
Submitted:
08 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
Phytoene desaturase (PDS) is a plant enzyme involved in carotenoid biosynthesis. The PDS gene has been used as a selective marker for genome editing in several plant species, including banana (Musa spp.). Its knockout promotes dwarfism and albinism, characteristics that are easily recognizable and highly favorable. In Musa spp., the A genome increases fruit production and quality, whereas the B genome is associated with tolerance to biotic and abiotic stresses. The objective of this study was to identify a molecular marker in the PDS gene to easily discriminate the A and B genomes of banana. A 2166 bp fragment for the ‘PDSMa’ marker was identified as polymorphic for the A genome (identification accuracy of 99.33%), whereas ~332 and ~225 bp fragments were detected for the ‘PDSMb’ primer with 100% accuracy using MedCalc software. To the best of our knowledge, this is the first study to use the PDS gene to determine doses of the A genome and identify the B genome in Musa spp., which will aid in evaluating the genomic constitution of banana hybrids and accessions at the seedling stage, and accelerating their classification in crop genetic improvement programs.
Keywords:
Gene composition
; phytoene desaturase
; molecular marker
; genetic improvement
; Musa spp.
1. Introduction
Bananas and plantains (Musaceae) are grown in all tropical and subtropical regions worldwide. They are the fourth largest food crop on the global market, after rice, wheat, and maize [1].
Most commercial banana cultivars originated from crosses between the wild subspecies Musa acuminata Colla (2n = 2x = 22; genome A) and Musa balbisiana Colla (2n = 2x = 22; genome B), which produced a series of diploid, triploid, and tetraploid bananas. The genomic groups resulting from these crosses are classified as AA, AB, AAA, AAB, ABB, AABB, AAAB, and ABBB [2]. The genome sequence of M. acuminata ssp. malaccensis, derived from a double haploid Pahang accession, represents the A genome (n = 11) [3,4], whereas that of M. balbisiana, derived from a Pisang Klutuk Wulung accession, represents the B genome (x = 11) [5,6].
The A genome is mostly related to improved production, yield, and fruit quality attributes, whereas the B genome lends robustness and tolerance/resistance to abiotic and biotic stresses [3,5,6]. The “B” genome is associated with the banana streak virus (BSV) [7], which influences the exchange of accessions between germplasm banks, field management, and in vitro cultivation. The virus has two forms of endogenous sequences (eBSV) in the “B” genome [8,9]: i) incomplete sequences that are considered evolutionary relics from previous infections and do not cause the disease, and ii) complete sequences that are initially dormant and activated to promote pathogenesis when the plant is challenged by biotic/abiotic stresses [10,11].
The genomic composition of banana is unpredictable, even in controlled crosses, owing to "unbalanced meiosis" and homologous recombination between “A” and “B” genomes, leading to a different number of sets or segments of each parent genome [12,13,14].
Ploidy is determined in banana using several methods, including morphological markers [15]. However, morphological markers are sensitive to environmental factors and are imprecise and impractical to measure at a large scale [13,16]. The use of molecular markers to distinguish the doses of “A” and “B” genomes in Musa spp. has been evaluated [13,15,17], however, despite their advantages over morphological markers, molecular markers are vulnerable to co-amplification with fungal DNA, if present, leading to misidentification, multiple copies, and ultimately, low accuracy.
Breeding programs seek effective and long-lasting techniques to improve crop characteristics, but are limited by the complex inheritance of most agronomic traits and strong genotype–environment interaction [18]. Recently, the CRISPR/Cas9 system has been widely used to induce specific genome mutations in several plant species, which has greatly contributed to the study of gene function in crop genetic improvement programs. This technique facilitates gene editing by cutting and replacing or adding sequences to the DNA of a given genotype [19]. To validate the use of CRISPR/Cas9 for tolerance to biotic and abiotic stresses in banana, the literature proposes initially using the knockout of the PDS (Phytoene desaturase) gene as a proof of concept [20,21,22].
The PDS gene has been widely used as a molecular marker for genome editing in several plant species, including bananas [20,21]. This gene plays a fundamental role in the carotenoid biosynthesis pathway, as it is highly conserved and has similar catalytic properties. PDS is a key enzyme in the carotenoid biosynthesis pathway, catalyzing the desaturation of phytoene (a transparent compound) into ζ-carotene, which is subsequently converted into lycopene, a colored compound [23]. PDS knockout affects photosynthesis, gibberellin production, and carotenoid biosynthesis, which leads to dwarfism and albinism in plants [24,25,26], suggesting that PDS can be a selective marker for the development of genetic engineering products.
The objective of this study was to develop a marker from the PDS gene capable of differentiating the A genome (M. acuminata) from the B genome (M. balbisiana) in banana. To validate its potential, the gene marker was tested on 150 banana accessions with different ploidy types collected from the Embrapa Mandioca e Fruticultura Germplasm Bank. This is the first report of a PDS gene-derived molecular marker that can identify “A” and “B” genomes in banana with 99.33% and 100% accuracy, respectively. Our study provides a foundation for the preliminary characterization of the genomic composition of banana accessions to predict agronomic, sensory, and resistance/tolerance characteristics that are desirable for genetic improvement programs.
2. Materials and Methods
2.1. Plant material
We evaluated 150 banana accessions associated with different genomic groups and levels of ploidy (AA, AAA, AAAA, AB, BB, AAB, ABB, AAAB, AABB, and ABBB), which were procured from the Banana Germplasm Bank of Embrapa Mandioca e Fruticultura, Cruz das Almas, Bahia, Brazil (12°40′48.03″S and 39°05′20.91″W). The collection consisted of four plants per accession, spaced 2.0 m between rows × 1.7 m between plants, and irrigation by micro-sprinklers was performed according to plant needs. This approach would minimize water stress, even during dry periods.
2.2. Primer construction
To design specific primers for M. acuminata and M. balbisiana, PDS gene sequences from the “A” genome (M. acuminata, gene Ma08_t16510.2) and “B” genome (M. balbisiana, gene Mba08_g16040.1) were initially obtained from the SouthGreen-Banana Genome Hub platform (https://banana-genome-hub.southgreen.fr/). “A” and “B” genome sequences were aligned to identify a conserved region in the “A” genome and polymorphic region in the “B” genome, and vice versa, using Clustal Omega software [27].
After identifying the conserved PDS regions of M. acuminata and M. balbisiana, specific PDS gene primers were constructed using Oligo Explorer software (version 1.2.) (PDSMaF ATTGTGAAAGAGGTCGAGGA, PDSMaR TGCGGTAAAAGAAGCTTCAA; PDSMbF GTGAGTTCATGGGTTGCCAA, PDSMbR ACCGGCTATGACAACCTTCA). The discriminatory power of the primers was assessed by polymerase chain reaction (PCR) analysis.
2.3. DNA extraction and PCR conditions
DNA was extracted from the young leaves of the 150 banana accessions as described by Doyle and Doyle [28], with modifications proposed by Ferreira et al. [29]. The samples of young banana leaves (300 mg) were immediately placed in plastic bags (20 × 10 cm) with 2 mL extraction buffer (2% CTAB; 100 mM Tris-HCl, pH 8.0; 50 mM EDTA, pH 8.0; 1.4 M NaCl; 2% PVP-40; and 1% sodium sulfite). They were then macerated in a drill press to release nuclear DNA until a homogeneous solution was obtained.
