.; Xing, F.; Wu, S.; Xiang, Z. Urine-derived stem cells loaded onto a chitosan-optimized biphasic calcium-phosphate scaffold for repairing large segmental bone defects in rabbits. J Biomed Mater Res B Appl Biomater 2021, 109, 2014–2029. [Google Scholar] [CrossRef]
- Wu, S.; Chen, Z.; Yu, X.; Duan, X.; Chen, J.; Liu, G.; Gong, M.; Xing, F.; Sun, J.; Huang, S. , et al. A sustained release of BMP2 in urine-derived stem cells enhances the osteogenic differentiation and the potential of bone regeneration. Regen Biomater 2022, 9, rbac015. [Google Scholar] [CrossRef]
- Xing, F.; Yin, H.M.; Zhe, M.; Xie, J.C.; Duan, X.; Xu, J.Z.; Xiang, Z.; Li, Z.M. Nanotopographical 3D-Printed Poly(epsilon-caprolactone) Scaffolds Enhance Proliferation and Osteogenic Differentiation of Urine-Derived Stem Cells for Bone Regeneration. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, J.L.; Xing, F.; Duan, X. Three-dimensional printed polylactic acid and hydroxyapatite composite scaffold with urine-derived stem cells as a treatment for bone defects. J Mater Sci Mater Med 2022, 33, 71. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fan, X.L.; Wang, Y.N.; Lu, W.; Wang, H.; Liao, R.; Zeng, M.; Yang, J.X.; Hu, Y.; Xie, J. Extracellular Vesicles from Human Urine-Derived Stem Cells Ameliorate Particulate Polyethylene-Induced Osteolysis. Int J Nanomedicine 2021, 16, 7479–7494. [Google Scholar] [CrossRef]
- Lu, W.; Zeng, M.; Liu, W.; Ma, T.; Fan, X.; Li, H.; Wang, Y.; Wang, H.; Hu, Y.; Xie, J. Human urine-derived stem cell exosomes delivered via injectable GelMA templated hydrogel accelerate bone regeneration. Mater Today Bio 2023, 19, 100569. [Google Scholar] [CrossRef]
- Hayashi, O.; Katsube, Y.; Hirose, M.; Ohgushi, H.; Ito, H. Comparison of osteogenic ability of rat mesenchymal stem cells from bone marrow, periosteum, and adipose tissue. Calcif Tissue Int 2008, 82, 238–247. [Google Scholar] [CrossRef]
- Stockmann, P.; Park, J.; von Wilmowsky, C.; Nkenke, E.; Felszeghy, E.; Dehner, J.F.; Schmitt, C.; Tudor, C.; Schlegel, K.A. Guided bone regeneration in pig calvarial bone defects using autologous mesenchymal stem/progenitor cells - a comparison of different tissue sources. J Craniomaxillofac Surg 2012, 40, 310–320. [Google Scholar] [CrossRef]
- Niemeyer, P.; Fechner, K.; Milz, S.; Richter, W.; Suedkamp, N.P.; Mehlhorn, A.T.; Pearce, S.; Kasten, P. Comparison of mesenchymal stem cells from bone marrow and adipose tissue for bone regeneration in a critical size defect of the sheep tibia and the influence of platelet-rich plasma. Biomaterials 2010, 31, 3572–3579. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B. , et al. Tissue source determines the differentiation potentials of mesenchymal stem cells: a comparative study of human mesenchymal stem cells from bone marrow and adipose tissue. Stem Cell Res Ther 2017, 8, 275. [Google Scholar] [CrossRef]
- Mohamed-Ahmed, S.; Yassin, M.A.; Rashad, A.; Espedal, H.; Idris, S.B.; Finne-Wistrand, A.; Mustafa, K.; Vindenes, H.; Fristad, I. Comparison of bone regenerative capacity of donor-matched human adipose-derived and bone marrow mesenchymal stem cells. Cell Tissue Res 2021, 383, 1061–1075. [Google Scholar] [CrossRef]
- Rodrigues, M.T.; Lee, S.J.; Gomes, M.E.; Reis, R.L.; Atala, A.; Yoo, J.J. Amniotic fluid-derived stem cells as a cell source for bone tissue engineering. Tissue Eng Part A 2012, 18, 2518–2527. [Google Scholar] [CrossRef]
- Mohammed, E.E.A.; El-Zawahry, M.; Farrag, A.R.H.; Aziz, N.N.A.; Sharaf-ElDin, W.; Abu-Shahba, N.; Mahmoud, M.; Gaber, K.; Ismail, T.; Mossaad, M.M. , et al. Osteogenic Differentiation Potential of Human Bone Marrow and Amniotic Fluid-Derived Mesenchymal Stem Cells in Vitro & in Vivo. Open Access Maced J Med Sci 2019, 7, 507–515. [Google Scholar] [CrossRef]
