
Submitted:
09 January 2024
Posted:
09 January 2024
You are already at the latest version
Abstract
Abstract: Amino acids are widely present as intermediates in marine nitrogen cycle. However, the information of amino acid distribution in deep-sea seawater, especially in abyss and hadal zone, is very limited. In the present study, we determined the concentrations of dissolved free amino acids (DFAA), dissolved combined amino acids (DCAA) and total hydrolysable amino acids (THAA) in the seawater from sea surface to hadal zone of the Yap Trench. Their occurrence, vertical variations and degradation behavior in the Yap Trench were investigated. The results showed that concentrations of ΣDFAA, ΣDCAA and ΣTHAA ranged from 0.09 to 1.78, 0.99 to 17.69 and 1.18 to 18.01 μmol/L, respectively, in the study area. In the seawater samples from the trench, Glycine (Gly) was the predominant DFAA, while the DCAA and THAA was dominated by threonine (Thr). Concentration of chlorophyll a (chl a) was not significantly correlated with any amino acids (DFAA, DCAA and THAA) in the euphotic layer. Mean concentrations for DFAA, DCAA and THAA in different water layers were in the order of: mesopelagic > euphotic > abyssal > bathyal. The concentrations of DCAA and THAA in sediment-seawater interface were higher than those from the euphotic to abyssal layer. Stations near the Yap Islands have the older organic matter (OM), while the organic matter in stations near the Yap Trench axis was relatively fresh above 1000-m depth. Degradation behavior of OM in the mesopelagic and abyssal layer are affected by transport of dissolved organic matter with water current. The organic matter in the sediment-seawater interface are older, especially on the west side of the trench. This is a survey of amino acids (DFAA, DCAA and THAA) in the Yap Trench, which provides information on the vertical variations and degradation behavior of amino acids from sea surface to hadal environment, and will be helpful to understand marine organic nitrogen cycle.
Keywords:
Amino acids
; Yap Trench
; Vertical variations
; Diagenetic
1. Introduction
Amino acids are the primary forms of organic nitrogen (ON) in most organisms (including both terrestrial and marine organisms) [1,2], play crucial roles as intermediates in marine nitrogen cycle [3,4]. As the fundamental component of protein, amino acids are present in all marine organisms [5,6], including both of living cells and residues [7,8]. As typical marine labile compounds, amino acids have attracted great attention of many scientific researchers.
Amino acids can be categorized based on their chemical bond states and physical forms in seawater [9]. These categories include particulate amino acids (PAA), dissolved free amino acids (DFAA), dissolved combined amino acids (DCAA) and total hydrolyzed amino acids (THAA = DFAA + DCAA) [10]. The production of amino acids depends on available light [5,11], so their concentration in the marine environment will decrease with depth in general [12,13]. For example, it is found that the concentrations of THAA and PAA decreased from 0.72 to 0.43 μmol/L and from 1.71 to 0.28 μmol/g, respectively, with increasing depth in the Southern Yellow Sea and the East China Sea [5,7]. Until now, the research on amino acids in marine environment is mainly focused on the surface seawater of coastal areas, while the study of the deep sea, especially the trench environment in open ocean is very limited [14]. In the west Pacific Ocean, previous study found that the concentration of DFAA in the Yap Trench generally decreased with depth [14]. However, the vertical distribution profiles of THAA and DCAA in the trench needs further study. In addition, amino acids are highly labile and easily degraded in seawater [15,16], and the production and consumption of DFAA and DCAA are closely related to bacteria and phytoplankton [17,18]. Amino acids are often used as biomarkers to provide information about decomposition and transport of organic carbon in marine environment [19,20]. For instance, the Asp/Gly ratio and serine (Ser) + Thr (mol%) can be used to evaluate the sources of diatomaceous and calcareous sources in organic matter [20]. As an important intermediate, the study of the decomposition and transportation of amino acids will help to understand the organic nitrogen and carbon cycle in the global oceans.
The Yap Trench is located at the Northwest Pacific Ocean, with the deepest depth of 8527 meters [21]. Its north part is the junction of the Pacific Plate, the Philippines Plate and the Caroline Plate, and its south part is between the Philippine Plate and the Caroline Plate, making the trench a unique area to study deep sea environment [22,23]. However, comparing the deepest Mariana Trench, there is relatively lack of attention and research on the Yap Trench. The “V-shape” structure of the Yap Trench enables it to have a weak convergence and sedimentation particle accumulation environment (i.e., funnel effect), which may become an accumulation place of amino acids and marine organism excreta [21,24,25]. The unique deep ocean currents may affect the distribution and composition of amino acids in its abyssal and hadal environment, and further affect the degradation of organic matter [3,26]. With the development of deep sea research method, more and more parameters in the Yap Trench were analyzed and their spatiotemporal distribution characteristics were studied [21,26,27]. However, the vertical distribution and degradation feature of amino acids in the trench seawater, especially DCAA and THAA, are still unclear. In addition, more research is necessary to investigate the impact of the funnel effect and bottom water current of the Yap Trench on the source and degradation of amino acids.
The overall objectives of the present study were to: (1) investigate the occurrence of amino acids in the seawater from the Yap Trench; (2) evaluate vertical variations of amino acids in the trench; (3) assess the different degradation behavior of various amino acids in the trench environment. This study will be helpful to better understand deep sea organic carbon and nitrogen cycle, especially in the abyss and hadal area.
2. Materials and Methods
2.1. Chemicals and reagents
Fourteen amino acids were selected as target analytes, including 2 acidic amino acids (aspartic acid (ASP) and glutamic acid (Glu)), 2 basic amino acids (arginine (Arg) and histidine (His)) and 10 neutral amino acids (serine (Ser), glycine (Gly), threonine (Thr), alanine (Ala), tyrosine (Tyr), valine (Val), methionine (Met), phenylalanine (Phe), isoleucine (Ile) and leucine (Leu)). All amino acid standards were obtained from Sigma (St. Louis, MO, USA). High-performance liquid chromatography (HPLC) grade acetonitrile, sodium acetate solution, tetrahydrofuran (THF) and methanol are from CNW (Germany). The derivatization reagents o-phthalaldehyde (OPA) and 3-mercaptopropionic acid (MPA) were also from Sigma. Ultra-pure water used in this study was produced by a Milli-Q purification system (Millipore Elix10, USA). Glass fiber filters (GFF, Whatman, OD 47 mm, 0.45 µm) were supplied by Whatman (Maidstone, England).
2.2. Sampling stations
Four CTD stations and five dive stations were selected in the Yap Trench (Figure 1). Detailed information on the sampling sites was listed in Table S1. The No.37 ocean cruises was conducted aboard the R/V Xiangyanghong 09, carrying “Jiaolong” manned submersible, in the Northern Yap Trench from May 13 to May 23, 2016. Seawater samples were collected from the four CTD stations in the study area through the shipborne seabird CTD equipment. Five diving stations have been set up at the sediment-seawater interface of the Yap Trench. SBE-911 plus CTD (Seabird Electronics), carried and controlled by “Jiaolong” manned submersible, collected the overlying water from sediment-seawater interface of the five stations. All the collected seawater samples were filtered by 0.45 µm glass fiber filters (Whatman GF/F) immediately on board, and stored in sample bottles at -20 °C until extraction. Temperature (T), salinity (S) and dissolved oxygen (DO) concentration were measured synchronously during seawater sampling. The pH and concentration of chlorophyll a (chl a) of the seawater samples were determined by spectrophotometric method and fluorescence spectrophotometric method, respectively [28,29]. The parameters related to temperature (T), pH, salinity (S), dissolved oxygen (DO) concentration and chlorophyll a (chl a for the euphotic layer) were shown in Table S2 and Figure S1.
2.3. Pretreatment and analysis of seawater samples
The analysis of amino acid concentrations was based on o-phthaldialdehyde-3-mercaptopropionic acid derivatization method [30]. Hydrolysis was required before THAA analysis. Specifically, the filtered seawater sample was added to an ampoule containing 6 mol/L HCl, sealed with flame in the nitrogen environment, and then the seawater sample was hydrolyzed at 110 °C for 22 h. After evaporation of the hydrolysate, the dried residue was dissolved in Milli-Q water for derivatization. The DFAA was directly derivatized without hydrolysis by the derivative reagent for 19 minutes.
Amino acids were analyzed by high-performance liquid chromatography (HPLC). The HPLC was a Waters e2695 system equipped with a quaternary pump, an online degasser, an auto sampler, a column heater and an e2475 fluorescence detector. 20 μL derivative samples were injected into the HPLC for qualitative and quantitative analysis. Amino acids separation was achieved on a ZORBAX Eclipse AAA column (150 × 3.0 mm, 5 μm; Agilent). The gradient elution of the mobile phase, are shown in Table S3, including 0.05-mol/L sodium acetate buffer solution (A) and acetonitrile: methanol꞉ Milli-Q (V/V/V) = 1꞉1꞉1 mixture (B). Specific instrumental parameters are provided in Text S1.