The quantity and quality of genomic DNA was assessed on 0.8% agarose gel in 0.5X TBE (90 mM tris, 90 mM boric acid, 2.5 mM EDTA, pH 8.3) and a microvolume spectrophotometer (GE NanoVue Plus, Biochrom, Holliston, MA, USA). The samples were then diluted in TE (1 M tris-HCl 1 M, pH 8.0; and 0.5 M EDTA, pH 8.0), and standardized to 5 ng/μL.
The DNA samples obtained from each genotype were amplified by PCR using the reference genes, β-tubulin, PDS_AB [26], PDSMa, and PDSMb primers (Table 1). Amplification reactions were conducted with 1.5 µL PCR buffer (10×), 1.5 µL MgCl2 (50 mM), 1.2 µL dNTP (2.5 mM), 1.0 µL of the F and R primers (10.0 μM), and 0.2 µL Taq DNA polymerase (5 U/µL), with a final volume of 20 μL.
The samples were then amplified in a Veriti thermal cycler (Applied Biosystems, Waltham, MA, USA) under the following programming conditions: one cycle at 95 °C for 15 min; 34 cycles at 94 °C for 30 s; one cycle at 55 °C for the PDS_AB primer, 60 °C for β-tubulin and PDSMa, and 65 °C for PDSMb for 30 s for primer annealing, and one cycle at 72 °C for 1 min; and a final cycle at 72 °C for 10 min (adapted from Ntui et al. [26]. Primer sequences and annealing temperatures are shown in Table 1.
The amplification products were separated by electrophoresis in 2% and 3% agarose gel (PDSMb) at 70 V in TBE buffer for 4 h and stained with Gel red (1 μg/mL). The amplified products were visualized and photographed under ultraviolet light in an (L-Pix Touch documentation system (Loccus, Cotia, Brazil).
2.4. Data analysis
The data obtained through electrophoresis was recorded as band presence or absence. The PDS gene marker for the “A” and “B” genomes (994 bp; [26]) was used to compare the efficiency of the new primers in identifying the “A” and “B” genomes, and the β-tubulin primer (110 bp) was used as an endogenous control for Musa spp. [30]. The base pairs of the fragments amplified by the PDS gene-derived primers were calculated on the log scale with the following regression models: i) identification of “A” genome, y = –0.003x + 9.599 and ii) identification of “A” and “B” genome, y=–0.0123+13.989. MedCalc software (https://www.medcalc.org/calc/diagnostic_test.php) [31] was used for statistical analysis. This software provides statistical information regarding the sensitivity, specificity, likelihood ratio (negative or positive), and accuracy of the polymorphic fragment detected by the screening of a set of genotypes.
3. Results and Discussion
The complete PDS gene sequences of M. acuminata (AA) and M. balbisiana (BB) were downloaded from the Banana Genome Hub (https://banana-genome-hub.southgreen.fr/) on the SouthGreen platform. The PDS gene (Ma08_g16510) of M. acuminata has 27 944 bp and 14 exons, and the PDS gene (Mba08_g16040.1) of M. balbisiana has 21 262 bp and 11 exons. The alignment of these sequences shared 96.40% nucleotide homology from the start to stop codons.
The PDS gene has often been used as a concept marker/proof in CRISPR/Cas9 gene editing experiments in many plant species such as maize [32], Arabdopsis [33], tomato [34], rice [35], and banana [26].
After downloading the material to construct the PDSMa- and PDSMb-specific markers, the coding regions of the PDS gene were aligned using Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/) to identify discriminatory/polymorphic regions between the “A” and “B”genomes (Figure 1).
The specific primers for PDSMa and PDSMb were evaluated by PCR. Two primers were used as controls—β-tubulin, as an endogenous gene, and the primer developed by Ntui et al. [26] to amplify the PDS gene in both the “A” and “B” genomes (PDS_AB) (Figure 2A). The amplification of β-tubulin, PDS_AB [26], and PDSMa in 12 banana samples with representative genomes of different ploidy types (Germplasm Bank of Embrapa Mandioca e Fruticultura) is shown in Figure 2A.
The cultivars Balbisiana Franca (BB), Butuhan (BB), Musa balbisiana (BB), BB Franca (BB), and Teparod (ABBB) showed band amplification only for the PDS_AB and β-tubulin primers, confirming the discriminatory power of the PDSMa primer (Figure 2B).
The PDSMa primer has 476 bp and was constructed without intron regions. The amplification of this primer in all “A” genome Musa accessions produced a 2166 bp fragment (Figure 2B) based on the regression model y = –0.003x + 9.5999 (R2 = 0.90). This band size reflects our use of total genomic DNA, which contains introns. The PDS_AB and β-tubulin primers produced fragments of 994 and 110 bp, corroborating the PDS gene sizes reported by Ntui et al. [26] and Podevin et al. [30], respectively.
The amplification of the PDSMb primer in six banana samples with different ploidy types is shown in Figure 3. A fragment of approximately 332 bp was observed in all samples with A and B ploidy (Figure 3), which was based on the regression model y = –0.0123 + 13.989 (R2 = 0.96). In addition to the ~322 bp band, the cultivars Zebrina (AA), Gros Michel (AAA), and Bucaneiro (AAAA) presented the amplification of a second specific band of ~225 bp (based on the same regression model), indicating that this band pattern only occurred in specimens with 100% A ploidy (Figure 3).
Table 2 shows the banana accessions from the Embrapa Mandioca e Fruticultura Germplasm Bank and their respective PCR amplification results with the four primers. The 150 genotypes evaluated were represented by different ploidy types: AA, AAA, AAAA, BB, AB, AAB, ABB, AAAB, AABB, and ABBB. Table 2 shows cultivars with bands amplified using the primers β-tubulin, PDS_AB, PDSMa, and PDSMb. All genotypes were positive for the primers β-tubulin and PDS_AB. For the PDSMa primer, amplification did not occur in genotypes with >75% of genome “B”, and all 100% “A” ploidy genotypes had amplified bands of ~225 bp in PDSMb.
Banana genetic improvement is based on crossing wild or improved diploids with commercial cultivars to generate hybrids resistant/tolerant to biotic and abiotic stresses and with agronomic characteristics consistent with market demands [36,37]. Nwakanma et al. [15], suggested that early determination of the banana genome composition can aid breeders in predicting the occurrence of useful agronomic characteristics and developing new varieties.
The development of molecular markers capable of discriminating high doses of the “B” genome in bananas is essential for determining gene composition and inferring important characteristics in hybrids [22]. The PDSMb primer developed in this study proved to be useful for detecting the ploidy of cultivars developed in the Embrapa breeding program. This marker effectively identified the “B” genome in the gene composition of the different accessions; even if the sample has only 25% of the B genome in its ploidy, the primer will not detect and not reveal the second ~225 bp band (Figure 3, Table 2).
MedCalc software (https://www.medcalc.org/calc/diagnostic_test.php) was used in the molecular analysis of the PDSMa and PDSMb markers. This software is used in the health sector for disease diagnosis and can be adapted for use in plants. In the program you need to fill in information about true positives, false negatives, false positives and true negatives. This way, it is possible to extract statistics on sensitivity, specificity, likelihood ratio (negative or positive) and accuracy of the polymorphic fragment found.
Of the 150 accessions subjected to PCR, 145 showed bands at 2166 bp for the PDSMa primer, indicating the presence of the “A” genome, and 92 showed bands at ~225 bp for the PDSMb primer. These samples were classified as true positive in the MedCalc analysis.