- Nakajima, K.; Kunimatsu, R.; Ando, K.; Ando, T.; Hayashi, Y.; Kihara, T.; Hiraki, T.; Tsuka, Y.; Abe, T.; Kaku, M. , et al. Comparison of the bone regeneration ability between stem cells from human exfoliated deciduous teeth, human dental pulp stem cells and human bone marrow mesenchymal stem cells. Biochem Biophys Res Commun 2018, 497, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Chan, Y.H.; Hsieh, S.C.; Lew, W.Z.; Feng, S.W. Comparing the Osteogenic Potentials and Bone Regeneration Capacities of Bone Marrow and Dental Pulp Mesenchymal Stem Cells in a Rabbit Calvarial Bone Defect Model. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Vater, C.; Mannel, C.; Bolte, J.; Tian, X.; Goodman, S.B.; Zwingenberger, S. Effectiveness of Dental Pulp-derived Stem Cells and Bone Marrowderived Mesenchymal Stromal Cells Implanted into a Murine Critical Bone Defect. Curr Stem Cell Res Ther 2022, 17, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.T.; Lee, W.F.; Chen, S.M.; Hao, L.T.; Hung, Y.T.; Lai, P.C.; Feng, S.W. Effect of Different Bone Grafting Materials and Mesenchymal Stem Cells on Bone Regeneration: A Micro-Computed Tomography and Histomorphometric Study in a Rabbit Calvarial Defect Model. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wei, S.M.; Yan, K.X.; Gu, Y.X.; Lai, H.C.; Qiao, S.C. Bovine-Derived Xenografts Immobilized With Cryopreserved Stem Cells From Human Adipose and Dental Pulp Tissues Promote Bone Regeneration: A Radiographic and Histological Study. Front Bioeng Biotechnol 2021, 9, 646690. [Google Scholar] [CrossRef]
- Nyberg, E.; Farris, A.; O'Sullivan, A.; Rodriguez, R.; Grayson, W. Comparison of Stromal Vascular Fraction and Passaged Adipose-Derived Stromal/Stem Cells as Point-of-Care Agents for Bone Regeneration. Tissue Eng Part A 2019, 25, 1459–1469. [Google Scholar] [CrossRef]
- Zhang, Y.; Grosfeld, E.C.; Camargo, W.A.; Tang, H.; Magri, A.M.P.; van den Beucken, J. Efficacy of intraoperatively prepared cell-based constructs for bone regeneration. Stem Cell Res Ther 2018, 9, 283. [Google Scholar] [CrossRef]
- Lough, D.; Swanson, E.; Sopko, N.A.; Madsen, C.; Miller, D.; Wang, H.; Guo, Q.; Sursala, S.M.; Kumar, A.R. Regeneration of Vascularized Corticocancellous Bone and Diploic Space Using Muscle-Derived Stem Cells: A Translational Biologic Alternative for Healing Critical Bone Defects. Plast Reconstr Surg 2017, 139, 893–905. [Google Scholar] [CrossRef]
- Agata, H.; Asahina, I.; Yamazaki, Y.; Uchida, M.; Shinohara, Y.; Honda, M.J.; Kagami, H.; Ueda, M. Effective bone engineering with periosteum-derived cells. J Dent Res 2007, 86, 79–83. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.B.; Lamo-Espinosa, J.M.; Muinos-Lopez, E.; Ripalda-Cemborain, P.; Abizanda, G.; Valdes-Fernandez, J.; Lopez-Martinez, T.; Flandes-Iparraguirre, M.; Andreu, I.; Elizalde, M.R. , et al. Periosteum-derived mesenchymal progenitor cells in engineered implants promote fracture healing in a critical-size defect rat model. J Tissue Eng Regen Med 2019, 13, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Gan, Y.; Shi, D.; Zhao, J.; Tang, T.; Dai, K. A novel cytotherapy device for rapid screening, enriching and combining mesenchymal stem cells into a biomaterial for promoting bone regeneration. Sci Rep 2017, 7, 15463. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hwang, M.P.; Wright, N.; Lu, A.; Ruzbarsky, J.J.; Huard, M.; Cheng, H.; Mullen, M.; Ravuri, S.; Wang, B. , et al. The use of heparin/polycation coacervate sustain release system to compare the bone regenerative potentials of 5 BMPs using a critical sized calvarial bone defect model. Biomaterials 2022, 288, 121708. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