2.4. Diagenetic indicator
Diagenetic indicator (DI) was used to indicate the degradation degree of amino acids in the seawater. The DI was calculated based on the principal component analysis (PCA) by the following equation [31]:
where vari is the mole percentage of individual amino acid in our data set; AVGvari and STDvari are its mean and standard deviation in all samples, respectively, and fac.coefi is the factor coefficient for individual amino acid. In seawater, the more positive the DI value is, the fresher the organic matter is, indicating the amino acids mainly come from newly generated organic matter. On the contrary, the more negative the DI value is, the older the organic matter is, suggesting the amino acids have greater degradation.
2.5. Quality assurance and control
Amino acid concentrations were calculated through the calibration curves constructed by analysis of standards solutions. The calibration curves displayed strong linearity with R2 values greater than 0.999. The blank sample was analyzed to monitor the background contamination, and all target amino acids were not detected in the blank samples. Each seawater sample had three replicates and the relative standard deviation was less than 7%. The detection limit of target amino acids ranged from 30 to 90 fmol. The mean recoveries of amino acids in all samples were in the range of 78.4% to 118.6%.
2.6. Statistical analysis
Normality of the data was evaluated by Shapiro-Wilk test before further statistical analysis. Mann-Whitney U test was used to study the relationships between different groups. Spearman’s rank correlation analysis was performed to investigate the possible correlations between different individual amino acid. Origin 2022 were employed for linear correlation analysis. PCA was performed by the mole percentage of amino acids as the original data matrix. Statistical analyses were analyzed using IBM SPSS Statistics (version, 25) with a statistical significance threshold of p < 0.05. Spatial maps for broadscale trench topography were drawn using ArcGIS mapping software.
3. Results and discussion
3.1. Compositions and concentrations of amino acids
All fourteen target amino acids were detected in the seawater from the Yap Trench. The ΣTHAA concentrations were in the range of 1.18 to 18.01 μmol/L, with the mean value of 2.67 μmol/L (Table S4). Overall, the ΣDFAA concentrations in seawater (0.09 to 1.78 μmol/L) were lower than that of ΣDCAA (0.99 to 17.69 μmol/L) (p < 0.05, Figure 2). This may be because DFAA have fast turnover rates in seawater and can be directly consumed by various microbe or adsorbed on particles to reduce their concentrations in the environment [32,33,34]. A significant positive correlation was found between the concentrations of THAA and DCAA (p < 0.05, n=70). This result was in line with previous studies that DCAA is the main control factor of THAA concentration change [10,35].
Gly (nd to 0.52 μmol/L) was the most abundant in DFAA, which contributed 19.13% to ΣDFAA in the seawater from the Yap Trench on average (Figure 3 and Table S4). Series studies have found similar results in other sea areas, in which Gly was also a main component of DFAA [1,35,36]. For example, Gly had the greatest contribution to the total concentration of DFAA in spring and fall seawater samples from the Gray's Reef National Marine Sanctuary [36]. Gly was the main product of photorespiration and was released from large marine zooplankton and algae cells [37,38]. In addition, Gly was more difficult to decompose than other amino acids in the process of microbial decomposition and zooplankton digestion [39,40]. Then, with the degradation of amino acids, the molar percentage of Gly increased [31,41]. Therefore, the molar percentages of Gly are often used to indicate the degree of degradation of amino acids. Thr was another dominant amino acid with similar concentration ranges (nd to 3.65 and 0.01 to 3.66 μmol/L) in DCAA and THAA in the seawater, which accounted for 15.40% and 13.68% of the ΣDCAA and ΣTHAA, respectively (Figure 3 and Table S4). This result is also consistent with previous studies [3,42]. Gly and Thr are easier to accumulate in the cell wall, while amino acids in the cell plasma are easier to be degraded [43], because the cell wall is relatively strong and is preserved during sinking and decomposition [31]. Then the protected amino acids are more difficult to be degraded.
The comparison of DFAA, DCAA and THAA concentrations in seawater from different marine environments was listed in Table S5. In general, concentrations of DFAA (0.46±0.33 μmol/L) in the waters from the Yap Trench were higher than those in the Izmir Bay, (0.23±0.17 and 0.11±0.11 μmol/L for Inner bays and outer bay, respectively) and the East China Sea (0.35±0.17 μmol/L) [9,35], but lower than those detected in the South Yellow Sea (0.94±0.08 μmol/L) [1]. The concentrations levels (2.20±2.04 and 2.67±2.05 μmol/L, respectively) of DCAA and THAA in the Yap Trench were similar, higher than those in the South Yellow Sea (1.18±0.35 and 1.91±0.28 μmol/L, respectively) [44] and the Izmir Bay (0.76±0.40 and 0.99±0.50 μmol/L, respectively) [9]. Overall, the concentration of DFAA in the water of the Yap Trench was at a medium level, while the concentrations of DCAA and THAA were at relatively high levels. This may be attributed to different sampling times [45]. The sampling time of this study is just the period of generally high primary productivity. In fact, previous studies have found that the concentrations of THAA and DCAA in summer seawater are significantly higher than those in autumn and winter [2,3]. Moreover, the transportation of other water bodies and funnel effects also increased the concentration of THAA and DCAA in the study area (see below for details).
Proportion for acidic, neutral and basic amino acids in the seawater from the Yap Trench were summarized in Figure 4. Percentages of acidic, neutral and basic amino acids in the DFAA, DCAA and THAA were different. The results showed that the percentage of neutral amino acids (> 50%) in the DFAA, DCAA and THAA was the highest, except for the DFAA in seawater of station CTD01. This result is consistent with previous studies that neutral amino acids occupied the largest fraction of amino acid in seawater [9,44]. High fraction of neutral amino acids may be related to their source of phytoplankton [3,4]. Some relatively stable neutral amino acids (such as Ser, Gly and Thr) have been proved to exist in the siliceous exoskeleton and cell walls of diatoms with much higher abundance than those in cell plasma and membrane [15,43,46]. Algae will release more Ser, Gly and Thr from cells into seawater through cell rupture to increase their concentration after cell death and sinking [14,47].
3.2. Vertical variations of amino acids
The vertical distributions of DFAA, DCAA and THAA concentrations in the seawater of the Yap Trench are similar. As shown in Figure 5a, the highest concentrations of DFAA, DCAA and THAA appear in the euphotic layer. This could be explained by the fact that amino acids were from phytoplankton in the euphotic zone through primary production [46,48]. Mean concentrations for DFAA, DCAA and THAA varied with depth in the euphotic layer (Table S6). The mean concentrations of DFAA, DCAA and THAA in different water layers followed the order: mesopelagic zone > euphotic zone > abyssal zone > bathyal zone. The concentration of amino acids below the mesopelagic layer began to decrease gradually, and the changes in the bathyal layer and abyssal layer were relatively flat. In general, the variation trend of amino acid concentrations in the four CTD stations is basically consistent and decreases with depth.
The vertical distributions of relevant environmental factors are shown in Figure S1, which may affect the distribution of organisms and the concentration of amino acids in different water layers [3,36]. In the four stations of the trench, the temperature and pH tended to decrease with depth, while the salinity had a slight increase trend with depth. Concentration of dissolved oxygen first decreased and then increased with depth, reaching its minimum value in the mesopelagic layer. Among these factors, only a positive linear relationship between DCAA and salinity was found in this study (Figure S1, p < 0.05). Similar results were observed in the Mississippi River and the Gray's Reef National Marine Sanctuary [4,36]. This linear relationship implies that sea salinity has important impact on the vertical distribution of DCAA in the water bodies.
3.2.1. Amino acids in euphotic layer
Mean concentrations among DFAA, DCAA and THAA in the euphotic layer were different (Table S6). It was found that the maximum concentrations of DCAA and THAA occurred at 125 m depth, while the maximum concentration of DFAA appeared at 50 m. As shown in Figure 5b, the maximum concentrations of DFAA, DCAA and THAA in the four CTD stations appeared in the euphotic layer (1.78, 6.15 and 6.63 μmol/L, respectively). This might be because amino acids mainly come from plankton [3,49], while the high value areas of amino acids and chl a were only limited to the euphotic layer [2]. However, the concentration changes of amino acids (DFAA, DCAA and THAA) in the euphotic layer were complex and affected by many factors [14,35]. There were no significant correlations between concentrations of chl a and any amino acids (DFAA, DCAA and THAA) in this study. This result was inconsistent with other research [5,50,51], indicating that impact of plankton biomass on amino acids was very limited in the euphotic layer of the Yap Trench [35]. Amino acids and chl a are labile components in seawater with high turnover rates [44,52], and their concentrations are affected by various processes, such as phytoplankton production, biodegradation and ocean current [3,35]. In addition, it was found that the concentrations of DFAA, DCAA and THAA in the euphotic layer changed rapidly with depth and had a wide range. Because plankton is not only the producer of amino acids in the seawater, but also their consumer [47], complicated variations of production and consumption of amino acids caused concentrations of amino acids could not be directly correlated with the biomass (reflected by concentration of chl a) of phytoplankton in the euphotic seawater of the study area.