Only one sample, the Teparod genotype (ABBB), was identified as a false negative for the PDSMa primer, because the “A” genome in its composition was not identified by band amplification in this region. This result corroborates the occurrence of homologous recombination between “A” and “B” genome cultivars, suggesting that, in this specific case, the Teparod genotype may not be carrying the full complement of the “A” genome [13,38,39]. There were no false negatives for PDSMb, as band amplification (~225 bp) occurred in all cultivars with 100% “A” ploidy.
None of the samples with the “B” genome showed the 2166 bp fragment in PDSMa, nor the second band of ~225 bp in PDSMb, representing a false positive. Four samples containing the “B” genome showed no amplification with PDSMa, and 58 with “B” ploidy genotypes showed no amplification for the second band with PDSMb, making them true negatives. We calculated the static parameters of the PDSMa and PDSMb markers, which showed 99.32% and 100% sensitivity, 100% specificity, 100% positive predictive value, 80% and 100% negative predictive value, and 99.33% and 100% accuracy, respectively, indicating that the PDSMa marker is highly effective in discriminating “B” genome doses >75% in banana genotypes and that the PDSMb marker can identify accessions with 100% of the “A” genome in their ploidy (Table 3).
The use of molecular markers to determine the genomic composition of Musa cultivars and other crops have many advantages over morphological markers [40]. Several studies have used molecular methods to identify the genomes of M. acuminata and M. balbisiana. Nwakanma et al. [15] and Jesus et al. [13] identified molecular markers based on internal transcribed spacers (ITS), which discriminated “A” from “B” genomes in bananas, but not very accurately. Hollingsworth [41] showed that markers based on ITS regions were vulnerable to co-amplification with fungal DNA, leading to misidentification and multiple, possibly divergent, ITS copies in a single specimen.
Mabonga and Pillay [42] developed a 500 bp SCAR marker based on a RAPD marker to identify the “A” genome in bananas and plantains. Although the marker was useful for identifying the “A” genome, a 700 bp fragment hybridized with all the genotypes and impeded the differentiation of “A” and “B” genomes. Many primers have been obtained by converting RAPD markers into SCAR markers. However, this conversion generally leads to a decreased level of polymorphism [43], particularly with different genetic backgrounds.
The identification of genotypes with B genome doses based on the absence of a band is also valuable for predicting BSV disease onset, which is mainly caused by three virus species: Goldfinger (eBSGFV), Imovè (eBSIMV), and Obinol’Ewai (eBSOLV). Because of viral introgression in the B genome (eBSV), caution must be exercised in the cultivation of “B”genome accessions since disease onset can be stimulated by the several in vitro subcultures required by the crop, which is vegetatively propagated, and external plant stresses, such as low temperatures [9,44]. Thus, early identification of the “B” genome could be instrumental in enhancing crop management practices in the agricultural field.
The highly accurate PDS gene markers developed in this study to discriminate the “A” and “B” genomes in bananas represents a useful new tool for the genetic improvement of Musaceae crops, particularly due to its origin from a highly conserved gene with few copies. These markers can potentially be used in the molecular characterization of germplasm collections and new accessions to expand the genetic diversity of the crop, which would be useful in discriminating between controlled and uncontrolled crosses and providing information for seedling exchange. These activities constitute the basis of genetic improvement programs for bananas.
5. Conclusions
The PDSMa and PDSMb markers derived from the PDS gene enable early prediction of genotypes with “A” and “B” genomes in banana hybrids. These markers represent useful tools for genetic improvement strategies, aimed at developing varieties with higher fruit quality, yield, and disease resistance, and could potentially predict BSV disease onset triggered by environmental conditions and biotic stresses. For CRISPR/Cas9-based genetic editing of banana crops in Brazil, the sequences of the PDSMa and PDSMa markers will be used in future studies as guide RNA for Cas9 in the CRISPR knockout of the PDS gene, which is involved in the biosynthesis of carotenoids and is associated with albinism. Subsequent studies will focus on the identification and manipulation of genes for resistance/tolerance to biotic and abiotic stresses.
Author Contributions
Conceptualization, F.d.S.N. and M.S.M.; methodology, F.d.S.N., M.S.M., A.d.J.R., J.M.d.S.S., C.F.F., T.A.d.O.M. and E.P.A.; software, F.d.S.N., M.S.M. and A.d.J.R.; validation, E.P.A. and C.F.F.; formal analysis, F.d.S.N. M.S.M. and E.P.A.; investigation, F.d.S.N., A.d.J.R., J.M.d.S.S., A.P.d.S.R., M.S.M., S.C.B. and C.C.H.d.S.; resources, E.P.A. and C.F.F.; data curation, F.d.S.N.; writing—original draft preparation, F.d.S.N., E.P.A. and C.F.F.; writing—review and editing, E.P.A., C.F.F. and L.E.C.D.; visualization, E.P.A. and C.F.F.; supervision, E.P.A., C.F.F., J.A.d.S.-S., T.A.d.O.M. and L.E.C.D.; project administration, E.P.A.; funding acquisition, E.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by IITA/The Bill and Melinda Gates Foundation—Accelerated Breeding of Better Bananas, ID OPP1093845.
Data Availability Statement
All data generated or analyzed during this study are included in this article.
Acknowledgments
The authors thank the Graduate Program in Biotechnology (PPGBiotec) of the State University of Feira de Santana, as well as CNPq (National Council for Scientific and Technological Development) for the research productivity grants for E.P.A. and C.F.F.; CAPES (Coordination for the Improvement of Higher Education Personnel) for granting DSc. scholarships to F.d.S.N. and M.S.M.; and Fapesb (Bahia Research Foundation) for granting DSc. scholarships to J.M.d.S.S., A.d.J.R. and M.d.S.F.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 10 August 2023).
- Simmonds, N.W.; Shepherd, K. The taxonomy and origins of the cultivated bananas. The journal of the Linean Society of London, London, v. 55, 1955, p. 302-12.