3.2.2. Amino acids in the mesopelagic to abyssal layer
Phytoplankton cannot carry out photosynthesis due to light limitation below 200 m depth. However, the highest mean concentrations of DFAA, DCAA and THAA occurred in the mesopelagic layer of the four CTD stations (0.52, 2.12 and 2.65 μmol/L, respectively Table S6). The concentration of amino acids in the mesopelagic seawater was affected by the degradation of dead organisms, the sinking of organic matters and microbial activity [53,54]. On the other side, prokaryotic biomass decreased with depth [55], resulting in a much lower consumption and degradation rate of amino acids by microorganisms in the mesopelagic layer than in the euphotic layer. Studies have reported that up to 91% of amino acids were degraded in the Equatorial Pacific euphotic zone [46]. Moreover, compared to the euphotic layer, lower oxygen and lower temperature in the mesopelagic layer also reduce microbial activity, thereby decreasing the direct utilization rate of amino acids [40,56]. Previous research has proven that low temperature (≤ 2℃) seemed to decrease the uptake of amino acids [17]. A previous study reported higher concentrations of amino acids (such as Ala and Ser) were found in deep water of the East Japan Sea, suggesting that bacteria from the mesopelagic waters contributed significantly to amino acids [19]. Our results show that Ala+Ser had a higher concentration in the mesopelagic seawater, accounting for 22.98%, 11.81%, and 13.89% of DFAA, DCAA, and THAA, respectively. These proportions were generally consistent with previous research results, with Ala+Ser accounting for 19% of THAA in seawater samples from the East Frisian islands [54].
Interestingly, the mean concentrations of DFAA, DCAA and THAA in the bathypelagic layer were lower than those in the abyssal layer of the trench, which might be attributed to the funnel effect and transportation of amino acids from other water bodies. In fact, the velocity of horizontal flow decreased with increasing water depth in typical V-shaped terrain of trenches [57,58], resulting in a large accumulation of organic matter in deeper water (abyssal layer) [21,27]. Moreover, the seawater of the abyssal layer in the Yap Trench come from the Submerged Water around the Antarctic [59]. A large amount of organic matter has been accumulated in the abyssal seawater of the Pacific Ocean under the action of the thermohaline circulation. Then, the organic matter in the abyssal layer were degraded by heterotrophic microorganisms to increase concentrations of amino acids [60]. Studies have found that the activity of heterotrophic microorganisms in deep trenches was significantly enhanced compared to shallow reference site [61], resulting in more effective decomposition of organic matter and release of amino acids. On the other hand, water bodies in the abyssal layer may be in progress vertical exchange with water bodies at the sediment-seawater interface. The water body at the sediment-seawater interface was rich in amino acids (see below for details), resulting in higher concentrations for amino acids in the abyssal layer than in the bathypelagic layer.
3.2.3. Amino acids in the sediment-seawater interface
The mean concentrations of DCAA and THAA (4.76 and 5.25 μmol/L, respectively, Table S6) in the seawater of the five diving station were higher than those in the seawater from the CTD stations, which might be ascribed to the sediment resuspension. The resuspension of sediment increased the vertical exchange capacity of pore water, resulting in the release of amino acids to overlying seawater, thus increasing the concentration of amino acids at the sediment-seawater interface [62,63]. Series previous studies have found that sediment resuspension (Abiotic factors) had the greatest impact on dissolved amino acids dynamic process in the Mississippi River Plume [4]. The significant contribution of resuspended material to THAA pools has been confirmed because the existence of a positive correlation between THAA and suspended particulate matter (SPM) was found in the seawater samples from the East Frisian islands [54]. Pore water directly and indirectly affected the concentration and composition of THAA, which is an important source of amino acids in water column [64]. Since the concentration of solute in sediment pore water and overlying water was different, solute could diffuse across the sediment-seawater interface to overlying water [13]. This diffusion process resulted in higher concentrations of DCAA and THAA in the seawater of the five diving stations close to the sediment than in the seawater of the CTD stations. The concentrations (17.69 and 18.01 μmol/L for Dive109, respectively) of DCAA and THAA on the west of the trench were all higher than that (1.29 and 1.49 μmol/L for Dive113, respectively) on the east at sediment-seawater interface of the trench. Similar result was also found at the sediment-seawater interface in the Yap Trench [14]. This phenomenon could be attributed to the action of the Coriolis force, the sediment on the western trench wall was washed away by the bottom water facing south, forming a resuspension of the surface sediment [14,63]. Resuspension of sediment released some amino acids into the seawater, increasing the concentration of amino acids on the seawater at the sediment-seawater interface at the west of the trench.
3.3. Sources and degradation of amino acids
Correlation analysis of various amino acids can be used to extrapolate quantitative similarities and differences of individual amino acids [3]. Spearman rank correlation was performed between individual amino acids in the seawater (Table S7). As the predominant DFAA, concentration of Gly was significantly negatively correlated with concentrations of Glu and Asp (p < 0.01). This negative correlation suggested that these amino acids might have different dynamic behavior in seawater [3]. In fact, the fundamental chemical properties of these amino acids were different because Gly was a neutral amino acid, while Glu and Asp were acidic amino acids. Gly is mainly related to structural substances and constitute degradation products of other amino acids (such as Asp and Glu) [15,65], but Glu tends to convert into non-protein amino acids and indicate freshness of POM (particulate organic matter) [66,67]. Therefore, the degradation of POM leaded to an increase in the contribution of Gly and a decrease in the contribution of Glu [62]. Moreover, some researchers have found that the contribution of Asp decreased during the degradation of organic matter [68,69]. Overall, Gly exhibits different dynamic behaviors from Asp and Glu in seawater [62]. Thr, as a main amino acid in DCAA and THAA, was significantly positively correlated with 5 amino acids (Ala, Arg, Gly, His and Ser) in DCAA and 7 amino acids (Ala, Arg, Asp, Glu, Gly, His and Ser) in THAA (p < 0.01). The positive correlationships indicated that these amino acids might be affected by similar mechanisms of biogeochemical alteration [1]. For instance, Ser and Thr have been confirmed to exist in diatom cell wall [43] and are enriched in diatom mediated silicification proteins [70]. This kind of coexistence might help them share similar biochemical processes and then formed a positive relationship. All of the above amino acids belong to the aliphatic group except for His, which can directly participate in cellular metabolic processes [71]. Among them, Thr, Gly and Ser can be easily transformed into each other in plant catabolism [72]. Therefore, it is reasonable to find positive relationships between concentrations of Thr and Gly and Ser.
PCA was further used to characterize the relationships among various amino acids. As shown in Figure 6, principal components 1 (PC1) for DFAA, DCAA and THAA explained 37.7%, 47.6% and 44.1% of the respective total variance, respectively. PC1 for DFAA exhibited high positive loadings for Ala, Tyr and Ser, and high negative loadings for acidic amino acids (Asp and Glu) (Table S8). This result revealed that the degradation processes of Ala, Tyr and Ser were exactly opposite to that of Asp and Glu. By contrast, PC1 for DCAA and THAA had positive high loading of Ala, Arg, Gly and Thr, and had negative loadings for Leu, Met and Val. These relationships indicated that the amino acids with approximate principal component loadings might share similar sources and degradation behavior in the Yap Trench [35]. In general, principal component loadings of individual amino acid may directly affect the DI value [31,42]. Because amino acids are the main source of carbon and nitrogen for microorganisms [44,73], some amino acids are accumulated in organism must lead to the consumption of others through selective utilization by microbes [35,74]. For instance, ornithine can be transferred to the plastid and re-participate in the synthesis of Arg during the nitrogen cycle [75]. Glu was a metabolite of His in animals, which undergoes four enzymatic steps involving histidine ammonia-lyase, urocanatehydratase, imidazole propionase, and formimidoylglutamase [72].