- D’hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Baurens, F.C.; Droc, G.; Rouard, M.; Cenci, A.; Kilian, A.; Hastie, A.; Doležel, J.; Aury, J.M.; Alberti, A.; et al. Improvement of the banana “Musa acuminata” reference sequence using NGS data and semi-automated bioinformatics methods. BMC Genomics 2016, 17, 243. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Gudimella, R.; Harikrishna, J.A.; Sin, L.W.; Khalid, N.; Keulemans, J. A draft Musa balbisiana genome sequence for molecular genetics in polyploid, inter-and intra-specific Musa hybrids. BMC Genom. 2013, 14, 683. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Miao, H.; Liu, J.; Xu, B.; Yao, X.; Xu, C.; Zhang, J. Musa balbisiana genome reveals subgenome evolution and functional divergence. Nat. Plants 2019, 5, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D.W.; Mcmichael, L.A.; Dietzgen, R.G.; Thomas, J.E. Genetic diversity among Banana streak virus isolates from Australia. Phytopathol 2000, 90, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Gayral, P.; Noa-Carrazana, J.C.; Lescot, M.; Lheureux, F.; Lockhart, B.E.L.; Matsumoto, T.; Piffanelli, P.; Iskra-Caruana, M.L. A Single Banana Streak Virus Integration Event in the Banana Genome as the Origin of Infectious Endogenous Pararetrovirus. J Virol 2008, 82, 6697–6710. [Google Scholar] [CrossRef] [PubMed]
- Chabannes, M.; Baurens, F.C.; Duroy, P.O.; Bocs, S.; Vernerey, M.S.; Goud, M.R.; Barbe, V.; Gayral, P.; Iskra-Caruana, M.L. Three infectious viral species lying in wait in the Banana Genome. J. Virol 2013, 87, 862–863. [Google Scholar] [CrossRef] [PubMed]
- Dallot, S.; Acuna, P.; Rivera, C.; Ramírez, P.; Cōte, F.; Lockhart, B.E.; Caruana, M.L. Evidence that the proliferation stage of micropropagation procedure is determinant in the expression of Banana streak virus integrated into the genome of the FHIA 21 hybrid (Musa AAAB). Archives of Virology 2001, 146, 2179–2190. [Google Scholar] [CrossRef] [PubMed]
- Côte, F.X.; Galzi, S.; Folliot, M.; Lamagnère, Y.; Teycheney, P.Y.; Iskra-Caruana, M.L. Micropropagation by tissue culture triggers differential expression of infectious endogenous Banana streak virus sequences (eBSV) present in the B genome of natural and synthetic interspecific banana plantains. Mol. Plant Pathol. 2010, 11, 137–144. [Google Scholar] [CrossRef]
- Perrier, X.; Bakry, F.; Carreel, F.; Jenny, C.; Horry, J.P.; Lebot, V.; Hippolyte, I. Combining biological approaches to shed light on evolution of edible bananas. Ethnobot Res Appl 2009, 7, 199–216. [Google Scholar] [CrossRef]
- Jesus, O.N.; Amorim, E.P.; Ferreira, C.F.; Campos, J.M.S.; Silva, G.D.G.; Figueira, A. Genetic diversity and population structure of Musa accessions in ex situ conservation. BMC plant biology 2013, 13, 41. [Google Scholar] [CrossRef]
- Santos-Serejo, J.A.; Amorim, E.P.; Jesus, O.N.; Silva, S.O. Germoplasma de Musa, conservação, caracterização e uso. In O agronegócio da banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, DF, 2016; pp. 113–136. [Google Scholar]
- Nwakanma, D.C.; Pillay, M.; Okoli, B.E.; Tenkouano, A. PCR-RFLP of the ribosomal DNA internal transcribed spacers (ITS) provides markers for the A and B genomes in Musa L. Theoretical and Applied Genetics 2003, 108, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Perrier, X.; Du Montcel, H.T. Musaid: a computerized determination system. In Proceedings of Identification of Genetic Diversity in the Genus Musa; Los Banos (PHL); Jarret, R., Ed.; INIBAP: Montpellier, França, 1990; pp. 76–91. [Google Scholar]
- Pillay, M.; Nwakanma, D.C.; Tenkouano, A. Identification de RAPD markers linked to A and B genome sequences in Musa L. Genome 2000, 43, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, G.P.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet 2016, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Molinari, H.B.C.; Vieira, L.R.; Silva, N.; Prado, G.S.; Lopes Filho, J.H. Tecnologia CRISPR na genômica de plantas: biotecnologia aplicada à agricultura; Embrapa: Brasília, Brazil, 2020; p. 207. [Google Scholar]
- Naim, F.; Dugdale, B.; Kleidon, J.; Brinin, A.; Shand, K.; Waterhouse, P.; Dale, J. Gene editing the phytoene desaturase alleles of Cavendish banana using CRISPR/Cas9. Transgenic research 2018, 27, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Alok, A.; Kaur, N.; Pandey, P.; Awasthi, P.; Tiwari, S. CRISPR/Cas9-mediated efficient editing in phytoene desaturase (PDS) demonstrates precise manipulation in banana cv. Rasthali genome. Funct. Integr. Genomics 2018, 18, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. Application of genetic modification and genome editing for developing climate-smart banana. Food Energy Secur. 2019, 8, e00168. [Google Scholar] [CrossRef]
- Bai, C.; Capell, T.; Berman, J.; Medina, V.; Sandmann, G.; Christou, P.; Zhu, C. Bottlenecks in carotenoid biosynthesis and accumulation in rice endosperm are influenced by the precursor product balance. Plant Biotechnol. J 2016, 195–205. [Google Scholar] [CrossRef]
- Wang, M.; Wang, G.; Ji, J.; Wang, J. The effect of pds gene silencing on chloroplast pigment composition, thylakoid membrane structure and photosynthesis efficiency in tobacco plants. Plant Science 2009, 177, 222–226. [Google Scholar] [CrossRef]
- Koschmieder, J.; Fehling-Kaschek, M.; Schaub, P.; Ghisla, S.; Brausemann, A.; Timmer, J.; Beyer, P. Plant-type phytoene desaturase: Functional evaluation of structural implications. PloS one 2017, 12, e0187628. [Google Scholar] [CrossRef]
- Ntui, V.O.; Tripathi, J.N.; Tripathi, L. Robust CRISPR/Cas9 mediated genome editing tool for banana and plantain (Musa spp.). Current Plant Biology 2020, 21, 100128. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Ferreira, C.F.; Gutierrez, D.; Kreuze, J.; Iskra Caruana, M.L.; Chabannes, M.; Barbosa, A.C.O.; Santos, T.A.; Silva, A.G.S.; Santos, R.M.F.; Amorim, E.P.; et al. Rapid plant DNA and RNA extraction protocol using a bench drill. GMR 2019, 18, gmr18394. [Google Scholar] [CrossRef]
- Podevin, N.; Krauss, A.; Henry, I.; Swennen, R.; Remy, S. Selection and validation of reference genes for quantitative RT-PCR expression studies of the non-model crop Musa. Molecular Breeding 2012, 30, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- MedCalc Software Ltd. Diagnostic test evaluation calculator. Version 22.009. Available online: https://www.medcalc.org/calc/diagnostic_test.php (accessed on 4 August 2023).
- Matthews, P.D.; Luo, R.; Wurtzel, E.T. Maize phytoene desaturase and ζ-carotene desaturase catalyse a poly-Z desaturation pathway: implications for genetic engineering of carotenoid content among cereal crops. J. Exp. Bot. 2003, 54, 2215–2230. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Gu, H.; Ma, L.; Peng, Y.; Deng, X.W.; Chen, Z.; Qu, L.J. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Res 2007, 17, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kyu, S.Y.; Pe, P.P.W.; Park, K.I.; Lee, J.M.; Lim, K.B.; Kim, C.K. Silencing of the phytoene desaturase (PDS) gene affects the expression of fruit-ripening genes in tomatoes. Plant methods 2019, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Banakar, R.; Schubert, M.; Collingwood, M.; Vakulskas, C.; Eggenberger, A.L.; Wang, K. Comparison of CRISPR-Cas9/Cas12a ribonucleoprotein complexes for genome editing efficiency in the rice phytoene desaturase (OsPDS) gene. Rice 2020, 13, 1–7. [Google Scholar] [CrossRef]
- Silva, S.O.; Santos-Serejo, J.A.; Amorim, E.P. Pré-melhoramento da banana. In Pré-melhoramento de plantas: estado da arte e experiências de sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informação Tecnológica: Brasília, DF, 2011; pp. 317–350. [Google Scholar]
- Amorim, E.P.; Santos-Serejo, J.A.; Amorim, V.B.O.; Silva, S.O. Melhoramento genético. In O agronegócio da banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, DF, 2016; pp. 171–200. [Google Scholar]
- Roux, N.; Dolezel, J.; Swennen, R.; Zapata-Arias, F.J. Effectiveness of three micropropagation techniques to dissociate cytochimeras in Musa spp. Plant Cell, Tissue and Organ Culture 2001, 66, 189–197. [Google Scholar] [CrossRef]
- De Langhe E, Hřibová E, Carpentier S, Doležel J, Swennen R: Didbackcrossing contribute to the origin of hybrid edible bananas? Ann Bot. 2010, 106, 849–857. [CrossRef] [PubMed]
- Dhivya, S.; Ashutosh, S.; Gowtham, I.; Baskar, V.; Harini, A.B.; Mukunthakumar, S.; Sathishkumar, R. Molecular identification and evolutionary relationships between the subspecies of Musa by DNA barcodes. BMC genomics 2020, 21, 659. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M. Refining the DNA barcode for land plants. Proc Natl Acad Sci USA. 2011, 108, 19451–19452. [Google Scholar] [CrossRef] [PubMed]
- Mabonga, L.; Pillay, M. SCAR Marker for the A Genome of Bananas (Musa spp. L.) Supports Lack of Differentiation between the A and B Genomes. Journal of Agricultural Science 2017, 9, 64. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoglu, R.; Ahmadg, F.; Alsalehh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: current status and recent advancements in genomic selection and genome editing. Biotechnology & Biotechnological Equipment 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Iskra-Caruana, M.L.; Baurens, F.C.; Gayral, P.; Chabannes, M. A four-partner plant–virus interaction: enemies can also come from within. Mol Plant Microbe Interact 2010, 23, 1394–1402. [Google Scholar] [CrossRef]
Figure 1.
Partial alignment of the PDS gene of Musa acuminata (AA) and M. balbisiana (BB) for the construction of specific M. acuminata (PDSMa) and M. balbisiana (PDSMb) primers. Blue and gray markings correspond to the forward and reverse primers of PDSMa, respectively; green and yellow markings to the forward and reverse primers of PDSMb, respectively. Red marking represents the polymorphic region of the F and R primers between the “A” and “B” genomes.
Figure 1.
Partial alignment of the PDS gene of Musa acuminata (AA) and M. balbisiana (BB) for the construction of specific M. acuminata (PDSMa) and M. balbisiana (PDSMb) primers. Blue and gray markings correspond to the forward and reverse primers of PDSMa, respectively; green and yellow markings to the forward and reverse primers of PDSMb, respectively. Red marking represents the polymorphic region of the F and R primers between the “A” and “B” genomes.
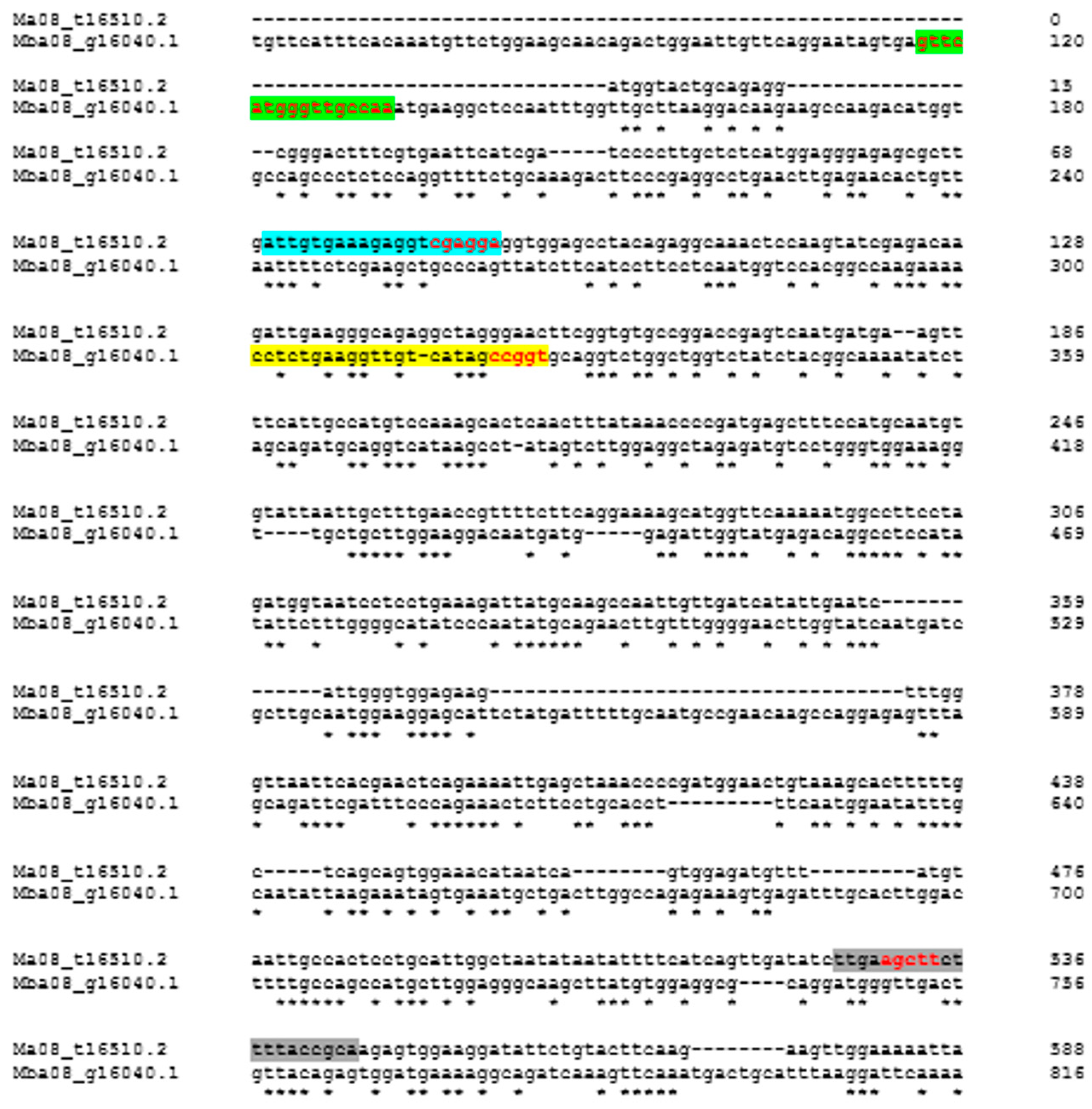
Figure 2.
PCR amplification with. (A) PDS_AB (994 bp) and β-tubulin (110 bp) primers; (B) PDSMa (2166 bp) in accessions 1–12 from the Embrapa Mandioca e Fruticultura Germplasm Bank; (C) Control; 1) Gros Michel (AAA); 2) Prata Anã (AAB); 3) Balbisiana Franca (BB); 4) Zebrina (AA); 5) IAC 1 (AB); 6) Bucaneiro (AAAA); 7) Butuhan (BB); 8) Musa balbisiana (BB); 9) BB Franca (BB); 10) Pelipita (ITC 0472) (ABB); 11) FC06-02 (AABB); 12) Teparod (ABBB), listed in Table 1. M = 1 kb marker (Invitrogen). Arrows of 110, 994, and 2166 bp correspond to the β-tubulin, PDS_AB, and PDSMa primers, respectively. The 500-bp fragment in genotype 4 for the PDSMa primer (B) corresponds to the non-specific band that only appeared in this case.