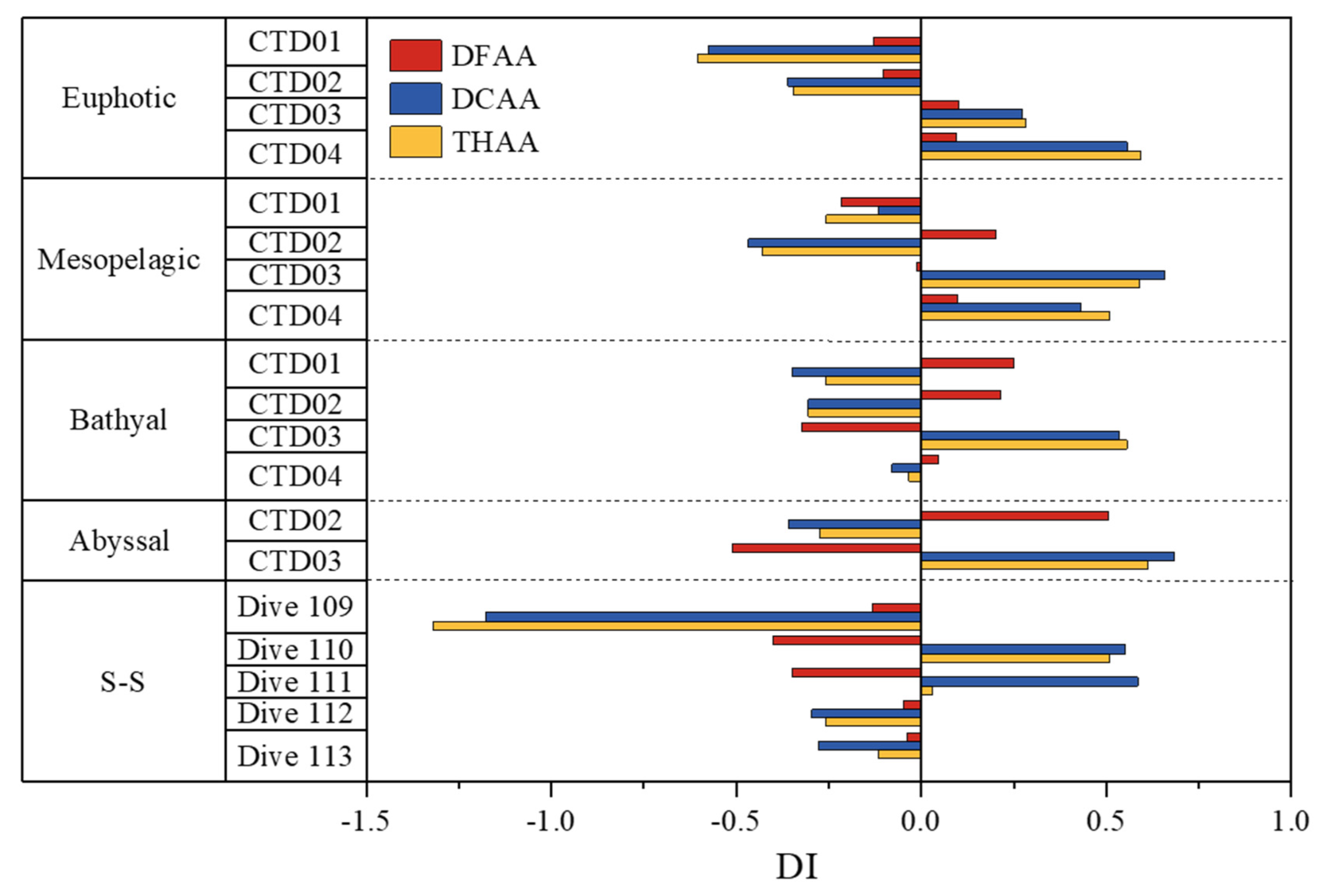
Based on the PC1 data, DI values of the seawater of the four CTD stations and the five dive stations were calculated. The DI values for the DFAA, DCAA and THAA were in the range of -0.73 to 1.52, -1.17 to 1.94 and -1.32 to 1.87, respectively (Table S9). Mean DI value for DFAA, DCAA and THAA were variable in different layers of the seawater from the Yap Trench (Figure 7). The DI values for DFAA, DCAA and THAA of the seawater in stations CTD01 and CTD02 presented negative values, while they were generally positive in the seawater of stations CTD03 and CTD04 in the euphotic and mesopelagic layers. These comparative relationships implied that the seawater from the euphotic and mesopelagic layers of stations CTD01 and CTD02 had the older organic matter, while the organic matter in stations CTD03 and CTD04 (near Yap Trench axis) was relatively fresh [31,35]. The stations CTD01 and CTD02 near the Yap Islands might be affected by input of old terrestrial organic matter. On the other side, Gly was stored within the refractory cell wall of diatoms, the molar percentage of Gly increases during the decay process of diatoms [40]. Therefore, the molar percentage of Gly can reflect the degree of organic matter degradation partially. In fact, the Gly molar percent of DCAA (mean 10.5% and 10.7%, respectively) in the seawater (above 1000m) of stations CTD01 and CTD02 were higher than those in the seawater of stations CTD03 and CTD04 (mean 5.1% and 3.5%, respectively). These results indicated that the degradation degree of organic matter in the seawater of stations CTD01 and CTD02 were relatively higher than that of stations CTD03 and CTD04. Unlike the euphotic and mesopelagic layers, the DI value of DFAA was exactly opposite to the DI value of DCAA and THAA in the seawater below 1000 m depth. These results indicated that seawater stratification at 1000 m depth impeded the vertical mixing process of seawater. Therefore, the dissolved organic matter in the deep layer of the trench might originate from the release of settled particulate organic matter to seawater and the transport of dissolved organic matter by water currents. Some previous studies have shown that the overlying water of the sediments and the seawater of the abyssal layer in the Yap Trench come from the bathypelagic seawater and submerged seawater around the Antarctic, respectively [59,76,77]. The deep water in the Yap Trench was a part of the west bar of the Lower Circumpolar Deep Water (LCPW) [26]. However, identifying the specific sources and degradation of organic matter in deep environments is extremely difficult and requires further exploration. More work is still necessary in the future.
The DI values of the seawater samples from the five diving stations were generally negative, with the minimum negative value (-1.32) for THAA appearing at station Dive 109, which could be explained by the effect of sediment resuspension and the LCPW. Negative DI values may be related to degradation of amino acids and old organic matter, and consistent results have previously been reported of the seawater from the Mississippi River and the St. Lawrence system (Canada) [4,64]. The sediment resuspension caused the diffusion of old organic matter in long-term pore water towards the sediment-seawater interface [13]. Under the influence of Coriolis force, the sediments in the western trench wall were severely scoured by the bottom water facing south [14,63]. This increased the resuspension of sediment in the western trench wall, resulting in an increase in the concentration of old organic matter in the abyss and hadal zone of the trench. Therefore, it is reasonable the minimum negative value (-1.32) for THAA appeared in station Dive 109 near the western trench. Moreover, the deep water in the Yap Trench originated in the westward propagating the LCPW [26]. The estimated net transport capacity of LCPW entering the Philippine Basin through the Yap Trench is 0.4 × 106 m3/s [78]. After a long period of transportation, the old and highly degraded organic matter carried by LCPW was transported to the sediment-seawater interface of the trench, resulting in negative DI values. In the seawater sample from the sediment-seawater interface, the molar percentage of Gly was positively correlated with DI for the DFAA, DCAA and THAA (p < 0.05), indicating that Gly in the Yap Trench originated from the new production of phytoplankton. Because Gly was considered to be more difficult to decompose than other amino acids during the degradation process of algae [39,40].
4. Conclusions
This study investigated the occurrence, vertical variations and degradation behavior of THAA, DFAA and DCAA in the seawater from the Yap Trench. Gly was the predominant DFAA in the seawater samples, while Thr was more abundant in the DCAA and THAA. Concentration of DFAA in the seawater of the Yap Trench seawater was equivalent to level around the world, while the levels of both DCAA and THAA were relatively high. The mean concentration of amino acids in the seawater from the Yap Trench followed the order: mesopelagic > euphotic > abyssal > bathypelagic. Compared to the CTD stations, the mean concentrations of DCAA and THAA in the seawater of the diving stations were higher. Above 1000-m depth, organic matter in the seawater from stations CTD01 and CTD02 was older, while the organic matter in the seawater from stations CTD03 and CTD04 was relatively fresh. The sources of amino acids in the mesopelagic and abyssal layer of the trench were related to transport of dissolved organic matter by water current. Compare to the seawater from the CTD stations, the seawater from the sediment-seawater interface had older organic matter, especially in the western side of the trench, attributed to the sediment resuspension and the LCPW transport. Further studies on sources and degradation of amino acids as they move with deep water mass in the Yap Trench are necessary.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, C.S.; data curation, C.Z.; investigation, J.N.; writing—original draft, S.X.; writing—review and editing, W.H., and H.D. All authors have read and agreed to the published version of the manuscript.