Figure 2.
PCR amplification with. (A) PDS_AB (994 bp) and β-tubulin (110 bp) primers; (B) PDSMa (2166 bp) in accessions 1–12 from the Embrapa Mandioca e Fruticultura Germplasm Bank; (C) Control; 1) Gros Michel (AAA); 2) Prata Anã (AAB); 3) Balbisiana Franca (BB); 4) Zebrina (AA); 5) IAC 1 (AB); 6) Bucaneiro (AAAA); 7) Butuhan (BB); 8) Musa balbisiana (BB); 9) BB Franca (BB); 10) Pelipita (ITC 0472) (ABB); 11) FC06-02 (AABB); 12) Teparod (ABBB), listed in Table 1. M = 1 kb marker (Invitrogen). Arrows of 110, 994, and 2166 bp correspond to the β-tubulin, PDS_AB, and PDSMa primers, respectively. The 500-bp fragment in genotype 4 for the PDSMa primer (B) corresponds to the non-specific band that only appeared in this case.
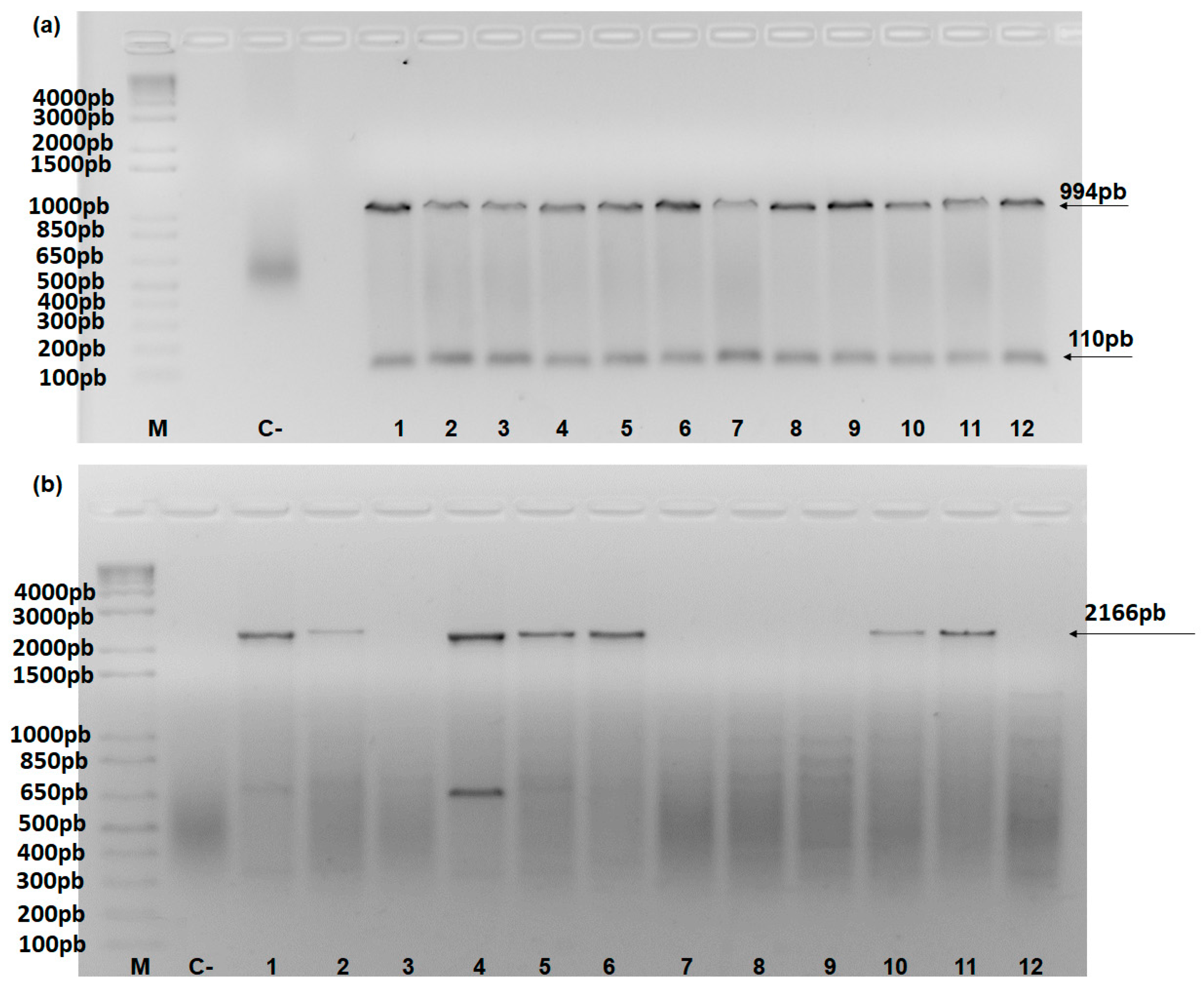
Figure 3.
PCR amplification with the PDSMb primer on accessions with different ploidy types from the Embrapa Mandioca e Fruticultura Germplasm Bank, in triplicate: 1- Zebrina (AA), 2- Gros Michel (AAA), 3- Bucaneiro (AAAA), 4- IAC 1 (AB), 5- Balbisiana Franca (BB), 6- Butuhan (ABBB), listed in Table 1. M = 50 bp marker (Promega). Arrows indicating ~332 bp fragments correspond to bands present in all samples, and ~225 bp fragments correspond only to A-genome accessions.
Figure 3.
PCR amplification with the PDSMb primer on accessions with different ploidy types from the Embrapa Mandioca e Fruticultura Germplasm Bank, in triplicate: 1- Zebrina (AA), 2- Gros Michel (AAA), 3- Bucaneiro (AAAA), 4- IAC 1 (AB), 5- Balbisiana Franca (BB), 6- Butuhan (ABBB), listed in Table 1. M = 50 bp marker (Promega). Arrows indicating ~332 bp fragments correspond to bands present in all samples, and ~225 bp fragments correspond only to A-genome accessions.