Acknowledgements
The author would like to thank the financial support by the National Key Research and Development Program of China (No. 2022YFC2803803), the National Natural Science Foundation of China (No. 42076040), and the 111 project (No. Fundamental Funds for Central Universities (No. B13030)
References
- Yang, G.P.; Chen, Y.; Gao, X.C. Distribution of Dissolved Free Amino Acids, Dissolved Inorganic Nitrogen and Chlorophyll a in the Surface Microlayer and Subsurface Water of the Yellow Sea, China. Cont. Shelf Res. 2009, 29, 1737–1747. [Google Scholar] [CrossRef]
- Zhang, P.Y.; Yang, G.P.; Chen, Y.; Zhang, J. Temporal and Spatial Distribution of Dissolved Amino Acids in the Surface Microlayer and Subsurface Water of the Bohai Sea and the Yellow Sea. J. Mar. Syst. 2021, 219, 103543. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, G.; Chen, Y.; Leng, W.; Ji, C. Temporal and Spatial Variations of Particulate and Dissolved Amino Acids in the East China Sea. Mar. Chem. 2016, 186, 133–144. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Grace, B.L.; Carman, K.R.; Maulana, I. Amino Acid Cycling in the Mississippi River Plume and Effects from the Passage of Hurricanes Isadore and Lili. J. Mar. Syst. 2014, 136, 10–21. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, P.; Shi, D.; Ji, C.X.; Chen, R.; Gao, X.C.; Yang, G.P. Distribution and Bioavailability of Dissolved and Particulate Organic Matter in Different Water Masses of the Southern Yellow Sea and East China Sea. J. Mar. Syst. 2021, 222, 103596. [Google Scholar] [CrossRef]
- Wei, J.E.; Chen, Y.; Wang, J.; Yan, S.B.; Zhang, H.H.; Yang, G.P. Amino Acids and Amino Sugars as Indicators of the Source and Degradation State of Sedimentary Organic Matter. Mar. Chem. 2021, 230. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, G.P.; Liu, L.; Zhang, P.Y.; Leng, W.S. Sources, Behaviors and Degradation of Dissolved Organic Matter in the East China Sea. J. Mar. Syst. 2016, 155, 84–97. [Google Scholar] [CrossRef]
- Niggemann, J.; Schubert, C.J. Sources and Fate of Amino Sugars in Coastal Peruvian Sediments. Geochim. Cosmochim. Acta 2006, 70, 2229–2237. [Google Scholar] [CrossRef]
- Alyuruk, H.; Kontas, A. Dissolved Free, Total and Particulate Enantiomeric Amino Acid Levels in Eutrophic and Oligotrophic Parts of a Semi-Enclosed Bay (İzmir, Aegean Sea). Reg. Stud. Mar. Sci. 2021, 44, 101750. [Google Scholar] [CrossRef]
- Xia, Q.; Yang, G.; Gao, X.; Chen, Y.; Pang, B. Distribution and Composition of Dissolved Amino Acids in Surface Water of Northern South China Sea. Mar. Environ. Sci. 2011, 30, 774–779. [Google Scholar]
- Shen, Y.; Fichot, C.G.; Liang, S.K.; Benner, R. Biological Hot Spots and the Accumulation of Marine Dissolved Organic Matter in a Highly Productive Ocean Margin. Limnol. Oceanogr. 2016, 61, 1287–1300. [Google Scholar] [CrossRef]
- Henrichs, S.M.; Farrington, J.W.; Lee, C. Peru Upwelling Region Sediments near 15°S. 2. Dissolved Free and Total Hydrolyzable Amino Acids. Limnol. Oceanogr. 1984, 29, 20–34. [Google Scholar] [CrossRef]
- Landén, A.; Hall, P.O.J. Benthic Fluxes and Pore Water Distributions of Dissolved Free Amino Acids in the Open Skagerrak. Mar. Chem. 2000, 71, 53–68. [Google Scholar] [CrossRef]
- Niu, J.; Sun, C.; Yang, B.; Xie, L.; Jiang, F.; Cao, W.; Chen, Y.; Ding, H.; Huang, Y.; Gao, X. Vertical Variations and Composition of Dissolved Free Amino Acid in the Seawater of the Yap Trench in the Western Pacific Ocean. J. Oceanol. Limnol. 2022. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, G.P.; Ji, C.X.; Zhang, H.H.; Zhang, P.Y. Sources and Degradation of Sedimentary Organic Matter in the Mud Belt of the East China Sea: Implications from the Enantiomers of Amino Acids. Org. Geochem. 2018, 116, 51–61. [Google Scholar] [CrossRef]
- Gupta, L.P.; Kawahata, H. Amino Acids and Hexosamines in the Hess Rise Core during the Past 220,000 Years. Quat. Res. 2003, 60, 394–403. [Google Scholar] [CrossRef]
- Coffin, R.B. Bacterial Uptake of Dissolved Free and Combined Amino Acids in Estuarine Waters. Limnol. Oceanogr. 1989, 34, 531–542. [Google Scholar] [CrossRef]
- Rosenstock, B.; Simon, M. Use of Dissolved Combined and Free Amino Acids by Planktonic Bacteria in Lake Constance. Limnol. Oceanogr. 1993, 38, 1521–1531. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, G.; Shen, Y.; Benner, R. Strong Linkages between Surface and Deep-Water Dissolved Organic Matter in the East/Japan Sea. Biogeosciences 2017, 14, 2561–2570. [Google Scholar] [CrossRef]
- Wei, J.E.; Chen, Y.; Zhang, N.; Yang, J.Q.; Chen, R.; Zhang, H.H.; Yang, G.P. Variability and Composition of Amino Acids and Amino Sugars in Sediment Cores of the Changjiang Estuary. Org. Geochem. 2022, 163, 104330. [Google Scholar] [CrossRef]
- Li, D.; Zhao, J.; Yao, P.; Liu, C.; Sun, C.; Chen, J.; Pan, J.; Han, Z.; Hu, J. Spatial Heterogeneity of Organic Carbon Cycling in Sediments of the Northern Yap Trench: Implications for Organic Carbon Burial. Mar. Chem. 2020, 223. [Google Scholar] [CrossRef]
- Ding, H.; Sun, C. Towards the Understanding from Sea Surface to Hadal Zone—A Multidisciplinary Study of the Yap Trench. J. Oceanol. Limnol. 2020, 38, 591–592. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, G. Geochemical and Chronological Evidence for Collision of Proto-Yap Arc/Caroline Plateau and Rejuvenated Plate Subduction at Yap Trench. Lithos 2020, 370–371. [Google Scholar] [CrossRef]
- Lee, S.M. Deformation from the Convergence of Oceanic Lithosphere into Yap Trench and Its Implications for Early-Stage Subduction. J. Geodyn. 2004, 37, 83–102. [Google Scholar] [CrossRef]
- Su, H.; Wu, C.; Han, P.; Liu, Z.; Liang, M.; Zhang, Z.; Wang, Z.; Guo, G.; He, X.; Pang, J.; et al. The Microbiome and Its Association with Antibiotic Resistance Genes in the Hadal Biosphere at the Yap Trench. J. Hazard. Mater. 2022, 439, 129543. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, X.; Lv, X.; Cao, W.; Sun, C.; Lu, J.; Wang, C.; Lu, B.; Yang, J. Watermass Properties and Deep Currents in the Northern Yap Trench Observed by the Submersible Jiaolong System. Deep. Res. Part I Oceanogr. Res. Pap. 2018, 139, 27–42. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, W.; Liu, Y.; Cai, M.; Luo, Z.; Li, M. Metagenomics Reveals Microbial Diversity and Metabolic Potentials of Seawater and Surface Sediment from a Hadal Biosphere at the Yap Trench. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.I. A Manual of Chemical & Btologwal Methods for 5eawater Analysts T R Parsons, Y Malta & C M Lalll Pergamon Press, Oxford, 1984 184 Pp, 7 Illustrations and 120 Literature References Hard-Backed £1225 ISBN 0 08 030288 2 Soft-Backed £5 50 ISBN 0 08 030287 4. Mar. Pollut. Bull. 1984, 15, 419–420. [Google Scholar] [CrossRef]
- Dickson, A.; Sabine, C.; Christian, J. Guide to Best Practices for Ocean C02 Measurments; 2007; Vol. 3; ISBN 1897176074.
- Lindroth, P.; Mopper, K. High Performance Liquid Chromatographic Determination of Subpicomole Amounts of Amino Acids by Precolumn Fluorescence Derivatization with O-Phthaldialdehyde. Anal. Chem. 1979, 51, 1667–1674. [Google Scholar] [CrossRef]
- Dauwe, B.; Middelburg, J.J.; Herman, P.M.J.; Heip, C.H.R. Linking Diagenetic Alteration of Amino Acids and Bulk Organic Matter Reactivity. Limnol. Oceanogr. 1999, 44, 1809–1814. [Google Scholar] [CrossRef]
- Lee, C.; Bada, J.L. Amino Acids in Equatorial Pacific Ocean Water. Earth Planet. Sci. Lett. 1975, 26, 61–68. [Google Scholar] [CrossRef]
- Middelboe, M.; Borch, N.H.; Kirchman, D.L. Bacterial Utilization of Dissolved Free Amino Acids, Dissolved Combined Amino Acids and Ammonium in the Delaware Bay Estuary: Effects of Carbon and Nitrogen Limitation. Mar. Ecol. Prog. Ser. 1995, 128, 109–120. [Google Scholar] [CrossRef]
- Svensson, E.; Skoog, A.; Amend, J.P. Concentration and Distribution of Dissolved Amino Acids in a Shallow Hydrothermal System, Vulcano Island (Italy). Org. Geochem. 2004, 35, 1001–1014. [Google Scholar] [CrossRef]
- Ji, C.X.; Yang, G.P.; Chen, Y.; Zhang, P.Y. Distribution, Degradation and Bioavailability of Dissolved Organic Matter in the East China Sea. Biogeochemistry 2019, 142, 189–207. [Google Scholar] [CrossRef]
- Lu, X.; Zou, L.; Clevinger, C.; Liu, Q.; Hollibaugh, J.T.; Mou, X. Temporal Dynamics and Depth Variations of Dissolved Free Amino Acids and Polyamines in Coastal Seawater Determined by High-Performance Liquid Chromatography. Mar. Chem. 2014, 163, 36–44. [Google Scholar] [CrossRef]
- Ogren, W.L.; Chollet, R. Photorespiration; 1982; ISBN 0122943023.
- Webb, L.; Johannes, R.E. Studies of the Release of Dissolved Free Amino Acids by Marine Zooplankton. Limnol. Oceanogr. 1967, 12, 376–382. [Google Scholar] [CrossRef]
- Cowie, G.L.; Hedges, J.I. Digestion and Alteration of the Biochemical Constituents of a Diatom (Thalassiosira Weissflogii) Ingested by an Herbivorous Zooplankton (Calanus Pacificus). Limnol. Oceanogr. 1996, 41, 581–594. [Google Scholar] [CrossRef]
- Nguyen, R.T.; Harvey, H.R. Protein and Amino Acid Cycling during Phytoplankton Decomposition in Oxic and Anoxic Waters. Org. Geochem. 1997, 27, 115–128. [Google Scholar] [CrossRef]
- Dittmar, T.; Fitznar, H.P.; Kattner, G. Origin and Biogeochemical Cycling of Organic Nitrogen in the Eastern Arctic Ocean as Evident from D- and L-Amino Acids. Geochim. Cosmochim. Acta 2001, 65, 4103–4114. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, G.P.; Wu, G.W.; Gao, X.C.; Xia, Q.Y. Concentration and Characterization of Dissolved Organic Matter in the Surface Microlayer and Subsurface Water of the Bohai Sea, China. Cont. Shelf Res. 2013, 52, 97–107. [Google Scholar] [CrossRef]
- Hecky, R.E.; Mopper, K.; Kilham, P.; Degens, E.T. The Amino Acid and Sugar Composition of Diatom Cell-Walls. Mar. Biol. 1973, 19, 323–331. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, G.P.; Xia, Q.Y.; Wu, G.W. Enrichment and Characterization of Dissolved Organic Matter in the Surface Microlayer and Subsurface Water of the South Yellow Sea. Mar. Chem. 2016, 182, 1–13. [Google Scholar] [CrossRef]
- Wedyan, M.A.; Preston, M.R. The Coupling of Surface Seawater Organic Nitrogen and the Marine Aerosol as Inferred from Enantiomer-Specific Amino Acid Analysis. Atmos. Environ. 2008, 42, 8698–8705. [Google Scholar] [CrossRef]
- Ingalls, A.E.; Lee, C.; Wakeham, S.G.; Hedges, J.I. The Role of Biominerals in the Sinking Flux and Preservation of Amino Acids in the Southern Ocean along 170°W. Deep. Res. Part II Top. Stud. Oceanogr. 2003, 50, 713–738. [Google Scholar] [CrossRef]
- Zhang, P.Y.; Chen, Y.; Zhang, J.; Yang, G.P. Temporal and Spatial Variations of Particulate and Dissolved Amino Acids in the East China Sea. Mar. Chem. 2015, 46, 329–339. [Google Scholar] [CrossRef]
- Kaiser, K.; Benner, R. Organic Matter Transformations in the Upper Mesopelagic Zone of the North Pacific: Chemical Composition and Linkages to Microbial Community Structure. J. Geophys. Res. Ocean. 2012, 117, 1–12. [Google Scholar] [CrossRef]
- Cowie, G.L.; Hedges, J.I. Sources and Reactivities of Amino Acids in a Coastal Marine Environment. Limnol. Oceanogr. 1992, 37, 703–724. [Google Scholar] [CrossRef]
- Meon, B.; Kirchman, D.L. Dynamics and Molecular Composition of Dissolved Organic Material during Experimental Phytoplankton Blooms. Mar. Chem. 2001, 75, 185–199. [Google Scholar] [CrossRef]
- Sabadel, A.J.M.; Browning, T.J.; Kruimer, D.; Airs, R.L.; Woodward, E.M.S.; Van Hale, R.; Frew, R.D. Determination of Picomolar Dissolved Free Amino Acids along a South Atlantic Transect Using Reversed-Phase High-Performance Liquid Chromatography. Mar. Chem. 2017, 196, 173–180. [Google Scholar] [CrossRef]
- Lee, C.; Wakeham, S.G.; I. Hedges, J. Composition and Flux of Particulate Amino Acids and Chloropigments in Equatorial Pacific Seawater and Sediments; 2000; Vol. 47; ISBN 0015166328820. 0015.
- Lee, C.; Bada, J.L. Dissolved Amino Acids in the Equatorial Pacific, the Sargasso Sea, and Biscayne Bay. Limnol. Oceanogr. 1977, 22, 502–510. [Google Scholar] [CrossRef]
- Behrends, B.; Liebezeit, G. Particulate Amino Acids in Wadden Sea Waters — Seasonal and Tidal Variations. J. Sea Res. 1999, 41, 141–148. [Google Scholar] [CrossRef]
- Sohrin, R.; Imazawa, M.; Fukuda, H.; Suzuki, Y. Full-Depth Profiles of Prokaryotes, Heterotrophic Nanoflagellates, and Ciliates along a Transect from the Equatorial to the Subarctic Central Pacific Ocean. Deep. Res. Part II Top. Stud. Oceanogr. 2010, 57, 1537–1550. [Google Scholar] [CrossRef]
- Veuger, B.; Middelburg, J.J.; Boschker, H.T.S.; Nieuwenhuize, J.; Van Rijswijk, P.; Rochelle-Newall, E.J.; Navarro, N. Microbial Uptake of Dissolved Organic and Inorganic Nitrogen in Randers Fjord. Estuar. Coast. Shelf Sci. 2004, 61, 507–515. [Google Scholar] [CrossRef]
- Turnewitsch, R.; Falahat, S.; Stehlikova, J.; Oguri, K.; Glud, R.N.; Middelboe, M.; Kitazato, H.; Wenzhöfer, F.; Ando, K.; Fujio, S.; et al. Recent Sediment Dynamics in Hadal Trenches: Evidence for the Influence of Higher-Frequency (Tidal, near-Inertial) Fluid Dynamics. Deep. Res. Part I Oceanogr. Res. Pap. 2014, 90, 125–138. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, S.; Gao, J.; Tian, L.; Yang, J.; Xu, Y. Geology of the Yap Trench: New Observations from a Transect near 10°N from Manned Submersible Jiaolong. Int. Geol. Rev. 2018, 60, 1941–1953. [Google Scholar] [CrossRef]
- Kaneko, I.; Takatsuki, Y.; Kamiya, H.; Kawae, S. Water Property and Current Distributions along the WHP-P9 Section (137°-142°E) in the Western North Pacific. J. Geophys. Res. Ocean. 1998, 103, 12959–12984. [Google Scholar] [CrossRef]
- Ianiri, H.L.; McCarthy, M.D. Compound Specific Δ15N Analysis of Amino Acids Reveals Unique Sources and Differential Cycling of High and Low Molecular Weight Marine Dissolved Organic Nitrogen. Geochim. Cosmochim. Acta 2023, 344, 24–39. [Google Scholar] [CrossRef]
- Glud, R.N.; Wenzhöfer, F.; Middelboe, M.; Oguri, K.; Turnewitsch, R.; Canfield, D.E.; Kitazato, H. High Rates of Microbial Carbon Turnover in Sediments in the Deepest Oceanic Trench on Earth. Nat. Geosci. 2013, 6, 284–288. [Google Scholar] [CrossRef]
- Pusceddu, A.; Grémare, A.; Escoubeyrou, K.; Amouroux, J.M.; Fiordelmondo, C.; Danovaro, R. Impact of Natural (Storm) and Anthropogenic (Trawling) Sediment Resuspension on Particulate Organic Matter in Coastal Environments. Cont. Shelf Res. 2005, 25, 2506–2520. [Google Scholar] [CrossRef]
- Tappin, A.D.; Millward, G.E.; Fitzsimons, M.F. Distributions, Cycling and Recovery of Amino Acids in Estuarine Waters and Sediments. Environ. Chem. Lett. 2007, 5, 161–167. [Google Scholar] [CrossRef]
- Fejjar, S.; Melanson, A.; Tremblay, L. Pore Waters as a Contributor to Deep-Water Amino Acids and to Deep-Water Dissolved Organic Matter Concentration and Composition in Estuarine and Marine Waters. Mar. Chem. 2021, 233. [Google Scholar] [CrossRef]
- Macko, S.A.; Estep, M.L.F. Microbial Alteration of Stable Nitrogen and Carbon Isotopic Compositions of Organic Matter. Org. Geochem. 1984, 6, 787–790. [Google Scholar] [CrossRef]
- Lee, C.; Cronin, C. Particulate Amino Acids in the Sea: Effects of Primary Productivity and Biological Decomposition. J. Mar. Res. 1984, 42, 1075–1097. [Google Scholar] [CrossRef]
- Dauwe, B.; Middelburg, J.J. Amino Acids and Hexosamines as Indicators of Organic Matter Degradation State in North Sea Sediments. Limnol. Oceanogr. 1998, 43, 782–798. [Google Scholar] [CrossRef]
- Medernach, L.; Grémare, A.; Amoureux, J.M.; Colomines, J.C.; Vétion, G. Temporal Changes in the Amino Acid Contents of Particulate Organic Matter Sedimenting in the Bay of Banyuls-Sur-Mer (Northwestern Mediterranean). Mar. Ecol. Prog. Ser. 2001, 214, 55–65. [Google Scholar] [CrossRef]
- Colombo, J.C.; Silverberg, N.; Gearing, J.N. Amino Acid Biogeochemistry in the Laurentian Trough: Vertical Fluxes and Individual Reactivity during Early Diagenesis. Org. Geochem. 1998, 29, 933–945. [Google Scholar] [CrossRef]
- Lobel, K.D.; West, J.K.; Hench, L.L. Computational Model for Protein-Mediated Biomineralization of the Diatom Frustule. Mar. Biol. 1996, 126, 353–360. [Google Scholar] [CrossRef]
- Fichtner, M.; Voigt, K.; Schuster, S. The Tip and Hidden Part of the Iceberg: Proteinogenic and Non-Proteinogenic Aliphatic Amino Acids. Biochim. Biophys. Acta - Gen. Subj. 2017, 1861, 3258–3269. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araújo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Reinthaler, T.; Sintes, E.; Herndl, G.J. Dissolved Organic Matter and Bacterial Production and Respiration in the Sea-Surface Microlayer of the Open Atlantic and the Western Mediterranean Sea. Limnol. Oceanogr. 2008, 53, 122–136. [Google Scholar] [CrossRef]
- Yamashita, Y.; Tanoue, E. Distribution and Alteration of Amino Acids in Bulk DOM along a Transect from Bay to Oceanic Waters. Mar. Chem. 2003, 82, 145–160. [Google Scholar] [CrossRef]
- Slocum, R.D. Genes, Enzymes and Regulation of Arginine Biosynthesis in Plants. Plant Physiol. Biochem. 2005, 43, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, A. England: Cambridge University Press. 2015, pp. 1–372.