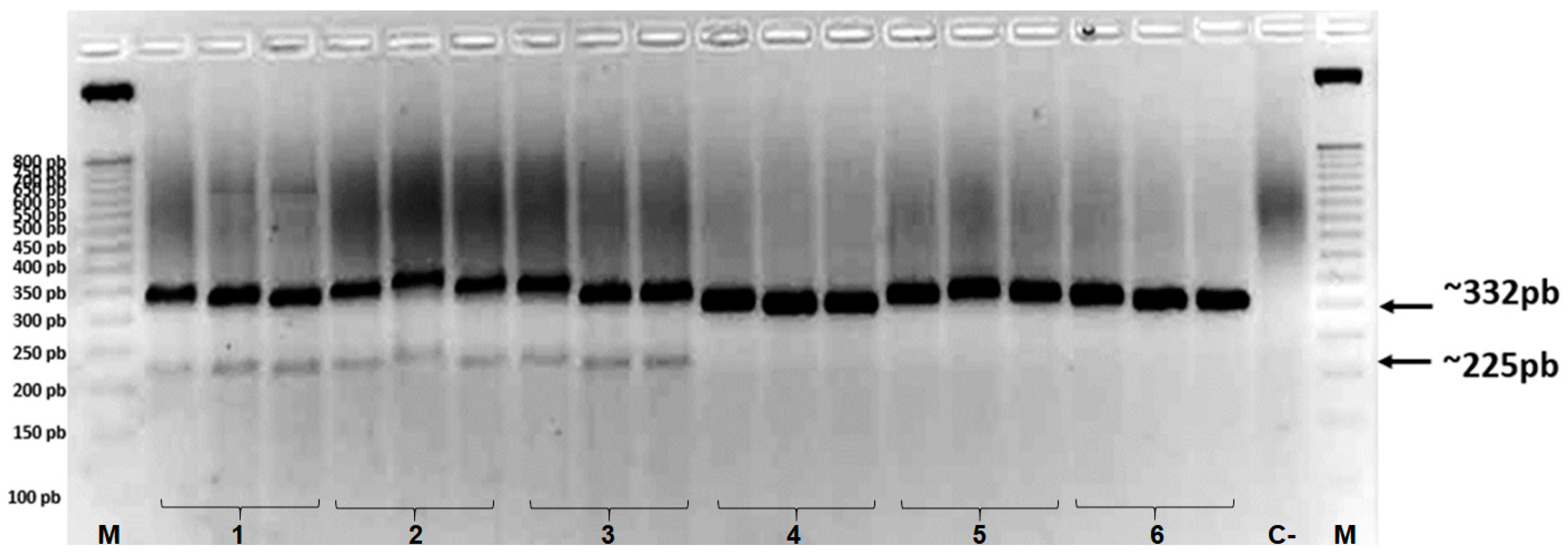

Table 1.
Primers used in the validation study of the PDS gene-derived PDSMa and PDSMb molecular markers of M. acuminata and M. balbisiana.
Table 1.
Primers used in the validation study of the PDS gene-derived PDSMa and PDSMb molecular markers of M. acuminata and M. balbisiana.
| Sequence | |||||
|---|---|---|---|---|---|
| Primers | F* (5´- 3´) | R* (5´- 3´) | At* | pb* | Reference |
| β-tubulin | ACATTGTCAGGT GGG GAGTT | CCTTTTGTTCCACACGAGATT | 60 ºC | 110 | [30] |
| PDS_AB | CAGCTAACTGAGATCAGTTT | AGATGGCTATATTTCGGTAC | 55 ºC | 994 | [26] |
| PDSMa | ATTGTGAAAGAGGTCGAGGA | TGCGGTAAAAGAAGCTTCAA | 60 ºC | 2166 | |
| PDSMb | GTGAGTTCATGGGTTGCCAA | ACCGGCTATGACAACCTTCA | 65 °C | 332 | |
Table 2.
Banana accessions from the Embrapa Mandioca e Fruticultura Germplasm Bank used in “A” and “B” genome differentiation by PCR with the β-tubulin, PDS_AB, PDSMa, and PDSMb primers.
Table 2.
Banana accessions from the Embrapa Mandioca e Fruticultura Germplasm Bank used in “A” and “B” genome differentiation by PCR with the β-tubulin, PDS_AB, PDSMa, and PDSMb primers.
| Accessions | Primer | ||||||
|---|---|---|---|---|---|---|---|
| Nº | Genotype/Cultivar | Genomic Group | Tubulin | PDS_AB | PDSMa | PDSMb ~320pb | PDSMb ~225pb |
| 1 | Gros Michel | AAA | + | + | + | + | + |
| 2 | Prata Anã | AAB | + | + | + | + | - |
| 3 | Balbisiana Franca | BB | + | + | - | + | - |
| 4 | Zebrina | AA | + | + | + | + | + |
| 5 | IAC 1 | AB | + | + | + | + | - |
| 6 | Bucaneiro | AAAA | + | + | + | + | + |
| 7 | Butuhan | BB | + | + | - | + | - |
| 8 | Musa balbisiana | BB | + | + | - | + | - |
| 9 | BB França | BB | + | + | - | + | - |
| 10 | Pelipita | ABB | + | + | + | + | - |
| 11 | FC06-02 | AABB | + | + | + | + | - |
| 12 | Teparod | ABBB | + | + | - | + | - |
| 13 | 028003-01 | AA | + | + | + | + | + |
| 14 | M53 | AA | + | + | + | + | + |
| 15 | Pisang Jaran | AA | + | + | + | + | + |
| 16 | Malbut | AA | + | + | + | + | + |
| 17 | Calcutta 4 | AA | + | + | + | + | + |
| 18 | PA Rayong | AA | + | + | + | + | + |
| 19 | Buitenzorg | AA | + | + | + | + | + |
| 20 | Khai Nai On | AA | + | + | + | + | + |
| 21 | Microcarpa | AA | + | + | + | + | + |
| 22 | Mambee Thu | AA | + | + | + | + | + |
| 23 | Malaccensis | AA | + | + | + | + | + |
| 24 | Pisang Tongat | AA | + | + | + | + | + |
| 25 | Pisang Pipit | AA | + | + | + | + | + |
| 26 | SF0751 | AA | + | + | + | + | + |
| 27 | Pisang Rojo Uter | AA | + | + | + | + | + |
| 28 | Tong Dok Mak | AA | + | + | + | + | + |
| 29 | Pisang Jari Buaya | AA | + | + | + | + | + |
| 30 | Pisang Lilin | AA | + | + | + | + | + |
| 31 | M48 | AA | + | + | + | + | + |
| 32 | NBA 14 | AA | + | + | + | + | + |
| 33 | Pisang Lidi | AA | + | + | + | + | + |
| 34 | Khai | AA | + | + | + | + | + |
| 35 | Pisang Berlin | AA | + | + | + | + | + |
| 36 | Khi Maeo | AA | + | + | + | + | + |
| 37 | Niyarma Yik | AA | + | + | + | + | + |
| 38 | Pisang Jari Buaya | AA | + | + | + | + | + |
| 39 | Tuu Gia | AA | + | + | + | + | + |
| 40 | Tjau Lagada | AA | + | + | + | + | + |
| 41 | Pisang Mas | AA | + | + | + | + | + |
| 42 | Ouro | AA | + | + | + | + | + |
| 43 | M61 | AA | + | + | + | + | + |
| 44 | Birmanie | AA | + | + | + | + | + |
| 45 | Borneo | AA | + | + | + | + | + |
| 46 | PA Musore 2 | AA | + | + | + | + | + |
| 47 | Pisang Cici | AA | + | + | + | + | + |
| 48 | 42049004 | AA | + | + | + | + | + |
| 49 | 42052004 | AA | + | + | + | + | + |
| 50 | 42079006 | AA | + | + | + | + | + |
| 51 | 42079013 | AA | + | + | + | + | + |
| 52 | 42085002 | AA | + | + | + | + | + |
| 53 | 50012002 | AA | + | + | + | + | + |
| 54 | 58054003 | AA | + | + | + | + | + |
| 55 | 73041001 | AA | + | + | + | + | + |
| 56 | 86079009 | AA | + | + | + | + | + |
| 57 | 86079010 | AA | + | + | + | + | + |
| 58 | 86094015 | AA | + | + | + | + | + |
| 59 | 86094020 | AA | + | + | + | + | + |
| 60 | 89087001 | AA | + | + | + | + | + |
| 61 | 91079003 | AA | + | + | + | + | + |
| 62 | 91087002 | AA | + | + | + | + | + |
| 63 | 91094004 | AA | + | + | + | + | + |
| 64 | SH3263 | AA | + | + | + | + | + |
| 65 | SH3362 | AA | + | + | + | + | + |
| 66 | TH03001 | AA | + | + | + | + | + |
| 67 | 42052003 | AA | + | + | + | + | + |
| 68 | 1916001 | AA | + | + | + | + | + |
| 69 | 13004006 | AA | + | + | + | + | + |
| 70 | BRS SCS Belluna | AAA | + | + | + | + | + |
| 71 | N´Jok Kon | AAB | + | + | + | + | - |
| 72 | CNPMF 0557 | AA | + | + | + | + | + |
| 73 | CNPMF0496 | AA | + | + | + | + | + |
| 74 | CNPMF0513 | AA | + | + | + | + | + |
| 75 | CNPMF0519 | AA | + | + | + | + | + |
| 76 | CNPMF0536 | AA | + | + | + | + | + |
| 77 | CNPMF0534 | AA | + | + | + | + | + |
| 78 | CNPMF0542 | AA | + | + | + | + | + |
| 79 | CNPMF0565 | AA | + | + | + | + | + |
| 80 | CNPMF0572 | AA | + | + | + | + | + |
| 81 | CNPMF0612 | AA | + | + | + | + | + |
| 82 | CNPMF0731 | AA | + | + | + | + | + |
| 83 | CNPMF0767 | AA | + | + | + | + | + |
| 84 | CNPMF0811 | AA | + | + | + | + | + |
| 85 | CNPMF0037 | AA | + | + | + | + | + |
| 86 | CNPMF0038 | AA | + | + | + | + | + |
| 87 | CNPMF 1102 | AA | + | + | + | + | + |
| 88 | CNPMF 0993 | AA | + | + | + | + | + |
| 89 | CNPMF 1323 | AA | + | + | + | + | + |
| 90 | CNPMF 1105 | AA | + | + | + | + | + |
| 91 | CNPMF 0998 | AA | + | + | + | + | + |
| 92 | CNPMF 1272 | AA | + | + | + | + | + |
| 93 | CNPMF1286 | AA | + | + | + | + | + |
| 94 | BRS Pacoua | AAAB | + | + | + | + | - |
| 95 | Yangambi Km 5 | AAA | + | + | + | + | + |
| 96 | Nanicão Cena 225 | AAA | + | + | + | + | + |
| 97 | Nanicão Jangada | AAA | + | + | + | + | + |
| 98 | IAC 2001 | AAA | + | + | + | + | + |
| 99 | IAC 504 | AAA | + | + | + | + | + |
| 100 | IAC 505 | AAA | + | + | + | + | + |
| 101 | Nanicão Viana | AAA | + | + | + | + | + |
| 102 | Rabo de Égua | AAA | + | + | + | + | + |
| 103 | Yangambi Nº2 | AAB | + | + | + | + | - |
| 104 | Saney | AAB | + | + | + | + | - |
| 105 | Thap maeo | AAB | + | + | + | + | - |
| 106 | Samura ‘B’ | AAB | + | + | + | + | - |
| 107 | Prata Jussara | AAB | + | + | + | + | - |
| 108 | Prata IAC | AAB | + | + | + | + | - |
| 109 | Tai | AAB | + | + | + | + | - |
| 110 | Pacovan | AAB | + | + | + | + | - |
| 111 | Walha | AAB | + | + | + | + | - |
| 112 | Prata Ponta Aparada | AAB | + | + | + | + | - |
| 113 | Prata Comum | AAB | + | + | + | + | - |
| 114 | Prata Manteiga | AAB | + | + | + | + | - |
| 115 | Terrinha | AAB | + | + | + | + | - |
| 116 | Curare Enano | AAB | + | + | + | + | - |
| 117 | Red Yade | AAB | + | + | + | + | - |
| 118 | D’Angola | AAB | + | + | + | + | - |
| 119 | Mongolo | AAB | + | + | + | + | - |
| 120 | Terra ponta aparada | AAB | + | + | + | + | - |
| 121 | Pinha | AAB | + | + | + | + | - |
| 122 | Tipo Velhaca | AAB | + | + | + | + | - |
| 123 | Terra maranhão | AAB | + | + | + | + | - |
| 124 | Trois Vert | AAB | + | + | + | + | - |
| 125 | Prata Baby | AAB | + | + | + | + | - |
| 126 | Prata graúda | AAB | + | + | + | + | - |
| 127 | Namwa Daeng | ABB | + | + | + | + | - |
| 128 | Ice Cream | ABB | + | + | + | + | - |
| 129 | Pitogo | ABB | + | + | + | + | - |
| 130 | Saba Honduras | ABB | + | + | + | + | - |
| 131 | Saba | ABB | + | + | + | + | - |
| 132 | Espermo | ABB | + | + | + | + | - |
| 133 | Poteau Naine | ABB | + | + | + | + | - |
| 134 | Figo | ABB | + | + | + | + | - |
| 135 | Fhia 02 | AAAA | + | + | + | + | + |
| 136 | Platina IAC | AAAB | + | + | + | + | - |
| 137 | Fhia 18 | AAAB | + | + | + | + | - |
| 138 | Prata Maçã | AAAB | + | + | + | + | - |
| 139 | BRS Japira | AAAB | + | + | + | + | - |
| 140 | BRS Pioneira | AAAB | + | + | + | + | - |
| 141 | BRS Garantida | AAAB | + | + | + | + | - |
| 142 | BRS Preciosa | AAAB | + | + | + | + | - |
| 143 | BRS Princesa | AAAB | + | + | + | + | - |
| 144 | BRS Vitoria | AAAB | + | + | + | + | - |
| 145 | YB 4203 | AAAB | + | + | + | + | - |
| 146 | Fhia 21 | AAAB | + | + | + | + | - |
| 147 | PA 42038 | AAAB | + | + | + | + | - |
| 148 | PA 42028 | AAAB | + | + | + | + | - |
| 149 | PA 42019 | AAAB | + | + | + | + | - |
| 150 | Fhia 03 | AABB | + | + | + | + | - |
(+): presence of a band; (-): absence of a band.
Table 3.
Statistical analysis, using MedCalc software, of the sensitivity, specificity, positive and negative predictive value, and accuracy of the specific markers for the PDS gene of Musa acuminata and M. balbisiana.
Table 3.
Statistical analysis, using MedCalc software, of the sensitivity, specificity, positive and negative predictive value, and accuracy of the specific markers for the PDS gene of Musa acuminata and M. balbisiana.
| Statistic | Value- PDSMa | Value- PDSMb | 95% CI- PDSMa | 95% CI- PDSMb |
|---|---|---|---|---|
| Sensitivity | 99.32% | 100.00% | 96.24% to 99.98% | 96.07% to 100.00% |
| Specificity | 100.00% | 100.00% | 39.76% to 100.00% | 93.84% to 100.00% |
| Positive Likelihood Ratio | ||||
| Negative Likelihood Ratio | 0.01 | 0.00 | 0.00 to 0.05 | |
| Disease prevalence (*) | 97.33% | 61.33% | 93.31% to 99.27% | 53.05% to 69.16% |
| Positive Predictive Value (*) | 100.00% | 100.00% | 96.07% to 100.00% | |
| Negative Predictive Value (*) | 80.00% | 100.00% | 36.19% to 96.58% | 93.84% to 100.00% |
| Accuracy (*) | 99.33% | 100.00% | 96.34% to 99.98% | 97.57% to 100.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