- Wu, B.; Li, D.; Zhao, J.; Liu, C.; Sun, C.; Chen, J.; Pan, J.; Han, Z.-B.; Hu, J. Vertical Distribution of Sedimentary Organic Carbon in the Yap Trench and Its Implications. China Environ. Sci. 2018, 38, 3502–3511. [Google Scholar] [CrossRef]
- Johnson, G.C.; Toole, J.M. Flow of Deep and Bottom Waters in the Pacific at 10°N. Deep. Res. Part I 1993, 40, 371–394. [Google Scholar] [CrossRef]
Figure 1.
Map showing the sampling sites in the Yap Trench. PHS: the Philippine Sea plate; PAC: the Pacific plate; CP: the Caroline plate; MT: the Mariana Trench; YT: the Yap Trench; PT: the Palau plate; CR: the Caroline Ridge.
Figure 1.
Map showing the sampling sites in the Yap Trench. PHS: the Philippine Sea plate; PAC: the Pacific plate; CP: the Caroline plate; MT: the Mariana Trench; YT: the Yap Trench; PT: the Palau plate; CR: the Caroline Ridge.
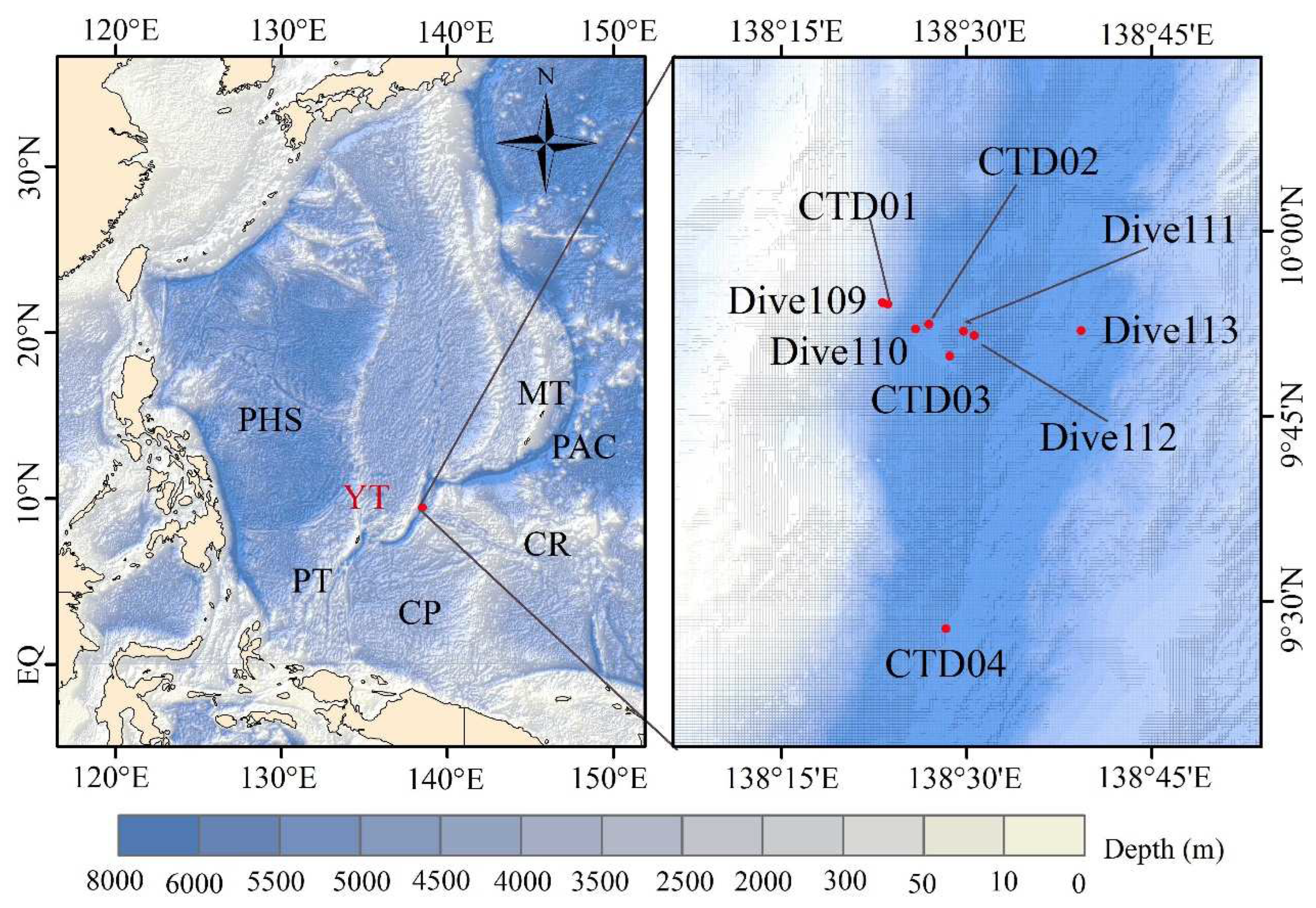
Figure 2.
Box and whisker plots of DFAA, DCAA and THAA in the seawater from the Yap Trench. * means significant (p < 0.05).
Figure 2.
Box and whisker plots of DFAA, DCAA and THAA in the seawater from the Yap Trench. * means significant (p < 0.05).
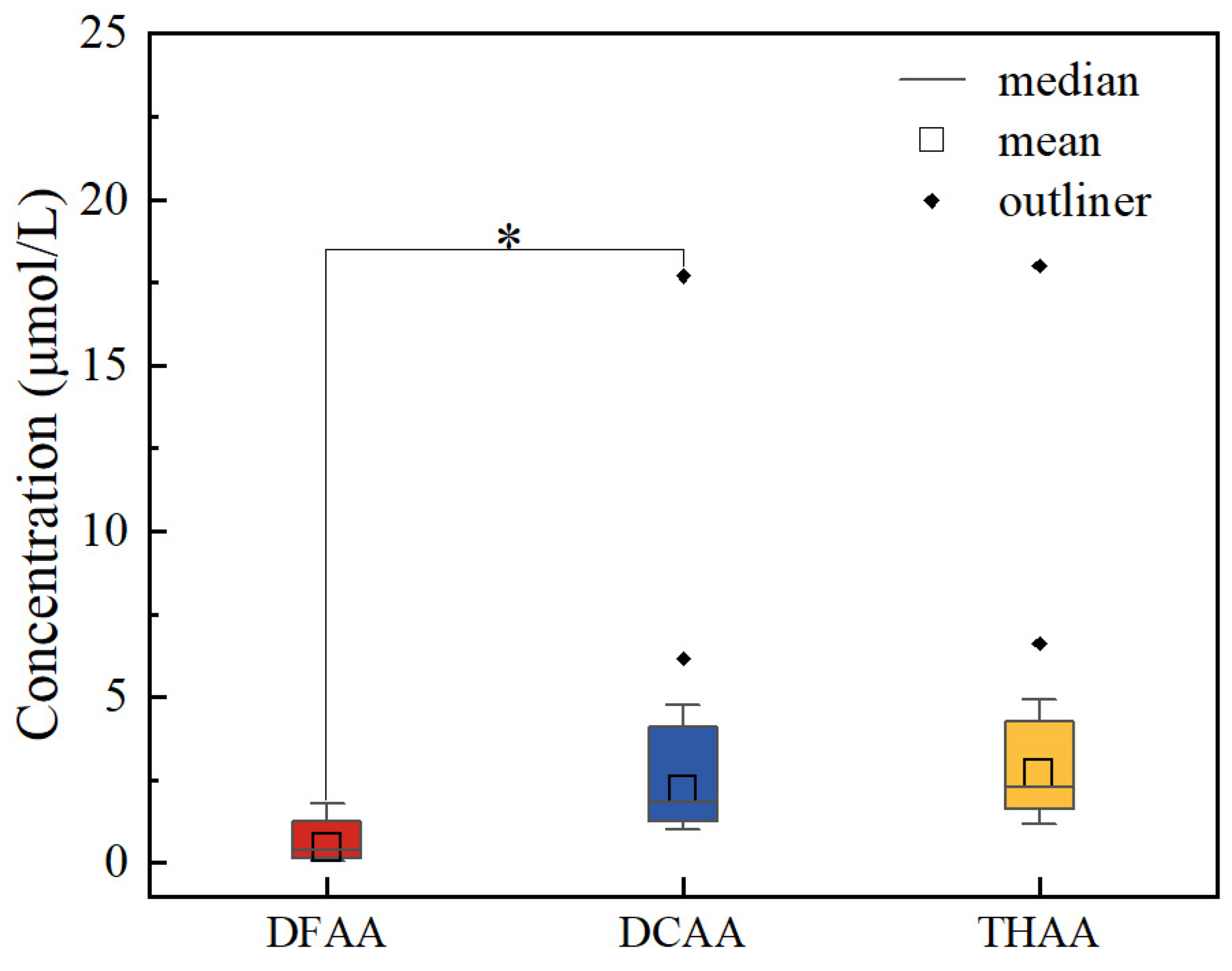
Figure 3.
Mean molar fraction of individual amino acids in DFAA, DCAA and THAA in the seawater from the Yap Trench.
Figure 3.
Mean molar fraction of individual amino acids in DFAA, DCAA and THAA in the seawater from the Yap Trench.
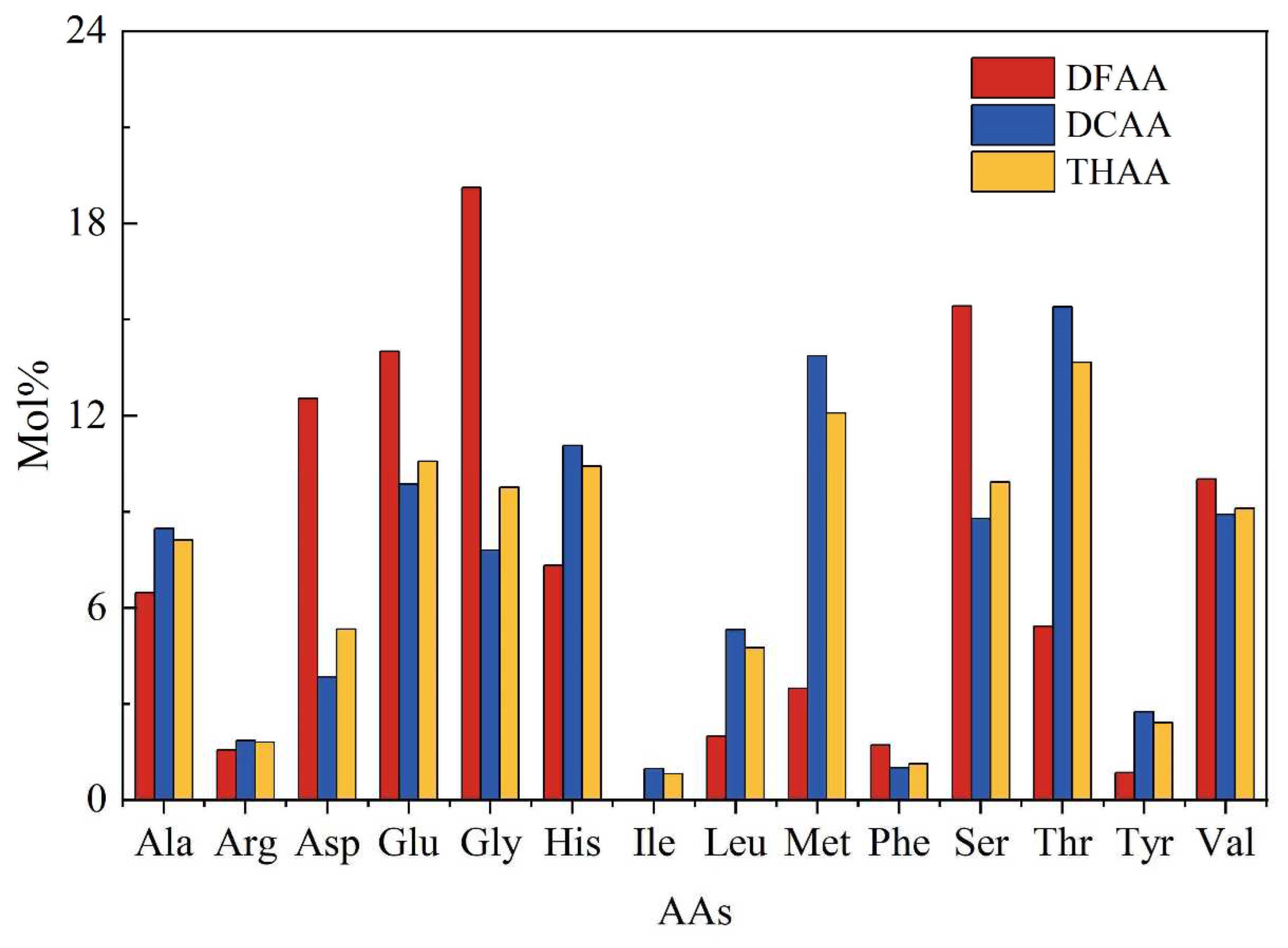
Figure 4.
The percentage content of acid, neutral and basic amino acids in the seawater of the Yap Trench.
Figure 4.
The percentage content of acid, neutral and basic amino acids in the seawater of the Yap Trench.
Figure 5.
The vertical variations of concentrations of DFAA, DCAA and THAA in the seawater from the Yap Trench.
Figure 5.
The vertical variations of concentrations of DFAA, DCAA and THAA in the seawater from the Yap Trench.
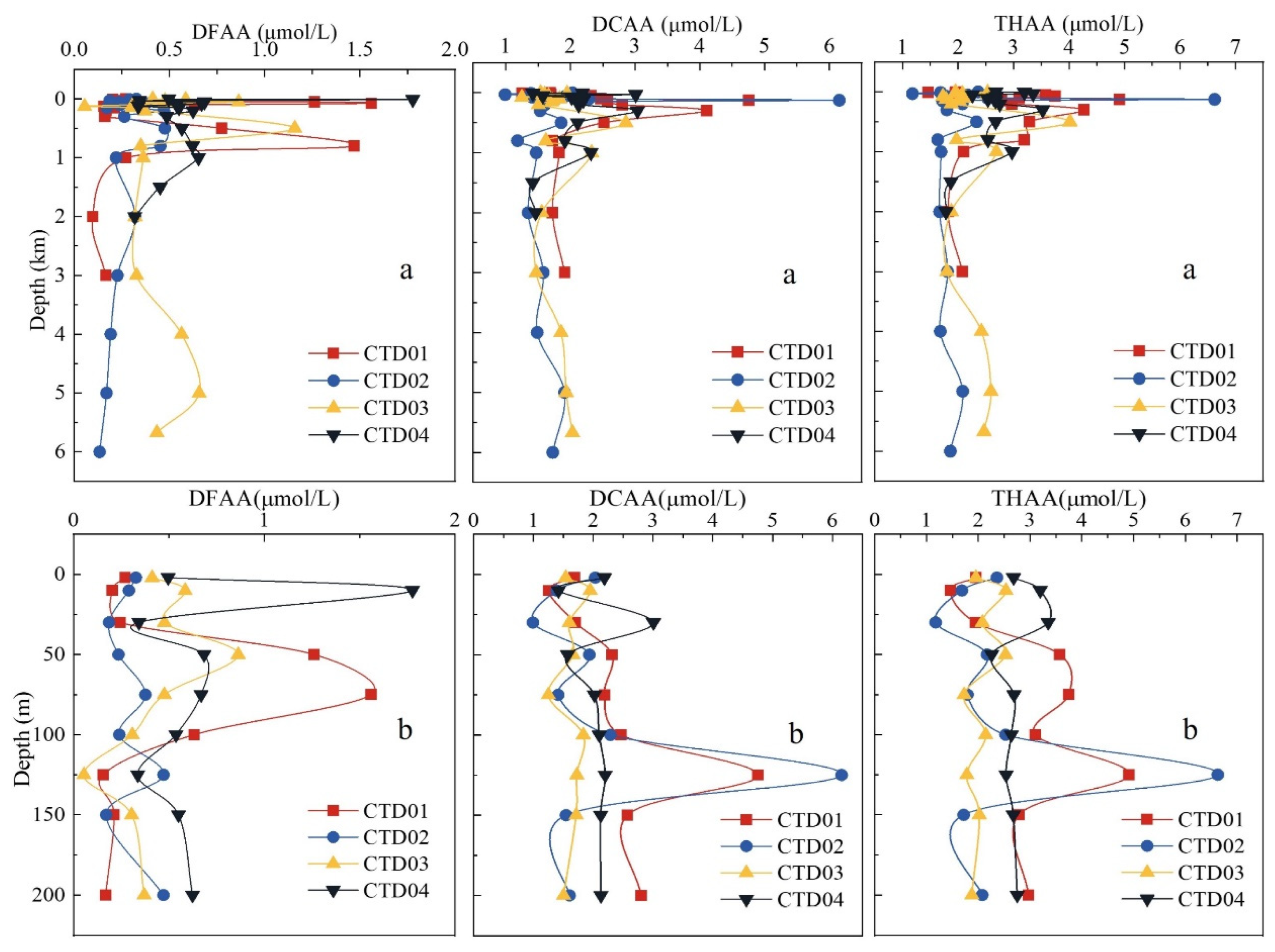
Figure 6.
Loading plot for DFAA, DCAA and THAA in the seawater from the Yap Trench.

Figure 7.
Mean DI values of DFAA, DCAA and THAA in different layers of the seawater in the Yap Trench. S-S represents sediment-seawater interface.
Figure 7.
Mean DI values of DFAA, DCAA and THAA in different layers of the seawater in the Yap Trench. S-S represents sediment-seawater interface.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



